
Methylation defects of imprinted genes in human testicular spermatozoa
10
Methylation defects of imprinted genes in human testicular spermatozoa C. Joana Marques, Ph.D., a T ^ ania Francisco, M.S., a S onia Sousa, M.S., b Filipa Carvalho, Ph.D., a Alberto Barros, M.D., Ph.D., a,b and M ario Sousa, M.D., Ph.D. a,b,c a Department of Genetics, Faculty of Medicine, University of Porto; b Centre for Reproductive Genetics Alberto Barros, Porto; and c Lab Cell Biology, ICBAS–Institute of Biomedical Sciences Abel Salazar, University of Porto, Porto, Portugal Objective: To study the methylation imprinting marks of two oppositely imprinted genes, H19 and MEST/PEG1, in human testicular spermatozoa from azoospermic patients with different etiologies. Testicular spermatozoa are often used in intracytoplasmic sperm injection for treatment of male factor infertility, but the imprinting status of these cells is currently unknown. Design: Experimental prospective study. Setting: University research laboratory and private in vitro fertilization (IVF) clinic. Patient(s): A total of 24 men, five with anejaculation, five with secondary obstructive azoospermia, five with pri- mary obstructive azoospermia, and nine with secretory azoospermia due to hypospermatogenesis. Intervention(s): Spermatozoa were isolated by micromanipulation from testicular biopsies. Main Outcome Measure(s): DNA methylation patterns were analyzed using bisulfite genomic sequencing with cloning analysis. Result(s): We found H19 complete methylation was statistically significantly reduced in secretory azoospermic patients with hypospermatogenesis, with one patient presenting complete unmethylation. Hypomethylation also affected the CTCF-binding site 6, involved in regulation of IGF2 expression. Regarding the MEST gene, all pa- tients presented complete unmethylation although this was statistically significantly reduced in the anejaculation group. Conclusion(s): Testicular spermatozoa from men with abnormal spermatogenesis carry methylation defects in the H19 imprinted gene which also affect the CTCF-binding site, further supporting an association between the occur- rence of imprinting errors and disruptive spermatogenesis. (Fertil Steril Ò 2010;94:585–94. Ó2010 by American Society for Reproductive Medicine.) Key Words: Azoospermia, DNA methylation, genomic imprinting, H19, MEST , male factor infertility, testicular spermatozoa Genomic imprinting leads to monoallelic parental-dependent expression of a subset of genes in mammals. Parental alleles are marked differentially, primarily by DNA methylation of CpG dinucleotides located on the differentially methylated re- gions of imprinted genes. These regulate the expression of im- printed genes and constitute the imprinting control regions (ICRs) (1). Methylation imprinting marks are erased in primordial germ cells (PGCs) (2) and are later reestablished during germ cell development according to parental sex. In murine spermatogenesis, paternal imprints start to be ac- quired prenatally and are definitively established before the onset of meiosis (3, 4). It is interesting that paternal imprints seem to be acquired earlier in the paternally inherited allele than in the allele of maternal origin, suggesting that, even when devoid of methylation, parental alleles can retain their identity (3). In humans, establishment of paternal imprints has been shown to be complete by the primary spermatocyte stage (5). On fertilization, a wave of DNA demethylation oc- curs, active in the paternal pronucleus and passive in the ma- ternal pronucleus, which does not affect the methylation imprinting marks previously set in gametes (6). There are only three known imprinting control regions that acquire methylation in the paternal germ line: H19-Igf2, Dlk1-Gtl2, and Rasgrf1 (7). H19 is one of the best character- ized imprinted genes; it is expressed from the maternal allele, it is imprinted in human (8) and mice (9), and it encodes an untranslated RNA (10) whose function remains unclear. It was previously suggested to have tumor-suppressor activity (11) but also to be an oncofetal RNA (12), and was recently demonstrated to give rise to a microRNA (miR-675) (13). Additionally, H19 is physically and functionally linked to an- other imprinted gene, IGF2 (insulin-like growth factor-2) through the sharing of common enhancers (14, 15). H19 and IGF2 are oppositely imprinted because H19 is expressed Received November 28, 2008; revised February 1, 2009; accepted February 16, 2009; published online April 1, 2010. C.J.M. has nothing to disclose. T.F. has nothing to disclose. S.S. has nothing to disclose. F.C. has nothing to disclose. A.B. has nothing to disclose. M.S. has nothing to disclose. Partially supported by the Portuguese Foundation for Science and Tech- nology (FCT) with a PhD fellowship (SFRH/BD/19967/2004 to CJM), a Young Researcher fellowship (POCI/SAU-MMO/60555/04 to T.F.) and research grants (POCI/SAU-MMO/60555/04, 60709/04, 59997/04; UMIB). Reprint requests: M ario Sousa, M.D., Ph.D., Lab Cell Biology, Institute of Biomedical Sciences Abel Salazar (ICBAS), University of Porto, Largo Prof Abel Salazar 2, 4099-003 Porto, Portugal (FAX: 00-351-22-206-22-32; E-mail: [email protected]). 0015-0282/$36.00 Fertility and Sterility â Vol. 94, No. 2, July 2010 585 doi:10.1016/j.fertnstert.2009.02.051 Copyright ª2010 American Society for Reproductive Medicine, Published by Elsevier Inc.
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Methylation defects of imprinted genes in human testicular spermatozoa

Received
Februar
C.J.M. ha
nothing
disclose
Partially su
nology
a Youn
and res
UMIB).
Reprint re
of Biom
Largo
00-351-
0015-028doi:10.10
Methylation defects of imprinted genes in humantesticular spermatozoaC. Joana Marques, Ph.D.,a Tania Francisco, M.S.,a S�onia Sousa, M.S.,b Filipa Carvalho, Ph.D.,a
Alberto Barros, M.D., Ph.D.,a,b and M�ario Sousa, M.D., Ph.D.a,b,c
a Department of Genetics, Faculty of Medicine, University of Porto; b Centre for Reproductive Genetics Alberto Barros, Porto;
and c Lab Cell Biology, ICBAS–Institute of Biomedical Sciences Abel Salazar, University of Porto, Porto, Portugal
Objective: To study the methylation imprinting marks of two oppositely imprinted genes, H19 and MEST/PEG1,in human testicular spermatozoa from azoospermic patients with different etiologies. Testicular spermatozoa areoften used in intracytoplasmic sperm injection for treatment of male factor infertility, but the imprinting status ofthese cells is currently unknown.Design: Experimental prospective study.Setting: University research laboratory and private in vitro fertilization (IVF) clinic.Patient(s): A total of 24 men, five with anejaculation, five with secondary obstructive azoospermia, five with pri-mary obstructive azoospermia, and nine with secretory azoospermia due to hypospermatogenesis.Intervention(s): Spermatozoa were isolated by micromanipulation from testicular biopsies.Main Outcome Measure(s): DNA methylation patterns were analyzed using bisulfite genomic sequencing withcloning analysis.Result(s): We found H19 complete methylation was statistically significantly reduced in secretory azoospermicpatients with hypospermatogenesis, with one patient presenting complete unmethylation. Hypomethylation alsoaffected the CTCF-binding site 6, involved in regulation of IGF2 expression. Regarding the MEST gene, all pa-tients presented complete unmethylation although this was statistically significantly reduced in the anejaculationgroup.Conclusion(s): Testicular spermatozoa from men with abnormal spermatogenesis carry methylation defects in theH19 imprinted gene which also affect the CTCF-binding site, further supporting an association between the occur-rence of imprinting errors and disruptive spermatogenesis. (Fertil Steril� 2010;94:585–94. �2010 by AmericanSociety for Reproductive Medicine.)
Key Words: Azoospermia, DNA methylation, genomic imprinting, H19, MEST, male factor infertility, testicularspermatozoa
Genomic imprinting leads to monoallelic parental-dependentexpression of a subset of genes in mammals. Parental allelesare marked differentially, primarily by DNA methylation ofCpG dinucleotides located on the differentially methylated re-gions of imprinted genes. These regulate the expression of im-printed genes and constitute the imprinting control regions(ICRs) (1). Methylation imprinting marks are erased inprimordial germ cells (PGCs) (2) and are later reestablishedduring germ cell development according to parental sex. Inmurine spermatogenesis, paternal imprints start to be ac-quired prenatally and are definitively established before the
November 28, 2008; revised February 1, 2009; accepted
y 16, 2009; published online April 1, 2010.
s nothing to disclose. T.F. has nothing to disclose. S.S. has
to disclose. F.C. has nothing to disclose. A.B. has nothing to
. M.S. has nothing to disclose.
pported by the Portuguese Foundation for Science and Tech-
(FCT) with a PhD fellowship (SFRH/BD/19967/2004 to CJM),
g Researcher fellowship (POCI/SAU-MMO/60555/04 to T.F.)
earch grants (POCI/SAU-MMO/60555/04, 60709/04, 59997/04;
quests: M�ario Sousa, M.D., Ph.D., Lab Cell Biology, Institute
edical Sciences Abel Salazar (ICBAS), University of Porto,
Prof Abel Salazar 2, 4099-003 Porto, Portugal (FAX:
22-206-22-32; E-mail: [email protected]).
2/$36.0016/j.fertnstert.2009.02.051 Copyright ª2010 American S
onset of meiosis (3, 4). It is interesting that paternal imprintsseem to be acquired earlier in the paternally inherited allelethan in the allele of maternal origin, suggesting that, evenwhen devoid of methylation, parental alleles can retain theiridentity (3). In humans, establishment of paternal imprintshas been shown to be complete by the primary spermatocytestage (5). On fertilization, a wave of DNA demethylation oc-curs, active in the paternal pronucleus and passive in the ma-ternal pronucleus, which does not affect the methylationimprinting marks previously set in gametes (6).
There are only three known imprinting control regions thatacquire methylation in the paternal germ line: H19-Igf2,Dlk1-Gtl2, and Rasgrf1 (7). H19 is one of the best character-ized imprinted genes; it is expressed from the maternal allele,it is imprinted in human (8) and mice (9), and it encodes anuntranslated RNA (10) whose function remains unclear. Itwas previously suggested to have tumor-suppressor activity(11) but also to be an oncofetal RNA (12), and was recentlydemonstrated to give rise to a microRNA (miR-675) (13).Additionally, H19 is physically and functionally linked to an-other imprinted gene, IGF2 (insulin-like growth factor-2)through the sharing of common enhancers (14, 15). H19and IGF2 are oppositely imprinted because H19 is expressed
Fertility and Sterility� Vol. 94, No. 2, July 2010 585ociety for Reproductive Medicine, Published by Elsevier Inc.

from the maternal allele and IGF2 is expressed from the pa-ternal allele. The H19 DMR (differentially methylated re-gion) is located upstream of the transcription start site andharbors several CTCF (CCCTC-binding factor) binding sites(16, 17); CTCF binds to the maternal unmethylated DMR andprevents IGF2 from accessing the common enhancers, thussilencing IGF2 expression. Oppositely, methylation on thepaternal DMR prevents the binding of CTCF, and IGF2 is ex-pressed. After fertilization, the promoter of H19 becomesmethylated and silences the H19 paternal allele while allow-ing the continued expression of IGF2 (18).
In humans, H19 is located in 11p15.5, a region linked toimprinting associated Beckwith-Wiedemann syndrome.This syndrome is characterized by congenital overgrowthand an increased risk of developing embryonic tumors suchas Wilms tumor. In some BWS patients, the genetic causehas been ascribed to the occurrence of H19 microdeletionsand hypermethylation of the maternal alleles, which leadsto IGF2 loss of imprinting and hyperexpression (19). Onthe other hand, paternal hypomethylation of H19, probablyleading to H19 biallelic expression and down-regulation ofIGF2, has recently been demonstrated to occur in the malegerm line, in association with oligozoospermia (20–22).Also, H19 hypomethylation was recently described as thecause of Silver-Russell syndrome, another congenital disor-der characterized by severe intrauterine and postnatal growthretardation (23).
MEST/PEG1 (mesodermal specific transcript/paternallyexpressed gene 1) is located in human chromosome 7q32,is paternally expressed (maternally methylated), and is im-printed in mice (24) and humans (25). MEST deficiency inmice, caused by a target mutation, was shown to cause gen-eral growth retardation of embryonic and extraembryonicstructures and abnormal maternal response to newborns(26). MEST was also found to encode an imprinted antisenseRNA (MESTIT1–MEST intronic transcript 1) that is only ex-pressed from the paternal allele which has been suggested toplay a role in the regulation of MESTexpression during devel-opment (27). This transcript was found to be predominantlyexpressed in the testis and in mature motile spermatozoa, in-dicating a possible role in human sperm physiology and fer-tilization (28).
The occurrence of imprinting syndromes in children bornafter assisted reproductive techniques (ART), usually present-ing loss of methylation of the maternal alleles, has raisedconcern about the safety of these techniques regarding the es-tablishment of imprints in the oocyte and maintenance duringin vitro embryo culture (7). However, imprinting methylationdefects have also been described in the paternal allele, in spermfrom patients with oligozoospermia, suggesting an associationbetween imprinting errors and infertility itself (20–22).
Spermatozoa retrieved from testis biopsies are often usedin ART, namely in intracytoplasmic sperm injection (ICSI),in cases of severe male factor infertility due to obstructiveor secretory azoospermia (29). However, the methylation sta-
586 Marques et al. Imprinting defects in testicular sperm
tus of imprinted genes in human testicular spermatozoa ofazoospermic patients is currently unknown. Both H19 andMEST were previously shown to be representative imprintedgenes for the evaluation of methylation errors present insperm from oligozoospermic patients (21). Additionally, in-creased expression of H19, IGF2, and MEST in the embryoand placenta suggests an important function for these genesin preimplantation and postimplantation embryo develop-ment (gene expression databases: SymAtlas at http://symat-las.gnf.org and Unigene at http://www.ncbi.nlm.nih.gov).
We describe H19 and MEST methylation patterns in testic-ular spermatozoa from patients with anejaculation (ANJ),secondary inflammatory obstructive azoospermia (OAZI),obstructive azoospermia due to congenital bilateral absenceof vas deferens (CBAVD), and secretory azoospermia dueto hypospermatogenesis (HP). We provide further evidencethat the occurrence of imprinting errors in the male germline is associated with impairment of spermatogenesis anddiscuss the possible implications of these new findings.
MATERIALS AND METHODS
Patient Recruitment and Classification
Under patients’ informed consent and according to the guide-lines of the medical ethics committee, testicular spermatozoawere retrieved from testicular biopsies of 24 cases, dividedinto four groups: ANJ (n ¼ 5) mainly due to spinal cordinjuries, secondary obstructive azoospermia due to inflam-matory epididymal disease (n ¼ 5), primary obstructiveazoospermia due to CBAVD (n ¼ 5), and secretory (nonob-structive) azoospermia due to hypospermatogenesis (n ¼9). Biopsies from normal fertile males were not includedbecause it was not possible to obtain testicular spermatozoafrom these cases. However, the present results were com-pared with cases with normal spermatogenesis, without(ANJ) or with (OAZI) epididymal obstruction, as well aswith data obtained from normoozoospermic ejaculated sper-matozoa (22).
All patients had normal karyotypes and absence of Y-chro-mosome AZF and DAZ microdeletions (30, 31). Ages rangedfrom 22 to 44 years with the average being of 32.3 (range: 22to 44 years) in ANJ, 37.2 years (range: 29 to 43 years) inOAZI, 35 years (range: 27 to 44 years) in CBAVD, and32.8 years (range: 27 to 42 years) in HP.
Isolation of spermatozoa by Micromanipulation fromTesticular Biopsies
Testicular biopsy samples were prepared as described previ-ously elsewhere (29). Briefly, each fragment was mechani-cally tweezed in sperm preparation medium (SPM withHEPES buffer; Medicult, Copenhagen, Denmark), and the re-sultant fluid was washed with SPM twice (5 minutes, 500� g).The pellet was resuspended in 2 mL of erythrocyte-lysingbuffer (155 mM NH4Cl, 10 mM KHCO3, 2 mM EDTA, pH7.2) (Sigma, Barcelona, Spain) during 5 minutes at 32�C,5% CO2 (32). After washing, samples were digested for 1
Vol. 94, No. 2, July 2010

hour at 32�C in SPM containing 25 mg/mL DNase and 1000IU/mL collagenase-IV (Sigma, cell cultured tested) (33). Afterwashing again, the pellet was resuspended in 50–100 mL ofIVF medium (Medicult) and incubated at 32�C, 5% CO2, untiluse. In a culture dish (cell culture tested; Nunc, Roskilde, Den-mark), 10 mL of cellular suspension were diluted in 100 mL ofSPM and covered with liquid light paraffin (Medicult, embryotested). Normal-appearing spermatozoa (300 to 400 cells)were isolated by micromanipulation in a Nikon research mi-croscope (Nikon, Tokyo, Japan) fitted with Hoffman optics,thermo-controlled stage, and Narishige micromanipulators(Narishige Scientific Instrument, Tokyo, Japan), using micro-pipettes of 4–5 mm final diameter (SweMed, Frolunda, Swe-den). After isolation, cells were transferred to a 10-mL dropof SPM and then to polymerase chain reaction (PCR) tubes us-ing mouth-driven super-thin Pasteur pipettes. Sperm cells werestored at –80�C until use.
DNA Isolation and Bisulfite Treatment
Sperm DNA was decondensed and extracted by adding 10 mlof alkaline lysis buffer—1 M KOH (Merck, Darmstadt,Germany) and 0.05 M DTT (Invitrogen, Carlsbad, CA)—to the sperm cells followed by incubation at 80�C for 20 min-utes. The reaction was stopped by adding 10 mL of neutrali-zation buffer—0.9 M Tris-HCl (Sigma, Steinheim,Germany), 0.3 M KCl, and 0.2 M HCl, pH 8.3 (Merck). Ex-tracted DNA was then treated and modified with a sodium bi-sulfite–based procedure using the Epitect Bisulfite kit(Qiagen, Hilden, Germany), according to the manufacturer’sinstructions. Briefly, this protocol enables to reduce DNAdegradation and maximize DNA recovery by reducing the in-cubation time with bisulfite solution to 5 hours and usingRNA carrier to recover modified DNA. The DNA was theneluted in 20 mL of elution buffer, with 10 mL used for ampli-fication of each gene. Bisulfite converts unmethylated cyto-sines to uracil while 5-methylcytosines (5-MeC) remainunchanged. The efficiency of the procedure was ensured bya conversion rate of non-CpG cytosines to uracil higherthan 95%.
PCR Amplification, Cloning, and Sequencing
Bisulfite-modified DNA was amplified by hemi-nested PCRfor H19 and MEST imprinted genes, with the primersdescribed elsewhere (5). For H19, a sequence with 231 bp(AF087017: 6098–6328) encompassing 18 CpGs was ana-lyzed. For MEST, the amplified region with 219 bp(Y10620: 609-827) and encompassing 22 CpGs is locatedin the DMR and includes part of the first exon and part ofthe first intron. A first-round PCR reaction in a final volumeof 50 mL was prepared for each gene and their respective neg-ative controls (DNA extraction, bisulfite treatment, and PCRreaction), containing 1X buffer with 1.5 mM MgCl2 (Qia-gen), 0.12 mM of each dNTP (Invitrogen), 0.5 mM of eachprimer (Thermo Electron, Ulm, Germany), 1.5 IU HotStar-Taq enzyme (5 IU/mL; Qiagen), and 10 mL of the eluted mod-ified DNA. The PCR conditions were initial strand
Fertility and Sterility�
denaturation (15 minutes, 95�C) followed by 30 amplifica-tion cycles (denaturation, 1 minute, 94�C; primer annealing,1 minute, 60�C; strand elongation, 1 minute, 72�C) and a finalextension (20 minutes, 72�C). Because the amount of startingmaterial is necessarily very low (300 to 400 haploid cells), wewere only able to perform one PCR reaction for each geneand sample. The second-round PCR reaction was preparedwith 1 mL of the first-round PCR product, using the sameconditions of the first PCR.
Amplified PCR products were purified with the GFXPCR-DNA and Gel Band Purification Kit (GE Healthcare,Buckinghamshire, UK), according to the manufacturer’s in-structions, and were cloned with the TOPO TA cloning kit(Invitrogen) using the E. coli Mach1-T1 (Invitrogen) bacteriastrain and the pCR II vector (Invitrogen), according to themanufacturer’s instructions. About 20 clones containing thefragment of interest were sequenced for each sample andgene by the use of the BigDye Terminator Cycle Sequencingv1.1 Ready Reaction kit (Applied Biosystems, Foster City,CA) and were analyzed in an ABI PRISM 310 Genetic Ana-lyzer (Applied Biosystems). Direct sequencing of all sampleswas also performed to assess the samples’ overall methyla-tion and to ensure the absence of cloning bias.
Statistics
Statistical analysis was performed with the use of the two-sided Fisher’s exact test (SPSS 15 for Windows; SPSS,Inc., Chicago, IL) for comparisons between groups, whichwere weighted by the number of clones analyzed. P<.05was considered statistically significant.
RESULTS
Methylation of the H19 DMR in Human TesticularSpermatozoa
We analyzed a sequence located within the DMR of H19 con-taining 18 CpGs including the human CTCF-binding site 6,represented in Figure 1 (16). This sequence was previouslyshown elsewhere to acquire the methylation imprintingmark in the male germ line (5). One of the analyzed CpGs(CpG 7, Fig. 2) was not considered for the quantitative anal-ysis because it was reported as a polymorphism between acytosine and a thymine, rendering it not informative in termsof methylation status (34).
In our study, a total of 500 clones from 24 cases were an-alyzed for H19; the results per individual and per group arerepresented in Table 1 and Figure 2. Of the 24 cases analyzed,20 had clones with H19 completely methylated (18 CpGsmethylated or only CpG 7 unmethylated) (see Fig. 2). The re-maining four patients did not present H19 complete methyl-ation and were all from the hypospermatogenesis group.Additionally, one of these patients (patient 16, see Fig. 2) pre-sented 9 of the 10 clones analyzed completely unmethylated.Further evaluation on the outcome of ART in this specific pa-tient revealed that this patient underwent ICSI treatment withthe resultant embryos showing developmental arrest.
587

FIGURE 1
Schematic representation of the H19 DMR, consisting of 3.5 repeats of 400 bp (50), the neighboring gene IGF2and the two shared endodermal enhancers (EE). The H19 region analyzed (18 CpGs), comprising the humanconserved CTCF site 6 (CpG 4–8), is represented. Positions relative to H19 transcription start site are depicted.
H19
-2.0 kb-3.4 kb
DMR
-2145/-2135/-2133/-2131/-2128
CpG
EE
CTCF-6
IGF2
Marques. Imprinting defects in testicular sperm. Fertil Steril 2010.
Comparing the different azoospermia groups, we observedthat the OAZI group presented the highest frequency ofclones with H19 completely methylated, followed by ANJ,CBAVD (P¼.011 to OAZI), and HP (P<.001 to ANJ andOAZI) (Table 1). Additionally, we observed H19 completeunmethylation in testicular sperm from one patient of theHP group, representing 5.4% of HP clones (P¼.012 forANJ; P¼.013 for OAZI and CBAVD). This imprinting defectwas also previously observed in oligozoospermic patientswith less than 10 million sperm per mL (22). The numberof unmethylated CpGs varied between one and two in ANJand CBAVD, one and three in OAZI and 1 and 17 in HP(see Fig. 2; Table 1). Considering the total number of meth-ylated CpGs, the HP group also presented the lowest value(P<.001 for OAZI, ANJ, and CBAVD) (see Table 1).Although this decreased value was clearly influenced byone single patient (patient 16) who presented 90% of clonescompletely unmethylated, the percentage of the total methyl-ation calculated without this patient was also statistically sig-nificantly different from the other groups (95%; P<.001 forANJ and OAZI and P¼.046 for CBAVD).
Comparing the level of unmethylation at each CpG in thetotal number of clones analyzed, we observed that CpG 6 wasthe most affected (P<.001 for all other CpGs) (Fig. 3). Thetotal frequency of unmethylation at each CpG varied between1.8% and 26.2%. Percentages of unmethylation at each CpGin the four groups of azoospermic patients and in ejaculatedsperm from normozoospermic males (22) are represented inFigure 3.
Methylation of the Human CTCF-Binding Site 6
The CTCF-binding site 6, one of the seven that are present inthe H19 DMR in humans (16), is located in the analyzed se-quence (CpG 4–8; see Figs. 1 and 2). Methylation of this spe-cific site was previously shown elsewhere to be involved inregulation of IGF2 expression (16). Complete methylationof this region was found in 20 of the 24 patients analyzed,with the four remaining patients belonging to the HP group(patients 16, 17, 19, and 21; see Fig. 1). The HP group pre-sented a decreased percentage of clones completely methyl-
588 Marques et al. Imprinting defects in testicular sperm
ated when compared with all other testicular sperm groups(P<.001 with ANJ, OAZI, and CBAVD) (see Table 2) andwith the results previously obtained in ejaculated spermato-zoa from oligozoospermic patients (67% of clones com-pletely methylated) (22) (P¼.006 with oligozoospermia).However, if we consider the results from oligozoospermic pa-tients with less than 10 million sperm per mL, then the per-centages are less different (54% in HP vs. 62% inoligozoospermia <10 M; P¼.089). This clearly correlateswith the severity of abnormal spermatogenesis. Comparingwith ejaculated sperm from normozoospermic controls, therewas no difference in CTCF-6 complete methylation withANJ, OAZI, and CBAVD, although HP was statistically sig-nificantly reduced (P<.001). The only group presenting com-plete unmethylation was HP in one patient and 5.4% of theclones (P¼.012 with ANJ; P¼.013 with OAZI and CBAVD).The number of unmethylated CpGs was of only one in allgroups except HP where it varied between one to four, withfour patients presenting two unmethylated CpGs and one pa-tient presenting four (see Fig. 2; Table 2).
Regarding the total number of methylated CpGs in thissite, all groups presented high levels of methylation exceptHP, which was statistically significantly lower than the others(P<.001 with ANJ, OAZI, and CBAVD) (see Table 2). Oncemore, if we discard patient 16 who showed complete unme-thylation, the HP value continued to be statistically signifi-cantly reduced (83.8%; P<.001 with ANJ, OAZI, andCBAVD).
Methylation Analysis of MEST DMR
A total of 502 clones of the same 24 patients analyzed forH19 were evaluated for the MEST gene (see Fig. 2; Table 3).
All 24 patients presented MEST completely unmethy-lated (see Fig. 2). However, we found a statistically signif-icant decrease in the percentage of completelyunmethylated clones in the ANJ group compared with tes-ticular sperm from all other groups (P¼.001 with CBAVD;P¼.002 with OAZI; and P¼.011 with HP) (see Table 3)but also with ejaculated sperm from normal men and oligo-zoospermic patients (22). One patient from the OAZI
Vol. 94, No. 2, July 2010
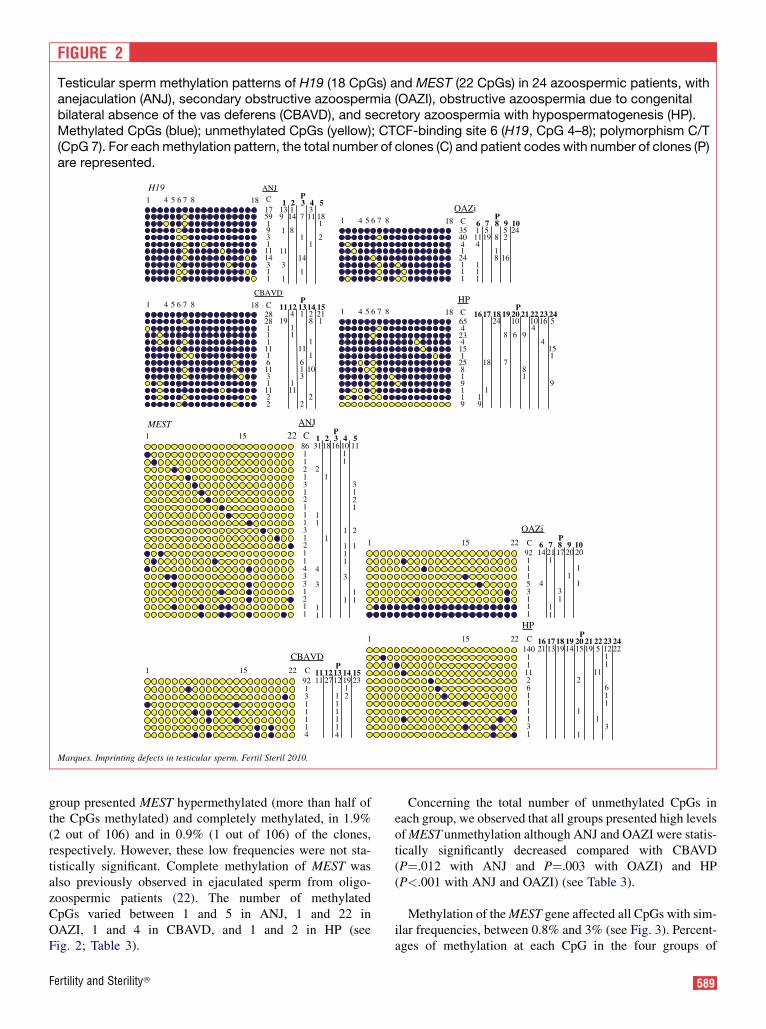
FIGURE 2
Testicular sperm methylation patterns of H19 (18 CpGs) and MEST (22 CpGs) in 24 azoospermic patients, withanejaculation (ANJ), secondary obstructive azoospermia (OAZI), obstructive azoospermia due to congenitalbilateral absence of the vas deferens (CBAVD), and secretory azoospermia with hypospermatogenesis (HP).Methylated CpGs (blue); unmethylated CpGs (yellow); CTCF-binding site 6 (H19, CpG 4–8); polymorphism C/T(CpG 7). For each methylation pattern, the total number of clones (C) and patient codes with number of clones (P)are represented.
H19P
1 2 3 4 513 1 3 9 14 7 11 18
1 1 8
1 2 1
1114
3 1
1
ANJ
1411
193
5917
1
113
8 185 641 7
P6 7 8 9 10 1 5 5 24 11 19 8 2 4
1 8 16
1 1 1
OAZi
11
4124
4035
1
C8 186541 7
P11 12 13 14 15
4 1 2 2119 8 1
1 1
1 11
1 6 1 10 3
1 11
2 2
CBAVD
C
C
8 186541 7
61
111
2828
11
1311
2211
HPC8 186541 7
825
23415
465
1
191
91
P16 17 18 19 20 21 22 23 24
24 10 10 16 5 4
8 6 9 4
15 1
18 7 8 1
9 1
1 9
MEST ANJ22151 C P
1 2 3 4 531 18 16 10 11
1 1
2 1
3121
1 1
1 21
1 11 1
4 3
3 1
1 11 1
86
31211
12
13111
21
13341
211
22151 C
OAZiP
6 7 8 9 1014 21 17 20 20
1 1
1 4 1
3 1
1 1
92
35111
111
CBAVD22151 C
P11 12 13 14 1511 27 12 19 23
1 1 2 1 1 1 1 4
92
11131
14
HP22151 C
140
621111
111
131
P16 17 18 19 20 21 22 23 24 21 13 19 14 15 19 5 12 22
1 1
11 2
6 1 1
1 1
3 1
Marques. Imprinting defects in testicular sperm. Fertil Steril 2010.
group presented MEST hypermethylated (more than half ofthe CpGs methylated) and completely methylated, in 1.9%(2 out of 106) and in 0.9% (1 out of 106) of the clones,respectively. However, these low frequencies were not sta-tistically significant. Complete methylation of MEST wasalso previously observed in ejaculated sperm from oligo-zoospermic patients (22). The number of methylatedCpGs varied between 1 and 5 in ANJ, 1 and 22 inOAZI, 1 and 4 in CBAVD, and 1 and 2 in HP (seeFig. 2; Table 3).
Fertility and Sterility�
Concerning the total number of unmethylated CpGs ineach group, we observed that all groups presented high levelsof MEST unmethylation although ANJ and OAZI were statis-tically significantly decreased compared with CBAVD(P¼.012 with ANJ and P¼.003 with OAZI) and HP(P<.001 with ANJ and OAZI) (see Table 3).
Methylation of the MEST gene affected all CpGs with sim-ilar frequencies, between 0.8% and 3% (see Fig. 3). Percent-ages of methylation at each CpG in the four groups of
589

TABLE 1Methylation status of H19 in human testicular sperm.
Number of unmethylated CpGs
Groups Clones (n) 0 1 2 3 17 Total methylation
ANJ 120 63.3a,b 32.5 4.2 97.6OAZI 107 70.1a 29 0.9 98.1CBAVD 107 52.3b,c 32.7 15 96.3a
HP 166 41.6c 25.9 25.9 1.2 5.4a 89.8b
Notes: Values are given in percentages (%) of clones (n, total number of clones analyzed for each group). Total methylationwas calculated dividing the total number of methylated CpGs by the total number of CpGs analyzed (except CpG7). ANJ,anejaculation; CBAVD, obstructive azoospermia due to congenital bilateral absence of vas deferens; HP, secretory azo-ospermia due to hypospermatogenesis; OAZI, secondary inflammatory obstructive azoospermia.
a,b,c Statistically significant differences between groups are represented by superscript letters in columns 3, 7, and 8. Dif-ferent letters denote statistically significant differences (Fisher’s exact test, two-sided).
Marques. Imprinting defects in testicular sperm. Fertil Steril 2010.
azoospermic patients and in ejaculated sperm from normo-zoospermic males (22) are represented in Figure 3.
DISCUSSION
Testicular sperm are often used for ICSI treatment of malefactor infertility due to azoospermia. Here we describe thefirst study of the methylation status of two imprinted genes,H19 and MEST, in these sperm cells, obtained from testicularbiopsies of patients with different causes of infertility: ANJ,secondary obstruction, CBAVD, and secretory azoospermiawith hypospermatogenesis (HP). We observed that H19 andCTCF-binding site 6 complete methylation was present inall groups but was statistically significantly reduced in thetesticular sperm from azoospermic patients presenting withHP, where four of the nine patients analyzed did not presentcompletely methylated clones. Hypomethylation and com-plete unmethylation of H19 as well as of the CTCF-bindingsite 6 also affected the HP group. We previously describedelsewhere that ejaculated sperm from oligozoospermicpatients with less than 10 million sperm per mL of semenalso present methylation errors at H19 gene which also affectthe CTCF-binding site (20, 22). Our current observations fur-ther support that H19 hypomethylation occurs in associationwith impaired spermatogenesis of idiopathic causes—because no abnormalities were found in the karyotype andY-chromosome of these patients—increasing in severityfrom moderate oligozoospermia to azoospermia with HP.
On the other hand, MEST gene correct demethylationseems not to be affected in sperm from azoospermic patients.However, we did find a decreased frequency of completelyunmethylated clones in the ANJ group, where the majorityof patients (four out of five) suffered from paraplegia dueto spinal cord injuries. It has been described that the qualityof sperm in the semen of these patients is low, with decreasedmotility and viability and with ultrastructural analysis reveal-ing defects similar to those described for epididymal necros-
590 Marques et al. Imprinting defects in testicular sperm
permia, such as abnormal flagella, mitochondria, andacrosome structures (35). Additionally, increased DNA frag-mentation has also been reported in these patients, possiblyrelated to elevated levels of cytokines and free radicals intheir seminal plasma (36). However, given that the formerstudies were performed on ejaculated sperm retrieved byelectroejaculation, it is not known if sperm retrieved fromthe testis of ANJ patients also present these defects Wehere show that abnormal methylation of the MEST geneoccurs in sperm from ANJ patients although the correlation,if any, with the previous findings is unclear.
The biological significance of few unmethylated CpGs atthe H19 gene and few methylated CpGs at MEST gene isnot known, except if it affects the CTCF-binding site inH19 DMR. However, we could speculate that, because theseare also present in spermatozoa from normal men (22), it isunlikely that it would interfere with sperm function or withthe correct expression of imprinted genes. On the other hand,in men with normal semen parameters but who also producespermatozoa with imprinting errors, these abnormal gametescould be selectively discarded and not be able to fertilize oo-cytes or give rise to embryos that would fail to develop nor-mally, as suggested by studies performed in mice usingazacytidine to induce hypomethylation in sperm (37). How-ever, when using the ICSI technique it is not possible to avoidbypassing the natural selection mechanisms, and selection ofspermatozoa with imprinting errors might thus be one of thecauses for poor embryo development. Presently ICSI is themethod of choice in cases of oligoasthenoteratozoospermia,and obstructive and secretory (nonobstructive) azoospermia.It has been reported that ICSI using testicular sperm frommen with secretory azoospermia leads to lower rates of fertil-ization and pregnancy compared with testicular sperm from pa-tients with obstructive azoospermia (38), which correlates withour results obtained for the occurrence of imprinting defects.
Some imprinting disorders have been reported in ARTchildren, such as Angelman syndrome (AS) and Beckwith-
Vol. 94, No. 2, July 2010

FIGURE 3
Percentage of unmethylation (white bars þ standard deviation) at each analyzed CpG (1–18) for H19 andpercentage of methylation (black bars þ standard deviation) at each analyzed CpG (1–22) for MEST in testicularsperm and in ejaculated sperm from normozoospermic patients (NZ) (22). Anejaculation (ANJ); secondaryobstructive azoospermia (OAZI); primary obstructive azoospermia due to congenital bilateral absence of the vasdeferens (CBAVD); secretory azoospermia with hypospermatogenesis (HP); CTCF-binding site 6 (H19 CpG 4–8).H19 CpG 7 is a polymorphism, not informative in terms of methylation status.
H19
02468101214
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
%
CpG
Total
0
2
4
6
8
10
12
14
1 2 3 4 5 6 7 8 9 10111213141516171819202122
%
CpG
ANJ
0
2
4
6
8
10
12
14
1 2 3 4 5 6 7 8 9 10 111213141516171819202122
%
CpG
OAZi
0
2
4
6
8
10
12
14
1 2 3 4 5 6 7 8 9 10111213141516171819202122
%
CpG
CBAVD
0
2
4
6
8
10
12
14
1 2 3 4 5 6 7 8 9 10111213141516171819202122
%
CpG
HP
0
2
4
6
8
10
12
14
1 2 3 4 5 6 7 8 9 10111213141516171819 202122
%
CpG
NZMEST
0102030405060708090100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
%
CpG
NZ
0102030405060708090100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
%
CpG
ANJ
0102030405060708090100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
%
CpG
OAZi
0102030405060708090100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
%
CpG
CBAVD
0102030405060708090100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
%
CpG
HP
0102030405060708090100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
%
CpG
Total
Marques. Imprinting defects in testicular sperm. Fertil Steril 2010.
Fertility and Sterility� 591

TABLE 2Methylation status of CTCF-binding site 6 in human testicular sperm.
Number of unmethylated CpGs
Groups Clones (n) 0 1 2 4 Total methylation
ANJ 120 85 15 96.3OAZi 107 77.6 22.4 94.4CBAVD 107 86.9 13.1 96.7HP 166 53.6a 19.9 21.1 5.4a 79.1a
Notes: Values are given in percentages (%) of clones (n, total number of clones analyzed for each group). Total methylationwas calculated dividing the total number of methylated CpGs at CTCF-6 by the total number of CpGs analyzed at theCTCF-6 (except CpG7). ANJ, anejaculation; CBAVD, obstructive azoospermia due to congenital bilateral absence ofvas deferens; HP, secretory azoospermia due to hypospermatogenesis; OAZI, secondary inflammatory obstructive azo-ospermia.
a Statistically significant difference between groups (Fisher’s exact test, two-sided).
Marques. Imprinting defects in testicular sperm. Fertil Steril 2010.
Wiedemann syndrome where an imprinting defect has beenfound in all cases related to abnormal methylation patternson the maternal allele (7). It was suggested that hormonalstimulation and/or in vitro culture of the early embryo coulddisturb the establishment and maintenance of maternal im-printing marks. However, we and others have also reportedan association between infertility itself and the occurrenceof imprinting errors in male gametes, such as H19 hypome-thylation and MEST hypermethylation (20–22). In this re-gard, a large Dutch population study observed a threefoldincrease in the incidence of imprinting syndromes (Prader-Willi, Angelman, and Beckwith-Wiedemann syndromes) inART children. However, after correction for impaired fertil-ity, the investigators concluded that the risk of transmittingimprinting disorders using ART is the same as in the generalpopulation, suggesting that imprinting syndromes in ARTchildren might be related to the type of infertility and notto the techniques used in assisted reproduction (39).
Hypomethylation of the H19 DMR has been reportedin patients with Silver-Russell syndrome, which is
TABLE 3Methylation status of MEST in human testicular spe
Number of meth
Groups Clones (n) 0 1 2 4
ANJ 123 69.9a 16.3 12.2 0.8OAZi 106 86.8 10.4 0.9CBAVD 104 88.5 3.8 3.8 3.8HP 169 82.8 14.2 3
Notes: Values are given in percentages (%) of clones (n, total nlation was calculated dividing the total number of unmethylajaculation; CBAVD, obstructive azoospermia due to congazoospermia due to hypospermatogenesis; OAZI, secondar
a Statistically significant differences between groups (Fisher’s
Marques. Imprinting defects in testicular sperm. Fertil Steril 2010.
592 Marques et al. Imprinting defects in testicular sperm
characterized by severe intrauterine and postnatal growthretardation among other clinical features (23, 40). Silver-Russell syndrome also has been recently reported insome cases of ART children (40–42), of which oneshowed H19 hypomethylation (40) and another a slighthypermethylation of MEST with no alterations on H19methylation (43).
The cause of H19 hypomethylation in sperm from infertilepatients is not known. We previously hypothesized that thismight be due to abnormal activity of the DNA methyltrans-ferases responsible for the establishment of methylationmarks (20), or eventually for the maintenance of these im-printing marks during adult spermatogenic cell divisions. Itwas recently shown that disruption of Dnmt3a and Dnmt3l,responsible for de novo methylation, in mice prospermatogo-nia causes H19 hypomethylation (44). Additionally, CTCFL(also known as BORIS), a testis-specific paralogue of theCTCF protein, has also been suggested to play a role in theestablishment of imprints in the male germ line (45) and par-ticularly in H19 DMR (46). The fact that the CTCF-binding
rm.
ylated CpGs
5 21 22 >11 Total unmethylation
0.8 97.8a
0.9 0.9 1.9 97.6a
98.899.1
umber of clones analyzed for each group). Total unmethy-ted CpG by the total number of CpGs analyzed. ANJ, ane-enital bilateral absence of vas deferens; HP, secretoryy inflammatory obstructive azoospermia.exact test, two-sided).
Vol. 94, No. 2, July 2010

site is frequently hypomethylated or completely unmethy-lated in cases with disturbed spermatogenesis suggests thatCTCF, or CTCFL, might bind during germ cell developmentand prevent the correct establishment of male imprints.
A recent study demonstrated that methylation of the firstand second CpGs at the 12-bp core sequence of the CTCF-binding site, which corresponds to CpG 5 and 6 in our study(as CpG 4 is located outside the core sequence), inhibited thebinding of CTCF, with the first CpG being the one with thestrongest effect (47). It is interesting that the most affectedCpG in our results was the CpG 6 of the CTCF-bindingsite, corresponding to the second CpG of the 12-bp coresite; the first CpG (CpG 5) appeared unmethylated only inthe HP group. Additionally, it was demonstrated that methyl-ation at the third CpG (CpG 7) had no effect in binding ofCTCF. This fact indicates that CTCF binding can be regu-lated by methylation at a single CpG.
Both H19 hypomethylation, possibly resulting in IGF2 in-activation, and MEST hypermethylation might be related tothe low birth weight observed in ART children (48) becauseboth IGF2 and MEST are growth promoter genes. Actually,decreased MEST expression has been observed in placentasof nonsyndromic human intrauterine growth restriction(IUGR) (49).
Our results suggest an increased risk of methylation errorsin testicular spermatozoa from patients with hypospermato-genesis, particularly at H19 imprinted gene and includingthe human CTCF-binding site 6. This is in accordance withprevious findings of imprinting methylation defects in ejacu-lated sperm from patients with low sperm counts, and it sup-ports the need for further research on the (epi)genetic causesof infertility and for follow-up studies on imprinting-relateddiseases in children born after ART.
Acknowledgments: The authors thank Dr. Susana Fernandes for Y-chromo-
some microdeletions analysis, Cristiano Oliveira, M.D., and Jos�e Manuel
Teixeira da Silva, M.D. (gynecology and obstetrics) for couple evaluation,
Luis Ferraz, M.D. (urology) for testicular surgery, and Joaquina Silva,
M.D. (embryology) for testicular biopsies preparation (CRGAB).
REFERENCES1. Reik W, Walter J. Genomic imprinting: parental influence on the
genome. Nat Rev Genet 2001;2:21–32.
2. Hajkova P, Erhardt S, Lane N, Haaf T, El-Maarri O, Reik W, et al. Epi-
genetic reprogramming in mouse primordial germ cells. Mech Dev
2002;117:15–23.
3. Davis TL, Yang GJ, McCarrey JR, Bartolomei MS. The H19 methylation
imprint is erased and re-established differentially on the parental alleles
during male germ cell development. Hum Mol Genet 2000;9:2885–94.
4. Ueda T, Abe K, Miura A, Yuzuriha M, Zubair M, Noguchi M, et al. The
paternal methylation imprint of the mouse H19 locus is acquired in the
gonocyte stage during foetal testis development. Genes Cells 2000;5:
649–59.
5. Kerjean A, Dupont JM, Vasseur C, Le Tessier D, Cuisset L, Paldi A, et al.
Establishment of the paternal methylation imprint of the human H19 and
MEST/PEG1 genes during spermatogenesis. Hum Mol Genet 2000;9:
2183–7.
6. Santos F, Hendrich B, Reik W, Dean W. Dynamic reprogramming of DNA
methylation in the early mouse embryo. Dev Biol 2002;241:172–82.
Fertility and Sterility�
7. Arnaud P, Feil R. Epigenetic deregulation of genomic imprinting in
human disorders and following assisted reproduction. Birth Defects
Res C Embryo Today 2005;75:81–97.
8. Zhang Y, Tycko B. Monoallelic expression of the human H19 gene. Nat
Genet 1992;1:40–4.
9. Bartolomei MS, Zemel S, Tilghman SM. Parental imprinting of the
mouse H19 gene. Nature 1991;351:153–5.
10. Brannan CI, Dees EC, Ingram RS, Tilghman SM. The product of the H19
gene may function as an RNA. Mol Cell Biol 1990;10:28–36.
11. Hao Y, Crenshaw T, Moulton T, Newcomb E, Tycko B. Tumour-suppres-
sor activity of H19 RNA. Nature 1993;365:764–7.
12. Ariel I, Ayesh S, Perlman EJ, Pizov G, Tanos V, Schneider T, et al. The
product of the imprinted H19 gene is an oncofetal RNA. Mol Pathol
1997;50:34–44.
13. Cai X, Cullen BR. The imprinted H19 noncoding RNA is a primary
microRNA precursor. RNA 2007;13:313–6.
14. Zemel S, Bartolomei MS, Tilghman SM. Physical linkage of two mam-
malian imprinted genes, H19 and insulin-like growth factor 2. Nat Genet
1992;2:61–5.
15. Leighton PA, Saam JR, Ingram RS, Stewart CL, Tilghman SM. An en-
hancer deletion affects both H19 and Igf2 expression. Genes Dev
1995;9:2079–89.
16. Takai D, Gonzales FA, Tsai YC, Thayer MJ, Jones PA. Large scale map-
ping of methylcytosines in CTCF-binding sites in the human H19
promoter and aberrant hypomethylation in human bladder cancer. Hum
Mol Genet 2001;10:2619–26.
17. Bell AC, Felsenfeld G. Methylation of a CTCF-dependent boundary con-
trols imprinted expression of the Igf2 gene. Nature 2000;405:482–5.
18. Tremblay KD, Duran KL, Bartolomei MS. A 5’ 2-kilobase-pair region of
the imprinted mouse H19 gene exhibits exclusive paternal methylation
throughout development. Mol Cell Biol 1997;17:4322–9.
19. Enklaar T, Zabel BU, Prawitt D. Beckwith-Wiedemann syndrome: mul-
tiple molecular mechanisms. Expert Rev Mol Med 2006;8:1–19.
20. Marques CJ, Carvalho F, Sousa M, Barros A. Genomic imprinting in dis-
ruptive spermatogenesis. Lancet 2004;363:1700–2.
21. Kobayashi H, Sato A, Otsu E, Hiura H, Tomatsu C, Utsunomiya T, et al.
Aberrant DNA methylation of imprinted loci in sperm from oligospermic
patients. Hum Mol Genet 2007;16:2542–51.
22. Marques CJ, Costa P, Vaz B, Carvalho F, Fernandes S, Barros A, et al.
Abnormal methylation of imprinted genes in human sperm is associated
with oligozoospermia. Mol Hum Reprod 2008;14:67–74.
23. Gicquel C, Rossignol S, Cabrol S, Houang M, Steunou V, Barbu V, et al.
Epimutation of the telomeric imprinting center region on chromosome
11p15 in Silver-Russell syndrome. Nat Genet 2005;37:1003–7.
24. Kaneko-Ishino T, Kuroiwa Y, Miyoshi N, Kohda T, Suzuki R,
Yokoyama M, et al. Peg1/Mest imprinted gene on chromosome 6 identi-
fied by cDNA subtraction hybridization. Nat Genet 1995;11:52–9.
25. Nishita Y, Yoshida I, Sado T, Takagi N. Genomic imprinting and chro-
mosomal localization of the human MEST gene. Genomics 1996;36:
539–42.
26. Lefebvre L, Viville S, Barton SC, Ishino F, Keverne EB, Surani MA.
Abnormal maternal behaviour and growth retardation associated with
loss of the imprinted gene Mest. Nat Genet 1998;20:163–9.
27. Nakabayashi K, Bentley L, Hitchins MP, Mitsuya K, Meguro M,
Minagawa S, et al. Identification and characterization of an imprinted
antisense RNA (MESTIT1) in the human MEST locus on chromosome
7q32. Hum Mol Genet 2002;11:1743–56.28. Li T, Vu TH, Lee KO, Yang Y, Nguyen CV, Bui HQ, et al. An imprinted
PEG1/MEST antisense expressed predominantly in human testis and in
mature spermatozoa. J Biol Chem 2002;277:13518–27.29. Sousa M, Cremades N, Silva J, Oliveira C, Ferraz L, Teixeira da Silva J,
et al. Predictive value of testicular histology in secretory azoospermic
subgroups and clinical outcome after microinjection of fresh and fro-
zen-thawed sperm and spermatids. Hum Reprod 2002;17:1800–10.
30. Fernandes S, Huellen K, Goncalves J, Dukal H, Zeisler J, Rajpert De
Meyts E, et al. High frequency of DAZ1/DAZ2 gene deletions in patients
with severe oligozoospermia. Mol Hum Reprod 2002;8:286–98.
31. Pinho MJ, Neves R, Costa P, Ferras C, Sousa M, Alves C, et al. Unique
t(Y;1)(q12;q12) reciprocal translocation with loss of the heterochromatic
593

region of chromosome 1 in a male with azoospermia due to meiotic
arrest: a case report. Hum Reprod 2005;20:689–96.
32. Verheyen G, De Croo I, Tournaye H, Pletincx I, Devroey P, van
Steirteghem AC. Comparison of four mechanical methods to retrieve
spermatozoa from testicular tissue. Hum Reprod 1995;10:2956–9.
33. Crabbe E, Verheyen G, Tournaye H, Van Steirteghem A. The use of enzymatic
procedures to recover testicular germ cells. Hum Reprod 1997;12:1682–7.
34. Frevel MA, Hornberg JJ, Reeve AE. A potential imprint control element:
identification of a conserved 42 bp sequence upstream of H19. Trends
Genet 1999;15:216–8.
35. Mallidis C, Lim TC, Hill ST, Skinner DJ, Brown DJ, Johnston WI, et al.
Necrospermia and chronic spinal cord injury. Fertil Steril 2000;74:221–7.
36. Brackett NL, Ibrahim E, Grotas JA, Aballa TC, Lynne CM. Higher sperm
DNA damage in semen from men with spinal cord injuries compared
with controls. J Androl 2008;29:93–9;discussion 100–1.
37. Oakes CC, Kelly TL, Robaire B, Trasler JM. Adverse effects of 5-aza-2’-
deoxycytidine on spermatogenesis include reduced sperm function and
selective inhibition of de novo DNA methylation. J Pharmacol Exp
Ther 2007;322:1171–80.
38. Vernaeve V, Tournaye H, Osmanagaoglu K, Verheyen G, Van
Steirteghem A, Devroey P. Intracytoplasmic sperm injection with testicular
spermatozoa is less successful in men with nonobstructive azoospermia
than in men with obstructive azoospermia. Fertil Steril 2003;79:529–33.
39. Doornbos ME, Maas SM, McDonnell J, Vermeiden JP, Hennekam RC.
Infertility, assisted reproduction technologies and imprinting distur-
bances: a Dutch study. Hum Reprod 2007;22:2476–80.
40. Bliek J, Terhal P, van den Bogaard MJ, Maas S, Hamel B, Salieb-
Beugelaar G, et al. Hypomethylation of the H19 gene causes not only Sil-
ver-Russell syndrome (SRS) but also isolated asymmetry or an SRS-like
phenotype. Am J Hum Genet 2006;78:604–14.
41. Kallen B, Finnstrom O, Nygren KG, Olausson PO. In vitro fertilization
(IVF) in Sweden: risk for congenital malformations after different IVF
methods. Birth Defects Res A Clin Mol Teratol 2005;73:162–9.
594 Marques et al. Imprinting defects in testicular sperm
42. Svensson J, Bjornstahl A, Ivarsson SA. Increased risk of Silver-Russell
syndrome after in vitro fertilization? Acta Paediatr 2005;94:1163–5.
43. Kagami M, Nagai T, Fukami M, Yamazawa K, Ogata T. Silver-Russell
syndrome in a girl born after in vitro fertilization: partial hypermethyla-
tion at the differentially methylated region of PEG1/MEST. J Assist
Reprod Genet 2007;24:131–6.
44. Kato Y, Kaneda M, Hata K, Kumaki K, Hisano M, Kohara Y, et al. Role
of the Dnmt3 family in de novo methylation of imprinted and repetitive
sequences during male germ cell development in the mouse. Hum Mol
Genet 2007;16:2272–80.
45. Loukinov DI, Pugacheva E, Vatolin S, Pack SD, Moon H, Chernukhin I,
et al. BORIS, a novel male germ-line-specific protein associated with
epigenetic reprogramming events, shares the same 11-zinc-finger
domain with CTCF, the insulator protein involved in reading imprinting
marks in the soma. Proc Natl Acad Sci USA 2002;99:6806–11.
46. Jelinic P, Stehle JC, Shaw P. The testis-specific factor CTCFL cooperates
with the protein methyltransferase PRMT7 in H19 imprinting control
region methylation. PLoS Biol 2006;4:e355.
47. Renda M, Baglivo I, Burgess-Beusse B, Esposito S, Fattorusso R,
Felsenfeld G, et al. Critical DNA binding interactions of the insulator
protein CTCF: a small number of zinc fingers mediate strong binding,
and a single finger-DNA interaction controls binding at imprinted loci.
J Biol Chem 2007;282:33336–45.
48. Schieve LA, Meikle SF, Ferre C, Peterson HB, Jeng G, Wilcox LS. Low
and very low birth weight in infants conceived with use of assisted repro-
ductive technology. N Engl J Med 2002;346:731–7.
49. McMinn J, Wei M, Schupf N, Cusmai J, Johnson EB, Smith AC, et al.
Unbalanced placental expression of imprinted genes in human intrauter-
ine growth restriction. Placenta 2006;27:540–9.
50. Jinno Y, Sengoku K, Nakao M, Tamate K, Miyamoto T, Matsuzaka T,
et al. Mouse/human sequence divergence in a region with a paternal-spe-
cific methylation imprint at the human H19 locus. Hum Mol Genet
1996;5:1155–61.
Vol. 94, No. 2, July 2010




















