
Production, purification and characterisation of thermostable metallo-protease from newly isolated...
11
1 Department of Biology, Science Faculty, Dicle University, 21280, Diyarbakir, Turkey 2 Department of Primary Education, Ziya Gökalp Education Faculty, Dicle University, 21280, Diyarbakir, Turkey *Corresponding author: [email protected] Production, purification and characterisation of thermostable metallo-protease from newly isolated Bacillus sp. KG5 1 EurAsian Journal of BioSciences Eurasia J Biosci 9, 1-11 (2015) http://dx.doi.org/10.5053/ejobios.2015.9.0.1 Abstract Background: Due to the importance of microbial proteases in biotechnological applications, a number of microorganisms are being explored. The production, purification and characterisation of extracellular metallo-proteases by producing Bacillus sp. KG5 was studied. Material and Methods: Bacterial strain KG5 was isolated from Kös (Bingöl) hot spring. The strain KG5 was identified by morphological, physiological, biochemical and 16S rRNA gene sequencing. The effects of various parameters on protease production, such as time, temperature, pH, carbon and nitrogen sources and CaCl2 were studied. The enzyme was purified by ammonium sulphate precipitation and Sephadex G-75 gel permeation chromatography. Molecular weight was calculated by sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) and zymographic analysis. The effects of some metal ions, chelators and inhibitors on enzyme activity were determined. Results: The optimum temperature, pH and incubation period for protease production were 40- 45°C, 7.0 and 24 h, respectively. It was determined that the best nitrogen sources were yeast extract and urea, while the best carbon sources were lactose and galactose. However, glucose as a source of carbon was found to inhibit the production of the enzyme. The maximum enzyme production was increased in the presence of CaCl2. The molecular weight of purified enzyme was found to be approximately 48 kDa. It was found that the enzyme was fully stable in the presence of 2 mM CaCl2 at 50°C after 120 min. Purified protease was significantly activated by Ca 2+ and Mg 2+ , while it was greatly inhibited by Cu 2+ , Zn 2+ , Hg 2+ and SDS as well as by the metal ion chelators ethylenediaminetetraacetic (EDTA) and 1,10-phenanthroline. Phenylmethylsulfonyl fluoride (PMSF) had a little effect on the enzyme. Conclusions: Our findings suggest the potential of this isolate for protease production and that this enzyme may be suitable for biotechnological applications. Keywords: Bacillus sp. KG5, biotechnology, protease production and characterisation. Abbreviations: BM: Basal medium; EDTA: Ethylenediaminetetraacetic; NB: Nutrient broth; OD: Optical density; phen: 1,10-phenanthroline; PMSF: Phenylmethylsulfonyl fluoride; SDS-PAGE: sodium dodecyl sulphate- polyacrylamide gel electrophoresis; TCA: trichloroacetic acid. Ahmetoglu N, Matpan Bekler F, Acer O, Guven RG, Guven K (2015) Production, purification and characterisation of thermostable metallo-protease from newly isolated Bacillus sp. KG5. Eurasia J Biosci 9: 1-11. http://dx.doi.org/10.5053/ejobios.2015.9.0.1 Nazenin Ahmetoglu 1 , Fatma Matpan Bekler 1* , Omer Acer 1 , Reyhan Gul Guven 2 , Kemal Guven 1 © EurAsian Journal of BioSciences Proteases are by far the most important groups of commercially and biotechnological enzymes, produced by various organisms such as bacteria, yeasts, moulds, plants and animal tissues, accounting for nearly 65% of the global industrial enzyme market (Anwar and Saleemuddin 1997, Banik and Prakash 2004, Annamalai et al. 2014). Microbial proteases, especially from Bacillus species, are the major industrial workhorses, and the use of proteases in several applications has increased in the last decade (Joo and Chang 2006). Proteases are used in a number of applications such as bioreme- diation, biosynthesis and biotransformation, brewing, dairy industries, detergent, diagnostics, food, meat, leather, photographic, and INTRODUCTION Received: January 2015 Received in revised form: February 2015 Accepted: February 2015 Printed: March 2015
Transcript of Production, purification and characterisation of thermostable metallo-protease from newly isolated...

1Department of Biology, Science Faculty, Dicle University, 21280, Diyarbakir, Turkey2Department of Primary Education, Ziya Gökalp Education Faculty, Dicle University, 21280, Diyarbakir, Turkey
*Corresponding author: [email protected]
Production, purification and characterisation ofthermostable metallo-protease from newly isolatedBacillus sp. KG5
1
EurAsian Journal of BioSciencesEurasia J Biosci 9, 1-11 (2015)http://dx.doi.org/10.5053/ejobios.2015.9.0.1
AbstractBackground: Due to the importance of microbial proteases in biotechnological applications, anumber of microorganisms are being explored. The production, purification and characterisation ofextracellular metallo-proteases by producing Bacillus sp. KG5 was studied.Material and Methods: Bacterial strain KG5 was isolated from Kös (Bingöl) hot spring. The strainKG5 was identified by morphological, physiological, biochemical and 16S rRNA gene sequencing.The effects of various parameters on protease production, such as time, temperature, pH, carbonand nitrogen sources and CaCl2 were studied. The enzyme was purified by ammonium sulphateprecipitation and Sephadex G-75 gel permeation chromatography. Molecular weight was calculatedby sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) and zymographicanalysis. The effects of some metal ions, chelators and inhibitors on enzyme activity weredetermined.Results: The optimum temperature, pH and incubation period for protease production were 40-45°C, 7.0 and 24 h, respectively. It was determined that the best nitrogen sources were yeast extractand urea, while the best carbon sources were lactose and galactose. However, glucose as a sourceof carbon was found to inhibit the production of the enzyme. The maximum enzyme productionwas increased in the presence of CaCl2. The molecular weight of purified enzyme was found to beapproximately 48 kDa. It was found that the enzyme was fully stable in the presence of 2 mM CaCl2
at 50°C after 120 min. Purified protease was significantly activated by Ca2+ and Mg2+, while it wasgreatly inhibited by Cu2+, Zn2+, Hg2+ and SDS as well as by the metal ion chelatorsethylenediaminetetraacetic (EDTA) and 1,10-phenanthroline. Phenylmethylsulfonyl fluoride (PMSF)had a little effect on the enzyme.Conclusions: Our findings suggest the potential of this isolate for protease production and that thisenzyme may be suitable for biotechnological applications.Keywords: Bacillus sp. KG5, biotechnology, protease production and characterisation.
Abbreviations: BM: Basal medium; EDTA: Ethylenediaminetetraacetic; NB: Nutrient broth; OD: Optical density;phen: 1,10-phenanthroline; PMSF: Phenylmethylsulfonyl fluoride; SDS-PAGE: sodium dodecyl sulphate-polyacrylamide gel electrophoresis; TCA: trichloroacetic acid.
Ahmetoglu N, Matpan Bekler F, Acer O, Guven RG, Guven K (2015) Production, purification andcharacterisation of thermostable metallo-protease from newly isolated Bacillus sp. KG5. Eurasia JBiosci 9: 1-11.
http://dx.doi.org/10.5053/ejobios.2015.9.0.1
Nazenin Ahmetoglu1, Fatma Matpan Bekler1*, Omer Acer1, Reyhan Gul Guven2,Kemal Guven1
©EurAsian Journal of BioSciences
Proteases are by far the most important groups of commercially and biotechnological enzymes, produced by various organisms such as bacteria, yeasts, moulds, plants and animal tissues, accounting for nearly 65% of the global industrial enzyme market (Anwar and Saleemuddin 1997, Banik and Prakash 2004, Annamalai et al. 2014). Microbial proteases, especially from Bacillus species, are the major industrial workhorses, and the use of
proteases in several applications has increased in the last decade (Joo and Chang 2006). Proteases are used in a number of applications such as bioreme-
diation, biosynthesis and biotransformation, brewing, dairy industries, detergent, diagnostics, food, meat, leather, photographic, and
INTRODUCTION
Received: January 2015Received in revised form: February 2015
Accepted: February 2015Printed: March 2015

pharmaceutical industries, in the production of protein hydrolysates, silk, and silver recovery, and in X-ray films (Joo and Chang 2006, Hmidet et al. 2009, Jellouli et al. 2011).
Many bacteria in the genus Bacillus, produce large amounts of protease extracellularly during the post-exponential and stationary growth phases into the culture medium (Lin et al. 2015). Several Bacillus species are involved in protease production i.e., Bacillus clausii (Christiansen and Nielsen 2002, Joo and Chang 2006, Oskouie et al. 2008), Bacillus cereus (Banik and Prakash 2004, Uyar et al. 2011), Bacillus circulans (Subba Rao et al. 2008, Benkiar et al. 2013), Bacillus lehensis (Joshi and Satyanarayana 2013), Bacillus licheniformis (Haki and Rakshit 2003, Sellami-Kamoun et al. 2008, Jellouli et al. 2011, Lin et al. 2015), Bacillus megaterium (Asker et al. 2013), Bacillus mojavensis (Beg and Gupta 2003, Haddar et al. 2009), Bacillus pumilus (Jaouadi et al. 2008), Bacillus sterothermophilus (Sookkheo et al. 2000), and Bacillus subtilis (Akcan and Uyar 2011, Maruthiah et al. 2013).
Extracellular microbial protease production is
known to be highly affected by media components
and conditions. In the present study, we describe the
optimal production, purification and characte-
risation of a new extracellular protease produced by
newly isolated Bacillus sp. KG5 from hot water spring
in Kös (Bingöl ) Turkey, which has not been previously
reported.
Bacterial strain and cell cultivation
The samples were collected from the mud of Kös
hot spring in Bingöl which is a city in the southeast of
Turkey and identified by Ahmetoglu et al. (2011).
Cultures were purifed from samples in the
enrichment media at 80°C, using the serial dilution
technique. In order to screen a potent proteolytic
bacteria, the selected colonies, which appeared on
the plates after incubation, were patched on skim
milk agar plate consisting of 0.1% peptone (w/v),
0.5% NaCl, 10% skim milk and 2% agar (pH 7.0) and
incubated at 60°C. Colonies with a surrounding clear
zone in skim milk agar were selected as protease
positive and streaked in a new skim milk agar plate
for further study. The isolate KG5 was characterised
morphologically, physiologically and biochemically
and using 16S rRNA gene sequence analysis. To
determine the temperature and pH range for
growth, the isolate was incubated at 15 to 75°C and
4.0-11.0 respectively. Cellular morphology was
determined by phase-contrast microscopy (Zeiss),
and colony morphology was determined by a Leica
M8 stereomicroscope using cultures grown on
Nutrient Broth (NB) agar (2%) plates for 24 h at 50°C.
Gram staining was carried out according to Dussault
(1955). Unless otherwise stated, the strain was
characterised using the modified methods of Gordon
and Pang (1973).
Genomic DNA extraction, PCR-mediated amplification of the 16S rDNA and purification of the PCR products were carried out as described by Rainey et al. (1994).
Cultures were grown in 100 mL Erlenmeyer flasks
containing 25 mL of basal medium (BM) containing
2% soluble starch, 0.2% yeast extract, 1% beef
extract, 0.02% CaCl2, and 0.01% MgSO4.7H2O. The
flasks were inoculated with 250 μL of a cell
suspension (1.1x108 cells/mL) and stirred in a water
bath at 120 rpm at 40°C for 24 h. After the desired
growth time, the cells were harvested by
centrifugation at 8,200×g for 10 min at 4°C. The
supernatant served as the enzyme source.
Enzyme assay
Proteolytic activity with azocasein (Sigma-
Aldrich) was determined according to the method of
Leighton et al. (1973). Here, 150 μL enzyme was
mixed with 250 μL 0.5% azocasein solution prepared
in 0.1 M Tris-HCl buffer pH 7.0. Then reaction mixture
was incubated at 45°C for 30 min. After the end of
incubation period, the reaction was stopped by
adding 1 mL of 10% trichloroacetic acid (TCA). After
the mixture was incubated for 15 min at 4°C, the
samples were centrifuged at 8,200×g for 10 min at
4°C. One milliliter supernatant was added in 500 μL
of 1.8 N NaOH solution and measured at 420 nm.
One Unit (U) is defined as the amount of enzyme
Ahmetoglu et al.
2
EurAsian Journal of BioSciences 9: 1-11 (2015)
MATERIALS AND METHODS

that breaksdown the amino acid of 1 micromole of
azocasein per minute. All protein content was
assayed according to the Lowry method (Lowry et al.
1951).
Effect of different incubation periods on
enzyme production
In order to determine the effect of different
incubation periods on enzyme production, the
cultures were grown in BM for 3-108 h. After
incubation, growth was measured at OD470 then the
samples were centrifuged at 8,200×g; the
supernatant was used for the protease activity.
Effect of various carbon and nitrogen sources
on enzyme production
In order to determine the effect of various carbon
and nitrogen sources on enzyme production, 250 μL
fresh medium with the isolate was inoculated into
soluble starch-free BM containing 2% concentration
of various carbon sources (glucose, sucrose, maltose,
lactose, galactose, fructose and glycerol ) and 1.2%
nitrogen sources (yeast extract, beef extract,
peptone, gelatine, urea, ammonium sulphate,
ammonium chloride and tryptone) in 100 mL
Erlenmeyer.
Effect of some metal ions on the enzyme
production
For the effects of some metal ions on the enzyme
production, the isolate was inoculated into BM
medium containing 0.5% CaCl2, NaCl, MgCl2, and
MnCl2 and incubated at 40°C in a shaker for 24 h.
Growth was followed by optical density
measurements at 470 nm. The cultures were
centrifuged at 8,200×g for 10 min at 4°C and the cell-
free supernatants were used for the proteolytic
enzyme activities.
Purification procedure of protease from
Bacillus sp. KG5
The crude extract with protease activity was
precipitated by ammonium sulphate added slowly
over period of time on ice with a constant stirring up
to a final concentration of 70% (w/v). The
centrifuged precipitate (8,200×g 20 min, 4°C) was re-
dissolved in a small volume of 0.05 M pH 7.0 Tris-HCl
buffer, dialysed twice at 4°C against 1 L volume of
the same buffer overnight. The dialysed enzyme
samples were loaded on a Sephadex G-75 (1.5X30
cm) column equilibrated with 0.1 M Tris-HCl, pH 7.0,
and eluted with the same buffer at a flow rate of 3
mL/min. The peaks exhibiting protease activity were
pooled together and the purified enzyme was used
for further characterisation. All purification
procedures were carried out at 4°C.
Properties of the purified protease
Sodium dodecyl sulphate-polyacrylamide gel
(SDS-PAGE) electrophoresis and zymography
SDS-PAGE was carried out for the determination
of purity and molecular weight of the protease as described by Laemmli (1970). SDS-denatured β-
galactosidase (116 kDa), fructose phosphate (80.5 kDa), and α-amylase (58 kDa) were used as standard protein molecular weight markers. For the zymogram of protease activity, the sample was not heated before electrophoresis. After electrophoresis, the gel containing 0.1% gelatine was submerged in 0.1 M Tris-HCl (pH 7.0) containing 2.5% Triton X-100 and 5mM CaCl2 for 45 min to
remove the SDS. Then, Triton X-100 was removed by
washing the gel with 0.1 M Tris-HCl buffer (pH 7.0).
Finally, the gel was stained with Coomassie Brilliant
Blue R-250. The development of clear zones on the
blue background of the gel indicated the presence of
protease activity.
Effect of pH and temperature on the activity of
the enzyme
The effect of pH on the purified protease activity
was performed at 45°C in 0.1 M Na-phosphate buffer
(6.0-6.5), 0.1 M Tris-HCl buffer (7.0-9.0) and 0.1 M
NaOH-Glycine (9.0-11.0) for 30 min. The effect of
temperature on activity was determined by assaying
enzyme activity between 20 and 70°C for 30 min.
Thermal stability of purified enzyme
The thermostability of the purified enzyme was
tested by pre-incubating the enzyme preparation in
the presence of 2 mM CaCl2 at 50°C for 120 min and
then determining the residual activity at regular
intervals of 30 min.
Effect of metal ions, inhibitors and detergents
on protease activity
To investigate of some metal ions (Cu2+, Hg2+,
Mn2+, Zn2+, Ca2+, and Mg2+), various chelating agents
and inhibitors such as ethylenediaminetetraacetic
3
Ahmetoglu et al.EurAsian Journal of BioSciences 9: 1-11 (2015)

acid (EDTA) and 1,10-phenanthroline (phen), phenyl-
methanesulfonylfluoride (PMSF) and some different
concentrations (0.1-1%) of detergents (SDS, Triton
X-100, Tween-80 and commercially detergent Alo)
on the purified protease activity, the enzyme was
pre-incubated with all agents for 15 min and then
the remaining activities were measured using the
enzyme assay under standard assay conditions.
Activity in the absence of the agents was taken as
the control (100%). Divalent metals, chelating
agents, chemicals, and detergents were dissolved in
0.1 M Tris-HCl buffer (pH 7.0). Only PMSF was
dissolved in ethanol and phen was dissolved in
methanol.
Isolation and characterisation of strain KG5
The strain KG5 was rod-shaped, Gram-positive, facultative anaerobic, motile and possessing oval and central endospores (Fig. 1). KG5 grows at a temperature of 15-45°C (optimum 40°C), meaning that it is considered a mesophilic strain. Due to the negative catalase and glucose utilisation, KG5 was found to be different from other Bacillus cereus strains (Table 1). The GenBank/EMBL/DDBJ accession number for the 16S rRNA sequence of Bacillus sp. KG5 is KP318029. The 16S rRNA gene sequence analysis of Bacillus sp. KG5 showed high pairwise sequence similarity to B. cereus ATCC 14579T (100%), Bacillus anthracis ATCC 14578T (99.9%) Bacillus thrungiensis ATCC 10792T (99.7%) and Bacillus toyonensis BCT-7112T.
Since no studies have been performed on the Kös
hot water spring, our study is significant for the
isolation of new microorganisms which are of
biotechnological importance. In addition, this study
presents novelty due to the isolation, purification
and characterisation of metallo-protease obtained
from Bacillus species.
Effects of different incubation periods on the
protease production
As can be seen from Fig. 2, the time course
experiments of protease synthesis in the cells show
that there is a sharp increase up to 24 h (7235.4
U/mL) at 40°C. Since it is important to produce
enzyme within a short time in the field of biotechnology, a 24 h incubation time is very important in terms of biotechnology. Shafee et al.
(2005), Sousa et al. (2007), Haddar et al. (2009) and Shivanand and Jarayaman (2009) reported the optimum protease production at 24 h for Bacillus cereus, Bacillus mojavensis A21 and Bacillus aquimaris VITP4, respectively.
Effects of various carbon and nitrogen sources
on the protease production
As shown in Fig. 3, lactose (8430.2 U/mL) as a carbon source increased the enzyme production to a great extent, while others except galactose inhibited enzyme production. It is already known that enzyme production is regulated by physiological mechanisms and the catabolites of glucose (catabolite repression) in liquid culture often repress the production of hydrolytic enzyme (Zambare et al. 2011). Similar results were reported by Johnvesly and Naik (2001) and VijayAnand et al.
(2009). Mabrouk et al. (1999), Oh et al. (2000) and Ibrahim and Al-Salamah (2009) all stated that lactose increases protease production. As shown in Fig. 4, yeast extract (9318 U/mL) increased the enzyme production to a great extent, while peptone (4729 U/mL) and tryptone (900 U/mL) inhibited enzyme production compared with the control (6420 U/mL). Among nitrogen sources, urea (7584 U/mL) caused a slight increase in enzyme activity. Johnvesly and Naik (2001), Frika et al. (2005) and Abidi et al. (2008) also reported that the maximum protease production was obtained in the presence of yeast extract.
The neutral protease produced from Bacillus sp.
KG5 can be conceived as a possible candidate for the
cost-effective enzyme due to the use of cheap
substrates such as lactose and yeast extract. In
addition, ammonium salts have no repressive effect
on protease production; this feature is important for
enzyme purification steps.
Effects of some metal ions on the protease
production
It has been found that protease production was
higly stimulated by CaCl2 (15400 U/mL), whereas it
was inhibited by NaCl (208 U/mL), MgCl2 (1742 U/mL)
Ahmetoglu et al.
4
EurAsian Journal of BioSciences 9: 1-11 (2015)
RESULTS AND DISCUSSION
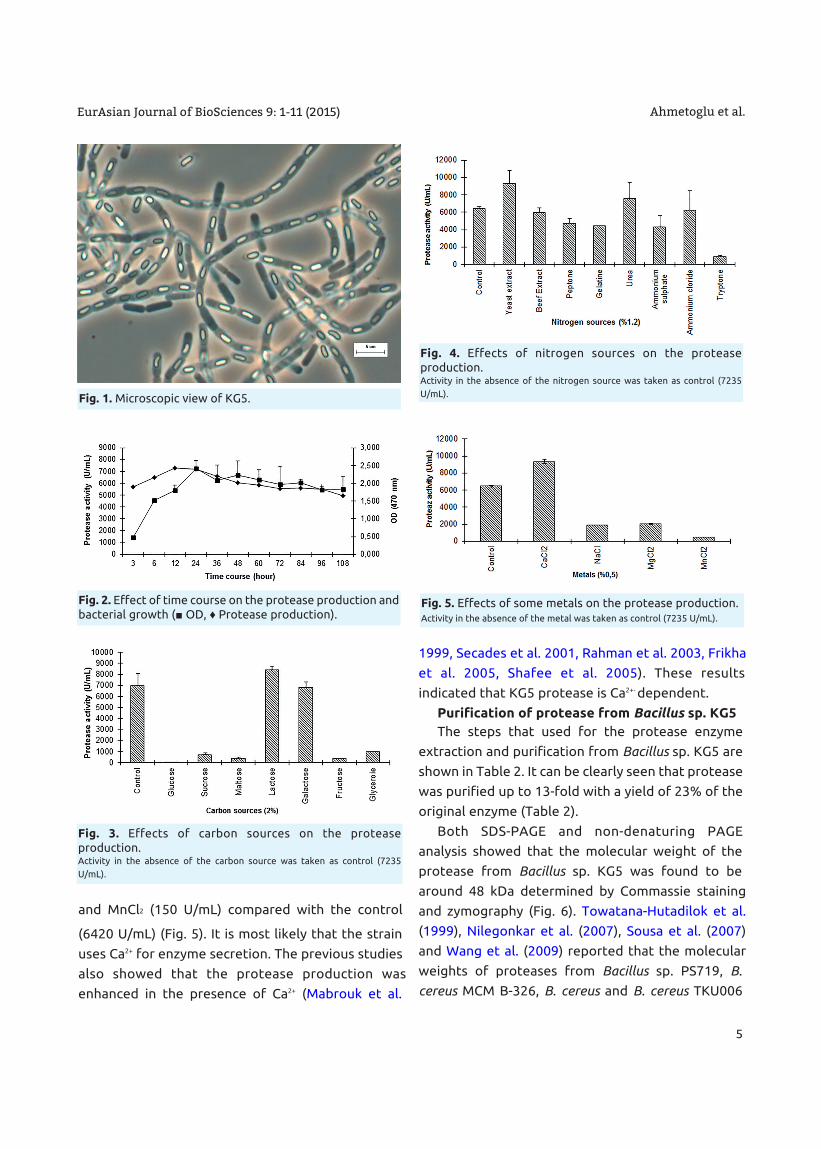
and MnCl2 (150 U/mL) compared with the control
(6420 U/mL) (Fig. 5). It is most likely that the strain
uses Ca2+ for enzyme secretion. The previous studies
also showed that the protease production was
enhanced in the presence of Ca2+ (Mabrouk et al.
1999, Secades et al. 2001, Rahman et al. 2003, Frikha
et al. 2005, Shafee et al. 2005). These results
indicated that KG5 protease is Ca2+- dependent.
Purification of protease from Bacillus sp. KG5
The steps that used for the protease enzyme
extraction and purification from Bacillus sp. KG5 are
shown in Table 2. It can be clearly seen that protease
was purified up to 13-fold with a yield of 23% of the
original enzyme (Table 2).
Both SDS-PAGE and non-denaturing PAGE
analysis showed that the molecular weight of the
protease from Bacillus sp. KG5 was found to be
around 48 kDa determined by Commassie staining
and zymography (Fig. 6). Towatana-Hutadilok et al.
(1999), Nilegonkar et al. (2007), Sousa et al. (2007)
and Wang et al. (2009) reported that the molecular
weights of proteases from Bacillus sp. PS719, B.
cereus MCM B-326, B. cereus and B. cereus TKU006
5
Ahmetoglu et al.EurAsian Journal of BioSciences 9: 1-11 (2015)
Fig. 1. Microscopic view of KG5.
Fig. 2. Effect of time course on the protease production andbacterial growth (■ OD, ♦ Protease production).
Fig. 3. Effects of carbon sources on the proteaseproduction. Activity in the absence of the carbon source was taken as control (7235
U/mL).
Fig. 4. Effects of nitrogen sources on the proteaseproduction.Activity in the absence of the nitrogen source was taken as control (7235
U/mL).
Fig. 5. Effects of some metals on the protease production. Activity in the absence of the metal was taken as control (7235 U/mL).
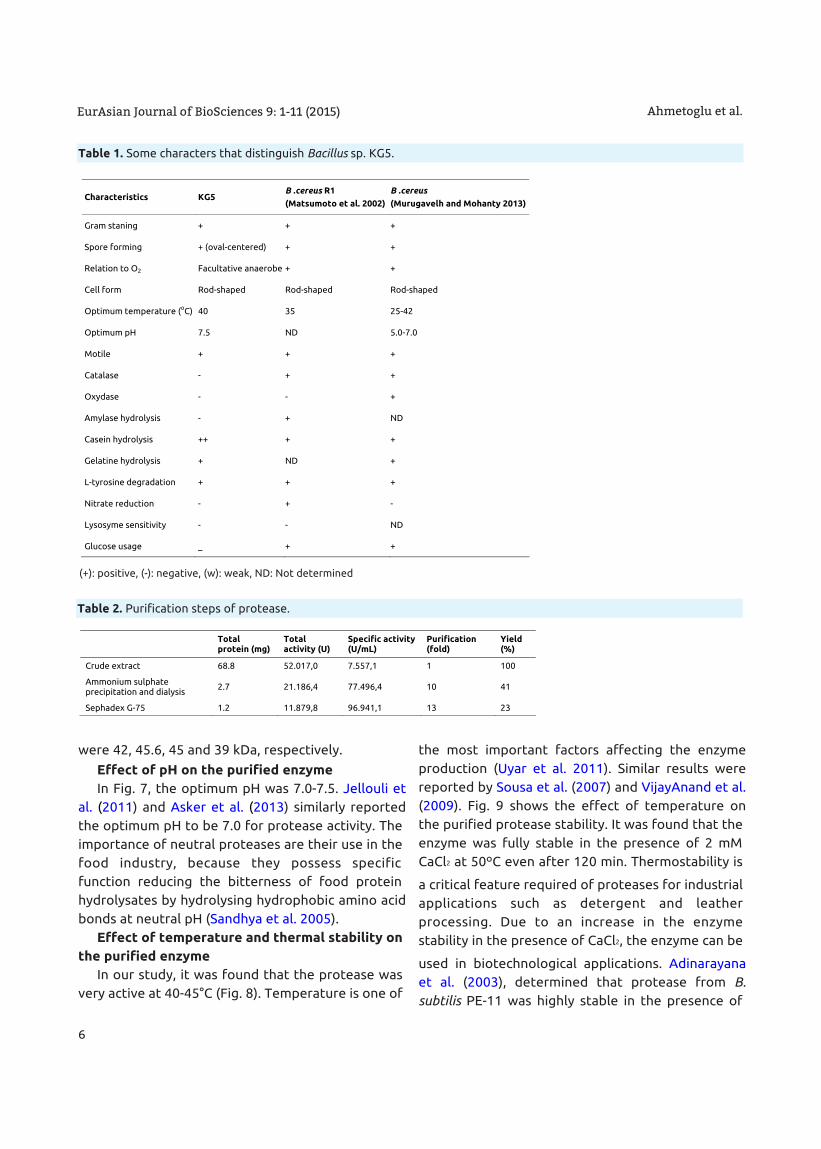
were 42, 45.6, 45 and 39 kDa, respectively.
Effect of pH on the purified enzyme
In Fig. 7, the optimum pH was 7.0-7.5. Jellouli et al. (2011) and Asker et al. (2013) similarly reported the optimum pH to be 7.0 for protease activity. The importance of neutral proteases are their use in the food industry, because they possess specific function reducing the bitterness of food protein hydrolysates by hydrolysing hydrophobic amino acid bonds at neutral pH (Sandhya et al. 2005).
Effect of temperature and thermal stability on
the purified enzyme
In our study, it was found that the protease was
very active at 40-45°C (Fig. 8). Temperature is one of
the most important factors affecting the enzyme production (Uyar et al. 2011). Similar results were reported by Sousa et al. (2007) and VijayAnand et al.
(2009). Fig. 9 shows the effect of temperature on the purified protease stability. It was found that the enzyme was fully stable in the presence of 2 mM CaCl2 at 50ºC even after 120 min. Thermostability is
a critical feature required of proteases for industrial
applications such as detergent and leather
processing. Due to an increase in the enzyme
stability in the presence of CaCl2, the enzyme can be
used in biotechnological applications. Adinarayana et al. (2003), determined that protease from B. subtilis PE-11 was highly stable in the presence of
Ahmetoglu et al.
6
EurAsian Journal of BioSciences 9: 1-11 (2015)
Table 1. Some characters that distinguish Bacillus sp. KG5.
(+): positive, (-): negative, (w): weak, ND: Not determined
Table 2. Purification steps of protease.

CaCl2 up to 60ºC. For this reason, the stability of
Bacillus sp. KG5 protease may have an advantage in
its use in laundry detergent formulations.
Effects of metal ions, inhibitors and
detergents on the purified protease activity
As shown in Table 3, protease was significantly activated by Ca2+ (242% at 2 mM ), Mg2+ (189% at 5 mM) and Mn2+ (129% at 2 mM), while strongly inhibited by Cu2+, Hg2+ and Zn2+ (at 10 mM). Adinarayana et al. (2003), Hmidet et al. (2009) and Annamalai et al. (2014) stated that Ca2+ acts as an activator for proteases. An increase in the activity in the presence of Ca2+ may be due to stabilisation of enzymes in its active conformation rather than it being involved in the catalytic reaction. It probably acts as a salt or an ion bridge via a cluster of carboxylic groups (Divakar et al. 2010). EDTA (96% at 10 mM) and 1,10 phenantroline (95% at 10 mM), which are metal chelating agents, heavily inhibited the protease activity. These data indicate that divalent metal ion cofactors are necessary for the
activity of protease; in other words, the enzyme is belonging to the metallo-protease family. In previous studies, Matta and Punj (1998), Kim et al.
(2001), Secades et al. (2001), Doddapaneni et al.
Ahmetoglu et al.EurAsian Journal of BioSciences 9: 1-11 (2015)
7
Table 3. Effects of metal ions, inhibitors and detergents onthe purified protease activity.
*Relative activity was determined as percentage of control with noadditions.ND: Not defined, -: Not tested
Fig. 6. a): SDS-PAGE(Lane 1: protein molecular weight of the standarts [β-galactosidase (116 kDa), fructose phosphate (80.5 kDa), α-amylase
(58 kDa)], Lane 2: Sephadex G-75 gel permeation chromatography.
b): Zymogram with the nondenatured polyacrylamide gelelectrophoresis containing 0.1% gelatine(Sample: Sephadex G-75 gel permeation chromatography).
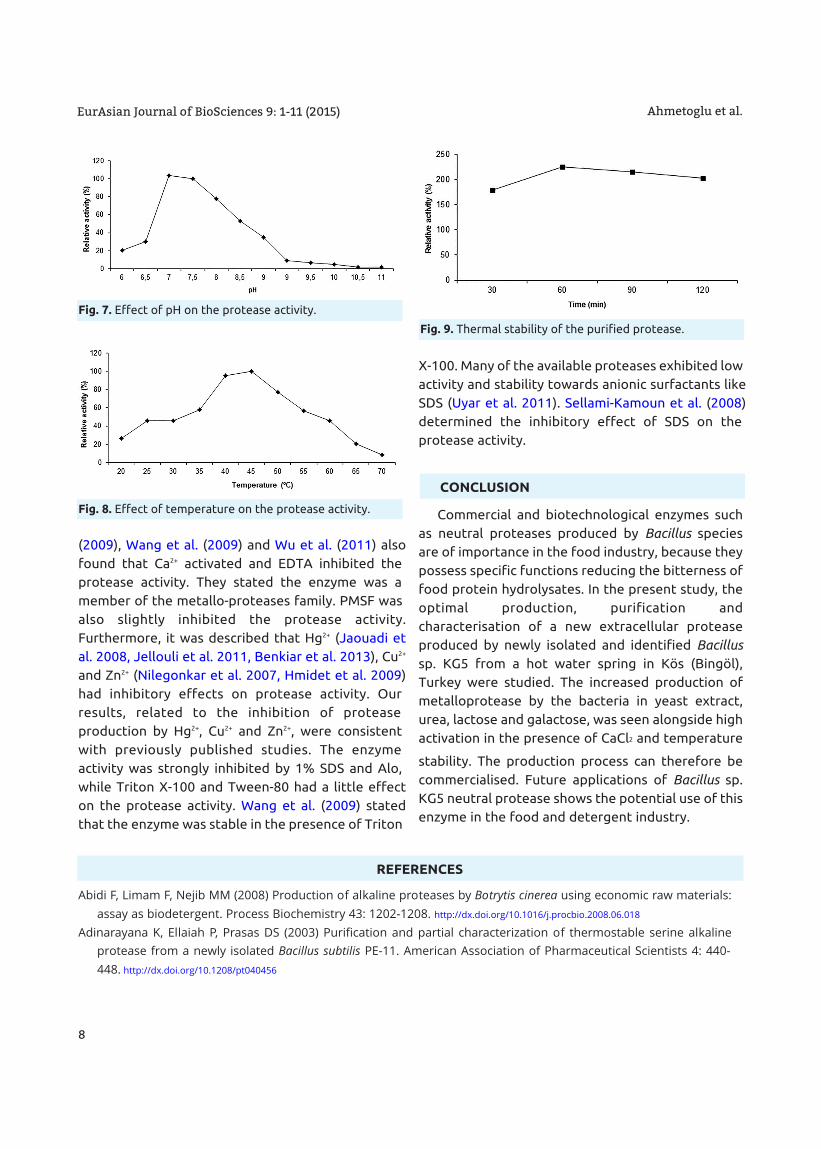
Ahmetoglu et al.EurAsian Journal of BioSciences 9: 1-11 (2015)
(2009), Wang et al. (2009) and Wu et al. (2011) also found that Ca2+ activated and EDTA inhibited the protease activity. They stated the enzyme was a member of the metallo-proteases family. PMSF was also slightly inhibited the protease activity. Furthermore, it was described that Hg2+ (Jaouadi et al. 2008, Jellouli et al. 2011, Benkiar et al. 2013), Cu2+
and Zn2+ (Nilegonkar et al. 2007, Hmidet et al. 2009) had inhibitory effects on protease activity. Our results, related to the inhibition of protease production by Hg2+, Cu2+ and Zn2+, were consistent with previously published studies. The enzyme activity was strongly inhibited by 1% SDS and Alo, while Triton X-100 and Tween-80 had a little effect on the protease activity. Wang et al. (2009) stated that the enzyme was stable in the presence of Triton
X-100. Many of the available proteases exhibited low activity and stability towards anionic surfactants like SDS (Uyar et al. 2011). Sellami-Kamoun et al. (2008) determined the inhibitory effect of SDS on the protease activity.
Commercial and biotechnological enzymes such
as neutral proteases produced by Bacillus species
are of importance in the food industry, because they
possess specific functions reducing the bitterness of
food protein hydrolysates. In the present study, the
optimal production, purification and
characterisation of a new extracellular protease
produced by newly isolated and identified Bacillus
sp. KG5 from a hot water spring in Kös (Bingöl),
Turkey were studied. The increased production of
metalloprotease by the bacteria in yeast extract,
urea, lactose and galactose, was seen alongside high
activation in the presence of CaCl2 and temperature
stability. The production process can therefore be
commercialised. Future applications of Bacillus sp.
KG5 neutral protease shows the potential use of this
enzyme in the food and detergent industry.
8
Fig. 7. Effect of pH on the protease activity.
Fig. 8. Effect of temperature on the protease activity.
Fig. 9. Thermal stability of the purified protease.
CONCLUSION
Abidi F, Limam F, Nejib MM (2008) Production of alkaline proteases by Botrytis cinerea using economic raw materials:assay as biodetergent. Process Biochemistry 43: 1202-1208. http://dx.doi.org/10.1016/j.procbio.2008.06.018
Adinarayana K, Ellaiah P, Prasas DS (2003) Purification and partial characterization of thermostable serine alkalineprotease from a newly isolated Bacillus subtilis PE-11. American Association of Pharmaceutical Scientists 4: 440- 448. http://dx.doi.org/10.1208/pt040456
REFERENCES

9
Ahmetoglu et al.EurAsian Journal of BioSciences 9: 1-11 (2015)
Ahmetoglu N, Matpan Bekler F, Gul-Guven R, Acer O, Guven K (2011) Characterisation of a partially purified proteasefrom Bacillus cereus KG5 isolated from a hot spring. Current Opinion of Biotechnology 22(S1): S88.http://dx.doi.org/10.1016/j.copbio.2011.05.267
Akcan N, Uyar F (2011) Production of extracellular alkaline protease from Bacillus subtilis RSKK96 with solid statefermentation. Eurasian Journal of Biosciences 5: 64-72. http://dx.doi.org/10.5053/ejobios.2011.5.0.8
Annamalai N, Rajeswari MV, Balasubramanian T (2014) Extraction, purification and application of thermostable andhalostable alkaline protease from Bacillus alveayuensis CAS 5 using marine wastes. Food and BioproductsProcessing 92: 335-342. http://dx.doi.org/10.1016/j.fbp.2013.08.009
Anwar A, Saleemuddin M (1997) Alkaline proteases: A review. Bioresource Technology 64: 175-183.http://dx.doi.org/10.1016/S0960-8524(97)00182-X
Asker MM, Mahmoud MG, El Shebwy K, Abd el Aziz MS (2013) Purification and characterization of two thermostableprotease fractions from Bacillus megaterium. Journal of Genetic Engineering and Biotechnology 11: 103-109.http://dx.doi.org/10.1016/j.jgeb.2013.08.001
Banik RM, Prakash M (2004) Laundry detergent compatibility of the alkaline protease from Bacillus cereus.Microbiological Research 159: 135-140. http://dx.doi.org/10.1016/j.micres.2004.01.002
Beg QK, Gupta R (2003) Purification and characterization of an oxidation-stable, thiol-dependent serine alkalineprotease from Bacillus mojavensis. Enzyme and Microbial Technology 32: 294-304. http://dx.doi.org/10.1016/S0141-
0229(02)00293-4
Benkiar A, Nadia ZJ, Badis A, Rebzani F, Soraya BT, Rekik H, Naili B, Ferradji FZ, Bejar S, Jaouadi B (2013) Biochemicaland molecular characterization of a thermo- and detergent-stable alkaline serine keratinolytic protease from Bacillus circulans strain DZ100 for detergent formulations and feather-biodegradation process. International Biodeterioration and Biodegradation 83: 129-138. http://dx.doi.org/10.1016/j.ibiod.2013.05.014
Christiansen T, Nielsen J (2002) Production of extracellular protease and glucose uptake in Bacillus clausii in steady-state and transient continuous cultures. Journal of Biotechnology 97: 265-273. http://dx.doi.org/10.1016/S0168-
1656(02)00109-8
Divakar K, Deepa Arul Priya J, Gautam P (2010) Purification and characterization of thermostable organic solvent-stableprotease from Aeromonas veronii PG01. Journal of Molecular Catalysis B: Enzymatic 66: 311-318. http://dx.doi.org/10.1016/j.molcatb.2010.06.008
Doddapaneni KK, Tatineni R, Vellanki RN, Rachcha S, Anabrolu N, Narakuti V, Mangaomoori LN (2009) Purification andcharacterization of a solvent and detergent- stable novel protease from Bacillus cereus. Microbiological Research164: 383-390. http://dx.doi.org/ 10.1016/j.micres.2007.04.005
Dussault HP (1955) An improve technique for staining for halophilic bacteria. Journal of Bacteriology 70: 484-485.http://dx.doi.org/10.1099/ijs.0.64482-0
Frikha B, Sellami-Kamoun A, Fakhfakh N, Haddar A, Manni L, Nasri M (2005) Production and purification of a calcium-dependent protease from Bacillus cereus BG1. Journal of Industrial Microbiology and Biotechnology 32: 186-194.http://dx.doi.org/10.1007/s10295-005-0228-z
Gordon RHWC, Pang CHN (1973) The Genus Bacillus. University of Washington, Washington, DC. Haddar A, Agrebi R, Bougatef A, Hmidet N, Sellami-Kamoun A, Nasri M (2009) Two detergent stable alkaline serine-
proteases from Bacillus mojavensis A21: Purification, characterization and potential application as a laundry detergent additive. Bioresource Technology 100: 3366-3373. http://dx.doi.org/10.1016/j.biortech.2009.01.061
Haki G, Rakshit S (2003) Developments in industrially important thermostable enzymes: a review. BioresourceTechnology 89: 17-34. http://dx.doi.org/10.1016/s0960-8524(03)00033-6
Hmidet N, El-Hadj Ali N, Haddar A, Kanoun S, Alya SK, Nasri M (2009) Alkaline proteases and thermostable α-amylaseco-produced by Bacillus licheniformis NH1: Characterization and potential application as detergent additive. Biochemical Engineering Journal 47: 71-79. http://dx.doi.org/10.1016/j.bej.2009.07.005

Ahmetoglu et al.
10
EurAsian Journal of BioSciences 9: 1-11 (2015)
Ibrahim ASS, Al-Salamah AA (2009) Optimization of media and cultivation conditions for alkaline pro tease productionby alkaliphilic Bacillus halodurans. Research Journal of Microbiology 4: 251-259. http://dx.doi.org/10.3923/jm.2009.251.259
Jaouadi B, Ellouz-Chaabouni S, Rhimi M, Bejar S (2008) Biochemical and molecular characterization of a detergent-stable serine alkaline protease from Bacillus pumilus CBS with high catalytic efficiency. Biochimie 90: 1291-1305http://dx.doi.org/10.1016/j.biochi.2008.03.004
Jellouli K, Ghorbel-Bellaaj O, Ayed HB, Manni L, Agrebi R, Nasri M (2011) Alkaline-protease from Bacillus licheniformisMP1: Purification, characterization and potential application as a detergent additive and for shrimp waste deproteinization. Process Biochemistry 46: 1248-1256. http://dx.doi.org/10.1016/j.procbio.2011.02.012
Johnvesly B, Naik GR (2001) Studies on production of thermostable alkaline protease from thermophilic and alkaliphilicBacillus sp. JB-99 in a chemically defined medium. Process Biochemistry 37: 139-144.http://dx.doi.org/10.1016/S0032-9592(01)00191-1
Joo HS, Chang CS (2006) Production of an oxidant and SDS-stable alkaline protease from an alkaophilic Bacillus clausiiI-52 by submerged fermentation: Feasibility as a laundry detergent additive. Enzyme and Microbial Technology 38: 176-183. http://dx.doi.org/10.1016/j.enzmictec.2005.05.008
Kim S, Jae Kim Y, Rhee IK (2001) Purification and characterization of a novel extracellular protease from Bacillus cereusKCTC 3674. Archives of Microbiology 17: 458-461. http://dx.doi.org/10.1007/s002030100282
Laemmli U (1970) Cleavage of structural proteins during the assembly of the head of bacteriphage T4. Nature 277:680-685. http://dx.doi.org/10.1038/227680a0
Leighton TJ, Doi RH, Warren RA J, Kelln RA (1973) The relationship of serine protease activity to RNA polymerasemodification and sporulation in Bacillus subtilis. Journal of Molecular Biology 76(1): 103-122.http://dx.doi.org/10.1016/0022-2836(73)90083-1
Lin S, Zhang M, Liu J, Jones GS (2015) Construction and application of recombinant strain for the production of analkaline protease from Bacillus licheniformis. Journal of Bioscience and Bioengineering 119: 284-288. http://dx.doi.org/10.1016/j.jbiosc.2014.08.002
Lowry OH, Rosebrough NJ, Farr AL (1951) Protein measurement with the folin phenol reagent. The Journal of BiologicalChemistry 193: 265-275. http://dx.doi.org/10.1074/jbc.270.46.27489
Mabrouk S, Hashem A, El-Shayeb N, Ismail A-M, Abdel-Fattah A (1999) Optimization of alkaline protease productivityby Bacillus licheniformis ATCC 21415. Bioresource Technology 69: 155-159. http://dx.doi.org/10.1016/S0960-8524(98)00165-5
Maruthiah T, Esakkiraj P, Prabakaran G, Palavesam A, Immanuel G (2013) Purification and characterization ofmoderately halophilic alkaline serine protease from marine Bacillus subtilis AP-MSU 6. Biocatalysis and AgriculturalBiotechnology 2: 116-119. http://dx.doi.org/10.1016/j.bcab.2013.03.001
Matsumoto M, De Bont JAM, Iskent S (2002) Isolation and characterization of the solvent-tolerant Bacillus cereus strainR1. Journal of Bioscience and Bioengineering 94: 45-51. http://dx.doi.org/10.1016/S1389-1723(02)80115-3
Matta H, Punj V (1998) Isolation and partial characterization of a thermostable extracellular protease of Bacilluspolymyxa B-17. International Journal of Food Microbiology 42: 139-145. http://dx.doi.org/10.1016/S0168-1605(98)00061-0
Murugavelh S, Mohanty K (2013) Isolation, identification and characterization of Cr(VI) reducing Bacillus cereus fromchromium contaminated soil. Chemical Engineering Journal 230: 1-9. http://dx.doi.org/10.1016/j.cej.2013.06.049
Nilegonkar SS, Zambare VP, Kanekar PP, Dhakephalkar PK, Sarnaik SS (2007) Production and partial characterization of dehairing protease from Bacillus cereus MCM B-326. Bioresource Technology 98: 1238-1245.http://dx.doi.org/10.1016/j.biortech.2006.05.003
Oh Y, Shih IL, Tzeng YM, Wang SL (2000) Protease produced by Pseudomonas aeruginosa K-187 and its application inthe deproteinization of shrimp and crab shell wastes. Enzyme and Microbial Technology 27: 3-10. http://dx.doi.org/10.1016/S0141-0229(99)00172-6
Oskouie SFG, Tabandeh F, Yakhchali B, Eftekhar F (2008) Response surface optimization of medium composition foralkaline protease production by Bacillus clausii. Biochemical Engineering Journal 39: 37-42. http://dx.doi.org/10.1016/j.bej.2007.08.016

Rahman R, Basri, M, Salleh AB (2003) Thermostable alkaline protease from Bacillus stearothermophilus F1; nutritionalfactors affecting protease production. Annals of Microbiology 53: 199-210. http://dx.doi.org/10.1016/j.biortech.2004.06.012
Rainey FA, Fritze D, Stackebrandt E (1994) The Phylogenetic diversty of thermophilic members of the genus Bacillus asrevealed by 16S rRNA analysis. FEMS Microbiology Letters 115: 205-212. http://dx.doi.org/10.1016/0378-1097(94)90015-9
Sandhya C, Sumantha A, Szakacs G, Pandey A (2005) Comparative evaluation of neutral protease production byAspergillus oryzae in submerged and solid-state fermentation. Process Biochemistry 40: 2689-2694.http://dx.doi.org/10.1016/j.procbio.2004.12.001
Secades P, Alvarez B, Guıjarro JA (2001) Purification and characterization of a psychrophilic, calcium-induced, growth-phase-dependent metalloprotease from The fish pathogen Flavobacterium psychrophilum. Applied and Environmental Microbiology 67: 2436-2444. http://dx.doi.org/10.1128/AEM.67.6.2436-2444.2001
Sellami-Kamoun A, Haddar A, Ali NE-H, Ghorbel-Frikha B, Kanoun S, Nasri M (2008) Stability of thermostable alkalineprotease from Bacillus licheniformis RP1 in commercial solid laundry detergent formulations. MicrobiologicalResearch 163: 299-306. http://dx.doi.org/10.1016/j.micres.2006.06.001
Shafee N, Aris SN, Rahman RNZRA, Basri M, Salleh AB (2005) Optimization of environmental and nutritional conditionsfor the production of alkaline protease by a newly isolated bacterium Bacillus cereus strain 146. Journal of AppliedSciences Research 1: 1-8.
Shivanand P, Jayaraman G (2009) Production of extracellular protease from halotolerant bacterium, Bacillus aquimarisstrain VITP4 isolated from Kumta Coast. Process Biochemistry 44: 1088-1094. http://dx.doi.org/10.1016/j.procbio.2009.05.010
Sookkheo B, Sinchaikul S, Phutrakul S, Chen ST (2000) purification and characterization of the highly thermostableproteases from Bacillus stearothermophilus TLS33. Protein Expression and Purification 20: 142-151.http://dx.doi.org/10.1006/prep.2000.1282
Sousa F, Jus S, Erbel A, Kokol V, Cavaco-Paulo A, Gubitz GM (2007) A Novel metalloprotease from Bacillus cereus forprotein fibre processing. Enzyme and Microbial Technology 40: 1772-1781. http://dx.doi.org/10.1016/j.enzmictec.2006.12.017
Subba Rao C, Madhavendra S, Sreenivas Rao R, Hobbs PJ, Prakasham R (2008) Studies on improving the immobilizedbead reusability and alkaline protease production by isolated immobilized Bacillus circulans (MTCC 6811) using
overall evaluation criteria. Applied Biochemistry and Biotechnology 150: 65-83.http://dx.doi.org/10.1007/s12010-008-8147-x
Joshi S, Satyanarayana T (2013) Characteristics and applications of a recombinant alkaline serine protease from anovel bacterium Bacillus lehensis. Bioresource Technology 131: 76-85. http://dx.doi.org/10.1016/j.biortech.2012.12.124
Towatana-Hutadilok N, Painupong A, Suntinanalert P (1999) Purification and characterization of an extracellularprotease from alkaliphilic and thermophilic Bacillus sp. PS7 19. Journal of Bioscience and Bioengıneering 87(5): 581-587. http://dx.doi.org/10.1016/S1389-1723(99)80118-2
Uyar F, Porsuk I, Kizil G, Yilmaz EI (2011) Optimal conditions for production of extracellular protease from newlyisolated Bacillus cereus strain CA15. Eurasian Journal of Biosciences 5: 1-9. http://dx.doi.org/10.5053/ejobios.2011.5.0.1
VijayAnand S, Hemapriya J, Selvinb J, Kiran S (2009) Production and optimization of haloalkaliphilic protease by anextremophile Halobacterium sp. Js1, isolated from Thalassohaline environment. African Journal of Basic & AppliedSciences 1(3-4): 49-54.
Wang S, Chao CH, Liang TW, Chen CC (2009) Purification and characterization of protease and chitinase from Bacilluscereus TKU006 and conversion of marine wastes by these enzymes. Journal of Marine Biotechnology 11: 334-344. http://dx.doi.org/10.1007/s10126-008-9149-y
Wu Y, Wang HX, Ng TB (2011) A novel metalloprotease from the wild Basidiomycete mushroom Lepista nuda. Journalof Microbiology and Biotechnology 21: 256-262. http://dx.doi.org/10.4014/jmb.1010.10060
Zambare V, Nilegaonkar S, Kanekar P (2011) A Novel extracellular protease from Pseudomonas aeruginosa MCM B-327:Enzyme production and its partial characterization. New Biotechnology 28: 173-181. http://dx.doi.org/10.1016/j.nbt.2010.10.002
11
Ahmetoglu et al.EurAsian Journal of BioSciences 9: 1-11 (2015)




















