
Interaction of divalent cations with peptide fragments from Parkinsons disease genes
Upload
independentCategory
view
2download
0
Gamete Research 15285-294 (1986)
Requirement of Monovalent Cations in the Acrosome Reaction of Guinea Pig Spermatozoa Amitabha Bhattacharyya, Eduardo R.S. Roldan, and Ryuzo Yanagimachi
Department of Anatomy and Reproductive Biology, University of Hawaii School of Medicine, Honolulu
The majority of the spermatozoa precapacitated in Ca2+-free medium underwent the acrosome raction rapidly when they were transferred to Ca2+-containing medium. The presence of Na+ and Ca2+ in the medium was essential for the acrosome reaction. The vast majority of spermatozoa failed to undergo the reaction in Ca2+ medium lacking monovalent ions, although they remained motile. At the concentration of 140 mM, Na+, K’, Rbt , and Cs’ all supported the reaction at the maximum level, but at 50 mM the latter three ions were not as effective as Naf . Lif was least effective in supporting the reaction. Virtually no acrosome reactions took place when precapacitated spermatozoa were first exposed to Naf medium (no Ca”) and then to Ca” medium (no Na+). On the other hand, a considerably higher proportion of spermatozoa acrosome reacted when they were exposed to these media in the reverse order. The most efficient acrosome reactions took place when the medium contained both a monovalent ion (Na+) and Ca2’ simultane- ously. Possible mechanisms by which monovalent and divalent cations participate in the acrosome reaction are discussed.
Key words: acrosome reaction, spermatozoa, monovalent cation
INTRODUCTION
Mammalian spermatozoa must undergo the acrosome reaction before effecting normal fertilization. It has been well established that extracellular cations regulate or control the initiation and completion of the acrosome reaction. In the absence of extracellular Ca2’, for instance, spermatozoa of all mammalian species so far studied do not initiate their acrosome reactions [Yangimachi and Usui, 1974; Talbot et al, 1976; Meizel, 1978; Yanagimachi, 1981 ; Fraser, 19841. The necessity of extracellular Naf for the acrosome reaction in mammalian (guinea pig) spermatozoa was reported first by Hyne et a1 [ 19841 and confirmed by Murphy et al [ 19861. We report here that it is a monovalent cation in the medium, not Na+ per se, that is essential for the acrosome reaction.
Received April 16, 1986; accepted June 11, 1986
Address reprint requests to Dr. R. Yanagimachi, Department of Anatomy and Reproductive Biology, University of Hawaii School of Medicine, 1951 East-West Road, Honolulu, HI 96822.
0 1986 Alan R. Liss, Inc.

286 Bhattacharyya, Roldan, and Yanagimachi
MATERIALS AND METHODS Reagents
Inorganic salts were purchased from either Mallinckrodt Chemical (St. Louis, MO) or Baker Chemical (Philadelphia, PA). D-glucose was purchased from Baker Chemical; Na-lactate, Na-pyruvate, sucrose, and Tris (Trizma-base and -HC1) from Sigma Chemical Co. (St. Louis, MO); lysolecithin from Avanti Biochemicals (Bir- mingham, AL); bovine serum albumin (BSA, fraction V, fatty acid-poor) from Calbiochem (La Jolla, CA); Ficoll-400 from Pharmacia (Piscataway, NJ); and mineral oil from Squibb and Sons (Princeton, NJ).
Composition of Media The composition of the media used are shown in Table 1 .The regular, modified
Tyrode's medium (mT) [Fleming and Yanagimachi, 19811 contained 144.8 mM Na' and 2.7 mM K+. Test media, mTs, were prepared by first omitting all of the Na+ and K+ salts, then incorporating known amounts of Na+, K+, Lif, Rb+, or Cs+ (as chloride salts). Tris (10 mM) was substituted for NaHC03. No antibiotics were included to avoid contamination of media with unidentified ions. All the media were prepared immediately before use.
Preparation of Precapacitated Spermatozoa Precapacitated guinea pig spermatozoa that were ready for the acrosome reac-
tion were prepared according to Fleming and Yanagimachi [1981] and Murphy et a1 [1986]. Briefly, mature spermatozoa collected from both the vas deferens and the distal segment of the cauda epididymis were suspended, at the concentration of approximately 4-7 X lo6/&, in 3 ml of Ca2+-free mT medium containing a low concentration of lysolecithin (85 pg/ml) . When the sperm suspension was incubated for 1 to 2 hr under mineral oil (37°C; gas phase, pure air), the majority of the spermatozoa were capacitated. A synchronous acrosome reaction could be induced by transferring these precapacitated spermatozoa to Ca2 +-containing regular media.
TABLE 1. ComDositions of Media Used
Concentration (mM) of components in Components Regular mT Caz+-free mT Test mTsa
NaCl 108.76 111.76 0-140 KCI 2.70 2.70 0-140 CaC12 2.00 - 2.00 MgC12.6H20 0.49 0.49 0.49 NaHC03 25.07 25.07 - D-glucose 5.56 5.56 Na-lactate 10.00 10.00 -
Na-pyruvate 1 .oo 1 .OO Sucrose - - 0-300 Tris base/HCl - - 10.00 BSA (mg/ml) 3 3 3 pH (under pure air) 7.8-7.9 7.8-7.9 7.8-8.0
-
-
Osmolality (mOsmol) 308 308 308
'Various test mTs were prepared by varying concentrations of NaCl and KCl, or replacing these salts with LiCl, RbC1, or CsC1. The osmolality of all test mTs was maintained constant by adjusting the amount of sucrose added to the medium.

Acrosome Reaction 287
Examination of the Effects of Monovalent Cations on the Acrosome Reaction of Precapacitated Spermatozoa
Precapacitated spermatozoa prepared as described above were washed twice in an isotonic (300 mM) sucrose solution by centrifugation (270g for 5 min each) and suspended in fresh isotonic sucrose solution at the concentration of approximately 4- 5 X 107/d. An aliquot (10 or 100 p1) of this sperm suspension was mixed with 300 p1 or 3 ml or a test medium that had been previously placed under mineral oil (37°C) in a plastic petri dish (10 X 30 mm; Falcon Plastics, Oxnard, CA). The final concentration of spermatozoa in each test medium was about 1.5 X 106/ml. After 10 to 15 rnin of further incubation at 37"C, spermatozoa were examined to determine the overall percentage of motile spermatozoa and the quality of sperm movement (graded 1 to 3) according to Roldan et al [ 19861. The percentage of acrosome-reacted spermatozoa in the entire sperm population was calculated as described by Murphy and Yanagimachi [1984] and Roldan et a1 [1986], and data were analyzed by the Student's t test.
Experiments To Reveal Whether the Concomitant Presence of Monovalent Cations (Na+ and K+) and Divalent Cations (Ca') in the Medium is Necessary for the Acrosome Reaction
When precapacitated spermatozoa in Ca2+-free medium are transferred to the regular medium containing Na+, K+, and Ca2+, the majority undergo their acrosome reaction promptly. Experiments were performed to determine whether both monova- lent and divalent cations must be present in the medium simultaneously to support the acrosome reaction. In the first series of experiments, precapacitated spermatozoa were washed once in isotonic sucrose by centrifugation (270g for 5 min) then incubated at 37°C for 10-15 min in Ca2+-containing, Na+/K+-free medium. After determining motility and the acrosome reaction, spermatozoa were washed again with isotonic sucrose and transferred to Ca2+-free, Na+/K+-containing medium (37 "C). After 10 to 15 min, spermatozoa were reexamined to evaluate motility and the acrosome reaction. The second series of experiments was performed in a similar manner except that the washed precapacitated spermatozoa were first exposed to Ca2+-free (Na+/K+-containing) medium and then to Ca2+-containing (Naf/Kf-free) medium. Precapacitated spermatozoa exposed twice to the regular medium served as the control.
Examination of Fertilizing Capacity of Acrosome-Reacted Spermatozoa To determine the fertilizing capacity of spermatozoa that had acrosome reacted
in media containing different monovalent cations, it was necessary to separate acro- some-reacted spermatozoa from acrosome-unreacted ones. This was done by using the method of Fleming and Yanagimachi [1982], with some modifications. The fertilizing capacity of the spermatozoa that had undergone the acrosome reaction in Ca2+-containing K+-rich (140 mM) medium, for instance, was evaluated as follows. Three milliliters of sperm suspension in the K+-rich medium containing both acro- some-reacted and acrosome-unreacted spermatozoa were centrifuged (270g for 5 rnin), and the pelleted spermatozoa were resuspended in 1 ml of Ca2+-free mT medium with 4% (w/v) Ficoll-400. This viscous sperm suspension was placed in the bottom of a test tube (15 x 50 mm, round bottomed) and gently overlaid with 2 ml of Ca2+-free mT without Ficoll. The tube, sealed with Parafilm, was inclined at a
288 Bhattacharyya, Roldan, and Yanagimachi
30" angle and incubated at 37°C. After 45 min of incubation, the tube was returned to the upright position and the upper 1 ml of the medium was carefully collected and transferred into a centrifuge tube. All spermatozoa collected in this manner were acrosome reacted. As guinea pig spermatozoa do not undergo the acrosome reaction in Ca2+-free medium [Yanagimachi and Usui, 19741, the acrosome-reacted sperma- tozoa collected in the Ca2+-free medium must have undergone the reaction while in the K+-rich, Ca2+-containing medium. The 1 ml sperm suspension in the tube was diluted with 10-15 ml regular mT medium and centrifuged (270g for 5 min). Pelleted spermatozoa were resuspended in the regular mT medium at the concentration of approximately 4-6 X lo4 sperm/ml. An aliquot (300 pl) of this sperm suspension was placed in a plastic petri dish (30 X 10 mm) and covered with mineral oil. Twenty to 30 zona-free hamster eggs, prepared as described by Yanagimachi [ 19721, were introduced into the sperm suspension. After 1 hr of incubation at 37°C (gas phase, pure air), eggs were examined for fertilization according to Yanagimachi [ 19721.
RESULTS Effects of Na+ and Other Monovalent Cations on the Acrosome Reaction
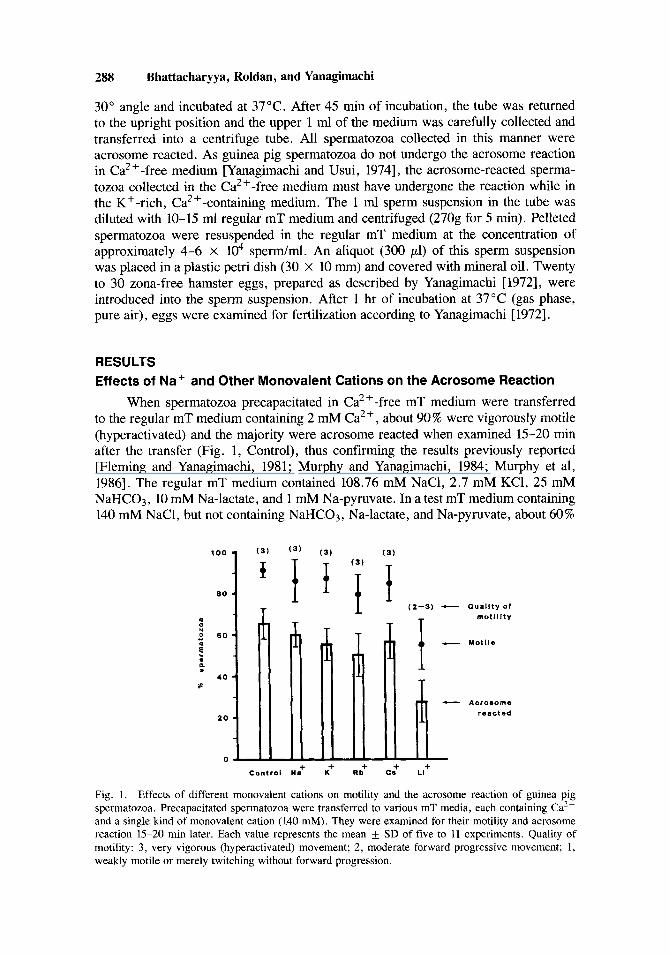
When spermatozoa precapacitated in Ca2+-free mT medium were transferred to the regular mT medium containing 2 mM Ca2+, about 90% were vigorously motile (hyperactivated) and the majority were acrosome reacted when examined 15-20 rnin after the transfer (Fig. 1, Control), thus confirming the results previously reported [Fleming and Yanagimachi, 1981; Murphy and Yanagimachi, 1984; Murphy et al, 19861. The regular mT medium contained 108.76 mM NaCl, 2.7 mM KC1, 25 mM NaHC03, 10 mM Na-lactate, and 1 mM Na-pyruvate. In a test mT medium containing 140 mM NaCl, but not containing NaHC03, Na-lactate, and Na-pyruvate, about 60%
Qual i ty O f
motll I t y
Yotl le
Acrosome reacted
+ + + Control Ha K R b C.+ Li+
Fig. 1. Effects of different monovalent cations on motility and the acrosome reaction of guinea pig spermatozoa. Precapacitated spermatozoa were transferred to various mT media, each containing Ca2+ and a single kind of monovalent cation (140 mM). They were examined for their motility and acrosome reaction 15-20 min later. Each value represents the mean + SD of five to 11 experiments. Quality of motility: 3, very vigorous (hyperactivated) movement; 2, moderate forward progressive movement; 1 , weakly motile or merely twitching without forward progression.

Acrosome Reaction 289
of spermatozoa underwent the acrosome reaction (Fig. 1). The mT media containing 140 mM of KCI, RbC1, or CsCl instead of NaCl all supported the acrosome reaction almost as effectively as the control medium (Fig. l), indicating that K + , Rbf , and Cs+ can replace Na' in this biological process. Li+ was not as effective as the other monovalent cations in supporting motility and the acrosome reaction of spermatozoa (P < 0.01) (Fig. 1).
Effects of Various Concentrations of a Single Monovalent Cation on the Acrosome Reaction
As stated above, K+, Cs+, and Rb' at the concentration of 140 mM were almost as effective as 140 mM Na' in supporting the acrosome reaction. To determine whether lower concentratins of these ions can support the acrosome reaction, mT media with various concentrations of Na+, K f , Cs+, and Rb+ were tested. Effects of various concentrations of Li' were also examined. The results obtained are summarized in Figure 2. In the mT medium with 2 mM Ca2+, but without any monovalent cations, about 70% of spermatozoa were motile and many were hyper- activated, but only 1.5% of spermatozoa of the entire population were acrosome reacted. The incidence of the acrosome reaction increased with increasing concentra- tion of monovalent cations. At the concentration of 140 mM, K+, Cs+, and Rb+ were almost as effective as Na' in supporting the acrosome reaction, but at 50 mM all of these ions were considerably less effective. Li+ was least effective in supporting the reaction.
Table 2 summarizes the results of experiments designed to reveal the effect of the Na+/K+ ratio of the medium on the acrosome reaction. It will be seen that high K+ concentrations relative to Na+ concentrations tend to reduce the incidence of the acrosome reaction. However, it should be noted that 140 mM K+ supports the reaction well in the absence of Na+ .
Monovalent and Divalent Cations Must Coexist in the Medium To Elicit Efficient Acrosome Reaction
Table 3 gives the results of experiments in which washed precapacitated sper- matozoa were exposed first to divalent cations (Ca2+) and then to monovalent cations (Na' plus K') (experiment B) or in the reverse order (experiment C). As can be seen from Table 3, many spermatozoa thus treated lost motility (Experiments B and C). It should be noted that the vast majority of spermatozoa were unable to acrosome react in response to Ca2+ in the absence of monovalent cations, but many of them acrosome reacted when they were transferred to Ca2+-free medium containing mon- ovalent cations (experiment B). Virtually no acrosome reaction took place when the spermatozoa were first exposed to Ca2+-free medium with monovalent cations, then transferred to Ca2+-containing medium without monovalent cations (Experiment C) .
Fertilizing Capacity of Acrosome-Reacted Spermatozoa
As stated above, precapacitated spermatozoa are able to undergo the acrosome reaction in various media, each containing both Ca2' and one of the monovalent cations (eg, 140 mM Rb+). The fertilizing capacity of spermatozoa that underwent the acrosome reaction in the presence of different monovalent cations were examined after washing them with isotonic sucrose and mixing them with zona-free hamster eggs in the regular mT medium. The results revealed that spermatozoa were able to

290 Bhattacharyya, Roldan, and Yanagimachi
N: K+
0 10 2 1 so 100 140
R t
0 3 0 1s so loo 140
CS+
loo. 131
100 140
- Quality of motility
C- Motile
o 10 as so 140
Concentration of ion (mM)
Fig. 2. Effects of various concentrations of different monovalent cations on motility and the acrosome reaction of guinea pig spermatozoa. Precapacitated spermatozoa were transferred to various media, each containing Caz+ and 0-140 mM of a single kind of monovalent cation. They were examined for their motility and acrosome reaction 15-20 min later. Each value represents the mean SD of four to 13 experiments. Quality of motility: 3 , very vigorous (hyperactivated) movement; 2 , moderate forward progression; 1 , weakly motile or merely twitching without forward progression.

Acrosome Reaction 291
TABLE 2. Effect of Na+/K+ Ratio on Motility and the Acrosome Reaction of Precapacitated Spermatozoa
Spermatozoa at 10-20 min after transfer to Ca2+-containing medium
Na+ K+ % Motile % Acrosome (mM) (mM) Na+:K+ (quality) reaction
140 - 80 f 11.7 (3) 55.8 f 5.02 137 3 1 :0.02 81 f 4.1 (3) 63.4 f 5.12a 93 47 1:0.5 73 f 10.9 (3) 43.1 f 8.29b 47 93 1:2 70 f 7.9 (3) 36.0 k 8.89b - 140 76 k 9.6 (3) 53.3 f 7.52
Results are expressed as mean f SD of five experiments using spermatozoa from five different males. aP < 0.01. bStatistically different from a (P < 0.01).
TABLE 3. Motility and Acrosome Reaction (AcR) of Precapacitated Spermatozoa Incubated Consecutively in Monovalent (Na+/K%Free and Divalent (Ca2%Free Media
Test mTs with
Incubation media Sperm after 10-15 min
of incubation (test mTs) containinga 1st incubation 2nd incubation
1st 2nd % Motile % Motile Exp. Naf K+ Ca2' Na' K+ Ca2+ (quality) % AcR (quality) % AcR
A + + + + + + 82 f. 2.7 63 f 3.9 79 +_ 1.9 61 f 2.2
B
C + + - - + 63 f 8.6 0 41 f 10.9 1 + 1.0
(3) (3) - + + + - 62 f 11.8 4 f 2.7 41 f 8.5 23 f 11.2 -
(2-3) (2-3) -
Results are mean f SD of six experiments using spermatozoa of four different males. "Concentrations of Na+, K + , and Ca2+ were 111.7 mM, 2.7 mM, and 2.0 mM, respectively.
TABLE 4. Fertilizing Capacity of Spermatozoa Acrosome Reacted in the Presence of Various Monovalent Cations
Monovalent cations (concentrations, mM) present in test mT during acrosome reaction
Na+ (108) and K + (2.7) Li+ (140) only Na+ (140) only K+ (140) only Rb+ (140) only Cs+ (140) only
No. of eggs inseminated
(No. of experiments)
59 (3) 63 (3) 78 (3)
100 (3) 71 (3)
70 (3)
Mean No. f SD of decondensing
No. (%)of sperm heads in in each fertilized egg eggs fertilized
59 (loo) 12 + 6.9 61 (97) 4 + 3.4 75 (96) 6 + 5.5 64 (91) 4 f. 3.0 93 (93) 4 f 2.3 70 (98) 7 f 4.9
fertilize the majority of the eggs regardless of the type of monovalent cation present in media during the acrosome reaction (Table 4).
DISCUSSION
Most of the media used for inducing capacitation and the acrosome reaction of guinea pig spermatozoa contain 130 mM or more of Na' [eg, Yanagimachi, 1972;

292 Bhattacharyya, Roldan, and Yanagimachi
Barros, 1974; Talbot et al, 1976; Green, 1978; Fleming and Yanagimachi, 1981; Hyne et al, 19841. According to Hyne et a1 [1984], 125 mM or more of Na+ in the medium is required for an efficient acrosome reaction of the precapacitated guinea pig spermatozoa; in media containing less than 100 mM Na', virtually no acrosome reaction takes place. Although we have confirmed the report by Hyne et a1 that spermatozoa fail to undergo the reaction in Na+-free media, we have observed 40- 50% of spermatozoa undergoing the reaction in the media containing only 40-50 mM Na+ [Murphy et al, 19861 (Fig. 2). This discrepancy could be due to differences in 1) the methods for capacitating spermatozoa and/or 2) the composition of the media used.
According to Hyne et a1 [1984], about 60% of precapacitated guinea pig spermatozoa undergo the acrosome reaction in a simple medium containing only 150 mM Na+, 5 mM Ca2+, 1 mM Mg2+, and 10 mM Tris (pH 7.8). In a similar medium with 150 mM K+ instead of 150 mM Na', about 44% of spermatozoa acrosome react, indicating that K+ can substitute for Na+in supporting the acrosome reaction. In the present study we have found that 140 mM K + , Cs+, and Rb+ are as effective as 140 mM Na+ in supporting the reaction (see Fig. 1). Although these ions are not as effective as Nat at lower concentrations (Fig. 2), it is obvious that all of these monovalent cations have the ability to support the acrosome reaction.
The regular mT medium (Table 1) that supports the maximum level of acrosome reactions of guinea pig spermatozoa contains 144.8 mM Na' and 2.7 mM K+ [Fleming and Yanagimachi, 1981; Murphy and Yanagimachi, 1984; Murphy et al, 1986; Roldan et al, 19861. Since 2.7 mM K + by itself is not sufficient to support the acrosome reaction (Fig. 2), Na+ must be the major monovalent cation in the regular medium that renders the spermatozoa capable of undergoing the reaction. Although it is obvious that Na+ plays an essential role in the acrosome reaction, the mechanisms of its action are not fully understood. That Na' is not just acting by means of ionic and osmotic strength is evidenced by the inability of choline chloride and sucrose to support the acrosome reaction [Murphy et al, 19861. Hyne et a1 [ 19841 have proposed that prior to the acrosome reaction the spermatozoon increases its intracellular Na+ (because of a reduction in the activity of the plasma membrane Na+,K+-ATPase). Increased intracellular Na+ reverses the direction of Na+/Ca2 + antiporter of the membrane, causing an influx of extracellular Ca2+. Elevated intracellular Ca2+ activates a series of events leading to the acrosome reaction.
Recently, Usui and Yanagimachi [ 19861 have demonstrated, by a cytochemical technique, the presence of an ATPase on the outer aspect of the outer acrosomal membrane and the inner aspect of the overlying plasma membrane of guinea pig spermatozoa. This ATPase, like the Na', K+-ATPase of the erythrocyte plasma membrane [Dowben, 19691, is activated by Mg2+ and inhibited by Ca2+. According to Usui and Yanagimachi [ 19861, the ATPase retains its activity during capacitation of spermatozoa in Ca2+-free medium containing both Na+ and K+. It loses its activity very rapidly when capacitated spermatozoa are exposed to Ca2+, perhaps even before fusion begins between the plasma and outer acrosomal membranes. Although the biochemical and physiological properties of this ATPase are yet to be studied, the enzyme associated with the plasma membrane could be the enzyme that is responsible for pumping Na+ out of (and K+ into) the cell during capacitation of spermatozoa. According to Hyne et a1 [1985], intracellular concentrations of Na+ and K + of guinea pig spermatozoa increase (approximately twofold) and decrease

Acrosome Reaction 293
(approximately fivefold), respectively, during a 2 hr incubation in a K+ and Ca2+- free medium. This is probably due to a decrease in the Na+/K+ pump activity because of the lack of extracellular K+ [Hyne et al, 19841. Although we did not measure ATPase activity or intracellular concentrations of Na' and K + directly, it is unlikely that the concentrations of intracellular Na+ and K+ changed prior to the acrosome reaction as drastically as reported by Hyne et a1 [1984], because the capacitation medium we used contained both Na+ and K', and we incubated sper- matozoa for only 1 hr for capacitation. In the hamster, Na+,K+-ATPase activity increases during capacitation in media containing both Na+ and K+ [Mrsny et al, 19841.
As reported previously [Murphy et al, 19861 and confirmed in this study, both Na+ and Ca2+ must be in the medium to ensure the acrosome reaction. Which of these ions acts on spermatozoa first? To answer this question we incubated capacitated spermatozoa first in Ca2+-containing, monovalent-free medium and then in Na+- containing, Ca2+-free medium and vice versa. According to our results (Table 3), Ca2+ apparently enters first. We tentatively propose that Ca2+ passing through the plasma membrane of capacitated spermatozoa inactivates a membrane-bound Na+ ,K+- ATPase. This inactivated ATPase would no longer pump Na' out, resulting in a rapid accumulation of intracellular Na+ that in turn would cause H+ efflux via a plasma membrane-associated Na+/H+ antiporter. As suggested by Hyne et a1 [1984], an increase in the intracellular Na+ concentration could reverse a plasma membrane- associated Na+/Ca2+ antiporter, allowing more Ca2+ to enter the spermatozoon. An increase in both intracellular pH and Ca2+ would induce a series of intracellular events leading to the acrosome reaction [Meizel, 1978, 1984; Yanagimachi, 19811. Na+ , the most abundant monovalent cation in physiological media, obviously plays crucial roles in the acrosome reaction under normal conditions. It should be noted that other monovalent cations, particularly K+, Rb+, and Cs', can substitute for Na+. In other words, Na+ functions as a monovalent cation, not as Na+ per se, at least in the acrosome reaction of guinea pig spermatozoa.
According to Mrsny and Meizel [ 19811 and Fraser [ 19831, hamster and mouse spermatozoa are unable to initiate their acrosome reactions in Na+-rich (150- 160 mM) media unless the media contain 2 mM or more of K+. These spermatozoa are apparently capable of capacitation in K+-free or K'-deficient media. It is the acro- some reaction that requires extracellular K+ [Mrsny and Meizel, 1981; Fraser, 19831. K+ must play some specific roles in the acrosome reaction of hamster and mouse spermatozoa. In the guinea pig, extracellular K+ is not essential for the acrosome reaction [Barros, 1974; Rogers et al, 1981; Hyne et al, 19841, although coexistence of Na+ and K+ seems to be beneficial for the reaction [Murphy et al, 19861 (Table 2). Apparently, K+ and Na+ act similarly or synergically in some biological systems and act quite differently in others.
ACKNOWLEDGMENTS
This study was supported by a grant from the National Institute of Child Health and Human Development (HD-03402). A.B. and E.R.S.R. are recipients of postdoc- toral fellowships from the Rockefeller Foundation. We thank Mr. T. Timothy Smith for his assistance in the preparation of the manuscript.

294 Bhattacharyya, Roldan, and Yanagimachi
REFERENCES
Barros C (1974): Capacitation of mammalian spermatozoa. In Coutinho EM, Fuchs F (eds): “Physiology and Genetics of Reproduction.” New York: Plenum Press, Part B, pp 3-24.
Dowben RM (1969): Composition and structure of membranes. In Dowben RM fed): “Biological Membranes.” Boston: Little, Brown and Co, pp 1-38.
Fleming AD, Yanagimachi R (1981): Effects of various lipids on the acrosome reaction and fertilizing capacity of guinea pig spermatozoa with special reference to the possible involvement of lyso- phospholipids in the acrosome reaction. Gamete Res 4:253-273.
Fleming AD, Yanagimachi R (1982): Fertile life of acrosome-reacted guinea pig spermatozoa. J Exp
Fraser LR (1983): Potassium ions modulate expression of mouse sperm fertilizing ability, acrosome reaction and hyperactivated motility in vitro. J Reprod Fertil69:539-553.
Fraser LR (1984): Mechanisms controlling mammalian fertilization. In Clarke JR (ed): “Oxford Reviews of Reproductive Biology.” Oxford: Clarendon Press, Vol6, pp 174-225.
Green DPL (1978): The induction of the acrosome reaction in guinea-pig sperm by the divalent metal cation ionophore A 23187. J Cell Sci 32:137-151.
Hyne RV, Edwards KP, Lopata A, Smith JD (1985): Changes in guinea pig sperm intracellular sodium and potassium content during capacitation and treatment with monovalent ionophores. Gamete Res 12:65-73.
Hyne RV, Higginson RE, Kohlman D, Lopata A (1984): Sodium requirement for capacitation and membrane fusion during the guinea-pig sperm acrosome reaction. J Reprod Fertil 70:83-94.
Meizel S (1978): The mammalian sperm acrosome reaction, a biochemical approach. In Johnson MH (ed): “Development in Mammals.” Amsterdam: ElseviedNorth-Holland Biomedical Press, Vol
Meizel S (1984): The importance of hydrolytic enzymes to an exocytotic event, the mammalian sperm acrosome reaction. Biol Rev 59: 125-157.
Mrsny RJ, Meizel S (1981): Potassium ion influx and Na+,K+-ATPase activity are required for the hamster sperm acrosome reaction. J Cell Biol91:77-82.
Mrsny RJ, Siiteri JE, Meizel S (1984): Hamster sperm Na+,K+-adenosine triphosphatase: Increased activity during capacitation in vitro and its relationship to cyclic nucleotides. Biol Reprod 30:573- 584.
Murphy SJ, Roldan ERS, Yanagimachi R (1986): Effects of extracellular cations and energy substrates on the acrosome reaction of precapacitated guinea pig spermatozoa. Gamete Res 14: 1-10.
Murphy SJ, Yanagimachi, R (1984): The pH dependence of motility and the acrosome reaction of guinea pig spermatozoa. Gamete Res 10: 1-8.
Rogers BJ, Ueno M, Yanagimachi R (1981): Fertilization by guinea pig spermatozoa requires potassium ions. Biol Reprod 25:639-648.
Roldan ERS, Shibata S, Yanagimachi R (1986): Effect of Ca2+ channel antagonists on the acrosome reaction of guinea pig and golden hamster spermatozoa. Gamete Res 13:281-292.
Talbot P, Summers RG, Hylander BL, Keough EM, Franklin LE (1976): The role of calcium in the acrosome reaction: An analysis using ionophore A23187. J Exp Zoo1 198:383-392.
Usui N, Yanagimachi R (1986): Cytochemical localization of membrane-bound Mg2+-dependent ATPase activity in guinea pig sperm head before and during the acrosome reaction. Gamete Res 13:271- 280.
Yanagimachi R (1972): Penetration of guinea-pig spermatozoa into hamster eggs in vitro. J Reprod Fertil
Yanagimachi R (1981): Mechanisms of fertilization in mammals. In Mastroianni L, Biggers JD (eds):
Yanagimachi R, Usui N (1974): Calcium dependence of the acrosome reaction and activation of guinea
Zoo1 220:109-115.
3, pp 1-64.
28 :477-480.
“Fertilization and Embryonic Development in Vitro.” New York: Plenum Press, pp 81-182.
pig spermatozoa. Exp Cell Res 89:161-174.
Copyright © 2022 FDOKUMEN




















