
Immunohistochemistry and in situ hybridization in the study of human skin melanocytes

www.elsevier.com/locate/jchemneu
Journal of Chemical Neuroanatomy 32 (2006) 127–142
Proliferative activity in the frog brain:
A PCNA-immunohistochemistry analysis
Franca Raucci a, Maria M. Di Fiore a, Claudia Pinelli a, Biagio D’Aniello b,Luciano Luongo b, Gianluca Polese b, Rakesh K. Rastogi b,*
a Department of Life Sciences, Second University of Naples, 81100 Caserta, Italyb Department of Structural and Functional Biology, University of Naples Federico II, Via Cinthia, MSA Campus, 80126 Naples, Italy
Received 12 April 2006; received in revised form 12 July 2006; accepted 5 August 2006
Available online 20 September 2006
Abstract
By means proliferating cell nuclear antigen (PCNA) immunohistochemistry, we have provided a detailed neuroanatomical mapping of
proliferative activity during development and adulthood in the frog (Rana esculenta) brain. Western blot analysis confirmed the presence of this
protein in brain extracts from adults and tadpoles. Proliferative activity was observed in the ventricular and subventricular zones throughout the
brain. The present study provides details as to which of the morphologically distinguishable brain region(s) has a long-lasting proliferative activity
and in which region this activity undergoes a progressive decrease during development. In the subventricular zones of the third ventricle, PCNA-
labeled cells were particularly abundant in the magnocellular preoptic nucleus and the ventromedial thalamic nucleus. It was observed that
proliferation zones are present practically in all major subdivisions of the forebrain, midbrain and hindbrain, including the cerebellum in which
PCNA-labeled cells were located in the outer granular layer and the inner molecular layer. The habenulae, epiphysis and isthmic nuclei never
showed the presence of PCNA-immunoreactive nuclei. The widespread proliferative activity implies that the frog brain has a great potential for
neurogenesis/gliogenesis not only during larval development but also in the adulthood.
# 2006 Elsevier B.V. All rights reserved.
Keywords: PCNA; Frog brain; Cell proliferation; Immunohistochemistry; Development
1. Introduction
The proliferating cells confer to the brain the capacity of
neurogenesis/gliogenesis and regeneration. Evidence accumu-
lated for over 30 years indicates that during adulthood, all
vertebrates possess potential to produce new neurons/astro-
cytes/glial cells in the central nervous system; this potential has
been observed within the framework of a seasonal or a
hormone-regulated behavioural rhythm, or in the replacement
of the damaged neurons by newly generated ones after
experimentally induced injuries (Nottebohm, 1985; Alvarez-
Buylla, 1990; Lois and Alvarez-Buylla, 1993; Zupanc and
Horschke, 1995; Perez-Canellas and Garcıa-Verdugo, 1996;
Chiasson et al., 1999; Dawley et al., 2000; Ino and Chiba, 2000;
Margotta et al., 2000, 2005; Ekstrom et al., 2001; Zupanc,
2001; Garcia-Verdugo et al., 2002; Alvarez-Buylla et al., 2002;
* Corresponding author. Tel.: +39 081 679209; fax: +39 081 679233.
E-mail address: [email protected] (R.K. Rastogi).
0891-0618/$ – see front matter # 2006 Elsevier B.V. All rights reserved.
doi:10.1016/j.jchemneu.2006.08.001
Rodrıguez-Perez et al., 2003; Rankin et al., 2004; Romero-
Aleman et al., 2004; Vidal Pizarro et al., 2004; Dewulf and
Bottjer, 2005). These studies have shown that neurogenesis/
gliogenesis occurs in the brain of adult teleost fishes,
amphibians, reptiles, birds and mammals. A neuroanatomically
more widespread potential for cell proliferation in the adult
brain has been suggested in some anamniote vertebrates, such
as teleost fishes (Ekstrom et al., 2001; Zupanc, 1999) and
amphibians (Richter and Kranz, 1981; Bernocchi et al., 1990;
Dawley et al., 2000; Margotta et al., 2000). In the adult
mammals, including humans, neurogenesis/gliogenesis has
been described to occur in a few circumscribed areas, viz.,
olfactory bulbs, hippocampus and cortex, although it is debated
that adult neurogenesis is far more widespread than these
studies have described (see Eriksson et al., 1988; Gould et al.,
1999; Rietze et al., 2000; Garcia-Verdugo et al., 2002;
Rodrıguez-Perez et al., 2003; Bedard and Parent, 2004).
Within the wide panorama of such studies in vertebrates,
amphibian models (urodeles and anurans) have had a rather
conspicuous role. While many authors focused their attention

F. Raucci et al. / Journal of Chemical Neuroanatomy 32 (2006) 127–142128
on the process of regeneration in the central nervous system
following lesions or thermal shocks (Minelli et al., 1982; Del
Grande et al., 1984, 1990; Franceschini et al., 1992; Benraiss
et al., 1999; Yoshino and Tochinai, 2004), several others
investigated upon the development/neurogenesis of specific
parts of brain such as the olfactory bulbs (Burd and Sein,
1998), vocal pathway (Gorlick and Kelley, 1987), cerebellum
(Filoni and Margotta, 1971; Gona, 1976; Uray et al., 1987,
1988), isthmic nucleus (Tay and Straznicky, 1980; Dann and
Beazley, 1989), optic tectum (Straznicky and Gaze, 1972;
Reznikov and Maliovanova, 1979; Dann and Beazley, 1988),
hypothalamus (Chetverukhin and Polenov, 1993; Polenov and
Chetverukhin, 1993), preoptic recess (Onishchenko et al.,
1983), telencephalon (Margotta et al., 1999b, 2000) or of the
entire brain (Bernocchi et al., 1990; Dawley et al., 2000;
Wullimann et al., 2005). These studies have demonstrated
that in the adult frogs, newts and salamanders the brain has
the potential for cell proliferation in certain localized areas
under normal and experimental conditions. It is surprising,
however, that a detailed neuroanatomical mapping of the
proliferative activity in the adult brain as well as during
development is not available for any single species. Regarding
the developmental changes in the proliferative activity within
the vertebrate brain there are only a few studies available:
cyclostomes (Villar-Cheda et al., 2002, 2005a,b), teleost fishes
(Wullimann and Puelles, 1999; Wullimann and Knipp, 2000;
Wullimann and Mueller, 2004), alligator hindbrain (Pritz,
2003) and mouse (Martinez and Puelles, 2000). Only recently
has data on the developmental pattern of the neurogenic and
proneural gene expression as well as a gross picture of the
proliferative activity in the brain of an amphibian, the African
clawed frog (Xenopus laevis), been published (Wullimann
et al., 2005). A more recent study has described the
proliferation pattern in the auditory medulla of the bullfrog
during development (Chapman et al., 2006).
Although cell proliferation in the nervous system of
vertebrates has been widely investigated by tritiated thymidine
labeling and 5-bromo-20-deoxyuridine (BrdU) immunocyto-
chemistry, a high degree of reliability consistently emphasized
for the more recent proliferating cell nuclear antigen (PCNA, an
auxillary protein to DNA-polymerase delta) has made of it the
cell cycle marker protein of choice (Rankin et al., 2004; Valero
et al., 2005). PCNA is a nuclear protein that is synthesized in the
G1 and S phases of the cell cycle and is, therefore, correlated
with the cell proliferative stage (Jaskulski et al., 1988;
Tsurimoto, 1999; Wullimann and Puelles, 1999; Rankin
et al., 2004). Additionally, PCNA analysis of cell proliferation
in the vertebrate nervous system has been found to be
comparable to the analysis using tritiated thymidine (see
Ramirez et al., 1997; Ekstrom et al., 2001). PCNA labeling has
been described in the ventricular and subventricular zones
(described as ependymal/subependymal cell areas in the adult
brain, and matrix areas in general by some other authors) of the
vertebrate brain (see Margotta and Morelli, 1996; Ino and
Chiba, 2000; Margotta et al., 1999a; Wullimann and Puelles,
1999; Wullimann et al., 2005). Thus, immunohistochemistry
for PCNA was our instrument for the present study to delineate
the developmental and adult profile of the proliferative activity
in the brain of a ranid frog, Rana esculenta.
2. Materials and methods
2.1. Animals
Tadpoles and adults of R. esculenta were collected from a pond, nearly
75 km northeast of Naples, at an altitude of 950 m ca. Tadpoles were identified
according to the staging table by Gosner (1960) and were divided into six
developmental groups as follows: stages 24–25 (hindlimb buds not yet visible)
as Group I, stages 27–28 (limb buds are visible) as Group II, stages 36–37 (toe
development) as Group III, stage 40 as Group IV, stage 42 (forelimbs emerged)
as Group V and stages 44–45 (tail resorption; metamorphic climax) as Group
VI. For immunocytochemical studies, 8–10 specimens were used for each
developmental group; only 6 adults (collected in the month of June), 3 males
and 3 females were used for this study, all in the weight range of 14 � 1 g.
Immediately after their collection, all samples were anesthetized with MS222
(tricaine methanesulfonate, Sigma Chemical Co.). Adult animals were perfused
intracardially first with cold PBS (pH 7.4) followed by Bouin’s fluid and the
brain was rapidly dissected out and immersed in Bouin’s fluid for 24 h at field/
room temperature. In tadpoles from Groups I to IV, the head was fixed in toto. In
more advanced stages of development, the brain was surgically removed from
the brain case and immersed in the fixative. Bouin-fixed material was then
dehydrated in ethanol, cleared in Bioclear, and embedded in paraffin. Sections
were cut at 5–7 mm in the sagittal or transverse plane and serially placed on
poly-L-lysin coated glass slides.
2.2. Immunohistochemistry
The paraffin sections were dewaxed in Bioclear, rehydrated in a decreasing
ethanol series, rinsed in deionized distilled water and PCNA-immunostained as
described. Briefly, the endogenous peroxidases were suppressed by incubation
of sections in 0.1% sodium azide with 0.3% hydrogen peroxide for 30 min at
room temperature. The sections were then incubated in normal goat serum
(1:20; Sigma Chemical Co., Italy) in 0.1 M phosphate buffer saline (PBS; pH
7.4) for 45 min at room temperature to reduce undesired background staining.
Intracerebral localization of PCNA was achieved by sequentially applying on
brain sections a monoclonal mouse anti-PCNA (1:10,000; Dako Corp., Den-
mark) in 10% bovine serum albumin (BSA) overnight at 4 8C in a dark moist
chamber, biotinylated secondary antibody (goat anti-mouse IgG diluted 1:500;
Sigma) for 45 min, and peroxidase-coupled streptavidin (1:200; Pierce, IL,
USA) for 1 h at room temperature. Then 3% DAB (3.30-diaminobenzidine
tetrahydrochloride; Sigma) with 0.03% hydrogen peroxide in Tris buffer
(0.05 M, pH 7.6) was used as chromogen. After each reaction step and after
DAB-peroxidase visualization, sections were washed thoroughly in PBS (two
changes of 10 min each). Sections were then dehydrated through a series of
graded alcohols, cleared in Bioclear, and coverslipped in a synthetic medium.
Control tests were done by: (a) omission of the primary antibody or (b)
substitution of the primary antiserum with PBS or normal goat serum
(1:500; Dako) in 10% BSA; no immunostaining was observed.
All experimental procedures were in line with the European Community’s
Council Directive (86/609/EEC).
2.3. Western blot analysis
Brains from tadpoles belonging to the developmental Groups III–VI, and
adults (male and female) were surgically removed under MS222 anaesthesia,
rapidly frozen in liquid nitrogen and stored at �80 8C. Frozen brains were
homogenized directly in lysis buffer containing 5 mM Tris–HCl, 150 mM
NaCl, 1 mM EDTA, 1 mM EGTA, 10% glycerol, 1% Triton X-100, 1 mM
phenylmethylsulfonyl fluoride (PMSF), 1 mg leupteptin, 0.5 mM sodium ortho-
vanadate, 20 mM sodium pyrophosphate and clarified by centrifugation at
14,000 � g for 10 min. Protein concentration was estimated using a modified
Bradford assay (Bio-Rad, Hercules, CA). Electrophoresis was performed on
SDS-PAGE by loading 30 mg proteins for each sample and proteins electro-
transferred onto nitrocellulose membrane (Immobilion Millipore Corporation,

F. Raucci et al. / Journal of Chemical Neuroanatomy 32 (2006) 127–142 129
Bedford, MA); completeness of transfer was assessed using prestained protein
standards (Bio-Rad). Nitrocellulose was treated for 2 h with a blocking solution
(5% non-fat powdered milk in 25 mM Tris, pH 7.4; 200 mM NaCl; 0.5% Triton
X-100) and incubated overnight with mouse monoclonal antibody against
recombinant PCNA, diluted 1:2000. After buffer washing the membranes were
incubated for 60 min with peroxidase-conjugated anti-mouse secondary anti-
body (1:5000; Chemicon, CA). Immunoreactivity was detected by enhanced
chemiluminescence system (Amersham Life Science, UK). The optical density
of the bands was obtained by densitometry and normalized using a SCAN
software.
Data were compared by analysis of variance followed by Duncan’s test for
multi-group comparison and the Student’s t-test for between-group comparison.
All data were expressed as mean � standard deviations. The level of signifi-
cance was taken at P < 0.01.
2.4. Photodocumentation
Digital photomicrographs were made with a Canon Digital camera on a
DMBR Leica photomicroscope. The images obtained were slightly adjusted for
brightness and contrast with Adobe Photoshop 6.0. Photomontages and lettering
were done using CorelDraw 9.0. Western blotting chart was drawn using
PowerPoint.
3. Results
3.1. Immunohistochemistry
The following description has been arranged in progressive
stages of development and adult.
3.1.1. Group I (stages 24–25)
Immediately prior to the appearance of limb buds, the
majority of cell nuclei in the developing brain were
Fig. 1. Photomicrograph of a medial sagittal section of the brain of a newly hatc
proliferative activity in the periventricular area as indicated by the abundance of PCN
distribution of PCNA-ir nuclei (arrows) in the telencephalon (A) and thalamic area (B
abbreviations as in Fig. 5. Scale bars: (inset) 100 mm; (A and B) 50 mm.
immunolabeled with the PCNA antibody (Fig. 1A and B).
All cells of the ventricular zone and the majority of those
located in the subventricular zone were strongly immunos-
tained. The brain ventricles were virtually lined with PCNA-
immunostained cells throughout the rostrocaudal axis (see inset
of Fig. 1). In other words, immunostaining occurred
uninterrupted throughout the ventricular lining, whereas more
distant cells showed decreasing intensity of immunoreactivity
and many were totally unstained. The greatest accumulation of
immunoreactive cells was seen in the pseudostratified tectum
mesencephali (optic tectum); all cells of the tectum were
immunostained with anti-PCNA.
3.1.2. Group II (stages 27–28)
As compared with the earlier group of tadpoles, the brain of
these tadpoles had undergone changes to the extent that it was
possible to make out the gross neuroanatomical limits of the
forebrain, midbrain and hindbrain regions, as well as the brain
ventricles, the developing cerebellum and the infundibular
stalk. In these tadpoles the proliferative zones of the brain could
be distinguished regionally. As yet, the PCNA-immunostained
cell lining was several layers thick in all brain ventricles (see
Fig. 2). In the telencephalon, through its entire length, the
lateral ventricles were lined with several layers (ventricular and
subventricular zones) of ir cells (Fig. 2A), indicative of a high
proliferative activity. Similarly, in the most dorsal, medial and
lateral parts of the diencephalon, several layers of ir cells lined
the third ventricle (Fig. 2B). The preoptic recess was easily
discernible (not seen in the section in Fig. 2) and was
surrounded by proliferating cells distributed in the ventricular
hed tadpole (prior to hindlimb bud; Group I) showing the occurrence of high
A-ir nuclei (arrows). Enlargements of the boxed areas in the inset showing the
). Empty triangles indicate some radially displaced ir nuclei. TH, thalamus; other
F. Raucci et al. / Journal of Chemical Neuroanatomy 32 (2006) 127–142130
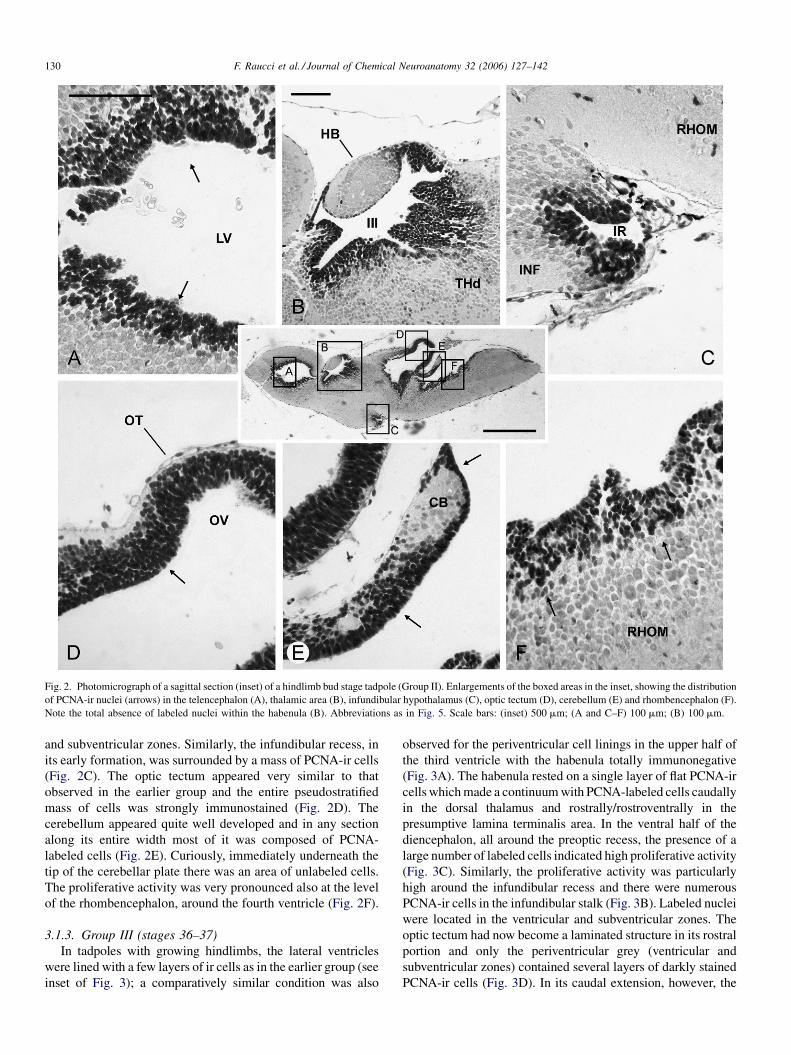
Fig. 2. Photomicrograph of a sagittal section (inset) of a hindlimb bud stage tadpole (Group II). Enlargements of the boxed areas in the inset, showing the distribution
of PCNA-ir nuclei (arrows) in the telencephalon (A), thalamic area (B), infundibular hypothalamus (C), optic tectum (D), cerebellum (E) and rhombencephalon (F).
Note the total absence of labeled nuclei within the habenula (B). Abbreviations as in Fig. 5. Scale bars: (inset) 500 mm; (A and C–F) 100 mm; (B) 100 mm.
and subventricular zones. Similarly, the infundibular recess, in
its early formation, was surrounded by a mass of PCNA-ir cells
(Fig. 2C). The optic tectum appeared very similar to that
observed in the earlier group and the entire pseudostratified
mass of cells was strongly immunostained (Fig. 2D). The
cerebellum appeared quite well developed and in any section
along its entire width most of it was composed of PCNA-
labeled cells (Fig. 2E). Curiously, immediately underneath the
tip of the cerebellar plate there was an area of unlabeled cells.
The proliferative activity was very pronounced also at the level
of the rhombencephalon, around the fourth ventricle (Fig. 2F).
3.1.3. Group III (stages 36–37)
In tadpoles with growing hindlimbs, the lateral ventricles
were lined with a few layers of ir cells as in the earlier group (see
inset of Fig. 3); a comparatively similar condition was also
observed for the periventricular cell linings in the upper half of
the third ventricle with the habenula totally immunonegative
(Fig. 3A). The habenula rested on a single layer of flat PCNA-ir
cells which made a continuum with PCNA-labeled cells caudally
in the dorsal thalamus and rostrally/rostroventrally in the
presumptive lamina terminalis area. In the ventral half of the
diencephalon, all around the preoptic recess, the presence of a
large number of labeled cells indicated high proliferative activity
(Fig. 3C). Similarly, the proliferative activity was particularly
high around the infundibular recess and there were numerous
PCNA-ir cells in the infundibular stalk (Fig. 3B). Labeled nuclei
were located in the ventricular and subventricular zones. The
optic tectum had now become a laminated structure in its rostral
portion and only the periventricular grey (ventricular and
subventricular zones) contained several layers of darkly stained
PCNA-ir cells (Fig. 3D). In its caudal extension, however, the

F. Raucci et al. / Journal of Chemical Neuroanatomy 32 (2006) 127–142 131
Fig. 3. Photomicrograph of a sagittal section (inset) of a premetamorphic tadpole with growing hindlimbs (Group III). Enlargements of boxed areas in the inset,
showing the distribution of PCNA-ir nuclei (arrows) in the dorsal thalamus (A), infundibular hypothalamus (B), preoptic area (C), rostral part of the optic tectum (D),
caudal part of the optic tectum (E) and rhombencephalon (F). Note the total absence of proliferating cells within the habenula (A) and the presence of labeled nuclei
exclusively in the periventricular grey layer of the rostrally stratified optic tectum (D) whereas the caudal part of the optic tectum is still a compact mass of labeled
nuclei (E). Abbreviations as in Fig. 5. Scale bars: (inset) 500 mm; (A and B) 100 mm; (C–F) 50 mm.
optic tectum was similar to that in the earlier stage of
development (no stratification or lamination had occurred)
and was entirely composed of a pseudostratified mass of ir cells
(Fig. 3E cf. Fig. 2D). The PCNA-labeled cells in the cerebellum
were numerous, a situation quite similar to that observed in the
earlier group of tadpoles. In the rhombencephalon, the PCNA-ir
periventricular lining, closely apposed to the posterior choroid
plexus (see Fig. 3, inset), was several cell-thick and numerous
scattered ir cells were observed at varying distances from it as if
they had migrated radially or tangentially (Fig. 3F).

F. Raucci et al. / Journal of Chemical Neuroanatomy 32 (2006) 127–142132
Fig. 4. Photomicrographs of a parasagittal (upper inset) and a midsagittal (lower inset) sections of an early prometamorphic tadpole (Group IV). Enlargements of the
boxed areas in the two insets illustrating the distribution of PCNA-labeled nuclei (arrows) in the optic tectum (A), telencephalon (B), infundibular hypothalamus (C)
and diencephalon (D). CP, pallial commissure; OC, optic chiasma; TL, lamina terminalis (terminal lamina); other abbreviations as in Fig. 5. Scale bars: (inset) 1 mm;
(A–D) 100 mm.
3.1.4. Group IV (stage 40)
In tadpoles with grown hindlimbs, the telencephalon
population of the PCNA-ir cells constituted a continuum, as
a pseudostratified periventricular lining (ventricular zone),
throughout the rostrocaudal and the dorsoventral axis of the
lateral ventricles (Fig. 4B). The optic tectum was very similar to
that observed in the tadpoles of Group III (Fig. 4A). Within the
thalamus, the majority of cells lining the ventricle dorsoven-
trally, were PCNA-ir. The lamina terminalis, now easily
distinguished, contained numerous PCNA-ir cells (Fig. 4B and
D). The proliferative activity around the preoptic recess was
very conspicuous as evidenced by the presence of a large
number of darkly stained PCNA-ir nuclei not only in the
ventricular and subventricular zones laterally and on the bottom
of the recess, but also in the magnocellular preoptic nucleus and
the suprachiasmatic nucleus (Fig. 4D). In sagittal sections, the
upward continuation of the PCNA-ir cells in the rostrolateral
ventricular zone of the preoptic recess appeared to be
interrupted by the pallial commissure; PCNA-ir cells were
again encountered along the bed nucleus of the pallial
commissure and the lamina terminalis (Fig. 4D). The posterior
tubercle is another region where there was abundance of
PCNA-ir cells denoting a high proliferative activity (Fig. 4D).
The infundibulum was comparatively more voluminous than
in the earlier stage of development and the cell layers
(ventricular and subventricular zones) lining the infundibular
recess was all PCNA-ir (Fig. 4C). The torus semicircularis area
was now easily distinguishable and its main body contained
numerous PCNA-ir cells; many ir cells surrounded it from
all directions.
3.1.5. Group V (stage 42)
In tadpoles with the forelimbs emerged, the distribution
profile of PCNA-ir nuclei within the brain was much modified
as compared to that observed in the earlier developmental
stages examined. The series of cross-sections of a brain in this
stage of development (Fig. 5) clearly showed that the
proliferating cells are located mainly along the ventricular
lining of the brain ventricles through the entire rostrocaudal
axis. It is also evident from this figure that there are regional
differences in the relative frequency of labeled cells along the
rostrocaudal or the dorsoventral axis in any given brain region.
Within the rostral telencephalon, the PCNA-ir cells forming the
pseudostratified lining of the ventral half of the lateral
ventricles appeared to be comparatively more abundant than
in the dorsal inner and lateral ventricular lining (Figs. 5A and

F. Raucci et al. / Journal of Chemical Neuroanatomy 32 (2006) 127–142 133
Fig. 5. Photomicrographs of cross-sections of the brain of a prometamorphic tadpole with forelimb emergence (Group V) at the levels indicated in the schematic
lateral view of the brain (inset). Arrows indicate the distribution of PCNA-ir nuclei. The left half of each brain section bears the labeling. ACP, anterior choroid plexus;
AMY, amygdala; AQ, aqueduct; CB, cerebellum; CG, central grey; E, epiphysis; EN, entopeduncular nucleus; HB, habenula; HR, hindbrain reticular formation; IN,
interpeduncular nucleus; INF, infundibulum; IR, infundibular recess; LEM, lemniscus; LV, lateral ventricle; MOT, medial optic tract; NA, nucleus accumbens; NI,
nucleus isthmi; ON, optic nerve; OT, optic tectum; OV, optic ventricle; PC, posterior commissure; PD, dorsal pallium; PL, lateral pallium; PM, medial pallium; POA,
preoptic area; PON, preoptic nucleus; PONm, magnocellular preoptic nucleus; POR, preoptic recess; PT, posterior tubercle; PTHN, posterior thalamic nucleus;
RHOM, rhombencephalon; SCN, suprachiasmatic nucleus; SCO, subcommissural organ; SGC, stratum griseum centrale; SGP, stratum griseum periventriculare; SL,
lateral septum; SM, medial septum; ST, striatum; TEGd, dorsal tegmentum; TEGr, rostral tegmentum; TEGv, ventral tegmentum; TEL, telencephalon; THd, dorsal
thalamus; THv, ventral thalamus; TS, torus semicircularis; TSL, laminar nucleus of the torus semicircularis; III, third ventricle; IV, fourth ventricle. Scale bar:
500 mm.
6A). At the level of the mid telencephalon, a pseudostratified
PCNA-ir ventricular lining was constantly observed particu-
larly along the bottom and the inner wall of the lateral
ventricles. Further caudally, the lateral ventricles begin to
narrow and continue more dorsolaterally in the posterior
extensions of the telencephalon; in this area there was a
conspicuous mass of PCNA-ir cells adjacent to the lateral
septum and medial septum and bulging into the lateral ventricle
(Fig. 5B). Dorsally, the lining of the lateral ventricles contained
comparatively fewer labeled cells than in an earlier stage of

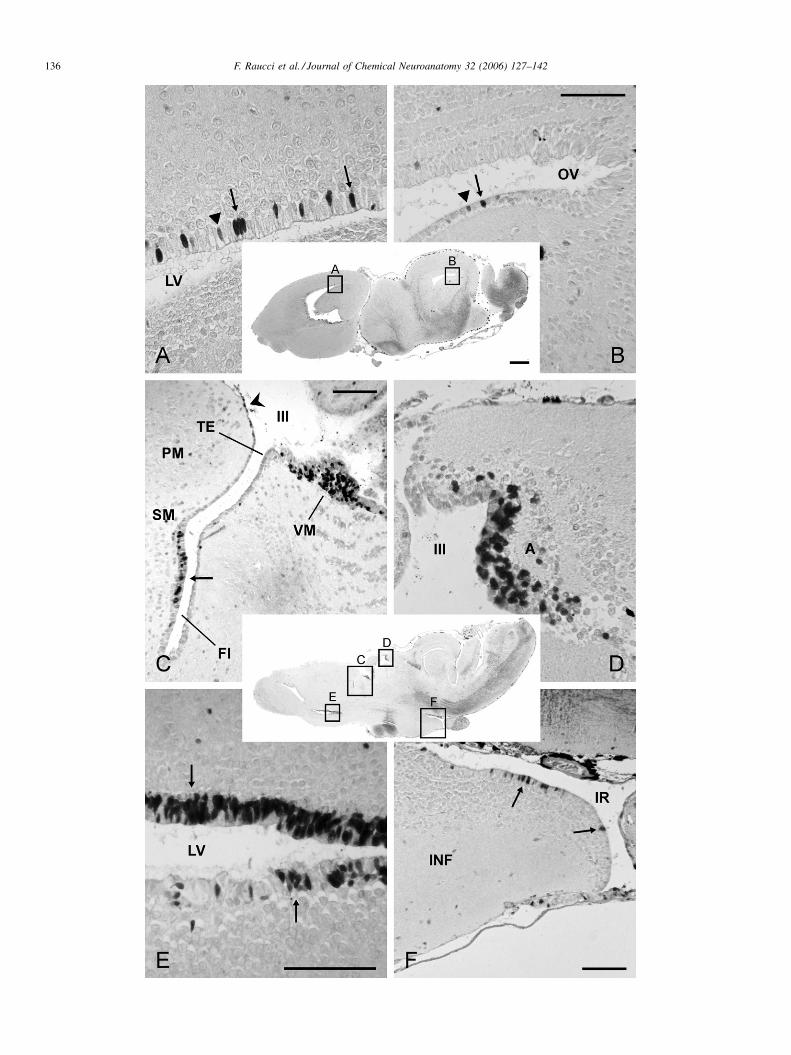
F. Raucci et al. / Journal of Chemical Neuroanatomy 32 (2006) 127–142134
Fig. 6. Photomicrographs of sagittal sections of late prometamorphic tadpoles with forelimbs grown (Group V). (B) The control reaction with no labeled nuclei. (C)
The enlargement of boxed area in (A), showing that labeled nuclei (arrows) are concentrated in the ependymal lining of the most caudal medial part of the lateral
ventricle (LV) and the dorsal part of the third ventricle (III). Note that there are no ir nuclei in the habenula (HB). (D) A parasagittal section from another brain
showing in greater details the distribution of PCNA-ir nuclei around the third ventricle, in the anterior thalamic nucleus (A, arrow), lamina terminalis (TL) and the
dorsocaudal extremity of the cerebral hemisphere (thick arrow). No labeled nuclei in the habenula. (E) From a midsagittal section of the same brain as in (D), showing
the great concentration of proliferating cells in the caudalmost optic tectum (OT), torus semicircularis (TS), tegmentum (TEG) and inner molecular (ML) and outer
granular layer (GL) of the cerebellum. (F) Midsagittal section as in (E), showing the labeled nuclei (arrows) below and around the preoptic recess in the
suprachiasmatic (SCN) and magnocellular preoptic (PONm) nuclei; the epiphysis (E) is totally negative. CP, pallial commessure; TEG, tegmentum; other
abbreviations as in Fig. 5. Scale bars: (inset) 1 mm; (C–H) 100 mm.
development; here the labeled cells did not form a continuous
lining of the ventricles. Exception to this distribution pattern
was the large number of PCNA-ir cells located at the dorsal
rostralmost extremity of the lateral ventricles (see Fig. 6A). The
ventricular lining of the third ventricle contained a large
number of PCNA-ir cells. Seen dorsoventrally, the major
portion of the cell lining of this ventricle was PCNA-ir; cells
were unlabeled in the vicinity of the habenula (Fig. 5C and D),
at a level between ventral thalamus and suprachiasmatic
nucleus (Fig. 5D), and along the midlateral of the rostral
tegmental area (Fig. 5E). No PCNA-ir cells were observed
within the habenulae (Figs. 5C and D and 6C and D); evidently
their growth is supported by the supply of new cells originating
in the anterior and anterodorsal thalamic nuclei, areas rich in
PCNA-ir cells. The preoptic recess was lined with PCNA-ir
cells in its basal, basolateral and middle portions with the ir
cells continuing into the magnocellular nucleus rostrally and
the suprachiasmatic nucleus caudally (Figs. 5B–D and 6F).
These nuclei contained comparatively less PCNA-ir cells than
in Group IV tadpoles (see Fig. 4D). At the level of the

F. Raucci et al. / Journal of Chemical Neuroanatomy 32 (2006) 127–142 135
subcommissural organ, which itself was totally devoid of any
proliferating cells, the ventricular lining at the height of the
posterior thalamic nucleus was rich in proliferating cells
(Fig. 5E). As compared to earlier stages, the ventricular lining
of the infundibular recess (mainly along the ventral hypotha-
lamus) was richly laid with PCNA-ir cells (Fig. 5E). The
periventricular cell lining of the optic ventricles showed further
progress in its process of stratification in a rostrocaudal
direction; while there were still some PCNA-ir cells in the
periventricular lining in the anterior medial and mediolateral
portions, a conspicuous mass of PCNA-ir cells was observed in
the posterior part of the optic lobes continuing downward into
the torus semicircularis (Fig. 6E). Indeed, in the caudal part of
the mesencephalon, while the lateral extremity of the optic
ventricle was lined by numerous ir cells, the big mass of PCNA-
ir cells was located medially, lining the roof and the floor (Figs.
5F and 6E). The ventromedial mass of labeled cells made a
continuum with the outer granular layer of the cerebellum
(Figs. 5F–H and 6E). In cross-sections, it was evident that the
majority of the ir cells are concentrated near the midline
(Fig. 5F). In the cerebellum, whereas the PCNA-ir molecular
layer was pseudostratified medially, more laterally the labeled
cells constituted a single layer (Figs. 5G and H and 6E). The
Fig. 7. Photomicrographs from a sagittal section of a brain from metamorphosing ta
head) of the lateral ventricle showing few ir nuclei (arrows) in the ependyma. (B) L
nuclei and the inner nuclei rather ovoidal. (C) Labeled nuclei are concentrated in th
labeled nuclei in the granular layer (GL) and pseudostratified molecular layer (ML) w
rare. Other abbreviations as in Fig. 5. Scale bars: (A, B and D) 100 mm; (C) 100
periventricular lining of the aqueduct also appeared as a
pluristratified mass of PCNA-ir cells, except in its midventral
portion (Figs. 5F and 6E). In the rhombencephalon, the lining of
the fourth ventricle still contained many ir cells (Fig. 5G and
H). In contrast with all other brain areas, however, here the
proliferating cells were mostly located laterally rather than
medially (Fig. 5G and H).
3.1.6. Group VI (stages 44–45)
During the metamorphic climax, the different brain regions
had further differentiated. The PCNA-ir cells appeared
comparatively less numerous in several parts of the brain as
compared with the relative frequency of such cells in the brain
of earlier stages of development. In the telencephalon, the cell
lining of the basal part of the lateral ventricles was still
composed of many ir cells. Dorsally, in the most rostral portion,
however, the proliferative activity had decreased to a great
extent as compared with some early stages of development
(Fig. 7A). The immunostained nuclei in this part of the brain
appeared ovoidal or elongated and many of them appeared
displaced in a radial fashion. The PCNA-ir cells do not form a
continuous lining of the lateral ventricles; the labeled nuclei
rather formed a sort of patchwork, i.e., they were intermingled
dpole (Group VI). Rostral to the left. (A) Rostralmost, dorsal extension (arrow
amina terminalis (TL) with the periventricular layer made of labeled columnar
e innermost periventricular grey layer (arrows). (D) A single layer of columnar
ith ovoidal ir nuclei. Labeled nuclei in the central Purkinje cell layer (PUL) are
mm.
F. Raucci et al. / Journal of Chemical Neuroanatomy 32 (2006) 127–142136

F. Raucci et al. / Journal of Chemical Neuroanatomy 32 (2006) 127–142 137
with immunonegative cells. The terminal lamina (lamina
terminalis) still contained a large number of PCNA-ir cells with
those in contact with the third ventricle being columnar in shape
whereas the more distant ones were rather ovoidal in shape
(Fig. 7B). The region of the neuroepithelium immediately
beneath the lamina terminalis was practically devoid of PCNA-
ir cells. In the optic tectum, the stratification of tectal cells had
advanced throughout the rostrocaudal axis and the PCNA-ir
cells were now restricted to the innermost layers and formed a
pseudostratified ventricular lining (Fig. 7C). The granular layer
of the cerebellum was now composed of a single layer of
PCNA-ir columnar cells (Fig. 7D). The inner, molecular layer
was composed of a pseudostratified palisade of ovoidal PCNA-
ir nuclei (Fig. 7D). Rare PCNA-ir cells were observed in the
central layers of the cerebellar plate. We never observed ir
nuclei belonging to Purkinje cell types.
3.1.7. Adult
In the telencephalon, the most rostrally located PCNA-ir
cells were present along the dorsal and dorsolateral extension
of the ventricular lining. Similarly, although not as numerous
as they were in premetamorphic tadpoles, ir cells were also
present in the most caudal, dorsolateral extension of the
lateral ventricles and were localized exclusively in the dorsal
ventricular lining (Fig. 8A); such cells were typically
columnar and distributed either singly or in very small groups,
of two to three cells, and there were strongly stained as well
weakly stained nuclei (Fig. 8A). In the ventromedial and
ventrocaudal extensions of the lateral ventricles the ventricular
lining was rich in ir cells; whereas the bottom lining of the
dorsoventrally flattened ventricles contained several ir cells
interspersed with immunonegative cells, the roof lining was
composed of a continuum (rather elongated cells arranged in a
pseudostratified layer) of PCNA-ir cells (Fig. 8E). Exception-
ally, the outer lining of the caudalmost part of the cerebral
hemispheres, in close contact with the third ventricle, just
above the thalamic eminence, contained a few ir cells with a
rather flattened appearance (Fig. 8C). A little ventrally, several
small and rounded PCNA-ir cells were also observed in the
posterior wall of the medial septum in the vicinity of the
interventricular foramen (Fig. 8C). Slightly elongated ir cells
were also present near the bed nucleus of the pallial
commissure (Fig. 9E). Of the entire brain, by far the most
conspicuous concentration of PCNA-ir cells was observed in
the diencephalon. Starting rostrally in the preoptic area,
besides many haphazardly scattered (laterally/rostrally/dor-
sally) ir cells, the entire magnocellular preoptic nucleus area
contained numerous ir cells (Fig. 9F). Laterally and below the
preoptic recess there were many labeled nuclei and more
caudally a rather small group of labeled cells was present in the
Fig. 8. Photomicrographs of two parasagittal sections from an adult male brain (in
(arrows) in several brain areas: (A) scattered, weakly (black triangle) or strongly (ar
weakly (black triangle) or strongly stained (arrow) PCNA nuclei in the ventral linin
thalamic nucleus (VM) just behind the thalamic eminence (TE), some ir nuclei at the
caudalmost boundary of the cerebral hemisphere (arrow head); (D) numerous ir nuc
dorsal ependyma of the ventrolateral extension of the lateral ventricle (upper arrow)
nuclei in the infundibular ependyma lining the infundibular recess (arrows). Other
suprachiasmatic nucleus in its ventralmost portion (Fig. 9F).
The postchiasmatic infundibular hypothalamus was totally
devoid of any ir cell in the most rostral extension of the
ventricular lining of the infundibular recess. Only in its caudal
extension there were some labeled cells of elongated shape
(Fig. 8F, cf. Figs. 3B and 4C). In the diencephalon,
furthermore, the anterior thalamic nucleus contained a rather
conspicuous group of PCNA-ir nuclei of variable shapes
(Figs. 8D and 9D). The habenular nuclei were totally devoid
of ir nuclei and very few solitary labeled nuclei were observed
along the ventricular lining. In the thalamus, the ventromedial
nucleus is another site where a conspicuous group of PCNA-ir
cells was observed in its rostral part (Fig. 8C). Here again, in
its immediate rostral, caudal and dorsal vicinity displaced ir
cells were also observed. In the mesencephalon, no ir nuclei
were observed along the most rostral portion of the
periventricular grey layer of the optic ventricles, neither
medially nor laterally. Caudally, however, most of the labeled
nuclei in the ventral portion of this lining were concentrated
medially, whereas only a few labeled cells were seen in the
dorsal portion (Fig. 9C) and in the lateral extensions of the
optic ventricles (Fig. 8B). There were a few solitary scattered
labeled cells in the laminar nucleus whereas no PCNA-ir cells
were observed in the main body of the torus semicircularis.
Similarly, the periventricular area of the dorsal tegmentum
contained very few labeled cells. The isthmic nuclei were
devoid of any proliferating cell. In the cerebellum, the granular
layer contained many darkly stained PCNA-ir cells (Fig. 9B);
the labeled cells were located medially while the lateral
extensions of the cerebellar plate were totally devoid of such
cells. Some ir cells, stained rather weakly, were scattered in the
medial cerebellar area. It is noteworthy that the molecular layer
totally lacks any proliferative activity. This characteristic has
been verified in all adult brains, males and females. The
proliferative activity along the lining of the fourth ventricle, in
the rhombencephalon, was drastically low in the sense that
none or only very few labeled cells were observed in any single
sagittal section.
3.2. Western blot analysis
Western blot analyses were performed on crude extracts of
the frog brain (developmental stages and adult males and
females). The anti-PCNA antibody detected a positive 36 kDa
band in all samples. The bands detected in the adult brain of
males and females were weaker and a comparatively lower
optical density was recorded in these bands (Fig. 10). Groups
corresponding to premetamorphic (Groups III–V poll and
metamorphic stages (1 and 2) showed significantly higher
values (P < 0.01).
sets). Enlargements of boxed areas in the two insets showing PCNA-ir nuclei
rows) stained PCNA nuclei in the caudalmost dorsal lateral ventricle; (B) rare,
g of the caudolateral optic ventricle; (C) strongly ir nuclei in the ventromedial
caudal extremity of the medial septum (SM) and a few small nuclei at the dorsal,
lei in the anterior thalamic nucleus (A); (E) numerous columnar ir nuclei in the
with less labeled cells in the floor ependyma (lower arrow); (F) only few labeled
abbreviations as in Fig. 5. Scale bars: (inset) 1 mm; (A–F) 100 mm.

F. Raucci et al. / Journal of Chemical Neuroanatomy 32 (2006) 127–142138
Fig. 9. Photomicrograph of a midsagittal section of the same brain as in Fig. 8. Boxed areas in (A) are enlarged in (B–F). (B) Labeled nuclei in the medial part of the
granular layer of the cerebellum (arrows); empty triangles indicate some weakly stained nuclei at some distance from the granular layer. (C) Caudal optic tectum with
ir nuclei (arrows) in the basal ependyma of the optic ventricle; weakly stained nuclei (black triangle) are sometimes observed. (D) Darkly stained ovoidal nuclei
(arrows) in the anterior thalamic nucleus and some displaced nuclei as well. (E) ir nuclei near the bed nucleus of the pallial commissure (BNCP) and a few at the
caudal boundary of the medial septum (uppermost arrow). (F) ir nuclei (arrows) are abundant in the magnocellular preoptic nucleus; there are ir nuclei just below the
preoptic recess (POR), and a small group of labeled nuclei in the posterior wall of the recess. LAM, nucleus laminaris of torus semicircularis; other abbreviations as in
Fig. 5. Scale bars: (inset) 1 mm; (A–F) 100 mm.
4. Discussion
For the past 30 years and more, the neurogenic and
regenerative capacities of the amphibian nervous system have
been investigated repeatedly involving frogs, newts and
salamanders (see Bernocchi et al., 1990; Margotta et al.,
2000; Wullimann et al., 2005). Nevertheless, the present study,
using a widely accepted and valid proliferating cell marker

F. Raucci et al. / Journal of Chemical Neuroanatomy 32 (2006) 127–142 139
Fig. 10. Western blot analyses of PCNA protein in brain extracts of tadpoles
and adults of R. esculenta. Protein (30 mg/lane) was resolved by SDS-PAGE,
transferred to nitrocellulose membrane and incubated with antibody raised
against PCNA protein. A specific band of about 36 kDa was observed by
comparison with co-migrating size markers (Bio-Rad). The amount of PCNA
was quantified by densiometry using the Scan program and normalized by total
PCNA protein. The values shown represent the mean of three separate deter-
minations. Significant values: *P < 0.01. (1) Premetamorphic developmental
stages (Groups III–V); (2) metamorphic climax (Group VI); (3) adult males; (4)
adult females.
technique, is to our knowledge the first detailed analysis of the
neuroanatomical picture of the putative proliferating areas
within the larval and adult brain of an amphibian.
That the developing and adult green frog (R. esculenta) brain
produces a PCNA protein was confirmed by its easy detection
in Western blot analysis. Although we have not used any
technique to characterize the neural or glial cell phenotype it is
implied that in our study cells stained with PCNA express
markers selective for neurons or glia. This aspect will not be
discussed here; suffice it to mention that past reports have
suggested that proliferating progenitor cells in the vertebrate
brain express neural- or glial-specific phenotypes (Alvarez-
Buylla et al., 2002; Gould and Gross, 2002; Chapman et al.,
2006).
Cell proliferation in the brain of R. esculenta continues
throughout larval development, during metamorphosis and in
the adult. In the early and late tadpoles (prior to metamor-
phosis) of R. esculenta, several areas of the brain along the
rostrocaudal course show abundance of PCNA-ir cells implying
a high proliferative potential; these areas include the caudal part
of the medial septum contiguous with the caudalmost dorsal
pallium, anterior preoptic area (magnocellular preoptic nucleus
and the suprachiasmatic nucleus in particular), posterior
tuberculum, dorsomedial caudal optic tectum contiguous with
the torus semicircularis and the tegmentum. While a long-
itudinal organization of PCNA-ir cells in telencephalon is
apparently seen, we are not in a position to unequivocally
demonstrate that this distribution pattern could fit the paradigm
of a neuromeric organization of the forebrain. The meta-
morphic climax is the period during which there occurs an
apparent reduction in the proliferative potential of the brain.
Indeed, in the adult brain a reduction in the proliferative
potential of these areas becomes evident because of a great
reduction in the relative frequency of the PCNA-labeled cells;
an exception is the anterior preoptic area where a conspicuous
mass of labeled cells was observed, in both sexes. No sexual
dimorphism as to the relative frequency or distribution of the
PCNA-labeled cells in the adult was observed by us. We have
not made a seasonal analysis of the proliferative activity in R.
esculenta brain; however, Minelli et al. (1982) had earlier
suggested that there may be seasonal fluctuations in the
magnitude of such activity.
Although a careful analysis of serial sections of R. esculenta
brain leads us to consider that the brain cells originating in the
ventricular cell lining migrate tangentially as well as radially in
all brain areas, we have no data to distinguish between cells
being born in situ rather than migrating from the ventricular
lining. Pulse-chase techniques may be required to determine
which cells may be migrating. The proliferating cells do not
always form a continuous lining around the brain ventricles;
often they are found as single or small groups of cells scattered
among the unlabeled cells. Although PCNA-ir cells were
observed throughout much of the brain during development, in
the advanced larval stages (stages 42–45) the labeled cells were
mainly confined to the ventricular/subventricular areas. The
only other amphibian species as yet investigated in this regard is
the African clawed frog, X. laevis. During the early larval life of
this frog the periventricular cell layers were described to be
composed of most cells labeled with PCNA and whose relative
abundance decreased drastically during the advanced stages of
development (Wullimann et al., 2005). Almost a decade ago,
Filoni et al. (1995) had described the presence of numerous
cycling cells around the lateral ventricles in the brain of X.
laevis larvae and observed that they became drastically low in
number in the adult brain. Unlike X. laevis, however, in R.
esculenta the PCNA-ir nuclei in the brain of tadpoles
immediately prior to and during metamorphosis as well as
of adults stages were rather abundant, although comparatively
less than in the early tadpole stages.
It is surprising that in the larval and adult R. esculenta the
nucleus isthmi was never found to contain in its neuroanato-
mical perimeter any PCNA-ir cell, a situation in contrast with
that of X. laevis (Tay and Straznicky, 1980) and Lymnodynastes
dorsalis (Dann and Beazley, 1989) in which proliferating cells
were described in the nucleus isthmi; in the nucleus isthmi of
both species active neurogenesis was described until the
midlarval life. No mention of this nucleus was made in the
paper by Wullimann et al. (2005), however. Similarities in the
occurrence of proliferating cells in the developing and/or adult
amphibian brain, as recognized by one or the other technique
(e.g., 3H-thymidine, BrdU, PCNA), have been described for the
telencephlon, preoptic recess, hypothalamus, optic tectum and

F. Raucci et al. / Journal of Chemical Neuroanatomy 32 (2006) 127–142140
cerebellum (Filoni and Margotta, 1971; Gona, 1976; Dann and
Beazley, 1988; Chetverukhin and Polenov, 1993; Margotta
et al., 2000). That cell proliferation undergoes changes in the
central nervous system was also recently confirmed in the
auditory medulla of R. catesbeiana (Chapman et al., 2006).
These authors used BrdU technique and observed that the
number of proliferating cells significantly increases during
metamorphic climax but decreases immediately thereafter in
the postmetamorphic froglets. It was also ascertained in this
study that newly generated cells in the auditory medulla express
either glial- or neural-specific phenotypes.
Our description of the distribution pattern of the proliferative
activity in the adult brain is, in general terms, consistent with
that of Bernocchi et al. (1990); in fact, using 3H-thymidine to
label dividing cells in the R. esculenta brain, these authors
described high proliferative activity in the ependymal
(ventricular) layers of the lateral ventricles, and preoptic and
infundibular recesses. Since we used PCNA immunohisto-
chemistry and Bernocchi et al. (1990) used 3H-thymidine
labeling, we do not have stringent criteria to make a detailed
comparison. These authors furnished a generalized picture of
the proliferating areas in the adult brain and provided very few
details and described that during winter the proliferative
activity is extremely low (cf. Minelli et al., 1982). It is obvious
that research needs to be done in order to recognize and
characterize the factors that progressively downregulate the
proliferative activity in the developing amphibian brain, and
those which upregulate the proliferative activity when it may be
required for regeneration and/or normal adult neurogenesis/
gliogenesis as has been ascertained in various vertebrate
groups.
A comparison with other vertebrate groups begins with the
cyclostomes. In this group of the anamniote vertebrates, cell
proliferation domains (ventricular zones) were observed in
different neuroanatomical regions of the forebrain, midbrain
and hindbrain during embryonic and larval development, with
the proliferative activity becoming extremely scarce or totally
absent in most parts of the brain in the adult (Vidal Pizarro et al.,
2004; Villar-Cheda et al., 2005a,b). The proliferative activity
continues in the spinal cord, however. Unlike the green frog (R.
esculenta), in the sea lamprey (Petromyzon marinus), the
habenula and the pineal complex showed PCNA-ir proliferating
cells (Villar-Cheda et al., 2002). Among teleost fishes, in the
Senegal sole, the PCNA-ir proliferation zones (periventricular
cell linings) in the forebrain of the premetamorphic animals
were similar to those observed by us in R. esculenta: pallium
and subpallium, preoptic region, pretectum, dorsal and ventral
thalamus and hypothalamus (Pinuela et al., 2004). During and
after metamorphosis, however, the frequency of PCNA nuclei
decreased. In some other teleosts, the proliferation zones of the
developing brain corresponded to the same areas as in the sole
(see Wullimann and Puelles, 1999; Ekstrom et al., 2001;
Wullimann and Mueller, 2004). In contrast with R. esculenta, in
some teleostean fish species, however, the molecular layer of
the cerebellum was described to contain proliferating cells
(Zupanc et al., 1996; Ekstrom et al., 2001). In the gymnotiform
teleost, Apteronotus leptorhynchus, Zupanc et al. (1996)
described that labeled cells in the molecular layer of the
cerebellum ‘‘migrate’’ to the granular layer. Whether a similar
phenomenon, including the presence of labeled cells in the
cerebellar molecular layer, occurs in adult R. esculenta during a
period of the year other than June is not known to us. Reptiles
have not been ignored in this context. In fact, not only has
neurogenesis been described in some species during develop-
ment, its occurrence in adulthood has been reported in the
major subdivisions of the lizard forebrain, including the
olfactory bulbs, and in one species even in the cerebellum (see
Perez-Canellas and Garcıa-Verdugo, 1996; Garcia-Verdugo
et al., 2002; Pritz, 2003; Romero-Aleman et al., 2004). Cell
proliferation in birds has been described in the brain during
development and adult neurogenesis has been emphasized in
several species of birds, restricted principally to the tele-
ncephalon (see Garcia-Verdugo et al., 2002; Margotta et al.,
2005; Dewulf and Bottjer, 2005). In the canary, for example,
cell proliferation in the adult brain involves most of the major
divisions of the telencephalon (a seasonally oriented neurogen-
esis occurring in the song nuclei) while it was totally absent in
the hypothalamus, septum, thalamus, cerebellum, optic tectum
and brain stem (see Nottebohm, 1985; Alvarez-Buylla, 1990).
Generation of new cells in the brain has been described
throughout embryonic and prenatal development of the
mammalian nervous system where it is dramatically reduced
after birth; in the adult brain it has been described to occur in the
subventricular zone of the lateral ventricle and of hippocampus
(see Lois and Alvarez-Buylla, 1993; Ino and Chiba, 2000;
Rodrıguez-Perez et al., 2003; Garcia-Verdugo et al., 2002;
Bedard and Parent, 2004; Rankin et al., 2004). While cell
proliferation occurs across a wide range of areas in the
developing brain, in the adult it appears to be restricted to the
subventricular (subependymal) zone which is defined to be the
major source of neural stem cells at least among the amniotes.
Thus, amphibians and teleost fishes are probably the only
vertebrates known today in which all major subdivisions of the
brain remain involved in generation of new cells in the
adulthood. In these two vertebrate groups, furthermore, the
ventricular and subventricular zones are apparently involved in
the generation of new cells. It is plausible to suggest that during
development of the frog brain, the vast majority of its
constituent cells arise from neuronal and non-neuronal
precursor cells located in the ventricular/subiventricular
(ependymal/subependymal) lining of the brain ventricles all
along the rostrocaudal and the dorsoventral axis. The normal
adult brain, male or female, as a whole, is composed of
quiescent tissue as well as proliferating cells located mainly in
the periventricular cell linings leading us to infer that there is
potential for neurogenesis and/or gliogenesis in the adult frog.
This is consistent with the repeatedly demonstrated regen-
erative capacity of the amphibian brain in the past (see
Margotta et al., 2000, for earlier literature). The persistence of
proliferative activity in the brain during adulthood may lead us
to assume that cell proliferation may also be related to changes
in the frequency of certain peptidergic neuronal phenotypes,
such as FMRFamide or gonadotropin-releasing hormone
neurons in the forebrain, midbrain and hindbrain (see Rastogi

F. Raucci et al. / Journal of Chemical Neuroanatomy 32 (2006) 127–142 141
et al., 1990, 1998, 2001; Iela et al., 1994). These considerations
imply that newly differentiated neuropeptidergic neurons in
different parts of the adult brain may derive from committed
progenitors located within the closely lying proliferation zones.
In conclusion, a detailed neuroanatomical map of the
proliferation zones of the brain during development and
adulthood may play a crucial role in future studies on whether
the presence of a prominent number of proliferating cells in the
adult brain is a sign of a sustained growth or simply a
replacement of neurons/glia lost by apoptosis.
Acknowledgements
This work was supported by the Second University of
Naples, the University of Naples Federico II and in part by
PRIN 2003 (to RKR).
References
Alvarez-Buylla, A., 1990. Mechanism of neurogenesis in adult avian brain.
Experientia 46, 948–955.
Alvarez-Buylla, A., Seri, B., Doetsch, F., 2002. Identification of neural stem
cells in the adult vertebrate brain. Brain Res. Bull. 57, 751–758.
Bedard, A., Parent, A., 2004. Evidence of newly generated neurons in the
human olfactory bulb. Dev. Brain Res. 151, 159–168.
Benraiss, A., Arsanto, J.P., Coulon, J., Thouveny, Y., 1999. Neurogenesis during
spinal cord regeneration in adult newts. Dev. Genes Evol. 209, 363–369.
Bernocchi, G., Scherini, E., Giacometti, S., Mares, V., 1990. Premitotic DNA
synthesis in the brain of the adult frog (Rana esculenta L.): an autoradio-
graphic 3H-thymidine study. Anat. Rec. 228, 461–470.
Burd, G.D., Sein, V., 1998. Influence of olfactory innervation on neurogenesis in
the developing olfactory bulb of the frog, Xenopus laevis. Ann. N. Y. Acad.
Sci. 855, 270–273.
Chapman, J.A., Weinstein, J.L., Simmons, A.M., 2006. Cell proliferation in the
Rana catesbeiana auditory medulla over metamorphic development. J.
Neurobiol. 66, 115–133.
Chetverukhin, V.K., Polenov, A.L., 1993. Ultrastructural radioautographic
analysis of neurogenesis in the hypothalamus of the adult frog, Rana
temporaria, with special reference to physiological regeneration of the
preoptic nucleus. I. Ventricular zone cell proliferation. Cell Tissue Res. 271,
341–350.
Chiasson, B.J., Tropepe, V., Morshead, C.M., van der Kooy, D., 1999. Adult
mammalian forebrain ependymal and subependymal cells demonstrate
proliferative potential, but only subependymal cells have neural stem cell
characteristics. J. Neurosci. 19, 4462–4471.
Dann, J.F., Beazley, L.D., 1988. Development of the optic tecta in the frog
Limnodynastes dorsalis. Dev. Brain Res. 44, 21–35.
Dann, J.F., Beazley, L.D., 1989. Neurogenesis and cell death in the isthmic
nuclei of the frog Limnodynastes dorsalis. J. Comp. Neurol. 283, 28–37.
Dawley, E.M., Fingerlin, A., Hwang, D., John, S.S., Stankiewicz, C.A., 2000.
Seasonal cell proliferation in the chemosensory epithelium and brain of red-
backed salamanders, Plethodon cinereus. Brain Behav. Evol. 56, 1–13.
Del Grande, P., Franceschini, V., Minelli, G., Ciani, F., 1984. Matrix area
activity in the regenerative optic tectum of Rana esculenta. Z. Mikrosk. -
Anat. Forsch. 98, 72–80.
Del Grande, P., Franceschini, V., Minelli, G., Ciani, F., 1990. Mitotic activity of
the telencephalic matrix areas following optic tectum or pallial cortex lesion
in newt. Z. Mikrosk. -Anat. Forsch. 104, 617–624.
Dewulf, V., Bottjer, S.W., 2005. Neurogenesis within the juvenile zebra finch
telencephalic ventricular zone: a map of proliferative activity. J. Comp.
Neurol. 481, 70–83.
Ekstrom, P., Carl-Magnus, J., Liss-Mari, O., 2001. Ventricular proliferation
zones in the brain of an adult teleost fish and their relation to neuromere and
migration (secondary matrix) zones. J. Comp. Neurol. 436, 92–110.
Eriksson, P.S., Perfilieva, E., Bjork-Eriksson, T., Alborn, A.M., Nordborg, C.,
Peterson, D.A., Gage, F.A., 1988. Neurogenesis in the adult human hippo-
campus. Nat. Med. 4, 1313–1317.
Filoni, S., Margotta, V., 1971. A study of the regeneration of the cerebellum of
Xenopus laevis (Daudin) in the larval stages and after metamorphosis. Arch.
Biol. (Liege) 8, 433–470.
Filoni, S., Bernardini, S., Cannata, S.M., 1995. Differences in the decrease in
regenerative capacity of various brain regions of Xenopus laevis are related to
differences in the undifferentiated cell populations. J. Brain Res. 36, 523–529.
Franceschini, V., Del Grande, P., Minelli, G., Ciani, F., 1992. 6-H3 thymidine
uptake in the central nervous system of newt following lesion and cold
shock. J. Hirnforsch. 33, 235–239.
Garcia-Verdugo, J.M., Ferron, S., Flames, N., Collado, L., Desfilis, E., Font, E.,
2002. The proliferative ventricular zone in adult vertebrates: a comparative
study using reptiles, birds, and mammals. Brain Res. Bull. 57, 765–775.
Gona, A.G., 1976. Autoradiographic studies of cerebellar histogenesis in the
bullfrog tadpole during metamorphosis: the external granular layer. J.
Comp. Neurol. 165, 77–87.
Gorlick, D.L., Kelley, D.B., 1987. Neurogenesis in the vocalization pathway of
Xenopus laevis. J. Comp. Neurol. 257, 614–627.
Gosner, K.L., 1960. A simplified table for staging anuran embryos and larvae
with notes on identification. Herpetology 16, 183–190.
Gould, E., Gross, C.G., 2002. Neurogenesis in adult mammals: some progress
and problems. J. Neurosci. 22, 619–623.
Gould, E., Reeves, A.J., Graziano, M.S.A., Gross, C.G., 1999. Neurogenesis in
the neocortex of adult primates. Science 286, 548–552.
Iela, L., D’Aniello, B., di Meglio, M., Rastogi, R.K., 1994. Influence of
gonadectomy and steroid hormone replacement therapy on the gonadotro-
pin-releasing hormone neuronal system in the anterior preoptic area of the
frog (Rana esculenta) brain. Gen. Comp. Endocrinol. 95, 422–431.
Ino, H., Chiba, T., 2000. Expression of proliferating cell nuclear antigen
(PCNA) in the adult and developing mouse nervous system. Mol. Brain
Res. 78, 163–174.
Jaskulski, D., deRiel, J.K., Mercer, W.E., Calabretta, B., Baserga, R., 1988.
Inhibition of cellular proliferation by antisense oligodeoxynucleotides to
PCNA cyclin. Science 240, 1544–1546.
Lois, C., Alvarez-Buylla, A., 1993. Proliferating subventricular zone cells in the
adult mammalian forebrain can differentiate into neurons and glia. Proc.
Natl. Acad. Sci. U.S.A. 90, 2074–2077.
Margotta, V., Morelli, A., 1996. Encephalic matrix areas and post-natal
neurogenesis under natural and experimental conditions. Anim. Biol. 5,
117–131.
Margotta, V., Morelli, A., Alfei, L., 1999a. PCNA positivity in the telencephalic
matrix areas in the adult of a lizard, Podarcis sicula. J. Brain Res. 3, 271–276.
Margotta, V., Morelli, A., Alfei, L., 1999b. PCNA positivity in the telencephalic
matrix areas in the adult of a newt, Triturus carnifex. J. Brain Res. 39, 523–
528.
Margotta, V., Morelli, A., Alfei, L., 2000. PCNA positivity in the telencephalic
matrix areas in the adult of a frog, Rana esculenta. Rend. Fis. Acc. Lincei
11, 185–196.
Margotta, V., Caronti, B., Morelli, A., Alfei, L., 2005. Signs of normal
proliferation in the telecephalon of adult male songbirds (Serinus serinus),
as shown by PCNA-positivity. Ital. J. Anat. Embryol. 110, 33–44.
Martinez, S., Puelles, L., 2000. Neurogenetic compartments of the mouse
diencephalon and some characteristic gene expression patterns. In: Goffinet,
A.M., Rakic, P. (Eds.), Mouse Brain Development. Springer, Berlin, pp.
91–106.
Minelli, G., Del Grande, P., Franceschini, P., 1982. Uptake of 6-H3 thymidine in
the normal and regenerating CNS of Rana esculenta. Z. Mikrosk. -Anat.
Forsch. 96, 201–213.
Nottebohm, F., 1985. Neuronal replacement in adulthood. Ann. N. Y. Acad. Sci.
457, 143–161.
Onishchenko, L.S., Tverskikh, N.M., Chetverukhin, V.K., 1983. Autoradio-
graphic study of the proliferative activity of the preoptic recess ependyma in
yearling and juvenile common frogs. Tsitologiia 25, 98–101.
Perez-Canellas, M.M., Garcıa-Verdugo, J.M., 1996. Adult neurogenesis in the
telencephalon of a lizard: a [3H] thymidine autoradio9graphic and bromo-
deoxyuridine immunocytochemical study. Dev. Brain Res. 93, 49–61.

F. Raucci et al. / Journal of Chemical Neuroanatomy 32 (2006) 127–142142
Pinuela, C., Rendon, C., Gonzales de Canales, M.L., Sarasquete, C., 2004.
Development of the Senegal sole, Solea senegalensis forebrain. Eur. J.
Histochem. 48, 377–384.
Polenov, A.L., Chetverukhin, V.K., 1993. Ultrastructural radioautographic
analysis of neurogenesis in the hypothalamus of the adult frog, Rana
temporaria, with special reference to physiological regeneration of the
preoptic nucleus. II. Types of neuronal cells produced. Cell Tissue Res. 271,
351–362.
Pritz, M.B., 2003. Cell proliferation during early hindbrain development in
Alligator. Brain Behav. Evol. 62, 193–200.
Ramirez, C., Nacher, J., Molowny, A., Sanchez-Sanchez, F., Irurzun, A., Lopez-
Garcia, C., 1997. Photoperiod-temperature and neuroblast proliferation-
migration in the adult lizard cortex. NeuroReport 8, 2337–2342.
Rankin, S.L., Partlow, G.D., McCurdy, R.D., Giles, E.D., Fisher, K.R., 2004.
The use of proliferating cell nuclear antigen immunohistochemistry
with a unique functional marker to detect postnatal neurogenesis in par-
affin-embedded sections of the mature pig brain. Brain Res. Protoc. 13,
69–75.
Rastogi, R.K., D’Aniello, B., Pinelli, C., Fiorentino, M., Di Fiore, M.M., di
Meglio, M., Iela, L., 2001. FMRFamide in the amphibian brain: a com-
prehensive survey. Microsc. Res. Tech. 54, 158–172.
Rastogi, R.K., di Meglio, M., Iela, L., 1990. Immunoreactive luteinizing
hormone-releasing hormone in the frog (Rana esculenta) brain: distribution
pattern in the adult, seasonal changes, castration effects, and developmental
aspects. Gen. Comp. Endocrinol. 78, 444–458.
Rastogi, R.K., Meyer, D.L., Pinelli, C., Fiorentino, M., D’Aniello, B., 1998.
Comparative analysis of GnRH neuronal systems in the amphibian brain.
Gen. Comp. Endocrinol. 112, 330–345.
Reznikov, K.I., Maliovanova, S.D., 1979. Cell proliferation and migration in the
roof of the mesencephalon (tectum) in Xenopus laevis. II. Cell proliferation
and differentiation of the tectum. Ontogenez 10, 350–358.
Richter, W., Kranz, D., 1981. Autoradiographische Untersuchungen der post-
natalen Proliferationsaktivitat in den Matrixzonen des Telencepahlons und
des Diencephalons neim Axolotl (Ambystoma mexicanum), unter Beruck-
sichtigung der Proliferation im ilfactorischen Organ. Z. Mikrosk. -Anat.
Forsch. 95, 883–904.
Rietze, R., Poulin, P., Weiss, S., 2000. Mitotically active cells that generate
neurons and astrocytes are present in multiple regions of the adult mouse
hippocampus. J. Comp. Neurol. 424, 397–408.
Rodrıguez-Perez, L.M., Perez-Martın, M., Jimenez, A.J., Fernandez-Llebrez,
P., 2003. Immunocytochemical characterisation of the wall of the bovine
lateral ventricle. Cell Tissue Res. 314, 325–335.
Romero-Aleman, M.M., Monzon-Mayer, M., Yanes, C., Lang, D., 2004. Radial
glial cells, proliferative periventricular cells, and microglia might contribute
structural repair in the cerebral cortex of the lizard Gallotia galloni. Exp.
Neurol. 188, 74–85.
Straznicky, K., Gaze, R.M., 1972. The development of the tectum in Xenopus
laevis: an autoradiographic study. J. Embryol. Exp. Morphol. 28, 87–115.
Tay, D., Straznicky, C., 1980. The development of the nucleus isthmi in
Xenopus: an autoradiographic study. Neurosci. Lett. 16, 313–318.
Tsurimoto, T., 1999. PCNA binding proteins. Front. Biosci. 4, 849–858.
Uray, N.J., Gona, A.G., Hauser, K.F., 1987. Autoradiographic studies of
cerebellar histogenesis in the premetamorphic bullfrog tadpole: I. Genera-
tion of the external granular layer. J. Comp. Neurol. 266, 234–246.
Uray, N.J., Gona, A.G., Hauser, K.F., 1988. Autoradiographic studies of
cerebellar histogenesis in the premetamorphic bullfrog tadpole: II. Forma-
tion of the interauricular granular band. J. Comp. Neurol. 269, 118–129.
Valero, J., Weruaga, E., Murias, A.R., Recio, J.S., Alonso, J.R., 2005. Pro-
liferation markers in the adult rodent brain: bromodeoxyuridine and pro-
liferating cell nuclear antigen. Brain Res. Protoc. 15, 127–134.
Vidal Pizarro, I., Swain, G.P., Selzer, M.E., 2004. Cell proliferation in the
lamprey central nervous system. J. Comp. Neurol. 469, 298–310.
Villar-Cheda, B., Perez-Costas, E., Melendez-Ferro, M., Abalo, X.M., Rodrı-
guez-Munoz, R., Anadon, R., Rodicio, M.C., 2002. Proliferating cell
nuclear antigen (PCNA) immunoreactivity and development of the pineal
complex and habenula of the sea lamprey. Brain Res. Bull. 57, 285–287.
Villar-Cheda, B., Abalo, X.M., Anadon, R., Rodicio, M.C., 2005a. The teg-
mental proliferation region in the sea lamprey. Brain Res. Bull. 66, 431–435.
Villar-Cheda, B., Perez-Costas, E., Melendez-Ferro, M., Abalo, X.M., Rodri-
guez-Munoz, R., Anadon, R., Rodicio, M.C., 2005b. Cell proliferation in the
forebrain and midbrain of the sea lamprey. J. Comp. Neurol. 494, 986–1006.
Wullimann, M.F., Knipp, S., 2000. Proliferation pattern changes in the zebrafish
brain from embryonic through early postembryonic stages. Anat. Embryol.
202, 385–400.
Wullimann, M.F., Mueller, T., 2004. Identification and morphogenesis of the
eminentia thalami in the zebrafish. J. Comp. Neurol. 471, 37–48.
Wullimann, M.F., Puelles, L., 1999. Postembryonic neural proliferation in the
zebrafish forebrain and its relationship to prosomeric domains. Acta
Embryol. 199, 329–348.
Wullimann, M.F., Rink, E., Vernier, P., Schlosser, G., 2005. Secondary neu-
rogenesis in the brain of the African clawed frog, Xenopus laevis, as
revealed by PCNA, delta-1, neurogenin-related-1, and NeuroD expression.
J. Comp. Neurol. 489, 387–402.
Yoshino, J., Tochinai, S., 2004. Successful reconstitution of the non-regenerat-
ing adult telencephalon by cell transplantation in Xenopus laevis. Dev.
Growth Differ. 46, 523–534.
Zupanc, G.K.H., 1999. Neurogenesis, cell death and regeneration in the adult
gymnotiform brain. J. Exp. Biol. 202, 1435–1446.
Zupanc, G.K.H., 2001. Adult neurogenesis and neuronal regeneration in the
central nervous system of teleost fish. Brain Behav. Evol. 58, 250–275.
Zupanc, G.K.H., Horschke, I., 1995. Proliferation zones in the brain of adult
gymnotiform fish: a quantitative mapping study. J. Comp. Neurol. 353, 213–
233.
Zupanc, G.K.H., Horschke, I., Ott, I., Rascher, G.S., 1996. Postemrbyonic
development of the cerebellum in gymnotiform fish. J. Comp. Neurol. 370,
443–464.
Copyright © 2022 FDOKUMEN














![[Proliferative lupus nephritis treatment: practice survey in nephrology and internal medicine in France]](https://static.fdokumen.com/doc/165x107/6336db7d20d9c9602f0b0be8/proliferative-lupus-nephritis-treatment-practice-survey-in-nephrology-and-internal.jpg)





