
L’ Ardèche à la fin de l’âge du Bronze et aux âges du Fer (IXe siècle-Ier siècle avant notre ère)

L'Anthropologie 105 (2001) 369-408
© 2001 ,~ditions scientifiques et m6dicales Elsevier SAS. Tous droits r6serv6s
Presentation pal~o-environnementale du remplissage de la Baume Moula-Guercy ~ Soyons (Ard~che) : implications pal~oclimatiques et chronologiques
Alban Defleur a*, l~velyne Cr6gut-Bonnoure b, Emmanuel Desclaux c, Michel Thinon d
R6sum6 - Les fouilles programm6es r6alis6es dans la Baume Moula-Guercy, depuis 1992 ont permis la mise au jour d'un tr6s abondant mat6riel faunique et d'une s6rie repr6sentative de charbons de bois dans les couches XIV et XV. L'6tude des grands mammif6res et des rongeurs permet de proposer une hypoth6se sur la chronologie de la raise en place des d6p6ts s6dimentaires. II est possible de distinguer trois phases climatiques majeures. A la base du remplissage, un ensemble froid qui peut &re paral- 161is6 avec la fin du P16istoc6ne moyen (stade 6 de la chronologie isotopique marine), au sommet une nouvelle phase fra]che fi froide, correspondant au d6but du stade 4, et, entre les deux, une formation homog6ne et 6paisse qui correspond a l'l~6mien sensu lato (stade 5). La richesse de cette formation interm6diaire apporte des donn6es in6dites et essentielles a la connaissance de cette p6riode actuellement fort mal connue en Europe m6diterran6enne. L'accent est plac6 sur la couche XV qui a livr6 une riche s~rie de restes humains n6andertaliens correspondant fi six individus cannibalis6s. ~') 2001 l~ditions scientifiques et m6dicales Elsevier SAS
biochronologie / faune / flore / Pleistocene moyen / France / c l imat
Abstract - Paleoenvironmental Presentation of the Filling from Baume Moula-Guercy, Soyons (Ard~che): Palaeoclimatic and Chronological Implications. Since 1992 excavations at Baume Moula-Guercy (Soyons, Ard6che, France) have been pro- viding abundant archeological, paleontological and anthracological remains, especially levels XIV and XV. Data concerning large mammals and micromammals enable us to elaborate a biochronological hypothesis. Three main climatic cycles can be dis- tinguished. Lower stratigraphical levels of deposits are representative to a cold period which can be related to the end of the Middle Pleistocene (stage 6 of the oceanic temperature curve, Saalien). Upper stratigraphical levels are also representative to a cold period corresponding to the start of the stage 4 of the oceanic temperature curve (Upper Pleistocene, Weichselian). Between this levels a homogeneous and thick stratigraphical level containing abundant temperate charcoal and faunal remains cor- responding to Eemian period sensu lato (Upper Pleistocene, stage 5 of the oceanic temperature curve) has been identified. The wealth of documentation of these intermediate stratigraphical levels provides main and necessary data concerning this period little-know in Mediterranean Europe. Level XV which provided a large number of Neanderthal remains corresponding to six individuals (MNI) is emphasised in this paper. (~ 2001 I~ditions scientifiques et m6dicales Elsevier SAS
biochronology / fauna / flora / Middle Pleistocene / France / c l imate
1. Presentation g6n6rale
Le site fut d~couvert en 1970, sond6 en 1972, puis fouill6 sans interruption entre 1975 et 1982,
sous la direction de Pierre Payen. En 1991, le gisement a fait l 'objet d 'un sondage qui permit de mettre au jour une s6rie de treize restes humains dont certains pr6sentaient des traces de
* Correspondance et tir6s fi part.
a UMR 6559 du CNRS Laboratoire d'anthropologie, facuit6 de m6decine de Marseille, secteur Nord, boulevard Pierre-Dramard, 13916 Marseille cedex 20, France.
Laboratory for human Evolutionary Studies, 3101 Valley Life Sciences Building, Berkeley, CA 94720-3160, l~tats-Unis.
b Museum Requien, 67, rue Joseph-Vernet, 84000 Avignon, France.
c Laboratoire de pr~histoire du Lazaret, Parc de la Villa "La C6te", 33 bis, boulevard Franc-Pilatte, 06300 Nice, France.
d Laboratoire de botanique et d'6cologie m6diterran6enne, facult6 des sciences de Saint-J6r6me, 13397 Marseille cedex 20, France.

370 A. Defleur et al.
Figure I. Localisation de la Baume Moula-Guercy.
Figure I. Baume Moula-Guercy, site placement.
Baume 1o0t ou,a!uerc! 0 500 1000 1500 2000 2500
ddcarnisation ainsi que des traces de fracturation sur os frais (Defleur et al., 1993a, b). Depuis 1992, des fouilles programm6es rdguliares sont r6alisdes ~ raison de 8 fi 10 semaines par an, sous la direction de l 'un d'entre nous (A.D.). Le site qui se pr6sentait/~ l'origine comme un abri sous roche/t fait place ~t une grotte de vastes dimen- sions. 11 est situ6 ~ environ 10 km au sud de Valence, sur la rive droite du Rh6ne et occupe le flanc est du Serre de Guercy, sur la commune de Soyons,/t moins de 100 m au dessus du fleuve et fi une vingtaine de m6tres de la Baume Ndron (figure 1) (Defleur et al., 1993c, 1994a).
Le remplissage est actuellement connu sur une 6paisseur de 8 m. Cependant, les niveaux de base (couches XVI ~ XIX) ne sont encore connus que par les r6sultats d 'un sondage pro- fond qui n 'a pas permis de rencontrer le substra- tum, ainsi, nos connaissances sur ces niveaux reposent essentiellement sur les rongeurs qui y sont rencontr6s en abondance. En revanche, la fouille des couches IV /i XV, a permis de recueillir un abondant matdriel paldontologique, archdologique et de mettre au jour une riche sdrie de restes humains ndandertaliens, cor- respondant fi six individus cannibalisds (Defleur 1995 ; Defleur et al., 1999).
2. Les grands mammif~res
En 1987, les niveaux sup6rieurs de ce gise- ment livraient une faune de grands mammif6res caract6ristique d'une p6riode froide du WOrm ancien (Cr6gut-Bonnoure et Gu6rin, 1986). Les nouvelles fouilles arch6ologiques r6alis6es entre 1992 et 1999 ont foumi un nouveau mat6riel pal6ontologique. L'attribution et la r6partition stratigraphique des grands mammif6res est la sui- vante:
- couche IV : Canis lupus, Vulpes vulpes, Ursus spelaeus, Panthera pardus, Crocuta spe- laea, Rhinocerotidae ind6t., Equus sp., Cervus elaphus, Rang(fer tarandus, Bos ou Bison, Capra sp., Rupicapra sp. ;
- couche V-VII : C. lupus, V. vulpes. U. spe- laeus, Felis silvestris, Equus sp., C. elaphus sub. sp., R. tarandus, Capra sp. ;
- couche VIII : C. lupus. V. vulpes, U. arctos, Mammuthus primigenius, Equus sp., C. elaphus, R. tarandus, Capra sp., Bos ou Bison ;
couche IX : R. tarandus, Equus sp., cl: Rhinocerotidae ;
- couche X : Equus sp., Sus scrq/& C. ela- phus, C. capreolus, R. tarandus ;
- couche X1 : ? Vulpes-? Alopex, C. spelaea,

Pr6sentation pal6o-environnementale du remplissage de la Baume Moula-Guercy 371
U. arctos, Equus sp., C. elaphus sub. sp., R. ta- randus, Bison sp., Capra sp. ;
- couche XII : Ursus cf. deningeri, V. vulpes, Equus sp., Rhinocerotidae ind6t., C. elaphus, C. capreolus, R. tarandus ;
- couche XIII : Cuon alpinus, Ursus cf. deningeri, U. arctos, U. thibetanus, Meles sp., R pardus, E silvestris, C. elaphus, R. tarandus, Bos ou Bison, Caprini ind6t. ;
- couche XIV : C. lupus, V. vulpes, Ursus cf. deningeri, U. arctos, Meles sp., R spelaea, P. pardus, E silvestris, Equus sp., Dicerorhinus sp., S. scrofa, C. elaphus cf. simplicidens, C. capreo- lus, R. tarandus, cf. Bison, C. ibex ainsi que Testudo hermanni-graeca pour les Reptiles ;
- couche XV : C. lupus, V. vulpes, C. alpinus, Ursus cf. deningeri, U. thibetanus, Ursus sp., Meles meles, Mustela palerminea, P. pardus, L. spelaea, Felis cf. minuta, Dicerorhinus sp., Equus cf. germanicus, S. scrofa, C. elaphus, Dama sp., C. capreolus, R. tarandus, Bos primi- genius, Capra aff. caucasica, Rupicapra sp.
Quelques unes de ces esp6ces apportent des donn6es pour la biochronologie du site.
Couche IV
La longueur de la M2 inf~rieure attribuable au loup (l 1,9 ram) s'int6gre dans l ' intervalle de variation de la population de Jaurens/~ Nespouls (Corr6ze) d'fige pl6istoc6ne sup6rieur moyen (n = 6 ; extremes - 10,1-13,9 ; m - 11,62 mm ; Ballesio, 1979). C 'est aussi le cas du mat6riel postcr~nien.
Les P4 inf6rieures de l 'ours sont 61argies pos- t6rieurement et multitubercul6es, ce qui est caract6ristique d 'un U. spelaeus 6volu6. Le morphotype est interm6diaire entre C1 et C2 d6fi- ni par Rabeder (1992). Toutefois, en l 'absence de P4 sup6rieure associ6e il est impossible de corn- parer la population de Moula avec les ours des cavernes europ6ens d'fige connu. La hy6ne a lais- s6 une camassibre inf6rieure sans m6taconide. Cette absence est r6v61atrice d 'un animal 6volu6 au sein de la lign6e Crocuta, la r6duction et la disparition de ce denticule 6tant plus fr6quentes chez les formes wfirmiennes. La diffdrence de longueur entre paraconide et m6taconide est importante (4,1 ram) et situe I 'individu de Moula dans les populations des hybnes wfirmiennes, cette diff'drence ne d6passant pas 2 mm au Riss.
Le degr6 d'6volution de l 'ours des cavernes et de la hy6ne ainsi que la massivit6 du loup concordent pour dater ce niveau de la fin du WOrm ancien ou du d6but du WOrm r6cent.
Couehe V-VII
Parmi les restes post-crfiniens du loup se trouve un m6tatarsien IV entier dont la longueur est de 102,2 mm. Cette valeur correspond fi celle des plus gros individus de Jaurens chez qui l ' am- plitude de variation est de 81 fi 103,7 mm (n = 9, m = 92,7 mm ; Ballesio, 1979).
La P2 sup6rieure du cerf 61aphe, fi face lin- guale clairement bilob6e, mesure au niveau occlusal 18,4 mm de long pour une largeur de 11,0 mm et fi la base 16,0 mm de large. Elle s ' in- t6gre dans le domaine de variabilit6 du Cervus elaphus sub. sp. du WOrm ancien sup6rieur de Combe Grenal (Domme, Dordogne) (respective- ment 14,9 fl 19,1 mm (n = 19, m = 17,12 ram), 11,3/l 18,0 mm (n = 16, m = 14,38 mm), 14,1 /t 20,0 mm (n = 17, m = 16,25 ram) ; Guadelli, 1987). La faune de ce niveau, comme pr6c6dem- ment, est caract6ristique du WOrm ancien sup6- rieur ou du tout d6but du WOrm r6cent.
Couehe VIII
Le loup poss6de des dents m6triquement pro- ches du maximum not6 chez le loup de Jaurens. La P4 inf6rieure est longue de 18,4 mm et large de 8,7 mm, la variation enregistr6e fi Jaurens 6tant respectivement de 14,8-17,4 (n = 15 ; m - 16,23) et 7,2-9,5 (n - 15 ; m = 8,37 ; Ballesio, 1979).
Les proportions des dents jugales du renne se situent dans l ' intervalle de variation de la popu- lation de Combe Grenal : la longueur de la M1 sup6rieure est de 17,5 mm, celle de M2 de 19,5 mm et celle de M3 de 19,9 ram, la valeur moyen- ne du cerf de Dordogne 6tant de 18,46 mm (n = 32 ; extr6mes = 15,0-20,9), 20,27 mm (n - 37 ; extr6mes - 16,1-23,2) et 19,72 mm (n - 120; extremes - 15,3-22,8).
Couches IX et X
La faune de ces couches est r~duite et frag- mentaire et ne fournit aucune donn6e biochrono- logique.
Couche XI
Si l 'on se base sur la molarisation de la P4 inf~rieure du cerf 61aphe, cet horizon stratigra- phique pourrait correspondre au Wfirm ancien. En effet, le paraconide et l 'entoconide sont fusionn6s mais ce dernier denticule est s6par6 de l 'entoconi- de par un sillon. Cette morphologie s 'observe chez Cervus elaphus sub. sp. des niveaux du WOrm ancien sup6rieur de Combe Grenal. Les proportions dentaires confirment cette analogie. En effet, la longueur occlusale de la P4 est de 21,4

372 A. Defleur et al.
mm, d6passant le maximum de C. e. simplicidens (extremes = 16,5-20,0 mm ; n = 31 ; m = 17,85), mais pas celui de C. elaphus sub. sp. (extremes = 16,9-22,4 ; n = 49 ; m = 19,68).
Couche XII
La MI inf6rieure de l'ours est r6duite/l un tri- gonide non us6. Le paraconide, puissant, poss6- de un flanc m6sial sub-vertical. Le m~taconide est d6doubl6, le denticule ant6rieur 6tant peu prononc6. Cette morphologie est typique d'U. deningeri (Prat, in : Laville et al., 1972). La P4 sup~rieure, non us6e et sans tubercules accessoi- res, pr6sente un parac6ne 61ev6 et un m6tac6ne bas dont les crates forment un angle 16g6rement obtu. Le deut6roc6ne, situs dans l'axe de l'inci- sure vestibulaire, est lingualement bien dilat6 ; en cons6quence l'indice de robustesse est de 81,28 % ce qui est conforme ~ l 'ours de Deninger, cet indice 6rant rarement sup6rieur/~ 75 % chez U spelaeus (Cr~gut-Bonnoure, in : Defleur et Cr6gut-Bonnoure, 1995).
L'ours de Deninger acquiert le stade 6volutif d'U. spelaeus ~ la fin du P16istoc6ne moyen. Cependant, les premi6res populations ont encore des caract6res archa'fques : c'est le cas ~ la grot- te de La Niche (Montmaurin, Haute-Garonne ; Tavoso et al., 1990), fi la grotte des C6dres (Le Plan d 'Aups, Vat ; Cr6gut-Bonnoure, in : Defleur et Cr6gut-Bonnoure, 1995) et/~ Az6 I-3 et 1-2 (Sa6ne-et-Loire ; Argant, 1991). Compte- tenu de la forte variabilit6 individuelle existant au sein de la lign6e deningeri-spelaeus et du peu de mat6riel disponible, il parait pr6f6rable, pour l'instant, de garder une certaine r~serve quant la position sp~cifique de l'ours de cette couche qui toutefois est plus primitif que celui des niveaux sus-jacents.
Couche XIII
Le talonide d'une carnassi6re inf6rieure, uni- cuspide et sans cingulum disto-lingual, permet d'identifier Cuon alpinus. La dur6e de vie de I'esp6ce couvre le Pl6istoc6ne moyen final (C. alpinus jbssilis) et le P16istoc6ne sup6rieur (C. alpinus europaeus). Actuellement, la position sous-sp6cifique des restes recueillis dans ce site ne peut 6tre pr6cis6e.
Un ours de la lign6e sp616enne est figur6 par une P4 sup6rieure trbs semblable au sp6cimen de la couche XII (indice de robustesse 82,20 %) ainsi que par deux P4 inf&ieures 6troites et peu multitubercul6es. La M2 inf&ieure, faiblement bilob6e, a un lobe m6sial plus long et plus 6troit que le lobe distal (18,1 et 18,6 ram). La cr6te
reliant le protoconide et le m6taconide est tran- chante, ee qui est conforme ~ U. deningeri (Prat, in : Prat et Thibault, 1976). La M3 inf6rieure, au protoconide aussi saillant que celui d'U. denin- geri, est longue de 26,3 mm. Cette dimension cadre avec la variabilit6 de I'U. deningeri de la br6che de Chateau/t Cluny (Sa6ne-et-Loire) et du site d 'Az6 I-l , dat6s du Pl6istoc6ne moyen m6dian, soit en moyenne respectivement 26,10 mm (n = 17, extr6mes = 23,2-29,6 mm) et 24,20 mm (n = 6, extremes = 21,0-26,0 ram). En cons6quenee, les donn6es morphom6triques per- mettent de rapprocher l'ours de Moula d'U. deningeri, sans qu'il soit ~ nouveau possible d'appr6cier la variabilit6 individuelle.
U. thibetanus est identifiable grace ~ la moi- ti6 proximale d 'un m6tatarsien Ill de petite taille et ~ diaphyse gracile. Sa morphologie est iden- tique au sp6cimen de la grotte des C6dres (Cr6gut-Bonnoure, in : Defleur et Cr6gut-Bon- noure, 1995).
C. alpinus indique que la couche XIII est plus r6cente que le Pl6istoc6ne moyen ancien et l'existence d'un ours proche, sinon identique, U. deningeri, montre qu'elle est ant6-wfirmien- he. Son association avec U. thibetanus rappelle la faune de la grotte de C6dres, dat6e de la fin du P16istoc6ne moyen ainsi que celle de Blanot 2, d'fige 66mien, qui a livr6 des restes de U. spe- laeus, U. arctos, U. thibetanus, Martes martes, M. meles, E minuta, E silvestris, P. pardus, E. caballus, E. hydruntinus, C. elaphus, C. capreo- lus, Bos ou Bison, S. scrofa (Argant, 1991 ).
Couehe XIV
Parmi les restes attribuables au loup se remarque un condyle occipital ~ peine plus allon- g6 (21,2 ram) que celui des petits Ioups du Pl6istoc6ne moyen rdcent de La Fage ~ Noailles (Corr6ze), de la grotte des C6dres et de la Niche (Cr6gut-Bonnoure in : Defleur et Crdgut-Bon- noure, 1995) tout en 6tant plus 6pais (13,9 ram). La Iongueur de la MI sup&ieure est, au niveau du mdtac6ne, de 21,9 mm. Cette dimension ddpasse la limite sup6rieure de la variation enregistrde chez C. lupus santenaisiensis de la fin du Pldistocbne moyen dont la longueur de la tuber- culeuse est comprise entre 15,0 et 17,0 mm (n - 18, Argant, 1991 ). 11 en est de marne pour les aut- res 616ments dentaires. Les mdtapodes, graciles, sont plus petits que ceux des loups du Pldistocbne supdrieur mais plus grands que ceux du Canidae de la Fage; ainsi, le m6tacarpien IV est long de 88 ram, le m6tacarpien II de 73,3 ram.
Pr6sentation pal6o-environnementale du remplissage de la Baume Moula-Guercy
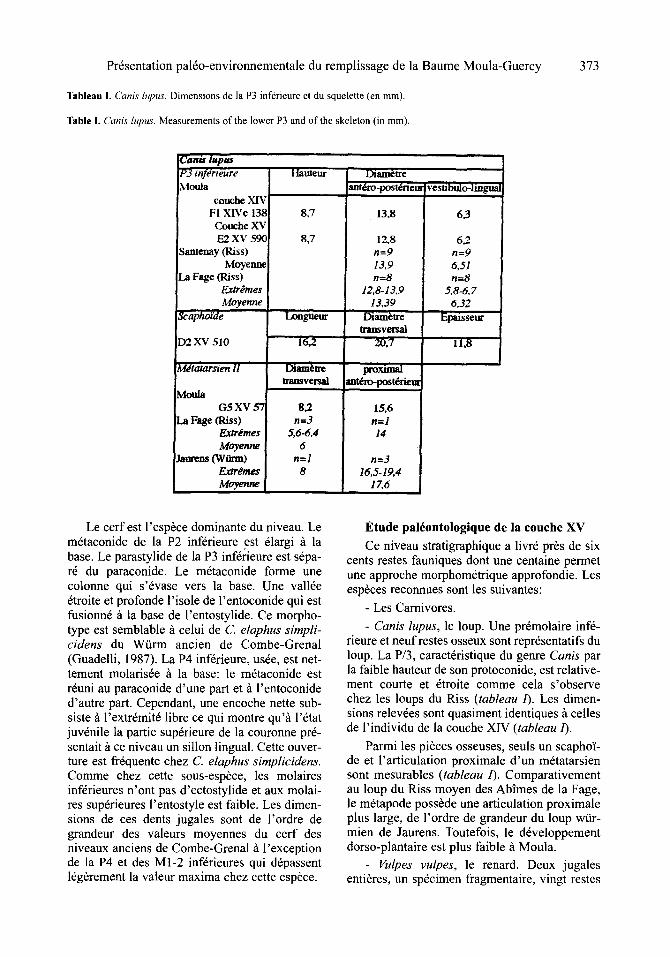
Tableau I. Canis lupus. Dimensions de la P3 infdrieure et du squelette (en ram).
Table I. Canis lupus. Measurements of the lower P3 and of the skeleton (in mm).
373
Canis l=pm P 3 i n f g r i e u r e
Moula couche XIV FI XIVc 13~ Couche XV E2 XV 59£
Santenay (Riss) Moyennc
La Fage (Riss) Extr&mes Moyenne
Scapho'fde
D2 XV 510
M#tatarsten I1
Moula G5 XV 5"~
La Fage fRiss) Extremes Moyenne
Jaurens (Wiiam) Extr#mes Moyenne
~uteur
8,7
8,7
, i , ,
l.xmgueur
16,2
Di~tre transversal
S~ n=3
5,6-6,4 6
n=l 8
Dimn~Are ant6rO-l~St/.~rleur
13,8
12,8 n=9 13,9 n=8
12.8-13,9 13,39
Diam~tre Iransvety~
,,, ,, ,, 20,7
proximal [ant~o-post6riem
15,6 n=1 14
n=3 16,5-19,4
17.6
vestibulo-lingual6,3 [
6,2 n=9 6,51 1 ' 1 = 8
5,8-6, 7 6,32
11,8
Le cerf est l'esp6ce dominante du niveau. Le m&aconide de la P2 inf6rieure est 61argi ~t la base. Le parastylide de la P3 inf6rieure est s6pa- r6 du paraconide. Le m6taconide forme une colonne qui s'6vase vers la base. Une vall6e 6troite et profonde l'isole de l'entoconide qui est fusionn6/l la base de l'entostylide. Ce morpho- type est semblable/t celui de C. elaphus simpli- cidens du Wtirm ancien de Combe-Grenal (Guadelli, 1987). La P4 inf6rieure, us6e, est net- tement molaris6e h la base: le m6taconide est r6uni au paraconide d'une part et/l l'entoconide d'autre part. Cependant, une encoche nette sub- siste/l l'extr6mit6 libre ce qui montre qu'/t l'6tat juv6nile la partie sup6rieure de la couronne pr6- sentait ~ ce niveau un sillon lingual. Cette ouver- ture est fr6quente chez C. elaphus simplicidens. Comme chez cette sous-esp6ce, les molaires inf6rieures n'ont pas d'ectostylide et aux molai- res sup6rieures l'entostyle est faible. Les dimen- sions de ces dents jugales sont de l'ordre de grandeur des valeurs moyennes du cerf des niveaux anciens de Combe-Grenal ~ l'exception de la P4 et des MI-2 inf6rieures qui d6passent 16g~rement la valeur maxima chez cette esp6ce.
Etude pal6ontologique de la couche XV
Ce niveau stratigraphique a livr6 pros de six cents restes fauniques dont une centaine perrnet une approche morphom6trique approfondie. Les esp~ces reconnues sont les suivantes:
- Les Carnivores.
- Canis lupus, le loup. Une pr6molaire inf6- rieure et neufrestes osseux sont repr6sentatifs du loup. La P/3, caract6ristique du genre Canis par la faible hauteur de son protoconide, est relative- ment courte et 6troite comme cela s'observe chez les loups du Riss (tableau 1). Les dimen- sions relev6es sont quasiment identiques ~ celles de l'individu de la couche XIV (tableau 1).
Parmi les pisces osseuses, seuls un scapho~- de et l'articulation proximale d'un m6tatarsien sont mesurables (tableau 1). Comparativement au loup du Riss moyen des Abimes de la Fage, le m6tapode poss6de une articulation proximale plus large, de l'ordre de grandeur du loup wfir- mien de Jaurens. Toutefois, le d6veloppement dorso-plantaire est plus faible ~ Moula.
Vulpes vulpes, le renard. Deux jugales enti6res, un sp6cimen fragmentaire, vingt restes

374 A. Defleur et al.
Tableau II. Vulpes vulpes. Dimensions des dents jugales inf6rieures et du squelette (en ram).
Table I1. Vulpes vulpes. Measurements of the lower cheek teeth and of the skeleton (in mm).
Prdmolaires 'tnfdrieures 1'2
DI XV 178 Gerde (mss ~
Extremes Moyenne
P3 G2VX 400
Lcs ~ s (Riss) F..ttr~Mtes
~ad¢ (Piss Extrg, mes Moyenne
'FI XV 358
U/no
I'-12 XV 51
Mdtapodes
M~tacarpien Ill JV XV tamis
~t4~tacarpien V Moula
GV XV tamis Les C~res (Piss) [4dtalarsien V
D2 XV 460
I 4 a ~
5 n=23
4,7-5,8 5,3
5,3 n=2
5d-5.6 n=3a ¢
4,9..6,1 5,6
I )hu~t~ b'llasv~,al
19,2
Dia~a~tre mt~o-pos~'ri©m
13.7
Loag~ur
3717 39,6
7,4 n=35
7,2-9,4 8,4
8~ n=2
1~1~1 n=48
~6
proximal ~ g ' t i e t a
25,6
tramversal 15,2
Extr6.mit~
ant(m3-post~a-iem"
82
6,5 5,9-7
8
¢xamv~al
2,8 n=36
2,9-3,7 3,3
3 3 n=2 3,5
n=47 ~0-4,0
~6
Ar~cula~on
I-lamct~ amculailt
10,7
i~roxi~e
mmsve~sal
52
7 6,9-7.6
72
Diam/:tl¢ transversal
9,3
Mtlicu Diam~trc
ml~o-pos~r, riem
3,4
3.2
dmphys¢
transvcvsa/
4.8
4,5
FAtxaSmit~ daslalc Dian~tre
mt6m--post6riem tramvers~
5,9 6,8
n=l n=l
osseux se rapportent au renard. Le mat6riel den- taire comprend deux prdmolaires inf6rieures ce qui est insuffisant pour une bonne caractdrisation de la variabilit6 populationnelle. On peut seule- ment indiquer que les proportions dentaires sont tout ~ fait compatibles avec celles du renard commun (tableau I1). Parmi les pi6ces osseuses, une moiti6 d'humdrus, un talus, une articulation proximale d'ulna, un mdtacarpien V entier, l'ar- ticulation proximale d 'un mdtatarsien V et deux phalanges enti6res peuvent ~tre mesurds (tableau I/). Les dimensions relevdes sont ~ nouveau comparables/l celles du renard commun.
- Cuon alpinus europaeus, le dhole europden. La P4 et la M1 supdrieures droites d 'un m6me individu permettent d'identifier le dhole euro- p6en. La M1/est caractdrisde par l'absence d 'hy- poc6ne, le talon interne ne possddant que le protoc6ne, et de torsion de la couronne vers l'ar- ri6re ainsi que par la pr6sence d'une seule
d6pression, arrondie et profonde, au centre du talon interne. Cette fosse est antagoniste ~ l 'hy- poconide de la carnassi6re inf6rieure. La conca- vit6 m6diane de la face vestibulaire est/~ peine esquiss6e. L'ensemble de la morphologie est conforme au genre Cuon. On ne rel6ve aucune d6pression r6v61atrice du cingulum disto-lingual de la M/1 antagoniste. Cette absence exclut C. priscus connu darts un petit nombre de gisements du P16istoc6ne moyen (Mosbach, Caune de l 'Arago, Montsaun6s, Hundsheim, Lunel-Viel, La Niche, Verz6 et Font6chevade ; Cr6gut- Bonnoure, 1996a) et permet d'identifier C. alpi- nus. II est int6ressant de noter que la couche Xlll a fourni le talonide d'une M/I dont la morpholo- gie est caract6ristique de cette esp6ce et pr6sente donc les caract6res morphologiques compl6men- taires de ceux de la M1 sup6rieure de la couche XV. Les dimensions des deux jugales sup6- rieures sont de l'ordre de grandeur des individus

PrOsentation palOo-environnementale du remplissage de la Baume Moula-Guercy
Tableau Ill. Cuon. Dimensions des dents jugales supOrieures (en mm).
Table III. Cuon. Measurements of the upper cheek teeth (in mm).
375
Cuon
P4 su#r leure
Cuon europaeus Moula
O1 15 221
CUOIZ pr~cIt$ Ltmel -Viel Catme de l'Arago
MI Sul~rieure Cuon europaeus Moula
Ol XCV 9-91 C~on priscus
Luncl-Viel Caune de l'Arago La Niche
c t t ~ a /p /n~ C-¢rde
[Ob~rvatoire Extr~rnes Moyenne
Oiam~Uae ~at ~ro-pos t~rieurJ ve stil~lo-lingual I
/ I
22,3
22,4 20.5
13,4
14.4 10,5 12,5
12,6 n=3
12.8-14 13,3
distal
11,5 8,6
10,8 8,3 10,9
16 14,2
16,5 15 12,7
17,3 15,3
13,3 13,9 n=3 n=3
13,3-15,9 13,5-13,9 t4,83 13.6
de la grotte de l'Observatoire (Monaco) et de la grotte de la Carri~re fi Gerde (Clot, 1980) qui ont servis de terme de comparaison (tableau II1).
C. alpinus est signal6 darts une trentaine de sites d 'Europe occidentale s'6chelonnant du Pl6istoc6ne moyen final (Riss) au Pl6istoc6ne sup6rieur. Au moins deux espbces sont pr6sentes: C. alpinus fossilis du Pl6istoc~ne moyen, d6crit des brbches d'Heppenloch, et C. alpinus euro- paeus du P16istoc6ne sup6rieur (caverne de Mars
Vence, Alpes-Maritimes ; grotte de l'Obser- vatoire) (Bonifay, 1971). Les diffOrences entre ces deux taxons concernent les jugales inf6rieu- res. Le mat6riel de la couche XV en est d6pour- vu. Cependant, si l 'on tient compte des propor- tions dentaires, l'individu de Moula est identique
C. alpinus europaeus.
- Ursus cf. deningeri, l'ours de Deninger. Parmi les restes attribuables aux Ursidae une M2 inf6rieure enti6re, un fragment m6sio-distal de M3 inf6rieure et une hemi-mandibule presque compl6te correspondent ~ un ours de la lign6e deningeri-spelaeus. La deuxi6me molaire est comparable fi celle de l'individu de la couche XIII. La face linguale est rectiligne et la face ves- tibulaire est bilob6e (figure 2) : l'allure g6n6rale
est donc moins 6trangl6e que chez U. spelaeus. Le lobe m6sial est plus ~troit que le lobe distal : respectivement 16,6 et 17,1 mm. Les m6mes pro- portions s'observent sur le sp6cimen de la couche XIII tandis que l'individu de la couche IV, typi- quement sp616en, poss6de des lobes de largeur sub-6gale (tableau IV). Le m6taconide est com- pos6 de trois tubercules : un m6dian, le plus 61ev6, un ant6rieur, en position plus basse et un denticule post6rieur bien d6velopp6, tout en 6tant plus faible que la cuspide principale. La cr&e reliant le protoconide et le m6taconide est nette- ment tranchante, ce qui est conforme ~ U. denin- geri (Prat, in : Prat et Thibault, 1976). I1 faut ajouter une premi6re phalange d'adulte. La dia- physe, moins massive que celle d'U. spelaeus pr6sente une 16g6re dissym6trie des faces lat6ra- les, ce qui permet d'6carter l'ours brun.
- Ursus thibetanus, l 'ours du Tibet. Un navi- culaire est attribu6/~ l'ours du Tibet. En rue dis- tale, le bord dorsal est clairement bilob6 (figures 3 et 4). En vue dorsale, l 'os est aminci au niveau de la facette pour le troisi6me cun6iforme. Cette morphologie est comparable/~ celle de I'U. thi- betanus des niveau rissiens de la Niche et claire- ment distincte de celle de l'ours brun (Cr6gut-

376 A. Defleur et al.
Tableau IV. Ursus deningeri. Dimensions de la M2 inf6rieure et du squelette (en mm).
Tab l e IV. Ursus deningeri . Measuremen t s o f the lower M2 and o f the skeleton (in ram) .
ffnus d e ~ , e n
M2 in~rieure Moula
DI XV lOq ft. sptk~u~
i Couche IV 1:5 IVc 41
iCouehe XIII D2 XIII 3- ~
Prera~re plmlaage
Moula U n ~ ~ e r ~ Couehe XV
D1 XV 13'~ U n ~ sp. Couche XV
F2 XV 344
U n ~ th i~ tan~
Navfculagre Moula
Couehe XV G2 XV 393
L~ Niche (Piss) T 175
Ungs arctos Los C~lres (piss)
Longueur totale
29,3
30,3
29,3
Longueur
44.3
38,5
Hauteur
Hauteur protoconide
11,1
us~
9,9
Extrerait~ Diamette
ant~ro-post~rieur
16,2
13
I~am6tr¢ tnmsversal
hypocomde
10,1
us~
9,4
proxtmale
transversgd
22.1
antero-pos~em
m*~taconide
11,1
US~
10,6
Milieu Diam6tre
mt~ro-post6rieu~
9,9
8.4
e.toconide
10,1
u~
11,5
diaphyse
transversal
13.4
12,6
/.~rg~ur tngomde
16,6
17.9
18,1
F ~ i t ~
wt~ro-post~rieur
11,5
8,5
13,4 34,8 32
17,9 33,6 35,4
15 34.3 39,9
talonide
17,1
18,2
18,6
distale
Iransversai
16,5
12.5
T a b l e a u V. Meles meles. Dimens ions de la mandibu le (en m m ) .
Tab l e V. Meles meles. Measu remen t s o f the mandibu le (in m m ) .
Mandibule ~Aoula
Couche XV H5 XV
Blanot 2 "E.ctr~mes
Ecartement exleme Longueur camnes incislves PI -P4
20.6 13.7 20,6
P3 - Diam~tr¢ m~r~p~ne~
5,5 n=2
5,7-5,8
vestlbulo-I ingtul
3,4 n=2
3.6-3. 7
P3 '- Dlamet~ snt6ro-pos~rieur
6,6
vestibalodingual
Bonnoure, 1996b). Les dimensions de ce tarsien sont fournies dans le tableau IV.
- Ursidae ind6termind. Quatre dents de lait, une canine corrodde et la racine d 'un autre spd- cimen ainsi qu'une premi6re phalange d'adulte sont laiss6s en nomenclature ouverte. Ces 61d- ments peuvent correspondre/~ rours du Tibet ou bien g rours brun.
- Meles meles, le blaireau. Quatre restes den- taires peuvent ~tre attribuds au blaireau: la partie rostrale d 'une mandibule avec les alv6oles des
incisives, canines, Pl et P2 gauche et possddant encore la P3 et la P4 gauches, le germe de la M 1 supdrieure gauche et droite ainsi que celui de la P4 gauche d 'un m6me animal. Un mdtacarpien II sans articulation distale correspond ~t ce dernier individu. Enfin, il existe une troisi6me phalange d'adulte. La fossilisation de ce matdriel est com- parable ~ celle de la faune de la couche. Aucune diffdrence morphologique n'existe par rapport au blaireau actuel. Les dimensions qu' i l est pos- sible de prendre sont fournies dans le tableau V

P r d s e n t a t i o n p a l d o - e n v i r o n n e m e n t a l e du r e m p l i s s a g e de la B a u m e M o u l a - G u e r c y 377
Figure 2. Ursus cf. deningeri, M2 inf6rieure gauche, de la couche XV de la Baume Moula-Guercy (clich6 A. Guerrand, Mus6e Calvet, Avignon). ~zchetle en mm.
Figure 2. Ursus cf. deningeri, lower second molar from level XV of la Baume Moula-Guercy (photo A. Guerrand, Mus6e Calvet, Avignon). Scale in ram.
Figure 3. Naviculaire d'Ursus thibetanus de la Niche Montmaurin (clich~ A. Guerrand, Musde Calvet, Avignon). l~chelle en ram.
Figure 3. Ursus thibetanus, navicular bone, from La Niche (Montmaurin), (photo A. Guerrand, Mus6e Calvet, Avignon). Scale in ram.
Figure 4. Naviculaire d'Ursus thibetanus de la couche XV de la Baume Moula-Guercy (clich6 A. Guerrand, Mus6e Calvet, Avignon). l~chelle en mm.
Figure 4. Ursus thibetanus, navicular bone (Moula-Guercy, layer XV) (photo A. Guerrand, Musde Calvet, Avignon). Scale in mm.

378 A. Defleur et al.
Tableau V1. Mustelapalermina et Mustela nivalis. Dimensions du cr~ne (en mm).
Table VI. Mustela palermina and Mustela nivalis. Measurements of the skull (in mm).
M estelidae C~.NE
Longueur C-MI (A) PI-P4 M1-M2
Ecartement zanines
extem¢ intem~
P4 e~[erne interne
MI externe interne
1:'4 - diam~o'e m~sio-distal (externe) vestibulo-lingual (m~sial)
M1 - diam~tre m~sio-distal externe mesio-distal interne vestibulo-lingual total vestibulo-lingual m&tian
Largeur museau (canines) (B) orbitaire Jalalin
Distance bord distal M1 - ~chancmre du t~a t in Foraraen infra-orbitaire largeur ((7) hauteur (D)
ltKlices A/B CID
M. palenninea
Moula C2XV 17
%26 6,54 3.15
7,25 3.95
10,64 4,8
11,2 4.66
3,9 2,15
1,15 1,56 3,33 1,09
8,14 7,95 3,72
3,23
1,78 1,7
113,75 95,5
hi. nivafis
acluel Extremes
n--A: 11-11,59 n--.4; 7.35-8.02 n-_At; 3,31-4.03
n=4:7,83-9.49 n--4; 4,36-5,21
n=4t; 12,09-13,94 n=4:5,6-6.81
n=4; 11,94-13,18 n--4; 5,67-6,42
n=4; 4,07-4,53 n--A: 2,09-2,41
n--4; 1,4-1,57 n--A; 1,3-1,52 n--~-: 3,2-3,9 n=4; 1,7-1,94
n=4:9,46-9,7
n=4; 4,18-4,58
n=4; 4,88-5,06
n=4; 1,86-2,83 n--4: 0,91-1,67
n--d; 115,3-I 19,48 n=4; 158,08-208,0~
Moyenne
11.17 %88 3,61
8,65 4,85
12.67 6,23
12,64 6,08
4,29 2,28
1,49 1,37 3,48 1,86
8,55
4,35
4,97
2,45 1,27
116.97 196,98
On note que les proportions de la P3 infdrieure sont similaires fi celles de l'individu 66mien de Blanot 2.
- Mustela palerminea, hermine. La pattie faciale d 'un crane est attribuable /l un Mus- telidae de la taille de la belette et de l'hermine. Les s6ries dentaires droite et gauche sont conser- v6es ainsi que la P4 et la M 1 des deux c6tds. Les autres dents sont tombdes post-mortem. Le pala- tin et son 6pine nasale caudale ainsi que la par- tie rostrale des orbites sont encore pr6sentes. Comparativement a la belette, M. nivalis, le museau est court et large ; l'incisure du palatin est aussi plus courte et dessine un V tr6s net; le foramen infra-orbitaire est plus volumineux et plus haut (tableau V[). La largeur du museau et la grosseur du foramen infra-orbitaire sont plut6t compatibles avec l'hermine, M. erminea. Toute-
fois, d'apr6s Toshi (1965), la belette poss6de un museau dont la largeur interorbitaire est infdrieu- re/l la largeur rostrale au niveau des canines : ces param6tres ayant des valeurs de 7,95 et 8,1 ram, ce cr~ne serait celui d'une belette, ce qui n'est pas compatible avec le reste de la morphologie. La MI sup6rieure est en vue distale tr~s forte- ment creusde, ce qui correspond ~ la morpholo- gie de M. palerminea (Rabeder, 1976, figure 1, p. 13). Ce fragment crfinien est digne d'int6r& : /t ma connaissance il s'agit du seul sp6cimen crfinien signal6 dans le Pldistocbne europ6en. On peut aussi indiquer que quatre esp6ces sont reconnues : M. praenivalis et M. palerminea d'fige Pldistoc6ne moyen, M. nivalis et M. ermi- nea d'fige Pldistoc6ne sup6rieur. La date d'dmer- gence des deux esp6ces actuelles n'est pas connue avec certitude. La prdsence de M. paler-

Pr6sentation pal6o-environnementale du remplissage de la Baume Moula-Guercy 379
Tableau VII. Panthera pardus. Dimensions de la canine et du squelette (in mm).
Table VII. Pantherapardus. Measurements of the canine and of the skeleton (in mm).
ema, ne~am
m~o-dlsml velcabttlo-lmgnal Ca~ime
DI XV 14.7. 12,8 10,8
Ulna FI XV 6~
Bluno~ 2
Pha/ange// G3-H3 XV terrie~
F2 XV 352
16,2 21.5 20,S
Loagttettr
an~ro--ptmeneur 25,8 30,1 32.9
Dmmetre ptoxJmale
tnm~.,erml
22,7 9,7 10' 1 25, t IL l IL7
Malieu dmphyse P.,gll~all t~ distale Dialn~tre Dtml~la"e.
mtero-postenem ~ tatt~o-posl6rie,tt Oansveml
6,7 6,8 7.2 9 7,4 7,6 7.9 11,3
T a b l e a u V I I I . Lynx spelaea. D i m e n s i o n s du squelet te (in m m ) .
T a b l e V I I I . Lynx spelaea. M e a s u r e m e n t s o f the ske le ton (in ram).
Lynx s p e ~ a Articulation transversal
~adms Moula
F1 XV 36£ 15.1 :~dres (Riss) t5.8 kazaret (Riss) u=2
F~tr,~mes 14.8-15,6 Hauteur
!eis~rme G3 XV tamis
Scaphotl/e H5RXV
$caphohma/re H4XV
Premiere phalange G3 XV 228
9,01
8,6
Longueur
23,1
diam&re aut6ro-I~t6rieur
10.8 11,4
Diamime transversal
8,29
9,4
16,3
Exlr~mit~ Diami~tre
am&o-post~rieur
6,9
ant~ro-post6neu~ 17,~ '
11,8
10"8.
~xlmate
~ a l s v ~ d
8.8
Milieu Dimnetre
lant~ro-pos~rieuJ
4,9
diaphyse
trm~sversa]
Exlr6mit~
alzt~ro-post6rieut
5,1
dislale
tr~lsversal
6,7
minea dans un contexte 66mien montre que la diff6renciation de l'hermine vraie est plus tardi- v e .
- Panthera pardus, la panth&e. La pr6sence de la panth~re est attest6e par sept restes frag- mentaires. Seuls la canine, l'ulna et deux deuxi& rues phalanges sont mesurables (tableau VII). La faible courbure de la canine permet de la posi- tionner sur l'arc dentaire inf6rieur. L'extr6mit6 proximale de l'ulna a pu &re compar6e fi celle de l'individu de Blanot 2, d'fige ddmien, et d'l~tri- gny d'fige worm ancien. Elle se r6v61e 16g~re- ment plus petite. Cette diff6rence pourrait 6tre
d'ordre sexuel ou li6e fi une technique de mesu- re diff6rente.
- Lynx spelaea, le lynx des cavernes. Le lynx est figur6 par sept restes osseux. L'articulation proximale du radius, seule mesurable, est large de 15 mm et longue de 11 ram, ce qui est de l'or- dre de grandeur des lynx de la fin du Pldistoc6ne moyen (tableau VIII).
- Felis cf. minuta, chat sauvage nain. Une h6mi-mandibule entibre atteste l'existence d 'un chat sauvage (figure 5). La morphologie dentai- re est identique fi celle de E silvestris. Compa- rativement aux sp6cimens pl6istoc6nes de la

380 A. Defleur et al.
Figure 5. Felis cf. minuta, hemi-mandibule gauche, de la couche XV de la Baume Moula-Guercy (clich6 A. Guerrand, Mus6e Calvet, Avignon). t~chelle en mm.
Figure 5. Felis cf. minuta, left half mandibule, from Moula-Guercy, layer XV (photo A. Guerrand, Mus6e Calvet, Avignon). Scale in mm.
Baume P6rigaud (Tourrette-Les Vens, Var) d'fige wiirmien III, du Saut-du-Loup (Bidon, Ard6che) d'~ge azilien, de Roquefure (Bonnieux, Vau- cluse) et de Chinchon II (Saumanes, Vaucluse ; Cr6gut-Bonnoure, 1992e) d'~ge castelnovien, nous avons un individu de petite taille, proche mais en dessous du minimum enregistr6 chez le chat sauvage actuel (tableau IX). Trois formes de Felis sont connues dans le P16istocbne d'Europe occidentale : E monspessulana des niveaux min- del-riss de Lunel-Viel (Bonifay, 1971), F. silves- tris rencontr6 depuis le P16istoc+ne moyen et F. minuta signal6 en France ~ Achenheim d'~ge ris- sien, /l Flavigny-sur-Ozerain (C6te-d'Or) et Blanot 2 dat6s de l'E6mien, ainsi qu'/t Germolles (Sa6ne-et-Loire) qui date du Wtirm III (Argant, 1991). Les deux premi6res esp+ces sont plus grandes que la derni6re. Malheureusement, aucune mandibule de E minuta n'a 6t6 d6cou- verte/1 ce jour. Seule est connue une canine inf6- rieure isol6e provenant du Grand Caveau /t Flavigny (Argant, 1991). La longueur (3,8 mm) est assez proche de ce qui est mesur6 /l Moula (4,1 mm) mais pas la largeur, la canine de Bourgogne 6tant plus gr61e (3,5 mm de largeur/l Moula pour 2,7 mm ~ Flavigny).
L'attribution sp6cifique de l'h6mi-mandibule de Moula est r6serv6e. Nous n'avons que peu de donn6es sur la variation de taille de E silvestris ~t l'l~6mien comme d'ailleurs de celle de F. minu-
ta. L'individu de Moula se trouvant au-delfi de la limite inf6rieure du chat sauvage actuel, il est raisonnable de le rapprocher de F. minuta plut6t que de E silvestris. I1 est fort possible que le talus entier corresponde au m~me individu. I1 est en effet plus petit que celui du chat sauvage ris- sien des C~dres qui a servi de terme de compa- raison (tableau X). La position syst6matique de F. minuta n'est pas claire. Pour Kurt6n (1965), il faut inclure les restes de petite taille/l l'int6rieur des limites de F. silvestris. Toutefois, les quelques diff6rences morphologiques existantes sont en faveur de la validit6 de ce taxon (Argant, 1991).
- Les P6rissodactyles.
- cf. Dicerorhinus, le rhinoc6ros. Deux frag- ments dentaires et trois fragments osseux per- mettent d'identifier un rhinoc6ros. Compte tenu du contexte stratigraphique, ce mat6riel est rap- port6 avec r6serve au genre Dicerorhinus. Aucune mesure n'est possible.
- Equus cf. germanicus, le cheval. Le cheval est identifiable grfice/l cinq fragments dentaires, une M2 inf6rieure, une D2 inf6rieure, un frag- ment de c6te et un grand s6samoi'de. La D2 est trop us6e pour permettre une examen morpholo- gique. La M/2 poss6de une double boucle de type caballin (m6taconide arrondi, p6doncule 6troit, m&astylide triangulaire, sillon lingual

Pr6sentation pal6o-environnementale du remplissage de la Baume Moula-Guercy
T a b l e a u IX. Felis cf. minuta. Dimensions de la mandibule (en mm).
Table IX. Felis cf. minuta. Measurements of the mandibule (in mm).
381
M a l d k l e Longueur
P3-MI 7ardne - diam~tre
m~sio-distal vestibulo-lingual
'3 - dt~n~tre m~sio-distal
vestilmlo-lingual P4 - d/am~tre
m6sio-distal vestibuto.lingual
~41 - diamdtre m6sio-distal
vestibule-lingual
F e ~ of. minutn ' Moula Favigny Roquefure Saut-du-Loup G2 XV
17,97
4.06 3,55
5.1 2.62
6.8 3.11
7.31 3;21
3,8 2.7
22.98
6.15 3.42
8,06 3.85
9,27 4,27
6.73 3.48
8,3] 3,71
9.42 4.44
Fel is $il~¢stris
P ~ gaud Ofinchon 2
23,33 23.6
6,24 6.9 3.22 3.5
8,46 8,6 3.9 4.3
9,12 9 4.14 4.6
Actuel Ext#cmes Moyera~
n=23; 18,5-22,4 20,64
n=22; 5.3-6.2 5.73 n=22; 2,5-3.1 2.83
n=22; 6.2-8,2 7.1 n=22; 2,9-3.5 3.26
n=23; 7.4-9 8,26 n=23; 3,2-3.9 3.54
T a b l e a u X. Felis cl: minuta et Felis sylvestris. Dimentions du talus (en mm).
Table X. Felis cf. minuta and Felis sylvestris. Measurements of the talus (in mm).
F d i s c f i
Moula C.XV
Les Ckdres (Riss) Eattr~rnes
Hauteur
totale troehl~e int~me troel, l l ~ e x t e r m i
16,5 9.9 10 n=2 n=2 n =l
19,7-19,8 13,4-13.7 12,6
Co~-Diam~tre ~rticulation distale - Diam~tre transversal a'ansversal ant~ro-pcst~new
Jl
6,1 7,8 5,8 n=2 n=2 n=l
6.4-7,6 9,6-10 7,4
large, parois vestibulaires rectilignes). Le rap- port entre la longueur de la double boucle et la longueur totale de la dent fournit une valeur de 53,15 % ce qui de l 'ordre de grandeur de I'E. germanicus des niveaux moust6riens de I 'A- daouste (Jouques, Bouches-du-Rh6ne) : 48,35 et 56,39 % (Crdgut-Bonnoure, in : Defleur et al., 1994b). Le rapport flexidique (longueur du post- flexide par rapport ~ la longueur totale) est de 48,3 %, ce qui d6passe 16g6rement la valeur maximale de I'E. taubachensis du Bau de l 'Au- besier (Monieux, Vaucluse) : de 31,03 ~ 48 % pour 29 individus (Fernandez, 1996) et le cheval de l 'Adaouste : 39,19 et 47,54 % (Cr6gut-Bon- noure, in : Defleur et al., 1994). Le s6samoi'de correspond ~ un animal de forte stature. Sa hau- teur de 29,8 mm, sa largeur de 27,5 mm et son 6paisseur de 23,8 mm sont plus fortes que sur le sp6cimen rissien des C6dres : respectivement 30,
24 et 21,6 mm (Cr6gut-Bonnoure, in : Defleur et Cr6gut-Bonnoure, 1995).
- Les Artiodactyles.
- Sus scrofa, le sanglier. Le sanglier est repr6- sent6 par un lobe distal de M3 fragmentaire et l'articulation proximale incompl&e d 'un m6ta- tarsien III. La seule mesure possible conceme le diam6tre du mdtapode qui est de 21,1 mm.
- Cervus elaphus nov. ssp., le cerf 61aphe (nouvelle sous-esp6ce). Le cerf 61aphe est connu par un mat6riel relativement abondant parmi lequel se trouvent sept jugales inf6rieures com- pl6tes, six j ugales supdrieures et six restes osseux mesurables. La P2 inf6rieure (trois exemplaires) n 'a pas de paraconide comme cela s 'observe sur le spdcimen rissien de la grotte des C~dres. A la diff6rence de cet animal, l 'entoconide est net, surtout sur E4 XV 259, isol6 du m6taconide et de

382 A. Defleur et al.
l'entostylide. L'unique P3 inf6rieure est totale- ment us6e et donc inexploitable. La P4 inf6rieure (un seul sp6cimen) se caract6rise par un m6taco- nide r6uni au paraconide et distinct de l'entoco- nide. Ce morphotype est similaire fi celui qui est observable ~. la grotte des C6dres (figure 54 n ° 5 et 6, p. 63 Cr6gut-Bonnoure, in : Defleur et Cr6gut-Bonnoure, 1995). La base de la P4 du niveau sus-jacent ~voque le m~me morphotype (cfsupra). La fusion m6taconide-paraconide dis- tingue clairement l'individu de la couche XV de C. elaphus simplicidens du Wt~rm ancien de Combe-Grenal (Guadelli, 1987) tandis que l'ab- sence de fusion m6taconide-entoconide est fr6- quente dans cette sous-esp6ce. Aucune analogie n'existe entre la P4 de Moula et celles du cerf de la couche G e t G' de Saint-Marcel-d'Ard6che, comparables au morphotype de C. elaphus sim- plicidens et dont l'fige le plus probable est wfir- mien ancien ["6pisode temp6r6 froid.., du Wfir- mien ancien (cycle 3 de la courbe isotopique)" Debard, 1988] : la reconnaissance de C. elaphus simplicidens va dans ce sens. J'indiquerai au pas- sage que les P3 inf6rieures de la couche G e t G' de Saint-Marcel ont une morphologie proche mais distincte de celle de l'individu rissien des C~dres (figure 54 n ° 3 et 4, p. 63, Cr6gut- Bonnoure, in : Defleur et Cr6gut-Bonnoure, 1995) : le m6taconide, isol6, est plus 6cras6 ves- tibulo-lingualement, le fond de la vallde en arri6- re du paraconide prdsente une dilatation absente aux Cedres, l'entoconide ne rejoint pas l'entosty- lide. Cette morphologie est ~ nouveau celle que l'on observe chez C. elaphus simplicidens. Sur la M3 inf6rieure F4 XV 263 l'ectostylide est bien d6velopp6 (5,1 mm de hauteur), il l'est moins sur E2 XV 585 (4,4 mm) et il est totalement absent sur E2 XV 599 comme sur la M2 infdrieure D2 XV 502. Les molaires de la couche G et G' de Saint-Marcel sont totalement d6pourvues de cette colonnette interlobaire fi l'exception du sp6cimen SM F2 134+E1 20, ce qui est caract6ristique de C. elaphus simplicidens. Les P2 supdrieures se distinguent de l'individu de la grotte des Cedres par la profondeur de la vall6e bordant distalement le c6ne vestibulaire. Elles ressemblent en cela au spdcimen de la couche O de Saint-Marcel (SM E3 422 0) datde de l'Eowtirm (Debard, 1988). Les molaires supdrieures ont toutes un entostyle net. Le pli protoconal et le pli hypoconal sont visibles sur les sp6cimens D3 XV 664 et 669 qui correspondent fi un m~me animal. En revanche sur la MI H2 XV 70 aucun des deux n'est expri- m6. Les dimensions compardes des dents ainsi
que du post-cr~nien sont fournies dans les tableaux XI-XIV. L'6chantillonnage est malheu- reusement trop faible pour une analyse biom6- trique. On peut simplement indiquer que le cerf de la couche XV de Moula est plus grand que celui des C6dres et des couches U et O de Saint- Marcel mais plus petit que celui de la couche G et G' de ce dernier site et dont la morphom6trie est conforme ~ C. elaphus simplicidens.
C'est donc une sous-espbce de morphom6trie interm6diaire entre le cerf de la fin du Riss et celui du ddbut du Wfirmien ancien qui peut 6tre reconnue dans les couches XIV et XV de Moula. Sa diagnose fera l'objet d 'un article ult~rieur.
- Dama sp., le daim. La D2, la D3 et la D4 infdrieure droite, un scaphoi'de gauche et un semi-lunaire droit sont attribuables au daim. Les dents ddciduales, de taille bien inf6rieure fi celle de C. elaphus mais plus forte que celle de C. capreolus, pr6sentent la m6me morphologie que le daim de la couche U de Saint-Marcel dat6e de "l'EowOrm" (Debard, 1988). Les proportions sont toutefois 16gerement supdrieures (tableau XV1). Le scaphoTde a pu 6tre compar6 /l son homologue de ce m6me gisement. Leur morpho- logie et leurs proportions sont identiques (tableau XVI).
- Rangifer tarandus, le renne. Une troisiame prdmolaire infarieure sur un fragment mandibu- laire et une deuxi6me phalange, fi profil tres 6trangl6 au milieu de la diaphyse (type "en bou- chon de champagne"), permettent d'identifier le renne. Leurs dimensions sont fournies dans le tableau XVII.
- Capreolus capreolus, le chevreuil. Parmi la dizaine de testes correspondant au chevreuil, seul un canon postdrieur droit incomplet et une deuxi6me phalange sont analysables. L'articu- lation proximale du m6tapode est large de 20 mm pour un diamatre dorso-plantaire de 23,4 mm, la troisi~me phalange est longue de 28,5 mm pour une largeur totale de 8,5 mm.
- Bos primigenius, l'aurochs. Un semi-lunai- re droit et un grand sdsamoTde sont les seuls ossements complets reprdsentatifs d 'un Bovinae. Les cinq autres pi6ces osseuses ne peuvent 6tre analysdes. Le semi-lunaire pr6sente (figure 6) : en vue plantaire, un bord externe anguleux dans sa pattie distale; en vue distale, un bord externe /t angle plantaire bien saillant; en vue proximale, un bord axial de courbe assez r6guli~re. Les deux premiers caracteres morphologiques permettent de diagnostiquer le genre Bos, le troisieme 6tant
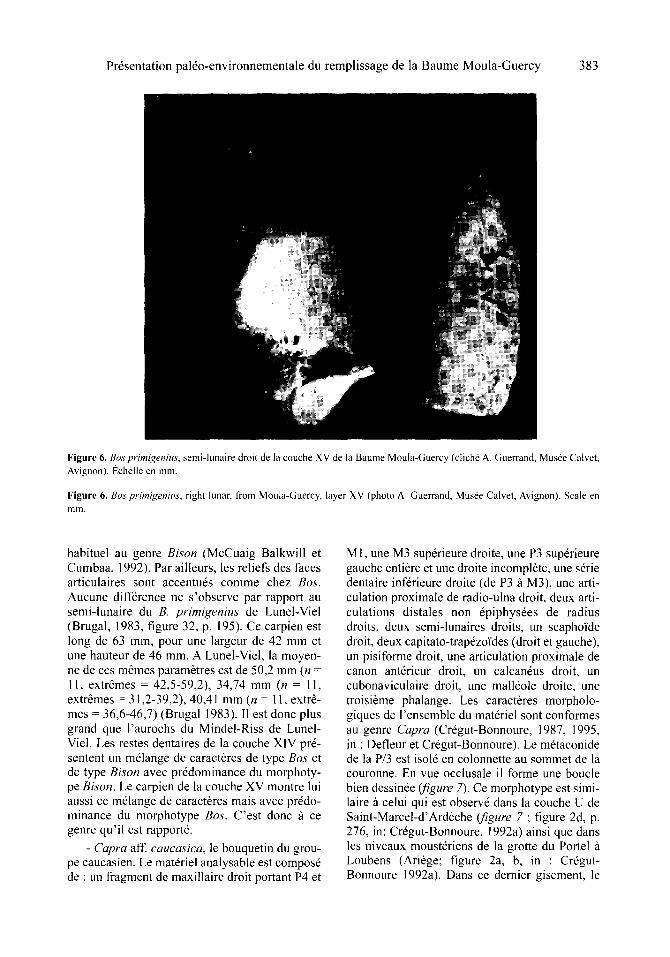
Pr6sentation pal6o-environnementale du remplissage de la Baume Moula-Guercy 383
Figure 6. Bos primigenius, semi-lunaire droit de la couche XV de la Baume Moula-Guercy (clich6 A. Guerrand, Mus6e Calvet, Avignon). l~chelle en mill.
Figure 6. Bos prim(~enius, right lunar, from Moula-Guercy, layer XV (photo A. Guerrand, Mus6e Calvet, Avignon). Scale en f i l m ,
habituel au genre Bison (McCuaig Balkwill et Cumbaa, 1992). Par ailleurs, les reliefs des faces articulaires sont accentu6s comme chez Bos. Aucune diff6rence ne s'observe par rapport au semi-lunaire du B. primigenius de Lunel-Viel (Brugal, 1983, figure 32, p. 195). Ce carpien est long de 63 ram, pour une largeur de 42 mm et une hauteur de 46 ram. A Lunel-Viel, la moyen- ne de ces m~mes param6tres est de 50,2 mm (n = 11, extremes = 42,5-59,2), 34,74 mm (n = 11, extr6mes = 31,2-39,2), 40,41 mm (n = 11, extr& rues = 36,6-46,7) (Brugal 1983). II est donc plus grand que l'aurochs du Mindel-Riss de Lunelo Viel. Les testes dentaires de la couche XIV pr6- sentent un m~lange de caractbres de type Bos et de type Bison avec pr6dominance du morphoty- pe Bison. Le carpien de la couche XV montre lui aussi ce m61ange de caract6res mais avec pr6do- minance du morphotype Bos. C'est donc fi ce genre qu'il est rapport&
- Capra aft: caucasica, le bouquetin du grou- pe caucasien. Le mat6riel analysable est compos6 de : un fragment de maxillaire droit portant P4 et
M l, une M3 sup6rieure droite, une P3 sup6rieure gauche enti6re et une droite incompl6te, une s6rie dentaire inf6rieure droite (de P3 fi M3), une arti- culation proximale de radio-ulna droit, deux arti- culations distales non 6piphys6es de radius droits, deux semi-lunaires droits, un scapho~'de droit, deux capitato-trap6zo'fdes (droit et gauche), un pisiforme droit, une articulation proximale de canon ant6rieur droit, un calcan6us droit, un cubonaviculaire droit, une mall6ole droite, une troisi6me phalange. Les caract6res morpholo- giques de l'ensemble du mat6riel sont conformes au genre Capra (Cr6gut-Bonnoure, 1987, 1995, in : Defleur et Cr6gut-Bonnoure). Le m6taconide de la P/3 est isol6 en colonnette au sommet de la couronne. En vue occlusale il forme une boucle bien dessin6e (figure 7). Ce morphotype est simi- laire ~ celui qui est observ6 dans la couche U de Saint-Marcel-d'Ard6che (figure 7 ; figure 2d, p. 276, in: Cr6gut-Bonnoure, 1992a) ainsi que dans les niveaux moust6riens de la grotte du Portel Loubens (Ari6ge; figure 2a, b, in : Cr6gut- Bonnoure 1992a). Dans ce dernier gisement, le

384 A. Defleur et al.
Tableau XI. Cervus elaphus. Dimensions des dents jugales inf6rieures (en mm).
Table XI. Cervus elaphus. Measurements of the lower cheek teeth (in mm).
Ceryus e~4~h~ Hauteur
m6sio-distal occlusal
Diam6tre vestibulo-lmgual
basal occlasal basal P2 in/drieure
Moula Couche XIV El YdV c 42 13,4 8,3 Couche XV E,4 XV 2.59 11.3 12,6 7,7 O1 XV 132 10,2 13,4 8,2
c. XV 13,4 8,3 n 2 3 3
Extremes 10,2-11.3 12.6-13,4 7,7-8,3 Moyenne 10,75 13,13 8,06
Saint-Marcel Couche G & G'
SM F2 95 G' 10 12,2 7.6 SMC3 170G 10,2 13,3 7,4
SP C2 G l 8,8 11,7 6,8 n 2 3 3
Extrdmes 10,2-11.3 11,7-13,3 6.8.7.6 Moyenne l 9,66 12,75 7,26
Les Cbdl'es 9,5 11,7 6,5 P3 infffrieure
:Moula Couclae XIV E3 XIVc 381 17,5 17,4 8,2 10,1
Couche XV E3 XV 601 16,6 16,6 10.9 10,9
Saint-Marcel Couche G & G'
SM 172.57 G 10,1 17,4 9,2 10,4 SM F2 95 G' 10,5 17,4 8,9 9,4 SM F2 508 G 7,4 17,6 9,9 11,6 SM (23 170 G 7,2 16.5 9,5 9,5 SM C23 72. G 16,5 17,1 7,7 8,6 SM C2 162 G 13 8,5 9,2
n 6 5 6 6 Extrbnes 7.2-16.5 16,5-17,6 7.7-9,9 9.2-11.6 Moyenne 10,78 17,2 9,04 9.78
Les C6dres (R/as) 13,6 16,2 14,9 8,9 9 P4 infdrieure
vloula Couche XI 21.4 21,3 13 13,9
Coueh¢ XIV D2 XIVd 306 17,1 17,9 10,9 11,4
I)3 X1V 307 16,7 16,7 11,9 11,9 Couche XV F2XV 315 14,2 19,7 18,9 11,4 11,5
e. XV 17,9 10,9 n 4 3 4 3
Extremes 16,7-19,7 16.7-18.9 10,9-11,9 11,5-11,9 Moyenne (cO(IV & XV) 17,85 17,83 11.27 11,6
Saint-Marcel Couche G & G'
SM F257G 8,6 20,1 11,4 12 SM F2 95 G' 11 9.20,5 11,3 11,4
SM F2 568 G 8,1 17,6 12,8 13,1 n 3 3 3 3
Extrdmes 8,1-11 17.6-20.5 11.3-12,8 11,4-12 Moyenne 9.23 20.3 11,83 12.16
Les C&tms (Riss) 15,9 17 10,9

Pr6sentation pal6o-environnementale du remplissage de la Baume Moula-Guercy 385
mat6riel dentaire est associ6 fi un crone de type caucasica. La M3/est assez us6e. Le m6tastyle est doric relativement large A la base mais demeu- re sub-rectiligne. Sa morphologie est similaire celle des troisi6mes molaires sup6rieures de Saint-Marcel-d'Ard6che mais aussi du Portel (figure 3a-d; figure 4 a-d, in : Cr6gut-Bonnoure, 1992a) et de l'Adaouste (Cr6gut-Bonnoure, in : Defleur et al., 1994b). Les proportions sont en revanche distinctes des individus du WOrm ancien. Les dimensions compar6es de l'ensemble du mat6riel se trouvent dans les tableaux XVIII et XIX.
- Rupicapra sp., le chamois. Le mat6riel est fragmentaire (n = 5). L'unique reste dentaire est une M2 inf6rieure gauche. Ses dimensions sont proches de celles des plus petits sp6cimens de la grotte des Gras fi Escragnolles (Var) d'fige wOr- mien r6cent (tableau XX). En l'absence de che- ville osseuse et de m6tapode, la position sp6ci- fique de ces pi6ces ne peut 6tre pr6cis6e.
Conclusion
Les premi6res analyses ont d6bouch6 sur des r6sultats significatifs en terme de biochronolo- gie. Deux ensembles fauniques sont, en effet, distinguables : celui des couches IV, V, VII1, IX, X et XI, de caract~re frais ~ froid, off se reneon- trent un loup dont le stade 6volutif est proche de celui qui s'observe au Pl6istoc6ne sup6rieur moyen, un ours et une hy6ne des cavernes typiques du WOrm et un cerf assez semblable au Cervus elaphus sub. sp. du Wtirm ancien sup6- rieur du Sud-Ouest de la France. Ces donn6es confirment celles qui sont fournies par l'analyse de la microfaune et de l'industrie et qui placent cet ensemble dans le Wtirm ancien; celui des couches XII, XIII et X1V, de caract6re nettement temp6r6. Le stade 6volutif du loup est interm6- diaire entre celui de la fin du Pl6istoc6ne moyen et celui du W~irm ancien. L'ours de la lign6e deningeri-spelaeus est d 'un type trbs primitif of a dominent les caract6res d'U. deningeri. Darts la couche XIII, on retiendra la pr6sence de l'ours du Tibet dont l'association avec l'ours brun, le blaireau, la panth6re et le chat sauvage est iden- tique fi celle de Blanot 2 d'fige l~6mien. L'ensemble peut en cons6quence ~tre dat6 de l'l~6mien, ce qui est fi nouveau confirm6 par la microfaune.
La faune de la couche XV qui fait suite l'ensemble pr6c6dent, ne montre pas de diff6ren- ce majeure. Les points remarquables sont les sui- rants : l'association des Carnivores est tr6s pro-
Figure 7. Capra aft. caucasica. De gauche fi droite : P3 inf6- rieure droite de la grotte de Saint-Marcel-d'Ard~che et P3 inf6rieure gauche de la couche XV de la Baume Moula- Guercy (clich6 A. Guerrand, Mus6e Calvet, Avignon). l~chel- le en mm.
Figure 7. Capra aft. caucasica, from left to right: lower right P3 from Saint-Marcel-d'Ard~che (level U) and lower left P3 from Moula-Guercy (photo A. Guerrand, Mus6e Calvet, Avignon). Scale en ram.
che de celle de Blanot 2 du fair de la pr6sence d'un Felis proche de Felis minuta et d'U. thibe- tanus ; pour la premi6re fois au sein du remplis- sage de l'abri Moula appara~t le daim. A l'ouest- du Rh6ne, l'esp6ce n'est connue que dans la cou- che U de Saint-Marcel-d'Ard6che dat6e de "l 'Eowtirm" par E. Debard (1988). A peu pros la m6me p6riode on la trouve au Bau de l'Aubesier (Cr6gut-Bonnoure et al., 1994). Par rapport ~ Saint-Marcel-d'Ard6che, l'esp6ce est rare/l Moula; le bouquetin est identique ~ celui de Saint-Marcel-d'Ard6che, de l 'Adaouste et du Portel. I1 correspond donc fi un bouquetin de type caucasien. D6s lors !'immigration de cette esp6- ce prend place h l'E6mien : elle est doric plus ancienne que pr6vue (Cr6gut-Bonnoure 1992a- d) ; le cerf 61aphe, proche des individus des cou- ches XllI et XIV, est diff6rent morphom6trique- ment de celui de la grotte des C6dres, de celui de la couche U de Saint-Marcel et de C. elaphus simplicidens. II s'agit d 'une nouvelle forme.

386 A. Defleur et al.
Tableau XII. Cervus elaphus. Dimensions des dents jugales inf6rieures (en mm).
Table XI1. Cervus elaphus. Measurements of the lower cheek teeth (in ram).
Cervus elap~us
M1-2 infdrieure Moula
Couche XlV! I)3 XIVb 30,1 E3 XIVb 158
Couche XV 1)2 XV 502
n
Extrdmes Moyenne C. XIV+ XV
Saint-Marcel Couehe G
M I : S M F 2 5 7 G M2 : SM F257G
n
Extr0mes Moyenne Gauche K
M2: SP F2 474 K M3 inf~rieure
Couche XV E2 XV 599 F4XV 263
c. XV n
Extremes
Hauteur
22,1
7,9 10,7 2
7,9-10,7 9.3
17,6
15,4 12,5 9,2 3
m~sio-distal oeclusal
23,4 26,3
29,7 3
23,4-29,7 24,46
22,4 26,5 2
22.4-26,5 24,45
25,6
30,9 30,7 32,5
3
basal
21,3 24,2
25.5 3
21,3-25,5 23,66
23,9
30,9 30
32,5 3
~)estibulo-lingual i
occlusal
11,1 12,3
13,6 3
11,1-13,6 12,33
12,9 14,8 2
12,9-14,8 13.85
13,2
9.2-15,4 30, 7-32.5 30-32.5
13,6 14,6 15.2 3
14,6-15,2 12,36 31,36 Moyenne
Saint-Marcel Couche G
SME2 124G 8,1 35,3 SM E1 20 G 22 33,3
n 2 2 ExtrOmes 8,1-22 33,3-35,3 Moyenne 15,05 34,3 Couche P'
SM E3 T P' 14,9 33,7
31,13
35,3 31,1 2
31.1-35,3 33,2
33,4
14.46
15,7 12,8 2
12,8-15,7 14,25
14,3
basal
13,3 15,4
15.5 3
13,3-15,5 14,73
14,1 15,2 2
14,1-15,2 14,65
16
14,2 15,5 15,2 3
14,2-15,5 14.96
15,7 14,7 2
14,7-15,7 15,2
14,1
Les donn6es disponibles permettent de pro- poser une nouvelle datation des couches U et O de Saint-Marcel-d'Ard6che et de positionner plus prdcis6ment les couches XIII fi XV de Moula : la couche U de Saint-Marcel, incluse fi la base de la sdquence paldoclimatique 3 de Saint-Marcel (Debard, 1988), par la pr6sence de H. cedrensis (Cr6gut-Bonnoure, 1989 et Cr6gut-
Bonnoure, in • Defleur et Cr6gut-Bonnoure, 1995) et d 'un cerf morphom&riquement diffd- rent de C. elaphus simplicidens et semble-t-il plus proche de celui des C6dres pourrait 6tre d 'gge rissien. Toutefois, l 'abondance du daim ainsi que la position stratigraphique de la couche U s 'oppose g cette hypoth6se. La composition faunique n 'a aucune analogie avec celle de la

Pr6sentation pal6o-environnementale du remplissage de la Baume Moula-Guercy
Tableau XIII. Cervus elaphus. Dimensions des dents jugales sup6rieures (en ram).
Table Xlll. Cervus elaphus. Measurements of the upper cheek teeth (in mm).
387
Cervus elaphus
P2 supgrieure Moula
Saint-Marcel
Couche IV E6 IV 14
Couehe X Couche XV E4XV 258
Hauteur n~sio-distal
occlusal
Couehe O SM E3 422 O
Les C6dres (Riss) P3 sup~rieure
Moula Couche IV
E1 IV 19 Cotw.he XV]
D1 XV 65 E4 XV 237 E4 XV 237
Saint-Marcel Couche O
SM E3 422 O Couche U
SM C2 454 U I.x,s C&lres (Riss)
15,3
12,8
18,6
10 13,5 13,5
18,1
19,4
15,6 18,4
16,6
18,1 15,7
15
16,3 16,8 16,8
16,7
14,8 14,9
Diam~tre
basal
15,3
16,1
16,8
14,9
16 16,3 16,3
13,8
vestibulo-liagu~ ' occlusal
i i
11,1
15,2
15,1 15,5
i i
18,5
17,3
16,3
18,4 17,2
basal
16
16,1
16,4
21,4
17,4
17,9
partie sup6rieure du remplissage de Saint- Marcel, ni avec celle de la grotte de l 'Adaouste situ6e clairement au d6but du Wtirm (Defleur et al., 1994b) et ni avec celle des niveaux de Moula coiffants les couches 66miennes. On peut donc difficilement placer la couche U au tout d6but du Wfirm comme l'envisageait Debard (1988) : elle parait correspondre fi l'E6mien. L'absence du tahr darts les niveaux 66miens de Moula, encore pr6sent fi Saint-Marcel (H. cedrensis ; Cr6gut- Bonnoure, in : Defleur et Cr6gut-Bonnoure, 1995), la raret6 du daim et les caract6res morphologiques du cerf sugg6rent que la couche U de Saint-Marcel est plus ancienne ; les cou- ches Ge t G' de Saint-Marcel se placeraient dans cette hypoth~se au dessus de l'ensemble XII ~. XV de Moula. Pour l'instant leur correspon- dance avec les couches IV ~ XI de Moula ne peut 6tre pr6cis6e. D'apr+s les analyses s6dimentolo-
giques, les couches G et G' sont plac6es dans la phase pal6oclimatique de Saint-Marcel 7 sem- blant correspondre fi la fin du Wfirm ancien (fin du stade isotopique 5 ; Debard, 1988). L'asso- ciation faunique est clairement temp6r6e (Cr6- gut-Bonnoure, in : Debard, 1988) et en cons6- quence diff6rente de celle de Moula.
II est important de signaler la pr6sence du renne dans tout le remplissage, m6me lorsque le reste de la grande faune est de type temp6r6. Un ph6nom6ne similaire a d6jfi. 6t6 observ6 dans les sites de la grotte des C6dres (Cr6gut-Bonnoure, in : Defleur et Cr6gut-Bonnoure, 1995) et de l 'Adaouste (Cr6gut-Bonnoure, in : Defleur et al., 1994b). D6s lors il est difficile d'envisager un biotope similaire fi celui qui existe aujourd'hui darts les zones temp6r6es. I1 convient de privil6- gier le maintien de zones de savane froide en relation avec une certaine instabilit~ du climat.

388 A. Defleur et al.
Tableau XIV. Cervus elaphus. Dimensions des dents jugales sup6rieures (en mm).
Tableau XIV. Cervus elaphus. Measurements of the upper cheek teeth (in mm).
Cer~ elaphus Hauteur
r~sio-distal occlusal basal
Dian~re vesttbulo-lmgual 1 ° lobe
occlusal basal vestibulo-lingual 2 ° lobe
occlusa/ basal M1- 2 sut~rieure
Moula Couche IV
F5IV 17 (MI 22,7 22,7 22,2 [:5 IV 17 (M2) 26,9 25,5 Z3,4
Couche XIV F_.2 XIV 390 21,5 21,5 23,2 23,2 22,4 22,4 F3 XIV 139 2A..1 21,8 18,1 22,5 Couche XV
1-12 XV 70 15 23,4 22 15,9 22,3 n 3 3 3 2
E.xtr&~qes 21.5-24,1 21,5-22 15,9-23,2 22,3-23.2 Moyenne CXV & XXIV 23 21,76 19,06 22,75
M3 supffrieure Moula
Couche XIV FI XIV 13,~ 25,8 24,9 21,3 25.5 18,1 23 F_.2 XIV 237[ 26,1 23,4 19,2 26 16,9 25,4 I)2 XIV 154 25,6 24,4 20,3 24,7 16,1 21,8 E4 XIV 127 23,1 22,4 16 243 133 21,8 D4 XIV 24 26,2 23,6 18.6 23,2 15,7
132 XIV 28"7 23,7 22.7 19.1 24,3 15,3 21,1 F3 XIV 173 24,3 23,6 19,4 25,5 D2 XIV 341 25,3 22,8 17,8 22,8 15,5 22,3
n 8 8 8 8 7 6 Extr~nes 23.1-26.2 22,4-24.9 16-21,3 22,8-26 13,3-18.1 21,2-25,4 Moyenne 25,01 23,47 18.96 24,53 15,84 22.56
Couche XV I)3 XV 669 15,7 24,5 23,2 17,7 23,2 16,5 22,6
Couche XIV ~ XV n=9 n=9 n=9 n=9 n=9 n=9 ExtrP.mes 23,1-26,2 22,4-24,9 16-21.3 22,8-26 13,3-18,1 21.2-25.4
Moyenne CXV 1 CXIV 24.95 23,44 18.82 24.38 15,92 22,57 Saint-Marcel
Couche G SM C2 82 G 14,5 26,8 25,2 19,4 22,7 15.5 20,5
3. L e s r o n g e u r s
3.1. In troduc t ion
Durant les fouilles rdcentes, le recueil des 616ments anatomiques des micromammiferes, o~ les rongeurs sont particuli6rement bien reprdsen- t6s, a fait l'objet d'une collecte exhaustive apr6s tamisage g l'eau sur un tamis fi mailles de 0,5 mm et tri sous loupe binoculaire. La documenta- tion demeure cependant hdt6rogene dtant donn6 que certaines couches, du fait des anciennes fouilles, n'ont pu etre fouilldes que sur une sur- face faible alors qu'elle est plus abondante pour les niveaux connus sur une surface plus dtendue. Si l'on se rdfere g I'ensemble de la s6quence, ce jour, plus de 25 000 616ments anatomiques ont
6t6 d6termin6s. Cinquante-deux genres et esp6- ces se rapportant ~ plus de 4 000 individus (d'a- pr6s le N.M.I.) ont fait l'objet d'une d&ermina- tion sp6cifique. La r6partition par couches est donn6e darts le tableau XXI (figures 8-10).
Couche IV : Apodemus sylvaticus, Eliomys quercinus, Arvicola sapidus, Terricola duodeci- micostatus, Clethrionomys glareolus, Microtus arvalis-agrestis, Chionomys nivalis, Dicrostonyx torquatus, M. oeconomus, M. gregalis, Sciurus vulgaris.
Couche V-VII : Eliomys quercinus, A. sapi- dus, C. glareolus, M. arvalis-agrestis, C. nivalis, M. oeconomus, M. gregalis.
Couche VIII : Marmota marmota, A. sylvati- cus, Glis glis, A. sapidus, C. glareolus, M. arva-

Pr6sentation pal6o-environnementale du remplissage de la Baume Moula-Guercy
Tableau XV. Cervus elaphus. Dimensions des carpiens et des phalanges (en ram).
Table XV. Cervus elaphus. Measurements of carpals and phalanxes (in ram).
389
Cervus elaphu$
SemLlunaire Vloula
E3 XV 579 D2 XV
~.daouste (WUrm ancien ScaphoFde
Moula E3 XV 5 ~
Adaouste (WUrm ancien
Hauteur Diam~ti'~ transversal
22.2 21.4 25,6 25.4 21.4
29.1 20,1 30,9 21.2
Longueur dorsale axiale
Premidre phalanse Moula
H2 XV 69 57,9 57,1 F4XV 285 FI XV 428 57,1 58,3
~.daouste (Wllrm ancien) l* phalanges ant&ieures a=3 n---3
F_xtr~mes 51,5-54,1 20.8-54,3 Moyenne 52.96 52,73
° phalanges post~rieures n--.4 a=4 F_xtr~tnes 47,3~55,3 47,l-55,9 Moyenne 51,67 51,77
mt~ro-post~ear
31,5
36
39,9 36,8
abaxiale Ext~'nut~ lm~ximale Diam~tre
mt£~-post~eur transversal
6O,5 28.8 27,8
60,5
n---3 n--3 51.9-55.3 24,3-26,8
54,1 25,83 n=4 n=4
48,4-5Z6 23,3-28,9 53,45 25,67
22.9 21,3 20,2
n--3 20,5-21,9
21,5 n=4
19,6-22.1 20,47
Mtlieu diaphyse Dianr~tte
imt~.,l~post~elll transversal
22,2 20.t
19,5 17,5
n=3 n-----~ 17,6-20,1 16,5-18,3
19,06 17,5 n=4 n=4
18,5-22,2 16.7-17,7 ' 20,07 17,32
E~h~,;i~ distale
I~ara~re
a,m&~-post~rJeu' transversal
18,7 22,3
19,1 19,9
n=3 n=3 17,6.19 20,2-21,6 18,36 21 n=4 u=4
16.1-19.1 19,4-21,4 17,32 20,1
lis-agrestis, C. nivalis, D. torquatus, M. oecono- mus, M. gregalis.
Couche IX : Citellus sp., A. sylvaticus, E. quercinus, G. glis, Muscardinus avellanarius, A. sapidus, C. glareolus, M. arvalis-agrestis, C. nivalis, D. torquatus, M. oeconomus, M. gregalis.
Couche X-XI : A. ~ylvaticus, E. quercinus, A. sapidus, T. duodecimcostatus, C. glareolus, M. arvalis-agrestis, M. oeconomus, M. gregalis.
Couche XII : S. vulgaris, A. sylvaticus, G glis, A. sapidus, Terricola subterraneus, T. mul- tiplex, T. pyrena't'cus, C. glareolus, M. arvalis- agrestis, C. nivalis, M. oeconomus.
Couche XIII : S. vulgaris, A. sylvaticus, E. quercinus, G glis, Muscardinus avellanarius, A. sapidus, T. subterraneus, T. multiplex, T. pyre- naicus, T. duodecimcostatus, C. glareolus, M. arvalis-agrestis, C. nivalis, M. oeconomus.
Couche XIV: Citellus sp, S. vulgaris, Allo- cricetus bursae, Cricetulus migratorius, A. sylva- ticus, A. microps, E. quercinus, G glis, M. avella- narius, A. sapidus, T. subterraneus, T. multiplex, T. pyrenaicus, T. duodecimcostatus, C. glareolus, M. arvalis-agrestis, C. nivalis, M. oeconomus, M. gregalis, M. brecciensis.
Couche XV : A. bursae, A. sylvaticus, E. quercinus, G glis, M. avellanarius, A. terrestris, A. sapidus, C. glareolus, M. arvalis-agrestis, C.
nivalis, M. oeconomus., M. gregalis, M. brec- ciensis, Pliomys lenki.
Couche XVI: A. bursae, C. migratorius, A. svlvaticus, S. vulgaris, Marmota marmota, C. nivalis, Cricetus major, Sicista cf. betulina, E. quercinus, M. avellanarius, M. brecciensis, T. subterraneus, T. multiplex, T. duodecimcostatus, G glis, Arvicola sp., C. glareolus, M. arvalis- agrestis, M. oeconomus., M. gregalis, P. lenki.
Couche XVII : C. superciliosus, S. vulgaris, M. marmota, A. bursae, C. migratorius, C. major, S. betulina, A. sylvaticus, A. microps, E. querci- nus, G. glis, M. avellanarius, Arvicola sp., T. sub- terraneus, T. multiplex, T. duodecimcostatus, C. glareolus, M. arvalis-agrestis, C. nivalis, D. tor- quatus, M. oeconomus, M. gregalis, M. breccien- sis, P. lenki.
Couche XVII1 : C. superciliosus, A. bursae, C. major, A. sylvaticus, E. quercinus, G. glis, Arvicola groupe cantiana/terrestris, T. pyrenai- cus, T. duodecimcostatus, C. glareolus, M. arva- lis-agrestis, M. oeconomus, M. gregalis, M. brecciensis, P. lenki.
Couche XIX : C. superciliosus, A. bursae, S. betulina, A. sylvaticus, E. quercinus, G. glis, Arvicola groupe cantiana/sapidus, T. pyrenai- cus, T. duodecimcostatus, C. glareolus, M. arva- lis-agrestis, Dicrostonyx torquatus, M. oecono- mus, M. gregalis, M. brecciensis, P. lenki.

390 A. Defleur et al.
IO
F
C
0 !
J
Figure 8. A. Chionomvs nivalis, M/I gauche, couche XV. B. C. nivalis. M/1 droite, couche XV. C. (~ nivalis, M/I droite, cou- che XVI. D. (5'. nivalis, M/I gauche, couche XVI. E. C. nivalis, M/1 droite, couche XVI. F. U. nivalis, M/I gauche, couche XV. G. Microtus oeconomus, M/I droite, couche XV. H. M. oeconomus, M/I droite, couche XV. 1. M. oeconomus, M/I gauche, cou- che XVI. J. M. oeconomus. M/1 droite, couche XVI. K. M. oeconomus, M/1 gauche, couche XVI. L. M. oeconomus, M/I Gauche, couche XV. ([~chelle - I mm).

Prdsentation pal6o-environnementale du remplissage de la Baume Moula-Guercy 391
I
K
Figure 9. A. Pliomys lenki, M/I droite, couche XV. B. P lenki, M/I gauche, couche XV. C. P lenki, M/I droite, couche XV1. D. Idem, vue latdrale. E. P lenki, M/I droite, couche XVI. F. P lenki, M/1 gauche, couche XV. G. ldem, vue lat6rale exteme. H. Microtus brecciensis, M/1 gauche, couche XV. I. M. brecciensis, M/1 droite, couche XVI. ,1. M. brecciensis, M/1 gauche, couche XVI. K. M. brecciensis, M/I gauche, couche XVI. (l~chelle - 1 mm).

392 A. Defleur et al.
B .3
F_, F
G Ii 11
Figure 10. A. Arvicola terrestris, M/1 gauche, couche XV. B. A. terrestris, M/I gauche, couche XV. C. A. sapidus, M/1 gauche, couche XV. D. A. sapidus, M/1 droite, eouche XV1. E. A. terrestris, M/1 droite, eouche XVI. F. A. terrestris, M/1 droite, cou- che XV|. G. Microtus gregalis, M/1 droite, couche XV. H. M. gregalis, M/I droite, couche XV. 1./1~ gregalis, M/I droite, cou- che XV. J. M. gregalis, M/1 droite, eouche XVI. K. Mgregalis, M/I droite, couche XVI. L. A~ gregalis, M/| droite, couche XV1. (l~chelle = 1 mm).

Prdsentation paldo-environnementale du remplissage de la Baume Moula-Guercy
Tableau XVI. Dama sp. Dimensions des dents jugales infdrieures et du scaphoi"de (en ram).
Table XVI. Dama sp. Measurements of the lower cheek teeth and of the scaphoid (in mm).
393
Dama sp.
D2 infgrieure Moula
DI XV 161 D3 infgrieure
Moula D1 XV 161
Saint-Marcel Couche UI C2631 U
D4 infgrieure Moula
DI XV 161 Saint-Marcel
Couche U C2631 U
Scaphoi'de Moula
G5XV 60 Saint-Marcel
Couche U
Hauteur
6,5
aa~sio-distal occlusai
9,5
7,5 14,4
8 13,9
11 23,1
20,7 Diarn~tre
transversal
Diam~tre
basal
8.3 Hauteur
18,7 25,4
13,1
13,1
21,5
20,7
~nt~ro-post~rieu~
12,5
vestibulo-lingual occlusal
6,7
5,5
8
8,5
Tableau XVII. Rangifer tarandus. Dimensions de la P3 inf6rieure et de la deuxi6me phalange (en mm).
Table XVII. Rangifer tarandus. Measurements of the lower P3 and of the second phalanx (in mm).
basal
5,5
7,1
6,7
9,5
8,8
P3 infgrieure Moula DI XV 179
Deuxi~me phalange FI XV 376+433
Dim~tre Hauteur m~s~o-di~al vlnltiblalo-lingwfl
occlusal basal oeclusal basal
10,8 12,9 11,1 7,7 8,3 L~lgl~ur ~Li~rm~ ~ l m s t e
dorsale axiale il~xiale Diam~tt~ aJat&o-post&iet~ tr~l~erslt
31 32.3 32,4 20.2 18,7
Milieu diaphyse Ext r~raitt[ distsle ~ ' Diam~tre Dian~te
mat~ro-po~riem tmnsvemd antCxo-po~rie~t transversal
13,5 12.7 16,4 16,7
Les premiers rdsultats concernant la syst6- matique, la biostratigraphie et la pal6o-6cologie des micromammif6res sont pr6sentds dans ce chapitre.
3.2. Syst~matique et biostratigraphie
Le degr6 d'6volution de certaines esp6ces identifi6es fi la Baume Moula-Guercy permet de pr6ciser le cadre chronologique de la s6quence.
Les rongeurs sont pr6sents dans t o u s l e s niveaux connus de la Baume Moula-Guercy. lls sont particuli6rement abondants dans les niveaux de base. Les esp6ces du sous-genre Terricola, 6tudi6es par Brunet-Lecomte (Brunet-Lecomte et al., 1995) correspondent/t des formes actuelles. Elles pr6sentent cependant une remarquable diversit6 qui a pu &re mise en ~vidence grfice une 6tude morphom6trique approfondie. En effet, l 'examen visuel (fi la loupe binoculaire) de la

394 A. D e f l e u r et al.
~ E
o g E
E
~ ~. ~. ~ ~ . ~ ,-2
I i i i i i
i i i
~. . i ~ . ~ ~
~ ~ > r, e ~ - ~ o -

Presentation paleo-environnementale du remplissage de la Baume Moula-Guercy 395
morphologie des M/I a montre que celles-ci pre- sentaient une morphologie "moderne" compara- ble/~ celle des especes actuellement presentes en France. C'est pourquoi la determination des Ter- ricola de la Baume Moula-Guercy a ete faite par classification obtenue/l partir d'une analyse dis- criminante effectuee entre les especes presentes dans la region Rhene-Alpes-Auvergne : Microtus (Terricola) subterraneus, M. (Terricola) multi- plex, M. (Terricola) pyrenaicus et M. (Terricola) duodecimcostatus, ~ partir de 21 mesures prises sur la surface occlusale de la M/l , selon la methode definie par Brunet-Lecomte (1988). La repartition des Terricola identifies dans la sequence est indiquee dans le tableau XXI.
Les populations de M. (Terricola) duodecim- costatus presentent une morphologie de M/1 moderne, semblable "2. celle des populations actuelles : longueur relative de la partie anterieu- re de la dent peu developpee, rhombe pytimien incline et boucle anterieure ouverte.
Globalement, la morphologie de la M/1 des populations de M. (Terricola) pyrenaicus est de type moderne avec un rhombe pytimien incline. La longueur relative de la pattie anterieure tend/t ~3tre moins ddveloppee que celle de l 'espece fos- sile du Pleistocene moyen du gisement d'Orgnac 3 en Ardbche, attribuee/t M. (Terricola) maria- claudiae par Brunet-Lecomte et Chaline (1991). On notera cependant que la boucle anterieure est moins ouverte que celle des populations actuelles de M. (Terricola) pyrenaicus.
Le rhombe pytimien de la population de M. (Terricola) multiplex presente une inclinaison intermediaire entre celle de la sous-espece des Alpes internes druentis et celle de la sous-espe- ce du Vercors niethammeri (Brunet-Lecomte et Volobouev, 1994). Enfin, M. (Terricola) subter- raneus presente une morphologie comparable b, l 'espece actuelle.
L'analyse des campagnols souterrains de la Baume Moula-Guercy presente un inter& d'ord- re paldobiogdographique. En effet, si les quatre especes reconnues dans le remplissage sont pre- sentes actuellement dans le Sud de la France, seuls M. (Terricola) duodecimcostatus et M. (Terricola) subterraneus se rencontrent encore dans les environs immediats de Soyons et sem- blent presents en continu dans cette zone depuis la fin du Pleistocene moyen. M. (Terricola) pyre- naicus parait avoir regress6 /t l 'ouest dans le Massif central et M. (Terricola) multiplex, qui est de nos jours absent en rive droite du Rhene, a une aire de repartition ldgbrement plus orientale.
I1 est 6galement interessant de prendre en consideration le degre d'evolution des autres ron- geurs presentant un inter& biostratigraphique.
La forme moderne Eliomys quercinus querci- nus est abondante dans les differents niveaux etu- dies. Une etude detaillee de l'indice d'epaisseur d'email, selon le principe defini par Heinrich (1978 et 1990) et de la morphologie des M/1 des Arvicola de la Baume Moula-Guercy (Abbassi et Desclaux, 1996 ; Abbassi et al., 1998 ; Desclaux et al., 2000) a permis d'identifier deux formes fossiles dans les couches XIX et XVII1 (Arvicola groupe cantiana/terrestris et Arvicola groupe cantiana/sapidus), deux especes dans la couche XV (A. terrestris et A. sapidus) et une seule espe- ce dans les niveaux superieurs (Arvicola cf. sap# dus, couches XIV ~ IV).
Les M/1 de petite taille, caracterisees par des triangles T4 et T5 opposes, une boucle anterieu- re dissymetrique et un col plus ou moins large (correspond au morphotype praeceptor defini initialement par Hinton, 1926) ont ete attribuees au morphotype terrestris.
Les M/1 de plus grande taille, presentant une boucle anterieure de forme triangulaire et syme- trique /l col generalement etroit (morphotype grenii selon Hinton, op. cit.) ont ete rapportees au morphotype sapidus.
On notera que, d'une faqon generale, la bou- cle anterieure presente une morphologie variable avec t o u s l e s intermediaires entre les formes decrites ci-dessus.
Les Arvicola (N - 31, SDQ = 87,41) des niveaux XIX et XVIII presentent, independam- ment de la forme identifiee, un indice d'epais- seur d'email comparable /t celui que van Kol- fschoten (1990) a observe dans le Nord de l 'Eu- rope durant le stade isotopique 6 (Saalien).
La valeur du SDQ des A. sapidus et A. ter- restris (N = 32, SDQ = 100,47) des couches medianes (XV/t XIII) indique que ces niveaux datent du debut du Pleistocene superieur (stade isotopique 5).
N.B. : On notera qu'A. sapidus domine lar- gement dans ces niveaux. Seuls, quelques indivi- dus recueillis dans les couches XV /t XVI1 ont et6 attribues ~ A. terrestris.
Enfin, les indices des Arvicola cf. sapidus (N - 5, SDQ = 84,25) observes dans les couches superieures (VIII /l IV) suggerent un ~.ge Pleistocene superieur plus recent.
Une telle hypothese biochronologique a pu. 6tre 6mise grace/t comparaison des Arvicola de

396 A. Defleur et al.
Age
10 000
50 000
100 000
130 000
S tade isotopique
Stratigraphie
H o l o c ~ n e
SDQ M 1
hO 70 80 90 100 110 120 130 140
Oigny Vl
G~gny X
3 ~ Moula Vlll-V ,=.. (~ ('~ Eglise m ::3
(~c su.rn
5 8 ~,~, xx-x,xc • ~ e ~ , c c 8
~(~ MInie
¢-- EL
E Moula XV.Xh/
LU
MOulII XVlII-XIX
Gigny XXII ¢,..
6 _ .1~ ~ Vaufrey VIii ~0 "(l)
~ Lazaret
• i"l
l 1 --r-t-q
I ,
__1 m
[ 1- } - -
£ t - t - -
- - 1 1 1
- - t 1- }
i I ~-
c t 1 - - -cb
- ~ m t
r 1 1
r 1 1 - -
Figure 11. Variation de l'indice d'6paisseur d'dmail (SDQ) des Arvicola de France et de Ligurie durant le Pldistoc6ne moyen et supdrieur.
Figure 11. Arvicola M/1 enamel quotient variation (SDQM1) in France and Liguria during Middle and Upper Pleistocene.
la Baume Moula-Guercy avec diff6rentes popula- tions du P16istoc6ne moyen et supdrieur du Sud de la France (figure 11). En effet, les Arvicola des couches XIX et XVIII de la Baume Moula- Guercy et de la couche XXII de la Baume Gigny prdsentent des indices inf6rieurs 5. 100, compara- bles 5. ceux que van Kolfschoten (1990) a obtenu pour les populations de la fin du Saalien (stade isotopique 6) en Hollande. On peut donc envisa- get que ces formes (ddnommdes Arvicola terres- tris ssp. B par cet auteur) ont eu une expansion importante en Europe occidentale lors d'une p6riode froide de la fin du P16istocbne moyen, pouvant correspondre 5. un maximum glaciaire. Les indices obtenus darts certains niveaux de Gi- gny (couche XXII) et de la Baume Moula-Guer- cy (couches XIX et XVII1) sugg6rent que le Jura et la r6gion Rh6ne-Alpes ont 6t6 affectds par cette migration.
N.B. : Les indices sup6rieurs 5' 100 obtenus 5' Vaufrey (Charente) et au Lazaret (Alpes-Mari- times) dans des niveaux suppos6s synchrones tendent 5, montrer que le domaine oc6anique et I'extr~me sud-est de la France n'ont pas 6t6 affect6s par cette migration.
Durant le Pl6istoc6ne sup6rieur, les popula- tions d'Arvicola semblent ~tre caract6ris6es par une diminution progressive de la valeur de l'in- dice d'6paisseur d'6mail, ll est par cons6quent possible de replacer chronologiquement ces populations les unes par rapport aux autres en se r&@ant 5. la valeur de I'indice d'@aisseur d'6- mail et aux donn6es pal6oclimatiques.
C'est ainsi que les valeurs obtenues pour les couches XV 5. Xlll de la Baume Moula-Guercy, proches de 100 et associ6es 5' une faune globale- ment temp6r6e, correspondent vraisemblable- merit 5. un 6pisode temp6r~ datant du d6but du
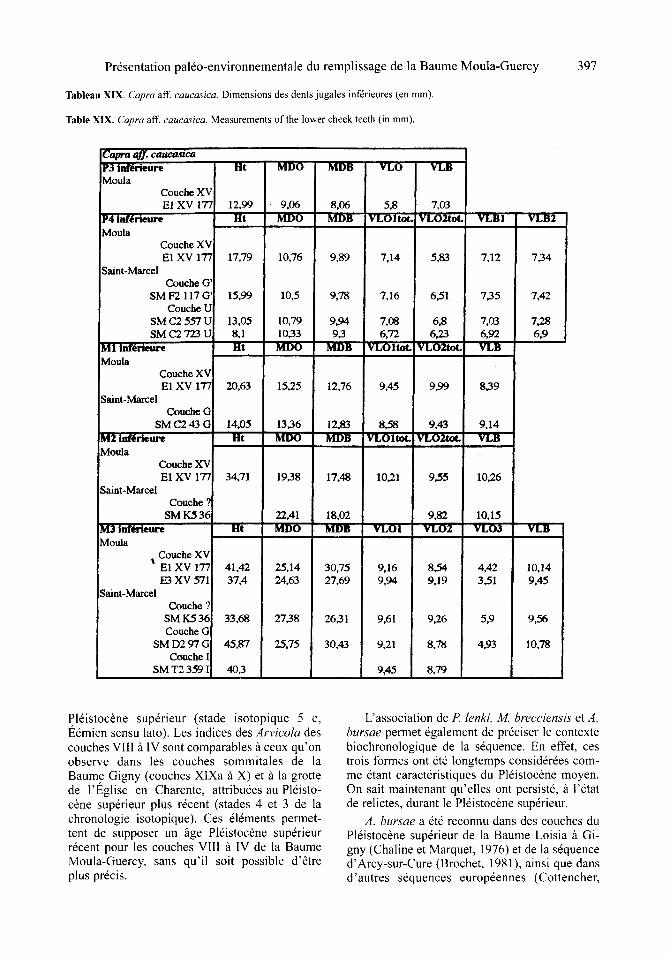
Pr6sentation pal6o-environnementale du remplissage de la Baume Moula-Guercy
Tableau XIX. Capra aft. caucasica. Dimensions des dents jugales inf6rieures (en mm).
Table XIX. Capra aft. caucasica. Measurements of the lower cheek teeth (in mm).
Capra aft. caucasica P3 lnfarieure Ht MDO MDB Moula
Couche XV E1 XV 177 12,99 i 9,06 8,06
?4 inf~rieure Ht MDO MDB vloula
Couche XV E1 XV 177 17,79 10,76 9,89
Saint-Marcel Couche G'
SM F2 117 G' 15,99 10,5 9,78 Couche U
SM C2 557 U 13,05 10,79 9,94 SM C2 723 U 8,1 10,33 9,3
g l inf~rteure Ht " MDO MDB
Moula Couche XV E1 XV 177 20,63 15,25 12,76
Saint-Marcel Couche G
SM C2 43 G 14,05 13,36 12,83 'M2 lnf~rleure Ht MDO MDB Moula
Couche XV) E1 XV 177! 34,71 19,38 17,48
Saint-Marcel Couche
SM K5 36 22,41 18,02 M3 inf~rieure Ht MDO MDB Vloula
Couche XV I
E1 XV 177 41,42 25,14 30,75 E3 XV 571 37,4 24,63 27,69
Saint-Marcel Couche ?
SM K5 36 33,68 27,38 26,31 Coaehe G
SM I22 97 G 45,87 25,75 30,43 Couche I
SM T2 3591 40,3
5,8 7,03 VLOltot . VLO2tot . VLBI VLB2
7,14 5,83 7,12 7,34
7,16 6,51 7,35 7,42
7,08 6,8 7,03 7,28 6,72 6,23 6,92 6,9
VLOltot . V L O ~ o t . VLB
9,45 9,99 8,39
8,58 9,43 9,14 VLOltot . VLO2tot. VLB
10,21 9,55 10,26
9,82 10,15 VLOI VLO2 VLO3 VLB
9,16 8,54 4,42 10,14 9,94 9,19 3,51 9,45
9,61 9,26 5,9 9,56
9,21 8,78 4,93 10,78
9,45 8,79
397
Pl6istocbne sup6rieur (stade isotopique 5 e, l~6mien sensu lato). Les indices des Arvicola des couches VIII ~. IV sont comparables ~, ceux qu'on observe dans les couches sommitales de la Baume Gigny (couches XIXa h X) et h la grotte de l'l~glise en Charente, attribu6es au Pl6isto- c6ne sup6rieur plus r6cent (stades 4 et 3 de la chronologie isotopique). Ces 616ments permet- tent de supposer un ~ge P16istoc6ne sup6rieur r6cent pour les couches VII! ~. IV de la Baume Moula-Guercy, sans qu' i l soit possible d'etre plus precis.
L'association de P. lenki, M. brecciensis et A. bursae permet 6galement de pr6ciser le contexte biochronologique de la s6quence. En effet, ces trois formes ont 6t6 longtemps consid6r6es corn- me 6tant caract6ristiques du Pl6istoc6ne moyen. On sait maintenant qu'elles ont persistS, ~ l'6tat de relictes, durant le Pl6istoc6ne sup6rieur.
A. bursae a 6t6 reconnu dans des couches du Pl6istoc6ne sup6rieur de la Baume Loisia ~. Gi- gny (Chaline et Marquet, 1976) et de la s6quence d'Arcy-sur-Cure (Brochet, 1981), ainsi que dans d 'autres s6quences europ6ennes (Cottencher,

398 A. Defleur et al.
Tableau XX. Rupicapra sp. Dimensions de la P2 inf6rieure (en mm).
Table XX. Rupicapra sp. Measurements of the lower P2 (in mm).
M2 inf~rieure Ht MDO MDB VLOlto¢. VLO24~ VLB Moula
Couche XV E3 XV 727
Saut-du-Loup (Azilien) Extremes
Les Gras Extremes Moyenne
22,4 13,4 n=2
114,3-15,2 n--.5
12,8-14,9 13,56
12,7 n=l 13,3 n=4
12,1-13 12,42
6,8 n=2
6,4-6,8 n=5
6,2-7,4 6,96
6,8 n=2 6,4-7 n=5 6-7,4 6,9
7,9 a = l 8,3 rt=4
7,5-9.2 8,17
Campana-del-Pinar et La Carihuela notamment) datant de la mfime p6riode.
Dans le remplissage de la Baume Moula- Guercy, A. bursae est pr6sent dans les couches XV et XIVb de la banquette centre, ainsi que dans les couches XIX ~ XVI du sondage. Selon Jeannet (1995), la morphologie et les dimensions des sp6cimens recueillis dans les couches XVIII fiXVI n'apportent pas d'616ment biostratigra- phique.
P. lenki a 6t6 identifi6 dans des couches attri- buses au d6but du P16istoc6ne sup6rieur de la grotte de Lezetxiki en Espagne (Chaline, 1970). D'autre part, Marquet (1989) a mis en 6vidence la pr6sence de cette esp6ce dans des couches r6put6es "wtirmiennes" de la grotte du Salp6tre de Pompignan. P. lenki persiste 6galement durant l 'interglaciaire "Riss-Wiirm" au Moulin du Milieu (Marquet, 1989).
Darts la baume Moula-Guercy, P. lenki a 6t6 identifi6 dans la couche XV de la banquette cen- tre et dans les couches XIX ~ XVI du sondage. Les longueurs des M/1 recueillies (N 6, moyenne = 3,30 ram) montrent qu'il s'agit d'une forme tr6s 6volu6e.
M. brecciensis (sous la d6nomination M. cf. brecciensis mediterraneus-dentatus) a 6t6 mis en 6vidence par Chaline (1972a) dans les d6p6ts du PI6istocbne sup6rieur de l'Hortus. Plus r6cem- ment, Marquet (1989) a identifi6 cette esp6ce dans les s6quences de Font Juv6nal (Holoc6ne) et d'Abeurador (Pl~istoc6ne sup6rieur r6cent).
M. brecciensis est pr6sent dans les couches XV, XIVd et XIVb de la banquette centre de la Baume Moula-Guercy, ainsi que dans les cou-
ches XIX ~ XVI du sondage. Les M/1 sont carac- t6risdes par des triangles T4 et T5 peu ou pas confluents et deux dents pr6sentent un BRA4 bien marqu6. Cette morphologie prdsente des similitudes avec celle des populations "6vo- ludes" de la grotte des C6dres (Radulescu in : Defleur et Crdgut-Bonnoure 1995) et du Lazaret (Desclaux et Abbassi, sous presse), deux sdquen- ces attribudes ~ la fin du Pl6istoc6ne moyen (stade isotopique 6). Les longueurs des M/1 (n = 5, moyenne - 3,22) indiquent 6galement que nous sommes en pr6sence d'une forme 6volude de M. brecciensis.
A. bursae, P. lenki et M. brecciensis sont associ6s dans les couches XIX fi XVI du sonda- ge et dans le niveau XV de la banquette centre. On trouve 6galement des 616ments anatomiques se rapportant/l l 'une ou l'autre de ces trois for- rues dans les couches X1Vb et XIVd. Ces esp6- ces n'ont par contre pas 6td identifides dans les niveaux supdrieurs (couches XIVa fi IV des ban- quettes sud et centre).
3.3. Conclusions concernant ia biostrati- graphie
Les rdcentes fouilles effectu6es dans la Baume Moula-Guercy ont livr6 des 616ments anatomiques se rapportant fi des formes 6volu6es caract6ristiques du PI6istocbne sup6rieur et fi des esp6ces g6n6ralement rencontr6es durant le P16istoc6ne moyen.
La pr6sence d'A. bursae, de M. brecciensis et de P. lenki darts les couches XIX et XVIII, asso- ci6es aux formes Arvicola groupe cantiana/ter-

Pr6sentation pal6o-environnementale du remplissage de la Baume Moula-Guercy 399
o
i5 '~

400 A. Defleur et al.
Tableau XXI1. Comparaison entre les couches XIV et XV (classement ponderal couches confondues, la presence du taxon est signalee par le symbole +).
Table XXII. Comparison between levels XIV and XV (weight distribution of charcoals in both levels, symbol + indicates the presence of each species).
Taxons ii
Cornus mas L. (comouiller m~e)
Pimt9 type -(pin du groupe sylvestfis)
(~uercus sp. caducitbli6 (chOae "h feuillaLge caduc)
Juniperus communis (~genevrier commun)
Carpinus betulus L. (charmc)
Fagus sylvatica.L. (h~tre)
llex aqui[olium L. (houx) . . . . .
Prunus spinosa L. (prmlellier)
Hedera helix L.(lierre)
Phillyrea lat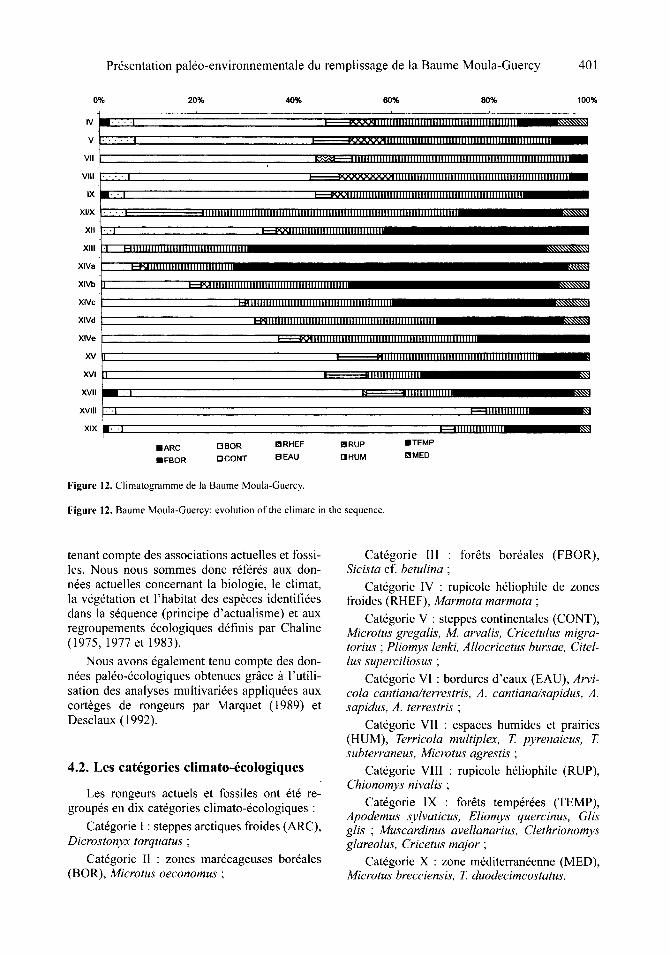
Pr6sentation pal6o-environnementale du remplissage de la Baume Moula-Guercy
0 % 20% 4 0 % 6 0 % 8 0 %
401
100%
IV
V
VII
VIII
IX
X I IX
Xl l "
Xll l
X l V a
XlVb il
X lVc ;
XIVd
X IVe I
X V 11 I XVI
XVl l
XVIII
XlX
I . . . . |
• i . . . . !
i
n , . I
. . . . I
- F i
-I
I . , . I
' 21
I I ~ , + % + % q l I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I "~\\\\'9,~I
I + I I I I I I I I I I I I I I I I I I I I I I I I I I I H I I I I I I I I I I I I I I H I I I I I I I I
IIIIIIIIIIIIIIIIIIIIIIIIit111111111111111111t11111t11111111111t1111111
I ~1111111111111t111111111111111111111111111111111111t11111
: . ~ ' . ) ( ~ l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l
••••••••••••••••••••••••|•••••••••••••••••••••••••••••••••••••••••••••••••••••••••• I ,~\\\\~1
1 J P r . ~ l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l
H I I I I I I I I t l I I I I I I I I I I I I I I I I I I I I l l l I t l l ~ \ \ \ \ \ \ \ \ \ \ X l
I~IF~ I I I I I I I I I I I I I l l l l l l l l l l l l l ,~ ,~,~.,t
~11111111111111111111111111111111111111111111 ~ .x.\\\\\\~l
HPl tlt111111111111111111111111111111111111111111 ~L%\\\\\~._~n
I - f q l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l k \ \ \ '~ : l
-" ; " . ~ q l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l t l l l l l l l l l l l l l
: --:31111111111111111111111111111111111111111111111111
n n U l l l l l l l l l l l l l l l
: t l l l l l l l l l l l l l
=
m A R C 13 BOR £1RHEF [ ] RUP • TEMP
I I FBOR r l CONT 13 EAU 131HU M m MED
=111111111111
= l l l l l l l l l l l l l i ~ l
Figure 12. Climatogramme de la Baume Moula-Guercy.
Figure 12. Baume Moula-Guercy: evolution of the climate in the sequence.
tenant compte des associations actuelles et fossi- les. Nous nous sommes donc r6f6r6s aux don- n6es actuelles concernant la biologie, le climat, la v6g6tation et l 'habitat des esp6ces identifi6es dans la s6quence (principe d'actualisme) et aux regroupements 6cologiques d6finis par Chaline (1975, 1977 et 1983).
Nous avons 6galement tenu compte des don- n6es pal6o-6cologiques obtenues grace ~ l'utili- sation des analyses multivari6es appliqu6es aux cort6ges de rongeurs par Marquet (1989) et Desclaux (1992).
4.2. Les categories climato-~cologiques
Les rongeurs actuels et fossiles ont ~t~ re- group6s en dix cat6gories climato-6cologiques :
Cat6gorie I : steppes arctiques froides (ARC), Dicrostonyx torquatus ;
Cat6gorie I1 : zones mar6cageuses bor6ales (BOR), Microtus oeconomus ;
Cat4gorie II1 : for6ts bor4ales (FBOR), Sicista cf. betulina ;
Cat6gorie IV : rupicole h61iophile de zones froides (RHEF), Marmota marmota ;
Cat6gorie V : steppes continentales (CONT), Microtus gregalis, M. arvalis, Cricetulus migra- torius ; Pliomys lenki, Allocricetus bursae, Citel- lus superciliosus ;
Cat6gorie VI : bordures d'eaux (EAU), Arvi- cola cantiana/terrestris, A. cantiana/sapidus, A. sapidus, A. terrestris ;
Cat6gorie VII : espaces humides et prairies (HUM), Terricola multiplex, T. pyrenaicus, T. subterraneus, Microtus agrestis ;
Cat6gorie VII1 : rupicole h61iophile (RUP), Chionomys nivalis ;
Cat6gorie IX : for~ts temp6r6es (TEMP), Apodemus sylvaticus, Eliomys quercinus, Glis glis ; Muscardinus avellanarius, Clethrionomys glareolus, Cricetus major ;
Cat6gorie X : zone m6diterran6enne (MED), Microtus brecciensis, T. duodecimcostatus.

402 A. Defleur et al.
4.3. Succession des pal~o-environnements
De la base connue du remplissage/l son som- met, le climatogramme obtenu (figure 12) per- met de distinguer trois phases climatiques prin- cipales.
- Phase Ill : phase d6couverte froide (ou phase steppique froide). Elle concerne les cou- ches XIX/t XV et peut ~tre divis6e en deux 6pi- sodes elimatiques distincts.
Sous-phase IIIa : couches XIX /~ XVI1 du sondage. Elle est caract6ris6e par l 'abondance des esp6ces des steppes continentales arides (M. gregalis, C. superciliosus, A. bursae). La pr6sen- ce des esp6ces des zones arctiques et bor6ales (D. torquatus, Sicista cf. betulina et M. oecono- mus) souligne le caract6re tr6s rigoureux du cli- mat. Ces 616ments permettent de sugg6rer que cette phase correspond/t un maximum glaciaire. Ce caract6re est cependant attdnu6 par la propor- tion non ndgligeable d'esp6ces des biotopes tem- p6r6s (A. sylvaticus, E. quercinus et C. glareo- lus). Le paysage devait correspondre/l une step- pe continentale tr6s aride avec persistance d'ilots forestiers dans des niches 6cologiques prot6g6es.
Sous-phase IIIb : phase de transition (cou- ches XVI et XV). Elle est caract6ris6e par une diminution progressive des proportions des esp6- ces des biotopes froids et des steppes continenta- les. Celles-ci sont remplacdes par des formes inf6od6es g des biotopes temp6r6s. Ce ph6nom6- ne est vraisemblablement lid ~ un r6chauffement climatique qui se traduit par l'installation de milieux d6couverts plus humides. On peut noter que les esp6ces des milieux forestiers sont peu reprdsentdes dans la couche XVI. Elles devien- nent plus abondantes dans la couche XV. Le pay- sage correspondait alors/l une steppe bois6e.
Ces deux niveaux sont vraisemblablement contemporains de la limite entre le Pl6istoc6ne moyen et le P16istoc~ne sup6rieur.
- Phase II : phase foresti6re (ou phase tempS- r6e). Elle concerne les couches X1Ve/l XI et peut 0tre divisde en deux 0pisodes climatiques.
Sous-phase IIa phase foresti6re tempdrde. La phase lla concerne les couches XIV (XIVe fi X1Va et XIII) de la banquette centre. Elle cor- respond/l l'installation d'une for~t tempdrde qui atteint son maximum d'extension dans la couche X1Va. Les esp6ces des espaces d6couverts humi- des et des steppes continentales sont encore bien reprdsent6es dans les couches XIVe fi XIVc. Par la suite, ces formes r6gressent progressivement
au profit des taxons forestiers. Les esp6ces des biotopes temp6r6s deviennent alors largement majoritaires. Cette remarquable fluctuation des pal6omilieux est lide fi un r6chauffement clima- tique de grande amplitude. La phase lIa cor- respond vraisemblablement/~ un stade intergla- ciaire.
Sous-phase I I b : phase de transition. Elle concerne les couches XII et XI. Cette deuxi6me phase de transition est caract~ris6e par une ldg6- re r6gression des esp6ces des for0ts temp6r6es qui s 'accompagne d'une augmentation des esp6- ces des zones bor6ales et des espaces ddcouverts humides. Ces couches correspondent vraisem- blablement aux pr6mices d'un changement cli- matique important.
- Phase I : phase ddcouverte. Cette phase con- cerne l 'ensemble des niveaux sup6rieurs de la baume Moula-Guercy (couches X/XI fi IV). Elle est caract6risde par la r6gression des esp6ces des milieux forestiers au profit des esp6ces des bioto- pes froids et des esp~ces des steppes continenta- les. La bonne repr6sentation des esp6ces des espaces ddcouverts humides tend cependant fi limiter l 'importance du caract6re froid du climat. Les zones de hauteur devaient 0tre plut6t arides et occup6es par la steppe. Les environs du Rh6ne constituaient une zone mar6cageuse. Les fonds de valldes devaient conserver un cachet humide.
4.4. Place de la Baume Moula-Guercy dans la chronologic climatique de l'Euro- pe de rOuest
Les associations de rongeurs permettent de replacer la s6quence de Moula-Guercy dans la chronologie climatique de l 'Europe occidentale et de proposer quelques corr61ations.
Phase !11
Comme nous l 'avons vu pr~c~demment, les donndes biostratigraphiques indiquent que les niveaux XIX fi XVII ont un fige Pldistoc6ne moyen final (phase Illa). La prdsence de D. tor- quatus et de Sicista cf. betulina darts la couche XIX peut 6tre corr616e avec la phase de migra- tion de ces deux esp6ces en Europe occidentale qui a 6t6 mise en 6vidence par Marquet (1989) dans la grotte Vaufrey (couches VII, VIII et XI) et qui correspond fi un 6pisode de froid majeur contemporain de la fin du P16istoc6ne moyen.
La phase IIIb (couches XVI et XV) de la Baume Moula-Guercy a 6t6 interprdtde comme

Pr6sentation pal6o-environnementale du remplissage de la Baume Moula-Guercy 403
6tant une p6riode de transition climatique. Des conditions climatiques similaires ont 6t6 mises en 6vidence dans trois s6quences pr6historiques du domaine m6diterran6en fran~ais attribu6es/l la fin du P16istoc6ne moyen. Les micromammi- f6res identifi6s dans les niveaux sup6rieurs de la grotte du Lazaret/~ Nice sugg6rent 6galement un climat temp6r6 mais plus froid que l'actuel et un environnement ouvert (Desclaux, 1996). En Ard6che, les niveaux de base de la grotte de Payre (Moncel et al., 1993) correspondent ~un milieu ouvert temp6r6 avec des zones foresti6res /t proximit6 de l'habitat. Enfin, les associations de mammif6res de la grotte des C6dres sont repr6sentatives d'un milieu ouvert et humide (Desclaux, 1995) et de conditions climatiques peu rigoureuses (Radulescu in : Defleur et Cr6gut-Bonnoure, 1995).
Phase II
Les niveaux XIV et XIII de la Baume Moula- Guercy correspondent ~ une phase forestbre et/l un climat temp6r6 (Phase IIa). ll peuvent ~tre mis en relation avec l'optimum forestier du stade 5e du diagramme pollinique de la Grande Pile (Woillard, 1980).
Dans le domaine mdditerran6en, on dispose de peu de donn~es concemant les associations de rongeurs durant cet interglaciaire. On notera cependant que les associations de rongeurs des couches XVII et XX du Salp~tre de Pompignan reconnues par Marquet (1989) peuvent dater de la m~me pdriode. Les couches XII et XI (Phase lib) de la Baume Moula-Guercy caract6ris6es par une diminution sensible des taxons forestiers cor- respondent vraisemblablement fi un 6pisode froid du stade 5 (5d ou 5b du diagramme de la Grande Pile).
Phase I La phase I de la Baume Moula-Guercy cor-
respond fi un refroidissement du climat, mais de moindre amplitude par rapport ~ celui de la phase Ilia (couches X1X fi XVII).
La faible repr6sentation des rongeurs dans les niveaux X/XI ~ IV rend cependant difficile la comparaison avec d'autres s6quences pr6histo- riques. Cette association de rongeurs est compa- rable /t celles de l'Hortus (Chaline, 1972a) et d 'Arma delle Manie (Abbassi et Desclaux, 1996b) qui sont attribu6es au Weichselien ("Wtirm II").
Les donn6es recueillies dans la Baume Moula-Guercy confirment les observations de Chaline (1972a) qui sugg6rait que cette p6riode
avait 6t6 moins froide dans les r6gions m6ridio- nales. On peut 6galement noter que la pr6sence ponctuelle de D. torquatus dans les niveaux IX et IV est li6e /~ la migration de cette esp6ce en France durant le Weichselien. Le lemming/t col- lier a en effet 6t6 identifi6 par Chaline (1972b) et Marquet (1989) darts de nombreuses s6quences pr6historiques datant de la m~me p6riode (Les Canalettes, La Quina, Marillac, Roc-en-Pail, la Roche-Cotard, la grotte Boccard et diff6rentes s6quences d'Arcy-sur-Cure).
On notera 6galement que l'analyse de la communaut6 des mammif~res (m6thodes des c6nogrammes) men6e par Montuire et Desclaux (1997a, b) a confirm6 cette interpr6tation clima- tique et environnementale du remplissage.
4.5. Conclusions
Les micromammif6res recueillis durant les r6centes campagnes de fouilles permettent de pr6ciser l'fige du remplissage de la Baume Moula-Guercy.
La morphologie, les dimensions et la valeur de l'indice d'6paisseur d'6mail des formes se rapportant au genre Arvicola, le degr6 d'6volu- tion de M. brecciensis et de P. lenki et la pr6sen- ce de ces deux demi6res formes associ6es fi A. bursae dans les couches XIX et XVIII du sonda- ge sugg6rent que ces niveaux datent de la fin du Pl6istoc6ne moyen.
La valeur de l'indice d'6paisseur d'6mail des Arvicola et le caractbre "moderne" des autres esp~ces identifi6es dans les couches m6dianes (XIV et XIII) indiquent que ces niveaux datent du d6but du Pl6istoc~ne sup6rieur.
Les micromammif6res identifi6s dans les cou- ches sup6rieures (XII fi IV) sont moins abondants. On peut noter qu'il s'agit d'une association carac- t6ristique du d6but du Pl6istoc6ne sup6rieur. Enfin, la pr6sence ponctuelle de P. lenki, d'A. bur- sae et de M. brecciensis dans les couches XIVa/t XV est remarquable. Elle confirme que ces esp6- ces, longtemps consid6r6es comme 6tant caract6- ristiques du Pl6istoc6ne moyen persistent au d6but du Pl6istoc6ne sup6rieur en Europe occi- dentale.
5. l{tude anthracologique
Cent-quatre-vingt charbons de bois prove- nant de la couche XV ont 6t6 examin6s. Ces

404 A. Defleur et al.
charbons, totalisant 1,32 g sont g6n6ralement en bon 6tat et pr6sentent, pour la plupart, une taille inf6rieure ~ 1-2 mm. Les taxons suivants ont 6t6 identifi6s:
Pinus type sylvestris (pin du m~me type ana- tomique que le pin sylvestre), Quercus sp. cadu- cifoli4 (chine/l feuillage caduc), Comus mas L. (cornouiller m~le), Juniperus communis L. (gen6- vrier commun), Prunus spinosa L. (prunellier), Phillyrea latifolia L. (filaire ft. larges feuilles), Ulmus sp. (orme, esp6ce ind~terminde), Ilex aqu-
folium L. (houx), Hedera helix L. (lierre), Rosa sp. (6glantier, esp6ce inddtermin6e), Rhamnus saxatilis Jacq. (nerprun des rochers), Angio- sperme ind6terminable, Rosac6e?
Ces taxons sont class6s par ordre d'abondan- ce. Pinus et Quercus, il 4galitS, sont majoritaires et constituent pr6s de la moiti6 de la masse tota- le, Pinus 6tant pr4sent dans 12 des 13 6chan- tillons prSlev6s, tandis que Quercus apparaTt dans 3. Comus mas vient en troisi6me position, bien avantJuniperus. 11 est difficile de distinguer les pins du groupe antomique sylvestris (Rol, 1932), cependant le relatif bon 6tat des charbons ainsi que les contextes floristique et g6ogra- phique permettent de consid6rer qu'il s 'agit vrai- semblablement ici de l 'esp~ce sylvestris L. L'incertitude est identique pour les chines cadu- cifoli6s qui, dans l'6tat actuel des connaissances, ne peuvent ~tre identifi6s. II est 6galement diffi- cile de distinguer les bois des divers ormes, en d6pit de la tr6s faible taille des charbons l 'esp6- ce Ulmus minor Miller semble la plus probable.
Interpr6tation Les taxons identifi6s appartiennent aux for-
mations foresti~res caducifolides tempSr6es sub- atlantiques et mSdio-europdennes. Ils t6moignent d'un climat proche de celui qui r6gne actuelle- ment dans la r6gion. On doit remarquer la prd- sence de Phillyrea latifolia, taxon m6diterran6en. Ce vdgStal apparait cependant aujourd'hui dans diverses localitds de l 'Ouest de la France et du Sud du Massif Central. 11 remonte 6galement lar- gement darts la vall6e du Rh6ne, une station sep- tentrionale 6tant signal6e dans le d6partement de l'Ain (Breistroffer, 1953-1954). L'4tude de la couche XIV sus-jacente, nettement plus riche en charbons de bois (environ 600 charbons prSsen- tant les m4mes caract6ristiques que pr6c6dem- ment), montre une composition floristique trbs proche qui permet de compenser la faiblesse de l'6chantillonnage de la couche XV (tableau XXI1). La couche XIV, en raison vraisemblable-
merit de son meilleur 6chantillonnage, r6v61e la prSsence d'autres arbres dominants (Fagus, Car- pinus) des for~ts caducifoli6es tempSr6es. Parmi le sous 6tage des domin6s (Comus, Ilex) apparait Taxus Salix et, dans une moindre mesure, Ulmus indiquent la prSsence d'un milieu humide. On doit remarquer que le cornouiller repr6sente plus des trois quarts des charbons de la couche XIV et montre/l l'Svidence une pr6f6rence pour la r6col- te du bois de cette esp6ce, tr6s dense et consti- tuant un excellent combustible. On peut noter que Phillyrea donne 6galement un bois tr6s dense fournissant des braises durables, ce qui peut expliquer sa r6colte dans un milieu off il devait 6tre assez rare. Pinus, Juniperus, Rhamnus et Phillyrea proviennent vraisemblablement de lieux plus ouverts comme les pentes escarp6es.
Les donn6es pal6obotaniques et pal6o-Scolo- giques concernant la r6gion pour la m6me p6rio- de sont relativement limit6es et repr6sentSes par les 6tudes palynologiques du marais des Echets, au nord-est de la ville de Lyon (Beaulieu et Reille, 1984), et la r6gion du Velay (Reille et al., 1998). Les analyses palynologiques, sans avoir la pr6ci- sion spatiale des charbons de bois, rendent comp- te des m6mes formations foresti6res caducifoli6es temp6r6es. L'6tude des charbons permet, par la pr6sence de Phillyrea espbce thermophile, de pr6- ciser que le climat 6tait au moins aussi chaud qu'actuellement. Par comparaison, les couches moust6riennes de la Baume N6ron (Defleur et al., 1994), situ6e ~_ une quinzaine de m6tres de la Bau- me Moula-Guercy, ont livr6 des charbons de pin sylvestre, gen6vrier et bouleau pour un contexte climatique froid (stade isotopique 3). En conclu- sion, l'6tude anthracologique de la couche XV indique un milieu forestier caducifoli6 caract6ri- sant un climat temp6r6 tr6s proche de l'actuel comme le confirme la prSsence d'une esp~ce m6diterran~enne existant de nos jours dans la r6gion.
6. Conclusions g6n6rales
Les nouvelles fouilles et analyses des docu- ments pal6ontologiques et anthracologiques r6ali- s6es dans la Baume Moula-Guercy entre 1992 et 1999 ont permis d'accumuler une somme impor- tante de donnSes nouvelles concernant la pal6o- 6cologie, la pal6oclimatologie et de pr6ciser la chronologie de la raise en place des d6p6ts.
Les donn6es convergentes fournies par l'6tu- de des rongeurs et des grands mammif6res per-

Pr6sentation pal6o-environnementale du remplissage de la Baume Moula-Guercy 405
mettent de diviser les 8 m de remplissage connus en une succession remarquable de 3 phases cli- matiques corr616es aux stades isotopiques 6/l 4 de la chronologie isotopique marine.
La phase Ilia (couches XIX /l XVII) cor- respond ~ une phase steppique et fi un climat tr6s froid. La pr6sence de D. torquatus, de Sicista cf. betulina et de C. superciliosus dans certains niveaux, ainsi que l'abondance de M. gregalis sugg6rent qu'il s'agit d'un maximum glaciaire qui correspond ~ la fin du Pl6istoc~ne moyen (stade isotopique 6, Saalien).
La phase IIIb (couches XVI et XV) cor- respond fi une p6riode de transition climatique. 11 s'agit vraisemblablement de la limite entre le Pl6istoc6ne moyen et le Pl6istoc6ne sup6rieur.
La phase II (couches XIVe fi XII) correspond /tune phase foresti6re et fi un climat temper6. II s'agit d'un interglaciaire datant du d6but du Pl6istoc6ne sup6rieur (stade isotopique 5, l~emien sensu lato).
La phase I (couches X/XI fi IV) correspond/t l'installation d'un climat froid et humide. Le pay- sage devait ~tre constitu6 de zones steppiques dans les hauteurs et de milieux humides dans les fonds de vall6es. Les conditions climatiques sont moins rigoureuses que celles de la phase IV. Cette seconde p6riode froide peut 6tre attribute au P16istoc6ne sup6rieur (stade isotopique 4, Weichselien).
Si les phases 6 et 4 sont connues darts plu- sieurs sites du Sud-Est de la France, l'6pisode 66mien se manifeste g6n6ralement dans les rem- plissages karstiques soit par des planchers stalag- mitiques soit par des lacunes s6dimentaires. Or, cette p6riode est repr6sent6e dans cette cavit6, par une formation homog6ne de plus d'un m~tre de puissance, livrant faune, industries lithiques, structures d'habitat mais aussi plusieurs dizaines de restes humains correspondant fi au moins six individus cannibalis6s (Defleur et al., 1999). Les r6sultats d6j~ acquis font de ce remplissage une s6quence de r6f6rence dans l'Europe m6diterra- n6enne principalement pour la connaissance de l't~6mien et de la p6riode de transition entre le P16istoc6ne moyen et le P16istoc6ne sup6rieur.
La poursuite des travaux devrait nous per- mettre de compl6ter nos donn6es sur certaines esp6ces rares du Pl6istoc6ne europ6en, recon- hues dans ce gisement : Cuon, U. thibetanus, M. palerminea, F. minuta, Allocricetus, C. major, Sicista etc. et d'affiner les premiers r6sultats pr6- sent6s darts cette 6tude.
Remerciements
Nos remerciements vont au Minist6re de la Culture (DRAC Rh6ne/Alpes) et au Conseil G6n6ral de l'Ard6che qui, entre 1992 et 1999, ont soutenu financi+rement le projet de fouilles et de recherches entrepris dans cette cavit6.
R~f~rences
Argant, A., 1991. Carnivores quaternaires de Bourgogne. Documents des Laboratoires de G6ologie de Lyon 115, 1-301.
Ballesio, R., 1979. Le gisement pl6istoc+ne sup6rieur de la grotte de Jaurens fi Nespouls Corr6ze, France : les Carnivores (Mammalia, Carnivora). In : Canidae et Hyaenidae. Nouv. Arch. Mus. Hist. Nat. Lyon 17, 25-55.
Abbassi, M., Desclaux, E., 1996. Arvicola Lac6p6de, 1799 (Rodentia, Mammalia) de quatre s6quences du Sud-Est de la France et de Ligurie datant de la fin du P16istoc6ne moyen et du d6but du Pl6istoc6ne sup6rieur. Quaternaire 7 (1), 29-37.
Abbassi, M., Desclaux, E., 1996. Biostra- tigraphie et pal6oenvironnements de quelques sites du Sud-Est de la France et de Ligurie d'apr6s l'6tude des rongeurs. XIII Int. Congr. of Prehistoric and Protohistoric Sciences, Abstracts T. 1, 8-14 September 1996, Forli, Italia, pp. 529-535.
Abbassi, M., Desclaux, E., Marquet, J-C., Chaline, J., 1998. R6partition et ~volution des Arvicola (Lac6p6de, 1799) au cours du Pl6istoc6ne moyen et sup6rieur en France et en Ligurie. Quaternaire 9 (2), 105-116.
Abbassi, M., Desclaux, E., 1998. Biostrati- graphic et pal6o6cologie de 5 s6quences pr6- historiques datant de la fin du P16istoc6ne moyen et du d6but du P16istoc6ne sup6rieur dans le Sud de la France et en Ligurie. XIII UISPP, Congr. Proc. vol. 1,529-535.
Beaulieu, J. de, Reille, M., 1984. A long Upper Pleistocene pollen record from Les Echets, near Lyon, France. Boreas, 111-132.
Bonifay, M.-F., 1971. Carnivores quaternaires du Sud-Est de la France. M6m. Mus. Hist. Nat. Paris n.s., C, 21, 43-377.
Breistroffer, M., 1953-1954. Les limites septen- trionales d'extension de la flore m6diterra- n6enne dans la Dr6me et l'Ard6che. Bull. Soc. Bot. Fr. m6moires, 62-95.

406 A. Defleur et al.
Brochet, G., 1981. Etude comparative des faunes d'Arcy-sur-Cure, la Cotte Sainte-Brelade et de Gigny ~ l'aide d'une methode d'ecologie quan- titative. DEA Universit6 de Dijon.
Brugal, J.-Ph., 1983. Application des analyses multidimensionnelles ~t l'etude du squelette des membres des grands Bovides Pleistocenes (Grottes de kunel-Viel, Herault) ; perspectives 6volutives. These, Universit6 d'Aix-Marseil- [e, Luminy.
Brunet-Lecomte, E, Chaline, J., 1991. Morpho- logical evolution and phylogenetic relations- hips of the ground voles (Terricola, Arvico- lidae, Rodentia). Lethaia 24, 69-80.
Brunet-Lecomte, R, Volobouev, V., 1994. Com- parative morphometry and cytogenetics of Microtus (Terricola) multiplex (Arvicolidae, Rodentia) of western French Alps. Z. S~iug- etierkunde 59, 116-125.
Brunet-Lecomte, E, Desclaux, E., Defleur, A., 1996. Les Terricola (Rodentia, Mammalia) du gisement P16istoc6ne de la Baume Moula- Guercy ~, Soyons (Ardache, France). Bull. Mus. Anthr. Prehist. Monaco 38, 9-15.
Chaline, J., 1970. Pliomys lenki, forme relique du Pleistocene superieur de la grotte de Lezetxiki (Guipuzcoa, Espagne). Munibe 22 (1-2), 43- 49.
Chaline, J., 1972. Les rongeurs du Wiirmien lI de la grotte de l'Hortus (Valflaunes, Herault). Etudes Quaternaires 1,233-240.
Chaline, J., 1975. Les rongeurs et la climatologie du Pleistocene superieur. In : L'Homme et son environnement pendant le Wtirm en Europe de l'Ouest. Palaeoecology of early man, Sous- groupe de travail Pleistocene superieur, Ouest de l'Europe. Universit6 de Bordeaux Ill, 5-16.
Chaline J.,1977. Les rongeurs au pilori? Analyse critique des reconstitutions paleoecologiques en microtheriodontologie. In : Laville, H., Renault-Miskovsky, J. (ed.), Approche 6colo- gique de l'Homme fossile, Bull. Ass. Fr. Et. Quat. 47, 75-81,
Chaline, J., 1983. Les rongeurs, les paleoenvi- ronnements et les climats du Pleistocene moyen de Vergranne (Doubs). Annales Scientifiques de l'Universit6 de Franche- Comte Besan~;on. Geologie, 5 (4), 31-45.
Chaline, J., Marquet, J.-C., 1976. Les conse- quences stratigraphiques de la persistance en France dans le Wtirm ancien des formes reli- ques Pliomys lenki et Allocricetus bursae
(Rodentia). C. R. Acad. Sc. Paris, 282 D, 1941-1942.
Clot, A., 1980. La grotte de la Carri6re (Gerde, Hautes-Pyrenees). Stratigraphie et paleontolo- gie des Carnivores. These, Toulouse.
Cregut-Bonnoure, E., 1987. Caract6res distinc- tifs du carpe et du tarse d'Hemitragus et de Capra (Mammalia, Bovidae, Caprinae). Bull. Mus. Anthropol, Prehist. Monaco 30, 43-94
Cregut-Bonnoure, E., 1989. Un nouveau Capri- ne, Hemitragus cedrensis nov. sp., (Mamma- Iia, Bovidae) des niveaux pleistocenes moyen de la Grotte des Cedres (Le Plan d'Aups, Vat): interat biostratigraphique. Geobios 22, (5), 653-663.
Cregut-Bonnoure, E., 1992a. lnterat biostratigra- phique de la morphologie dentaire de Capra (Mammalia, Bovidae). Ann. Zool. Fenn. Hel- sinki 28,273-290.
Cregut-Bonnoure, E., 1992b. Pleistocene tahrs, ibexes and chamois of France. In : Spitz, F., Janeau, G., Gonzalez, G., Aulagnier, S. (dir.) Ongules/Ungulates 91. S.F.E.P.M.-I.R.G.M. (Ed.), Paris-Toulouse, pp. 49-56.
Cregut-Bonnoure, E., 1992c. Dynamics of Bovid migration in Western Europe during the midd- le and the late Pleistocene. Cour. Forsch.-Inst. Senckenberg, Frankfurt a.M., 153, 177-185.
Cregut-Bonnoure, E., 1992d. Les Caprinae (Mammalia, Bovidae) du Pl6istocene d'Eu- rope. Interet biostratigraphique, palaoecolo- gique et archeozoologique. Mere. Soc. Geol. Fr. 160, 85-93.
Cregut-Bonnoure, E., 1992e. La faune de mam- miferes de Chinchon !I (Saumanes, Vauclusc). Bull. Archeologique de Provence 21, 9-13.
Cregut-Bonnoure, E., 1996a. Ordre des Carni- vores. II : Famille des Canidae. In : Les grands Mammiferes plio-pleistocenes d'Europe, col- lection "Prehistoire", Masson, Paris, pp. 156- 166
Cregut-Bonnoure, E., 1996b. A review of small Middle Pleistocene bears from France. Acta Zool. Cracov. 39 (21 ), 89-101.
Cregut-Bonnoure, E., Guerin, C., 1986. La faune de mammiferes de l'Abri Moula (Soyons, Ar- deche). Bull. Soc. Et. Sci. Nat. Vaucluse 2, 41- 87.
Cregut-Bonnoure, E., Faure, M., Guerin, C., 1994. Faune et vegetation: le Bau de I'Aube- sier. In : Le Paleolithique moyen en Vaucluse. Notices d'Archeologie vauclusienne 3, 116- 118.

Pr6sentation pal6o-environnementale du
Debard, E., 1988. Le Quaternaire du Bas-Viva- rais d'apr6s l'dtude des remplissages d'avens, de porches de grotte et d'abris sous roche. Dynamique sddimentaire, pal6oclimatologie et chronologic. Documents des Laboratoires de Gdologie de Lyon 103, 1-317.
Defleur, A., 1995. Nouvelles ddcouvertes de res- tes humains moustdriens dans les ddp6ts de la Baume Moula-Guercy (Soyons, Ard6che). Bull. M6m. Soc. Anthropol. Paris 7 (3-4), 185-190.
Defleur, A., 1998. Des fossiles pr6-ndanderta- liens dans la moyenne vallde du Rh6ne. La Science au pr6sent 1999 Encyclopedia Universalis, pp. 41-42.
Defleur, A., Dutour, O., Valladas, H., Vander- meersch, B., 1993a. Cannibals among the Neanderthals? Nature 362 (6417), 214.
Defleur, A., Dutour, O., Valladas, H., Combier, J., Vandermeersch, B., 1993b. Ddcouvertes de testes humains darts les niveaux moustdriens de l'abri Moula (Soyons, Ardeche). C. R. Acad. Sci. Paris 1I, 316, 1005-1010.
Defleur, A., Dutour, O., Vandermersch, B., 1993c. t~tude de deux dents humaines prove- nant des niveaux moustdriens de la Baume Ndron (Soyons, Ard6che). Bull. M6m. Soc. Anthropol. Paris 1992, 4 (2), 127-134.
Defleur, A., Bez, J.-F., Crdgut-Bonnoure, E., Fontugne, M., Jeannet, M., Magnin, F., Talon, B., Thinon, M., Combier, J., 1994a. Industries, biostratigraphie, testes humains et datation du gisement moust6rien de la Baume Ndron (Soyons, Ardeche). C. R. Acad. Sci. Paris II, 318, 1409-1414.
Defleur, A., Bez, J.-F., Crdgut-Bonnoure, E., Desclaux, E., Onoratini, G., Radulescu, C., Thinon, M., Vilette, Ph., 1994b. Le niveau moust6rien de la grotte de l'Adaouste (Jou- ques, Bouches du Rh6ne). Approche culturelle et paldoenvironnements. Mus. Anthrop. Prdhist. Monaco 37, 11-48.
Defleur, A., Cr6gut-Bonnoure, E., 1995. Le gise- merit Pal6olithique moyen de la grotte des C6dres (Var). Documents d'Arch6ologie Fran- qaise. Maison des Sciences de l'Homme, Paris.
Defleur, A., Crdgut-Bonnoure, E., Desclaux, E., 1998. Premi6re raise en 6vidence d'une sdquence ddmienne ~ restes humains dans le remplissage de la Baume Moula-Guercy (Soyons, Ardeche). C. R. Acad. Sci. II, 326 (6), 453-458.
remplissage de la Baume Moula-Guercy 407
Defleur, A., White, T., Valensi, E, Slimak, L., Crdgut-Bonnoure, E., 1999. Neanderthal Can- nibalism at Moula-Guercy, Ard6che, France. Science 286, 128-131.
Desclaux, E., 1992. Les petits vertdbr6s de la Caune de l'Arago (Pyr6ndes-Orientales, France). Biostratigraphie, pal6o6cologie et taphonomie. Bull. Mus. Anthrop. Prdhist. Monaco 35, 35-64.
Desclaux, E., 1996. Contribution des micro- mammif6res a la connaissance des pal6oenvi- ronnements des chasseurs de la grotte du Lazaret fi Nice (France, Alpes-Maritimes). Arch6am 4, 7-11.
Desclaux, E., Abbassi, M., (sous-presse). Biostratigraphie des microvert6br6s de la grot- te du Lazaret fi Nice (France, Alpes-Mari- times). In : Lumley, H. de (ed.), Cadre chro- nologique du remplissage de la grotte du Lazaret (g6omorphologie, stratigraphie, bio- stratigraphie, pal6o6cologie, 6volution cultu- relle, gdochronologie).
Desclaux, E., Abbassi, M., Marquet, J-C., Chaline, J., Van Kolsfschoten, T., 2000. Dis- tribution and evolution ofArvicola Lac6p6de, 1799 (Mammalia, Rodentia) in France and Liguria (Italy) during the Middle and Upper Pleistocene. Acta Zool. Cracow, (sous presse).
Fernandez, Ph., 1996. Le cheval et l'aurochs : biostratigraphie et strat6gie alimentaire des n6andertaliens au Bau de l'Aubesier Monieux (Vaucluse). M6moire de DEA, Universit~ Lumi~re Lyon I1.
Guadelli, J.-L., 1987. Contribution ~ l'6tude des zooc6noses pr~historiques en Aquitaine (WOrm ancien et interstade wtirmien). Th6se, Universit6 Bordeaux 1.
Heinrich, W.D., 1978. Zur biometrischen Erfassung eines Evolutionstrends bei Arvicola (Rodentia, Mammalia) aus dem Pleistoz~in Th~iringens. S~iugetierkdl. Inform. 2/3, 21.
Heinrich, W.-D., 1990. Some aspects of the evo- lution and biostratigraphy of Arvicola (Ro- dentia, Mammalia) in the central european Pleistocene. In : Int. Fefjar, O., Heinrich, W.- D. (Eds.), Symp. Evol. Phyl. Biostr. Arvico- lids. Praha, 165-182.
Hinton, M.-A.-C., 1926. Monograph of the Voles and Lemmings (Microtinae), Living and Extinct. Vol. I. British Museum (Natural His- tory), London.
Jeannet, M., 1995. Les Cric6tid6s (Mammalia, Rodentia) des gisements moust6riens de

408 A. Defleur et al.
Soyons (Ard6che, France). Bull. mens. Soc. Linn. Lyon 64 (1), 41-48.
Kolsfschoten, T. van, 1990. The evolution of the mammal fauna in the Netherlands and the middle Rhine area (Western Germany) during the late middle Pleistocene. Medelingen Rijks Geologische Dienst 43 (3), 1-69.
Kurt6n, B., 1965. On the evolution of the European wild cat, Felis silvestris Schreber. Acta Zool. Fenn. 111, 1-26.
Laville, H., Prat, F., Thibault, C. 1972. Un gise- ment/~ faune du P16istoc6ne moyen : la grotte de l'l~glise /l C6nac-et-Saint-Julien (Dordo- gne). Quaternaria XV1, 79-119.
Marquet, J.-C., 1989. Paldoenvironnement et chronologie des sites du domaine atlantique frangais d'fige Pldistocbne moyen et sup6rieur d'aprbs l'6tude des rongeurs. Th6se de Doctorat, Universit6 de Dijon.
McCuaig Balkwill, D., Cumbaa, S.-I., 1992. A guide to the identification ofpostcranial bones of Bos taurus and Bison bison. Syllogeus, Ottawa.
Montuire, S., Desclaux, E., 1997a. Analyse paldo- 6cologique des faunes de mammif6res et envi- ronnements darts le Sud de la France au cours du Pleistoc6ne. Quaternaire 8 (1), 13-20.
Montuire, S., Desclaux, E., 1997b. Palaeo- ecological analysis of mammalian faunas and environmental evolution in the South of France during the Pleistocene. Boreas 26, 355-265.
Prat, F., Thibault, C., 1979. Le gisement de Nau- terie/~ La Romieu (Gets). Fouilles de 1967/~ 1973. M6m. Mus. Nat. Hist. Nat, n.s. C, XXXV, 1-82.
Rabeder, R., 1976. Die Carnivoren (Mammalia) aus dem Altpleistoz~in von Deutsch-Altenburg 2 mit Beitragen zur Systemetik einiger Musteliden und Caniden. Beitr. Palaont. Osten'. 1, 5-119.
Rabeder, G., 1992. Gli orsi spel6i delle Conturines. Scavi paleozoologici in una caverna delle Dolomiti (Val Badia) a 2800 metri. Athesia edit., Bolzano.
Reille, M., Andrieu, V., Beaulieu, J. de, Guenet, R, Goeury, C., 1998. A long pollen record from Lac du Bouchet, Massif Central, France: for the period ca. 325 to 100 ka BP (OIS 9c to OIS 5e). Quat. Sci. Rev. 17, 1107-1123.
Rol, R., 1932. Note sur un essai de classification du genre Pinus d'apr6s des caract6res tir6s de I'anatomie du bois. Congr. Soc. Savantes 65, 333-341.
Tavoso, A., Cr6gut-Bonnoure, E., Gudrin, C., Pernaud-Orliac, J., Cammas, R. 1990. La grot- te de La Niche/t Montmaurin (Haute-Garon- he, France). Nouvelles donn6es biostratigra- phiques et approche taphonomique. C. R. Acad. Sci. Paris [l, 310, 95-100.
Toshi, A., 1965. Mammalia (Lagomorpha, Ro- dentia, Carnivora, Ungulata, Cetacea). Fauna d'Italia. Calderini, Bologna.
Copyright © 2022 FDOKUMEN




















