
Computational prediction of microRNAs from Oil Palm (Elaeis guineensis Jacq.)expressed sequence tags
7
Asia Pacific Journal of Molecular Biology and Biotechnology, 2007 Vol. 15 (3) : 107-113 *Author for Correspondence. Mailing address: Genetics and Molecular Biology, Institute of Bio- logical Sciences, Faculty of Science, University of Malaya, 50603 Kuala Lumpur, Malaysia. Tel: (603)79675896; Fax: (603)79675908; Email: [email protected] Computational prediction of microRNAs from Oil Palm (Elaeis guineensis Jacq.)expressed sequence tags Nadirah Md Nasaruddin 1 , K.Harikrishna 1 , Rofina Yasmin Othman 2 , Lim Saw Hoon 3 and Jennifer Ann Harikrishna 2 * 1 Sime Darby Technology Centre Sdn Bhd, 2 Jalan Tandang, 46050 Petaling Jaya, Selangor 2 Institute of Biological Sciences, Faculty of Science, University of Malaya, 50603 Kuala Lumpur 3 Biotechnology, Malaysia University of Science and Technology, Block C Kelana Square, 47301 Petaling Jaya, Selangor Received 4 September 2007 / Accepted 5 October 2007 Abstract. MicroRNAs (miRNAs) are defined as ~22nt non-coding RNAs derived from one arm of longer hairpin stem-loop precursors through the action of Dicer, that are capable of silencing gene activities at the transcriptional or post-transcriptional level. miRNA plays important regulatory roles in animal and plant development and as such the identification of miRNA en- coding regions of genomes is an important part of gene annotation and functional genomics. A total of 7,28 7,28 Elaeis guineensis Jacq. expressed sequence tags (EST) were compared to previously known Arabidopsis thaliana and Oryza sativa miRNA, to search for novel miRNA from oil palm. Twenty-five EST contigs with homology to previously known miRNAs were identified. After predicting the RNA secondary structure, five new potential miRNA were identified in oil palm. One of the most promising potential miRNA was miR160 which has very strong characteristics of an miRNA precursor structure and 100% identity with the mature miR160 from other plant species. Keywords. miRNA, oil palm, Elaeis guineensis Jacq., expressed sequence tags INTRODUCTION MicroRNAs (miRNAs) are endogenous ~22 nucleotide small non-coding RNAs that are processed from one arm of their stem-loop hairpin RNA precursors by the action of a Ribonuclease III - like enzyme named Dicer (Reinhart Dicer (Reinhart et al., 2002; Bartel, 200; Adai et al., 2005). miRNA plays important miRNA plays important regulatory roles in animal and plants and acts by gene silenc- ing via either the destruction or blocking of translation of homologous mRNA (Bartel, 200). In plants there is also evidence of miRNA involvement in gene silencing at the transcriptional level through DNA methylation (Bao et al., 2004). The first miRNA, known as The first miRNA, known as lin-4, a gene known to control the timing of Caenorhabditis elegans larval development, was discovered by Victor Ambros and colleagues, Rosalind Lee and Rhonda Feinbaum (Lee et al., 1993). More recently miRNAs have been described from many eukaryotic organ- isms including a wide range of plant species (Bartel, 200). Bartel, 200). In plants the confirmed roles for miRNA include the regu- lation of root development, leaf development and polarity, floral organ identity, flowering time, embroyogenesis, auxin signaling and various stress responses (Mallory and Vaucheret 2006; Yang et al., 2007). Plant miRNAs are also involved in auto-regulation of the miRNA pathway and the biogenesis of transacting siRNAs (ta-siRNAs) (Yang et al., 2007). Plant miRNAs are generally conserved in evolution and Plant miRNAs are generally conserved in evolution and are found in regions of the genome distinct from previously annotated genes (Reinhart et al., 2002; Bartel, 200). Unlike their animal counterparts, many plant miRNAs have perfect many plant miRNAs have perfect or near-perfect homology to their target mRNAs and gen- erally have only a single site within their target (Rhoades et al., 2002). These characteristics facilitate the computational prediction of plant miRNAs and their targets which has resulted in a rapidly growing amount of such sequence data. The miRNA Registry is a web based, searchable database for the storage of published miRNA sequences and for the assignment of miRNA gene names prior to publication (Griffiths-Jones, 2004; Griffiths-Jones et al., 2005). It was . It was created by The Wellcome Trust Sanger Institute and includes more than 5,071 entries of miRNAs found in animals and plants (Release 10.0, August 2007). Out of 1,220 miRNA
Transcript of Computational prediction of microRNAs from Oil Palm (Elaeis guineensis Jacq.)expressed sequence tags

AsPac J. Mol. Biol. Biotechnol., Vol. 15 (3), 2007 107 Computational prediction of microRNAs Asia Pacific Journal of Molecular Biology and Biotechnology, 2007Vol. 15 (3) : 107-113
*Author for Correspondence.Mailing address: Genetics and Molecular Biology, Institute of Bio-logical Sciences, Faculty of Science, University of Malaya, 50603 Kuala Lumpur, Malaysia. Tel: (603)79675896; Fax: (603)79675908; Email: [email protected]
Computational prediction of microRNAs from Oil Palm (Elaeis guineensis Jacq.)expressed sequence tags
Nadirah Md Nasaruddin1, K.Harikrishna1, Rofina Yasmin Othman2, Lim Saw Hoon3 and Jennifer Ann Harikrishna2*
1Sime Darby Technology Centre Sdn Bhd, 2 Jalan Tandang, 46050 Petaling Jaya, Selangor2Institute of Biological Sciences, Faculty of Science, University of Malaya, 50603 Kuala Lumpur
3Biotechnology, Malaysia University of Science and Technology, Block C Kelana Square, 47301 Petaling Jaya, Selangor
Received 4 September 2007 / Accepted 5 October 2007
Abstract. MicroRNAs (miRNAs) are defined as ~22nt non-coding RNAs derived from one arm of longer hairpin stem-loop precursors through the action of Dicer, that are capable of silencing gene activities at the transcriptional or post-transcriptional level. miRNA plays important regulatory roles in animal and plant development and as such the identification of miRNA en-coding regions of genomes is an important part of gene annotation and functional genomics. A total of 7,28��7,28�� Elaeis guineensis Jacq. expressed sequence tags (EST) were compared to previously known Arabidopsis thaliana and Oryza sativa miRNA, to search for novel miRNA from oil palm. Twenty-five EST contigs with homology to previously known miRNAs were identified. After predicting the RNA secondary structure, five new potential miRNA were identified in oil palm. One of the most promising potential miRNA was miR160 which has very strong characteristics of an miRNA precursor structure and 100% identity with the mature miR160 from other plant species.
Keywords. miRNA, oil palm, Elaeis guineensis Jacq., expressed sequence tags
IntroductIon
MicroRNAs (miRNAs) are endogenous ~22 nucleotide small non-coding RNAs that are processed from one arm of their stem-loop hairpin RNA precursors by the action of aa Ribonuclease III - like enzyme named Dicer (ReinhartDicer (Reinhart et al., 2002; Bartel, 200��; Adai et al., 2005). miRNA plays importantmiRNA plays important regulatory roles in animal and plants and acts by gene silenc-ing via either the destruction or blocking of translation of homologous mRNA (Bartel, 200��). In plants there is also evidence of miRNA involvement in gene silencing at the transcriptional level through DNA methylation (Bao et al., 2004). The first miRNA, known asThe first miRNA, known as lin-4, a gene known to control the timing of Caenorhabditis elegans larval development, was discovered by Victor Ambros and colleagues, Rosalind Lee and Rhonda Feinbaum (Lee et al., 1993). More recently miRNAs have been described from many eukaryotic organ-isms including a wide range of plant species (Bartel, 200��).Bartel, 200��).. In plants the confirmed roles for miRNA include the regu-lation of root development, leaf development and polarity, floral organ identity, flowering time, embroyogenesis, auxin signaling and various stress responses (Mallory and Vaucheret 2006; Yang et al., 2007). Plant miRNAs are also involved in
auto-regulation of the miRNA pathway and the biogenesis of transacting siRNAs (ta-siRNAs) (Yang et al., 2007). Plant miRNAs are generally conserved in evolution andPlant miRNAs are generally conserved in evolution and are found in regions of the genome distinct from previously annotated genes (Reinhart et al., 2002; Bartel, 200��). Unlike their animal counterparts, many plant miRNAs have perfectmany plant miRNAs have perfect or near-perfect homology to their target mRNAs and gen-erally have only a single site within their target (Rhoades et al., 2002). These characteristics facilitate the computational prediction of plant miRNAs and their targets which has resulted in a rapidly growing amount of such sequence data. The miRNA Registry is a web based, searchable database for the storage of published miRNA sequences and for the assignment of miRNA gene names prior to publication (Griffiths-Jones, 2004; Griffiths-Jones et al., 2005). It was. It was created by The Wellcome Trust Sanger Institute and includes more than 5,071 entries of miRNAs found in animals and plants (Release 10.0, August 2007). Out of 1,220 miRNA

108 AsPac J. Mol. Biol. Biotechnol., Vol. 15 (3), 2007 Computational prediction of microRNAs
genes predicted and identified from the phylum Viridiplantae, 18�� miRNA gene entries are from Arabidopsis thaliana and 2��3 miRNA gene entries are from Oryza sativa. However, there were no entries for miRNA genes predicted or identi-fied from Elaeis guineensis Jacq. (oil palm) at the time of the present study.
Oil Palm (Elaeis guineensis Jacq.) is a monocotyledonous plant of the palm family (Arecaceae). It is an economically important plant species and provides the second largest source of edible oil in the world, after soy-bean (Jouannic et al., 2005). As a step towards gene identification and discovery in this crop, characterization of the oil palm gene transcrip-tome was undertaken by sequencing cDNA libraries to gener-ate ESTs (Jouannic et al., 2005; Ho et al., 2007). Identifying miRNAs within oil palm ESTs is possible because miRNAs are transcribed by RNA polymerase II as longer precursor molecules and they also may occur within protein coding regions of the genome (Zhang et al., 2005; Jones-Rhoades and Bartel, 200��; Bonnet et al., 200��).
Research using computational methods for the identifica-tion of miRNA genes with sequence data has been increasing, with the development of new algorithms and applications for this specific area of interest (Adai et al., 2005; Qui et al., 2007; Xie et al., 2007; Zhang et al., 2005). Computational methods have been proven to be successful for the discovery of new miRNAs from various animal and plant species including C.elegans (GradGrad et al., 2003), Arabidopsis thaliana, Oryza sativa (Li et al., 2005), Gossypium hirsutum (QiuQiu et al., 2007) and Bras-sica napus (Xie et al., 2007). It has been estimated that thereIt has been estimated that there should be between 1 to 1.67 miRNA per 10,000 plant EST (Zhang et al., 2006; Qiu et al., 2007, XieXie et al., 2007). Thus it). Thus it was reasonable to expect to identify one or more miRNA in the collection of the 7,28�� EST from oil palm.
MAterIAls And Methods
Sequences of known miRNAs and ESTs. The sequences of previously sequenced and published Arabidopsis thaliana and Oryza sativa miRNAs were obtained from the microRNA Registry (Release 9.1, February 2007), resulting in a collectionesulting in a collection of 18�� miRNA sequences of Arabidopsis thaliana and 2��2 sequences from Oryza sativa, representing ��2 miRNA fami-lies. To avoid redundancy, any duplicated miRNA sequences (present within both species) were excluded on the basis of sequence identity. The remaining sequences were defined as the reference set of miRNA sequences, and used as the query for BLAST searches.
The ESTs of oil palm used were previously derived by Ho et al. (2007), from cDNA libraries constructed from shoot apical meristem, immature flowers and mature flowers. The 7,284 ESTs were sorted in a flat file database to be used in the BLAST searches.
Software utilization. BLAST-2.2.15 was downloaded from NCBI ftp site (publicly available at ftp://ncbi.nih.nlm.gov) (Altschul et al., 1997). BLAST searches were used to compare the known mature miRNAs listed in the miRNA Registry against the EST database of oil palm using BLAST parameter settings from Zhang et al., (2005). MFold 3.1 (Mat-thews et al., 1999; Zuker, 2003) was used to predict RNA secondary structure folding of the sequences (accessed at http://frontend.bioinfo.rpi.edu/applications/mfold/cgi-bin/rna-form1.cgi)
EST analysis and prediction of potential miRNAs. Figure 1 shows the workflow of the search for potential miR-NAs in oil palm. Oil palm ESTs which are homologous to previously known miRNAs (n/n, (n-1)/n, (n-2)/n nucleotide matches, n=length of previously known miRNA) from the BLAST searches where chosen and their secondary struc-tures were predicted and generated using mFold. Oil palm ESTs with a maximum of three (≤3) nucleotide mismatches within sequences of known mature miRNAs from Arabidopsis thaliana or Oryza sativa were selected for secondary structure prediction.
The sequences’ secondary structure output were analysed analysed and scored for their potential to form miRNA precursors with the following characteristics (Ambros et al., 2003; Zhang et al., 2006; Xie et al., 2007; Qiu et al., 2007): 1) The RNA sequences should be able to fold into a hairpinThe RNA sequences should be able to fold into a hairpin
structure, such that one arm of the hairpin contains the ~22nt mature miRNA sequence.
2) The predicted secondary structure of the miRNA pre-The predicted secondary structure of the miRNA pre-cursor should have lower minimal free energy (MFE) and minimal free energy index (MFEI) then other types of RNA (e.g. tRNA, rRNA).
3) The predicted mature miRNA should have an A�UThe predicted mature miRNA should have an A�U content of 30-70%.
��) The mature miRNA sequences within the hairpin loopThe mature miRNA sequences within the hairpin loop
Figure 1. Workflow for bioinformatic identification of potential miRNA gene sequences.
AsPac J. Mol. Biol. Biotechnol., Vol. 15 (3), 2007 109 Computational prediction of microRNAs
segment must have less than six mismatches to the op-posite miRNA* sequence in the other arm.
5) Any loop or bulge in the mature miRNA:RNA* dimerAny loop or bulge in the mature miRNA:RNA* dimer should contain a maximum of 3 nucleotides not involved in canonical base pairing.
results And dIscussIon
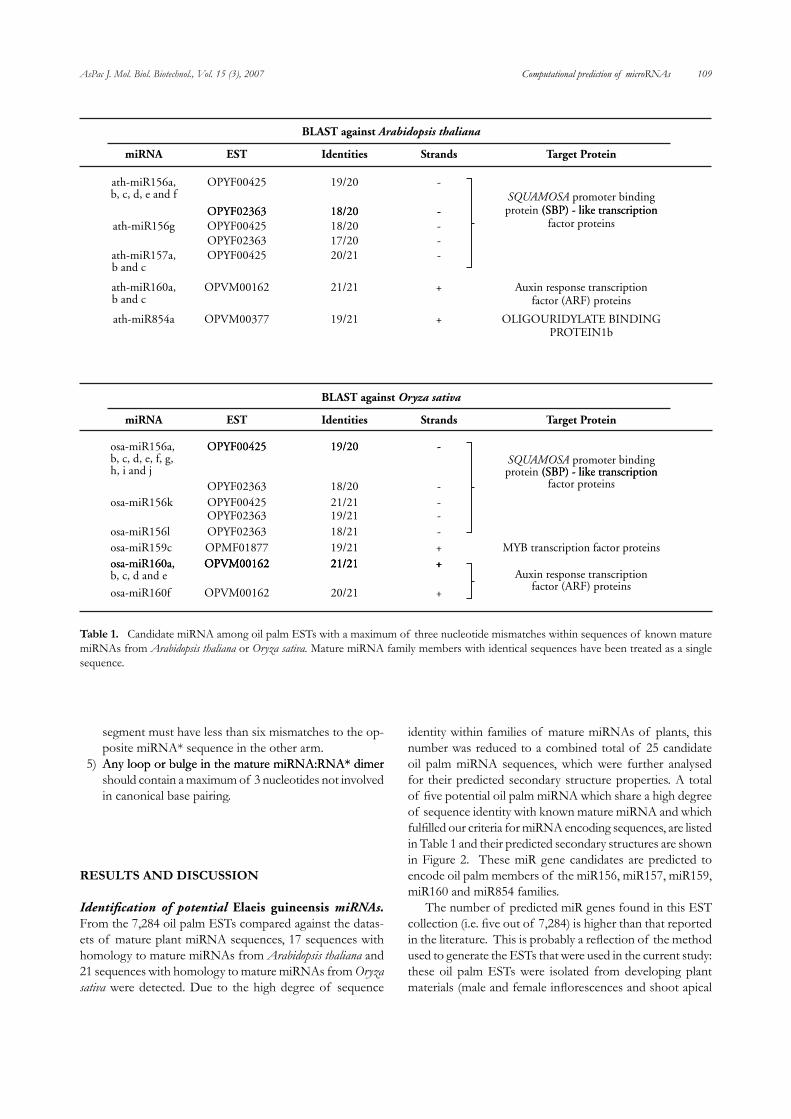
Identification of potential Elaeis guineensis miRNAs. From the 7,28�� oil palm ESTs compared against the datas-ets of mature plant miRNA sequences, 17 sequences with homology to mature miRNAs from Arabidopsis thaliana and 21 sequences with homology to mature miRNAs from Oryza sativa were detected. Due to the high degree of sequence
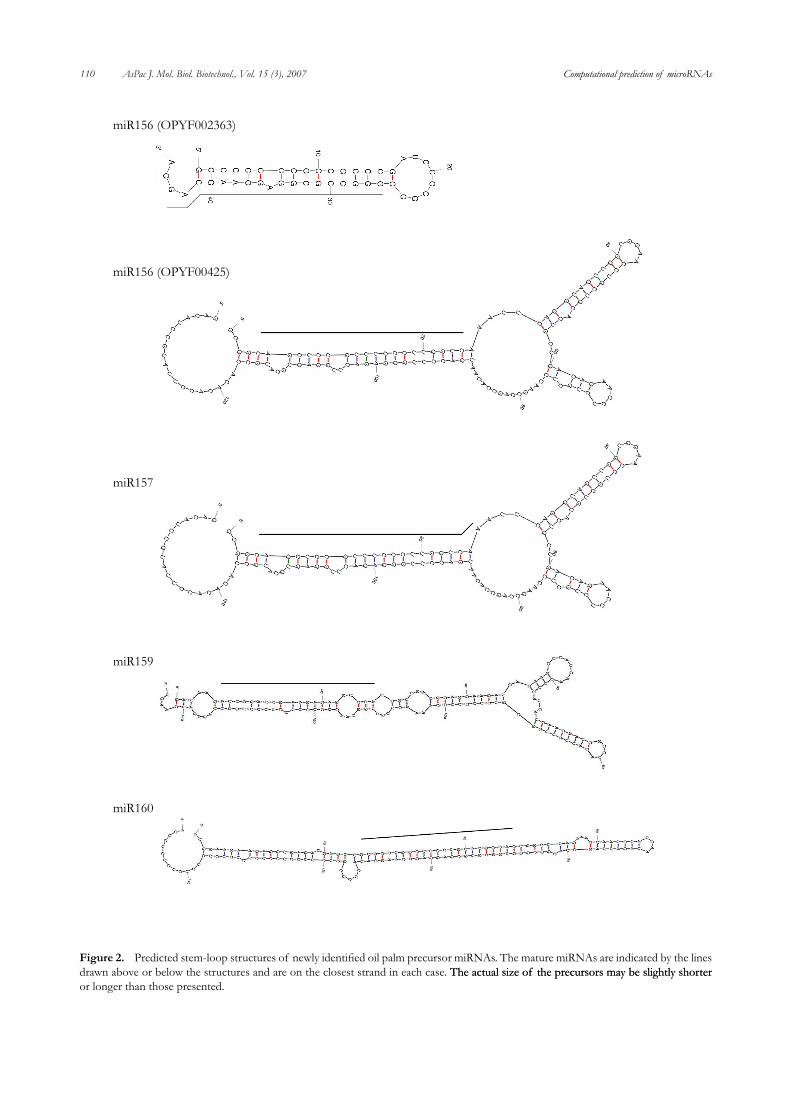
identity within families of mature miRNAs of plants, this number was reduced to a combined total of 25 candidate oil palm miRNA sequences, which were further analysed for their predicted secondary structure properties. A total of five potential oil palm miRNA which share a high degree of sequence identity with known mature miRNA and which fulfilled our criteria for miRNA encoding sequences, are listed in Table 1 and their predicted secondary structures are shown in Figure 2. These miR gene candidates are predicted to encode oil palm members of the miR156, miR157, miR159, miR160 and miR85�� families. The number of predicted miR genes found in this EST collection (i.e. five out of 7,284) is higher than that reported in the literature. This is probably a reflection of the method used to generate the ESTs that were used in the current study: these oil palm ESTs were isolated from developing plant materials (male and female inflorescences and shoot apical
BLAST against Arabidopsis thaliana
miRNA EST Identities Strands Target Protein
ath-miR156a, OPYF00425 19/20 - b, c, d, e and f SQUAMOSA promoter binding OPYF02363 1�/20 -OPYF02363 1�/20 - protein (SBP) - like transcription (SBP) - like transcription ath-miR156g OPYF00425 1�/20 - factor proteins
OPYF02363 17/20 - ath-miR157a, OPYF00425 20/21 - b and c ath-miR160a, OPVM00162 21/21 + Auxin response transcription b and c factor (ARF) proteins ath-miR�54a OPVM00377 19/21 + OLIGOURIDYLATE BINDING PROTEIN1b
BLAST against Oryza sativa
miRNA EST Identities Strands Target Protein
osa-miR156a, OPYF00425 19/20 -OPYF00425 19/20 - b, c, d, e, f, g, SQUAMOSA promoter binding h, i and j protein (SBP) - like transcription (SBP) - like transcription OPYF02363 1�/20 - factor proteins osa-miR156k OPYF00425 21/21 - OPYF02363 19/21 - osa-miR156l OPYF02363 1�/21 - osa-miR159c OPMF01�77 19/21 + MYB transcription factor proteins osa-miR160a, OPVM00162 21/21 +osa-miR160a, OPVM00162 21/21 +OPVM00162 21/21 + b, c, d and e Auxin response transcription osa-miR160f OPVM00162 20/21 + factor (ARF) proteins
table 1. Candidate miRNA among oil palm ESTs with a maximum of three nucleotide mismatches within sequences of known mature miRNAs from Arabidopsis thaliana or Oryza sativa. Mature miRNA family members with identical sequences have been treated as a single sequence.
110 AsPac J. Mol. Biol. Biotechnol., Vol. 15 (3), 2007 Computational prediction of microRNAs
miR156 (OPYF002363)
miR156 (OPYF00425)
miR157
miR159
miR160
Figure 2. Predicted stem-loop structures of newly identified oil palm precursor miRNAs. The mature miRNAs are indicated by the lines drawn above or below the structures and are on the closest strand in each case. The actual size of the precursors may be slightly shorterThe actual size of the precursors may be slightly shorter or longer than those presented.

AsPac J. Mol. Biol. Biotechnol., Vol. 15 (3), 2007 111 Computational prediction of microRNAs
meristem) and were from libraries that had been enriched by subtraction and/or screened using cold plaque hybridisation (Ho et al., 2007). This could be expected to improve the representation of low abundance and differentially regulated transcripts from the selected oil palm tissues used to create the ESTs, thus enriching the data for miRNA sequences.
miR156 and miR157. The miR156 and miR157 families are very closely related and target SQUAMOSA promoter binding protein (SBP) - like transcription factor proteins (Rhoades et al., 2002; Jones-Rhoades and Bartel, 200��), which are involved in the regulation of flowering time (Mallory and Vaucheret 2006). The predicted oil palm members of this family were EST clones OPYF00425 and OPYF02363 both of which were isolated from subtracted cDNA librar-ies made from young male and female inflorescences of oil palm, tissues which would be predicted to express this miR super-family.
miR159. The miR159 family reportedly targets the GAMYB members of the MYB transcription factor fam-ily (Rhoades et al., 2002; Jones-Rhoades and Bartel, 200��), members of which play roles in the regulation of flowering time and leaf development (Mallory and Vaucheret 2006). Achard et al. (200��) demonstrated that miR159 was involved in regulation of the plant hormone gibberellin (GA) and that it’s over expression could result in flowering time delay, LEAFY transcript level reduction and altered floral develop-ment (Zhang et al., 2005b; Achard et al., 200��). The predicted
oil palm member of this family was EST clone OPMF01877 which was isolated from cold plaque screening of a cDNA library made from mature female inflorescences of oil palm, suggesting expression during late stage floral development, although this expression may be at a low level as the cold plaque screening is designed to identify rare transcripts.
miR854. The miR�54 family is thought to target multiple sites in the 3’ untranslated region of OLIGOURIDYLATE BINDING PROTEIN1b (At UBP1b) and to be involved as regulators of basal transcriptional mechanisms in both animals and plants (Arteaga-Vázquez et al., 2006). In Arabi-dopsis thaliana, miR85�� is expressed in stems and developingiR85�� is expressed in stems and developing expressed in stems and developing inflorescences and is thought to be expressed at very low and is thought to be expressed at very low levels in plants (Arteaga-Vázquez et al., 2006). The predictedThe predicted oil palm member of this family was EST clone OPVM00377 which was isolated from an oil palm cDNA library derived from young shoot apical meristem, a tissue which would beyoung shoot apical meristem, a tissue which would be expected to show expression of this transcript.
miR160. The miR160 family is reported to target the Auxin Response Factors (ARF transcription factors) (Rhoades et al., 2002; Jones-Rhoades and Bartel, 200��) which regulate auxin responses, leaf development and floral identity (Mallory and Vaucharet, 2006). Potential targets of miR160 include ARF10, ARF16 and ARF17 (Zhang et al., 2005b; Rhoades et al., 2002). Oil palm EST OPVM00162 contains a 21 nucleotide region which is identical to the sequence of the mature miR160 of Arabidopsis thaliana and Oryza sativa. This EST also has the
miR85��
Figure 2. (continued)

112 AsPac J. Mol. Biol. Biotechnol., Vol. 15 (3), 2007 Computational prediction of microRNAs
strongest characteristics of an miRNA amongst the oil palm EST candidates; The folding free energy of -73.1 kcal mol-1 is within the range for known Arabidopsis thaliana precur-sors which have an average value of around -57 kcal mol-1 (BonnetBonnet et al., 200��). Nineteen nucleotides of the of the 21 Nineteen nucleotides of the of the 21 nucleotide homolog sequence are engaged in canonical (CG, AU or GU) base pairings between the mature miRNA and the opposite arm (miRNA*) in the hairpin loop structure and the stem–loop precursor has no internal loops or bulges greater than three nucleotides. These combined character-istics indicate that OPVM00162 is a strong candidate to encode the miR160 of oil palm. MiR160 is one of the mostMiR160 is one of the most highly conserved families in plants (Yang et al., 2007) so it is not unexpected to find an oil palm sequence with 100% identity to its orthologs. Oil palm miR160 candidate, EST clone OPVM00162, was isolated from a subtractive library from oil palm young shoot apical meristem. The oil palm apical meristem can be expected to be auxin responsive, as a tissue that is under strong developmental control in plants and that may develop into leaf or floral meristems in response to transcriptional regulators. MiR160 regulation of the auxin response is critical in plant development (reviewed in Mallory and Vaucharet, 2006) and thus can be expected to be important for plant productivity in a crop plant such as oil palm.
CONCLUSION
The high degree of evolutionary conservation of mature miRNAs within the plant kingdom permits a bioinformatic approach to prediction of new miRNA orthologs or ho-mologs in other plant species. In this study a collection of 7,28�� oil palm EST sequences expressed in apical meristem, immature and mature flowers were found to contain five potential miRNA encoding sequences by a combined homol-ogy and structural analysis approach. The potential roles of these miRNAs in oil palm include regulation of the auxin response, floral development and the regulation of basal tran-scription. The annotation of these oil palm EST as miR gene candidates provides useful data for the functional analysis of these sequences, which are likely to play important roles in the developmental biology of this major crop plant.
reFerences
Achard P., Herr A., Baulcombe D.C. and Harberd N.P. 200��. Modulation of floral development by a gibberellin-regu-lated microRNA. Development, 131: 3357-3365.
Adai A., Johnson C., Mlotshwa S., Archer-Evans S., Manocha V., Vance V. and Sundaresan V.. 2005. Computational prediction of miRNAs in Arabidopsis thaliana. Genome Research, 15: 78-91.
Altschul S.F., Madden T.L., Schäffer A.A., Zhang J., Zhang Z., Miller W. and Lipman D.J. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Research, 25(17): 3389-3��02.
Ambros V., Bartel B., Bartel D.P., Burge C.B., Carrington J.C., Chen X., Dreyfuss G., Eddy S.R., Griffiths-Jones S., Marshall M., Matzke M., Ruvkun G and Tuschl T. 2003. A uniform system for microRNA annotation. RNA, 9(1-2): 277-279.
Arteaga-Vázquez M., Caballero-Pérez J. and Vielle-Calzada J. 2006. A family of microRNAs present in plants and2006. A family of microRNAs present in plants and animals. The Plant Cell, 18: 3355-3369.
Bao, N., Lye K-W. and M. Barton M.K. 200��. MicroRNA Binding Sites in Arabidopsis Class III HD-ZIP mRNAs Are Required for Methylation of the Template Chromo-some. Developmental Cell, 7: 653–662.
Bartel D.P. 200��. MicroRNAs: Genomics, biogenesis, mecha-nism, and function. Cell, 116: 281-297.
Bonnet E., Wuyts J., Rouze P. and Van de Peer Y. 2004. Evi-dence that microRNA precursors, unlike other non-cod-ing RNAs, have lower folding free energies than random sequences. Bioinformatics, 20(17): 2911-2917.
Grad Y., Aach J., Hayes G.D., Reinhart B.J., Church G.M., Ruvkun G. and Kim J.. 2003. Computational and experi-mental identification of C.elegans microRNAs. Molecular Cell, 11(5): 1253-1263.
Griffiths-Jones S. 2004. The microRNA Registry. Nucleic Acids Research, 32: D109- D111.
Griffiths-Jones S., Grocock R.J. van Dongen S., Bateman A. and Enright J. 2005. miRBase: microRNA sequences, targets and gene nomenclature. Nucleic Acids Research, 3��: 1��0-1����.
Ho C.L., Kwan Y.Y., Choi M.C., Tee S.S., Ng W.H., Lee K.A., Lee Y.P., Ooi S.E., Lee W.W., Tee J.M.,Tan S.H., H. Ku-laveerasingam, Syed Alwee S.S.R. and Ong Abdullah M. 2007. Analysis and functional annotation of expressed sequence tags (ESTs) from multiple tissues of oil palm (Elaeis guineensis Jacq.). BMC Genomics 8:381.
Jones-Rhoades M.W. and Bartel D.P. 200��. ComputationalComputational identification of plant microRNAs and their targets, including a stress-induced miRNA. Molecular Cell. 1��:787-799.
Jouannic S., Argout X., Lechauve F., Fizames C., Borgel A.,

AsPac J. Mol. Biol. Biotechnol., Vol. 15 (3), 2007 113 Computational prediction of microRNAs
Morcillo F., Aberlenc-Bertossi F., Duval Y. and Tregear J.. 2005. Analysis of expressed sequence tags from oil palm (Elaies guineensis). FEBS Letters, 579: 2709-271��.
Lee R.C., Feinbaum R.L. and Ambros V. 1993. TheThe C. elegans heterochronic gene lin-4 encodes small RNAs wih anti-sense complementarity to lin-14. Cell, 75: 8��3-85��.
Li Y., Li W. and Jin Y.X. 2005. Computational identifica-tion of novel family members of microRNA genes in Arabidopsis thaliana and Oryza sativa. Acta Biochim Biophys Sin, 37:75-87.
Mallory A.C. and Vaucheret H. 2006. Functions of micro-RNAs and related small RNAs in plants. Nat Genetics, 38: S31-S37.
Matthews D.H., Sabina J., Zuker M. and Turner D.H. 1999. Expanded sequence dependence of thermodynamic parameters improves prediction of RNA secondary structure. J. Mol. Biol, 288: 911-9��0.
Qiu C.X., Xie F.L., Zhu Y.Y., Guo K., Huang S.Q., Nie L. and Yang Z.M.. 2007. Computational identification of microRNAs and their targets in Gossypium hirsutum expressed sequence tags. Gene, 395: ��9 – 61.
Reinhart B.J., Weinstein E.G., Rhoades M.W., Bartel B. and Bartel D.P. 2002. MicroRNA in plants.MicroRNA in plants. Genes & Develop-ment, 16: 1616-1626.
Rhoades M.W., Reinhart B.J., Lim L.P., Burge C.B., Bartel B. and Bartel D.P. 2002. Prediction of plant miRNA targets.Prediction of plant miRNA targets. Cell, 110: 513-520.
Xie F.L., Huang S.Q., Guo K., Xiang A.L. Zhu Y.Y., Nie L. and Yang Z.M.. 2007. Computational identification of novel microRNAs and targets in Brassica napus. FEBS Letters, 581: 1��6��-1��7��.
Yang T., Xue L. and An L. 2007. Functional diversity of miRNA in plants. Plant Science, 172(3): ��23-��32.
Zhang B.H., Pan X., Cannon C.H., Cobb G.P. and Ander-son T.A.. 2006. Conservation and divergence of plant microRNA genes. Plant Journal, ��6: 2��3-259.
Zhang B.H., Pan X.P., Wang Q.L., Cobb G.P. and Anderson T.A.. 2005. Identification and characterization of new plant microRNAs using EST analysis. Cell Research, 15 (5): 336-360.
Zhang B.H., Pan X.P., Cobb G.P. and Anderson T.A. 2005b. Plant microRNA: A small regulatory molecule with big impact. Dev Biology, 289: 3-16.
Zucker M. 2003. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Research, 31: 3��06-3��15.




















