
Identification of the ovine mannose receptor and its possible role in Visna/Maedi virus infection
Upload
independentCategory
view
2download
0
1992 80: 2363-2373
N Harris, M Super, M Rits, G Chang and RA Ezekowitz mediated by interferon-gamma occurs at the level of transcriptiondemonstration that the downregulation of receptor expression Characterization of the murine macrophage mannose receptor:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
reserved.Copyright 2011 by The American Society of Hematology; all rightsWashington DC 20036.by the American Society of Hematology, 2021 L St, NW, Suite 900, Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly
For personal use only. by guest on July 26, 2011. bloodjournal.hematologylibrary.orgFrom

Characterization of the Murine Macrophage Mannose Receptor: Demonstration That the Downregulation of Receptor Expression Mediated by Interferon-y
Occurs at the Level of Transcription
By Neil Harris, Michael Super, Miriam Rits, Grace Chang, and R. Alan B. Ezekowitz
The macrophage mannose receptor (MMR) is a 175-Kd cell- surface transmembrane glycoprotein that is expressed on tissue macrophages where it functions both to mediate the uptake of mannose-rich glycoproteins and as a phagocytic receptor for bacteria, yeasts, and other pathogenic microor- ganisms. In this report we describe the cloning of the full-length cDNA of the mouse macrophage mannose recep- tor and we investigate the level at which interferon y (IFN-y) downregulates mannose receptor expression. The latter is a marker of the functional state of the cell as high levels are expressed on resident and inflammatory macrophages, whereas cells activated by treatment with IFNr have de- creased-to-absent cell-surface mannose receptor expression. The murine MMR cDNA contains an open reading frame that predicts a protein of 1,456 amino acids. Transient expression of the protein in heterologous cells shows that this cDNA encodes a functional mannose receptor. The deduced amino acid sequence of this protein has an overall 82% homology with the human mannose receptor and as such, the
ACROPHAGES play a central role in host defense M in collaboration with antibody, complement, and T lymphocyte^.'-^ The macrophages function as effector cells and contribute to the induction of the immune response. Tissue macrophages often form a lattice beneath epithelial surfaces in the lung, gut, kidney, and skin and as such play a role in first line host defense.'J Direct recognition of foreign antigens is a primary function of tissue macro- phages and it is these cells that express phagocytic receptors such as F2j4 and complement receptor^,^ and high levels of mannose receptor? Although recognition of complex sur- faces that decorate the cell walls of pathogens may well involve cooperation between a number of macrophage surface receptors, recent experiments have shown that each of the major phagocytic receptors, viz, Fc1496,7 complement,s and mannose receptor^,^ can function alone when ex- pressed in heterologous cells. Transient transfection of the human mannose receptor into COS-1 cells showed that the receptor is sufficient to mediate the binding of Candida albicans and Pneumocystis carinii. 9 ~ 1 0 These microorganisms, as well as other pathogens, have cells walls that contain mannose-rich glycans. The latter are rarely, if ever, present as exposed moieties in eukaryotic proteins. Binding of pathogens via the mannose receptor on the macrophage surface initiates secretion of a wide array of inflammatory mediators including arachidonic acid metabolites, reactive oxygen intermediates, and neutral proteinases.'S2
The cDNA of the human mannose receptor has been characterized9J1 and was found to encode a transmem- brane protein of 1,438 amino acids after cleavage of the signal peptide. The receptor possesses a short transmem- brane region and a C-terminal cytoplasmic tail of 28 and 45 residues, respectively. There are also eight tandem carbohy- drate recognition domains (CRDs), each bearing the fea- tures of a calcium-dependent or C-type animal lectin.12 The C-type lectin family is very d ive r~e . '~ J~ Some members are
ectodomain contains an N-terminus that is cysteine-rich followed by a fibronectin type II domain and eight carbohy- drate recognition domains (CRDs). The ectodomain is linked to a hydrophobic transmembrane region and a 46-amino acid cytoplasmic tail. All of the eight CRDs are particularly well conserved, especially CRD4, which shows 92% homology with the equivalent region of the human protein. Steady- state levels of murine MMR mRNA were measured in the macrophage cell line J774E, which is known to express the protein a t the cell surface. These levels were decreased by a 4- to 8-hour incubation with IFN-y, but were almost abolished by overnight treatment with this cytokine. Nuclear run-on experiments showed that IFN-y inhibits MMR gene transcrip- tion. Therefore, the regulation of mannose receptor expres- sion by IFNr provides a novel system in which to study the mechanisms by which this cytokine represses gene expres- sion. o 1992 by The American Society of Hematology.
transmembrane proteins, eg, the asialoglycoprotein recep- tor13-15 and the selectins,16-'s while others are secreted humoral proteins, eg, the serum mannose-binding pro-
The mannose receptor is the only member of this family in which a monomer contains more than one CRD.
In this study we report the characterization of the murine macrophage mannose receptor D N A . Both the cDNA and the predicted sequence of the murine protein display a high degree of homology with the human macrophage mannose receptor (MMR) sequence. The CRDs, the N-terminal cysteine-rich region, and the domain with the features of a fibronectin type I1 repeat are particularly well conserved. Because the latter two regions are known not to participate in the binding to carbohydrate,21 their sequence conserva- tion suggests a hitherto unrecognized function. In addition to its physiologic role as a p i n o c y t i ~ ~ , ~ ~ - ~ ~ and phagocytic re~eptor,5,'~." the expression of the MMR is a sensitive marker of the functional state of the macrophage in that high levels are expressed on nonactivated macrophages.
From the Division of HematologvlOncology, Children's Hospital and Dana-Farber Cancer Institute; and the Department of Pediatrics, Harvard Medical School, Boston, MA.
Submitted May 28,1992; accepted July 7, 1992. Supported by National Institutes of Health Grant No. ROI
AI23786-05 (R.A.B.E.), the South Afncan Medical Research Council (N.H.), and R.A.B.E. is an Established Investigator of the American Heart Association.
Address reprint requests to Neil Hams, MD, Enders Building, 7th Floor, The Children's Hospital, 300 Longwood Ave, Boston, MA 021 1.5.
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. section I734 solely to indicate this fact.
0 1992 by The American Society of Hematology. 0006-4971 /92/8009-0026$3.00/0
Blood, Vol80, No 9 (November 1). 1992: pp 2363-2373 2363
For personal use only. by guest on July 26, 2011. bloodjournal.hematologylibrary.orgFrom

2364 HARRIS ET AL
Treatment of macrophages with interferon y (IFN-y) induces the activated phenotype which, while it includes the upregulation of many genes, causes downregulation of MMR cell surface e x p r e s s i ~ n . ~ ~ , ~ ~ Although much attention has focussed on studying IFN-7 inducible genes, little is known about IFN-y-modulated gene repression. To investi- gate whether decreased mannose receptor surface expres- sion is regulated transcriptionally or posttranscriptionally, we chose to examine the murine macrophage cell line J774E,28 which is known to express cell-surface mannose receptor. We first established, by means of a nuclease protection assay, that IFN-y treatment results in a decrease in the stable accumulation of MMR-specific mRNA. A nuclear run-on analysis then demonstrated that this de- crease can be accounted for by a decrease in the transcrip- tion of the mannose receptor gene.
MATERIALS AND METHODS
Identijication of Murine cDNA An oligo-dT-primed mouse peritoneal macrophage hgtll cDNA
library (Clonetech Laboratories Inc, Cat. No. ML1005b; Palo Alto, CA) was plated on Escherichia coli strain Y1088 and was screened as previously d e ~ c r i b e d ~ , ~ ~ , ~ ~ by plaque hybridization with a 32P- labeled 1.9-kb PstI fragment derived from the full-length human macrophage mannose receptor cDNA.~ Briefly, prehybridization and overnight hybridization of the nylon filters (Amersham, Hy- bond N Arlington Heights, IL) was performed at 42°C in 6X SSC, 5X Denhardt’s solution, 0.5% sodium dodecyl sulfate (SDS), and 100 Kg/mL denatured salmon sperm DNA. Filters were washed at 42°C in 1X SSC, 0.1% SDS.
The screening of approximately 500,000 plaques from the murine cDNA library yielded a single positive phage clone (hgtll.1) that was isolated by two rounds of amplification. This library was rescreened at a higher stringency using a radiolabeled 3’ 274-bp Hind I11 fragment derived from the single positive phage. This procedure identified a second positive overlapping cDNA phage clone (hgtll.2).
A 292-bp BamHI fragment of hgtll.1 provided a probe to screen a XzapII random-primed mouse peritoneal macrophage cDNA library (MACZAP3, kindly provided by Dr Paul Crocker). This library was plated on E coli strain BB4. Plaque hybridization was performed at 65°C in 6X SSC, 5X Denhardt’s, 0.5% SDS, and 100 pg/mL salmon sperm DNA. The nylon filters were washed at 65°C in 0.2X SSC. Four overlapping positive phage clones were obtained after screening approximately 300,000 plaques.
Analysis of cDNA Clones Agtll.1 and xzAP3.3. Phage DNA was isolated from 500-mL
large-scale liquid lysate cultures by cesium chloride density gradi- ent ultracentrif~gation.~~ The cDNA inserts were excised by EcoRI digestion and subcloned into pUC18 (Pharmacia LKB Biotechnol- ogy Inc, Piscataway, NJ). This procedure had to be used for the hZAP3.3 clone because of the repeated failure of the automatic AZAP phage excision.
The phage insert was amplified from 4 pL of purified phage stock using Taq polymerase and oligonucleotide primers specific for the hgtll arms.31 The amplified product was obtained after 30 cycles (30 seconds melting at 94”C, 30 seconds of primer annealing at 55”C, and a 3-minute extension at 72°C). The product was digested with EcoRI and was subcloned into pUC18.
The nucleotide sequence of the cDNA clones was determined by double-stranded sequencing of the DNA using a Sequenase
Agfll.2.
version 2 kit (US Biochemical Corp, Cleveland, OH), which uses a recombinant T7 DNA polymerase in the Sanger dideoxy chain- termination protocol.32
First-Strand cDNA Synthesis One microgram of total RNA from the macrophage cell line
J774E was briefly heat-denatured, snap-cooled, and primed with 50 pmol of an antisense 26-mer complementary to nucleotides 509 to 534 of the murine mannose receptor cDNA. Reverse transcription proceeded at 42°C for 1 hour and then at 52°C for 30 minutes in the presence of 25 U AMV reverse transcriptase (Boehringer Mann- heim Biochemicals, Indianapolis, IN), 50 mmol/L Tris-HC1 pH 8.3, 40 mmol/L KCI, 6 mmol/L MgC12, 2.5 mmol/L dithiothreitol (DlT), 10 U Ribonuclease inhibitor (Promega Corp, Madison, WI), and 2.5 mmol/L of each dNTP (Pharmacia). The total volume of the reaction was 50 pL.
After first-strand cDNA synthesis, excess primers and nucle- otides were removed by passing the sample through a Magic Miniprep column (Promega). The sample was eluted in water and concentrated to 10 KL by lyophilization.
Rapid Amplijication of cDNA Ends (RACE) This was performed according to a modification (K. Sastry,
personal communication, 1990) of the protocol of Frohman et al.33 First-strand murine mannose receptor cDNA was tailed with poly-G as follows: 10 pL of the cDNA preparation was added to 4.6 pL of 5X Terminal Transferase Buffer (Boehringer Mannheim), 4 pL of 1 mmol/L dGTP, and 2 pL (50 U) of terminal transferase (Boehringer Mannheim). The final reaction volume was 20 pL. The reaction was incubated at 37°C for 15 minutes and then inactivated at 65°C for 5 minutes. Finally, the reaction mixture was diluted to 500 pL with Tris-EDTA (TE) pH 7.5. Ten microliters of this product was then amplified with 3 oligonucleotide primers by means of the polymerase chain reaction (PCR). Two of the primers, KS1 and KS2, were sense “adaptor” primers.33 These primers both possess a number of useful 5’ restriction sites (including EcoRI) for the purposes of subcloning. KS1 was the “adaptor-dC” primer while KS2 was the “adaptor” primer (KS1 5’ GCATGCGCGCGGCCGCGGAATTCCCCCCCCCCCCCC 3’; KS2 5’ GCATGCGCGCGGCCGCGGACC 3’). The third primer was the antisense 26-mer complementary to nucleotides 509 to 534 (primer A). This primer contained a 5’ BamHI site.
Ten microliters of the polyG-tailed first-strand cDNA was mixed with 10 pmol KS1, 25 pmol KS2, and 25 pmol of primer A. PCR was performed in a final reaction volume of 50 pL in the presence of 250 pmol/L of each dNTP, 2 mmol/L DTT, 1X PCR buffer (Boehringer Mannheim), and 2.5 U Taq polymerase (Boeh- ringer Mannheim). Amplification was performed over 30 cycles. Melting was at 94°C for 30 seconds, annealing at 55°C for 30 seconds, and extension at 72°C for 3 minutes. The PCR product was purified and was digested with EcoRI and BamHI and was subcloned into the polylinker site of pUC18.
Mammalian Cell Cultures Cell cultures were performed at 37°C in 5% C02/95% humidi-
fied air. J774E cells were obtained as a kind gift of Dr Philip Stahl (Washington University School of Medicine, St Louis, MO). These cells were maintained in RPMI-1640 (GIBCO BRL, Grand Island, NY) containing 10% fetal calf serum (FCS) (GIBCO). COS-1 cells were grown in Dulbecco’s Modification of Eagle’s Minimal Essen- tial Medium (DMEM; GIBCO) and supplemented with either 10% FCS or 10% newborn calf serum. The former was used to maintain the cells in culture while the latter was used in the transfection protocol.
For personal use only. by guest on July 26, 2011. bloodjournal.hematologylibrary.orgFrom

MURINE MANNOSE RECEPTOR 2365
All tissue culture media were supplemented with penicillin (50 U/mL) and streptomycin (50 pg/mL).
Construction of a Full-length Mannose Receptor cDNA The 5-kb hZAP3.3 cDNA clone was the longest that was isolated
and it contained the nucleotides that corresponded to in number from 175 to 5,100 of the full-length cDNA. A uniqueBspH1 site was present 121 bp from the 5‘ end of this clone. This corresponded to the BspHI site at position 296 of the full-length cDNA. The hZAP3.3 clone was excised from the pUC18 vector and was subcloned into the EcoRI site of a mammalian expression vector (obtained as a kind gift of Dr Brian Seed, Massachusetts General Hospital, Boston). This vector contained a unique MZuI site in the polylinker immediately upstream of the EcoRI cloning site. Restric- tion digest analysis of these plasmids containing subcloned hZAP3.3 allowed selection of those with the correct 5’ to 3’ orientation. These constructs were then sequentially digested with BspHI and MZUI.
The region encompassing nucleotides 1 to 489 of the 534 bp 5’ RACE product was amplified by a PCR. The 5’ PCR sense primer contained a unique MZuI site that was absent from the RACE template. Melting was at 94°C for 1 minute, annealing at 55°C for 2 minutes, and extension was performed at 72°C for 3 minutes. The PCR proceeded over 30 cycles. The product of this reaction was sequentially digested with BspHI (which cuts at position 296) and with MluI that would cut at the new 5’ MZuI site. The 296-bp fragment that was generated by this digest was then vectorially subcloned into the BspHIIMZuI-cut pZAP3 expression vector. This generated a full-length cDNA construct termed pMMR. Partial sequence analysis confirmed that the ends of the construct were as predicted.
Expression of the Mannose Receptor in Heterologous Cells COS-1 cells were transfected as previously described34 with the
full-length murine mannose receptor construct pMMR. The day after the transfection, the cells were trypsinized and were replated into 6-well 35-mm diameter cluster plates (Costar). The next day, a suspension of zymosan particles conjugated with the fluorescent probe BODIPY (BODIPY-Zymosan; Molecular Probes Inc, Eu- gene, OR) that was added to a final concentration of 20 pg/mL. The cells were incubated with the zymosan particles for 30 to 60 minutes at 37°C in 5% C02/95% humidified air. The medium containing nonadherent cells was then gently aspirated and the cells were pelleted by centrifugation. Both these nonadherent cells as well as the adherent monolayer of COS-1 cells were treated with trypsin-EDTA at 37°C for 10 minutes (0.05% trypsin, 0.53 mmol/L EDTA GIBCO). The cells that had been previously adherent were now aspirated from the wells after addition of fresh medium containing serum and were pooled with the nonadherent fraction. The cells were then pelleted and resuspended in DMEM plus 10% serum. This suspension was used to make cytospin preparations onto glass microscope slides. The cytospin preparations were fixed in methanol and were stained by the Wright-Giemsa procedure. Because the BODIPY fluorescence was not quenched, this treat- ment allowed easy scoring of particles using both epi-fluorescence and low direct light.
Isolation of Total RNA From Cells This was performed according to the single-step acid guandin-
ium thiocyanate phenol-chloroform extraction procedure as de- s~ribed.3~ RNA samples were stored at -80°C.
IFN-?, Recombinant murine IFN-y was obtained as a kind gift of
Genentech Inc (South San Francisco, CA). Just before use, the IFN was diluted in RPMI containing 10% fetal bovine serum (FBS). The IFN was used at concentrations of 200 to 500 U/mL in all experiments with J774E cells.
Ribonuclease Protection Assay cDNA clone hgtll.1 in pUC18 was digested with Hind 111 and
Pst I. A 299-bp fragment of this digest was subcloned into the vector pGEM-4Z (Promega). This 299-bp fragment encompassed the area corresponding to nucleotides 3660 to 3923 (264 bp) of the full-length cDNA (the extra 35 bp consisted of a fragment of the pUC18 plylinker). The orientation of the subcloned fragment was determined by partial sequencing. PluII digestion of the construct yielded a linear 681-bp blunt-ended fragment that contained both the insert as well as the SP6 and T7 RNA polymerase promoter sites. This fragment was used as a template for in vitro transcrip- tion. Radiolabeled 570-bp antisense mannose receptor RNA transcripts were generated from 0.5 pg of template in the presence of 15 U of SP6 polymerase (Promega). In a similar fashion, a human y-actin control riboprobe was transcribed off a pSP6 y-actin vector.36 In vitro transcription was performed in a final reaction volume of 20 pL and in the presence of 1X transcription buffer (Promega), 10 mmol/L DTT, 30 U ribonuclease inhibitor (Prome- ga), 0.5 mmol/L of ATP/GTP/UTP, 0.015 mmol/L CTP, and 100 pCi of 0132P CTP (Amersham PB 40382). Transcription proceeded at 37°C for 1 hour and was terminated by the addition of 1 U of DNAse for an additional 15 minutes at 37°C (RQDNAse, RNAse- free; Promega). An extra 30 U RNAse inhibitor was added at this stage to protect the riboprobe.
The riboprobes were then extracted in phenol/chloroform plus isoamyl-alcohol and were ethanol precipitated in the presence of ammonium acetate (1 vol extract, 1 vol 4 mol/L ammonium acetate, 4 ~0195% ethanol). The probes were then washed several times in 70% ethanol and dried briefly before being finally resuspended in 100 pL sterile RNAse-free water.
For the protection assay, 20 pg of each sample of total cellular RNA was resuspended in 24 pL of hybridization buffer comprising 40 mmol/L PIPES pH 6.4,400 mmol/L NaCI, 1 mmol/L EDTA, and 80% (vol/vol) formamide. Freshly prepared mannose receptor and y-actin riboprobes, 2 to 5 x 105 cpm, were added to each of the RNA suspensions. The samples were heated at 95°C for 5 minutes and were immediately transferred to 37°C for overnight hybridiza- tion. The total volume of the reaction mix was 25 pL. After hybridization, 350 pL of RNAse buffer (RNAse A 40 pg/mL, RNAse TI 2 U/mL, 10 mmol/L Tris-Hcl pH 7.5,5 mmol/L EDTA, 300 mmol/L NaCI) was added to each sample. Digestion was allowed to proceed at 37°C for 20 minutes. This digestion was terminated by the addition of SDS and proteinase K (Boehringer Mannheim) to final concentrations of 0.5% (wt/vol) and 0.27 mg/mL, respectively, with a further 15-minute incubation at 37°C. Samples were then extracted by phenol/chloroform plus isoamyl- alcohol, ethanol precipitated, and dried without additional ethanol washes. They were then reconstituted in 10 pL of gel loading buffer (US Biochemicals, Sequencing Stop Buffer), heated at 90°C for 5 minutes, and snap-cooled on ice. The samples were electrophore- sed on a 5% polyaclylamide gel containing 7 mol/L urea in the presence of Tri-Borate EDTA buffer (TBE, 89 mmol/L Tris- Borate, 2 mmol/L EDTA). The gel was dried and was subjected to autoradiography at -80°C. 4x174 DNA (GIBCO-BRL) that had been digested with HaeIII and labeled with dGTP in the presence of Klenow fragment DNA polymerase sewed as the molecular size standards in the electrophoresis.
For personal use only. by guest on July 26, 2011. bloodjournal.hematologylibrary.orgFrom

2366 HARRIS ET AL
Transcription Run-on Assay DNA, 2 to 5 pg, was used at each position of the dot-blot. The
appropriate plasmid was denatured by hiling for I O minutes in 200 pL of TE pH 7.4 containing 0.2 mollL NaOH. After boiling, the sample was made up to 400 pL with IOX SSC and was neutralized with 44 pL of 2 mol/L HEPES. The sample was loaded onto a prewetted nylon membrane (Amersham, Hybond N) using a dot-blot apparatus (Schleicher & Schuell, Minifold I; Keene, NH). Each well of the apparatus was rinsed with 6X SSC. The membrane was dried and the DNA was linked to the nylon by UV irradiation.
The run-on assay was performed according to previously de- scribed protocol^.^^^^ Subconfluent J774E mouse macrophage cells were cultured overnight in the presence or absence of 500 U/mL of recombinant murine IFNq (Genentech Inc). The cells were resuspended in a hypotonic lysis buffer containing 1% NP-40W and were briefly homogenized to release the nuclei that were stored in a glycerol buffer at -7O’C. Transcription run-on was performed at 30°C for 30 minutes in a buffer containing 1 mmollL ATP, CTP. and GTP plus 125 pCi of a”?P UTP (Amersham, 3,ooO Cilmmol). The sample was then digested sequentially with DNase1 and with Proteinase K. An extraction with phenollchloroform was followed by TCA precipitation onto Millipore HA filters (Millipore, Bed- ford, MA). The samples were eluted from the filters, alkalinized briefly, and then neutralized according to the protocol.”x The RNA was then precipitated and partially dried before being resuspended in the hybridization solution.
The labeled nascent RNA transcripts were hybridized with nylon strips containing the various plasmids described in the legend to Fig 6. Hybridization and washing of the nylon strips was performed at 65°C.37.3x
RESULTS AND DISCUSSION
Mannose Receptor cDNA and Primary Sequence A human mannosc rcccptor cDNA fragmcnt cncompass-
ing thc region coding for Carbohydrate rccognition domains (CRDs) 3 through 7 was uscd as a probc to scrccn an
(A)
oligo-dT primcd mouse pcritoncal macrophage hgt 1 1 cDNA library. Two positivc cloncs wcrc sclcctcd from - 5 x los plaqucs and wcrc purificd, subcloncd, and scquenccd. The two cloncs, hgt 1 1. I and hgt 1 1.2, wcrc found to contain an ovcrlapping nuclcotidc scqucncc (Fig 1 B). The combincd scquencc of 3.8 kb had a significant dcgrcc of homology (75% to 80%) with thc nuclcotidc scquencc downstrcam of position 1368 of thc human mannosc rcccptor cDNA. Clonc hgt11.2 containcd a long poly-A stretch consistcnt with a poly-A tail. A 5’ fragmcnt of clone hgtll.1 corrc- sponding to nuclcotidcs 1368 to 1660 of the human cDNA was uscd to scrccn a random-primcd AZAP murinc macro- phage cDNA library. Four positivc overlapping clones, hZAP3.2, hZAP3.3, hZAP3.5, and hZAP3.10, were sc- lcctcd from - 4 x 10s plaques. hZAP3.3 of - 5 kb was both thc longest clone as well as that with thc grcatcst 5‘ cxtcnt (Fig 1B). This clonc was homologous to nuclcotidcs 179 through 5150of the human cDNA and it containcd most of the scquencc of the previous two hgtll cloncs, with thc exception of thc most 3’ untranslatcd region and thc poly-A tail.
Because further screening of the availablc mouse cDNA libraries yielded no furthcr cloncs, we uscd thc prcviously dcscribcd PCR-bawd RACE proccdurc3’ to obtain an additional 5’ murinc cDNA scqucncc. This approach (scc Matcrials and Mcthods) gcncratcd a 538-bp product with 75% to 80% homology to thc human mannosc rcceptor cDNA from nuclcotidc 1 to 538 (Fig IB). Whcn thc scqucncc of thc RACE product was ovcrlappcd with thc hZAP3.3 clonc, thc combincd scqucncc had significant homology with thc full-lcngth human scqucncc. Thc first nuclcotidc of the RACE product corrcspondcd to thc most 5’ cxtcnt of the human cDNA scquencc.”J’ Translation of the dcrivcd full-lcngth murinc scqucncc showcd a long
N:c
CYS Fn II 1 2 3 4 5 6 7 8 T M RICH
I I 500 1000 1500 2000 2500 3000 3500 4000 4500 5000
x g t 11.1 +
x g f 11.2
XZAP 3.3
RACE
Fig 1. Murine mannose recep- tor cDNA. (A) Schematic view of the macrophage mannose re- ceptor showing the structural domains. The numbering 1 to 8 refers to the carbohydrate recog- nition domains. Abbreviations: CYS RICH, cysteine-rich; FN It, fibronectin type II repeat; TM, transmembrane segment. (E) Mu- rine mannose receptor cDNA clones. The restriction map of the full-length cDNA is given at the top. The protein coding re- gion is shown in this map as an open box. The extent of the cDNA clones are depicted as lines be- low the main restriction map. The arrowhead on clone Agtll.2 indicates a poly-A tail. The no- menclature of these clones is de- scribed in the text. The nucle- otide sequence can be obtained via EMBL accession no. 211974.
For personal use only. by guest on July 26, 2011. bloodjournal.hematologylibrary.orgFrom

MURINE MANNOSE RECEPTOR 2367
open reading frame that terminated with a stop codon at position 4467. The ATG at position 100 through 103 of the murine cDNA was determined to be the translation start site, based on the very close homology with the previously sequenced human cDNA.~,"
Analysis of the deduced primary amino acid sequence of the murine macrophage mannose receptor protein shows a striking homology with its human counterpart. The two molecules show an overall 82% identity at the amino acid level. The first 18 amino acids following the initiator methionine have the features of a signal sequence, viz, a hydrophobic core and a charged residue following the methionine. A leucine residue 18 amino acids downstream of this methionine was deduced to be the N-terminus of the mature protein. This again was based on the homology between the human and the mouse sequence.
The mature murine macrophage mannose receptor pos- sesses 1,438 amino acids and is therefore identical in this regard to the human receptor. The sequence displays seven potential N-glycosylation sites while the human equivalent has eight." Like the human p r~ te in ,~J l the murine man- nose receptor possess an N-terminal cysteine-rich domain followed by a domain with close homology to the type I1 repeat found in fibronectin (see below). The latter is in turn followed by eight CRDs, a transmembrane region, and a C-terminal cytoplasmic tail (Fig 1A). The extent of the eight CRDs, as well as their alignment, was mapped using previous data from the human receptor sequence and with the assistance of the recently published crystallographic analysis of the rat MBP-A carbohydrate recognition do-
main.39 This allowed us firstly to define the position of each of the domains together with their connecting stalks. Secondly, we were able to align the eight murine CRDs both with one another as well as with the single CRD of rat MBP-A (Fig 2). Weis et aP9 have demonstrated that when a number of animal C-type lectins are aligned, 37 amino acid positions are generally conserved. In some cases, a particu- lar type of amino acid is conserved, while in other cases it is the class of amino acid that is conserved, eg, aliphatic or aromatic. All the CRDs of the mouse mannose receptor have at least 27 of these conserved residues (Fig 2). CRD5 has 34, CRD2 has 33, while CRD4 and CRD6 each have 31.
In Fig 3 the close homology between the murine and human mannose receptors is displayed. These data are summarized in Table 1. Almost all regions of the two proteins show a high degree of amino acid identity and many of the substitutions are conservative. CRD4 is the most highly conserved of the carbohydrate recognition domains with 92% identity between the human and mouse proteins. This is consistent with the recent observation that CRD4 is important in determining both the carbohydrate- binding affinity and specificity of the intact mannose recep- tor.21 Indeed, CRD4 appears to be the only CRD that can bind to carbohydrate in isolation. However, our data demonstrate significant conservation of all the carbohy- drate recognition domains despite evidence that not only are CRDs 1 through 3 nonessential for carbohydrate recognition,21 they also fail to bind to mannose-sepharose affinity columns, unlike CRDs 4 through 8 or 5 through 8?l
The issue of functional importance also arises when one
A K T . . . . . . .
S S E P G K S . S L N P G K N A
R . L T Y S
X G X % W Z P B B ~ n c e x G W N D C % C
Fig 2. The alignment of the eight CRDs of the murine mannose receptor. The eight CRDs of the murine macrophage mannose receptor have been aligned with one another and with the single CRD of rat MBP-A according to the criteria of conserved amino acid positions set out in ref 39. There are 37 such positions, some of which refer to a single invariant amino acid while others encompass a particular class of amino acids. The conservation is indicated by the single letters on the bottom line of the figure. The predicted secondary structures within the CRDs, viz, crhelices, B-sheets, and loops (L) are depicted on the line below the mannose receptor domains. Note that no single domain fulfils all 37 criteria, which were based on the alignment of a number of C-type lectin carbohydrate recognition domains.= The following code is used (see ref 39): e, aliphatic; 4, aromatic; x. aliphatic or aromatic; 2, E or Q; B. D or N; 0, D. N. E, or Q (carbonyl oxygen atom in side chain). Amino acids that have been conserved in five or more mannose receptor CRDs have been shaded. These have been selected independently of the abovementioned criteria.
For personal use only. by guest on July 26, 2011. bloodjournal.hematologylibrary.orgFrom

2368 HARRIS ET AL
CRD 8
Fig 3. Alignment of the murine and human mannose receptor domains. Selected regions of the mature murine and human mannose receptor amino acid sequences have been aligned with respect to one another. The murine sequence is shown above the human sequence and conserved amino acids are shaded. The regions shown include the two N-terminal domains of the mature protein (cysteine-rich and fibronectin type II) as well as the eight CRDs together with the transmembrane region and the C-terminal cytoplasmic tail. The stalks refer to the segments which connect the CRDs. The amino acid numbering is given at the beginning of each domain. Both the mouse and human numbering are given at the points where these differ.
examines the two N-terminal domains of the mannose receptor. The human and murine N-terminal domains are extensively conserved (Table 1) with the fibronectin type I1 repeat showing the greatest sequence conservation of the entire protein (93% identity). Although this suggests some essential function, none has yet been assigned to these N-terminal domains. Like the first three CRDs, neither the N-terminal cysteine-rich domain nor the fibronectin type I1 repeat are required for carbohydrate binding.2I The cysteine- rich domain appears to be unique to the mannose receptor in that a similar region has not been identified in any other known protein sequence. The fibronectin type I1 repeat, which is found in a number of proteins (see below), was named for its presence in the extracellular matrix protein fibronectin."O The latter is a dimer of two large 250-Kd subunits, each containing multiple copies of three types of homology units, types I, 11, and III.@ Each subunit contains two type I1 repeats and these are present in the region of the molecule known to interact with collagen.@ The type I1 repeats are related to the Kringle domains of blood clotting and fibrinolytic proteins and appear to be derived from the same ancestral protein."* The fibronectin type I1 repeat is present not only in fibronectin and the mannose receptor, but also in clotting factor XII,42 the bovine seminal plasma protein PDC-109," as well as in the cation-independent mannose-6-phosphate re~eptor.''''.~5 Despite its ability to interact with phosphorylated carbohydrate residues, the
latter is not a member of the C-type lectin family. It functions to direct lysosomal enzymes tagged with mannose- 6-phosphate to the appropriate lysosomal ~ompartment44,~~ and also serves as a receptor for the growth factor IGF II.& The only protein domain common to the macrophage mannose receptor and the cation-independent mannose-6- phosphate receptor appears to be the fibronectin type I1 repeat. However, the extensive conservation of this domain in the macrophage mannose receptor as well as its presence in a variety of secreted, transmembrane, and extracellular proteins suggests that it may have an important function. One postulated function is that this domain plays a role in macrophage adherence to the extracellular matrix and in the subsequent spreading of the ce11.9 In addition, it is possible that the interaction with certain classes of patho- gens may be enhanced by this region of the ectodomain. The high degree of conservation between the mouse and the human receptors provide support for this concept.
Expression of the Mannose Receptor in Heterologous Cells
The human mannose receptor has been shown to retain its phagocytic properties when expressed in heterologous cells.9 Therefore, we investigated whether the same was true for the murine equivalent. A full-length murine man- nose receptor cDNA construct was generated by joining the 5' RACE product to the longest cDNA clone XZAP3.3 (see
For personal use only. by guest on July 26, 2011. bloodjournal.hematologylibrary.orgFrom

MURINE MANNOSE RECEPTOR 2369
D W A S V T I V A C D S K S E V O K W C C K N D T i F E I 1: o T E L v F N Y O N n o r K N I L L V K G S ~ L W S I I W K V I o T T D D i c s R O V E A M D W V A I T t V A C D S K S E F O X W E C K N D T L L 0 I It 0 E D L F F N V O N R O E K N I M L Y K O S O L W S I I W K V 0 T T O h L C S R O V E A M
O W A G 0 9 P F R V L h L P G S P S S E E B K S C V S L N P 0 % N A X W Cb! L E C V D K L O Y I C K K O W S D R 9 P F R V L N L P G S P 9 A E P O K S C V S L N P O X N A L W E N L E C V D K L O Y I C K K
t o L N D t x I o M v F E W s D o t t V T F r K W L P a E P s H c N N n o t Q c v v u L O X D E Y W A D R A C E O P L o Y t c 6 t G L N 0 : K I a M Y f ~ w s D O T P V T F T K W L R a E P s H E N Y R o e D C V V M K O K D Q V W A D n o c e w p L E v : c e
V O N K O T F R W T V D E O V O F T I I W N A D M P ~ ~ X A G c V A M X T o V A Q E L W D V L S C E I K A K F V E X H I O T K O T F ~ W T I E E E V R F T ~ W N S O M P O R L P O c V A U R T o A P G L W D V L K C D I K A K f v c ~ m
v 8 E L F w L G L T Y O S P s E o r T W S D O S P v s v E N w A V o E P N u v o N v E V c a P L K O D P O M S W ~ D I N c E n L N Y W I c o I v H L L F w a L T v O S P s E o F T W ~ D O S P V S v f N w AI G E P N N V o N v E v C Q E L R O D P T M S W I P D : N c E H L N N W I c a I
f I O M L I S M D K K F I W Y D O S U V D F V A W A T Q E P N F A N D D E n c v r u v T N S O F W N O I Y C O V P N N P I c a n f i G L L I S L D K K F A W M D O P K V D V V S W A T O ~ P N F A N F D E U C V T Y V S N B ~ P W U D I N C ~ V P N A ~ I C O R
N A W T o L N D I n A E ~ M c L W T A G C I O V ~ V T N W G K o v P O O R n 8 s L s v E D A O C V V V I s o N a n f A O T w M o O T c o S K o e ~ t c D T s A W T o L N D V I( s E ~ T f L W T D O R O V ~ V T N W Q K B v P Q Q R n s n L s v E O A O C Y V I I G O A S N f A Q K w M D D T c D s u R O Y : C O T
N N E v T W T D R w R v n v T N W O A D E P K L K S A C v v M 0 v o o v w n T s v c N i s ~ y F L C K L D N O V T W T 0 % W R V R Y T N W A A O f t K L U S A C V V L 0 L D Q V W 6 f A H C N E S F V f L C I R
W I O U F n N V E o L W ~ W L N D N P V S P V N W K T O D P e O E R N O C V V L A S S S Q ~ W N N I H C S S V K ~ F I c K M W I E. F n N V E O T W L W I N N S P V S F V N W N T G O P $ O E R N D C V T L H A S S G F W S N I H C S S Y K O V I c K n
1438 P B T S D I K D L M O N I E O N E h A l l P B T S D M * D L V G W I E O N E H S V I
1438
Fig 3. (Cont'd).
Table 1. Comparison of the Amino Acid Sequences of the Individual Domains of the Murine and Human Mannose Receptors
Domain % Identity
Signal peptide Cysteine-rich Fibronectin type 11 Carbohydrate Recognition Domains
CRD 1 CRD 2 CRD 3 CRD 4 CRD 5 CRD 6 CRD 7 CRD 8
Transmembrane section Cytoplasmic tail Stalk regions connecting CRDs
Stalk 1 Stalk 2 Stalk 3 Stalk 4 Stalk 5 Stalk 6 Stalk? Stalk 8 Stalk 9
67 86 93
88 89 81 92 84 80 76 84 68 74
82 79 56 79 88 74 57 86 67
The identities between the human and murine sequences were determined using the alignment function of the Microgenie sequence analysis program (SciSoft, Inc, Distributed by Spinco Division, Beck- man Instruments, Inc, Palo Alto, CA).
Materials and Methods). This construct was inserted into a mammalian expression vector and was transfected into COS 1 cells. Transfected cells displayed cell surface fluores- cence when stained with a mannose receptor-specific anti- body (results not shown) and were able to phagocytose zymosan particles (Fig 4). This figure shows both the adherence of the zymosan particles to the cell surface as well as the apparent engulfment of these particles by plasma membrane processes. Interestingly, the BODIPY fluorescence was not quenched by the cytospin or by staining with Wright-Giemsa. Therefore, examination of the cells by fluorescence microscopy together with low levels of direct tungsten lamp illumination was possible and this enabled a clear demarcation of the stained cell pro- cesses around the fluorescent zymosan particle as shown in Fig 4. Previous experiments have shown that such actively phagocytosing COS-1 cells are absent from mock-trans- fected cultures or even from cultures transfected with the high-affinity F, receptor CD64.9 Therefore, the full-length cDNA appears to encode a functional mannose receptor.
While the sequence data 3' of nucleotide 175 was derived from the hZAP3.3 clone, the region encompassing nucleoti- des 175 through 296 of the full-length construct originate from the RACE product and it replaces the equivalent region of the hZAP3.3 clone. Sequence analysis of the full-length construct showed that it contains two nucleotide substitutions when compared with the hZAP3 clone. Two T + C substitutions were present, one at nucleotide position 179 and the other at position 202. These differ-
For personal use only. by guest on July 26, 2011. bloodjournal.hematologylibrary.orgFrom
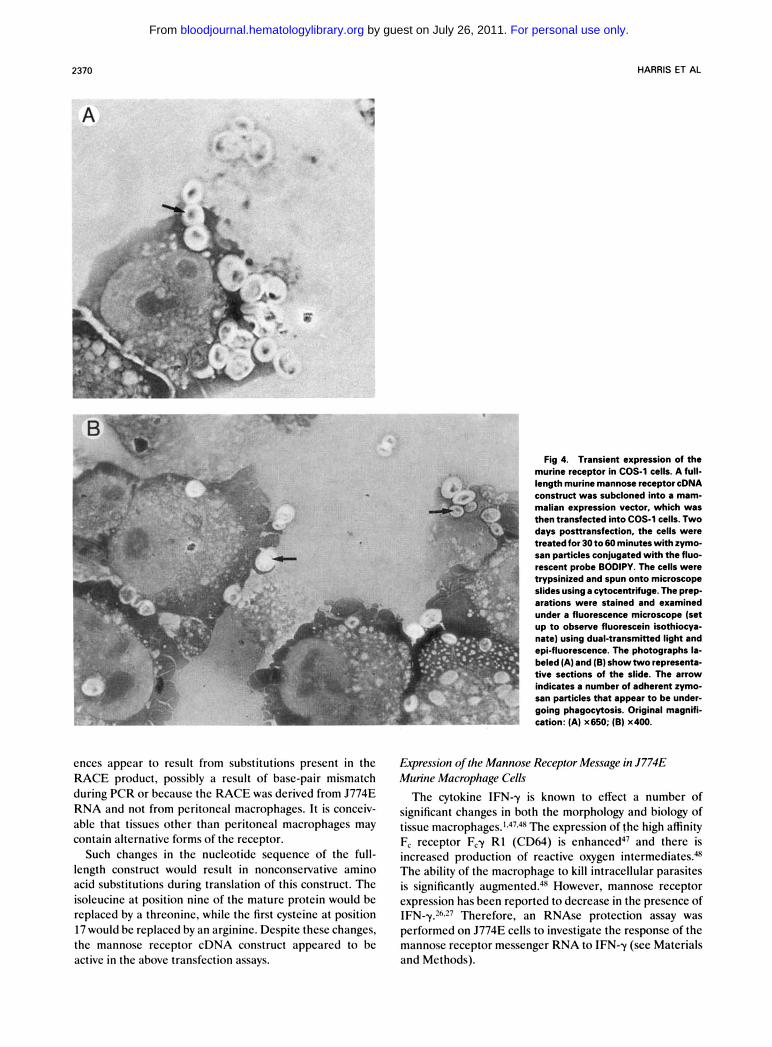
2370 HARRIS ET AL
A \-I I
Fig 4. Transient expression of the murine receptor in COS-1 cells. A full- length murine mannose receptor cDNA construct was subcloned into a mam- malian expression vector, which was then transfected into COS-1 cells. Two days posnransfection, the cells were treated for 30 t o 60 minutes with zymo- san particles conjugated with the fluo- rescent probe BODIPY. The cells were hypsinized and spun onto microscope slides using a cytocentrifuge. The prep- arations were stained and examined under a fluorescence microscope (set up t o observe fluorescein isothiocya- nate) using dual-transmitted light and epi-fluorescence. The photographs la- beled (A) and (6) show two representa- tive sections of the slide. The arrow indicates a number of adherent zymo- san particles that appear to be under- going phagocytosis. Original magnifi- cation: (A) x650; (B) x400.
cnccs appcar to rcsult from substitutions prescnt in the RACE product, possibly a rcsult of base-pair mismatch during PCR or bccausc t hc RACE was dcrivcd from J774E RNA and not from pcritoncal macrophagcs. I t is concciv- ablc that tissucs other than peritoneal macrophagcs may contain altcmativc forms of thc rcccptor.
Such changcs in thc nuclcotidc scqucncc of thc full- lcngth construct would rcsult in nonconscrvativc amino acid substitutions during translation of this construct. Thc isolcucinc at position ninc of the maturc protein would bc replaced by a threonine, whilc the first cysteine at position 17 would be replaced by an argininc. Despite thcsc changcs, thc mannosc rcccptor cDNA construct appcarcd to bc active in thc abovc transfcction assays.
Expression of the Mannose Receptor Message in J774E Mitrine Macrophage Celh The cytokinc IFNy is known to cffcct a numbcr of
significant changcs in both the morphology and biology of tissuc macrophag~s. ' .~~.~~ The cxprcssion of thc high affinity Fc rcccptor Fcy R1 (CD64) is ~ n h a n c c d ~ ~ and thcrc is incrcascd production of rcactivc oxygen intermcdiatcsU Thc ability of the macrophage to kill intracellular parasites is significantly augmcntcd.lx However, mannosc rcccptor cxprcssion has bccn reported to dccrcase in thc prcscncc of IFN-y.2h.27 Thercforc, an RNAsc protection assay was pcrformcd on J774E cclls to invcstigatc the response of the mannosc rcccptor mcsscnger RNA to IFN-y (scc Matcrials and Mcthods).
For personal use only. by guest on July 26, 2011. bloodjournal.hematologylibrary.orgFrom
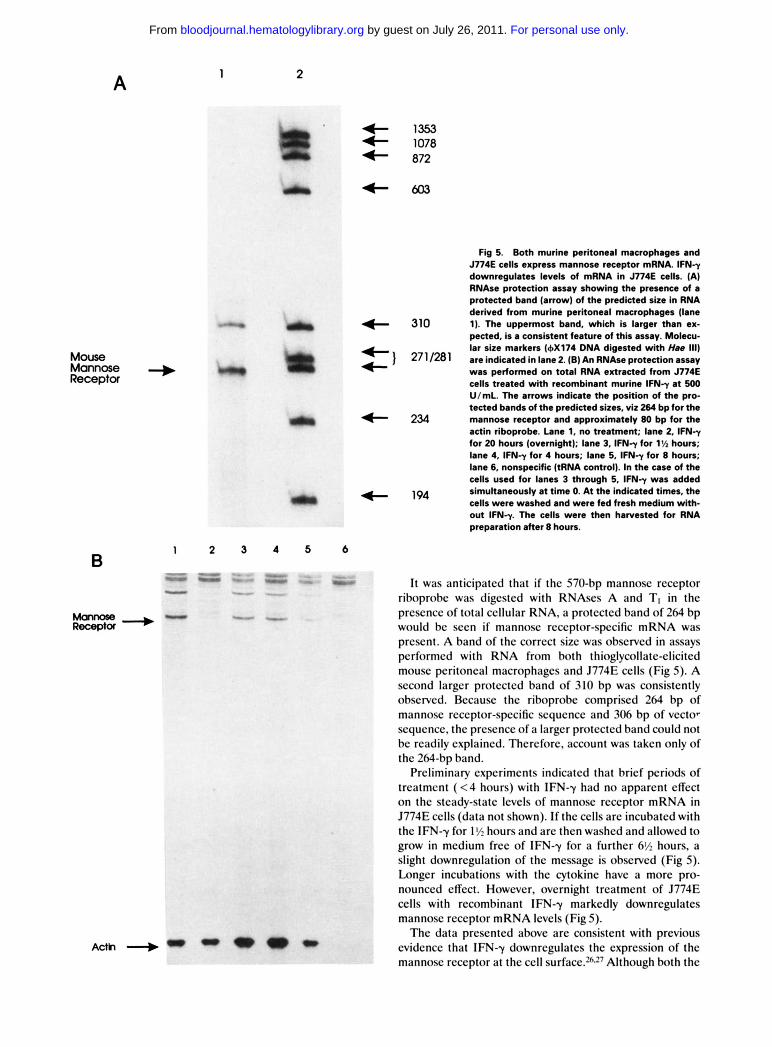
A 1 2
MOW0
Receptor Mannose + 111,
B 1 2 3
-4- Receptor
4 5
Fig 5. Both murine peritoneal macrophages and J774E cells express mannose receptor mRNA. lFNy downregulates levels of mRNA in J774E cells. (A) RNAse protection assay showing the presence of a protected band (arrow) of the predicted size in RNA derived from murine peritoneal macrophages (lane 1). The uppermost band, which is larger than ex- pected. is a consistent feature of this assay. Molecu- lar size markers (6x174 DNA digested with Ha8 111) are indicated in lane 2. (B) An RNAse protection assay was performed on total RNA extracted from J774E cells treated with recombinant murine lFNy at 500 UlmL. The arrows indicate the position of the pro- tected bands of the predicted sizes, viz 264 bp for the
4- 234 mannose receptor and approximately 80 bp for the actin riboprobe. Lane 1. no treatment; lane 2, IFNy for 20 hours (overnight); lane 3, IFNy for 1% hours; lane 4, IFNy for 4 hours; lane 5, IFNy for 8 hours; lane 6, nonspecific (tRNA control). In the case of the cells used for lanes 3 through 5, lFNy was added simultaneously at time 0. At the indicated times, the cells were washed and were fed fresh medium with- out IFNy. The cells were then harvested for RNA preparation after 8 hours.
+ 310
} 2711281 4-
+ 194
6
- -- I t was anticipated that if thc 570-bp mannosc rcccptor riboprobc was digested with RNAscs A and TI in the prcscncc of total ccllular RNA, a protcctcd band of 264 bp would bc sccn if mannosc rcccptor-spccific mRNA was prcscnt. A band of thc corrcct s i x was obscrvcd in assays pcrformcd with RNA from both thioglycollatc-clicitcd mousc pcritoncal macrophagcs and J774E cclls (Fig 5). A second largcr protcctcd band of 310 bp was consistently obscrvcd. Bccausc thc riboprobc compriscd 264 bp of mannosc rcccptor-spccific scqucncc and 306 bp of vcctor scqucncc, thc prcscncc of a largcr protected band could not bc rcadily explained. Thcrcforc, account was takcn only of the 2Wbp band.
Prcliminary cxpcrimcnts indicatcd that bricf periods of trcatmcnt ( < 4 hours) with IFN-y had no apparcnt cffcct on thc stcady-statc lcvcls of mannosc rcccptor mRNA in J774E cclls (data not shown). If thc cclls arc incubatcd with the IFN-y for 1% hours and arc thcn washcd and allowed to grow in medium frcc of IFN-y for a further 6% hours, a slight downrcgulation of thc mcssagc is obscrvcd (Fig 5) . Longer incubations with the cytokinc havc a morc pro- nounced cffcct. Howevcr, overnight treatment of J774E cclls with rccombinant IFN-y markedly downregulates mannosc rcccptor mRNA levels (Fig 5 ) .
Thc data prcscntcd above arc consistent with previous cvidcncc that IFN-y downrcgulatcs the expression of the mannosc rcceptor at thc ccll Although both thc
For personal use only. by guest on July 26, 2011. bloodjournal.hematologylibrary.orgFrom

2372 HARRIS ET AI.
1 2
nn 1
@ t I ?e 0
Fig 6. The effect of 1FN-y on mannose receptor expression in J774E cells appears to be at the level of transcription. cDNA probes for the murine mannose receptor, pMMR (1) or murine p-actin (2) were attached to nylon fitters using a dot-blot apparatus as described in Materials and Methods. In the case of the mannose receptor, the full-length cDNA construct was used. These fitters were hybridized at 6 5 C in the presence of nascent 3zP-labeled RNA from J774E cells either untreated (A) or treated (B) overnight with I F N r .
mannosc rcccptor and F,yRI appear to mcdiatc phagocyto- sis, thcir oppositc rcsponsc to IFN-y makes it plausihlc that whilc thc high-affinity F, rcccptor interacts with opsoniscd microorganisms as part of thc spccific immune rcsponsc, thc mannosc rcccptor functions at an carlicr stagc during
the innatc immune rcsponsc, ic, in thc abscncc of spccific antibodics or activatcd T-hclpcr cclls. To addrcss the qucstion of thc lcvcl of action of the IFN
in the abovc cxpcrimcnts, a nuclcar run-on assay (scc Matcrials and Mcthods) was pcrformcd with J774E cclls that wcrc cithcr untrcatcd or trcatcd ovcrnight with IFN-y at 500 U/mL (Fig 6). This assay dcmonstratcs that the ratc of transcription of thc mannosc rcccptor gcnc is signifi- cantly dccrcascd in the prescnce of IFN-y whcrcas transcrip- tion of actin is unaltcrcd. Rcprcssion of mannose rcccptor gcnc cxprcssion by IFN-y appcars to be cycloheximide scnsitivc (rcsults not shown), and this implics that this effect may be mcdiatcd via a newly synthcsizcd IFN-y- induciblc protcin that would bc a candidatc repressor. We arc at prcscnt attcmpting to map thc cis-acting elements that may bc targcts for this putative trans-acting protcin to tcst this hypothcsis.
ACKNOWLEDGMENT
We thank the members of the laboratory for helpful discussions and Marsha Kartzman for her assistance in preparing this manu- script. We are grateful to Dr Celeste Simon for providing the human y-actin probe and to Dr Nancy Andrews for the murine p-actin probe.
REFERENCES 1. Papadimitriou JM, Ashman RB: Macrophages: Current views
on their differentiation, structure, and function. Ultrastruct Pathol 13343. 1989
2. Ezekowitz RAB: Macrophages-Biological aspects, in Nel- son DS (ed): Natural Immunity. Sydney, Australia, Academic, 1989, p 15
3. Gordon S, Perry VH. Rabinowitz S, Chung L-P, Rosen H: Plasma membrane receptors of the mononuclear phagocyte sys- tem. J Cell Sci Suppl9:1, 1988
4. Ravetch JV, Kinet JP: Fc receptors. Annu Rev Immunol 9457,1991
5. Ezekowitz RAB, Stahl PD: The structure and function of vertebrate mannose lectin-like proteins. J Cell Sci Suppl 9:121, I988
6. Indik Z, Kelly C. Chein P, Levinson AI, Schreiber AD: Human Fc-gamma R11, in the absence of other Fc-gamma recep- tors, mediates a phagocytic signal. J Clin Invest 88:1766, 1991
7. Tuijnman WD, Capel PJA. van de Winkel JGJ: Human low-affinity IgG receptor Fc-gamma Rlla (CD32) introduced into mouse fibrohlasts mediates phagocytosis of sensitized erythrocytes. Blood 79:1651,1992
8. Mosser DM. Springer TA. Diamond MS: Leishmania promas- tigotes require opsonic complement to bind to the human leuko- cyte integrin Mac-I (CDllb/CD18). JCell Biol I l k S I l , 1992
9. Ezekowitz RAB. Sastry K, Bailly P, Warner A: Molecular characterization of the human macrophage mannose receptor: Demonstration of multiple carbohydrate recognition-like domains and phagocytosis of yeasts in Cos-I cells. J Exp Med 172:1785, 1990
IO. Ezekowitz RAB, Williams D, Koziel H. Armstrong M. Warner A, Richards FF, Rose R: Uptake of Pneumocysris curitii mediated by the macrophage mannose receptor. Nature 351: 155, 1991
1 I . Taylor ME, Conary JT, Lennartz MR. Stahl PD, Drickamer K: Primary structure of the mannose receptor contains multiple
motifs resembling carbohydrate recognition domains. J Biol Chem 265:12156,1990
12. Drickamer K Two distinct classes of carbohydrate-recogni- tion domains in animal lectins. J Biol Chem 263:9557,1988
13. Drickamer K: Membrane receptors that mediate glycopro- tein endocytosis: Structure and biosynthesis. Kidney In1 2 3 167, 1987 (suppl32)
14. Ashwell G. Harford J: Carbohydrate-specific receptors of the liver. Annu Rev Biochem 51:531.1982
15. Breitfeld PP, Simmons CF Jr, Strous GJ, Geuze HJ. Schwartz A L Cell hiology of the asialoglycoprotein-receptor system: A model for receptor-mediated endocytosis. Int Rev Cytol 9247. I985
16. Johnston GI, Cook RG, McEver R P Cloningof GMP-140, a granule membrane protein of platelets and endothelium: Sequence similarity to proteins involved in cell adhesion and inflammation. Cell 56: 1033, 1989
17. Lasky LA. Singer MS, Yednock TA, Dowbenko D, Fennie C. Rodriguez H, Nguyen T, Stachel S, Rosen SD: Cloning of a lymphocyte homing receptor reveals a lectin domain. Cell 56:1045, I989
18. Springer TA: Adhesion receptors of the immune system. Nature 34k425.1990
19. Ezekowitz RAB: Ante-antibody immunity. Curr Biol 1:M. 1991
20. Ezekowitz RAB: Antigens coming to a sticky end. Current Biology 2 147, 1992
21. Taylor ME, Bezouska K, Drickamer K: Contribution to ligand binding by multiple carbohydrate-recognition domains in the macrophage mannose receptor. J Biol Chem 267:1719, 1992
22. Stahl P, Schlesinger PH, Sigardson E, Rodman JS, Lee YS: Receptor-mediated pinocytosis of mannose glycoconjugates by macrophages. Characterization and evidence for receptor recy- cling. Cell 19:207, 1980
For personal use only. by guest on July 26, 2011. bloodjournal.hematologylibrary.orgFrom

MURINE MANNOSE RECEPTOR 2373
23. Stahl PD: The macrophage mannose receptor: Current status. Am J Respir Cell Mol Biol2317,1990
24. Drickamer K: Clearing up glycoprotein hormones. Cell 67:1029,1991
25. Sun-Sang J, Nelson RS, Silverstein S C Yeast mannans inhibit binding and phagocytosis of zymosan by mouse peritoneal macrophages. J Cell Biol96:160,1983
26. Mokoena T, Gordon S: Human macrophage activation: Modulation of mannosyl, fucosyl receptor activity in vitro by lymphokines, gamma and alpha interferons, and dexamethasone. J Clin Invest 75:624,1985
27. Ezekowitz RAB, Gordon S: Interaction and regulation of macrophage receptors, in Evered D, Nugent J, O’Connor M (eds): Biochemistry of Macrophages. London, UK, Pitman (Ciba Founda- tion Symposium 118), 1986, p 127
28. Diment S, Leech MS, Stahl PD: Generation of macrophage variants with 5-azacytidine: Selection for mannose receptor expres- sion. J Leuk Biol42485,1987
29. Sambrook J, Fritsch EF, Maniatis T Molecular Cloning, A Laboratory Manual. Cold Spring Harbor, NY, Cold Spring Harbor Laboratory, 1989
30. Sastry K, Herman G, Day L, Deignan E, Burns G, Morton C, Ezekowitz RAB: The human mannose-binding protein gene. Exon structure reveals its evolutionary relationship to a human pulmo- nary surfactant gene and its localization to chromosome 10. J Exp Med 1701175,1989
31. Dorfman DM, Zon LI, Orkin SH: Rapid amplification of 1ambdaGTll bacteriophage library reaction. Biotechniques 7568, 1989
32. Sanger F, Nicklen AS, Coulson AR: DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci USA 745463, 1977
33. Frohman MA, Dush MK, Martin GR: Rapid production of full-length cDNAs from rare transcripts: Amplification using a single gene-specific oligonucleotide primer. Proc Natl Acad Sci USA 85:8998,1988
34. Aruffo A, Seed B: Molecular cloning of a CD28 cDNA by a high efficiency COS cell expression system. Proc Natl Acad Sci USA 84:8573,1987
35. Chomczynski P, Sacchi N Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform ex- traction. Anal Biochem 162156,1987
36. Enoch T, Zinn K, Maniatis T Activation of the human beta-interferon gene requires an interferon-inducible factor. Mol Cell Biol6:801,1986
37. Greenberg ME, Ziff EB: Stimulation of 3T3 cells induces transcription of the c-fos proto-oncogene. Nature 311:433,1984
38. Greenberg ME, Bender TP: Identification of newly tran- scribed RNA, in Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K (eds): Current Protocols in Molecular Biology. New York, NY, Greene Publishing Associates & Wiley-Interscience, 1990, p 4.10.1
39. Weis IW, Kahn R, Foume R, Drickamer K, Henrickson W A Structure of the calcium-dependent lectin domain from a rat mannose-binding protein determined by MAD phasing. Science 254:1608,1991
40. Petersen TE, Thogersen HC, Skorstengaard K, Vibe- Pedersen K, Sahl P, Sottrup-Jensen L, Magnussen s: Partial primary structure of bovine plasma fibronectin: Three types of internal homology. Proc Natl Acad Sci USA 80:137,1983
41. Holland SK, Harlos K, Blake C C F Deriving the generic structure of the fibronectin type I1 domain from the prothrombin Kringle 1 crystal structure. EMBO J 6:1875, 1987
42. McMullen BA, Fujikawa K Amino acid sequence of the heavy chain of human alpha-factor XIIa (activated Hageman Factor). J Biol Chem 260:5328,1985
43. Esch FS, Ling NC, Bohlen P, Ying SY, Guillemin R: Primary structure of PDC-109, a major protein constitutent of bovine seminal plasma. Biochem Biophys Res Commun 113:861,1983
44. Pohlmann R, Nagel G, Schmidt B, Stein M, Lorkowski G, Krentler C, Cully J, Meyer HE, Grzeschik K-H, Mersmann G, Hasilik A, von Figura K Cloning of the cDNA encoding the human cation-independent mannose-6-phosphate-specific receptor. Bio- chemistry 84375,1987
45. Lobe1 P, Dahms NM, Kornfeld S: Cloning and sequence analysis of the cation-independent mannose-6-phosphate receptor. J Biol Chem 263:2563,1988
46. Morgan DO, Edman JC, Standring DN, Fried VA, Smith MC, Roth RA, Rutter WJ: Insulin-like growth factor I1 receptor as a multifunctional binding protein. Nature 329:301,1987
47. Perussia B, Dayton ET, Lazarus R, Fanning V, Trinchieri G: Immune interferon induces the receptor for monomeric IgGl on human moncytic and myeloid cells. J Exp Med 158:1092,1983
48. Nathan CF, Murray HW, Wiebe ME, Rubin B Y Identifica- tion of interferon gamma as the lymphokine that activates macro- phage oxidative metabolism and antimicrobial activity. J Exp Med 158:670,1983
For personal use only. by guest on July 26, 2011. bloodjournal.hematologylibrary.orgFrom
Copyright © 2022 FDOKUMEN




















