
Interaction of divalent cations with peptide fragments from Parkinsons disease genes
12
1477-9226(2013)42:17;1-M ISSN 1477-9226 Dalton Transactions An international journal of inorganic chemistry www.rsc.org/dalton Volume 42 | Number 17 | 7 May 2013 | Pages 5945–6236 COVER ARTICLE Remelli, Zoroddu et al. Interaction of divalent cations with peptide fragments from Parkinson’s disease genes Themed issue: Chemistry and applications of metal complexes
Transcript of Interaction of divalent cations with peptide fragments from Parkinsons disease genes

1477-9226(2013)42:17;1-M
ISSN 1477-9226
DaltonTransactionsAn international journal of inorganic chemistry
www.rsc.org/dalton Volume 42 | Number 17 | 7 May 2013 | Pages 5945–6236
COVER ARTICLERemelli, Zoroddu et al. Interaction of divalent cations with peptide fragments from Parkinson’s disease genes
Themed issue: Chemistry and applications of metal complexes

DaltonTransactions
PAPER
Cite this: Dalton Trans., 2013, 42, 5964
Received 23rd September 2012,Accepted 12th November 2012
DOI: 10.1039/c2dt32222f
www.rsc.org/dalton
Interaction of divalent cations with peptide fragmentsfrom Parkinson’s disease genes†
Maurizio Remelli,*a Massimiliano Peana,b Serenella Medici,b Lucia Gemma Delogub
and Maria Antonietta Zoroddu*b
Protected Ac-PDEKHEL-NH2 (PK9-H) and Ac-FCGDGANDCG-NH2 (PK9-C) peptide fragments correspond-
ing to sequences from residues 1165 to 1171 and 1184 to 1193, respectively, in the Park9 encoded
protein from Parkinson’s disease gene were tested for their protonation and complex formation capabili-
ties with Cu(II), Zn(II) and Mn(II) ions by potentiometric and UV-Vis measurements. The effects of peptide
titration with the metal ions have been followed by mono- and bi-dimensional NMR spectroscopy in
order to support the potentiometric results and to understand the details of metal binding. Only mono-
nuclear complexes have been evidenced for all the checked metal ions with PK9-H peptide. Mononuclear
and bis-complexes with PK9-C peptide have been evidenced with Cu(II) and Zn(II) metal ions. From the
dissociation-constants and pM values obtained for the binary competition diagrams for the systems con-
taining Cu(II), Zn(II) or Mn(II) and the two ligands, the Cu(II) ion is able to bind more efficiently than Zn(II)
and Mn(II) metal ions to both ligands.
Introduction
A connection between genetic and environmental causes ofParkinson’s disease (PD) has been recently reported.1–6 Thereis considerable evidence that excess of environmental andoccupational exposure to Mn(II)7–10 (i.e. in miners and welders)induces symptoms that resemble Parkinson’s disease, calledParkinsonism or manganism,7,11 by attacking the centralnervous system. It has been reported that a human PD gene,Park9 (a member of the P-type 5 ATPase family), and its homo-logue in yeast, YPk9, can prevent manganese induced PD andprotect neurons and cells from manganese poisoning besidesfrom α-synuclein’s toxicity.4 These genes encode a proteinwhich, considering its richness in coordinating residues, lookslike a metal transporter protein.12
YPk9 protein is localized in the cell vacuole membrane, abag which captures toxins, possibly taking in manganese andsequestering it for detoxification.4
Following our interest in the study of metal ions binding torelevant fragments in proteins,13–17 we have decided to verifythe possibility of metal binding to some simplified portions ofPark9 and YPk9. From the inspection within the protein
amino acid sequence, we have found very promising sites formetal interaction and, excluding the membrane-crossing andthe alpha-helical rich segments not easily accessible by metals,we focused on the highly conserved binding motifs betweenPark9 and YPk9. We start our study from some of its frag-ments, located from 1165 to 1171 and from 1184 to 1193 resi-dues in the protein, in order to have insight into the possibleregions that could be implicated in ion binding.
Here we report on the structurally accessible PDEKHEL(PK9-H) and FCGDGANDCG (PK9-C) protein fragments; theycontain histidine and cysteine residues that are surrounded byamino acids bearing coordinating side chains, such as aspar-tate and glutamate residues which can be relevant in thehomeostasis, transport or detoxification of manganeseions.18–20
In the present paper, formation equilibria of Cu(II), Zn(II)and Mn(II) complexes with the protected peptide models Ac-PDEKHEL-NH2 and Ac-FCGDGANDCG-NH2 have been investi-gated. The study has been carried out in aqueous solution andin a wide pH range, at I = 0.1 M and T = 298.2 K. Protonationand complex-formation constants have been potentiometri-cally determined; complex-formation models and species stoi-chiometry have been carefully checked by means of UV-Visabsorption spectrophotometry.
The effects of peptide titration with Cu(II), Zn(II) and Mn(II)have been followed by mono- and bi-dimensional NMR spec-troscopy in order to support the potentiometric results and tohave more details about the metal binding sites, at differentpH values and at different ligand to metal molar ratios.
†Electronic supplementary information (ESI) available. See DOI: 10.1039/c2dt32222f
aDepartment of Chemical and Pharmaceutical Sciences, University of Ferrara,
Ferrara, Italy. E-mail: [email protected]; Tel: +39 0532 455150bDepartment of Chemistry and Pharmacy, University of Sassari, Sassari, Italy.
E-mail: [email protected]; Tel: +39-079-229529
5964 | Dalton Trans., 2013, 42, 5964–5974 This journal is © The Royal Society of Chemistry 2013
Publ
ishe
d on
13
Nov
embe
r 20
12. D
ownl
oade
d by
Uni
vers
ita' d
egli
Stud
i di S
assa
ri o
n 03
/02/
2014
06:
08:2
8.
View Article OnlineView Journal | View Issue

ExperimentalPeptide synthesis
Peptides were chemically synthesized using solid-phase Fmoc(fluoren-9-ylmethoxycarbonyl) chemistry in an Applied Biosys-tems Synthesizer.21 Peptides were N-terminally acetylated andC-terminally amidated in order to mimic this region of YPK9within the full-length protein. Peptides were removed from theresin and deprotected before purification by reverse-phaseHPLC. Fractions were collected and analyzed by MALDI-TOFMS. Fractions containing the peptide of the expected molecu-lar weight were then pooled and lyophilized.
Potentiometric measurements
Potentiometric titrations were performed with the followingapparatus: Orion EA 940 and Orion 720A pH meters (resolu-tion 0.1 mV, accuracy 0.2 mV) equipped with a combinedglass electrode (Metrohm EA125); Hamilton MicroLab M andMicroLab300 motor burettes (resolution 0.1 μl, accuracy 0.2 μl)equipped with Hamilton syringes (delivery volume 250 and500 μl, respectively). Constant-speed magnetic-stirring wasapplied throughout. The temperature of the titration cell waskept at 298.2 ± 0.1 K by use of a Haake F3C circulation thermo-stat. UPP grade nitrogen, previously saturated with H2O (bub-bling into a solution of KNO3 0.1 M, at 298.2 K), was blownover the test solution in order to maintain an inert atmos-phere. The electrode couple was standardized on the pH =−log cH+ scale by titration of HNO3 (0.01 M) with standardKOH at 298.2 K and I = 0.1 M (KNO3). Aliquots (2 ml) of thesample solution, containing suitable amounts of metal,ligand, HCl and KCl, were first titrated with standard KOH andthen back-titrated with standard HCl, without opening thereaction vessel, in order to check the reversibility of the metal-complex formation reactions. The concentration rangeemployed for the metal ions was 0.6–1.0 mM with a metal–ligand ratio ranging from 1 : 1 to 1 : 3. The Hyperquad 2006program22 was used for stability constant calculations andtheir standard deviations, which represent random errors only.The speciation and competition diagrams were plotted withthe Hyss 2006 program.23
Reported log β values refer to the overall equilibrium:
pMþ qLþ rH ⇄ MpLqHr ð1Þwhile log Kstep values refer to the protonation process:
MpLqHr�1 þH ⇄ MpLqHr ð2Þ(charges omitted; p might also be 0 and r can be negative). Ifthe dissociation of a proton is taken into account, the pKstep
value is considered.The stability constants of complexes with different stoichi-
ometry and protonation degree cannot be directly compared.In the case of 1 : 1 species, an effective way to perform such acomparison is to calculate the log K* values, where K* refers tothe equilibrium:
Mþ LHn ⇄ MLHj þ ðn� jÞH ð3Þ
where n is the maximum protonation degree of the ligand andj is the protonation degree of the complex; charges are omittedfor the sake of simplicity.
In order to compare the global binding ability of ligands,under the given experimental conditions (e.g. at physiologicalpH), other parameters can be employed. Among them, themost popular are (i) the dissociation constant (Kd)
24 and (ii)the pM value.25 The first one refers to a hypothetical overallequilibrium:
ML ⇄ Mþ L ð4Þwhere the protonation degree of the ligand (and the complex)is not explicitly considered. The second parameter is the nega-tive logarithm of the concentration of the free metal ion (pM),in the presence of the ligand, under normalized experimentalconditions, i.e. [L]total = 10−5 M and [M]total = 10−6 M, at pH7.4. Finally, the overall binding ability of different ligandstowards the same metal ion (or of one ligand towards differentmetals) in the whole pH range can be compared by plottingthe “competition diagrams”. These plots refer to a hypotheti-cal solution where all the ligands are present at the same timeand “compete” to bind the metal ion; they can be calculatedby means of the speciation models obtained by potentiometryand assuming that no mixed-ligand species is formed.
UV-Vis measurements
The absorption spectra were recorded with a Varian Cary 50spectrophotometer, in the 300–900 nm range; solutions wereof similar concentrations to those used in the potentiometricstudies. Absorptivities (ε, M−1 cm−1) were calculated at the pHvalue of maximum concentration of the considered species, asindicated by distribution diagrams.
NMR measurements
NMR experiments were performed on a Bruker Ascend™400 MHz spectrometer equipped with a 5 mm automatedtuning and matching broad band probe (BBFO) withz-gradients.
Samples used for NMR experiments were 2.5 mM in con-centration and dissolved in 90 : 10 (v/v) H2O–D2O solutions. AllNMR experiments were performed at 298 K in 5 mm NMRtubes. 2-D 1H–13C heteronuclear correlation spectra (HSQC)were acquired using a phase-sensitive sequence employingEcho-Antiecho-TPPI gradient selection with a heteronuclearcoupling constant JXH = 145 Hz, and shaped pulses for all 180°pulses on the f2 channel with decoupling during acquisition;sensitivity improvement and gradients in back-inept were alsoused.26–28
Relaxation delays of 2 s and 90° pulses of about 10 μs wereapplied for all experiments. Solvent suppression for 1-D andTOCSY experiments was achieved using excitation sculptingwith gradients. The spin-lock mixing time of the TOCSY experi-ment was obtained with MLEV17.29,30
1H–1H TOCSYs were performed using mixing times of60 ms. A combination of 1-D, TOCSY and HSQC experiments
Dalton Transactions Paper
This journal is © The Royal Society of Chemistry 2013 Dalton Trans., 2013, 42, 5964–5974 | 5965
Publ
ishe
d on
13
Nov
embe
r 20
12. D
ownl
oade
d by
Uni
vers
ita' d
egli
Stud
i di S
assa
ri o
n 03
/02/
2014
06:
08:2
8.
View Article Online

was employed to assign the spectra of both free and metal-bound peptides at different pH values. All NMR data were pro-cessed with TopSpin (Bruker Instruments) software and ana-lyzed by Sparky 3.1131 and MestRe Nova 6.0.2 (MestrelabResearch S.L.) programs.
Results and DiscussionProtonation equilibria
The two peptides here investigated were protected at both theirN- and C-ends. As far as PK9-H is concerned, its protonationequilibria involve, in order of basicity, the following residues:Lys-4, His-5, Glu-3,6 and Asp-2. Potentiometric data cannotdistinguish between the two Glu residues. In the case of PK9-C,two basic cysteines and two acidic Asp residues are presentin the chain; also in this case it is not possible, with potentio-metric data alone, to distinguish between the protonations oftwo amino acids of the same type. The thermodynamic para-meters of ligand protonation are reported in Table 1: they arein good agreement with the literature values of similarsystems.32
Copper(II) complexes
Both ligands form various Cu(II) mono-nuclear complexes ofdifferent stoichiometry and protonation degree. PK9-H formsonly 1 : 1 species while PK9-C forms also bis-complexes; nobinuclear species was observed in the whole pH range. Thecomplete set of complex-formation constants, along with theUV-Vis results, is reported in Table 2; the corresponding distri-bution diagrams are shown in Fig. 1. UV-Vis spectra atdifferent pH values are given as ESI (Fig. S1†).
The formation of Cu(II) complexes with PK9-H begins justbefore pH 3 with the [CuLH3]
2+ species. Most likely the metalis linked to the nitrogen of His imidazole, the only donor-sitepresent in the peptide which, in the absence of the terminalamino group, is able to coordinate the copper ion at this pH.The three protonated sites should be the carboxylic groups ofthe two Glu residues and the side-amine of Lys. Starting frompH 3.5, two protons are released, leading to the formation of[CuLH2]
+ and [CuLH] complexes: the corresponding pKstep
values (4.2 and 4.9) suggest that these protons derive from thelateral carboxylic groups of the two Glu residues, without any
interaction with the metal and thus without any change in thecopper coordination. The [CuLH] complex is the predominantspecies between pH 5 and 6.5; the wavelength of maximumabsorption, 757 nm at pH 5.75, is in excellent agreement withthe value expected for the coordination of one imidazole toCu(II) (758 nm).33
These results are in accordance with 1D and 2D NMR exper-iments carried out in the same range of pH. In Fig. 2, obtainedat pH 5 and a ligand to Cu(II) 1 : 0.025 molar ratio, it is possibleto see remarkable paramagnetic broadening and disappearingor lowering of the intensity of selected 1H–13C 2D HSQC corre-lations. The disappearance of both aromatic Hε1 and Hδ2from His-5 (inset in the figure), together with its aliphaticHβ2/3 signals, confirms the coordination through the His-5residue; the disappearance of Hβ2/3 from Asp-2 and Qγ fromGlu-3 and Glu-6 and the lowering in intensity of Hβ2/3 signalsfrom both glutamate residues suggest moreover that, at thispH and at substoichiometric amounts of the metal, the invol-vement also of the carboxylic group from Asp and Glu residuesin the coordination, with the formation of a macrochelatesystem, which is a favoured process at low metal concen-tration,34 cannot be excluded.
Starting from pH 5, two protons are released in a rapidsequence. It is well-known that, at this pH, the copper ion,
Table 1 Protonation constants at 298.2 K and I = 0.10 mol dm−3. Precision onthe last significant figure in parentheses
Species log β log K Residue
PK9-H LH2− 10.69(2) 10.69 LysLH2
− 17.50(3) 6.81 HisLH3 22.29(4) 4.79 GluLH4
+ 26.33(4) 4.04 GluLH5
2+ 29.62(4) 3.29 AspPK9-C LH3− 9.35(1) 9.35 Cys
LH22− 17.94(1) 8.59 Cys
LH3− 22.38(1) 4.44 Asp
LH4 25.83(2) 3.45 Asp
Table 2 Complex-formation constants and spectrophotometric data for Cu(II)complexes, at 298.2 K and I = 0.1 mol dm−3. Precision on the last significantfigure in parentheses
Species log β log Kstep log K* λ/nm ε/M−1 cm−1
PK9-H[CuLH3]
2+ 25.1(1) 4.2 −4.5 — —[CuLH2]
+ 20.9(1) 4.9 −8.7 — —[CuLH] 16.0(1) 6.9 −13.6 757 26[CuL]− 9.1(1) 6.2 −20.5 — —[CuLH−1]
2− 2.9(1) 8.9 −26.7 609 93[CuLH−2]
3− −6.0(1) 10.0 −35.6 594 97[CuLH−3]
4− −16.0(1) — −45.6 534 110PK9-C[CuL2H2]
4− 39.7(2) 4.7 — — —[CuL2H]5− 35.0(2) 8.3 — — —[CuL2]
6− 26.7(3) 9.3 — — —[CuL2H−1]
7− 17.4(3) — — — —[CuLH2] 26.0(2) 3.4 0.2 832 11[CuLH]− 22.6(2) 4.9 −3.2 788 13[CuL]2− 17.7(2) 10.8 −8.1 585 76[CuLH−1]
3− 6.9(3) — −18.9 564 104
Fig. 1 Representative distribution diagrams. [Cu(II)]tot = 1.0 mM; Cu(II)/L ratio =1 : 1.
Paper Dalton Transactions
5966 | Dalton Trans., 2013, 42, 5964–5974 This journal is © The Royal Society of Chemistry 2013
Publ
ishe
d on
13
Nov
embe
r 20
12. D
ownl
oade
d by
Uni
vers
ita' d
egli
Stud
i di S
assa
ri o
n 03
/02/
2014
06:
08:2
8.
View Article Online

once anchored to the peptide through a His residue, is able todisplace the amide protons.35 The corresponding pKstep values(6.9 and 6.2) are compatible with this hypothesis, suggesting acooperative effect that facilitates the coordination of thesecond amide nitrogen, once the conformation of the peptideis adapted to “wrap” the metal ion.
From NMR spectra obtained above pH 5 it is possible torecognize the involvement of N amides from the backbone ofthe peptide in the coordination to the paramagnetic metal ion.Starting from pH 6.5 the disappearance of HN of histidine andof lysine confirms the involvement of both N− amides in thecoordination.
The [CuLH−1]2− species is the predominant complex at
physiological pH; it is characterized by a wavelength ofmaximum absorption, recorded at pH 7.5, of 609 nm, inreasonably good agreement with the expected value (581 nm)33
for a {Nim, 2N−, O} coordination.
The [CuLH−1]2− species was clearly evidenced in the NMR
spectra obtained at pH 7.7, where its maximum formation wasdetermined from potentiometric measurements. In Fig. 3 thedisappearance of HN signals from His-5 and Lys-4 suggests the3N coordination. In addition, from the 1H–13C HSQC NMRspectrum (Fig. S2, ESI†), at the same pH, the almost total dis-appearance of Cβ–Hβ2/3 from Asp-2 and the probable disap-pearance of Cγ–Qγ signals from Glu-3, though in part inoverlap with Glu-6, could confirm the {Nim, 2N
−, O} chromo-phore with the involvement of a carboxylic group in the coordi-nation, either from Asp-2 or from Glu-3 residues, which couldbe in a dynamic equilibrium between them.
At basic pH, a further proton is released, leading to the for-mation of the [CuLH−2]
3− complex, the dominant species atpH 9.5. The Vis spectra clearly show, in the pH range 9–10, thecoexistence of two absorption bands: the first one is the samedescribed above for [CuLH−1]
2− species; the second one,increasing in intensity when pH is raised, is characterized by amarked blue-shift which moves the position of the maximum
to 534 nm. This value, which characterizes both [CuLH−2]3−
and [CuLH−3]4− complexes, is typical of a {Nim, 3N
−} species.In particular, the [CuLH−2]
3− complex should derive from thedeprotonation of a third amide nitrogen and its equatorialcoordination to the metal ion; the expected wavelength ofmaximum absorption for a complex of this type is 521 nm.33
NMR data obtained in the pH range 10–11 are totally inagreement with the results from potentiometric measure-ments. In fact, from the NMR spectrum recorded at pH 11 thedisappearance of Hα and Hβ signals from Glu-3 confirms afurther N amide deprotonation from the backbone of thepeptide and the direction of coordination towards theN-terminus.
From the NMR results, the differences in the chemicalshifts of aliphatic side chain protons of the Lys-4 residue(Fig. 4), together with the disappearance of Cγ–Hγ obtained atpH 10, whereas Qδ signals are still present, confirm the possibledeprotonation of the side amino-group of the lysine residue andexclude its involvement in the coordination to the metal.
In fact, as already mentioned, the formation of [CuLH−3]4−
species does not involve any spectral variations: this obser-vation, together with the corresponding pKstep value (10.0),makes reasonable, for this process, the hypothesis of thesimple deprotonation of the side amino-group of lysine. ThispKstep value is a bit lower than that of the free ligand, but spec-trophotometric data do not support any interaction with themetal ion.
Fig. 2 Selected aliphatic regions of 1H–13C HSQC NMR spectra of the PK9-Hpeptide, 2.5 mM, pH 5, T 298 K, in the absence (orange) and in the presence(blue) of 0.025 equivalents of Cu(II). The residues with major changes areorange-labeled. Insets: the aromatic protons of His-5 with total disappearanceof the signals in the bound status of peptide in the 1N coordination type withCu(II).
Fig. 3 Selected aromatic regions of 1H–1H TOCSY NMR spectra for PK9-Hpeptide, 2.5 mM, pH 7.7, T 298 K, in the absence (orange) and in the presence(blue) of 0.05 equivalents of Cu(II). Disappearing peaks are orange-labeled.Signals that experience chemical shift changes are black-labeled with the newspin system indicated by blue color and star. Green labels are for unchangedsignals.
Dalton Transactions Paper
This journal is © The Royal Society of Chemistry 2013 Dalton Trans., 2013, 42, 5964–5974 | 5967
Publ
ishe
d on
13
Nov
embe
r 20
12. D
ownl
oade
d by
Uni
vers
ita' d
egli
Stud
i di S
assa
ri o
n 03
/02/
2014
06:
08:2
8.
View Article Online

The behavior of PK9-C is rather different. At a 1 : 1 ratiowith Cu(II), PK9-C forms almost exclusively mono-complexes(Fig. 1, right): the metal ion is already completely complexedat pH 3.0 where the neutral [CuLH2] species is formed. Since(i) the donor groups available for this ligand are the side thioland carboxylic groups of Cys and Asp residues, respectively,and (ii) the affinity of Cu(II) for the latter is normally very low,it is easy to assume that the metal ion is bound here to the twodeprotonated thiol groups, while the side carboxylic groups ofthe two Asp residues are protonated; the remaining two coordi-nation positions in the equatorial plane of the complex areoccupied by water molecules (complex {2S, 2O}). The Asp resi-dues lose their carboxylic proton when pH is increased,leading to the formation of the [CuLH]− complex (pKstep = 3.4),the most abundant species in the pH range 3.5–5.0, and [CuL]2−
(pKstep = 4.9) which dominates in the alkaline range; thesepKstep values are slightly higher than the corresponding valuesof log Kstep of protonation of the free ligand, confirming thatthe carboxylic groups are not directly involved in thecoordination.
The 1H–13C HSQC NMR spectrum at physiological pH 7.4(Fig. 5) confirms the involvement of the two thiol groups inthe formation of the complex and at the same time excludesany involvement of the two residues Asp-4 and Asp-8: thesignals relating to the correlations Cβ–Qβ and Cα–Hα from
Cys-2 and Cys-9 are the only ones affected by the presence ofthe metal ion.
Finally, starting from pH 7.4 on, the [CuLH−1]3− species
begins to form. It is worth noting that the Vis absorptionmaximum drastically moves to shorter wavelengths, suggestingdramatic changes in the coordination geometry and, in par-ticular, the involvement of nitrogen donors, most likely fromthe peptide chain (Fig. S1, ESI†).
In the TOCSY NMR spectrum (Fig. 6) starting from pH 7.4on, a variation in the aromatic region related to both spinsystem HN from Asn-7, as well as HN from Cys-2 and Cys-9occurs: HN–Hα and HN–Hβ2/3 cross correlations from the Asnresidue disappear, suggesting the involvement of N− amidefrom the Asn residue in the coordination to Cu(II) ions, inagreement with the blue shift of the λmax of absorptionobserved in the visible spectrum.
When PK9-C is present in solution in excess with respect toCu(II), bis-complexes are also formed (see Fig. S3, ESI†): thecoordination begins again, at very acidic pH, with [CuLH2]species, but bis-complexes becomepredominant alreadyat pH5.The first bis-complex detected is [CuL2H2]
4− species: its for-mation constant (39.7) is significantly lower than that expectedfor two ligands bound to copper in the same fashion as in the[CuLH]− complex. A reasonable hypothesis is that this is a{3S, 1O} complex, where one thiol and one carboxylic groupare protonated. The formation of a {4S} species could be pre-vented by steric hindrance and even by the high negativecharge. It cannot be excluded that the oxygen coordinated to
Fig. 4 Aliphatic region of 1H–13C HSQC NMR spectra of the PK9-H peptide,2.5 mM, pH 11, T 298 K, in the absence (orange) and in the presence of 0.05equivalents of Cu(II) at both pH 11 (blue) and pH 10 (green). Only the cross cor-relations 1H–13C of the side chain of Lys-4 are indicated for simplicity. In orangecolor are the labels for the peaks experiencing disappearance of their signals inthe bound status and in blue color are the peaks shifting after Nεdeprotonation.
Fig. 5 Aliphatic region of the 1H–13C HSQC NMR spectrum of PK9-C peptide,2.5 mM, pH 7.4, T 298 K, in the absence (red) and in the presence (green) of 0.1equivalents of Cu(II). Disappearing peaks are red-labeled.
Paper Dalton Transactions
5968 | Dalton Trans., 2013, 42, 5964–5974 This journal is © The Royal Society of Chemistry 2013
Publ
ishe
d on
13
Nov
embe
r 20
12. D
ownl
oade
d by
Uni
vers
ita' d
egli
Stud
i di S
assa
ri o
n 03
/02/
2014
06:
08:2
8.
View Article Online

the equatorial plane belongs to the deprotonated carboxylategroup substituting a water molecule. The protonated carboxylicgroup begins to release its proton at pH about 3.5, leading tothe formation of [CuL2H]5− species: this complex, whose modeof coordination would be again {3S, 1O}, is the main species atphysiological pH and dominates between pH 5 and pH 8. ThepKstep values characterizing the next two deprotonations (8.3and 9.3) give rise to two different interpretations. It can beobserved that the first pKstep value is very close to that of onecysteine (8.6 for the free ligand): as a consequence, the firstdeprotonation could be attributed to the unbound SH groupwithout any further interaction with copper: the mode ofcoordination would still remain {3S, 1O}. Under these circum-stances, the second deprotonation could be attributed to theloss of a proton from the equatorial water molecule. Thesecond hypothesis is the following: the first deprotonationinvolves an amide nitrogen of the peptide chain with its simul-taneous coordination to copper, leading to an equatorial {3S,1N} donor-atom set: the pKstep value (8.3) is compatible withthis hypothesis. The second pKstep value (9.3) correspondsexactly to that of the second cysteine, still protonated.
The formation constant value is quite in agreement withthe values found for complexes with a cysteine–thiol groupamong donor atoms.36–38
It is important to note that no reduction of the Cu(II) ionwas observed during the titration time, as confirmed by thefact that direct (with NaOH) and reverse (with HCl) titrations(see the Experimental section) were almost perfectly super-imposable, taking into account the dilution.
From NMR experiments, although carried out by using lowconcentrations and substoichiometric amounts of Cu(II) ions,it was not possible to highlight bis-complexes.
Zinc(II) complexes
The behavior of the two ligands towards the Zn(II) ion is rathersimilar to that towards copper, described above. The complex-formation constants are reported in Table 3, while the corre-sponding distribution diagrams are shown in Fig. 7 and S4(ESI†).
Like copper, zinc forms only mononuclear complexes withPK9-H. The coordination of Zn(II) to the imidazole nitrogentakes place at pH 4.5: at this pH, all the carboxylic groups arealmost completely deprotonated, while the Lys residue is stillprotonated. The formation constant of the formed complex,[ZnLH], is two orders of magnitude lower than the correspond-ing one with copper (14.1 and 16.0, respectively), as expectedfor complexes of this type. It is not possible to say whethersome interaction between the metal ion and the carboxylategroups of the ligand takes place. Two protons are then releasedin a sequence so rapid that potentiometry records a singlestep, leading to the species with stoichiometry [ZnLH−1]
2−.Given the inability of zinc to displace amide protons, only the
Fig. 6 Selected aromatic regions of the 1H–1H TOCSY NMR spectrum of PK9-Cpeptide, 2.5 mM, pH 7.4, T 298 K, in the absence (red) and in the presence(green) of 0.1 equivalents of Cu(II). Disappearing peaks are red-labeled.
Table 3 Complex-formation constants for Zn(II) complexes, at 298.2 K and I =0.1 mol dm−3. Precision on the last significant figure in parentheses
Species log β log Kstep log K*
PK9-H[ZnLH] 14.1(1) — −15.5[ZnLH−1]
2− −1.8(1) 10.5 −31.4[ZnLH−2]
3− −12.3(1) — −41.9PK9-C[ZnL2H2]
4− 30.6(2) 5.5 —[ZnL2H]5− 25.1(1) 9.0 —[ZnL2]
6− 16.1(1) — —[ZnLH2] 20.4(1) 5.1 −5.4[ZnLH]− 15.3(1) 6.2 −10.5[ZnL]2− 9.1(1) 9.2 −16.7[ZnLH−1]
3− −0.1(1) — −25.9
Fig. 7 Representative distribution diagrams. [Zn(II)]tot = 1.0 mM; Zn(II)/L ratio =1 : 1.
Dalton Transactions Paper
This journal is © The Royal Society of Chemistry 2013 Dalton Trans., 2013, 42, 5964–5974 | 5969
Publ
ishe
d on
13
Nov
embe
r 20
12. D
ownl
oade
d by
Uni
vers
ita' d
egli
Stud
i di S
assa
ri o
n 03
/02/
2014
06:
08:2
8.
View Article Online

deprotonation of two water molecules can be suggested, withthe formation of the hydroxylated species: in this case, thecomplex is better described as [Zn(LH)(OH)2]
2−. The corre-sponding cumulative pK value (15.9) is not far from the globalformation constant for [Zn(OH)2] species (17.34)39 and the pHrange of existence of [Zn(LH)(OH)2]
2− is the same as [Zn(OH)2](see Fig. 7). Finally, starting from pH 9, an additional proton isreleased, most likely from the side amino-group of lysine, withno interaction with the metal: in fact, the corresponding pKstep
(10.50) is very close to that of the free ligand (10.69).NMR data confirm the binding of Zn(II) ion to the imida-
zole ring of histidine. In Fig. 8 and 9, in addition to the shiftsof histidine protons (HN, Hδ2, and Hε1), major shifts are thosefor HN from Glu-6 and Asn-7 together with HN from Lys-4, pre-cisely the residues that are just around the anchoring site,His-5.
While no precipitation was observed at the 1 mM concen-tration used for potentiometric measurements, in the wholepH range, it occurred in NMR experiments (where a higher,2.5 mM, concentration was employed), due to the formation of
Zn(OH)2; therefore only the free ligand could be seen in theNMR spectra at pH 9.
The speciation model provided by potentiometry for thecomplexes formed by Zn(II) with PK9-C is totally similar to thatfound with Cu(II), as far as the species stoichiometry is con-sidered. The constant values are instead much lower (morethan 7 orders of magnitude for [ZnL]2− species), as alreadyobserved for PK9-H. This is also especially evident from thecomparison of the corresponding distribution diagrams: thecomplex formation begins at pH 3 for zinc, while at the samepH value, copper is already fully complexed. Starting from thebis-complexes (formed in excess of the ligand) and taking intoaccount that the Zn(II) ion is not able to displace amideprotons, one might think that the [ZnL2]
6− complex (prevailingat basic pH) is a {4S} species. However, its formation constant(16.1) is three orders of magnitude lower than that found forthe analogous complex of Zn(II) with Ac-CGC-NH2 (18.81).40
The protonation constants of cysteine residues are very similarfor the two peptides; the chelate ring formed by Ac-CGC-NH2
is smaller, but anyhow 10 terms. Such a significant differencecould be explained by assuming that, in this case, the coordi-nation is of {3S, 1O} type, with the oxygen belonging to a car-boxylate group of an Asp residue or to a water molecule; as aconsequence, the thiol group of one cysteine would be free. Atneutral pH the protonated [ZnL2H]5− species prevails: the cor-responding log Kstep value for the protonation of [ZnL2]
6− isvery close to that of a cysteine in the free ligand, confirmingthe fact that this should be not bound to the metal. When pHis lowered, a further protonation can be observed: the corre-sponding log Kstep value (5.5) is too high to be simply ascribedto the protonation of a side carboxylic-group and a bit too low
Fig. 9 Selected aromatic regions of 1H–1H TOCSY NMR spectra of the PK9-Hpeptide, 2.5 mM, pH 7, T 298 K, in the absence (red) and in the presence (blue)of 1 equivalent of Zn(II). The signals experiencing chemical shift changes areblue-labeled.
Fig. 8 Aromatic region of 1H–13C HSQC NMR spectra of PK9-H peptide,2.5 mM, pH 7, T 298 K, in the absence (red) and in the presence (blue) of 1equivalent of Zn(II).
Paper Dalton Transactions
5970 | Dalton Trans., 2013, 42, 5964–5974 This journal is © The Royal Society of Chemistry 2013
Publ
ishe
d on
13
Nov
embe
r 20
12. D
ownl
oade
d by
Uni
vers
ita' d
egli
Stud
i di S
assa
ri o
n 03
/02/
2014
06:
08:2
8.
View Article Online
for the process of deprotonation/complexation of a cysteine. Ahypothesis which may explain this intermediate value wouldbe the presence of different structures in solution, in equili-brium with each other, having the same stoichiometry butdifferent donor atom sets, like {3S, 1O} or {2S, 2O}.
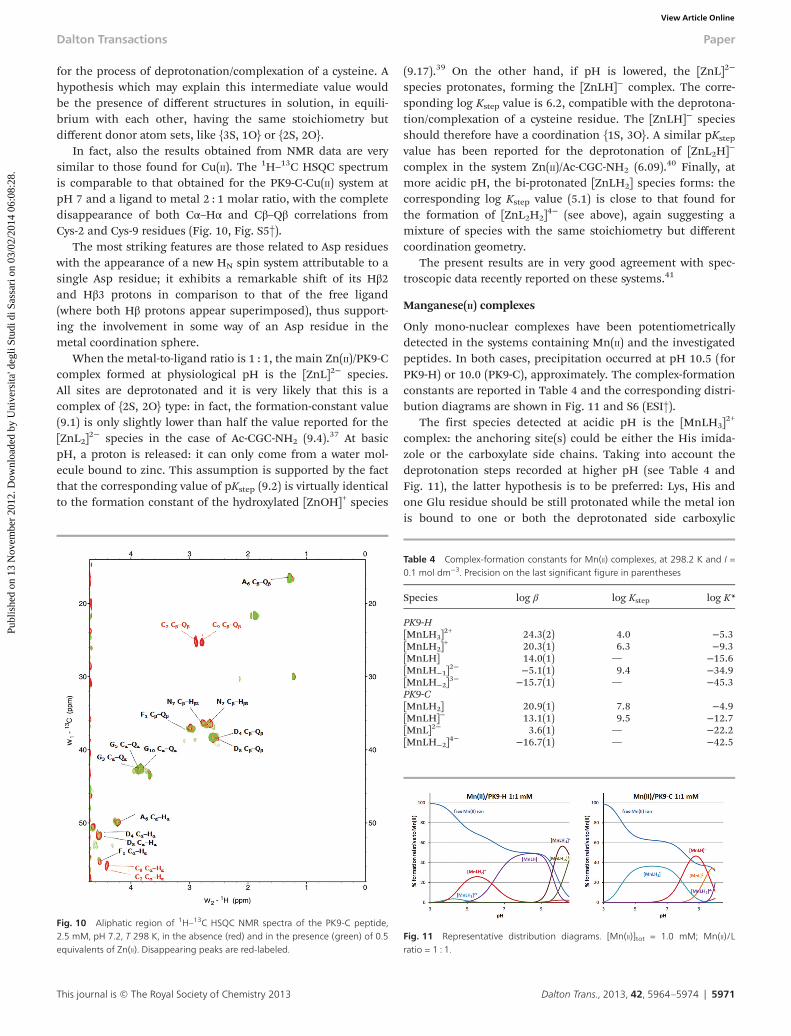
In fact, also the results obtained from NMR data are verysimilar to those found for Cu(II). The 1H–13C HSQC spectrumis comparable to that obtained for the PK9-C-Cu(II) system atpH 7 and a ligand to metal 2 : 1 molar ratio, with the completedisappearance of both Cα–Hα and Cβ–Qβ correlations fromCys-2 and Cys-9 residues (Fig. 10, Fig. S5†).
The most striking features are those related to Asp residueswith the appearance of a new HN spin system attributable to asingle Asp residue; it exhibits a remarkable shift of its Hβ2and Hβ3 protons in comparison to that of the free ligand(where both Hβ protons appear superimposed), thus support-ing the involvement in some way of an Asp residue in themetal coordination sphere.
When the metal-to-ligand ratio is 1 : 1, the main Zn(II)/PK9-Ccomplex formed at physiological pH is the [ZnL]2− species.All sites are deprotonated and it is very likely that this is acomplex of {2S, 2O} type: in fact, the formation-constant value(9.1) is only slightly lower than half the value reported for the[ZnL2]
2− species in the case of Ac-CGC-NH2 (9.4).37 At basicpH, a proton is released: it can only come from a water mol-ecule bound to zinc. This assumption is supported by the factthat the corresponding value of pKstep (9.2) is virtually identicalto the formation constant of the hydroxylated [ZnOH]+ species
(9.17).39 On the other hand, if pH is lowered, the [ZnL]2−
species protonates, forming the [ZnLH]− complex. The corre-sponding log Kstep value is 6.2, compatible with the deprotona-tion/complexation of a cysteine residue. The [ZnLH]− speciesshould therefore have a coordination {1S, 3O}. A similar pKstep
value has been reported for the deprotonation of [ZnL2H]−
complex in the system Zn(II)/Ac-CGC-NH2 (6.09).40 Finally, atmore acidic pH, the bi-protonated [ZnLH2] species forms: thecorresponding log Kstep value (5.1) is close to that found forthe formation of [ZnL2H2]
4− (see above), again suggesting amixture of species with the same stoichiometry but differentcoordination geometry.
The present results are in very good agreement with spec-troscopic data recently reported on these systems.41
Manganese(II) complexes
Only mono-nuclear complexes have been potentiometricallydetected in the systems containing Mn(II) and the investigatedpeptides. In both cases, precipitation occurred at pH 10.5 (forPK9-H) or 10.0 (PK9-C), approximately. The complex-formationconstants are reported in Table 4 and the corresponding distri-bution diagrams are shown in Fig. 11 and S6 (ESI†).
The first species detected at acidic pH is the [MnLH3]2+
complex: the anchoring site(s) could be either the His imida-zole or the carboxylate side chains. Taking into account thedeprotonation steps recorded at higher pH (see Table 4 andFig. 11), the latter hypothesis is to be preferred: Lys, His andone Glu residue should be still protonated while the metal ionis bound to one or both the deprotonated side carboxylic
Fig. 10 Aliphatic region of 1H–13C HSQC NMR spectra of the PK9-C peptide,2.5 mM, pH 7.2, T 298 K, in the absence (red) and in the presence (green) of 0.5equivalents of Zn(II). Disappearing peaks are red-labeled.
Table 4 Complex-formation constants for Mn(II) complexes, at 298.2 K and I =0.1 mol dm−3. Precision on the last significant figure in parentheses
Species log β log Kstep log K*
PK9-H[MnLH3]
2+ 24.3(2) 4.0 −5.3[MnLH2]
+ 20.3(1) 6.3 −9.3[MnLH] 14.0(1) — −15.6[MnLH−1]
2− −5.1(1) 9.4 −34.9[MnLH−2]
3− −15.7(1) — −45.3PK9-C[MnLH2] 20.9(1) 7.8 −4.9[MnLH]− 13.1(1) 9.5 −12.7[MnL]2− 3.6(1) — −22.2[MnLH−2]
4− −16.7(1) — −42.5
Fig. 11 Representative distribution diagrams. [Mn(II)]tot = 1.0 mM; Mn(II)/Lratio = 1 : 1.
Dalton Transactions Paper
This journal is © The Royal Society of Chemistry 2013 Dalton Trans., 2013, 42, 5964–5974 | 5971
Publ
ishe
d on
13
Nov
embe
r 20
12. D
ownl
oade
d by
Uni
vers
ita' d
egli
Stud
i di S
assa
ri o
n 03
/02/
2014
06:
08:2
8.
View Article Online

chains of Asp and the second Glu. The third carboxylic grouploses early its proton, to form the [MnLH2]
+ complex, the mainspecies in the pH range 4–6. The corresponding pKstep value(4.0) does not clearly indicate whether there is an interactionof this group with the metal ion or not.
From 1H–13C HSQC and TOCSY spectra (Fig. 12), obtainedat pH 5 and a ligand to metal 1 : 0.1 molar ratio, it appearsvery clearly that oxygens from carboxyl groups from Asp-2 andGlu-3 and Glu-6 bind the metal ion. Moreover the analysis ofthe aromatic region of the TOCSY spectrum (data not shown)suggests that the Glu-3 residue can be preferred by the metalcompared to the Glu-6 residue; probably, its proximity to theAsp-2 residue makes it more available for interaction with themetal or, anyway, it seems positioned closer to the coordi-nation site. In fact, while the HN signal from Glu-3 completelydisappears, the HN signal from Glu-6 is still present.
At neutral pH, till pH 9, the prevailing complex is theneutral [MnLH] species: the corresponding deprotonation stepis characterized by a pKstep value of 6.3, slightly lower than thatof the imidazole side-chain of His in the free ligand. Thisvalue suggests a coordination geometry involving the imida-zole nitrogen along with one or more carboxylic oxygensaround Mn(II), in agreement with NMR and EPR spectroscopicresults already reported for the pH range 5.5–6.8.41
From pH 9 to pH 10.5, where precipitation was observed,other deprotonation steps were detected. The correspondingpKstep values suggest the formation of hydroxylated speciesthrough either the deprotonation of water molecules bound tomanganese or the substitution of oxygen donors of carboxylicside chains with OH− ions. The side amino group of Lys(pKstep = 10.7 for the free ligand) most likely does not interactwith the metal ion and remains protonated.
From the 1H–13C HSQC spectrum, obtained at pH 9, itseems that Glu-3 and Glu-6 residues are more weakly bound athigh pH rather than at lower pH. In fact, signals fromunbound Glu residues reappear (while their Cβ–Qβ cross corre-lations totally disappeared at pH 7.8). Maybe, for some geo-metric arrangement, the Glu residue becomes again freefollowing the concomitant deprotonation of the water
molecule and its coordination to the metal. In contrast,signals from Asp-2 remain unaltered compared to those atlower pH (Fig. S7, ESI†).
As far as PK9-C is concerned, only one complex forms insignificant amounts in the whole acidic pH range, i.e. theneutral [MnLH2] species (see Fig. 11, right). Differently to theother metal ions, no bis-complex was detected in the systemMn(II)/PK9-C. This species is doubly protonated: in this pH
Fig. 12 Aliphatic region of 1H–1H TOCSY NMR spectra of the PK9-H peptide,2.5 mM, pH 5, T 298 K, in the absence (orange) and in the presence (blue) of0.1 equivalents of Mn(II). Disappearing peaks are orange-labeled.
Fig. 13 Aliphatic region of 1H–13C HSQC NMR spectra of the PK9-C peptide,2.5 mM, pH 5, T 298 K, in the absence (orange) and in the presence of 0.1equivalents of Mn(II) at pH 5 (blue) and pH 7.8 (green). Disappearing peaks areorange-labeled.
Table 5 Dissociation constants and pM values for the studied systems (seetext)
System Kd/M−1 pM
Cu(II)/PK9-H 9.94 × 10−08 7.97Cu(II)/PK9-C 8.91 × 10−19 16.69Zn(II)/PK9-H 4.61 × 10−04 6.01Zn(II)/PK9-C 4.63 × 10−09 7.35Mn(II)/PK9-H 5.69 × 10−04 6.01Mn(II)/PK9-C 8.09 × 10−04 6.01
Paper Dalton Transactions
5972 | Dalton Trans., 2013, 42, 5964–5974 This journal is © The Royal Society of Chemistry 2013
Publ
ishe
d on
13
Nov
embe
r 20
12. D
ownl
oade
d by
Uni
vers
ita' d
egli
Stud
i di S
assa
ri o
n 03
/02/
2014
06:
08:2
8.
View Article Online

range only Cys residues can be protonated and thus theanchoring sites for manganese can only be the carboxylateside-groups.
From the analysis of the 1H–13C HSQC NMR spectra(Fig. 13) carried out in the pH range 5–7.8, it is possible, inaddition to [MnLH2], where only carboxylic groups from Asp-4and -8 are involved, to detect also [MHL] species. From poten-tiometric measurements it starts to form at pH 5 on, thoughits maximum formation is seen at pH 8.8, thus supporting thedisappearance of signals from cysteine residues evidenced inthe NMR spectrum starting at pH 5 on.
In the alkaline pH range, the first thiol group loses itsproton: the corresponding pKstep value (7.8) is compatible withthe coordination of sulphur to Mn(II), as already evidenced inprevious spectroscopic investigations.41
As pH is raised, the [MnL]2− complex forms, with a pKstep
value of 9.5 which suggests the deprotonation of the secondCys residue, without any interaction with the metal. Finally, atpH higher than 9, hydroxylated species are most likely formedand precipitation was observed.
Conclusions
From the dissociation-constants and pM values (Table 5) forthe binary systems containing either Cu(II), Zn(II) or Mn(II) andone of the two ligands, it is possible to conclude that the Cu(II)ion is capable of binding both ligands more efficiently thanZn(II) and Mn(II).
The competition diagrams reported in Fig. 14 and S8 (ESI†)show that PK9-C is a stronger ligand towards both Cu(II) andZn(II) in the whole pH range (especially at acidic and neutralpH). This is certainly due to the availability of two Cys resi-dues, a more effective anchoring site than the single His con-tained in PK9-H. As far as the Mn(II) ion is concerned, due toits affinity for oxygen donors which are present in bothpeptide fragments, the two ligands show a very similar bindingability, independently of pH value.
The structurally accessible PDEKHEL and FCGDGANDCGprotein fragments, here studied, contain histidine and cysteineresidues that are surrounded by amino acids bearing coordi-nating side chains, such as aspartate and glutamate residueswhich can be relevant in the homeostasis, transport or detoxi-fication of manganese ions.18–20
It has been found that YPk9 protein, the yeast equivalent toPark9, a vacuolar protein with a possible role in sequesteringheavy metals, is localized in the vacuole that walls off toxicsubstances, keeping toxins away from other organelles, buthaving a mutation in the YPk9 gene causes problems for thisprocess in yeast and possibly in humans.
In fact, by exposing YPk9-deficient yeast cells to an excessof different metal ions such as copper, zinc or manganese todetermine which metal it might transport, it has been foundthat in the presence of manganese the YPk9-deficient yeast didnot grow as well: cells were hypersensitive to manganese.
Taking into account the presence of an extremely largenumber of coordinating residues in the whole protein and inparticular those bearing oxygen donor atoms such as Asp orGlu residues (7.6, 2.1 and 1.9% for Glu plus Asp, Cys and Hisresidues, respectively, in the human protein; 11, 1.6 and 2%for Glu plus Asp, Cys and His residues, respectively, in theyeast protein), our results could support the idea that theentire protein might be more suitable to bind manganese thancopper or zinc ions and then to be more specific to transportand uptake manganese more efficiently than the other metals.
Further studies using bigger fragments particularly rich inAsp and Glu residues may help to shed light on the possibledetoxifying role of protein and in particular on its specific rolein relation to manganese, which is an environmental riskfactor for a Parkinson’s disease-like syndrome, thus preventingtoxicity.
Acknowledgements
We would like to acknowledge the University of Ferrara (FAR2011), the CIRCMSB (Consorzio Interuniversitario di Ricercain Chimica dei Metalli nei Sistemi Biologici). This work wasalso supported by the Regione Autonoma Sardegna FSE Sar-degna 2007–2013 L.R.7/2007 “Promozione della ricerca scienti-fica e dell’innovazione tecnologica in Sardegna” programme,the L.R.7/2010 programme and the Fondazione Banco diSardegna.
References
1 F. D. Dick, P. G. De, A. Ahmadi, N. W. Scott, G. J. Prescott andJ. Bennett, et al.,Occup. Environ. Med., 2007, 64, 666–672.
Fig. 14 Representative competition diagrams for a ternary solution containing the metal ion (either Cu(II) or Zn(II) or Mn(II)) and the two ligands. The total concen-trations of the three components are equal (1.0 mM).
Dalton Transactions Paper
This journal is © The Royal Society of Chemistry 2013 Dalton Trans., 2013, 42, 5964–5974 | 5973
Publ
ishe
d on
13
Nov
embe
r 20
12. D
ownl
oade
d by
Uni
vers
ita' d
egli
Stud
i di S
assa
ri o
n 03
/02/
2014
06:
08:2
8.
View Article Online

2 A. Elbaz, J. Clavel, P. J. Rathouz, F. Moisan, J. P. Galanaudand B. Delemotte, et al., Ann. Neurol., 2009, 66, 494–504.
3 B. C. Lai, S. A. Marion, K. Teschke and J. K. Tsui, Parkinson-ism Relat. Disord., 2002, 8, 297–309.
4 A. D. Gitler, A. Chesi, M. L. Geddie, K. E. Strathearn,S. Hamamichi, K. J. Hill, K. A. Caldwell, G. A. Caldwell,A. A. Cooper, J.-C. Rochet and S. Lindquist, Nat. Genet.,2009, 41(3), 308–315.
5 K. Schmidt, D. M. Wolfe, B. Stiller and D. A. Pearce,Biochem. Biophys. Res. Commun., 2009, 383, 198–202.
6 J. P. Covy and B. I. Giasson, NeuroToxicology, 2011, 32, 622–629.
7 M. Aschner, T. R. Guilarte, J. S. Schneider and W. Zheng,Toxicol. Appl. Pharmacol., 2007, 221, 131–147.
8 C. Au, A. Benedetto and M. Aschner, NeuroToxicology, 2008,29, 569–576.
9 R. Squitti, G. Gorgone, V. Panetta, R. Lucchini, S. Bucossiand E. Albini, et al., J. Neural Transm., 2009, 116, 1281–1287.
10 D. P. Perl and C. W. Olanow, J. Neuropathol. Exp. Neurol.,2007, 66, 675–682.
11 P. Zhang, K. M. Lokuta, D. E. Turner and B. Liu, J. Neuro-chem., 2010, 112, 434–443.
12 NCBI (http://blast.ncbi.nlm.nih.gov/Blast.cgi) ReferenceSequence: NP_014934.1.
13 M. A. Zoroddu, S. Medici, M. Peana and R. Anedda, DaltonTrans., 2010, 39, 1282–1294.
14 M. A. Zoroddu, S. Medici and M. Peana, J. Inorg. Biochem.,2009, 103, 1214–1220.
15 M. A. Zoroddu, M. Peana, S. Medici and R. Anedda, DaltonTrans., 2009, 5523–5534.
16 M. A. Zoroddu, T. Kowalik-Jankowska, M. Peana, S. Mediciand H. Kozlowski, Dalton Trans., 2008, 6127–6134.
17 M. A. Zoroddu, M. Peana and S. Medici, Dalton Trans.,2007, 379–384.
18 V. C. Culotta, M. Yang and M. D. Hall, Eukaryotic Cell,2005, 4, 1159–1165.
19 T. Tanaka, A. Shinkai, Y. Bessho, T. Kumarevel andS. Yokoyama, Proteins: Struct., Funct., Bioinf., 2009, 77, 741–746.
20 J. I. Kliegman, S. L. Griner, J. D. Helmann, R. G. Brennanand A. Glasfeld, Biochemistry, 2006, 45, 3493–3505.
21 J. Meienhofer, M. Waki, E. P. Heimer, T. J. Lambros,R. C. Makofske and C. D. Chang, Int. J. Pept. Protein Res.,1979, 13, 35–42.
22 G. Gans, A. Sabatini and A. Vacca, Talanta, 1996, 43, 1739–1753.
23 L. Alderighi, P. Gans, A. Ienco, D. Peters, A. Sabatini andA. Vacca, Coord. Chem. Rev., 1999, 184, 311–318.
24 H. Kozlowski, M. Luczkowski and M. Remelli, DaltonTrans., 2010, 39, 6371–6385.
25 G. Crisponi and M. Remelli, Coord. Chem. Rev., 2008, 252,1225–1240.
26 A. G. Palmer III, J. Cavanagh, P. E. Wright and M. Rance,J. Magn. Reson., 1991, 93, 151–170.
27 L. E. Kay, P. Keifer and T. Saarinen, J. Am. Chem. Soc., 1992,114, 10663–10665.
28 J. Schleucher, M. Schwendinger, M. Sattler, P. Schmidt,O. Schedletzky, S. J. Glaser, O. W. Sorensen andC. Griesinger, J. Biomol. NMR, 1994, 4, 301–306.
29 T. L. Hwang and A. J. Shaka, J. Magn. Reson., Ser. A, 1995,112, 275–279.
30 A. Bax and D. G. Davis, J. Magn. Reson., 1985, 65, 355–360.
31 T. D. Goddard and D. G. Kneller, SPARKY 3.114, Universityof California, San Francisco, 2007.
32 L. D. Pettit and H. K. J. Powell, The IUPAC StabilityConstants Database, Royal Society of Chemistry, London, 1.
33 H. Sigel and R. B. Martin, Chem. Rev., 1982, 82, 385–426.
34 H. Kozlowski, A. Janicka-Klos, P. Stanczak, D. Valensin,G. Valensin and K. Kulon, Coord. Chem. Rev., 2008, 252,1069–1078.
35 L. D. Pettit, J. E. Gregor and H. Kozlowski, in PerspectivesOn Bioinorganic Chemistry, ed. R. W. Hay, J. R. Dilworth andK. B. Nolan, JAI Press, London, 1991, vol. 1, pp. 1–41.
36 P. S. Hallman, D. D. Perrin and A. E. Watt, Biochem. J.,1971, 121, 549–555.
37 R. N. Patel, N. Singh, R. P. Shrivastava, K. K. Shukla andP. K. Singh, Proc. Indian Acad. Sci. (Chem. Sci.), 2002, 114,115–124.
38 L. D. Pinto, P. A. L. Puppin, V. M. Behring, D. H. Flinker,A. L. Ramalho Merce, A. S. Mangrich, N. A. Rey andJ. Felcman, Inorg. Chim. Acta, 2010, 363, 2624–2630.
39 A. E. Martell and R. M. Smith, Critical Stability Constants,Plenum Press, London, New York, 1989, vol. 6.
40 K. Kulon, D. Wozniak, K. Wegner, Z. Grzonka andH. Kozlowski, J. Inorg. Biochem, 2007, 101, 1699–1706.
41 S. Medici, M. Peana, L. G. Delogu and M. A. Zoroddu,Dalton Trans., 2012, 41, 4378–4388.
Paper Dalton Transactions
5974 | Dalton Trans., 2013, 42, 5964–5974 This journal is © The Royal Society of Chemistry 2013
Publ
ishe
d on
13
Nov
embe
r 20
12. D
ownl
oade
d by
Uni
vers
ita' d
egli
Stud
i di S
assa
ri o
n 03
/02/
2014
06:
08:2
8.
View Article Online




















