
Protein kinase C activation during progesterone-stimulated acrosomal exocytosis in human spermatozoa
7
Molecular Human Reproduction vol.2 no.12 pp. 921-927, 1996 Protein kinase C activation during progesterone-stimulated acrosomal exocytosis in human spermatozoa Christine M.B.OToole 1 , Eduardo R.S.Roldan 2 and Lynn R.Fraser 1 - 3 Anatomy and Human Biology, King's College London, Strand, London WC2R 2LS, UK, and 2 Departamento de Reproducci6n Animal, Centro de Investigacidn y Tecnologla, INIA, Ctra de La Coruna Km 5.9, 28040 Madrid, Spain 3 To whom correspondence should be addressed The involvement of protein kinase C (PKC) in exocytosis of the mammalian sperm acrosome is still a controversial issue. Work carried out thus far has failed to provide direct evidence for the activation of this enzyme upon stimulation with natural agonists of acrosomal exocytosis. We have therefore used progesterone stimulation of the acrosome reaction in human spermatozoa to clarify this issue. In spermatozoa preincubated under conditions known to support capacitation and fertilization in vitro, treatment with progesterone caused a time-dependent stimulation of phosphorylation of at least eight proteins ranging in size from -20-220 kDa. The inclusion of the PKC inhibitors chelerythrine chloride or calphostin C reduced the observed phosphorylation in a concentration-dependent manner. Exogenously supplied phorbol 12-myristate-13-acetate (PMA) or the permeant diacylglycerol 1-oleoyl-2-acetyl-sn-glycerol (OAG), synthetic activators of PKC, also stimulated phosphorylation in preincubated spermatozoa, but inclusion of calphostin C diminished the response. Furthermore, the prior inclusion of the 1,4-dihydropyridine Ca 2+ channel antagonist nifedipine also inhibited phosphorylation, suggesting that PKC is activated downstream of Ca 2+ channel opening. Exocytosis triggered by progesterone was significantly inhibited by chelerythrine chloride or calphostin C. Both PMA and OAG triggered exocytosis, but the inclusion of chelerythrine chloride significantly inhibited the response; exocytotic responses were seen only in capacitated cells. These results provide the first direct evidence that PKC activation plays a role in the signal transduction pathway underlying acrosomal exocytosis in progesterone- stimulated capacitated spermatozoa. Key words: Ca 2+ channels/diacylglycerol/fertilization/membrane fusion/phorbol ester Introduction The mammalian sperm acrosome is analogous to a secretory granule and the 'acrosome reaction', an exocytotic process whereby acrosomal contents are released from the spermato- zoon during interaction with the oocyte, is a prerequisite for successful fertilization (Yanagimachi, 1994). Stimulation of spermatozoa to undergo exocytosis causes an increase in intracellular Ca 2+ concentrations, due to influx from the extracellular space, and the activation of an array of signal transduction pathways important for the sequence ending in membrane fusion (F6nichel and Parinaud, 1995). One important component in this chain of events could be the activation of protein kinase C (PKC) because, in somatic cells, this enzyme appears to play a modulatory role during exocytotic responses (Kikkawa and Nishizuka, 1986; Cheek, 1993). However, an unequivocal role for PKC during the acrosome reaction has yet to be demonstrated. The presence of detectable PKC activity in spermatozoa has been disputed (Kimura et al, 1984; Roldan and Harrison, 1988; Rotem et al, 1990a,b; Breitbart et al, 1992). It now seems clear that PKC activity in mammalian spermatozoa is extremely low, ~ 3 % of that seen in the rat brain enzyme, or -20% of that found in PC 12 cells or in rat pituitary cells (based on |Xg protein) (Rotem et al., 1990a,b; Breitbart et al., © European Society for Human Reproduction and Embryology 1992). Furthermore, when activity is compared taking into account the number of cells used to obtain enzyme preparations, the activity of sperm PKC is only 0.4-1.3% of that in PC12 or in pituitary cells (Rotem et al., 1990a). Several studies employing either PKC antagonists or exogen- ous synthetic activators of PKC have provided indirect (i.e. circumstantial) evidence that PKC may function during acrosomal exocytosis (DeJonge et al., 1991; Breitbart et al., 1992; Rotem et al., 1992). However, the major problem with these studies is that they have relied on the use of PKC activators (e.g. phorbol esters) that may have other effects (Ahmed et al, 1993; Areces et al, 1994), and non-selective inhibitors such as H7 or staurosporine (see Wilkinson and Hallam, 1994, for review). Therefore, the results presented so far do not allow one to draw any firm conclusion on the likely role of PKC in intact spermatozoa. Moreover, no evidence of protein phosphorylation in response to either activators of PKC or natural agonists of acrosomal exocytosis has been presented. In somatic cells, a series of PKC isoforms is regulated by Ca 2+ and diacylglycerol (DAG) (Asaoka et al, 1992; Dekker and Parker, 1994). We have previously demonstrated that DAG is produced in human spermatozoa undergoing acrosomal exocytosis in response to progesterone treatment (O'Toole 921
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Protein kinase C activation during progesterone-stimulated acrosomal exocytosis in human spermatozoa

Molecular Human Reproduction vol.2 no.12 pp. 921-927, 1996
Protein kinase C activation during progesterone-stimulatedacrosomal exocytosis in human spermatozoa
Christine M.B.OToole1, Eduardo R.S.Roldan2 and Lynn R.Fraser1-3
Anatomy and Human Biology, King's College London, Strand, London WC2R 2LS, UK, and 2Departamento deReproducci6n Animal, Centro de Investigacidn y Tecnologla, INIA, Ctra de La Coruna Km 5.9, 28040 Madrid, Spain3To whom correspondence should be addressed
The involvement of protein kinase C (PKC) in exocytosis of the mammalian sperm acrosome is still acontroversial issue. Work carried out thus far has failed to provide direct evidence for the activation of thisenzyme upon stimulation with natural agonists of acrosomal exocytosis. We have therefore used progesteronestimulation of the acrosome reaction in human spermatozoa to clarify this issue. In spermatozoa preincubatedunder conditions known to support capacitation and fertilization in vitro, treatment with progesterone causeda time-dependent stimulation of phosphorylation of at least eight proteins ranging in size from -20-220 kDa.The inclusion of the PKC inhibitors chelerythrine chloride or calphostin C reduced the observed phosphorylationin a concentration-dependent manner. Exogenously supplied phorbol 12-myristate-13-acetate (PMA) or thepermeant diacylglycerol 1-oleoyl-2-acetyl-sn-glycerol (OAG), synthetic activators of PKC, also stimulatedphosphorylation in preincubated spermatozoa, but inclusion of calphostin C diminished the response.Furthermore, the prior inclusion of the 1,4-dihydropyridine Ca2+ channel antagonist nifedipine also inhibitedphosphorylation, suggesting that PKC is activated downstream of Ca2+ channel opening. Exocytosis triggeredby progesterone was significantly inhibited by chelerythrine chloride or calphostin C. Both PMA and OAGtriggered exocytosis, but the inclusion of chelerythrine chloride significantly inhibited the response; exocytoticresponses were seen only in capacitated cells. These results provide the first direct evidence that PKCactivation plays a role in the signal transduction pathway underlying acrosomal exocytosis in progesterone-stimulated capacitated spermatozoa.Key words: Ca2+ channels/diacylglycerol/fertilization/membrane fusion/phorbol ester
IntroductionThe mammalian sperm acrosome is analogous to a secretorygranule and the 'acrosome reaction', an exocytotic processwhereby acrosomal contents are released from the spermato-zoon during interaction with the oocyte, is a prerequisite forsuccessful fertilization (Yanagimachi, 1994). Stimulation ofspermatozoa to undergo exocytosis causes an increase inintracellular Ca2+ concentrations, due to influx from theextracellular space, and the activation of an array of signaltransduction pathways important for the sequence ending inmembrane fusion (F6nichel and Parinaud, 1995). One importantcomponent in this chain of events could be the activation ofprotein kinase C (PKC) because, in somatic cells, this enzymeappears to play a modulatory role during exocytotic responses(Kikkawa and Nishizuka, 1986; Cheek, 1993). However, anunequivocal role for PKC during the acrosome reaction hasyet to be demonstrated.
The presence of detectable PKC activity in spermatozoa hasbeen disputed (Kimura et al, 1984; Roldan and Harrison,1988; Rotem et al, 1990a,b; Breitbart et al, 1992). It nowseems clear that PKC activity in mammalian spermatozoa isextremely low, ~ 3 % of that seen in the rat brain enzyme, or-20% of that found in PC 12 cells or in rat pituitary cells(based on |Xg protein) (Rotem et al., 1990a,b; Breitbart et al.,
© European Society for Human Reproduction and Embryology
1992). Furthermore, when activity is compared taking intoaccount the number of cells used to obtain enzyme preparations,the activity of sperm PKC is only 0.4-1.3% of that in PC12or in pituitary cells (Rotem et al., 1990a).
Several studies employing either PKC antagonists or exogen-ous synthetic activators of PKC have provided indirect (i.e.circumstantial) evidence that PKC may function duringacrosomal exocytosis (DeJonge et al., 1991; Breitbart et al.,1992; Rotem et al., 1992). However, the major problem withthese studies is that they have relied on the use of PKCactivators (e.g. phorbol esters) that may have other effects(Ahmed et al, 1993; Areces et al, 1994), and non-selectiveinhibitors such as H7 or staurosporine (see Wilkinson andHallam, 1994, for review). Therefore, the results presented sofar do not allow one to draw any firm conclusion on the likelyrole of PKC in intact spermatozoa. Moreover, no evidence ofprotein phosphorylation in response to either activators ofPKC or natural agonists of acrosomal exocytosis has beenpresented.
In somatic cells, a series of PKC isoforms is regulated byCa2+ and diacylglycerol (DAG) (Asaoka et al, 1992; Dekkerand Parker, 1994). We have previously demonstrated that DAGis produced in human spermatozoa undergoing acrosomalexocytosis in response to progesterone treatment (O'Toole
921

C.M.B.OToole, E.R.S.Roldan and L.R.Fraser
et ai, 1996b), and that DAG production, in fact, depends ona prior activation of Ca2+channels (O'Toole et ai, 1996a). Theinclusion of nifedipine, a 1,4-dihydropyridine Ca2+ channelantagonist, either at time zero or just prior to addition ofprogesterone, significantly inhibited DAG production. Further-more, the same treatment inhibited acrosomal exocytosis asdetermined by chlortetracycline fluorescence (O'Toole et al,1996b).
Progesterone is known to be synthesized and secreted bythe cumulus cells surrounding the oocyte (Meizel, 1995).Acting via a non-genomic receptor (reviewed by Revelliet ai, 1994), it has been shown to stimulate specific signaltransduction pathways that result in the triggering of acrosomalexocytosis (Meizel, 1995). In the present study we demonstratethat stimulation of spermatozoa with progesterone results inphosphorylation of endogenous proteins, this response beingelicited also by synthetic activators of PKC and inhibited byspecific inhibitors of PKC. Acrosomal exocytosis could bestimulated both by progesterone and by various syntheticactivators of PKC, and inhibited by the same inhibitors ofPKC. Taken together, these results strongly indicate that PKCactivation plays an important role in the signal transductionpathway underlying acrosomal exocytosis in spermatozoa.
Materials and methods
Medium and reagentsThe standard medium used was Earle's medium (TCN Row, HighWycombe, Bucks, UK) with added benzylpenicillin (100 IU/ml,Sigma, Poole, Dorset, UK) and human serum albumin (HSA; Sigma)at 4 mg/ml; this medium contains 1.8 U.M CaCl2. Percoll was obtainedfrom Pharmacia LKB (Uppsala, Sweden), [32P]Pj (specific activity370 MBq/ml, ~10 mCi/ml) from Amersham International (Amersham,UK), and both chelerythrine chloride and calphostin C were obtainedfrom Calbiochem (Nottingham, UK). All other reagents were pur-chased from Sigma.
Preparation of sperm suspensionsSemen obtained by masturbation was provided by volunteer researchdonors at King's College London. The use of human semen for thisresearch has received ethical approval from the King's CollegeResearch Ethics Committee. Motile cells were prepared using mini-Percoll (Ord et ai, 1990) gradients (300 ul each of 95, 70 and 50%v/v solutions prepared from a substock containing 90 ml of 100%Percoll, 10 ml 10X Earle's concentrate, 0.37 ml sodium lactate, 2 mlHEPES buffer, 3 mg sodium pyruvate and HSA at 4 mg/ml). Aftercentrifugation at 600 g for 5 min all the supernatant was removedand the pelleted cells were resuspended in Earle's medium, centrifugedagain at 600 g for 5 min and resuspended in fresh medium. Thesperm concentration was estimated using a haemocytometer andadjusted to 5X 106 cells/ml. The percentage of motile cells in a samplewas estimated microscopically; usually >90% exhibited progressivemotility. All incubations were carried out in an atmosphere of 5% CO2/5% 02/90% N2. The medium and conditions used for experimentalincubations are similar to those frequently used in successful humanin-vitro fertilization (Stock et ai, 1989). In all experiments, spermsuspensions were preincubated for a total of 5 h before exocytosiswas stimulated with 10 u.g/ml progesterone (31.8 uM). A stocksolution of progesterone was prepared in dimethylsulphoxide(DMSO), diluted 10-fold in Earle's medium and then added at 1/100to suspensions (final concentration of DMSO was 0.1%).
Assessment of acrosomal exocytosisThe vital dye Hoechst bis-benzimide 33258 was used to assess cellvitality and the functional state of sperm was assessed using achlortetracycline (CTC) fluorescence assay (DasGupta et ai, 1993).Cells were assessed first for live/dead status using ultraviolet light.The Hg excitation beam was passed through a 334 nm band passfilter and fluorescence emission was observed through a DM 400dichroic mirror. The same cells were then assessed using violet light(the Hg excitation beam was passed through a 405 nm band passfilter and fluorescence emission observed through a DM 455 dichroicmirror) and classified as expressing one of the three main CTC stainingpatterns. These patterns are: F, with uniform bright fluorescence overthe entire head, which is characteristic of uncapacitated, acrosome-intact cells; B, with a fluorescence-free band in the post-acrosomalregion, which is characteristic of capacitated, acrosome-intact cells;AR, with dull or no fluorescence in the head region, which ischaracteristic of capacitated, acrosome-reacted cells. An earlier study(DasGupta et ai, 1993) ascertained that cells assessed as expressingthe AR pattern had undergone exocytosis.
The Percoll-prepared samples used had >90% motile cells and theproportion of dead (Hoechst-positive) cells was similar to that ofimmotile cells. As the numbers of dead cells were low and randomlydistributed among the assessment categories, all our CTC data arefor live (Hoechst-negative) cells only (200 assessed per replicate).
Labelling of intact cellsA modified method of Roldan and Harrison (1989) was used to labelintact sperm cells with [32P]P,. Sperm suspensions were preincubatedfor 3 h 45 min at 37°C, [32P]P, (carrier free, 10 mCi/ml) was addedto give a final concentration of 500 uCi/ml and the suspensions wereincubated in a water bath at 37°C for a further 1 h. Samples werethen centrifuged at 600 g for 5 min, the supernatant was discardedand the pellet was resuspended to its original volume in Earle'smedium. After stimulation ± inhibitors (see individual experiments)all suspensions were centrifuged at 2600-g for 5 min. The supernatantwas discarded and each pellet was resuspended in 50 ul Earle'smedium + 50 u.1 sodium dodecyl sulphate (SDS) loading buffer[containing 2.7% SDS, 13.5% glycerol, 0.14 uM dithiothreitol (DTT),0.1 uM Tris pH 6.8 and 0.03% Bromphenol Blue]; the samples wereboiled for 5 min and centrifuged at 10 500 g for 5 min to sedimentDNA before applying 15 ul (40 ug protein) to each lane forelectrophoresis.
Electrophoresis and autoradiography
SDS-polyacrylamide gel electrophoresis (PAGE) (Laemmli, 1970)was carried out using a Mighty Small II electrophoresis system(Hoefer Pharmacia, St Albans, Herts, UK). Proteins were separatedusing either a 7.5-12.5% linear gradient or a non-gradient 10%acrylamide resolving gel, with a 4% acrylamide stacking gel; [14C]methylated protein high molecular weight standards (AmershamInternational) were run on each gel and electrophoresis was carriedout at 20 mA per gel for ~2 h. After electrophoresis gels were stainedwith Coomassie R250. Gels were then dried, exposed to HRX film(Fujii) for up to 14 days at -70°C, and developed.
Statistical analysisCTC results were analysed using Cochran's modification of the X"test (Snedecor and Cochran, 1980). This is a stringent test thatcompares responses within replicates; significant differences indicatethat responses were both consistent and of a reasonable magnitudein all replicates.
922

kOa
220-
97-6 6 -
4 6 -
3 0 -
a b c d
Protein kinase C activation
k Da s a b e d
2 2 0 -
< 9 7 -
-* 6 6 -
4 6 -
2 2 -
1 4 -
30-
22E
Figure 1. Autoradiographic analysis of human sperm phosphoproteins following sodium dodecyl sulphate-polyacrylamide gelelectrophoresis (SDS-PAGE). Sperm suspensions were preincubated for 4 h in Earle's medium ± nifedipine (final concentration 100 nM),then labelled with [32P]Pi for 1 h. After centrifugation the cells were resuspended in fresh medium ± nifedipine and then exposed to31.8 uM progesterone for 20 min. Sperm proteins were solubilized in SDS loading buffer and separated using either (A) a 10% non-gradient or (B) a 7.5-12.5% linear gradient resolving gel with a 4% stacking gel. The bands were visualized after up to 14 days exposure ofthe gel to a Fuji HRX X-ray film at —70°C. Lanes show: a, control cells (not exposed to progesterone); b, progesterone-only treated cells;c, nifedipine-only treated cells and d, cells treated with nifedipine and progesterone. In this figure, the molecular weight standards are shownin the far left lane (S). In this and Figures 2 and 3, the phosphoproteins consistently seen are indicated by arrowheads.
Results
Phosphorylation patterns in progesterone-stimulatedsuspensions
Sperm suspensions preincubated under capacitating conditionswere labelled with [32P]Pi, washed and then exposed to 10 ugprogesterone/ml. Phosphorylation patterns were evaluated after10, 20 and 30 min. We observed a time-dependent stimulationin phosphorylation of an array of proteins, with the maximalresponse at 20 min (Figure 1). A total of at least eightphosphoproteins, of -20, 24, 29, 38, 46, 64, 98, and 220 kDa,could be detected. In order to evaluate low and high molecularweight proteins adequately, we used both non-gradient 10%(Figure 1A) and 7.5-12.5% linear gradient acrylamide gels(Figure IB). The 10% gels have the advantage of showingthe full range of phosphoproteins in terms of size, but thedisadvantage of relatively poor resolution of the high molecularweight phosphoproteins. As these are the most heavily-labelledproteins, we have included Figure IB to demonstrate clearlythe presence of the ~98 and 220 kDa phosphoproteins. Underthe conditions used, the smaller and relatively less heavily-labelled proteins are not visible.
To investigate the possible involvement of Ca2+ influx viaCa2+ channels in the activation of PKC, we included nifedipine,a 1,4-dihydropyridine Ca2+ channel antagonist, at time 0.Nifedipine inhibited the phosphorylation observed at 5 h inresponse to progesterone. Little change over the 30 minassessment period was detected in either the control suspen-sions (no added progesterone) or those receiving nifedipineonly, but the maximal response to progesterone was observedat 20 min (Figure 1). In subsequent experiments, suspensionswere incubated for 20 min following addition of progesterone
or PKC activators and then evaluated for phosphorylation.Coomassie-stained gels indicated that equal amounts of proteinwere added to each lane; proteins of sizes similar to several(~29, 38, 46, 220 kDa) but not all of the phosphoproteinscould be identified (not shown).
In a second series we examined the effects of two PKCinhibitors, chelerythrine chloride (which interacts with thecatalytic domain; Ko et ai, 1990) and calphostin C (whichinteracts with the regulatory domain; Kobayashi et at., 1989),on progesterone-induced stimulation of phosphorylation.Again, progesterone stimulated phosphorylation of an array ofproteins (Figure 2), but the inclusion of either chelerythrinechloride (0.6 or 1.8 jiM) or calphostin C (0.15 iM) 15 minbefore the addition of progesterone inhibited the response; allphosphoproteins appeared to be equally affected. Thus PKCactivity would appear to be responsible for the observedphosphorylation.
Phosphorylation patterns in PMA- or OAG-stimulatedspermatozoa
Since PKC activity in a wide range of cells is stimulated bypermeant DAG and phorbol esters, we then investigated theeffects of l-oleoyl-2-acetyl-j/i-glycerol (OAG), phorbol-12-myristate-13-acetate (PMA), and the inactive phorbol ester(Neidel and Blackshear, 1986) 4a-phorbol-12, 13-didecanoate(4a-PDD) (Figure 3). In spermatozoa preincubated and labelledas indicated above, exposure to 1 (iM PMA or 10 uM OAGstimulated phosphorylation, but 1 \JM 4a-PDD had little effect.The inclusion of calphostin C (0.3 jlM) inhibited the responseto PMA, again indicating that the phosphorylation eventsobserved involved PKC activity.
923
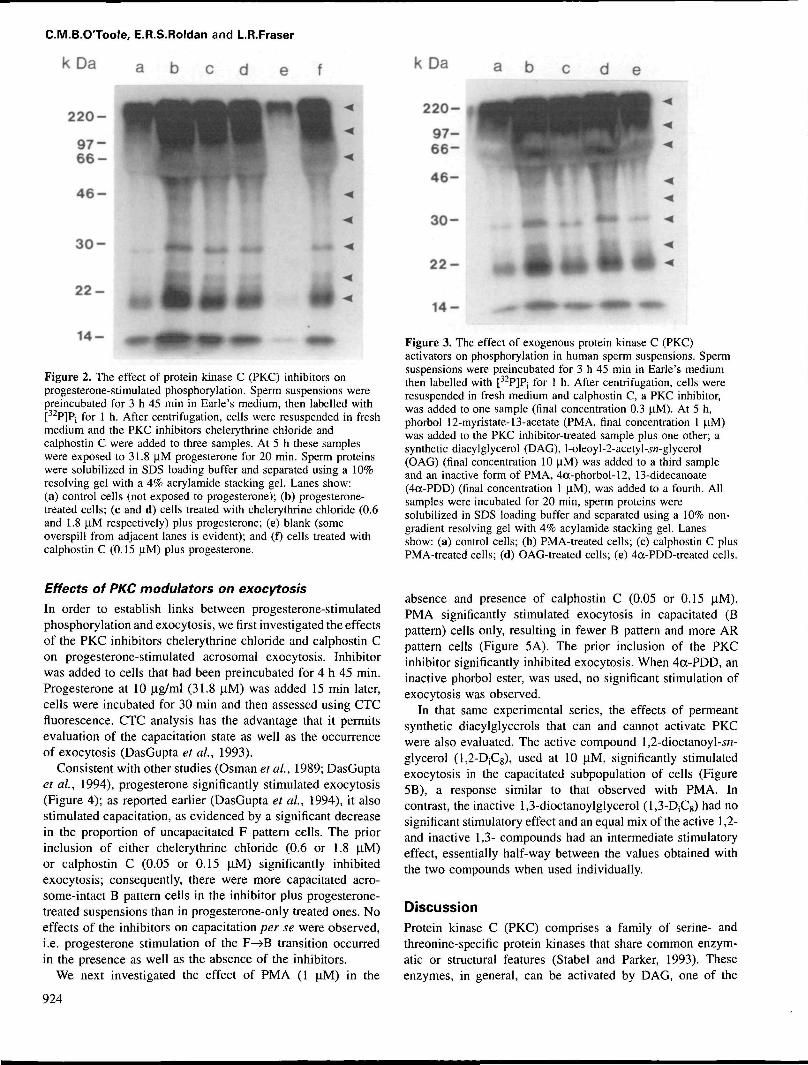
C.M.B.OToole, E.R.S.Roldan and L.R.Fraser
k D a a b c d e f
220-
4 6 -
3 0 -
2 2 -
14 -
Figure 2. The effect of protein kinase C (PKC) inhibitors onprogesterone-stimulated phosphorylation. Sperm suspensions werepreincubated for 3 h 45 min in Earle's medium, then labelled with[32P]Pj for 1 h. After centrifugation, cells were resuspended in freshmedium and the PKC inhibitors chelerythrine chloride andcalphostin C were added to three samples. At 5 h these sampleswere exposed to 31.8 uM progesterone for 20 min. Sperm proteinswere solubilized in SDS loading buffer and separated using a 10%resolving gel with a 4% acrylamide stacking gel. Lanes show:(a) control cells (not exposed to progesterone); (b) progesterone-treated cells; (c and d) cells treated with chelerythrine chloride (0.6and 1.8 uM respectively) plus progesterone; (e) blank (someoverspill from adjacent lanes is evident); and (f) cells treated withcalphostin C (0.15 u,M) plus progesterone.
Effects of PKC modulators on exocytosis
In order to establish links between progesterone-stimulatedphosphorylation and exocytosis, we first investigated the effectsof the PKC inhibitors chelerythrine chloride and calphostin Con progesterone-stimulated acrosomal exocytosis. Inhibitorwas added to cells that had been preincubated for 4 h 45 min.Progesterone at 10 (ig/ml (31.8 u,M) was added 15 min later,cells were incubated for 30 min and then assessed using CTCfluorescence. CTC analysis has the advantage that it permitsevaluation of the capacitation state as well as the occurrenceof exocytosis (DasGupta et al., 1993).
Consistent with other studies (Osman et al., 1989; DasGuptaet al., 1994), progesterone significantly stimulated exocytosis(Figure 4); as reported earlier (DasGupta et al., 1994), it alsostimulated capacitation, as evidenced by a significant decreasein the proportion of uncapacitated F pattern cells. The priorinclusion of either chelerythrine chloride (0.6 or 1.8 u,M)or calphostin C (0.05 or 0.15 U,M) significantly inhibitedexocytosis; consequently, there were more capacitated acro-some-intact B pattern cells in the inhibitor plus progesterone-treated suspensions than in progesterone-only treated ones. Noeffects of the inhibitors on capacitation per se were observed,i.e. progesterone stimulation of the F—>B transition occurredin the presence as well as the absence of the inhibitors.
We next investigated the effect of PMA (1 U.M) in the
924
kDa
220-
a b
4 6 -
3 0 -
2 2 -
1 4 - —•
Figure 3. The effect of exogenous protein kinase C (PKC)activators on phosphorylation in human sperm suspensions. Spermsuspensions were preincubated for 3 h 45 min in Earle's mediumthen labelled with [32P]P, for 1 h. After centrifugation, cells wereresuspended in fresh medium and calphostin C, a PKC inhibitor,was added to one sample (final concentration 0.3 U.M). At 5 h,phorbol 12-myristate-13-acetate (PMA, final concentration 1 uM)was added to the PKC inhibitor-treated sample plus one other; asynthetic diacylglycerol (DAG), l-oleoyl-2-acetyl-.yrc-glycerol(OAG) (final concentration 10 uM) was added to a third sampleand an inactive form of PMA, 4a-phorbol-12, 13-didecanoate(4oc-PDD) (final concentration 1 uM), was added to a fourth. Allsamples were incubated for 20 min, sperm proteins weresolubilized in SDS loading buffer and separated using a 10% non-gradient resolving gel with 4% acylamide stacking gel. Lanesshow: (a) control cells; (b) PMA-treated cells; (c) calphostin C plusPMA-treated cells; (d) OAG-treated cells; (e) 4<x-PDD-treated cells.
absence and presence of calphostin C (0.05 or 0.15 \iM).PMA significantly stimulated exocytosis in capacitated (Bpattern) cells only, resulting in fewer B pattern and more ARpattern cells (Figure 5A). The prior inclusion of the PKCinhibitor significantly inhibited exocytosis. When 4a-PDD, aninactive phorbol ester, was used, no significant stimulation ofexocytosis was observed.
In that same experimental series, the effects of permeantsynthetic diacylglycerols that can and cannot activate PKCwere also evaluated. The active compound 1,2-dioctanoyl-in-glycerol (1,2-DjCg), used at 10 (iM, significantly stimulatedexocytosis in the capacitated subpopulation of cells (Figure5B), a response similar to that observed with PMA. Incontrast, the inactive 1,3-dioctanoylglycerol (l^-DjCg) had nosignificant stimulatory effect and an equal mix of the active 1,2-and inactive 1,3- compounds had an intermediate stimulatoryeffect, essentially half-way between the values obtained withthe two compounds when used individually.
Discussion
Protein kinase C (PKC) comprises a family of serine- andthreonine-specific protein kinases that share common enzym-atic or structural features (Stabel and Parker, 1993). Theseenzymes, in general, can be activated by DAG, one of the

Protein kinase C activation
Figure 4. Inhibition of progesterone-induced acrosomal exocytosisby protein kinase C (PKC) inhibitors. Sperm suspensions werepreincubated for 4 h 45 min in Earle's medium; PKC inhibitorschelerythrine chloride and calphostin C were then added to foursamples. At 5 h these samples plus an untreated one were exposedto 31.8 U.M progesterone for 30 min. The sperm suspensions werethen stained with chlortetracycline (CTC) and examined byfluorescence microscopy. Results are mean ± SE (n = 4).Histograms show: (a) untreated control cells; (b) progesterone-treated cells; (c and d), cells treated with chelerythrine chloride (0.6and 1.8 |iM respectively) plus progesterone; (e and f) cells treatedwith calphostin C (0.05 and 0.15 U.M respectively) plusprogesterone. • = F-uncapacitated acrosome-intact;• =B-capacitated acrosome-intact; • = AR-capacitated acrosome-reacted.**P <0.01, ***P <0.001 compared with untreated controls; + P <0.05,+ +P <0.01 compared with progesterone-only treated cells.
products of phosphatidylinositol turnover. This suggests a rolefor PKC in the signal transduction pathway involving activationof polyphosphoinositide-specific phosphoinositidase C (PIC)and subsequent hydrolysis of polyphosphoinositides to generateinositol 1,4,5-trisphosphate and DAG. Tumour-promotingphorbol esters can also activate PKC, suggesting that theenzyme may be important in both normal and abnormal cellmetabolism (Nishizuka, 1984; Martelly and Castagna, 1989).
Activation of PIC and PKC has been demonstrated in awide variety of somatic cell systems where Ca2+-dependentexocytosis occurs (Kikkawa and Nishizuka, 1986; Cheek,1993). The mammalian sperm acrosome reaction is a Ca2+-dependent exocytotic event and activation of PIC is known tobe an early response to Ca2+ entry from the extracellularcompartment (Roldan and Harrison, 1989). Immunohisto-chemical analyses of human spermatozoa using both light(Rotem et al, 1990a,b; 1992) and electron microscopy (Kalinaet al., 1995) indicate the presence of at least some isoformsof PKC in both the head and tail. However, a role for PKC inthe exocytotic pathway has remained equivocal, with availableevidence being indirect in nature. Phorbol esters and permeantsynthetic DAGs have been reported capable of initiatingacrosomal exocytosis in capacitated human sperm suspensions(DeJonge et al., 1991; Rotem et al., 1992), but such treatmentseffectively circumvent the signal transduction pathways thatappear to function during fertilization (Yanagimachi, 1994) andhence may not mimic normal endogenous events. Moreover, itis now recognized that both phorbol esters and DAGs have
other targets and, therefore, may be acting via mechanismsnot involving PKC (Wilkinson and Hallam, 1994).
As stated earlier, human spermatozoa stimulated to undergoacrosomal exocytosis are known to generate DAG, with bothPIC and phospholipase C contributing to the overall pool ofDAG detected (Roldan and Harrison, 1989; O'Toole et al.,1996b); no evidence for phospholipase D activity in humanspermatozoa has been obtained (O'Toole et al., 1996b). There-fore, it is possible that this endogenously-generated DAGmight activate PKC; the experimental evidence obtained inthis study supports a role for PKC in acrosomal exocytosis.
Direct evidence for PKC activation in response to a naturalagonist of exocytosis was obtained by investigating phos-phorylation of endogenous proteins in response to progesterone.This agonist interacts with the sperm plasma membrane andactivates a receptor-mediated signal transduction pathwayleading to membrane fusion (Meizel et al., 1990; Meizel,1995; Roldan, 1995). The human sperm response to pro-gesterone resulted in a time-dependent increase in proteinphosphorylation. At least eight phosphoproteins, ranging insize from ~20 to 220 kDa, could be detected in stimulatedcells; proteins of a size similar to four of these phosphoproteins(-29, 38, 46 and 220 kDa) could be seen on Coomassie Blue-stained gels (data not shown), suggesting that some of theprotein substrates may be reasonably abundant. Several resultssuggest that phosphorylation of these proteins in response toprogesterone is due to PKC: (i) a permeant synthetic DAG(OAG) and a tumour-promoting phorbol ester (PMA), knownto activate PKC, stimulated phosphorylation of the sameproteins; and (ii) a phorbol ester which is incapable ofactivating PKC did not stimulate phosphorylation; (iii) bothprogesterone- and PMA-stimulated phosphorylation was inhib-ited by inclusion of two different and highly specific PKCinhibitors.
To explore the role of Ca2+ in the activation of PKC weexamined the pattern of protein phosphorylation in spermatozoaexposed to nifedipine, a dihydropyridine Ca2+ channelantagonist, before stimulation with progesterone. Inclusion ofnifedipine inhibited the phosphorylation response elicited byprogesterone, indicating that an influx of Ca2+ from theextracellular compartment is required for the activation ofPKC. However, the fact that treatment with nifedipine alsoblocks DAG formation and acrosomal exocytosis (O'Tooleet al., 1996a) raises the question of which mechanism isactually being inhibited. It is possible that PKC activation,and hence phosphorylation, does not take place due to theabsence of a rise in DAG, the endogenous activator of PKC(Rando, 1988). If this is the case, then exogenously addedDAG should cause phosphorylation and this proved to bethe case: a permeant synthetic DAG (OAG) stimulated thisresponse, as did PMA. These results then raise the questionof whether an elevation in intracellular Ca2+ is required incells stimulated only with OAG or phorbol esters. Currentevidence suggests that changes in sperm intracellular Ca2+ aredue primarily to influx, rather than to mobilization frominternal stores (Meizel, 1995). It could be argued that activationof PKC may not require an elevation of intracellular Ca2+
(DAG would lower the PKC requirement for Ca2+; Kaibuchi
925

C.M.B.OToole, E.R.S.Roldan and L.R.Fraser
80-
a b c d e
Figure 5. Stimulation of acrosomal exocytosis by a phorbol ester and a synthetic diacylglycerol and inhibition by inclusion of a proteinkinase C (PKC) inhibitor. (A) Sperm suspensions were preincubated for 4 h 45 min in Earle's medium and the PKC inhibitor caJphostin Cwas then added to two samples. At 5 h, phorbol 12-myristate-13-acetate (PMA) (final concentration 1 uM) was added to the inhibitor-treated samples plus one other; a fourth sample was exposed to an inactive form of PMA, 4a-phorbol-12, 13-didecanoate (4a-PDD) (finalconcentration 1 |iM). All samples were incubated for 30 min, then stained with chlortetracycline (CTC) and assessed. Results are mean ±SE {n = 8). Histograms show: (a) untreated control cells; (b) PMA-treated cells; (c and d) cells treated with calphostin C (0.05 and0.15 pM respectively) plus PMA; (e) 4a-PDD-treated cells. • = F-uncapacitated acrosome-intact; • = -capacitated acrosome-intact;• = AR-capacitated acrosome-reacted. *P <0.05, ***P <0.001 compared with untreated controls; + +P <0.01 compared with PMA-treated cells. (B) Sperm suspensions preincubated for 5 h in Earle's medium were incubated in the presence of: (a) no added compounds(control cells); (b) a synthetic active form of diacylglycerol (DAG), 1,2-dioctanoyl-^-glycerol (final concentration 10 nM); (c) a 1:1 mix of1,2-dioctanoyl-w-glycerol and 1,3-dioctanolyglycerol (final concentration 10 nM); (d) an inactive DAG, 1,3-dioctanolyglycerol (finalconcentration 10 uM) for 30 min. All samples were then stained with CTC and assessed. Results are mean ± SE (n = 8). *P <0.05,**P <0.01, ***P <0.001 compared with untreated controls.
et al, 1981). Since either OAG or phorbol esters alone canstimulate capacitated spermatozoa to complete the sequenceleading to membrane fusion (DeJonge et al, 1991; Rotemetal, 1992; this study), a process which requires an intracellularCa2+ rise (Roldan and Harrison, 1989; Roldan and Fragio,1993), some mechanism presumably provides the requiredCa2+. One possible explanation is that the rise in intracellularCa2+ occurring during capacitation (Florman, 1994; Suarezand Dai, 1995) provides sufficient Ca2+ to support eventsoccurring downstream of PKC activation. For example, mousespermatozoa preincubated for 30 min (insufficient time forcomplete capacitation) in reduced Ca2+-containing medium(0.09 (lM; sufficient to support capacitation but not spontaneousacrosomal exocytosis) did not undergo exocytosis in responseto A23187 (Fraser and McDermott, 1992), but this sametreatment applied to 120 min preincubated capacitated cellstriggered a significant exocytotic response (Forsdike and Fraser,1996). This altered response suggests a capacitation-dependentrise in intracellular Ca2+, resulting in cells able to respondto ionophore despite the low extracellular Ca2+. Thereforecapacitated cells might 6e equally responsive to activatorsof PKC.
Whether PKC itself might control additional changes inintracellular Ca2+ is unclear at present. Foresta et al. (1995)reported that inclusion of two non-specific inhibitors of PKC(H-7 and staurosporine) inhibited the progesterone-stimulatedCa2+ rise in human spermatozoa, whereas Bonaccorsi et al.(1995) found that a different inhibitor, sangivamycin, did not.Further research is required to address this specific point.
A clear link between PKC-mediated phosphorylation of a
series of sperm proteins and exocytosis was evident. Conditionsthat stimulated phosphorylation (e.g. treatment with progester-one or with either PMA or exogenous DAG, both well knownactivators of PKC) also resulted in exocytosis in capacitatedcells, whereas the use of reagents incapable of activating PKCcaused neither. Also, inhibition of progesterone and phorbolester-induced phosphorylation was paralleled by inhibition ofexocytosis. These results are consistent with an essential rolefor PKC in the exocytotic pathway.
As noted in an earlier study, progesterone treatment ofpreincubated suspensions not only triggered exocytosis incapacitated cells but also stimulated capacitation, as determinedby the significant decrease in the proportion of F pattern(uncapacitated) cells, even after a short (30 min) exposure toprogesterone. Interestingly, the presence of either PKC inhibitorhad no inhibitory effect on this response to progesterone,suggesting that PKC may not play a role in capacitationitself but only during exocytosis. Furthermore, treatment ofpreincubated suspensions with PMA or OAG only caused atransition from the B (capacitated) to the AR pattern, but notfrom the F (uncapacitated) to the B pattern; consistent withthis, an earlier study demonstrated that uncapacitated suspen-sions were unable to undergo exocytosis in response to OAG(O'Toole et al, 1996b). The present data indicate that PKCcan only be activated in capacitated cells, suggesting that thereare capacitation-related changes in the availability of PKC foractivation, e.g. alterations in PKC association with the spermplasma membrane.
In conclusion, the evidence gathered in this study demon-strates a fundamental role for PKC activation, with its con-
926

Protein kinase C activation
sequent phosphorylation of endogenous proteins, duringprogesterone-induced acrosomal exocytosis in human sperma-tozoa and therefore during fertilization.
AcknowledgementsThis work was supported by The Wellcome Trust.
ReferencesAhmed, S., Lee, J., Kozma, R. et al. (1993) A novel functional target for
tumour-promoting phorbol esters and lysophosphatidic acid: the p21 rac-GTPase activaung protein N-chimaerin. J. Bwl. Chem., 268, 10709-10712.
Arcces, L.B., Kazanietz, M.G. and Blumberg, P.M. (1994) Close similarity ofbaculovirus-expressed N-chimaerin and protein kinase C-a as phorbol esterreceptors. /. Biol. Chem., 269, 19553-19558.
Asaoka, Y., Nakamura, S., Yoshida, K. and Nishizuka, Y. (1992) Proteinkinase C, calcium and phospholipid degradation. Trends Biochem. 5c/., 17,414-417.
Bonaccorsi, L., Luconi, M., Forti, G. and Baldi, E. (1995) Tyrosine kinaseinhibition reduces the plateau phase of the calcium increase in response toprogesterone in human sperm. FEBS Leu., 364, 83-86.
Breitbart, H., Lax, J., Rotem, R. and Naor, Z. (1992) Role of protein kinaseC in the acrosome reaction of mammalian spermatozoa. Biochem. J., 281,473-476.
Cheek, T.R. (1993) Calcium signalling and the triggering of secretion inadrenal chromaffin cells. In Taylor, C.W. (ed.), Intracellular Messengers.Pergamon Press, Oxford, pp. 427^46.
DasGupta, S., Mills, C.L. and Fraser, L.R. (1993) Ca2+-related changes inthe capacitation state of human spermatozoa assessed by a chlortetracyclinefluorescence assay. /. Reprod. Fertil., 99, 135-143.
DasGupta, S., O'Toole, C, Mills, C.L. and Fraser, L.R. (1994) Effect ofpentoxifylline and progesterone on human sperm capacitation and acrosomalexocytosis. Hum. Reprod., 9, 2103-2109.
De Jonge, C.J , Han, H-L., Mack, S.R. and Zaneveld, LJ.D. (1991) Effect ofphorbol diesters, synthetic diacylglycerols and a protein kinase C inhibitoron the human sperm acrosome reaction. J. Androl., 12, 62-70.
Dekker, L.V. and Parker, PJ. (1994) Protein kinase C-a question of specificity.Trends Biochem. Sci., 19, 73-77.
Finichel, P. and Pannaud, J. (eds) (1995) Human Sperm Acrosome Reaction.John Libbey Eurotext, Montrouge.
Florman, H.M. (1994) Sequential focal and global elevations of spermintracellular Ca2+ are initiated by the zona pellucida during acrosomalexocytosis. Dev. Biol., 165, 152-164.
Foresta, C , Rossato, M. and DiVirgilio, F. (1995) Differential modulation byprotein kinase C of progesterone activated responses in human sperm.Biochem. Biophys. Res. Commn., 206, 408-413.
Forsdike, R.A. and Fraser, L.R. (1996) Relative importance of Ca2+ andHCO3" for capacitation and acrosomal exocytosis in mouse spermatozoa.J. Reprod. Fertil. (Abslr. Ser), 17, 4.
Fraser, L.R. and McDermott, C.A. (1992) Ca2+-related changes in the mousesperm capacitation state: a possible role for Ca2+-ATPase. J. Reprod.Fertil., 96, 363-377.
Kaibuchi, K., Takai, Y. and Nishizuka, Y. (1981) Cooperative roles ofvarious membrane phospholipids in the activation of calcium-activated,phospholipid-dependent protein kinase. J. Biol. Chem., 256, 7146-7149.
Kalina, M., Socher, R., Rotem R. and Naor, Z. (1995) Ultrastructurallocalization of protein kinase C in human sperm. J. Histochem. Cylochem.,43, 439-445.
Kikkawa, U. and Nishizuka, Y. (1986) The role of protein kinase C intransmembrane signalling. Ann. Rev. Cell Biol., 2, 149—178.
Kimura, K., Katoh, N., Sakurada, K. and Kubo, S. (1984) Phospholipid-sensitive Ca2+-dependent protein kinase system in testis-localization andendogenous substrates. Endocrinology, 115, 2391-2399.
Ko, F.N., Chen, I.S., Wu, SJ. et aL (1990) Antiplatelet effects of chelerythrinechloride isolated from Zanthoxylum simulans. Biochim. Biophys. Acta,1052, 360-365.
Kobayashi, E., Nakano, H., Morimoto, M. and Tamaoki, T. (1989) CalphostinC (UCN-1028C), a novel microbial compound, is a highly potent andspecific inhibitor of protein kinase C. Biochem. Biophys. Res. Commn.,159, 548-553.
Laemmli, U.K. (1970) Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature, 227, 680-685.
Martelly, I. and Castagna, M. (1989) Protein kinase C and tumour promotors.Curr. Opinion Cell Biol., 1, 206-210.
Merzel, S. (1995) Initiation of human sperm acrosome reaction by progesterone.In Finichel, P. and Parinaud, J. (eds), Human Sperm Acrosome Reaction.John Libbey Eurotext, Montrouge, pp. 151-164.
Meizel, S., Pillai, M.C., Dfaz-P6rez, E. and Thomas, P. (1990) Initiation ofthe human sperm acrosome reaction by components of human follicularfluid and cumulus secretions including steroids. In Bavister, B.D., Cummins,J. and Roldan, E.R.S. (eds), Fertilization in Mammals. Serono SymposiaUSA, Norwell, MA, pp. 205-222.
Neidel, J.E. and Blackshear, PJ. (1986) Protein kinase C In Putney, J.W.(ed.), Phosphoinositides and Receptor Mechanisms. Alan R.Liss Inc., NewYork, pp. 47-88.
Nishizuka, Y. (1984) The role of protein kinase C in cell surface signaltransduction and rumour promotion. Nature, 308, 693-698.
O'Toole, C.M.B., Roldan, E.R.S. and Fraser, L.R. (1996a) Role for Ca2+
channels in the signal transduction pathway leading to acrosomal exocytosisin human spermatozoa. Mol. Reprod. Dev., 45, 204—211.
OToole, C.M.B., Roldan, E.R.S., Hampton, P. and Fraser, L.R. (1996b) Arole for diacylglycerol in human sperm acrosomal exocytosis. Mol. Hum.Reprod., 2, 317-326.
Ord, T, Patrizio, P., Marello, E. et al. (1990) Mini Percoll: a new method ofsemen preparation for FVF in severe male factor infertility. Hum. Reprod.,5, 987-989.
Osman, R.A., Andria, M.L., Jones, A.D. and Meizel, S. (1989) Steroid inducedexocytosis: the human sperm acrosome reaction. Biochem. Biophys. Res.Commun,, 160, 828-833.
Rando, R.R. (1988) Regulation of protein kinase C activity by lipids. FASEBJ., 2, 2348-2355.
Revelli, A., Modotti, M., Piffaretti-Yanez, A. et al. (1994) Steroid receptorsin human spermatozoa. Hum. Reprod., 9, 760-766.
Roldan, E.R.S. (1995) Role of phosphoinositides in the mammalian spermacrosome reaction. In F^nichel, P. and Parinaud, J. (eds), Human SpermAcrosome Reaction. John Libbey Eurotext, Montrouge, pp. 225-243.
Roldan, E.R.S. and Harrison, R.A.P. (1988) Absence of active protein kinaseC in ram spermatozoa. Biochem. Biophys. Res. Commun., 155,901-906.
Roldan, E.R.S. and Harrison, R.A.P. (1989) Polyphosphoinositide breakdownand subsequent exocytosis in the Ca2+ ionophore induced acrosome reactionof mammalian spermatozoa. Biochem. J., 259, 397-406.
Roldan, E.R.S. and Fragio, C. (1993) Phospholipase A2 activity and exocytosisof the ram sperm acrosome: regulation by bivalent cations. Biochem.Biophys. Acta, 1210, 48-54.
Rotem, R., Paz, G.F., Homonnai, Z.T. et al. (1990a) Further studies on theinvolvement of protein kinase C in human flagellar motility. Endocrinology,127,2571-2577.
Rotem, R., Paz, G.F., Homonnai, Z.T. et al. (1990b) Protein kinase C ispresent in human sperm: possible role in flagellar motility. Proc. Natl.Acad. Sci. USA, 87, 7305-7308.
Rotem, R., Paz, G.F., Homonnai, Z.T. etal (1992) Ca2+-independent inductionof acrosome reaction by protein kinase C in human sperm. Endocrinology,131, 2235-2243.
Snedecor, G.W. and Cochran, W.G. (1980) Statistical Methods, 7th edn. IowaState University Press, Ames, USA.
Stabel, S. and Parker, PJ. (1993) Protein kinase C. In Taylor, C. W. (ed.),Intracellular Messengers. Pergamon Press, Oxford, pp. 167-198.
Stock, C.E., Bates, R., Lindsay, K.S., et al. (1989) Human oocyte-cumuluscomplexes stimulate die human acrosome reaction. J. Reprod. Fertil., 86,723-730.
Suarez, S.S. and Dai, X. (1995) Intracellular calcium reaches different levelsof elevation in hyperactivated and acrosome-reacted hamster sperm. MotReprod. Dev., 42, 325-333.
Wilkinson, S.E. and Hallam, TJ. (1994) Protein kinase C-is its pivotal rolein cellular activation overstated? Trends Pharmacol. Sci., 15, 53-57.
Yanagimachi, R. (1994) Mammalian fertilization. In Knobil, E. and Neill,J.D. (eds.), The Physiology of Reproduction. Vol. 1, 2nd edn. Raven Press,New York, pp. 189-317.
Received on August 2, 1996; accepted on October 18, 1996
927




















