
Finite Element Analysis and Design of Crest-fixed Trapezoidal ...
Upload
independentCategory
view
5download
0
Prosencephalic Neural Folds Give Rise to NeuralCrest Cells in the Australian Lungfish, Neoceratodusforsteri
MARTIN KUNDRAT1,2�, JEAN M. P. JOSS2, AND LENNART OLSSON3
1Geological Institute, Slovak Academy of Sciences, Banska Bystrica, SlovakRepublic2Department of Biological Sciences, Macquarie University, Sydney, Australia3Institut fur Spezielle Zoologie und Evolutionsbiologie mit PhyletischemMuseum, Friedrich-Schiller-Universitat Jena, Jena, Germany
ABSTRACT Here we present a fate map of the prosencephalic neural fold (PNF) for theAustralian lungfish. The experimental procedures were carried out on lungfish embryos at Kemp’sstage 24 using three different approaches. First, either medial PNF (MPNF) or lateral PNF (LPNF)were ablated and the embryos cultured until they reached Kemp’s stage 42 and 44. Ablation of theLPNF provided phenotypes with arrested development of the eye, reduction of periocularpigmentation, frontonasal deformity, and a slightly reduced olfactory organ, whereas the MPNF-ablated phenotypes resulted in arrested development of the cornea and frontonasal deformity.Second, we labeled the mid-axial level of the PNF with vital DiI and traced the migration of labeledcells following culture to Kemp’s stage 33. Labeled PNF-derived cells populated a basal layer of theolfactory placode, migrated into the frontonasal region, the antero-dorsal periocular quadrant, andalso terminated at positions where the forebrain meninges form at later stages. Third, we examinedHNK-1 immunoreactivity in the forebrain-related region. We conclude that in the Australianlungfish: (1) LPNF-derived neuroepithelium gives rise to the basal layer and contributes to the apicallayer of the olfactory placode; (2) PNF-derived NC cells appear to give rise to meningeal, periocular,and frontonasal ectomesenchyme and likely infiltrate the olfactory placode as developmentalprecusors of the terminal nerve; (3) HNK-1 epitope is temporarily expressed in cells of the neuraltube, NC cells, and neurogenic placodal cells. Our experiments have provided the first evidence for apremandibular NC stream (sensu Kundrat, 2008) in a fish. J. Exp. Zool. (Mol. Dev. Evol.)312B:83– 94, 2009. r 2008 Wiley-Liss, Inc.
How to cite this article: Kundrat M, Joss JMP, Olsson L. 2009. Prosencephalic neuralfolds give rise to neural crest cells in the Australian lungfish, Neoceratodus forsteri.J. Exp. Zool. (Mol. Dev. Evol.) 312B:83–94.
Shortly after neural induction, embryonic ecto-derm becomes subdivided into domains withdifferent fates: most dorsally, the neural plate,immediately adjacent to the neural plate, theneural crest (NC), and beyond this, the placodeprecursors and future epidermis (Keller, ’75;Rosenquist, ’81; Couly and Le Douarin, ’85,’87;Garcia-Martinez et al., ’93; Woo and Fraser, ’95;Baker and Bronner-Fraser, 2001; Fernandez-Garre et al., 2002; Knecht and Bronner-Fraser,2002; Streit, 2004). The NC is believed to havebeen a significant evolutionary innovation en-abling the rapid adaptive radiation of craniates(Gans and Northcutt, ’83; Northcutt, 2005).
Toward the end of neurulation, NC cells migrateaway from the dorsal region adjacent to the neuraltube to prescribed regions of the embryo (LeDouarin and Kalcheim, ’99). The plethora offeatures to which the NC cells contribute includespigment and neuroendocrine cells, meninges,
Published online 8 October 2008 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/jez.b.21248
Received 5 May 2008; Revised 30 September 2008; Accepted 6October 2008
Grant sponsor: Macquarie University.�Correspondence to: Martin Kundrat, Geological Institute, Slovak
Academy of Sciences, Severna 5, Banska Bystrica, SK-97401, SlovakRepublic. E-mail: [email protected]
r 2008 WILEY-LISS, INC.
JOURNAL OF EXPERIMENTAL ZOOLOGY (MOL DEV EVOL) 312B:83–94 (2009)

pericytes, most elements of the peripheral nervoussystem, connective tissue, and skeletal elements ofthe face, including odontoblasts (Le Douarin, ’83;Hall, ’99; Le Douarin and Kalcheim, ’99).
The most complicated patterning of the NCoccurs in the head region. The cephalic NCcells migrate in distinctly recognizable streams,designated here as premandibular (fromcaudal forebrain), mandibular (from caudal mid-brain1rostral hindbrain), hyoid (from caudalpreotic hindbrain), and branchial (fromcaudal postotic hindbrain). Each stream of NCcells has a somewhat different pattern of migra-tion and contribution to features in the head(Le Douarin and Kalcheim, ’99). The mostanterior of the NC streams, the premandibular(Kundrat, 2008), provides a variety of cellprecursors that participate in the formationof adenohypophyseal and olfactory placodesand other NC-derived features of amniotevertebrates (Couly and Le Douarin, ’85,’87; Hall,’99; Le Douarin and Kalcheim, ’99; Baker andBronner-Fraser, 2001; Schlosser and Ahrens,2004; Schlosser, 2005).
In the mouse, ultrastructural investigations(Nichols, ’86) indicated that there are emigratingNC cells at the level of the forebrain. Followinginjections at the level of the mouse caudalprosencephalon, DiI-labeled cells were observedto migrate, as a sheet of cells, ventrally ashort distance from the neural tube, where theywere found to populate the mesenchyme aroundthe eye (Serbedzija et al., ’92). Subsequent experi-ments with cultured mouse embryos showed thatDiI-labeled NC cells derived from the prosence-phalon migrated into the periocular region wherethey became mixed with mesencephalic NC cells(Osumi-Yamashita et al., ’94). Furthermore, la-beled NC cells also contributed to mesenchymalcells in both the lateral and medial areas of thefrontonasal mass (Osumi-Yamashita et al., ’94).Recent data from 0 to 3-somite stage in miceindicate that not only the neural fold, but also thelateral part of the prosencephalic neuroepitheliumproduces NC cells; these NC-generating domainsare gradually confined to narrower areas duringdevelopment as only a few populations are foundto contribute to the NC cell lineage when thedomains are labeled at the 5–7-somite stage (Inoueet al., 2000).
Studies on chick embryos, tend to corroboratethe mouse findings, that there is a population ofNC cells migrating away from the forebrain(prosencephalon) region. The chick studies are
mostly based on quail-chick chimaeras, ratherthan vital dye tracing of migratory cells. In chick,NC cells derived from the forebrain zone gave riseto rostral membraneous bones (premaxillary,maxillary, nasal, and vomer), the interorbitalseptum, the sclerotic and supraorbital bone, aswell as the corneal endothelium, stromal cells, andleptomeningeal tissues (pericytes and smoothmuscle cells) (Noden, ’78; Couly and Le Douarin,’87; Couly et al., ’93; Etchevers et al., ’99, 2001).Recently, precursors of the ophthalmic and oculo-motor nerves, and of the ciliary ganglion in thechicken, were also shown to come from the caudalforebrain/rostral midbrain (Lee et al., 2003).Whether both prosencephalic and mesencephalicpopulations contribute to the skeletogenic tissuesin anterior craniofacial structures has not yet beenclarified.
As far as we know, there is still no evidencethat NC cells are derived from the prosencephalonin amphibians (Epperlein et al., 2000; Olssonet al., 2002; Olsson, 2003). Ambiguous informationis coming from studies on other anamniotes,including agnathans. NC cells have been observedat the border of the prosencephalon and mesence-phalon in platyfish embryos (Sadaghiani andVielkind, ’89). However, despite considerableeffort, no equivalent of the premandibular NChas been demonstrated in any anamniotevertebrate (teleosts studied include medaka: Lan-gille and Hall, ’88; and zebrafish: Woo andFraser, ’95; Whitlock and Westerfield, 2000;Whitlock, 2004).
To investigate whether premandibular NC cellsmay have existed in more basal osteichthyans, weused the living sarcopterygian lungfish, Neocer-atodus forsteri, as our experimental model. Wehave examined the fate of a group of ectodermalcells adjacent to the procencephalic neural folds(PNF) of developing lungfish, using three differentapproaches: (1) ablation, (2) vital dye labeling ofthis presumptive NC-forming site, and (3) HNK-1staining properties.
MATERIAL AND METHODS
Embryo collection
Eggs containing embryos of the Australianlungfish, N. forsteri, were collected from ourbreeding ponds at Macquarie University inSydney. The embryos were maintained in ovoindividually in sterile pond-water until theyreached the required stages, which we determined
M. KUNDRAT ET AL.84
J. Exp. Zool. (Mol. Dev. Evol.)

according to Kemp’s table of normal develop-ment for Neoceratodus (Kemp, ’82). All experi-mental procedures were conducted accordingto protocols approved by the MacquarieUniversity Animal Ethics Committee (AECapproval ] 2003-001).
Immunohistochemistry
Embryos (n 5 14, stage 32–34; n 5 2, stage 44) ofN. forsteri fixed in Bouin’s fixative were dehy-drated in a graded series of ethanol, methylsalicylate, and benzene, embedded in paraffinwax and sectioned using a rotary microtome(Spencer, Model ]820) at 10mm. Sections weremounted on slides, dewaxed and treated withMeOH and 1% (v/v) hydrogen peroxide for 30 minat room temperature (RT), followed by washing inTS1/3T/DMSO. An HNK-1 (anti Leu-7, BectonDickinson, San Jose, CA) monoclonal antibody(Ab) was used to identify neurogenic tissues, suchas NC and placodal cells. The HNK-1 Ab wasdiluted 1/50 in TS1/3T/M/DMSO (5% skim milk inTS1/3T/DMSO) and applied to the sections for 1 hrat RT. After washing in TS1/3T/DMSO, thesecondary Ab, a horseradish peroxidase (HRP)-conjugated goat anti-mouse IgM, (JacksonImmuno Research Laboratories, Inc., West Grove,PA), was diluted 1/200 in TS1/3T/M/DMSO andapplied to the sections for 1 hr. The sectionswere finally washed in TS1/3T/DMSO, and pre-incubated using 3,30-diaminobenzidine diluted inTS1/3T/DMSO. After the peroxidase reaction,the sections were counterstained with HarrisHematoxylin.
In vivo dye labeling
All injections were made with a 0.05% solution(w/v) of 1,1-dioctadecyl-3,3,30,30-tetramethylindo-carbocyanine perchlorate (DiI, Molecular Probes,Eugene, OR), made from a 0.5% stock solution in100% ethanol, diluted immediately before injec-tion 1:10 with 0.3 M sucrose. Micropipets werebackfilled with DiI solution and attached to aPicospritzer micro-injector (Narishige, Japan). Tolabel cephalic NC cells at specific levels, themicropipet was inserted by hand at an acute anglethrough the egg envelopes into the developingprosencephalic and mesencephalic neural folds atKemp’s stage 24 (7–9-somites). With the pipetpositioned at the junction of the neuroectodermand the ectoderm, a small amount of the DiIsolution was expelled. The fidelity of these injec-tions was determined by examining the embryos
at the time of injection. The labeled site wasvisible through the dissecting microscope as a pinkspot of dye in the tissue. Only embryos in whichthe DiI was clearly contained within the solelyprosencephalic (nP 5 3), prosencephalic1mesen-cephalic regions (nP/M 5 1) and solely mesence-phalic region (nM 5 3) were reared to Kemp’sstage 33 (nP1P/M 5 4) and 44 (nM 5 3). We usedthe same embryonic stage 24 for DiI injections asfor the ablation experiments. Moreover, DiI wasinjected into the same site centrally located insidethe PNF (Fig. 2C), from which tissue was removedin the ablation experiments. Based on the chickfate map (Couly and Le Douarin, ’87), this site isnext to the anlage of the olfactory placode rostrallyand separated from the most rostral extension ofthe cephalic NC. The embryos were cultured in20% Holtfreter solution in the dark at RT. At theend of the culture period (stage 33 or 44), embryoswere anaesthetized with 3-aminobenzoic acidethyl ester (MS222, Sigma) and fixed in 2%paraformaldehyde/1.25% glutaraldehyde in 0.1 Mphosphate-buffered saline (PBS) at least overnightand processed for vibratome sectioning (Leica VT1000S, Germany).
Ablation experiments
Lungfish embryos at Kemp’s stage 24 (n 5 5)were operated in ovo. Sharpened glass needleswere used to make small incisions to removethe prosencephalic neuroectoderm and underlyingpresumptive NC cells. Ablations, either lateral(n 5 2) or medial (n 5 3), were limited to thedorsal part of the prosencephalic fold and didnot expose the lumen of the neural tube.During this operation, the lungfish eggs werestabilized in a hole made inside an agar-coateddish. All operated embryos survived and werecultured in 20% Holtfreter solution in thedark at RT until they reached Kemp’s stage42–44, when they were anaesthesized with 3-aminobenzoic acid ethyl ester (MS222, Sigma)and fixed in 4% paraformaldehyde in 0.1 M PBS atleast overnight. The unoperated side of each fishserved as its own control. In addition, unoperatedlungfish (n 5 3) were cultured from stage 24to 44 under the same conditions as the operatedfish. Following imaging of the external features ofthe fixed lungfish, the specimens were stainedwith Alcian blue and cleared for viewing anydifferences in development of the internal skele-ton according to the method of Dingerkus andUhler (’77).
PROSENCEPHALIC NEURAL FOLD IN LUNGFISH 85
J. Exp. Zool. (Mol. Dev. Evol.)
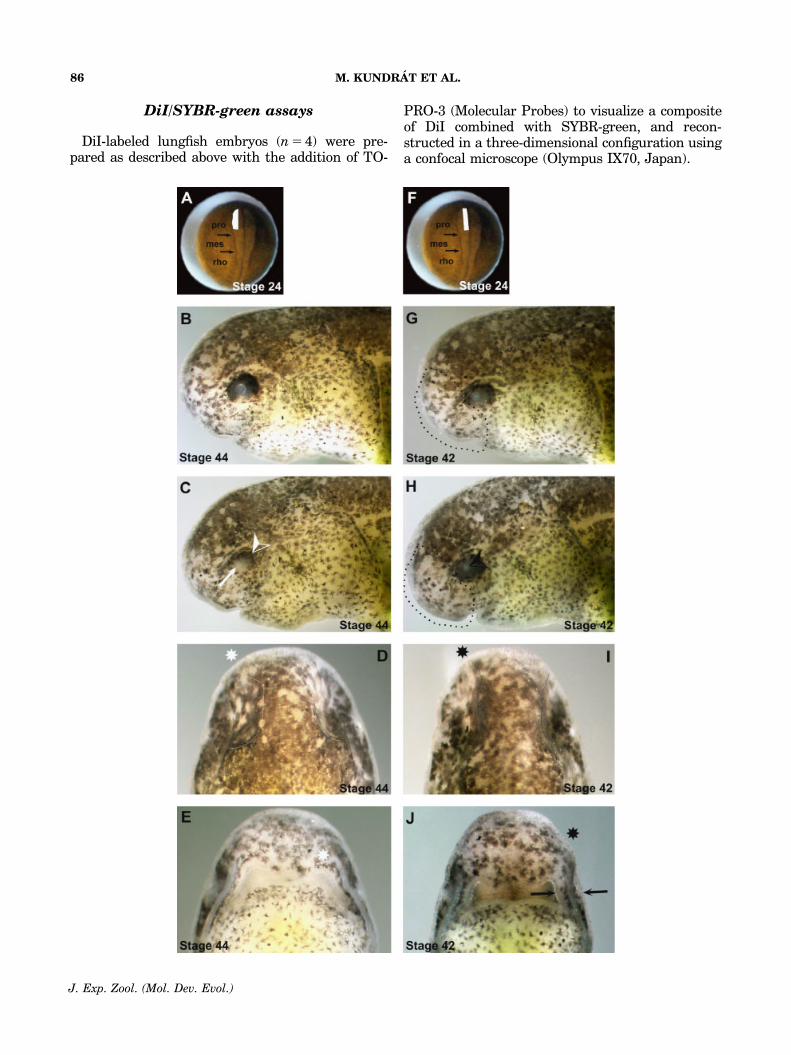
DiI/SYBR-green assays
DiI-labeled lungfish embryos (n 5 4) were pre-pared as described above with the addition of TO-
PRO-3 (Molecular Probes) to visualize a compositeof DiI combined with SYBR-green, and recon-structed in a three-dimensional configuration usinga confocal microscope (Olympus IX70, Japan).
M. KUNDRAT ET AL.86
J. Exp. Zool. (Mol. Dev. Evol.)

DiI/HNK-1/TO-PRO-3 or HNK-1/TO-PRO-3composite assays
DiI-labeled lungfish embryos (stage 33; n 5 3)and intact wild type lungfish embryos (stage 44)were fixed by immersion in 2% paraformaldehyde/1.25% glutaraldehyde in 0.1 M PBS at 41C. Theywere serially sectioned at 50mm using a vibratome(Leica VT 1000S). The sections were immunos-tained with HNK-1 and Alexa Fluor 488 goat anti-mouse IgM (Molecular Probes), and stained withTO-PRO-3 (Molecular Probes), washed with PBS,and mounted in gel/mount (ProSciTech) for view-ing first in the epifluorescent field to detect thedistribution of DiI signal and then using a laserscanning confocal microscope (Olympus IX70) tovisualize a composite three-dimensional view ofthese three labels in different colors.
RESULTS
Ablation of prosencephalic neural foldcauses developmental deficiencies in NC-
derived and placodal tissues
Our first series of ablation experiments targetedthe removal of a dorsal two-thirds of the lateralPNF (LPNF) (Fig. 1A). We were careful not tocontaminate our removals with non-neural ecto-derm and avoided breaking into the neural canal.All embryos in which a breakage into a neuraltube was observed visually, were removed fromfurther experimental procedure (these specimenswere not included in counts of operated embryosgiven in Methods). Operated embryos were cul-tured until they reached stage 44 (Fig. 1B). Weobserved the following phenotypic differences inthese embryos when compared with the controlsand the unoperated side of each embryo: (1)reduction of periocular pigmentation and arresteddevelopment of the eye (microphthalmia),particularly of the cornea (Fig. 1C); (2)frontonasal deformity (Fig. 1D); and (3) a slight
reduction of the olfactory organ on the operatedside (Fig. 1E).
Our second series of ablation experimentsremoved the dorsal two-thirds of the medialPNF (MPNF) (Fig. 1F). Again, we were carefulto avoid penetrating the neural canal. Operatedembryos were cultured until they reached eitherstage 42 (Fig. 1G) or 44. In this case, phenotypicdifferences from controls were: (1) slightly under-developed cornea and reduction of intensity anddensity of pigmentation fields in the frontonasalregion (Fig. 1H); (2) a deformation of the fronto-nasal region (Fig. 1I); and (3) reduction of tissuelateral to the olfactory organ on the operated side(Fig. 1J).
There were no differences between the internalcranial skeletal features, in particular, the trabe-culae of both ablated specimens and controls haddeveloped equivalently.
Fate of prosencephalic NC cell andplacodal precursors
Vital dye labeling with the lipophilic dye, DiI,was used to fate map PNF cells to substantiate ourdata from microsurgery (Fig. 1).
Detection of DiI-labeled cells demonstrated thepresence of a population of migratory cells posi-tioned rostral to the mandibular (trigeminal) NCstream (Fig. 2A, B). This premandibular migra-tory population is derived exclusively from theforebrain. This is the first direct evidence thatmigratory cells may be derived from the rostral-most segment of the neural tube, the prosence-phalon, in a fish. These cells migrated into thefrontonasal region of the stage 33 lungfish embryo(Fig. 2D, E, G).
DiI-labeled cells also ended up in the peripheryof the neural tube (Fig. 2G) where meninges areformed at later stages and populated the perio-cular mesenchyme (Fig. 2F), specifically a regionassociated with the antero-dorsal quadrant to theoptic vesicle (Fig. 2A).
Fig. 1. Ablation of prosencephalic NC cells in the Australian lungfish, N. forsteri. (A) Unilateral ablation of a lateralprosencephalic fold (white area) before emigration of the NC cells at stage 24, and the corresponding phenotypes (B–E) afterculturing operated embryos until stage 44. (B) Intact right side of the embryo as the control. (C) Operated left side of the embryoshows arrested development of the eye (arrow) and reduction of the periocular pigmentation (arrowhead). (D) Dorsal view of theembryo with a deformation in the frontonasal region on the operated left side (asterisk). (E) Ventral view of the embryo with thereduced olfactory organ on the operated side (asterisk). (F) Unilateral ablation of a medial prosencephalic fold (white area)before emigration of the NC cells at stage 24, and the corresponding phenotypes (G–J) after culturing to stage 42. (G) Intactright side of the embryo as the control. (H) Operated left side of the embryo shows arrested development of the cornea(arrowhead) and reduction of pigmentation in the frontonasal region (area boxed by dashed line in G and H). (I) Dorsal view ofan embryo with a deformation in the frontonasal region on the operated left side (asterisk). (J) Ventral view of the embryo witha reduction of tissue (asterisk) lateral to the olfactory opening (arrows) on the operated side. mes, mesencephalon; pro,prosencephalon; rho, rhombencephalon; NC, neural crest.
PROSENCEPHALIC NEURAL FOLD IN LUNGFISH 87
J. Exp. Zool. (Mol. Dev. Evol.)
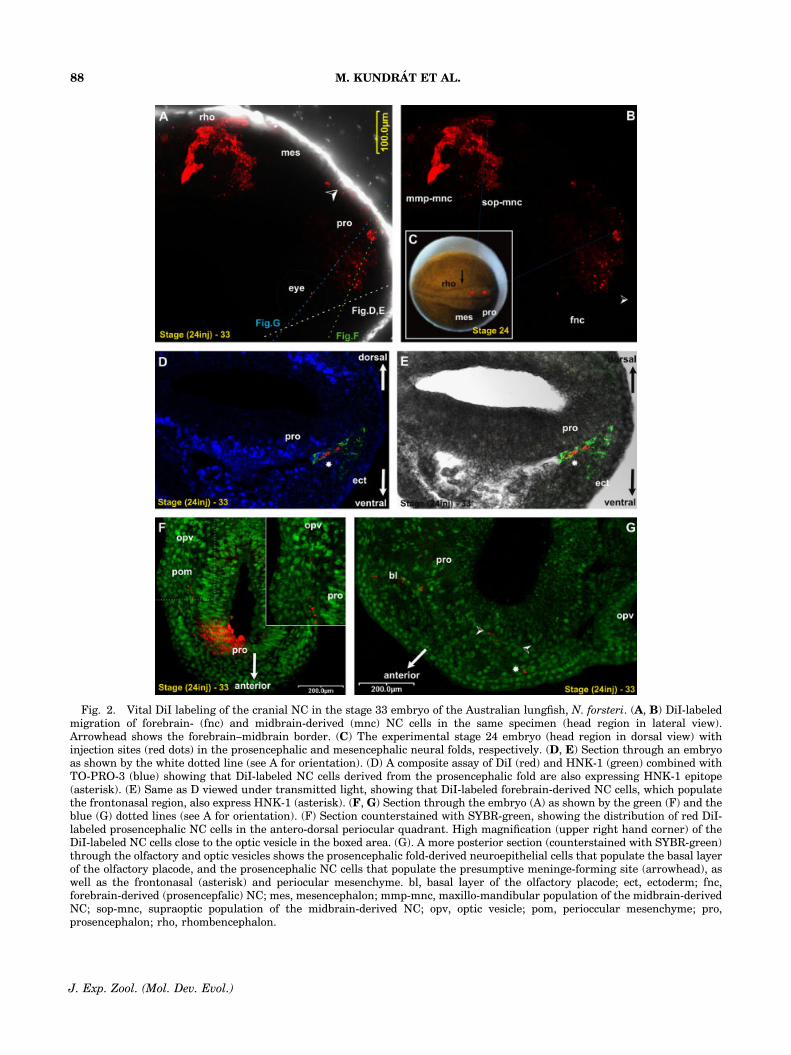
Fig. 2. Vital DiI labeling of the cranial NC in the stage 33 embryo of the Australian lungfish, N. forsteri. (A, B) DiI-labeledmigration of forebrain- (fnc) and midbrain-derived (mnc) NC cells in the same specimen (head region in lateral view).Arrowhead shows the forebrain–midbrain border. (C) The experimental stage 24 embryo (head region in dorsal view) withinjection sites (red dots) in the prosencephalic and mesencephalic neural folds, respectively. (D, E) Section through an embryoas shown by the white dotted line (see A for orientation). (D) A composite assay of DiI (red) and HNK-1 (green) combined withTO-PRO-3 (blue) showing that DiI-labeled NC cells derived from the prosencephalic fold are also expressing HNK-1 epitope(asterisk). (E) Same as D viewed under transmitted light, showing that DiI-labeled forebrain-derived NC cells, which populatethe frontonasal region, also express HNK-1 (asterisk). (F, G) Section through the embryo (A) as shown by the green (F) and theblue (G) dotted lines (see A for orientation). (F) Section counterstained with SYBR-green, showing the distribution of red DiI-labeled prosencephalic NC cells in the antero-dorsal periocular quadrant. High magnification (upper right hand corner) of theDiI-labeled NC cells close to the optic vesicle in the boxed area. (G). A more posterior section (counterstained with SYBR-green)through the olfactory and optic vesicles shows the prosencephalic fold-derived neuroepithelial cells that populate the basal layerof the olfactory placode, and the prosencephalic NC cells that populate the presumptive meninge-forming site (arrowhead), aswell as the frontonasal (asterisk) and periocular mesenchyme. bl, basal layer of the olfactory placode; ect, ectoderm; fnc,forebrain-derived (prosencepfalic) NC; mes, mesencephalon; mmp-mnc, maxillo-mandibular population of the midbrain-derivedNC; sop-mnc, supraoptic population of the midbrain-derived NC; opv, optic vesicle; pom, perioccular mesenchyme; pro,prosencephalon; rho, rhombencephalon.
M. KUNDRAT ET AL.88
J. Exp. Zool. (Mol. Dev. Evol.)
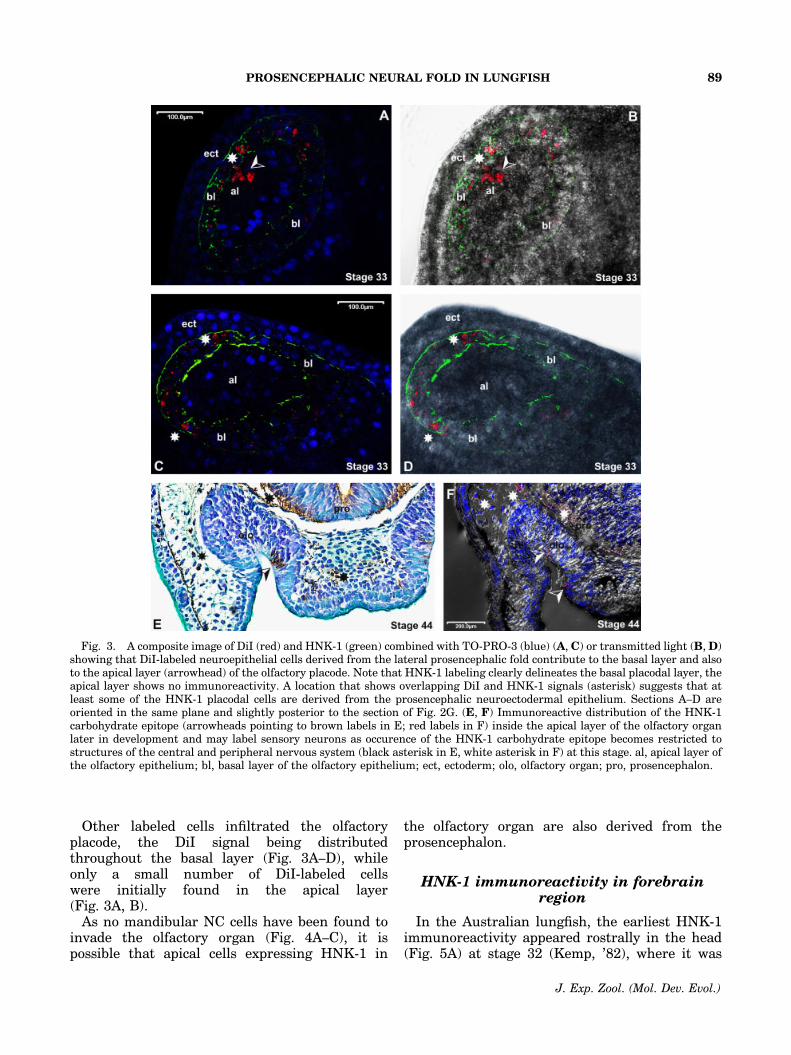
Other labeled cells infiltrated the olfactoryplacode, the DiI signal being distributedthroughout the basal layer (Fig. 3A–D), whileonly a small number of DiI-labeled cellswere initially found in the apical layer(Fig. 3A, B).
As no mandibular NC cells have been found toinvade the olfactory organ (Fig. 4A–C), it ispossible that apical cells expressing HNK-1 in
the olfactory organ are also derived from theprosencephalon.
HNK-1 immunoreactivity in forebrainregion
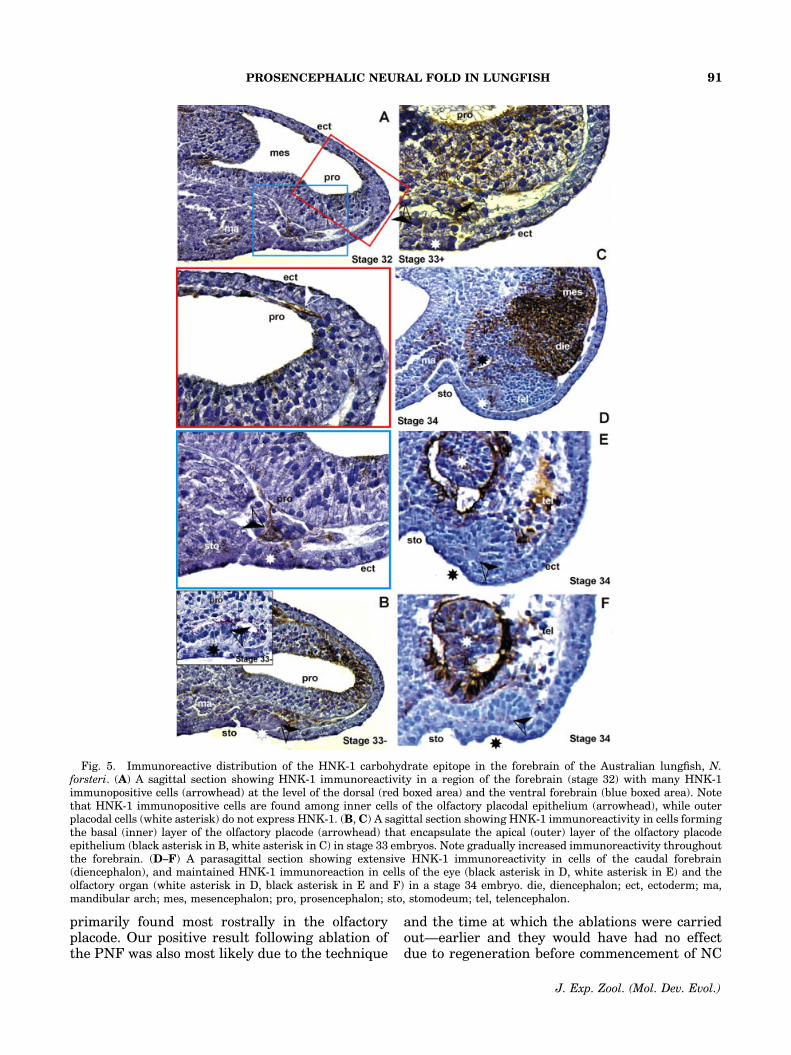
In the Australian lungfish, the earliest HNK-1immunoreactivity appeared rostrally in the head(Fig. 5A) at stage 32 (Kemp, ’82), where it was
Fig. 3. A composite image of DiI (red) and HNK-1 (green) combined with TO-PRO-3 (blue) (A, C) or transmitted light (B, D)showing that DiI-labeled neuroepithelial cells derived from the lateral prosencephalic fold contribute to the basal layer and alsoto the apical layer (arrowhead) of the olfactory placode. Note that HNK-1 labeling clearly delineates the basal placodal layer, theapical layer shows no immunoreactivity. A location that shows overlapping DiI and HNK-1 signals (asterisk) suggests that atleast some of the HNK-1 placodal cells are derived from the prosencephalic neuroectodermal epithelium. Sections A–D areoriented in the same plane and slightly posterior to the section of Fig. 2G. (E, F) Immunoreactive distribution of the HNK-1carbohydrate epitope (arrowheads pointing to brown labels in E; red labels in F) inside the apical layer of the olfactory organlater in development and may label sensory neurons as occurence of the HNK-1 carbohydrate epitope becomes restricted tostructures of the central and peripheral nervous system (black asterisk in E, white asterisk in F) at this stage. al, apical layer ofthe olfactory epithelium; bl, basal layer of the olfactory epithelium; ect, ectoderm; olo, olfactory organ; pro, prosencephalon.
PROSENCEPHALIC NEURAL FOLD IN LUNGFISH 89
J. Exp. Zool. (Mol. Dev. Evol.)
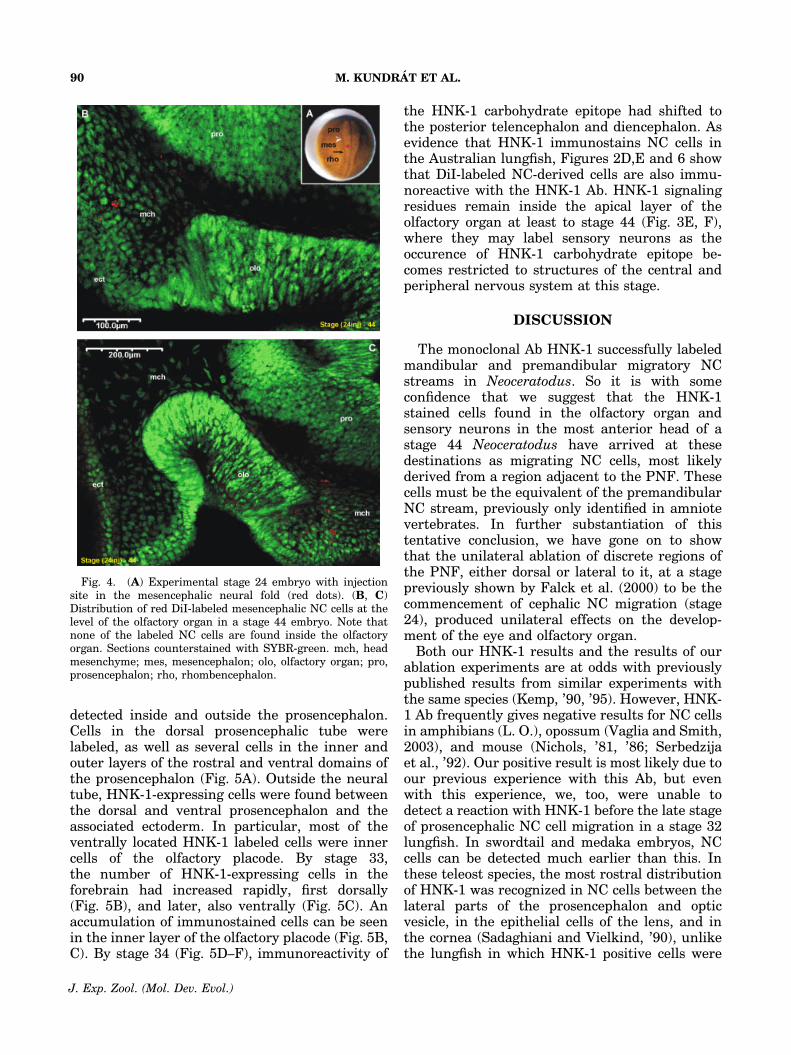
detected inside and outside the prosencephalon.Cells in the dorsal prosencephalic tube werelabeled, as well as several cells in the inner andouter layers of the rostral and ventral domains ofthe prosencephalon (Fig. 5A). Outside the neuraltube, HNK-1-expressing cells were found betweenthe dorsal and ventral prosencephalon and theassociated ectoderm. In particular, most of theventrally located HNK-1 labeled cells were innercells of the olfactory placode. By stage 33,the number of HNK-1-expressing cells in theforebrain had increased rapidly, first dorsally(Fig. 5B), and later, also ventrally (Fig. 5C). Anaccumulation of immunostained cells can be seenin the inner layer of the olfactory placode (Fig. 5B,C). By stage 34 (Fig. 5D–F), immunoreactivity of
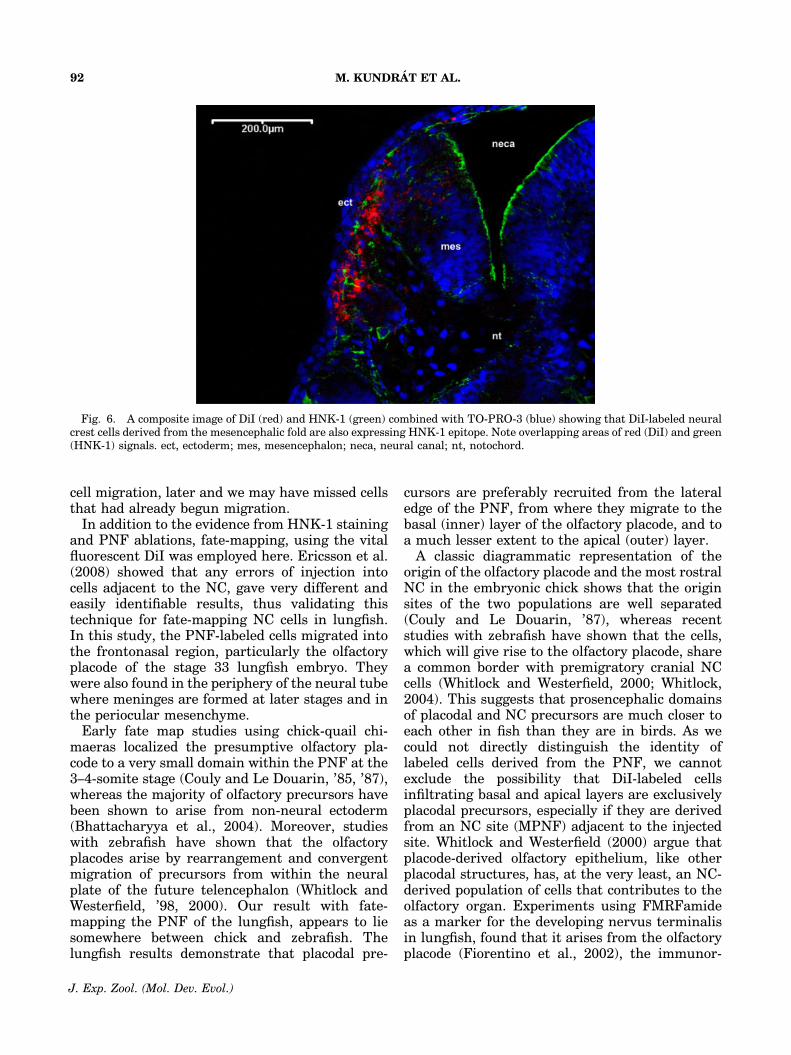
the HNK-1 carbohydrate epitope had shifted tothe posterior telencephalon and diencephalon. Asevidence that HNK-1 immunostains NC cells inthe Australian lungfish, Figures 2D,E and 6 showthat DiI-labeled NC-derived cells are also immu-noreactive with the HNK-1 Ab. HNK-1 signalingresidues remain inside the apical layer of theolfactory organ at least to stage 44 (Fig. 3E, F),where they may label sensory neurons as theoccurence of HNK-1 carbohydrate epitope be-comes restricted to structures of the central andperipheral nervous system at this stage.
DISCUSSION
The monoclonal Ab HNK-1 successfully labeledmandibular and premandibular migratory NCstreams in Neoceratodus. So it is with someconfidence that we suggest that the HNK-1stained cells found in the olfactory organ andsensory neurons in the most anterior head of astage 44 Neoceratodus have arrived at thesedestinations as migrating NC cells, most likelyderived from a region adjacent to the PNF. Thesecells must be the equivalent of the premandibularNC stream, previously only identified in amniotevertebrates. In further substantiation of thistentative conclusion, we have gone on to showthat the unilateral ablation of discrete regions ofthe PNF, either dorsal or lateral to it, at a stagepreviously shown by Falck et al. (2000) to be thecommencement of cephalic NC migration (stage24), produced unilateral effects on the develop-ment of the eye and olfactory organ.
Both our HNK-1 results and the results of ourablation experiments are at odds with previouslypublished results from similar experiments withthe same species (Kemp, ’90, ’95). However, HNK-1 Ab frequently gives negative results for NC cellsin amphibians (L. O.), opossum (Vaglia and Smith,2003), and mouse (Nichols, ’81, ’86; Serbedzijaet al., ’92). Our positive result is most likely due toour previous experience with this Ab, but evenwith this experience, we, too, were unable todetect a reaction with HNK-1 before the late stageof prosencephalic NC cell migration in a stage 32lungfish. In swordtail and medaka embryos, NCcells can be detected much earlier than this. Inthese teleost species, the most rostral distributionof HNK-1 was recognized in NC cells between thelateral parts of the prosencephalon and opticvesicle, in the epithelial cells of the lens, and inthe cornea (Sadaghiani and Vielkind, ’90), unlikethe lungfish in which HNK-1 positive cells were
Fig. 4. (A) Experimental stage 24 embryo with injectionsite in the mesencephalic neural fold (red dots). (B, C)Distribution of red DiI-labeled mesencephalic NC cells at thelevel of the olfactory organ in a stage 44 embryo. Note thatnone of the labeled NC cells are found inside the olfactoryorgan. Sections counterstained with SYBR-green. mch, headmesenchyme; mes, mesencephalon; olo, olfactory organ; pro,prosencephalon; rho, rhombencephalon.
M. KUNDRAT ET AL.90
J. Exp. Zool. (Mol. Dev. Evol.)
primarily found most rostrally in the olfactoryplacode. Our positive result following ablation ofthe PNF was also most likely due to the technique
and the time at which the ablations were carriedout—earlier and they would have had no effectdue to regeneration before commencement of NC
Fig. 5. Immunoreactive distribution of the HNK-1 carbohydrate epitope in the forebrain of the Australian lungfish, N.forsteri. (A) A sagittal section showing HNK-1 immunoreactivity in a region of the forebrain (stage 32) with many HNK-1immunopositive cells (arrowhead) at the level of the dorsal (red boxed area) and the ventral forebrain (blue boxed area). Notethat HNK-1 immunopositive cells are found among inner cells of the olfactory placodal epithelium (arrowhead), while outerplacodal cells (white asterisk) do not express HNK-1. (B, C) A sagittal section showing HNK-1 immunoreactivity in cells formingthe basal (inner) layer of the olfactory placode (arrowhead) that encapsulate the apical (outer) layer of the olfactory placodeepithelium (black asterisk in B, white asterisk in C) in stage 33 embryos. Note gradually increased immunoreactivity throughoutthe forebrain. (D–F) A parasagittal section showing extensive HNK-1 immunoreactivity in cells of the caudal forebrain(diencephalon), and maintained HNK-1 immunoreaction in cells of the eye (black asterisk in D, white asterisk in E) and theolfactory organ (white asterisk in D, black asterisk in E and F) in a stage 34 embryo. die, diencephalon; ect, ectoderm; ma,mandibular arch; mes, mesencephalon; pro, prosencephalon; sto, stomodeum; tel, telencephalon.
PROSENCEPHALIC NEURAL FOLD IN LUNGFISH 91
J. Exp. Zool. (Mol. Dev. Evol.)
cell migration, later and we may have missed cellsthat had already begun migration.
In addition to the evidence from HNK-1 stainingand PNF ablations, fate-mapping, using the vitalfluorescent DiI was employed here. Ericsson et al.(2008) showed that any errors of injection intocells adjacent to the NC, gave very different andeasily identifiable results, thus validating thistechnique for fate-mapping NC cells in lungfish.In this study, the PNF-labeled cells migrated intothe frontonasal region, particularly the olfactoryplacode of the stage 33 lungfish embryo. Theywere also found in the periphery of the neural tubewhere meninges are formed at later stages and inthe periocular mesenchyme.
Early fate map studies using chick-quail chi-maeras localized the presumptive olfactory pla-code to a very small domain within the PNF at the3–4-somite stage (Couly and Le Douarin, ’85, ’87),whereas the majority of olfactory precursors havebeen shown to arise from non-neural ectoderm(Bhattacharyya et al., 2004). Moreover, studieswith zebrafish have shown that the olfactoryplacodes arise by rearrangement and convergentmigration of precursors from within the neuralplate of the future telencephalon (Whitlock andWesterfield, ’98, 2000). Our result with fate-mapping the PNF of the lungfish, appears to liesomewhere between chick and zebrafish. Thelungfish results demonstrate that placodal pre-
cursors are preferably recruited from the lateraledge of the PNF, from where they migrate to thebasal (inner) layer of the olfactory placode, and toa much lesser extent to the apical (outer) layer.
A classic diagrammatic representation of theorigin of the olfactory placode and the most rostralNC in the embryonic chick shows that the originsites of the two populations are well separated(Couly and Le Douarin, ’87), whereas recentstudies with zebrafish have shown that the cells,which will give rise to the olfactory placode, sharea common border with premigratory cranial NCcells (Whitlock and Westerfield, 2000; Whitlock,2004). This suggests that prosencephalic domainsof placodal and NC precursors are much closer toeach other in fish than they are in birds. As wecould not directly distinguish the identity oflabeled cells derived from the PNF, we cannotexclude the possibility that DiI-labeled cellsinfiltrating basal and apical layers are exclusivelyplacodal precursors, especially if they are derivedfrom an NC site (MPNF) adjacent to the injectedsite. Whitlock and Westerfield (2000) argue thatplacode-derived olfactory epithelium, like otherplacodal structures, has, at the very least, an NC-derived population of cells that contributes to theolfactory organ. Experiments using FMRFamideas a marker for the developing nervus terminalisin lungfish, found that it arises from the olfactoryplacode (Fiorentino et al., 2002), the immunor-
Fig. 6. A composite image of DiI (red) and HNK-1 (green) combined with TO-PRO-3 (blue) showing that DiI-labeled neuralcrest cells derived from the mesencephalic fold are also expressing HNK-1 epitope. Note overlapping areas of red (DiI) and green(HNK-1) signals. ect, ectoderm; mes, mesencephalon; neca, neural canal; nt, notochord.
M. KUNDRAT ET AL.92
J. Exp. Zool. (Mol. Dev. Evol.)

eactive cells subsequently moving outside theolfactory placode to form the ganglion of thenervus terminalis. These cells appear first in thepart of the olfactory mucosa that corresponds tothe basal layer of the olfactory placode. In thezebrafish, using a combination of DiI labeling andsingle cell lineage tracing, Whitlock et al. (2003)demonstrated that GnRH-positive cells of thenervus terminalis originated from the cranialNC. This suggests that in lungfish, at least someDiI-labeled cells, derived from the investigateddomain producing prosencephalic, placodal, andNC precursors, could represent NC-derived pre-cursors of the terminal nerve. This suggestionwould support the idea that the nervus terminaliscould be NC-derived (amphioxus and lamprey:Fritzsch and Northcutt, ’93; Northcutt, ’93;sharks: Locy, ’05; reptiles and mammals: John-ston, ’09, ’13).
In conclusion, our experiments have providedstrong evidence for a premandibular NC stream(sensu Kundrat, 2008) in lungfish. We have shownthat PNF-derived migratory cells have a largerange of developmental potentialities, similar tothose for the premandibular NC in mouse andchick. These include differentiation of cell pre-cursors that form sensory placodal epithelium, aswell as NC cells that infiltrate the frontonasalregion of the lungfish head, and likely contributeto placodal precursors of the terminal nerve.These NC cells were not found in the Falck et al.(2000) SEM study, so this study has extended thecranial NC of lungfish further rostrally thanpreviously thought. Because lungfish are basalsarcopterygians, the lack of a premandibular NCstream in other anamniote tetrapods (amphibians)must now be questioned.
ACKNOWLEDGMENTS
We thank Debra Birch for assistance withconfocal microscopy, Libby Eyre for collection oflungfish eggs, the sub-editor Shigeru Kuratani andanonymous referees for their valuable comments.M. K. thanks his family for support and dedicatesthis finding and research paper in memory of hisfather.
LITERATURE CITED
Baker CVH, Bronner-Fraser M. 2001. Vertebrate cranialplacodes. I. Embryonic induction. Dev Biol 232:1–61.
Bhattacharyya S, Bailey AP, Bronner-Fraser M, Streit A.2004. Segregation of lens and olfactory precursors from acommon territory: cell sorting and reciprocity of Dlx5 andPax6 expression. Dev Biol 271:403–414.
Couly GF, Le Douarin NM. 1985. Mapping of the early neuralprimordium in quail-chick chimeras. I. Developmentalrelationships between placodes, facial ectoderm, and pro-sencephalon. Dev Biol 110:422–439.
Couly GF, Le Douarin NM. 1987. Mapping of the early neuralprimordium in quail-chick chimeras. II. The prosencephalicneural plate and neural folds: implications for the genesis ofcephalic human congenital abnormalities. Dev Biol 120:198–214.
Couly GF, Coltey PM, Le Douarin NM. 1993. The triple originof skull in higher vertebrates–A study in quail-chickchimeras. Development 117:409–429.
Dingerkus G, Uhler LD. 1977. Enyzme clearing of alcian bluestained whole small vertebrates for demonstration ofcartilage. Stain Technol 52:229–232.
Epperlein H, Meulemans D, Bronner-Fraser M, SteinbeisserH, Selleck MA. 2000. Analysis of cranial neural crestmigratory pathways in axolotl using cell markers andtransplantation. Development 127:2751–2761.
Ericsson R, Joss J, Olsson L. 2008. The fate of cranial neuralcrest cells in the Australian lungfish, Neoceratodus forsteri.J Exp Zoolog B Mol Dev Evol 310B:345–354.
Etchevers HC, Couly G, Vincent C, Le Douarin NM. 1999.Anerior cephalic neural crest is required for forebrainviability. Development 126:3533–3543.
Etchevers HC, Vincent C, Le Douarin N, Couly GF. 2001. Thecephalic neural crest provides pericytes and smooth musclecells to all blood vessels of the face and forebrain.Development 128:1059–1068.
Falck P, Joss JM, Olsson L. 2000. Cranial neural crest cellmigration in the Australian lungfish, Neoceratodus forsteri.Evol Dev 4:179–185.
Fernandez-Garre P, Rodriguez-Gallardo L, Gallego-Diaz V,Alvarez IS, Puelles L. 2002. Fate map of the chicken neuralplate at stage 4. Development 129:2807–2822.
Fiorentino M, D’Aniello B, Joss J, Polese G, Rastogi RK. 2002.Ontogenic organization of the FMRFamide immunore-activity in the nervus terminalis of the lungfish, Neocer-atodus forsteri. J Comp Neurol 450:115–121.
Fritzsch B, Northcutt RG. 1993. Cranial and spinal nerveorganization in amphioxus and lamprey: evidence for anancestral craniate pattern. Acta Anat 148:96–109.
Gans C, Northcutt RG. 1983. Neural crest and the origin ofvertebrates: a new head. Science 220:268–274.
Garcia-Martinez V, Alvarez IS, Schoenwolf GC. 1993. Loca-tions of the ectodermal and nonectodermal subdivisions ofthe epiblast at stages 3 and 4 of avian gastrulation andneurulation. J Exp Zool 267:431–446.
Hall BK. 1999. The neural crest in development and evolution.New York: Springer-Verlag.
Inoue T, Nakamura S, Osumi N. 2000. Fate mappingof the mouse prosencephalic neural plate. Dev Biol 219:373–383.
Johnston JB. 1909. The morphology of the forebrain vesicle invertebrates. J Comp Neurol Psychol 19:457–539.
Johnston JB. 1913. Nervus terminalis in reptiles andmammals. J Comp Neurol 23:97–120.
Keller RE. 1975. Vital dye mapping of the gastrula andneurula of Xenopus laevis. I. Prospective areas andmorphogenetic movements of the superficial layer. Dev Biol42:222–241.
Kemp A. 1982. The embryological development of theQueensland lungfish, Neoceratodus forsteri (Krefft). MemQueensland Mus 20:553–597.
PROSENCEPHALIC NEURAL FOLD IN LUNGFISH 93
J. Exp. Zool. (Mol. Dev. Evol.)

Kemp A. 1990. Involvement of the neural crest in thedevelopment of the Australian lungfish, Neoceratodusforsteri (Frefft, 1980). Mem Queensland Mus 28:101–102.
Kemp A. 1995. On the neural crest cells of the Australianlungfish. Bull Mus Natl Hist Nat Paris 17:343–357.
Knecht AK, Bronner-Fraser N. 2002. Induction of the neuralcrest: a multigene process. Nat Rev Genet 3:453–461.
Kundrat M. 2008. HNK-1 immunoreactivity during earlymorphogenesis of the head region in a model-vertebrate,crocodile embryo. Naturwissenschaften DOI: 10.1007/s00114-008-0426-4.
Langille RM, Hall BK. 1988. Role of the neural crest indevelopment of the cartilaginous cranial and visceralskeleton of the medaka, Oryzias latipes (Teleostei). AnatEmbryol 177:297–305.
Le Douarin NM. 1983. The Neural Crest. Cambridge:Cambridge University Press.
Le Douarin NM, Kalcheim C. 1999. The Neural Crest.Cambridge: Cambridge University Press.
Lee VM, Sechrist JW, Luetolf S, Bronner-Fraser M. 2003.Both neural crest and placode contribute to the ciliaryganglion and oculomotor nerve. Dev Biol 263:176–190.
Locy WA. 1905. On a newly recognized nerve connected withthe forebrain of selachians. Anat Anz 26:33–36.
Nichols DH. 1981. Neural crest formation in the head of themouse embryo as observed using a new histologicaltechnique. J Embryol Exp Morphol 64:105–120.
Nichols DH. 1986. Formation and distribution of neural crestmesenchyme to the first pharyngeal arch region of themouse embryo. Am J Anat 176:221–231.
Noden DM. 1978. The control of avian cephalic neural crestcytodifferentiation. I. Skeletal and connective tissues. DevBiol 67:296–312.
Northcutt RG. 1993. A reassessment of Goodrich’s model ofcranial nerve phylogeny. Acta Anat 148:71–80.
Northcutt RG. 2005. The new head hypothesis revisited. J ExpZoolog B Mol Dev Evol 304B:274–297.
Olsson L. 2003. Cell migration, pattern formation and cell fateduring head development in lungfishes and amphibians.Theory Biosci 122:252–265.
Olsson L, Moury JD, Carl TF, Hastad O, Hanken J.2002. Cranial neural crest-cell migration in the direct-developing frog, Eleutherodactylus coqui: molecular hetero-
geneity within and among migratory streams. Zoology105:3–13.
Osumi-Yamashita N, Ninomiya Y, Doi H, Eto K. 1994. Thecontribution of both forebrain and midbrain crest cells tothe mesenchyme in the frontnasal mass of mouse embryos.Dev Biol 164:409–419.
Rosenquist GC. 1981. Epiblast origin and early migrationof neural crest cells in the chick embryo. Dev Biol67:201–211.
Sadaghiani B, Vielkind JR. 1989. Neural crest development inXiphophorus fishes: scanning electron and light microscopicstudies. Development 105:487–504.
Sadaghiani B, Vielkind JR. 1990. Distribution and migrationpatways of HNK-1-immunoreactive neural crest-cells inteleost fish embryos. Development 110:197–209.
Schlosser G. 2005. Evolutionary origins of vertebrate placodes:insights from developmental studies and from comparisonswith other deuterostomes. J Exp Zoolog B Mol Dev Evol304B:347–399.
Schlosser G, Ahrens K. 2004. Molecular anatomy of placodedevelopment in Xenopus laevis. Dev Biol 271:439–466.
Serbedzija GN, Bronner-Fraser M, Fraser SE. 1992. Vital dyeanalysis of cranial crest cell migration in the mouse embryo.Development 116:297–307.
Streit A. 2004. Early development of the cranial sensorynervous system: from a common field to individual placodes.Dev Biol 276:1–15.
Vaglia JL, Smith KK. 2003. Early differentiation and migra-tion of cranial neural crest in the opossum, Monodelphisdomestica. Evol Dev 5:121–135.
Whitlock KE. 2004. Development of the nervus terminalis:origin and migration. Microsc Res Tech 65:2–12.
Whitlock KE, Westerfield M. 1998. A transient population ofneurons pioneers the olfactory pathway in the zebrafish. JNeurosci 18:8919–8927.
Whitlock KE, Westerfield M. 2000. The olfactory placodes ofthe zebrafish form by convergence of cellular fields at theedge of the neural plate. Development 127:3645–3653.
Whitlock KE, Wolf CD, Boyce ML. 2003. Gonadotropin-releasing hormone (GnRH) cells arise from cranial neuralcrest and adenohypophyseal regions of the neural plate inthe zebrafish, Danio rerio. Dev Biol 257:140–152.
Woo K, Fraser SE. 1995. Order and coherence in the fate map ofthe zebrafish nervous system. Development 121:2595–2609.
M. KUNDRAT ET AL.94
J. Exp. Zool. (Mol. Dev. Evol.)
Copyright © 2022 FDOKUMEN




















