
Production and detoxification of H 2 O 2 in lettuce plants exposed to selenium
10
RESEARCH ARTICLE Production and detoxification of H 2 O 2 in lettuce plants exposed to selenium J.J. Rı´os, B. Blasco, L.M. Cervilla, M.A. Rosales, E. Sanchez-Rodriguez, L. Romero & J.M. Ruiz Department of Plant Physiology, Faculty of Science, University of Granada, Granada, Spain Keywords Antioxidant enzymes; hydrogen peroxide; Lactuca sativa; lipid peroxidation; selenate; selenite. Correspondence J.J. Rı´os, Department of Plant Physiology, Faculty of Science, University of Granada. Avd. Fuente Nueva s/n, E-18071 Granada, Spain. Email: [email protected] Received: 5 March 2008; revised version accepted: 28 May 2008. doi:10.1111/j.1744-7348.2008.00276.x Abstract Selenium is considered an essential element for animals. Despite that it has not been demonstrated to be essential for higher plants, it has been attributed with a protective role against reactive oxygen species in plants subjected to stress. In this study, lettuce plants (Lactuca sativa cv. Philipus) received different applica- tion rates (5, 10, 20, 40, 60, 80 and 120 lM) of selenite or selenate, with the aim of testing the effect of Se on the production and detoxification of H 2 O 2 in non-stressed plants. The results indicate that the form selenate is less toxic than selenite; that is, the plants tolerated and responded positively to this element, and even increasing in growth up to a rate of 40 lM for the form selenate. On the contrary, the application of selenite triggered a higher foliar concentration of H 2 O 2 and a higher induction of lipid peroxidation [malondialdehyde content and lipoxygenase activity] in comparison to that observed after the selenate application. Also, the plants treated with selenate induced higher increases in enzymes that detoxify H 2 O 2 , especially ascorbate peroxidase and glutathione (GSH) peroxidase, as well as an increase in the foliar concentration of antioxidant compounds such as ascorbate and GSH. These data indicate that an application of selenate at low rates can be used to prevent the induction in plants of the antioxidant system, thereby improving stress resistance. Introduction Selenium has been considered essential to animal nutri- tion since 1957, and humans have a daily requirement of 50–70 lg day 21 (U.S. Department of Agriculture, 2003) as a component of the enzymes glutathione peroxidase (GSH-Px), selenoprotein and tetraiodothyronine 5#-de- iodinase. The studies have recommended a dietary Se supplement of 100–200 lg day 21 to decrease the inci- dence of lung and prostate cancer (Ip et al., 1991; Ip & Ganther, 1992; La ¨ uchli, 1993; Clark et al., 1996). Fur- thermore, although Se is not considered to be required by higher plants, there are indications that it exerts posi- tive effects on plants. Recent research has identified sev- eral beneficial effects of Se in plants which include antioxidant properties that can stimulate plant growth (Hartikainen et al., 2000), delay plant senescence (Djanaguiraman et al., 2005), protect plants from phloem-feeding aphids by means of deterrence and tox- icity to aphids (Hanson et al., 2004), protect against fun- gal infection and from herbivory (Hanson et al., 2003) and protect plants against different types of abiotic stress (Hartikainen & Xue, 1999). The beneficial effect of Se in plants subjected to stress conditions has in most cases been attributed to increased antioxidant activity. Thus, Xue et al. (2001) and after- wards Djanaguiraman et al. (2005) observed the effect of Se application in the form of selenate on senescence in lettuce and soy, confirming that the decline in antioxi- dant enzyme activity was milder in plants treated with this element, which offsets oxidative damage by boosting growth in plants treated with Se. Annals of Applied Biology ISSN 0003-4746 Ann Appl Biol 154 (2009) 107–116 ª 2008 The Authors Journal compilation ª 2008 Association of Applied Biologists 107
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Production and detoxification of H 2 O 2 in lettuce plants exposed to selenium

RESEARCH ARTICLE
Production and detoxification of H2O2 in lettuce plantsexposed to seleniumJ.J. Rıos, B. Blasco, L.M. Cervilla, M.A. Rosales, E. Sanchez-Rodriguez, L. Romero & J.M. Ruiz
Department of Plant Physiology, Faculty of Science, University of Granada, Granada, Spain
Keywords
Antioxidant enzymes; hydrogen peroxide;
Lactuca sativa; lipid peroxidation; selenate;
selenite.
Correspondence
J.J. Rıos, Department of Plant Physiology,
Faculty of Science, University of Granada. Avd.
Fuente Nueva s/n, E-18071 Granada, Spain.
Email: [email protected]
Received: 5 March 2008; revised version
accepted: 28 May 2008.
doi:10.1111/j.1744-7348.2008.00276.x
Abstract
Selenium is considered an essential element for animals. Despite that it has not
been demonstrated to be essential for higher plants, it has been attributed with
a protective role against reactive oxygen species in plants subjected to stress. In
this study, lettuce plants (Lactuca sativa cv. Philipus) received different applica-
tion rates (5, 10, 20, 40, 60, 80 and 120 lM) of selenite or selenate, with the
aim of testing the effect of Se on the production and detoxification of H2O2 in
non-stressed plants. The results indicate that the form selenate is less toxic
than selenite; that is, the plants tolerated and responded positively to this
element, and even increasing in growth up to a rate of 40 lM for the form
selenate. On the contrary, the application of selenite triggered a higher
foliar concentration of H2O2 and a higher induction of lipid peroxidation
[malondialdehyde content and lipoxygenase activity] in comparison to that
observed after the selenate application. Also, the plants treated with selenate
induced higher increases in enzymes that detoxify H2O2, especially ascorbate
peroxidase and glutathione (GSH) peroxidase, as well as an increase in the
foliar concentration of antioxidant compounds such as ascorbate and GSH.
These data indicate that an application of selenate at low rates can be used to
prevent the induction in plants of the antioxidant system, thereby improving
stress resistance.
Introduction
Selenium has been considered essential to animal nutri-
tion since 1957, and humans have a daily requirement of
50–70 lg day21 (U.S. Department of Agriculture, 2003)
as a component of the enzymes glutathione peroxidase
(GSH-Px), selenoprotein and tetraiodothyronine 5#-de-iodinase. The studies have recommended a dietary Se
supplement of 100–200 lg day21 to decrease the inci-
dence of lung and prostate cancer (Ip et al., 1991; Ip &
Ganther, 1992; Lauchli, 1993; Clark et al., 1996). Fur-
thermore, although Se is not considered to be required
by higher plants, there are indications that it exerts posi-
tive effects on plants. Recent research has identified sev-
eral beneficial effects of Se in plants which include
antioxidant properties that can stimulate plant growth
(Hartikainen et al., 2000), delay plant senescence
(Djanaguiraman et al., 2005), protect plants from
phloem-feeding aphids by means of deterrence and tox-
icity to aphids (Hanson et al., 2004), protect against fun-
gal infection and from herbivory (Hanson et al., 2003)
and protect plants against different types of abiotic stress
(Hartikainen & Xue, 1999).
The beneficial effect of Se in plants subjected to stress
conditions has in most cases been attributed to increased
antioxidant activity. Thus, Xue et al. (2001) and after-
wards Djanaguiraman et al. (2005) observed the effect of
Se application in the form of selenate on senescence in
lettuce and soy, confirming that the decline in antioxi-
dant enzyme activity was milder in plants treated with
this element, which offsets oxidative damage by boosting
growth in plants treated with Se.
Annals of Applied Biology ISSN 0003-4746
Ann Appl Biol 154 (2009) 107–116 ª 2008 The Authors
Journal compilation ª 2008 Association of Applied Biologists
107

Kong et al. (2005), studying the effect of applying sele-
nate to sorrel plants under saline stress, reported an
increase in antioxidant enzymes and furthermore an
improvement in the integrity of plasma, mitochondrial
and chloroplast membranes. These data, therefore, apart
from suggesting an antioxidant function of Se, define
this element as a strengthener of osmotic capacity for its
role in the maintenance of cell membranes.
Recently, a beneficial effect of Se on heavy metal toxic-
ity has been reported. Pedrero et al. (2008) studied the
response of broccoli submitted to Cd toxicity together
with the application of Se. This study reported that Se
application diminished the malondialdehyde (MDA)
content and decreased the translocation of Cd towards
the shoot, thereby reducing oxidative stress provoked by
this heavy metal.
The greater antioxidant activity after Se application has
been related by several authors to its possible action on the
enzyme GSH-Px. Although the existence of Se-dependent
GSH-Px has not been demonstrated in plants, as opposed
to the situation in animals, it appears that this enzyme
could be influenced by the presence of Se. Support of
this idea comes from several works such as that of
Shigeoka et al. (1991), who observed after purifying the
GSH-Px in the alga Chlamydomonas that it was very simi-
lar to the one characterised in humans. Afterwards,
Takeda et al. (1997) demonstrated in this same alga that
H2O2 was eliminated primarily by the enzyme ascorbate
peroxidase (APX) in the absence of Se; however, when
this trace element was applied, the H2O2 was detoxified
mainly by GSH-Px. Afterwards, work on higher plants,
such as those by Hartikainen et al. (1997, 2000), credit
Se with antioxidant properties at low concentrations for
the inducing power of GSH-Px.
However, apart from GSH-Px, other enzymes such as
superoxide dismutase (SOD) form an integral part in oxi-
dative metabolism and function to transform the radical
superoxide into H2O2 (Gratao et al., 2005). In this res-
pect, Kong et al. (2005), in sorrel plants under saline
stress found stronger SOD activity. On the contrary, in
the same work, the authors failed to find the same behav-
iour for the enzyme catalase (CAT), which was not af-
fected by the presence or absence of Se.
In short, the effect of Se in promoting antioxidant activ-
ity has been reported basically in plants subjected to any
type of stress, while the possible action of this trace ele-
ment (different application rates as well as forms selenate
versus selenite) in the oxidative metabolism of non-
stressed plants has hardly been documented. Notable
among the few works on this subject, the study by
Cartes et al. (2005) reported that lipid peroxidation de-
pended on the shoot Se concentration rather than the Se
source (selenite or selenate) in ryegrass plants. In this
study, the shoot Se concentration was significantly
greater in selenate than in selenite-treated plants, and
the pro-oxidant effect of Se was detected only in plants
treated with selenate. Recently, Gomes-Junior et al.
(2007) found that in coffee cell suspensions the applica-
tion of selenite at low concentrations (0.05 mM) pro-
duces lipid peroxidation and alterations in antioxidant
enzymes including a severe reduction in APX activity.
These types ofwork are of great importance at present in
the sense that Se is vital to mammalian nutrition because
Se deficiency may promote cancer (Diwadkar-Navsar-
iwala et al., 2006). Dietary Se deficiency in humans is
caused by the ingestion of plant foods with an impercep-
tible concentration of this element because of its low
bioavailability in most crop soils (Smorklji et al., 2005;
Pedrero et al., 2006). Therefore, as a result of this low
bioavailability and the role of plants as the main dietary
source of this element, more studies have appeared in
recent years concerning ways to increase the Se content
in plants used for human consumption through bio-
fortification programmes (Rıos et al., 2008). Thus, the
main aim of the present work was to determine, in non-
stressed plants, the way in which Se applied at different
rates and in different forms (selenate vs. selenite) affects
the formation and detoxification of H2O2.
Material and methods
Plant material and growing conditions
Seeds of Lactuca sativa L. cv. Philipus were germinated
and grown for 35 days in cell flats (cell size, 3 � 3 � 10
cm) filled with perlite mixture, and the flats were placed
on benches in an experimental greenhouse in southern
Spain (Saliplant S.L., Motril, Granada, Spain). The 35-
day-old seedlings were transferred to a cultivation cham-
ber under controlled environmental conditions with
relative humidity of 60–80%, temperature 25�C/15�C(day/night) and 12/12 h photoperiod at a photosynthetic
photon-flux density (PPFD) of 350 lmol m22 s21 (mea-
sured at the top of plants with a 190 SB quantum sensor;
LI-COR Inc., Lincoln, NE, USA). The plants were grown
in individual pots (25 cm upper diameter, 17 cm lower
diameter, 25 cm in height) of 8 L volume, filled with ver-
miculite. Until the end of experiment, the plants received
a growth solution composed of 4 mM Ca(NO3)2, 6 mM
KNO3, 2 mM MgSO4�7H2O, 1 mM NaH2PO4�2H2O, 50 lMH3BO3, 2 lM MnCl2�4H2O, 1 lM ZnSO4�7H2O, 0.1 lMNa2MoO4�2H2O, 0.25 lM CuSO4�5H2O and 10 lM Fe-ED-
DHA. The nutrient solution (pH 5.5–6.0) was renewed
every 3 days, and the vermiculite partly rinsed with
Millipore-filtered water to avoid nutrient accumulation.
Selenium and antioxidant system in lettuce plants J.J. Rıos et al.
108 Ann Appl Biol 154 (2009) 107–116 ª 2008 The Authors
Journal compilation ª 2008 Association of Applied Biologists

At 45 days after germination, the different Se dosages
were applied together with the nutrient solution de-
scribed above and maintained to the end of the experiment
(for 21 days). The concentrations of Se added as SeO4 =
(Na2SeO4) or SeO3 = (Na2SeO3) were 5, 10, 20, 40, 60, 80
and 120 lmol L21. The choice of the different Se ap-
plication rates was to cover a broad range, from the rec-
ommended rate for biofortification programmes to rates
causing phytotoxicity (Rıos et al., 2008). In addition to
these treatments, a control treatment consisted of ap-
plying the complete growth solution without a Se sup-
plement. The experimental design was a randomised
complete block with 15 treatments, arranged in individ-
ual pots with six plants per treatment and three repli-
cations. The experiment was repeated three times under
the same conditions (n = 9).
Plant sampling
Lettuce leaves were sampled on day 66 after sowing. Leaf
samples were standardised using only fully expanded
leaves from the middle part of plants in each replicate, as
these reflect most clearly, from the nutritional and meta-
bolic standpoint, the effect of the treatment applied. The
material was rinsed three times in distilled water after dis-
infection with non-ionic 1% detergent and then blotted
onfilter paper. At each sampling, freshmatterwas used for
analysis of enzyme activity: lipoxygenase (LOX), SOD,
CAT, APX and GSH-Px; MDA, H2O2, ascorbate and
glutathione (GSH) content. The rest of the plant material
(edible leaves) was lyophilised and used to determine
the biomass and Se concentration.
Plant analysis
Measurement of lipid peroxidation
For the assay of MDA, 0.5 g of lettuce leaf was homoge-
nised in 5 mL of 50 mM buffer solution (containing
0.07% of NaH2PO4�2H2O and 1.6% Na2HPO4�12H2O),
ground with a mortar and pestle on ice, and centrifuged
at 20 000 g for 25 min (4�C). The MDA concentration
was calculated using its extinction coefficient of
155 mM21 cm21 (Fu & Huang, 2001). LOX activity was
measured according to Minguez-Mosquera et al. (1993),
using 50 mM K-phosphate buffer (pH 6.0) for extrac-
tion. The LOX activity was calculated following the rise
in the extinction at A234 using an extinction coefficient
of 25 000 M21 cm21 (Egert & Tevini, 2002).
H2O2 concentration
The H2O2 content of leaf samples was colorimetrically
measured as described by Mukherjee & Choudhuri
(1983). Leaf samples were extracted with cold acetone
to determine the H2O2 levels. An aliquot (1 mL) of the
extract solution was mixed with 200 lL of 0.1% titan-
ium dioxide in 20% (v/v) H2SO4 and the mixture was
then centrifuged at 6000 g for 15 min. The intensity of
the yellow colour of the supernatant was measured at
415 nm. The H2O2 concentration was calculated from
a standard curve plotted within the range of 100–
1000 nmol H2O2.
Measurement of total ascorbate and glutathione content
The total ascorbate was determined spectrophotometri-
cally following Gossett et al. (1994). Lettuce leaves [1–
1.5 g fresh weight (FW)] were homogenised over ice
with 0.5 g inert sand and 10 mL of ice-cold 5% (w/v)
m-phosphoric acid freshly prepared with a mortar and
a pestle. The homogenate was centrifuged at 21 800 g
for 15 min at 2�C. Total ascorbate was determined by
incubating at 25�C for 30 min of a reaction mixture con-
sisting of 100 lL supernatant, 500 lL 110 mM KH2PO4,
100 lL 3.6 mM ethylenediaminetetraacetic acid (EDTA)
and 100 lL 1.5 mM dithiothreitol (DTT), this latter
compound applied to reduce all dehydroascorbate and
ascorbate to avoid ascorbate oxidation. After incuba-
tion, 100 lL of 0.5% (w/v) N-ethylmaleimide was
added to remove excess DTT. Colour was developed
by the addition of 400 lL 10% (w/v) trichloroacetic
acid, 400 lL 44% o-phosphoric acid, 400 lL of 65 mM
a-a-1-dipyridyl in 70% ethanol and 200 lL of 110 mM
FeCl3. The reaction mixture was then incubated at 37�Cfor 45 min in a water bath and quantified at 525 nm.
Total GSH was determined following Gossett et al.
(1994). The extraction was made by homogenisation in
metaphosphoric acid 5% (w/v). The homogenate was
centrifuged at 21 800 g for 15 min. Afterwards, the reac-
tion mixture was made of 50 lL extract, 250 lL buffer,
HEPES-HCl 50 mM (pH 7.6) that contained 330 mM
betaine and 150 lL sulphosalicylic acid at 10 % (v/v).
Afterwards, in a test tube, 150 lL of the above reaction
mixture, 700 lL of NADPH 0.3 mM, 100 lL of 5,5#-dithiobis-(2-nitrobenzoic acid) (DNTB) 6 mM and 50 lLof glutathione reductase (10 U mL21) were added.
Finally, the absorbances of the samples were read at
412 nm. A standard curve was drawn by preparing sol-
utions of 0.002–0.0001 g mL21 GSH in 60 mL meta-
phosphoric acid (pH 2.8) containing 1 mM EDTA and
determining in the same manner as for the extracts.
Activity of antioxidant enzymes
Superoxide dismutase (EC 1.15.1.1) activity was assayed
by monitoring the inhibition of the photochemical
J.J. Rıos et al. Selenium and antioxidant system in lettuce plants
Ann Appl Biol 154 (2009) 107–116 ª 2008 The Authors
Journal compilation ª 2008 Association of Applied Biologists
109

reduction of nitroblue tetrazolium (NBT), according to
the methods of Giannopolitis & Ries (1977) and Beyer &
Fridovich (1987), with some modifications. A 5-mL reac-
tion mixture was used, containing 50 mM Na2CO3 (pH
10.0), 13 mM methionine, 0.025% (v/v) Triton X-100,
63 lM NBT, 1.3 lM riboflavin and an appropriate aliquot
of enzyme extract. The reaction mixtures were illumi-
nated for 15 min at a PPFD of 380 lmol m22 s21. Mix-
tures not illuminated were used to correct for background
absorbance. One unit of SOD activity was defined as the
amount of enzyme required to cause 50% inhibition of
the reduction of NBT as monitored at 560 nm.
Catalase (EC 1.11.1.6) activity was determined by fol-
lowing the consumption of H2O2 at 240 nm for 5 min
(Nakano & Asada, 1981; Rao et al., 1997). The reaction
mixture (3 mL total volume) contained 25 mM Tris–
acetate buffer (pH 7.0), 0.8 mM EDTA-Na and 20 mM
H2O2, and the enzyme assay was performed at 25�C.The enzyme APX (EC 1.11.1.11) was assayed following
Rao et al. (1996). APX activity was determined by register-
ing the absorbance change at 290 nm for 3 min of a reac-
tion mixture (3.75 mL) containing 100 mM of potassium
phosphate buffer (pH 7.5), 0.5 mM of reduced ascorbate,
0.2 mM of H2O2 and 0.75 mL of enzyme extract.
The GSH-Px (EC 1.11.1.9) activity was measured by
a modification of the method of Flohe & Gunzler (1984)
using H2O2 as substrate. The enzyme was extracted by
prechilled KNaHPO4 buffer, pH 7.0, with homogeniser.
The supernatant obtained by centrifugation at 1100 g for
10 min was used as a coarse enzyme extract. For the
enzyme reaction, 0.2 mL of the supernatant was placed
in tube and mixed with 0.4 mL GSH (0.1 mM) and
0.2 mL KNaHPO4 (0.067 M). The above reagents with-
out supernatant extract were used for the non-enzyme
reaction. After preheating the mixture on water bath at
25�C for 5 min, 0.2 mL H2O2 (1.3 mM) was added to
initiate the reaction. The reaction lasted 10 min and was
stopped by adding 1 mL 1% trichloroacetic acid and the
mixture was put into an ice bath for 30 min. Then the
mixture was centrifuged for 10 min at 1100 g, 0.48 mL
the supernatant was placed into a cuvette and 2.2 mL of
0.32 M Na2HPO4 and 0.32 mL of 1.0 mM DNTB were
added for colour development. The absorbance at wave-
length 412 nm was measured after 5 min. The enzyme
activity was calculated as a decrease in GSH within the
reaction time when compared with that in the non-
enzyme reaction.
Measurement of total Se content
For the determination of the Se concentration, 25 mg of
sample were digested with 2.5 mL of concentrated HNO3
and 1 mL of H2O2 in an analytical microwave oven. The
resulting solution was diluted to 25 mL with deionised
water and the metal concentration determined by induc-
tively coupled plasma mass spectroscopy (ICP-MS) ac-
cording to Pedrero et al. (2006).
Statistical analysis
The data were subjected to a simple ANOVA at 95% con-
fidence, using the program Statgraphics 6.1. A two-way
ANOVA was applied to ascertain whether the Se appli-
cation rate, and forms applied, significantly affected the
results, and the means were compared by Fisher’s least
significant differences (LSD). The data shown are mean
values � standard errors (SE). The significance levels for
both analyses were expressed as: *P < 0.05; **P < 0.01;
***P < 0.001 and ns, not significant.
Results
Selenium, despite not being an essential element for
plants, is absorbed and accumulated by them depending
on the application rate and the form available in the
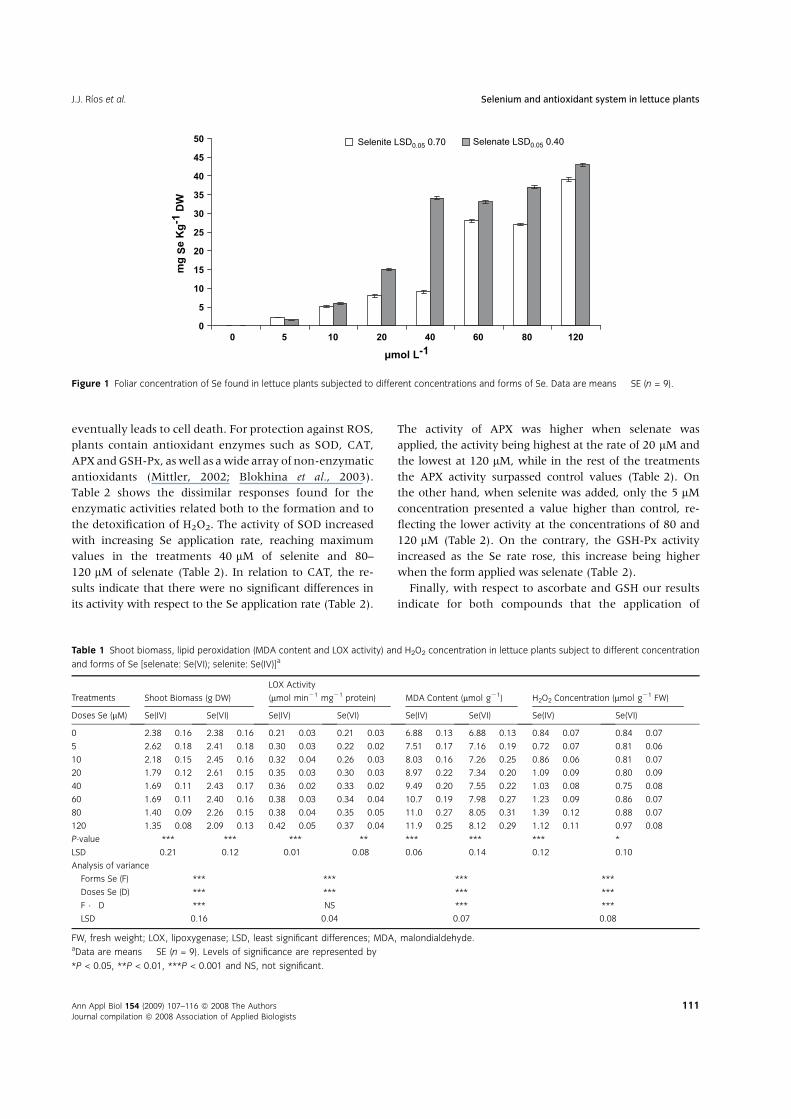
culture medium. As expected, foliar Se concentration
increased with increasing application rate (P < 0.001),
peaking in the 120 lM treatment for both Se forms
applied (Fig. 1). It is striking that for most of the applica-
tion rates, the foliar concentration of Se was higher with
selenate than with selenite (Fig. 1).
In relation to the shoot biomass, the highest valueswere
registered at the rate of 20 lMfor selenate and 5 lMin the
case of selenite. These concentrations resulted in greater
plant growth than in control (Table 1). Finally, plant
growth diminished to a concentration of 120 lM where
lower shoot biomass production was found for both Se
forms applied (Table 1).
With respect to stress indicators used in this experiment,
we found that the external application of the different
rates of Se augmented LOX activity, foliar concentration
of MDA, with the treatment of 120 lM showing the high-
est values, and the control plants the lowest (Table 1). As
for these parameters, at all rates selenite application
induced higher values than did selenate (Table 1). In
addition, Table 1 shows an increase in the H2O2 concen-
tration as the Se application rate increases, its concentra-
tion being again higher when the form used was
selenite. The dissimilar effect of the two Se forms on the
H2O2 concentration is striking because, when selenite
was applied at a concentration of 20 lM, the H2O2 con-
centration rose significantly with respect to control,
while for selenate this did not occur until at the concen-
tration of 120 lM (Table 1).
During oxidative stress, the excess reactive oxygen spe-
cies (ROS) production causes membrane damage that
Selenium and antioxidant system in lettuce plants J.J. Rıos et al.
110 Ann Appl Biol 154 (2009) 107–116 ª 2008 The Authors
Journal compilation ª 2008 Association of Applied Biologists
eventually leads to cell death. For protection against ROS,
plants contain antioxidant enzymes such as SOD, CAT,
APX andGSH-Px, aswell as awide array of non-enzymatic
antioxidants (Mittler, 2002; Blokhina et al., 2003).
Table 2 shows the dissimilar responses found for the
enzymatic activities related both to the formation and to
the detoxification of H2O2. The activity of SOD increased
with increasing Se application rate, reaching maximum
values in the treatments 40 lM of selenite and 80–
120 lM of selenate (Table 2). In relation to CAT, the re-
sults indicate that there were no significant differences in
its activity with respect to the Se application rate (Table 2).
The activity of APX was higher when selenate was
applied, the activity being highest at the rate of 20 lM and
the lowest at 120 lM, while in the rest of the treatments
the APX activity surpassed control values (Table 2). On
the other hand, when selenite was added, only the 5 lMconcentration presented a value higher than control, re-
flecting the lower activity at the concentrations of 80 and
120 lM (Table 2). On the contrary, the GSH-Px activity
increased as the Se rate rose, this increase being higher
when the form applied was selenate (Table 2).
Finally, with respect to ascorbate and GSH our results
indicate for both compounds that the application of
0
5
10
15
20
25
30
35
40
45
50
0 5 10 20 40 60 80 120
µmol L-1
mg
Se
Kg
-1 D
W
Selenate LSD0.05 0.40Selenite LSD0.05 0.70
Figure 1 Foliar concentration of Se found in lettuce plants subjected to different concentrations and forms of Se. Data are means � SE (n = 9).
Table 1 Shoot biomass, lipid peroxidation (MDA content and LOX activity) and H2O2 concentration in lettuce plants subject to different concentration
and forms of Se [selenate: Se(VI); selenite: Se(IV)]a
Treatments Shoot Biomass (g DW)
LOX Activity
(lmol min21 mg21 protein) MDA Content (lmol g21) H2O2 Concentration (lmol g21 FW)
Doses Se (lM) Se(IV) Se(VI) Se(IV) Se(VI) Se(IV) Se(VI) Se(IV) Se(VI)
0 2.38 � 0.16 2.38 � 0.16 0.21 � 0.03 0.21 � 0.03 6.88 � 0.13 6.88 � 0.13 0.84 � 0.07 0.84 � 0.07
5 2.62 � 0.18 2.41 � 0.18 0.30 � 0.03 0.22 � 0.02 7.51 � 0.17 7.16 � 0.19 0.72 � 0.07 0.81 � 0.06
10 2.18 � 0.15 2.45 � 0.16 0.32 � 0.04 0.26 � 0.03 8.03 � 0.16 7.26 � 0.25 0.86 � 0.06 0.81 � 0.07
20 1.79 � 0.12 2.61 � 0.15 0.35 � 0.03 0.30 � 0.03 8.97 � 0.22 7.34 � 0.20 1.09 � 0.09 0.80 � 0.09
40 1.69 � 0.11 2.43 � 0.17 0.36 � 0.02 0.33 � 0.02 9.49 � 0.20 7.55 � 0.22 1.03 � 0.08 0.75 � 0.08
60 1.69 � 0.11 2.40 � 0.16 0.38 � 0.03 0.34 � 0.04 10.7 � 0.19 7.98 � 0.27 1.23 � 0.09 0.86 � 0.07
80 1.40 � 0.09 2.26 � 0.15 0.38 � 0.04 0.35 � 0.05 11.0 � 0.27 8.05 � 0.31 1.39 � 0.12 0.88 � 0.07
120 1.35 � 0.08 2.09 � 0.13 0.42 � 0.05 0.37 � 0.04 11.9 � 0.25 8.12 � 0.29 1.12 � 0.11 0.97 � 0.08
P-value *** *** *** ** *** *** *** *
LSD 0.21 0.12 0.01 0.08 0.06 0.14 0.12 0.10
Analysis of variance
Forms Se (F) *** *** *** ***
Doses Se (D) *** *** *** ***
F � D *** NS *** ***
LSD 0.16 0.04 0.07 0.08
FW, fresh weight; LOX, lipoxygenase; LSD, least significant differences; MDA, malondialdehyde.aData are means � SE (n = 9). Levels of significance are represented by
*P < 0.05, **P < 0.01, ***P < 0.001 and NS, not significant.
J.J. Rıos et al. Selenium and antioxidant system in lettuce plants
Ann Appl Biol 154 (2009) 107–116 ª 2008 The Authors
Journal compilation ª 2008 Association of Applied Biologists
111
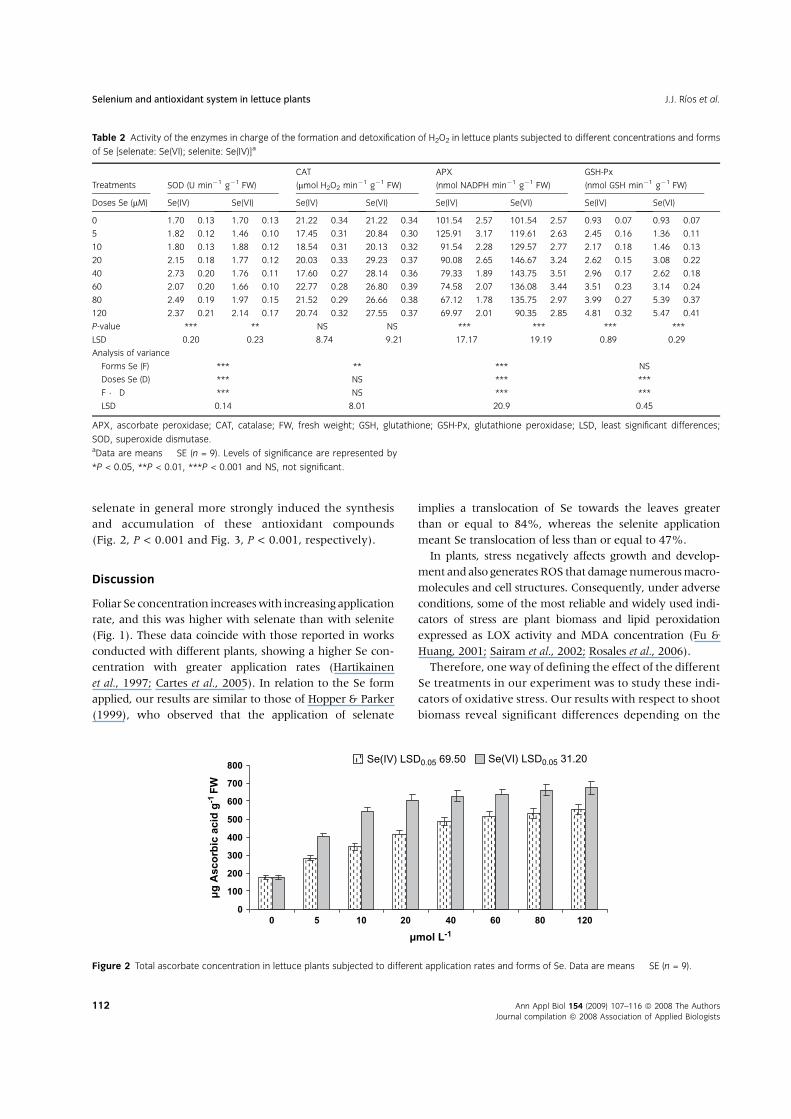
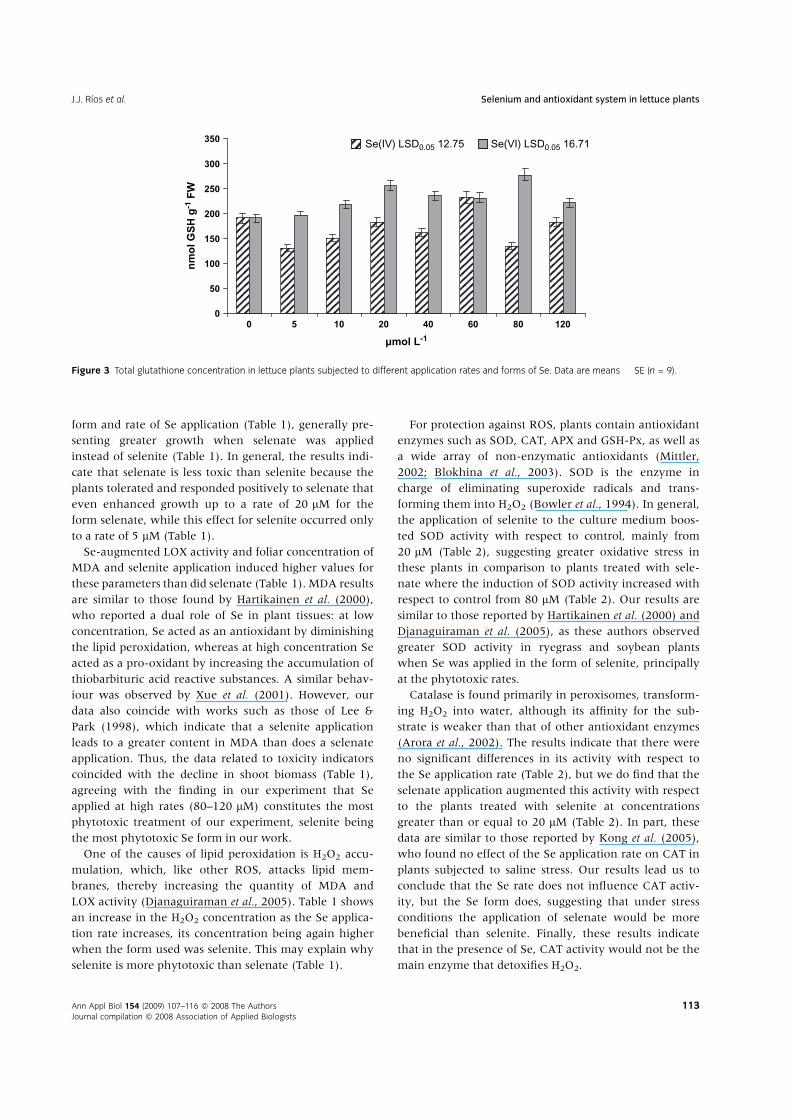
selenate in general more strongly induced the synthesis
and accumulation of these antioxidant compounds
(Fig. 2, P < 0.001 and Fig. 3, P < 0.001, respectively).
Discussion
Foliar Se concentration increaseswith increasing application
rate, and this was higher with selenate than with selenite
(Fig. 1). These data coincide with those reported in works
conducted with different plants, showing a higher Se con-
centration with greater application rates (Hartikainen
et al., 1997; Cartes et al., 2005). In relation to the Se form
applied, our results are similar to those of Hopper & Parker
(1999), who observed that the application of selenate
implies a translocation of Se towards the leaves greater
than or equal to 84%, whereas the selenite application
meant Se translocation of less than or equal to 47%.
In plants, stress negatively affects growth and develop-
ment and also generates ROS that damagenumerousmacro-
molecules and cell structures. Consequently, under adverse
conditions, some of the most reliable and widely used indi-
cators of stress are plant biomass and lipid peroxidation
expressed as LOX activity and MDA concentration (Fu &
Huang, 2001; Sairam et al., 2002; Rosales et al., 2006).
Therefore, oneway of defining the effect of the different
Se treatments in our experiment was to study these indi-
cators of oxidative stress. Our results with respect to shoot
biomass reveal significant differences depending on the
Table 2 Activity of the enzymes in charge of the formation and detoxification of H2O2 in lettuce plants subjected to different concentrations and forms
of Se [selenate: Se(VI); selenite: Se(IV)]a
Treatments SOD (U min21 g21 FW)
CAT
(lmol H2O2 min21 g21 FW)
APX
(nmol NADPH min21 g21 FW)
GSH-Px
(nmol GSH min21 g21 FW)
Doses Se (lM) Se(IV) Se(VI) Se(IV) Se(VI) Se(IV) Se(VI) Se(IV) Se(VI)
0 1.70 � 0.13 1.70 � 0.13 21.22 � 0.34 21.22 � 0.34 101.54 � 2.57 101.54 � 2.57 0.93 � 0.07 0.93 � 0.07
5 1.82 � 0.12 1.46 � 0.10 17.45 � 0.31 20.84 � 0.30 125.91 � 3.17 119.61 � 2.63 2.45 � 0.16 1.36 � 0.11
10 1.80 � 0.13 1.88 � 0.12 18.54 � 0.31 20.13 � 0.32 91.54 � 2.28 129.57 � 2.77 2.17 � 0.18 1.46 � 0.13
20 2.15 � 0.18 1.77 � 0.12 20.03 � 0.33 29.23 � 0.37 90.08 � 2.65 146.67 � 3.24 2.62 � 0.15 3.08 � 0.22
40 2.73 � 0.20 1.76 � 0.11 17.60 � 0.27 28.14 � 0.36 79.33 � 1.89 143.75 � 3.51 2.96 � 0.17 2.62 � 0.18
60 2.07 � 0.20 1.66 � 0.10 22.77 � 0.28 26.80 � 0.39 74.58 � 2.07 136.08 � 3.44 3.51 � 0.23 3.14 � 0.24
80 2.49 � 0.19 1.97 � 0.15 21.52 � 0.29 26.66 � 0.38 67.12 � 1.78 135.75 � 2.97 3.99 � 0.27 5.39 � 0.37
120 2.37 � 0.21 2.14 � 0.17 20.74 � 0.32 27.55 � 0.37 69.97 � 2.01 90.35 � 2.85 4.81 � 0.32 5.47 � 0.41
P-value *** ** NS NS *** *** *** ***
LSD 0.20 0.23 8.74 9.21 17.17 19.19 0.89 0.29
Analysis of variance
Forms Se (F) *** ** *** NS
Doses Se (D) *** NS *** ***
F � D *** NS *** ***
LSD 0.14 8.01 20.9 0.45
APX, ascorbate peroxidase; CAT, catalase; FW, fresh weight; GSH, glutathione; GSH-Px, glutathione peroxidase; LSD, least significant differences;
SOD, superoxide dismutase.aData are means � SE (n = 9). Levels of significance are represented by
*P < 0.05, **P < 0.01, ***P < 0.001 and NS, not significant.
0
100
200
300
400
500
600
700
800
0 5 10 20 40 60 80 120
µmol L-1
µg
Asc
orb
ic a
cid
g-1
FW
Se(VI) LSD0.05 31.20 Se(IV) LSD0.05 69.50
Figure 2 Total ascorbate concentration in lettuce plants subjected to different application rates and forms of Se. Data are means � SE (n = 9).
Selenium and antioxidant system in lettuce plants J.J. Rıos et al.
112 Ann Appl Biol 154 (2009) 107–116 ª 2008 The Authors
Journal compilation ª 2008 Association of Applied Biologists
form and rate of Se application (Table 1), generally pre-
senting greater growth when selenate was applied
instead of selenite (Table 1). In general, the results indi-
cate that selenate is less toxic than selenite because the
plants tolerated and responded positively to selenate that
even enhanced growth up to a rate of 20 lM for the
form selenate, while this effect for selenite occurred only
to a rate of 5 lM (Table 1).
Se-augmented LOX activity and foliar concentration of
MDA and selenite application induced higher values for
these parameters than did selenate (Table 1). MDA results
are similar to those found by Hartikainen et al. (2000),
who reported a dual role of Se in plant tissues: at low
concentration, Se acted as an antioxidant by diminishing
the lipid peroxidation, whereas at high concentration Se
acted as a pro-oxidant by increasing the accumulation of
thiobarbituric acid reactive substances. A similar behav-
iour was observed by Xue et al. (2001). However, our
data also coincide with works such as those of Lee &
Park (1998), which indicate that a selenite application
leads to a greater content in MDA than does a selenate
application. Thus, the data related to toxicity indicators
coincided with the decline in shoot biomass (Table 1),
agreeing with the finding in our experiment that Se
applied at high rates (80–120 lM) constitutes the most
phytotoxic treatment of our experiment, selenite being
the most phytotoxic Se form in our work.
One of the causes of lipid peroxidation is H2O2 accu-
mulation, which, like other ROS, attacks lipid mem-
branes, thereby increasing the quantity of MDA and
LOX activity (Djanaguiraman et al., 2005). Table 1 shows
an increase in the H2O2 concentration as the Se applica-
tion rate increases, its concentration being again higher
when the form used was selenite. This may explain why
selenite is more phytotoxic than selenate (Table 1).
For protection against ROS, plants contain antioxidant
enzymes such as SOD, CAT, APX and GSH-Px, as well as
a wide array of non-enzymatic antioxidants (Mittler,
2002; Blokhina et al., 2003). SOD is the enzyme in
charge of eliminating superoxide radicals and trans-
forming them into H2O2 (Bowler et al., 1994). In general,
the application of selenite to the culture medium boos-
ted SOD activity with respect to control, mainly from
20 lM (Table 2), suggesting greater oxidative stress in
these plants in comparison to plants treated with sele-
nate where the induction of SOD activity increased with
respect to control from 80 lM (Table 2). Our results are
similar to those reported by Hartikainen et al. (2000) and
Djanaguiraman et al. (2005), as these authors observed
greater SOD activity in ryegrass and soybean plants
when Se was applied in the form of selenite, principally
at the phytotoxic rates.
Catalase is found primarily in peroxisomes, transform-
ing H2O2 into water, although its affinity for the sub-
strate is weaker than that of other antioxidant enzymes
(Arora et al., 2002). The results indicate that there were
no significant differences in its activity with respect to
the Se application rate (Table 2), but we do find that the
selenate application augmented this activity with respect
to the plants treated with selenite at concentrations
greater than or equal to 20 lM (Table 2). In part, these
data are similar to those reported by Kong et al. (2005),
who found no effect of the Se application rate on CAT in
plants subjected to saline stress. Our results lead us to
conclude that the Se rate does not influence CAT activ-
ity, but the Se form does, suggesting that under stress
conditions the application of selenate would be more
beneficial than selenite. Finally, these results indicate
that in the presence of Se, CAT activity would not be the
main enzyme that detoxifies H2O2.
0
50
100
150
200
250
300
350
0 5 10 20 40 60 80 120
µmol L-1
nm
ol G
SH
g-1
FW
Se(IV) LSD0.05 12.75 Se(VI) LSD0.05 16.71
Figure 3 Total glutathione concentration in lettuce plants subjected to different application rates and forms of Se. Data are means � SE (n = 9).
J.J. Rıos et al. Selenium and antioxidant system in lettuce plants
Ann Appl Biol 154 (2009) 107–116 ª 2008 The Authors
Journal compilation ª 2008 Association of Applied Biologists
113

The enzymes APX and GSH-Px are in charge of detoxi-
fication of H2O2 produced by SOD in the chloroplast
through transformation of superoxide ion during photo-
synthesis (Arora et al., 2002). In general, the activity of
APX was higher with selenate than with selenite (Table 2).
In short, the lower APX activity and the absence of sig-
nificant variations in the CAT activity with respect to
control in plants treated with selenite (Table 2) together
with the increased SOD activity in these plants (Table 2)
could explain the higher concentration of H2O2 and
enhanced lipid peroxidation and, therefore, of the
decline in shoot biomass (Table 1) found after applying
rates higher than 20 lM of selenite. By contrast, sele-
nate application up to 60 lM caused variations in the
SOD activity and also increased the APX and GSH-Px
activities (Table 2), prevented greater foliar concen-
trations of H2O2 and lipid peroxidation (Table 1) and
caused a significant reduction in shoot biomass with
respect to control (Table 1).
The results for GSH-Px coincide with several studies
made with Se in which an increase in this trace element
augmented its activity (Hartikainen et al, 2000; Xue et al.,
2001; Djanaguiraman et al. 2005; Cartes et al., 2005).
However, our results indicate that the GSH-Px activity did
not have an essential role in H2O2 detoxification, at least
in the plants treated with selenite; such an increase in this
enzyme activity (Table 2) did not diminish the foliar con-
centration of H2O2 (Table 1). Finally, the different behav-
iour found in our study in terms of the GSH-Px activity
could be defined as a Se-dependent protein for which
activity depends exclusively on the presence of Se.
Both theAPX and theGSH-Px activities required reduced
compounds for their functioning. These compounds are
ascorbate and GSH, respectively (Arora et al. 2002). Pre-
viously, Rıos et al. (2008) observed that Se application
induced the synthesis of antioxidant compounds such as
total phenols, flavonoids, phenylpropanoids and glyco-
sides in lettuce plants, this induction being higher with
the application of selenate. In our experiment, the greater
induction of the synthesis and accumulation of different
antioxidants, such as ascorbate and GSH by selenate
(Figs 2 and 3), could also be explained by the fact that the
plants treated with this form of Se had lower oxidative
stress than those treated with selenite.
The results of GSH are important in this work, as they
indicate the degree of S assimilation and therefore the pos-
sibility of Se toxicity. According to Anderson (1993), Se
metabolism follows the same pathway as S, due to the
chemical similarity of the two molecules. Most enzymes
involved in S metabolism can catalyse the analogue reac-
tion with the corresponding Se substrates, forming Se ana-
logues of S compounds that are substrates for S enzymes
or replacing S in amino acids and thereby disrupting the
protein catalytic activity causing Se toxicity in plants.
Therefore, the greater formation of GSH in plants treated
with selenate (Fig. 3), together with a greater concentra-
tion in these plants of total S and total SH (data not
shown), could also explain why the application of this Se
form proved less phytotoxic than the application of sele-
nite. Possibly the highest incorporation of Se to amino
acids, especially Se-cysteine in plants treated with selenite
could provoke an alteration in the transport of e2 in pho-
tosynthesis, in consideration of the role of cysteine in this
process (Leustek et al., 2000), which would boost ROS
production in these plants. On the contrary, the lesser for-
mation of Se-cysteine in plants treated with selenate
together with a greater induction of the antioxidant sys-
tem could explain the lower ROS production and there-
fore less reduction of biomass production with respect to
plants treated with selenite.
In conclusion, we can indicate that the effect of Se on
the formation and detoxification of H2O2 in our work
depends largely on the form in which this trace element
is applied to the culture medium. Thus, the application
rate of selenite greater than 10 lM provoked a decline
in shoot biomass, associated with an increase in SOD
activity and a decrease in the enzymes in charge of H2O2
detoxification, mainly APX, which provoked a rise in
lipid peroxidation. On the contrary, in our work, the ap-
plication of selenate to 80 lM did not mean an increase
in the generation and accumulation of H2O2. Further-
more, the antioxidant system (CAT, APX, GSH-Px activi-
ties and total ascorbate and GSH concentrations) was
significantly improved with respect to control, which
could explain the beneficial effect of selenate found in
plants subjected to diverse abiotic stress. Finally, our re-
sults are novel and could define the application of sele-
nate at low application rates as a treatment to prevent
the induction of the antioxidant system in plants, help-
ing to improve the resistance responses to stress. How-
ever, it would be necessary to confirm these results in
future with studies that focus on the effect of Se during
the application time and the response of the different
isoenzymes that make up the antioxidant system in
plants.
Acknowledgements
This work was supported by PAI (Plan Andaluz de Inves-
tigacion, AGR-161).
References
Anderson J.W. (1993) Selenium interactions in sulfur
metabolism. In Sulfur Nutrition and Assimilation in Higher
Selenium and antioxidant system in lettuce plants J.J. Rıos et al.
114 Ann Appl Biol 154 (2009) 107–116 ª 2008 The Authors
Journal compilation ª 2008 Association of Applied Biologists

Plants: Regulatory, Agricultural and Environmental Aspects,
pp. 49–60. Ed. L.J. De Kok. The Hague, The Netherlands:
SPB Academic Publishing.
Arora A., Sairam R.K., Srivastava G.C. (2002) Oxidative
stress and antioxidative system in plants. Current Science,
82, 1227–1238.
Beyer W.F., Fridovich I. (1987) Assaying for superoxide dis-
mutase activity: some large consequences of minor
changes in conditions. Analytical Biochemistry, 161,
559–566.
Blokhina O., Virolainen E., Fagerstedt K.V. (2003) Anti-
oxidants, oxidative damage and oxygen deprivation stress:
a review. Annals of Botany, 91, 179–194.
Bowler C., Van Camp W., Van Montagu M., Inze D. (1994)
Superoxide dismutase in plants. Critical Reviews in Plant
Sciences, 13, 199–218.
Cartes P., Gianfreda L., Mora M.L. (2005) Uptake of sele-
nium and its antioxidant activity in ryegrass when applied
as selenate and selenite forms. Plant and Soil, 276,
359–367.
Clark L.C., Combs G.F. Jr, Turnbull B.W., Slate E.H.,
Chalker D.K., Chow J., Davis L.S., Glover R.A., Graham
G.F., Gross E.G., Krongrad A., Lesher J.L. Jr, Park H.K.,
Sanders B.B. Jr, Smith C.L., Taylor J.R. (1996) Effects of
selenium supplementation for cancer prevention in patients
with carcinoma of the skin. A randomised controlled trial.
JAMA, 276, 1957–1963.
Diwadkar-Navsariwala V., Prins G.S., Swanson S.M., Birch
L.A., Ray V.H., Hedayat S., Lantvit D.L., Diamond A.M.
(2006) Selenoprotein deficiency accelerates prostate carci-
nogenesis in a transgenic model. Proceedings of the National
Academy of Sciences of the United States of America, 103,
8179–8184.
Djanaguiraman M., Devi D.D., Shanker A.K., Sheeba J.A.,
Bangarusamy U. (2005) Selenium – an antioxidative
protectant in soybean during senescence. Plant and Soil,
272, 77–86.
Egert M., Tevini M. (2002) Influence of drought on some
physiological parameters symptomatic for oxidative stress
in leaves of chives (Allium schoenoprasum). Environmental
and Experimental Botany, 48, 43–49.
Flohe L., Gunzler W.A. (1984) Assays of glutathione
peroxidase. In Methods in Enzymology. Vol. 105, pp.
114–121. Ed. L. Packer. New York: Academic Press.
Fu J.M., Huang B.R. (2001) Involvement of antioxidants and
lipid peroxidation in the adaptation of two cool-season
grasses to localized drought stress. Environmental and
Experimental Botany, 45, 105–114.
Giannopolitis C.N., Ries S.K. (1977) Superoxide dismutases.
1. Occurrence in higher plants. Plant Physiology, 59,
309–314.
Gomes-Junior R.A., Gratao P.L., Gaziola S.A., Mazzafera P.,
Lea P.J., Azevedo R.A. (2007) Selenium-induced oxidative
stress in coffee cell suspension cultures. Functional Plant
Biology, 34, 449–456.
Gossett D.R., Millhollon E.P., Lucas M.C. (1994) Antioxidant
response to NaCl stress in salt-tolerant and
salt-sensitive cultivars of cotton. Crop Science, 34,
706–714.
Gratao P.L., Polle A., Lea P.J., Azevedo R.A. (2005) Making
the life of heavy metal-stressed plants a little easier. Func-
tional Plant Biology, 32, 481–494.
Hanson B., Garifullina G.F., Lindblom S.D., Wangeline A.,
Ackley A., Kramer K., Norton A.P., Lawrence C.B., Pilon-
Smits E.A.H. (2003) Selenium accumulation protects
Brassica juncea from invertebrate herbivory and fungal
infection. New Phytologist, 159, 461–469.
Hanson B., Lindblom S.D., Loeffler M.L., Pilon-Smits E.A.H.
(2004) Selenium protects plants from phloem-feeding
aphids due to both deterrence and toxicity. New Phytologist,
162, 655–662.
Hartikainen H., Xue T. (1999) The promotive effect of sele-
nium on plant growth as triggered by ultraviolet radiation.
Journal of Environmental Quality, 28, 1372–1375.
Hartikainen H., Ekholm E., Piironen V., Xue T., Koivu T.,
Yli-Halla M. (1997) Quality of the ryegrass and lettuce
yields as affected by selenium fertilization. Agricultural and
Food Science in Finland, 6, 381–387.
Hartikainen H., Xue T., Piironen V. (2000) Selenium as an
anti-oxidant and pro-oxidant in ryegrass. Plant and Soil,
225, 193–200.
Hopper J.L., Parker D.R. (1999) Plant availability of selenite
and selenate as influenced by the competing ions phos-
phate and sulfate. Plant and Soil, 210, 199–207.
Ip C., Ganther H.E. (1992) Comparison of Se and S analogs
in cancer prevention. Carcinogenesis, 13, 1167–1170.
Ip C., Hayes C., Budnick R.M., Ganther H.E. (1991) Chem-
ical form of Se, critical metabolites, and cancer prevention.
Cancer Research, 51, 595–600.
Kong L.A., Wang M., Bi D. (2005) Selenium modulates the
activities of antioxidant enzymes, osmotic homeostasis and
promotes the growth of sorrel seedlings under salt stress.
Plant Growth Regulation, 45, 155–163.
Lauchli A. (1993) Selenium in plants: uptake, function, and
environmental toxicity. Botanica Acta, 106, 455–468.
Lee G.P., Park K.W. (1998) Effect of Selenium concentration
in the Nutrient solution on the Growth and internal
quality of Endive. Journal Korean of Society and Horticultural
Science, 39, 391–396.
Leustek T., Martin M.N., Bick J.-A., Davies J.P. (2000)
Pathways and regulation of sulfur metabolism revealed
through molecular and genetic studies. Annual Review of
Plant Physiology and Plant Molecular Biology, 51, 141–165.
Minguez-Mosquera M.I., Jaren-Galen M. Garrido-Fernandez
J. (1993) Lipoxygenase activity during pepper ripening
and processing of paprika. Phytochemistry, 32, 1103–1108.
Mittler R. (2002) Oxidative stress, antioxidants and stress
tolerance. Trends in Plant Science, 7, 405–410.
Mukherjee S.P., Choudhuri M.A. (1983) Implications of
water stress-induced changes in the levels of endogenous
J.J. Rıos et al. Selenium and antioxidant system in lettuce plants
Ann Appl Biol 154 (2009) 107–116 ª 2008 The Authors
Journal compilation ª 2008 Association of Applied Biologists
115

ascorbic acid and hydrogen peroxide in Vigna seedlings.
Physiologia Plantarum, 58, 166–170.
Nakano Y., Asada K. (1981) Hydrogen peroxide scavenged
by ascorbate specific peroxidase in spinach chloroplast.
Plant and Cell Physiology, 22, 867–880.
Pedrero Z., Madrid Y., Camara C. (2006) Selenium species
bioaccessibility in enriched radish (Raphanus sativus):
a potential dietary source of selenium. Journal of
Agricultural and Food Chemistry, 54, 2412–2417.
Pedrero Z., Madrid Y., Hartikainen H., Camara C. (2008)
Protective effect of selenium in broccoli (Brassica oleracea)
plant subjected to cadmium exposure. Journal of
Agricultural and Food Chemistry, 56, 266–271.
Rao M.V., Paliyath C., Ormrod D.P. (1996) Ultraviolet-B
radiation and ozone-induced biochemical changes in
the antioxidant enzymes of Arabidopsis thaliana. Plant Phys-
iology, 110, 125–136.
Rao M.V., Paliyath C., Ormrod D.P., Murr D.P., Watkins
C.B. (1997) Influence of salicylic acid on H2O2 production,
oxidative stress and H2O2-metabolizing enzymes: salicylic
acid-mediated oxidative damage requires H2O2. Plant
Physiology, 115, 137–149.
Rıos J.J., Rosales M.A., Blasco B., Cervilla L.M., Romero L.,
Ruiz J.M. (2008) Biofortification of Se and induction
of the antioxidant capacity in lettuce plants. Scientia
Horticulturae, 116, 248–255.
Rosales M.A., Ruiz J.M., Hernandez J., Soriano T., Castilla
N., Romero L. (2006) Antioxidant content and ascorbate
metabolism in cherry tomato exocarp in relation to tem-
perature and solar radiation. Journal of the Science of Food
and Agriculture, 86, 1545–1551.
Sairam R.K., Rao K.V., Srivastava G.C. (2002) Differential
response of wheat genotypes to long term salinity stress
in relation to oxidative stress, antioxidant activity and
osmolyte concentration. Plant Science, 163, 1037–1046.
Shigeoka S., Takeda T., Hanaoka T. (1991) Characterization
and immunological properties of selenium-containing glu-
tathione peroxidase induced by selenite in Chlamydomonas
reinhardtii. Biochemical Journal, 275, 623–627.
Smorklji P., Pograjc L., Hlastan-Ribic C., Stibilj V. (2005)
Selenium content in selected Slovenian foodstuffs and
estimated daily intakes of selenium. Food Chemistry, 90,
691–697.
Takeda T., Ishikawa T., Shigeoka S. (1997) Metabolism of
hydrogen peroxide by scavenging system in Chlamydomo-
nas reinhardtii. Physiologia Plantarum, 99, 49–55.
U.S. Department of Agriculture. (2003) Dietary reference intakes:
elements [WWW document]. URL http://warp.nal.usda.gov/
fnic/etext/000105.html%3E [accessed on 5 March 2006].
Xue T., Hartikainen H., Piironen V. (2001) Antioxidative and
growth-promoting effect of selenium on senescing lettuce.
Plant and Soil, 27, 55–61.
Selenium and antioxidant system in lettuce plants J.J. Rıos et al.
116 Ann Appl Biol 154 (2009) 107–116 ª 2008 The Authors
Journal compilation ª 2008 Association of Applied Biologists




















