
Transthyretin Accelerates Vascular Aβ Deposition in a Mouse Model of Alzheimer's Disease
Upload
independentCategory
view
0download
0
Neuron, Vol. 46, 541–554, May 19, 2005, Copyright ©2005 by Elsevier Inc. DOI 10.1016/j.neuron.2005.04.008
Presenilin-Dependent Transcriptional Controlof the A�-Degrading Enzyme Neprilysinby Intracellular Domains of �APP and APLP
Raphaëlle Pardossi-Piquard,1 Agnès Petit,1
Toshitaka Kawarai,2 Claire Sunyach,1
Cristine Alves da Costa,1 Bruno Vincent,1
Sabine Ring,3 Luciano D’Adamio,4,5 Jie Shen,6
Ulrike Müller,3 Peter St. George Hyslop,2
and Frédéric Checler1,*1Institut de Pharmacologie Moléculaire et CellulaireCentre National de la Recherche ScientifiqueUMR6097 CNRS/UNSAValbonne 06560France2Centre for Research in Neurodegenerative DiseasesDepartment of MedicineUniversity of Toronto and University Health NetworkToronto Western Hospital Research Institute6 Queen’s Park CrescentToronto, Ontario M5S 3H2Canada3 Institute for Pharmacy and Molecular BiotechnologyUniversity of Heidelberg69120 HeidelbergGermany4Albert Einstein College of MedicineNew York, New York5Dipartimento di Biochimica e Biotecnologie MedicheUniversita’ Degli Studi di Napoli Federico IINapoliItaly6Center for Neurologic DiseasesHarvard Medical SchoolBoston, Massachusetts
Summary
Amyloid �-peptide (A�), which plays a central role inAlzheimer’s disease, is generated by presenilin-dependent �-secretase cleavage of �-amyloid precur-sor protein (�APP). We report that the presenilins(PS1 and PS2) also regulate A� degradation. Preseni-lin-deficient cells fail to degrade A� and have drasticreductions in the transcription, expression, and activ-ity of neprilysin, a key A�-degrading enzyme. Nepri-lysin activity and expression are also lowered by�-secretase inhibitors and by PS1/PS2 deficiency inmouse brain. Neprilysin activity is restored by tran-sient expression of PS1 or PS2 and by expressionof the amyloid intracellular domain (AICD), which iscogenerated with A�, during �-secretase cleavage of�APP. Neprilysin gene promoters are transactivatedby AICDs from APP-like proteins (APP, APLP1, andAPLP2), but not by A� or by the �-secretase cleavageproducts of Notch, N- or E- cadherins. The presenilin-dependent regulation of neprilysin, mediated byAICDs, provides a physiological means to modulateA� levels with varying levels of �-secretase activity.
*Correspondence: [email protected]
Introduction
One of the two main histopathological hallmarks in Alz-heimer’s disease (AD) is the senile plaque, an extracel-lular protein deposit composed in part by fibrillar aggre-gates of amyloid β-peptides (Aβ) (Haass and Selkoe,1993). Aβ is a 40–42 amino acid peptide that is gener-ated from the β-Amyloid Precursor Protein (βAPP) bytwo sequential cleavages. The first of these cleavagesoccurs in the extracellular domain and is mediated bya membrane-bound aspartyl protease termed β-secre-tase (Vassar and Citron, 2000). The second set of cleav-ages occurs at residues 40–42 (termed γ-site) and atresidues 48–52 (termed �-site) within the transmem-brane domain of the βAPP stub generated by β-secre-tase. The γ-site cleavage generates Aβ, while the con-current �-site cleavage generates a cytosolic stubreferred to as ICD (Passer et al., 2000) or AICD (βAPPIntraCellular Domain). The exact role of AICD remainsunclear.
Both the γ- and the �-site cleavages are mediated bypresenilin (PS)-independent and dependent proteases(De Strooper et al., 1998; Armogida et al., 2001). Thepresenilin-dependent γ-secretase and �-site proteolyticactivities (which are often generically collectively termedγ-secretase) are dependent upon a multimeric complexof at least four different membrane proteins includingPresenilin 1(PS1) or Presenilin 2 (PS2), nicastrin, Aph-1,and Pen-2 (Yu et al., 2000; Francis et al., 2002). In thesecomplexes, the presenilins have been proposed as anovel type of transmembrane aspartyl protease bearingthe catalytic core of the γ-secretase (Wolfe et al., 1999).
This novel type of intramembranous proteolysis ap-parently governs the function of βAPP and several TypeI transmembrane proteins including Notch, cadherins,ErbB-4, CD44, or p75NTR. Many of these proteins areinvolved in a variety of vital cellular functions such asintracellular signaling in development and adulthood,cell adhesion, cell growth and proliferation, and kinaseactivities (for review see Sisodia and St. George-Hys-lop, 2002; Pollack and Lewis, 2005). Thus, γ-secretasecleavage of Notch releases an intracellular fragmentcalled NICD (Notch IntraCellular Domain), which actsas a transcription factor mediating signal transductionin the Notch-Delta pathway, a critical intercellular sig-naling mechanism, during both embryonic develop-ment and adulthood (Kopan et al., 1996; Shen et al.,1997; De Strooper et al., 1998; Kopan and Goate, 2000).
Under normal conditions, Aβ occurs as a solublefragment, the concentration of which is normally tightlycontrolled below the threshold for its self-aggregationinto β sheet fibrils (Burdick et al., 1992). Aβ is activelydegraded by several enzymes including neprilysin(NEP), insulin-degrading enzyme (IDE), and endothelin-converting enzyme (ECE) (Carson and Turner, 2002). Al-though these Aβ-degrading enzymes have been wellcharacterized, very little is known about the regulatorymechanisms that govern their expression and/or activ-ity. Nevertheless, under normal physiological circum-stances, the balance between the rates of production
Neuron542
and clearance of Aβ is likely to be delicately regulated, nobreaking down only in circumstances that lead to theronset of Alzheimer’s disease. We show here that al-ithough γ-secretase cleavage produces Aβ, the otherdproduct of γ-secretase cleavage (AICD) specificallylupregulates the transcription of NEP, which in turn, ac-ncelerates the degradation of Aβ. This transcriptionalcsignaling pathway therefore provides a simple and ele-tgant physiological mechanism for the regulation of Aβ3levels following physiological activation of γ-secretase7cleavage of βAPP.0l
Results 27
Neprilysin Expression and Activity Are Reduced 0in Cells Devoid of Presenilins sAβ40 immunoreactivity decreases in a time-dependent imanner upon exposure of exogenous Aβ40 peptide to 5wild-type fibroblasts and blastocysts (PS+/+ and BD6, bFigure 1A). This decrease could be blocked by phos- kphoramidon (Suda et al., 1973), a specific inhibitor of nneprilysin (not shown). However, we observed that bAβ40 was not efficiently degraded by PS-deficient fi- 1broblasts or by PS-deficient blastocysts (PS−/− and
bBD8, Figure 1A). These results raise the possibility that
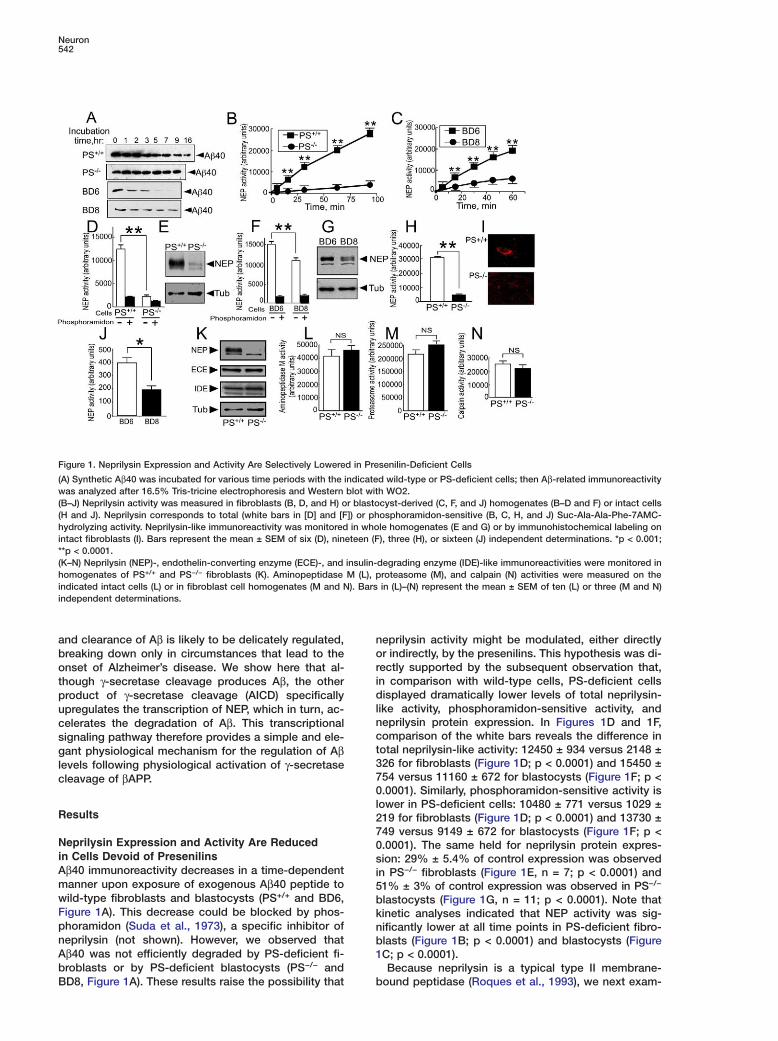
Figure 1. Neprilysin Expression and Activity Are Selectively Lowered in Presenilin-Deficient Cells
(A) Synthetic Aβ40 was incubated for various time periods with the indicated wild-type or PS-deficient cells; then Aβ-related immunoreactivitywas analyzed after 16.5% Tris-tricine electrophoresis and Western blot with WO2.(B–J) Neprilysin activity was measured in fibroblasts (B, D, and H) or blastocyst-derived (C, F, and J) homogenates (B–D and F) or intact cells(H and J). Neprilysin corresponds to total (white bars in [D] and [F]) or phosphoramidon-sensitive (B, C, H, and J) Suc-Ala-Ala-Phe-7AMC-hydrolyzing activity. Neprilysin-like immunoreactivity was monitored in whole homogenates (E and G) or by immunohistochemical labeling onintact fibroblasts (I). Bars represent the mean ± SEM of six (D), nineteen (F), three (H), or sixteen (J) independent determinations. *p < 0.001;**p < 0.0001.(K–N) Neprilysin (NEP)-, endothelin-converting enzyme (ECE)-, and insulin-degrading enzyme (IDE)-like immunoreactivities were monitored inhomogenates of PS+/+ and PS−/− fibroblasts (K). Aminopeptidase M (L), proteasome (M), and calpain (N) activities were measured on theindicated intact cells (L) or in fibroblast cell homogenates (M and N). Bars in (L)–(N) represent the mean ± SEM of ten (L) or three (M and N)independent determinations.
eprilysin activity might be modulated, either directlyr indirectly, by the presenilins. This hypothesis was di-ectly supported by the subsequent observation that,n comparison with wild-type cells, PS-deficient cellsisplayed dramatically lower levels of total neprilysin-
ike activity, phosphoramidon-sensitive activity, andeprilysin protein expression. In Figures 1D and 1F,omparison of the white bars reveals the difference inotal neprilysin-like activity: 12450 ± 934 versus 2148 ±26 for fibroblasts (Figure 1D; p < 0.0001) and 15450 ±54 versus 11160 ± 672 for blastocysts (Figure 1F; p <.0001). Similarly, phosphoramidon-sensitive activity is
ower in PS-deficient cells: 10480 ± 771 versus 1029 ±19 for fibroblasts (Figure 1D; p < 0.0001) and 13730 ±49 versus 9149 ± 672 for blastocysts (Figure 1F; p <.0001). The same held for neprilysin protein expres-ion: 29% ± 5.4% of control expression was observedn PS−/− fibroblasts (Figure 1E, n = 7; p < 0.0001) and1% ± 3% of control expression was observed in PS−/−
lastocysts (Figure 1G, n = 11; p < 0.0001). Note thatinetic analyses indicated that NEP activity was sig-ificantly lower at all time points in PS-deficient fibro-lasts (Figure 1B; p < 0.0001) and blastocysts (FigureC; p < 0.0001).Because neprilysin is a typical type II membrane-
ound peptidase (Roques et al., 1993), we next exam-

Presenilins Regulate Neprilysin Transcription543
ined neprilysin activity on the surface of intact cells,using a cell-impermeable fluorimetric substrate. In thisassay, the substrate is cleaved only by enzymes thatare present at the cell surface with their catalytic sitesfacing the extracellular space. In agreement with thestudies on whole-cell lysates described above, PS-deficient fibroblasts exhibited a significant 80% reduc-tion of cell membrane neprilysin activity compared tothat in wild-type fibroblasts (31130 ± 582 versus 4609± 359, Figure 1H; p<0.0001). Furthermore, neprilysin im-munoreactivity was poorly detectable at the surface ofintact PS-deficient fibroblasts, although it was readilydetectable on the surface of wild-type fibroblasts (Fig-ure 1I). A similar reduction in cell membrane neprilysinactivity was also observed in PS-deficient blastocysts(BD8), but not in wild-type blastocysts (BD6) (380.7 ±34 versus 199 ± 25, Figure 1J; p < 0.001).
Presenilin Deficiency Selectively Affects NeprilysinTo assess whether PS deficiency specifically alteredneprilysin activity, or whether it also affected other pu-tative Aβ-degrading activities or proteases, we mea-sured the expression of endothelin-converting enzymeand insulin-degrading enzyme (Figure 1K) and theactivities of aminopeptidase M, another ectoenzyme(Figure 1L) (Checler, 1993), proteasome (Figure 1M),and calpain (Figure 1N). In sharp contrast to the effectsof PS-deficiency on neprilysin, none of these other en-zymes were affected by the absence of PS1 and PS2.
Presenilin 1 and Presenilin 2 AffectNeprilysin TranscriptionThe presenilins directly interact with several unrelatedproteins such as nicastrin, Aph-1, and Pen-2, and manyof these proteins are destabilized by the absence of thepresenilins (for review, see De Strooper, 2003). How-ever, six lines of evidence indicate that the reductionsin neprilysin in PS-deficient cells are not due to the lossof a direct, stabilizing interaction between the preseni-lin and neprilysin proteins, but rather arise from a re-duction of neprilysin transcription. First, anti-NEP im-munoprecipitates from wild-type fibroblasts do notcontain PS1 (Figure 2A) or PS2 (not shown). Second,PS1 and PS2 immunoprecipitation does not deplete su-pernatants of NEP activity (not shown). Third, residualNEP expression in PS−/− fibroblasts partitioned withincell compartments with the same distribution as inwild-type fibroblasts (Figure 2B), suggesting that PSdeficiency did not alter trafficking of NEP in PS−/− fibro-blasts. Fourth, neprilysin stability is not affected by PSdeficiency (Figure 2C). Fifth, both neprilysin expressionand cell surface neprilysin activity can be fully restoredby transfection of neprilysin cDNA into PS-deficient fi-broblasts (Figure 2D) and into PS-deficient blastocysts(120% ± 4.6% above control mock-transfected cells;p < 0.01; data not shown). Finally, quantitative RT-PCRanalyses revealed an approximately 80% reduction inneprilysin mRNA (Figure 3A) (note that overexposure ofthe gel [lower panel] shows residual mRNA expression).
Taken together, these data suggest that the loss ofneprilysin expression in PS-deficient cells arises fromimpairment in neprilysin transcription. Intriguingly, nep-rilysin transcription, protein expression, and enzymatic
Figure 2. Presenilin Deficiency Does Not Affect Neprilysin at a Post-transcriptional Level
(A) NEP was immunoprecipitated from PS+/+ and PS−/− fibroblasthomogenates. Immunological complexes were analyzed for theirendogenous NEP and PS1-like immunoreactivities. Note that PS1-like immunoreactivity corresponds to the C-terminal maturationproduct of PS1 (CTF-PS1; Checler, 1999).(B) Endogenous cell distribution of NEP was analyzed by sucrosedensity gradient. Note that NEP immunoreactivity is drasticallylower in PS−/− fibroblasts (while control β-actin is identical) andthat residual NEP behaves as in PS+/+, i.e., like the ectoenzymeADAM10.(C) PS+/+ and PS−/− fibroblasts were treated with cycloheximide toprevent NEP neosynthesis as described in the Experimental Pro-cedures. At the indicated times, NEP activity was fluorimetricallyrecorded. Note the identical slopes (−1.65 ± 0.22 [PS+/+] and−1.35 ± 0.31 [PS−/−]), indicating a PS-independent similar decay ofNEP (each point is the mean ± SEM of three independent determin-ations). Upper panel shows NEP immunoreactivity decrease inPS+/+ fibroblasts.(D) Forty-eight hours after transfection in PS+/+ and PS−/− fibro-blasts, NEP expression (upper panel) and activity (lower panel)were monitored as described in the Experimental Procedures. Barsrepresent the mean of two independent experiments carried outin duplicate.
activity were not affected in cells devoid of either PS1only or PS2 only. Thus, normal levels of neprilysinmRNA (Figure 3A), enzymatic activity (Figure 3B), andprotein expression (Figure 3C) were observed in PS1−/−
fibroblasts (expressing only endogenous PS2) and inPS2−/− fibroblasts (expressing only endogenous PS1).Identical results were achieved when exogenous PS1or exogenous PS2 were transfected into PS-deficientfibroblasts. Thus, neprilysin mRNA expression, proteinexpression, and enzymatic activity were equivalentlyand fully restored in PS-deficient fibroblasts by tran-sient transfection of PS1 and PS2, PS1 only, or PS2only (Figures 3D and 3E). Control experiments indicatethat aminopeptidase M activity was not affected byPS1 or PS2 complementation in PS-deficient fibro-blasts (Figure 3F). These data were fully confirmed
Neuron544
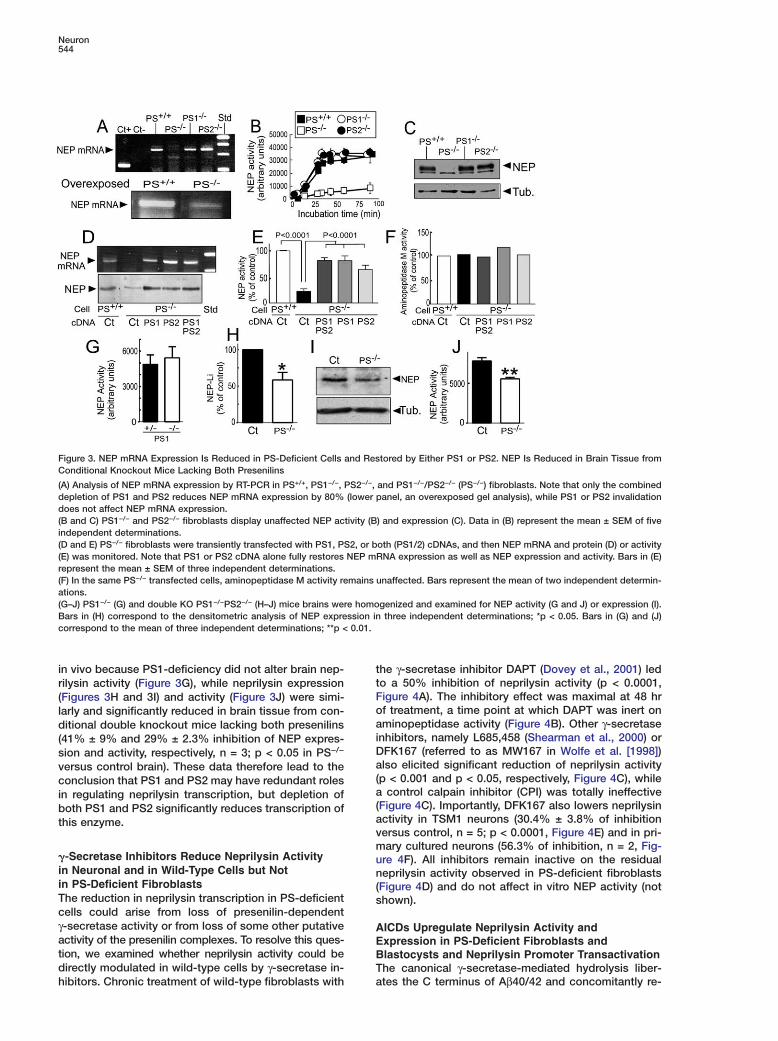
Figure 3. NEP mRNA Expression Is Reduced in PS-Deficient Cells and Restored by Either PS1 or PS2. NEP Is Reduced in Brain Tissue fromConditional Knockout Mice Lacking Both Presenilins
(A) Analysis of NEP mRNA expression by RT-PCR in PS+/+, PS1−/−, PS2−/−, and PS1−/−/PS2−/− (PS−/−) fibroblasts. Note that only the combineddepletion of PS1 and PS2 reduces NEP mRNA expression by 80% (lower panel, an overexposed gel analysis), while PS1 or PS2 invalidationdoes not affect NEP mRNA expression.(B and C) PS1−/− and PS2−/− fibroblasts display unaffected NEP activity (B) and expression (C). Data in (B) represent the mean ± SEM of fiveindependent determinations.(D and E) PS−/− fibroblasts were transiently transfected with PS1, PS2, or both (PS1/2) cDNAs, and then NEP mRNA and protein (D) or activity(E) was monitored. Note that PS1 or PS2 cDNA alone fully restores NEP mRNA expression as well as NEP expression and activity. Bars in (E)represent the mean ± SEM of three independent determinations.(F) In the same PS−/− transfected cells, aminopeptidase M activity remains unaffected. Bars represent the mean of two independent determin-ations.(G–J) PS1−/− (G) and double KO PS1−/−PS2−/− (H–J) mice brains were homogenized and examined for NEP activity (G and J) or expression (I).Bars in (H) correspond to the densitometric analysis of NEP expression in three independent determinations; *p < 0.05. Bars in (G) and (J)correspond to the mean of three independent determinations; **p < 0.01.
in vivo because PS1-deficiency did not alter brain nep- ttrilysin activity (Figure 3G), while neprilysin expressionF(Figures 3H and 3I) and activity (Figure 3J) were simi-olarly and significantly reduced in brain tissue from con-aditional double knockout mice lacking both presenilinsi(41% ± 9% and 29% ± 2.3% inhibition of NEP expres-Dsion and activity, respectively, n = 3; p < 0.05 in PS−/−
aversus control brain). These data therefore lead to the(conclusion that PS1 and PS2 may have redundant rolesain regulating neprilysin transcription, but depletion of(both PS1 and PS2 significantly reduces transcription ofathis enzyme.vm
�-Secretase Inhibitors Reduce Neprilysin Activity uin Neuronal and in Wild-Type Cells but Not nin PS-Deficient Fibroblasts (The reduction in neprilysin transcription in PS-deficient scells could arise from loss of presenilin-dependentγ-secretase activity or from loss of some other putative Aactivity of the presenilin complexes. To resolve this ques- Etion, we examined whether neprilysin activity could be Bdirectly modulated in wild-type cells by γ-secretase in- T
ahibitors. Chronic treatment of wild-type fibroblasts with
he γ-secretase inhibitor DAPT (Dovey et al., 2001) ledo a 50% inhibition of neprilysin activity (p < 0.0001,igure 4A). The inhibitory effect was maximal at 48 hrf treatment, a time point at which DAPT was inert onminopeptidase activity (Figure 4B). Other γ-secretase
nhibitors, namely L685,458 (Shearman et al., 2000) orFK167 (referred to as MW167 in Wolfe et al. [1998])lso elicited significant reduction of neprilysin activityp < 0.001 and p < 0.05, respectively, Figure 4C), while
control calpain inhibitor (CPI) was totally ineffectiveFigure 4C). Importantly, DFK167 also lowers neprilysinctivity in TSM1 neurons (30.4% ± 3.8% of inhibitionersus control, n = 5; p < 0.0001, Figure 4E) and in pri-ary cultured neurons (56.3% of inhibition, n = 2, Fig-
re 4F). All inhibitors remain inactive on the residualeprilysin activity observed in PS-deficient fibroblasts
Figure 4D) and do not affect in vitro NEP activity (nothown).
ICDs Upregulate Neprilysin Activity andxpression in PS-Deficient Fibroblasts andlastocysts and Neprilysin Promoter Transactivationhe canonical γ-secretase-mediated hydrolysis liber-tes the C terminus of Aβ40/42 and concomitantly re-

Presenilins Regulate Neprilysin Transcription545
Figure 4. Effect of γ-Secretase Inhibitors and γ-Secretase-Derived βAPP Fragments on NEP Activity and Promoter Transactivation
(A–F) Wild-type (PS+/+) fibroblasts were chronically treated by successive additions of DAPT for a total time period of 8, 24, and 48 hr (seeExperimental Procedures) (A) or for 48 hr (B) with 2 �M of DAPT, and then NEP (A) or aminopeptidase M (B) activities were fluorimetricallyrecorded on intact cells. Error bars represent the mean ± SEM of three to six independent determinations. PS+/+ (C), PS−/− fibroblasts (D),TSM1 neurons (E), and primary cultured neurons (F) were treated with the indicated inhibitor (DAPT, 48 hr, 2 �M; L685,458, 8 hr, 1 �M;DFK167, 8 hr, 100 �M; calpain inhibitor [CPI], 8 hr, 100 �M), and then the NEP activity of intact cells was measured. Error bars represent themean ± SEM of three to five independent determinations; ***p < 0.0001.(G–K) PS-deficient (PS−/−) fibroblasts or PS-deficient (BD8) blastocyst-derived cells (G) were treated for various time periods with 10 ng/ml ofAβ42, and then NEP activity was fluorimetrically assayed on intact cells. Error bars represent the mean ± SEM of three experiments. PS−/−
fibroblasts (H and J) or BD8 cells (I and K) were transiently transfected with empty vector or with the indicated AICD cDNA , and then NEPactivity (H and I) or expression (J and K) was monitored. Bars in (H) and (I) represent the mean ± SEM of five independent experiments.(L and M) The indicated AICD was cotransfected in fibroblasts (L) or in TSM1 neurons (M) with β-gal cDNA and either renal NEP promotersrNEPP1 and rNEPP2 (L) or neuronal NEP promoter nNEPP (M), and then β-galactosidase and luciferase activities were monitored.(N) HEK293 cells were transfected with AICDC59, Fe65, and Tip60 cDNA , and then nuclear extracts and binding experiments with the 219bp probe derived from rNEPP were carried out as described in the Experimental Procedures. Lane 1, labeled probe + nuclear extract; lane 2,labeled probe + nuclear extract + Anti-myc; lane 3, labeled probe + nuclear extract + unrelated antibody.
leases the 59 amino acid stub composed of the cyto-plasmic C-terminal tail of βAPP referred to as AICDC59(see Introduction and Figure 8). An additional prese-nilin-dependent proteolytic cleavage of βAPP andNotch occurs several amino acids downstream (re-ferred to as � cleavage). This � cleavage event liberatesAICDC50 from APP and a Notch IntraCellular Domain(NICD) from Notch (Gu et al., 2001; Sastre et al., 2001;Weidemann et al., 2002). Because NICD is known tomodulate the transcription of several genes, we rea-soned that neprilysin transcription might be modulatedby one of the γ/�-secretase-derived products. Exoge-nous Aβ42 did not modify neprilysin activity in PS-defi-cient fibroblasts or in PS-deficient blastocysts (Figure4G). However, transient transfections of AICDC50 orAICDC59 cDNAs increased both neprilysin activity (Fig-ures 4H and 4I) and neprilysin expression (Figures 4Jand 4K) in PS-deficient fibroblasts and blastocysts.
In order to link our observation of AICD-induced in-crease of neprilysin activity and expression to our ob-servation of PS-dependent neprilysin mRNA upregula-tion, we examined the effect of AICDC50 and AICDC59on neprilysin promoter transactivation, using various
neprilysin promoter elements upstream of a luciferasereporter minigene. Both AICDs dramatically increasedthe transactivation of two renal neprilysin promoters,rNEPP1 (−385 bp to +147 bp) and rNEPP2 (−263 bpto +145 bp, Figure 4L) in fibroblasts. To confirm AICD-induced transactivation of the neprilysin promoter in aneural cell line, we also cloned the neuronal neprilysinpromoter and repeated the luciferase reporter assay. Inagreement with the above observations, AICDC50 alsotransactivated neprilysin promoter in TSM1 neurons(Figure 4M). Supergel shift assay analysis demon-strated that the AICD-potentiated transactivation of theneprilysin promoter indeed appears to be mediated bya direct physical interaction of AICD with the neprilysinpromoter (Figure 4N).
AICD-Induced Complementation of NeprilysinActivity Is Potentiated by Fe65 and Tip60in Fibroblasts and in HEK293 CellsSeveral lines of evidence have indicated that the adap-tor protein Fe65 modulates the stability of AICD (Kim-berly et al., 2001; Kinoshita et al., 2002), thereby po-tentiating its subsequent nuclear translocation and
Neuron546
interaction with the histone acetyltransferase Tip60 TF(Cao and Südhof, 2001). We therefore examined whether
Fe65 and Tip60 could (1) influence neprilysin by modu- ralating endogenous AICD in wild-type fibroblasts and in
HEK293 cells and (2) potentiate AICD-induced increase tiin neprilysin activity.
Fe65 and Tip60 transfection enhanced neprilysin ex- Hwpression and activity in PS+/+ fibroblasts (Figure 5A [up-
per panel] and Figure 5C; p < 0.0005) and in HEK293 picells (Figure 5A [lower panel]), but not in PS-deficient
cells (Figures 5B and 5C), indicating that Fe65 and isTip60 augment neprilysin expression through functional
interaction with an endogenous PS-dependent prod- i2uct. In wild-type PS+/+ fibroblasts, the cotransfection of
Fe65 and Tip60 with either AICDC50 or AICDC59 cDNA ifincreased neprilysin expression when compared to
AICDs cDNA transfection alone (Figure 5A). This was pcaccompanied by an augmentation of AICDC50 and
AICDC59 immunoreactivities (Figure 5A [lower panel])Xand by a clear translocation of AICDC59 (Figure 5D)
and AICDC50 (not shown) into the nuclei of HEK293 w1cells and PS+/+ fibroblasts (not shown). Similar potenti-
ation of neprilysin expression by Fe65 and Tip60 trans- Hrfection was also observed in AICD-transfected PS-defi-
cient fibroblasts (136% ± 6%, AICDC50+Fe65+Tip60 Xversus AICDC50 alone and 135% ± 6.2%, AICDC59+Fe65+Tip60 versus AICDC59 alone; p < 0.0005). Inter- �
testingly, AICDC50 and AICDC59 increase renal nepri-lysin promoter transactivation in HEK293 cells (not T
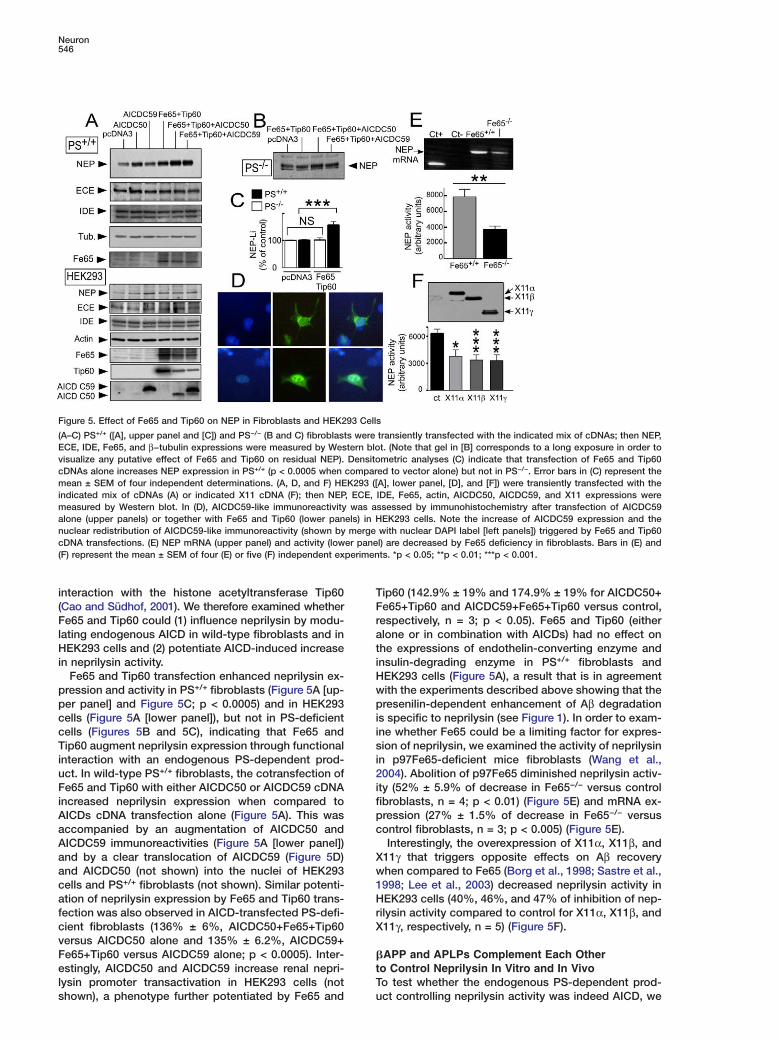
Figure 5. Effect of Fe65 and Tip60 on NEP in Fibroblasts and HEK293 Cells
(A–C) PS+/+ ([A], upper panel and [C]) and PS−/− (B and C) fibroblasts were transiently transfected with the indicated mix of cDNAs; then NEP,ECE, IDE, Fe65, and β−tubulin expressions were measured by Western blot. (Note that gel in [B] corresponds to a long exposure in order tovisualize any putative effect of Fe65 and Tip60 on residual NEP). Densitometric analyses (C) indicate that transfection of Fe65 and Tip60cDNAs alone increases NEP expression in PS+/+ (p < 0.0005 when compared to vector alone) but not in PS−/−. Error bars in (C) represent themean ± SEM of four independent determinations. (A, D, and F) HEK293 ([A], lower panel, [D], and [F]) were transiently transfected with theindicated mix of cDNAs (A) or indicated X11 cDNA (F); then NEP, ECE, IDE, Fe65, actin, AICDC50, AICDC59, and X11 expressions weremeasured by Western blot. In (D), AICDC59-like immunoreactivity was assessed by immunohistochemistry after transfection of AICDC59alone (upper panels) or together with Fe65 and Tip60 (lower panels) in HEK293 cells. Note the increase of AICDC59 expression and thenuclear redistribution of AICDC59-like immunoreactivity (shown by merge with nuclear DAPI label [left panels]) triggered by Fe65 and Tip60cDNA transfections. (E) NEP mRNA (upper panel) and activity (lower panel) are decreased by Fe65 deficiency in fibroblasts. Bars in (E) and(F) represent the mean ± SEM of four (E) or five (F) independent experiments. *p < 0.05; **p < 0.01; ***p < 0.001.
shown), a phenotype further potentiated by Fe65 and
ip60 (142.9% ± 19% and 174.9% ± 19% for AICDC50+e65+Tip60 and AICDC59+Fe65+Tip60 versus control,espectively, n = 3; p < 0.05). Fe65 and Tip60 (eitherlone or in combination with AICDs) had no effect onhe expressions of endothelin-converting enzyme andnsulin-degrading enzyme in PS+/+ fibroblasts andEK293 cells (Figure 5A), a result that is in agreementith the experiments described above showing that theresenilin-dependent enhancement of Aβ degradation
s specific to neprilysin (see Figure 1). In order to exam-ne whether Fe65 could be a limiting factor for expres-ion of neprilysin, we examined the activity of neprilysin
n p97Fe65-deficient mice fibroblasts (Wang et al.,004). Abolition of p97Fe65 diminished neprilysin activ-
ty (52% ± 5.9% of decrease in Fe65−/− versus controlibroblasts, n = 4; p < 0.01) (Figure 5E) and mRNA ex-ression (27% ± 1.5% of decrease in Fe65−/− versusontrol fibroblasts, n = 3; p < 0.005) (Figure 5E).Interestingly, the overexpression of X11α, X11β, and
11γ that triggers opposite effects on Aβ recoveryhen compared to Fe65 (Borg et al., 1998; Sastre et al.,998; Lee et al., 2003) decreased neprilysin activity inEK293 cells (40%, 46%, and 47% of inhibition of nep-
ilysin activity compared to control for X11α, X11β, and11γ, respectively, n = 5) (Figure 5F).
APP and APLPs Complement Each Othero Control Neprilysin In Vitro and In Vivoo test whether the endogenous PS-dependent prod-
uct controlling neprilysin activity was indeed AICD, we
Presenilins Regulate Neprilysin Transcription547
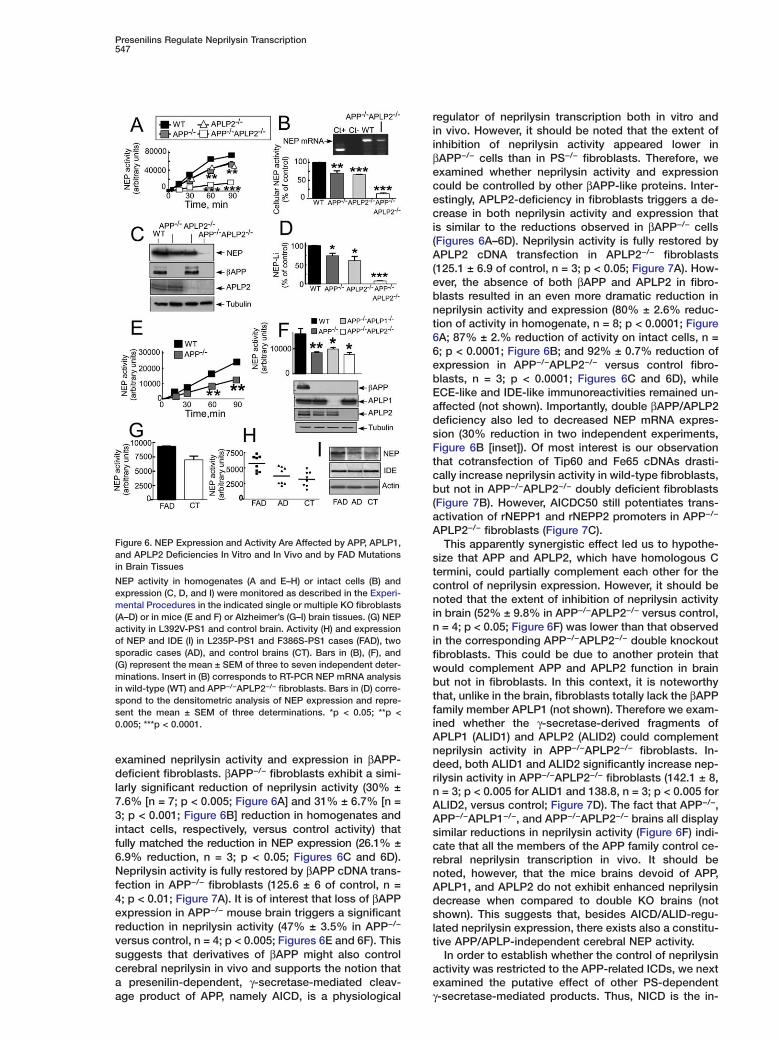
examined neprilysin activity and expression in βAPP-deficient fibroblasts. βAPP−/− fibroblasts exhibit a simi-larly significant reduction of neprilysin activity (30% ±7.6% [n = 7; p < 0.005; Figure 6A] and 31% ± 6.7% [n =3; p < 0.001; Figure 6B] reduction in homogenates andintact cells, respectively, versus control activity) thatfully matched the reduction in NEP expression (26.1% ±6.9% reduction, n = 3; p < 0.05; Figures 6C and 6D).Neprilysin activity is fully restored by βAPP cDNA trans-fection in APP−/− fibroblasts (125.6 ± 6 of control, n =4; p < 0.01; Figure 7A). It is of interest that loss of βAPPexpression in APP−/− mouse brain triggers a significantreduction in neprilysin activity (47% ± 3.5% in APP−/−
versus control, n = 4; p < 0.005; Figures 6E and 6F). Thissuggests that derivatives of βAPP might also controlcerebral neprilysin in vivo and supports the notion thata presenilin-dependent, γ-secretase-mediated cleav-
Figure 6. NEP Expression and Activity Are Affected by APP, APLP1,and APLP2 Deficiencies In Vitro and In Vivo and by FAD Mutationsin Brain Tissues
NEP activity in homogenates (A and E–H) or intact cells (B) andexpression (C, D, and I) were monitored as described in the Experi-mental Procedures in the indicated single or multiple KO fibroblasts(A–D) or in mice (E and F) or Alzheimer’s (G–I) brain tissues. (G) NEPactivity in L392V-PS1 and control brain. Activity (H) and expressionof NEP and IDE (I) in L235P-PS1 and F386S-PS1 cases (FAD), twosporadic cases (AD), and control brains (CT). Bars in (B), (F), and(G) represent the mean ± SEM of three to seven independent deter-minations. Insert in (B) corresponds to RT-PCR NEP mRNA analysisin wild-type (WT) and APP−/−APLP2−/− fibroblasts. Bars in (D) corre-spond to the densitometric analysis of NEP expression and repre-sent the mean ± SEM of three determinations. *p < 0.05; **p <0.005; ***p < 0.0001.
age product of APP, namely AICD, is a physiological
regulator of neprilysin transcription both in vitro andin vivo. However, it should be noted that the extent ofinhibition of neprilysin activity appeared lower inβAPP−/− cells than in PS−/− fibroblasts. Therefore, weexamined whether neprilysin activity and expressioncould be controlled by other βAPP-like proteins. Inter-estingly, APLP2-deficiency in fibroblasts triggers a de-crease in both neprilysin activity and expression thatis similar to the reductions observed in βAPP−/− cells(Figures 6A–6D). Neprilysin activity is fully restored byAPLP2 cDNA transfection in APLP2−/− fibroblasts(125.1 ± 6.9 of control, n = 3; p < 0.05; Figure 7A). How-ever, the absence of both βAPP and APLP2 in fibro-blasts resulted in an even more dramatic reduction inneprilysin activity and expression (80% ± 2.6% reduc-tion of activity in homogenate, n = 8; p < 0.0001; Figure6A; 87% ± 2.% reduction of activity on intact cells, n =6; p < 0.0001; Figure 6B; and 92% ± 0.7% reduction ofexpression in APP−/−APLP2−/− versus control fibro-blasts, n = 3; p < 0.0001; Figures 6C and 6D), whileECE-like and IDE-like immunoreactivities remained un-affected (not shown). Importantly, double βAPP/APLP2deficiency also led to decreased NEP mRNA expres-sion (30% reduction in two independent experiments,Figure 6B [inset]). Of most interest is our observationthat cotransfection of Tip60 and Fe65 cDNAs drasti-cally increase neprilysin activity in wild-type fibroblasts,but not in APP−/−APLP2−/− doubly deficient fibroblasts(Figure 7B). However, AICDC50 still potentiates trans-activation of rNEPP1 and rNEPP2 promoters in APP−/−
APLP2−/− fibroblasts (Figure 7C).This apparently synergistic effect led us to hypothe-
size that APP and APLP2, which have homologous Ctermini, could partially complement each other for thecontrol of neprilysin expression. However, it should benoted that the extent of inhibition of neprilysin activityin brain (52% ± 9.8% in APP−/−APLP2−/− versus control,n = 4; p < 0.05; Figure 6F) was lower than that observedin the corresponding APP−/−APLP2−/− double knockoutfibroblasts. This could be due to another protein thatwould complement APP and APLP2 function in brainbut not in fibroblasts. In this context, it is noteworthythat, unlike in the brain, fibroblasts totally lack the βAPPfamily member APLP1 (not shown). Therefore we exam-ined whether the γ-secretase-derived fragments ofAPLP1 (ALID1) and APLP2 (ALID2) could complementneprilysin activity in APP−/−APLP2−/− fibroblasts. In-deed, both ALID1 and ALID2 significantly increase nep-rilysin activity in APP−/−APLP2−/− fibroblasts (142.1 ± 8,n = 3; p < 0.005 for ALID1 and 138.8, n = 3; p < 0.005 forALID2, versus control; Figure 7D). The fact that APP−/−,APP−/−APLP1−/−, and APP−/−APLP2−/− brains all displaysimilar reductions in neprilysin activity (Figure 6F) indi-cate that all the members of the APP family control ce-rebral neprilysin transcription in vivo. It should benoted, however, that the mice brains devoid of APP,APLP1, and APLP2 do not exhibit enhanced neprilysindecrease when compared to double KO brains (notshown). This suggests that, besides AICD/ALID-regu-lated neprilysin expression, there exists also a constitu-tive APP/APLP-independent cerebral NEP activity.
In order to establish whether the control of neprilysinactivity was restricted to the APP-related ICDs, we nextexamined the putative effect of other PS-dependent
γ-secretase-mediated products. Thus, NICD is the in-
Neuron548
Figure 7. Neprilysin Activity Is Modulated by ALID1 and ALID2 but Not by NICD or E-Cad/CTF2
APP−/− (A), APLP2−/− (A), or APP−/−APLP2−/− (B–D) fibroblasts were transfected with empty pcDNA3 vector or indicated cDNAs; then NEPactivity (A, B, and D) or transactivation of rNEPP1 and rNEPP2 promoters (C) were monitored as described in the Experimental Procedures.Error bars in (A), (B), and (D) represent the mean ± SEM of three to four independent experiments. *p < 0.05; **p < 0.005. (E–G) The indicatedfibroblasts were transiently cotransfected with 4XCBF-luciferase and β-gal cDNAs in combination with either empty vector (Ct) or cDNAsencoding myc-tagged m�ENotch or myc-tagged NICD, and then m�ENotch and NICD (E) or NEP (F) expressions were assayed by Westernblot in the indicated cell lines, and luciferase (E) or NEP (G) activities were measured as described in the Experimental Procedures. Bars in(E) and (G) represent the mean ± SEM of three to four independent experiments. NICD expression was also estimated by immunohistochem-istry in PS+/+ and PS-deficient fibroblasts (H). Note a clear dense label of NICD in the nucleus. (I) E-Cad/CTF2 does not modulate NEPexpression and activity in HEK293 cells. Bars in (I) represent the mean ± SEM of seven independent experiments.
tracellular fragment liberated from Notch upon γ-secre- actase cleavage. To test whether NICD might also affect
neprilysin activity, we created fibroblast- and blasto- γIcyst-derived cell lines transiently expressing the 4XCBF-
luciferase reporter gene. In agreement with previouslypublished works (Herreman et al., 2000; Zhang et al., N
i2000), the transient cotransfection of m�ENotch (whichgenerates NICD upon γ-secretase cleavage) led to a ro- W
mbust transcriptional activation in wild-type fibroblasts(Figure 7E) and blastocyst-derived cells (not shown), s
ubut not in the equivalent PS-deficient cells (Figure 7E).However, transfection of NICD itself into wild-type and s
APS-deficient cells induced NICD protein expression(Figure 7E), favored nuclear localization of NICD (Figure b
i7H), and increased luciferase activity (Figure 7E). Thesecontrol experiments therefore established that NICD e
twas indeed functionally expressed in our experimentalsystem. However, regardless of the cell system used,m�ENotch and NICD were unable to restore neprilysin Dexpression (Figure 7F) or neprilysin activity (Figure 7G)in PS-deficient cells. Furthermore, the γ-secretase- S
lderived C-terminal products of E-Cadherin (E-Cad/CTF2, Figure 7I) (Marambaud et al., 2002) and N-Cad- 2
aherin (N-Cad/CTF2, not shown) (Marambaud et al.,2003) were also unable to affect neprilysin expression e
eand activity. Thus, unlike AICDs and ALIDs, NICD andE- and N-Cad/CTF2 did not complement neprilysin p
ctivity in PS−/− cells. Taken together, these data indi-ate that the complementation of neprilysin function by-secretase-derived products is specifically elicited byCDs of the βAPP family members.
eprilysin Expression and Activity Are Increasedn Alzheimer’s Brains of Genetic Origin
e have examined the neprilysin activity of control hu-an brains and compared them to brain samples of
poradic or genetic AD origin. Neprilysin activity (Fig-res 6G and 6H) and expression (Figure 6I) were nottatistically different between control (CT) and sporadicD brains, while Familial Alzheimer’s disease (FAD)rains harboring PS1 mutations displayed higher activ-
ty (Figures 6G and 6H) and expression (Figure 6I). Inter-stingly, insulin-degrading enzyme (IDE) was not al-ered in FAD brains (Figure 6I).
iscussion
everal lines of evidence strongly suggest that nepri-ysin (Hauss-Wegrzyniak and Wenk, 2002; Hama et al.,001; Marr et al., 2003; Leissring et al., 2003; Iwata etl., 2001), endothelin-converting enzyme (ECE; Eckmant al., 2003), and insulin-degrading enzyme (IDE; Farrist al., 2003) could participate in the catabolism of Aβeptides. We show here that neprilysin transcription,

Presenilins Regulate Neprilysin Transcription549
expression, and enzymatic activity are dramatically re-duced in non-neuronal and neuronal cells when theyare devoid of PS1 and PS2 and that this phenotype canbe mimicked by γ-secretase inhibitors. We have shownthat the presenilins control NEP activity at a transcrip-tional level and that PS deficiency does not affect theexpression of IDE, ECE, or other widely distributed in-tracellular (calpain and proteasome) or ectopeptidase(aminopeptidase M) activities. We have also shown thatneprilysin expression and activity are significantly re-duced in brain tissue from mice lacking PS1 and PS2(Saura et al., 2004), indicating that the presenilins alsocontrol cerebral neprilysin transcription, expression,and activity in vivo. This highly specific and physiologi-cal relationship between the presenilins and neprilysinstrongly argues for a major evolutionary and functionalrole of neprilysin in the physiological catabolism of Aβ,but does not preclude a role for other enzymes in Aβcatabolism.
The molecular mechanisms by which the presenilinsregulate neprilysin transcription would appear, from thedata presented here, to involve the C-terminal intracel-lular domains of APP and APLP proteins (AICDs andALIDs). Thus, we have clearly established that defi-ciency of βAPP, APLP1, or APLP2 drastically reducesneprilysin expression and activity in both fibroblastsand in brain tissues. We have specifically shown thatAβ42 and the γ-secretase-derived products of Notch,E-cadherins, and N-cadherins do not support this activ-ity. Furthermore, the Fe65 adaptor protein (which bindsto the GYENPTY motif in the cytoplasmic tail of βAPP/AICD [King and Turner, 2004]) and Tip60 clearly potenti-ate the AICD-induced upregulation of neprilysin in wild-type fibroblasts but not in PS−/− and APP−/−/APLP2−/−
cells.It has been previously reported that Fe65 and Tip60
stabilize AICD and favor its translocation to the nucleus(Kimberly et al., 2001; Kinoshita et al., 2002). Our exper-iments have shown that AICD translocates to the nu-cleus in the presence of Fe65 and Tip60 and transacti-vates neuronal and renal neprilysin promoters in TSM1neurons, HEK293 cells, and APP−/−/APLP2−/− fibro-blasts, respectively. Furthermore, Supergel shift analy-sis indicates that AICDC59 interacts physically with theneprilysin promoter in the presence of Fe65 and Tip60.Interestingly, X11, which clearly reduces γ-secretasecleavage of βAPP, also drastically reduces neprilysinactivity. Therefore, Fe65 and X11, which trigger oppo-site effects on Aβ production, also elicit an oppositephenotype in neprilysin regulation.
Altogether, the similar decrease in neprilysin activityand expression triggered by the absence of PS or APP/APLPs, the analogous absence of control of neprilysinby Fe65 and Tip60 in these invalidated fibroblasts, andthe opposite phenotype observed with Fe65 and X11all lead to the firm suggestion that the endogenousγ-secretase-dependent fragments controlling neprilysinare indeed AICD/ALIDs.
The redundant role of PS1 and PS2 in regulating nep-rilysin transcription contrasts with their overlapping butnot redundant roles in modulating Aβ peptide andNotch signaling. Thus, PS2 deficiency alone does notaffect γ-secretase-mediated production of Aβ or Notchin vivo (Herreman et al., 1999). However, PS1 deficiency
drastically reduces both Aβ production and Notch sig-naling (De Strooper et al., 1998). We hypothesize that inPS1−/− fibroblasts and brain tissue, APLP1 and APLP2could undergo γ-secretase-like cleavage by residualendogenous PS2, thus complementing AICD function,and thereby control neprilysin expression and activity.Because APLPs lack the Aβ sequence (although Aβ-likepeptides seem to be generated from APLP2), their PS2-dependent cleavage would not compensate for Aβ re-duction that is triggered by PS1 deficiency. In thiscontext, the resulting phenotype would be a reductionin Aβ production but with unaltered or weakly modifiedneprilysin levels. Alternatively, we hypothesize thatthere is likely to be both a constitutive (AICD-indepen-dent) basal expression of NEP and an inducible (AICD-dependent) expression of NEP, as suggested by resid-ual NEP activity in triple KO (APP−/−APLP1−/−APLP2−/−)mice brains (see Results). In PS1−/− fibroblasts and inPS1−/− brain tissue, this constitutive level of expression,together with the cumulative effects of residual AICDand ALIDs generated from APP, APLP1, and APLP2 bythe PS2 γ-secretase activity, could be sufficient tomaintain neprilysin expression and activity in PS1−/−
cells. In contrast, for Notch, there is both absence ofconstitutive basal expression of Notch-targeted genesand a lack of redundancy in alternate signaling mole-cules. Another possibility could be that NEP tran-scription is preserved in PS1−/− cells because APLPsγ-secretase cleavage is not affected to the same extentas APP by PS1 deficiency. However, to our knowledge,there is currently no data to support the notion thatPS1 and PS2 have differential/preferential effects onγ-secretase cleavage of APP, APLP1, and APLP2.
The experiments described here do more than simplyconfirm the previously and widely held suspicion thatAICD, like NICD, might act as a signaling molecule in-volved in transcriptional activation. The experimentshere depict an elegant and unusual mechanism bywhich Aβ levels are controlled. Thus, βAPP is proteolyti-cally processed by presenilin-dependent γ-secretase,which generates two distinct types of catabolites, Aβand AICD (Figure 8). One of these products, AICD, thencontrols the lifetime of the other product, Aβ, by selec-tively activating the transcription of an enzyme (nepri-lysin) capable of degrading that other product. Thus,γ-secretase activity directly controls Aβ production andthen indirectly modulates its degradation. To our knowl-edge, a similar “self-contained” mechanism for regulat-ing the degradation of enzyme products has not beendescribed previously; (NB: this does not precludeAICDs from also activating other genes).
If Aβ production and degradation are tightly linked,this raises the question of why Aβ accumulates in AD.The net accumulation of Aβ in AD pathology likely re-flects the cumulative effect of multiple events acting onproduction, fibrillogenesis, and degradation. In manyforms of AD, especially the late-onset sporadic forms,it has not been shown that there is increased β- andγ-secretase activity. In fact, some have suggested thatthese forms may reflect defective degradation of Aβ(Iwata et al., 2001; Leissring et al., 2003). Therefore,AICD levels are likely to be unchanged in these late-onset forms of AD, and as a result, the AICD-mediatedability to upregulate neprilysin activity would not be ef-

Neuron550
Figure 8. Model for PS-Dependent Transcriptional Activation of NEP by the βAPP-Intracellular Domain
βAPP undergoes proteolysis by various secretases. Aβ peptide is released by the sequential cleavages triggered by β- and γ-secretases inthe intracellular compartment from which it can be secreted. Once secreted, Aβ is degraded by the ectopeptidase named neprilysin (NEP), atype II integral protein with catalytic domain facing the extracellular space. Presenilin-dependent γ-secretase cleavage is triggered by amultiproteic complex composed of presenilin 1 (PS1) or presenilin 2 (PS2), nicastrin (NCT), Aph1, and Pen2. γ-secretase contributes to anadditional cleavage, referred to as � cleavage, that takes place slightly downstream from the γ-secretase site. The action at γ and � sites givesrise to AICDC59 and AICDC50, respectively. AICDs interact with Fe65 and Tip60 to form an active complex activating NEP mRNA transcription.Increased NEP activity, in turn, leads to increased Aβ degradation.
ficiently brought into play to protect the brain. In con- Aptrast, in those cases of AD arising from mutations in
APP and PS1, which activate γ-secretase and AICD csproduction, the principal effect is to produce longer Aβ
isoforms such as Aβ42. However, although Aβ40 is effi- aiciently degraded by NEP, Aβ42 is degraded by NEP
both in vitro and in vivo at a 6-fold lower rate, (Shirotani odet al., 2001). As a result, the upregulation of AICD (and
thus, NEP expression), which would be anticipated insubjects with presenilin mutations, would not com-
Epletely abolish the accumulation of Aβ42 in thesecases. It should be noted that, in agreement with the Cabove hypothesis, neprilysin expression and activity P
(were higher only in brain tissues with familial Alzheim-per’s disease linked to various presenilin-1 mutations,vwhile sporadic AD cases displayed neprilysin levelsZ
similar to those exhibited by normal brain tissues (Fig- Lures 6G–6I). Interestingly, PS1 mutations selectively af- bfect neprilysin and do not alter insulin-degrading en- a
lzyme expression.cThe above observations are also of direct practicalainterest because they indicate the possibility of new av-o
enues for controlling Aβ levels without directly affectingγ-secretase. This latter concept is important because ofthe various developmental and postnatal side-effects F
Nassociated with the inhibition of γ-secretase-mediatedwcleavage of other signaling molecules, including Notchp(Sisodia and St. George-Hyslop, 2002; Haass and Dea
Strooper, 1999). Our work now suggests that Aβ levels imight be modulated by directly increasing neprilysin p
wexpression, using AICD or small molecule mimics of
ICD. Upregulation of neprilysin by transgenic overex-ression, at least to modest levels, appears to be suffi-ient to reduce brain Aβ levels and to pose few toxicide effects (Leissring et al., 2003). This strategy wouldlso circumvent the other side effects of γ-secretase
nhibitors, including the potentially self-defeating effectf reducing AICD and thus preventing NEP-mediatedegradation of Aβ.
xperimental Procedures
ell Culture and Transfectionsrimary cultured neurons, HEK293 cells, telencephalon murine
TSM1) cell lines, blastocyst-derived cells, PS-, βAPP-, and97Fe65-deficient fibroblasts were obtained and cultured as pre-iously described (Vincent et al., 1996; De Strooper et al., 1999;hang et al., 2000; Herreman et al., 2000 Armogida et al., 2001;eissring et al., 2002; Wang et al., 2004). Mouse embryonic fibro-lasts derived from APLP2 or APP/APLP-deficient mouse embryosnd their littermates (Heber et al., 2000) were immortalized with the
arge T antigen of SV40. Several clonal lines were established andultured in DMEM/10% fetal calf serum containing 2 mM glutaminend 50 �M β-mercaptoethanol. Transient transfections were carriedut with DAC 30 (Eurogentec).
luorimetric Assays of Enzymatic ActivitiesEP activity was measured on intact cells or in cell homogenatesith Suc-Ala-Ala-Phe-7AMC in the absence or presence of phos-horamidon as described previously (Checler, 1993). When NEPctivity was measured in brain tissues, brains were homogenized
n Tris 10 mM (pH, 7.5) and centrifuged; then pellets were resus-ended in initial volumes of homogenization buffer, and activityas measured as above. Aminopeptidases B and M, calpain, and

Presenilins Regulate Neprilysin Transcription551
proteasome activities were measured as previously described(Checler, 1993).
Effect of A�42 and �-Secretase Inhibitors on NEP Activityon Intact CellsChronic treatment of wild-type PS+/+ fibroblasts with DAPT wasachieved by addition of 2 �M of the inhibitor at 0, 4, 8, 16, 20, 24,40, 44, and 48 hr, and NEP activity was measured on intact cells at8, 24, or 48 hr as described above. The effect of L685,458 (1 �M),DFK167 (100 �M), or Aβ42 (10 ng/ml) on NEP activity was alsoassessed, as indicated above, in intact wild-type fibroblasts, TSM1neurons, and primary cultured neurons.
Coimmunoprecipitation Experiments of EndogenousNEP and PS1Cells were harvested in lysis buffer (50 mM Tris-HCl [pH, 7.5], 150mM NaCl, 5 mM EDTA, 0.5% Triton X-100, and 0.5% Deoxycholate),and then 500 �g of protein was diluted in 1:2 RIPA 2X buffer andincubated overnight at 4°C with a 2000-fold dilution of anti-nepri-lysin 18B5 antibody together with protein A-sepharose. After cen-trifugation, pellets were resuspended in loading buffer containingSDS, submitted to 8% (NEP) or 12% (PS1) SDS-polyacrylamide gelelectrophoresis, and Western blot analysis. Membranes wereprobed with 18B5 or anti-PS1 antibody. Immunoreactive bandswere identified with anti-rabbit peroxidase (Immunotech) or anti-mouse peroxidase (Amersham Life Science) antibodies, followedby electrochemiluminescence (ECL) as previously described (Alvesda Costa et al., 2002) (Amersham Pharmacia Biotech).
Subcellular Fractionation by Sucrose GradientFour 100 mm diameter dishes of confluent PS+/+ and PS−/− fibro-blasts were homogenized with a dunce homogenizer in 0.25 Msucrose and 10 mM Tris-HCl (pH, 7.4) containing 1 mM MgAc2 anda protease inhibitor mixture (Sigma). Equal amounts of proteins (3.8mg) of each homogenate were loaded on top of a step gradient.After centrifugation, eleven 1 ml fractions were collected from thetop of each gradient. Proteins contained in 100 �l of each fractionwere precipitated with 5 volumes of methanol overnight at 4°C,resuspended in sample buffer, and heated for 5 min at 95°C. Nepri-lysin was assayed by Western blot as described below. The fullcharacterization of cell markers has been described elsewhere (Luoet al., 2003).
SDS/PAGE and Western Blot AnalysesNEP, β-tubulin, β actin, ECE, IDE, m�ENotch, NICD, βAPP, APLP1/2,X11α, β, and γ, Fe65, and Tip60 were separated on 8% Tris-glycinegels, while Aβ40, AICDC50, and AICDC59 were analyzed on 16.5%Tris-tricine gels. Proteins were transferred onto Hybond-C mem-branes and then probed with the following antibody: 18B5 (anti-neprilysin; Dr. G. Boileau), anti-mouse ECE 32 and anti-human ECE27 (Dr. K. Isobe), polyclonal anti-mouse IDE and anti-APLP1 andAPLP2 (EMD Biosciences), anti-human IDE (Dr. F. Authier), 9E10anti-myc antibody (myc-tagged m�ENotch, NICD, AICDC50, andAICDC59), monoclonal 22C11 (βAPP), polyclonal anti-Fe65 (Dr. L.Mercken), anti-HA (Tip60-HA), and WO2 antibody (Aβ40). Immuno-logical complexes were revealed with anti-rabbit peroxidase oranti-mouse peroxidase antibodies, followed by electrochemilum-inescence (Alves da Costa et al., 2002).
RT-PCR Analysis of NEP mRNATotal RNA was extracted and purified with the SV Total RNA Isola-tion System (Promega). For each RT-PCR reaction, 500 ng of RNAwas used. To amplify mouse neprilysin cDNA, the forward primerwas 5#-AGCCTCTCTGTGCTTGTCTTGC-3# and the backward primerwas 5#-CACTCATAGTAGCCT-CTGGAAGGG-3#, yielding a 614-bpproduct. RT-PCR reactions were performed with the Access RT-PCR System (Promega). The reverse transcription was done at48°C for 45 min, followed by a denaturation step at 94°C for 2 min.PCR reactions were performed at 94°C for 30 s, 55°C for 1 min,and 68°C for 2 min during 40 cycles, followed by a final extensionof 7 min at 68°C. RT-PCR products were analyzed on a 1% agarosegel stained with ethidium bromide.
Cloning of AICDC50 and AICDC59pcDNA4-AICDC59 was constructed by PCR amplification of the in-dicated coding sequence of βAPP from pcDNA4-βAPP (Chen et al.,2002) using the primers: 5#-TTTGGTGGAATTCATGATAGCGACAGTGATC GTCATCACC-3# and 5#-TTTACCTCG-AGCGTTCTGCATCTGCTCAAAG AAC-3#. The initial methionine was artificially intro-duced (underlined). The product was digested with EcoRI/XhoI andsubcloned into the EcoRI/XhoI site of pcDNA4 (Invitrogen) contain-ing the coding sequence for the Myc epitope at C terminus. Theconstruct was confirmed by sequencing. pcDNA4-AICDC50 wasconstructed using the primers 5#-TTTGGTGGAATTCATGGTGATGCTGAAGAAGAAA-CAGTAC-3# and 5#-TTTACCTCGAGCGTT CTGCATCTGCTCAAAGAAC-3#. The amplified product was ligated intothe pcDNA4 vector as described above.
Neuronal NEP Promoter Cloning and NEP-LuciferaseReporter ConstructsThe longest 5#-UTR for neprilysin is 98 bp, suggesting that the tran-scription start site is at least 4452 bp from the ATG site. To clonethe promoter regulatory elements, a 2500 bp DNA fragment up-stream of the presumed transcription start site was amplified fromhuman genomic DNA using the forward primer 5#-GCACTATAGCATTTTTAAAGG-3# and the reverse primer 5#-TGCTCCAGCCTGCTCTCGGTC-3# (NCBI accession number AC117384, positions 71,677–74,176, which corresponds to −6951 to –4452 from the ATG site).The fragment was inserted into the pCR2.1 vector (Invitrogen) andsubcloned between BamHI and XbaI restriction sites into pBlue-script vector (Stratagene) to introduce an additional XhoI site. NEPpromoter containing insert was then excised by XhoI digestion andreligated into pGL-2 vector (Promega). The subcloned fragmentwas sequenced and its direction and the absence of mutationalsequences were confirmed. Two renal NEP promoter constructsrNEPP1 (−385 bp to +147 bp) and rNEPP2 (−263 bp to +145 bp) inframe with luciferase have been previously reported (Ishimaru etal., 1997).
Measurements of NEP Promoter TransactivationNeuronal and renal neprilysin promoter-luciferase constructs werecotransfected with βgal reporter cDNA in fibroblasts, TSM1 neu-rons, and HEK293 cells in the absence or presence of AICDs cDNA.Forty-eight hours after transfection, luciferase and βgal activitieswere measured according to previously described procedures(Paitel et al., 2004).
Immunofluorescence AnalysisCells (grown on glass coverslips) were transfected with AICDs orNICD cDNAs. Forty eight hours after transfection, cells were fixedfor 20 min with 1.5% paraformaldehyde and permeabilized for 5min with 0.1% Triton X-100. After three washes, cells were incu-bated with the primary antibody for 4 hr at room temperature. Cellswere then washed three times with PBS and incubated for 1 hrat room temperature with the Alexa488-labeled goat anti-mouseantibody (Interchim). Coverslips were washed and mounted with orwithout DAPI. Staining was visualized with a Leica fluorescencemicroscope.
NICD Transcriptional Activity AssayFibroblasts were transiently cotransfected with 4XCBF-luciferasecDNA and empty vector (pCS2), myc-tagged m�ENotch-codingvectors, or NICD-coding vectors. A β-gal reporter cDNA was co-transfected to normalize data for transfection efficiency. Forty-eight hours after transfection, cells were rinsed, gently scraped inPBS (pH, 7.4), and spun for 5 min at 4000 rpm. Cells were thenhomogenized in 100 �l of lysis buffer, and luciferase activity wasmeasured by the Luciferase Assay System as described (Petit etal., 2001).
Gel Shift AssayElectrophoretic mobility shift assays (EMSA) were performed usinga commercial DNA binding-protein detection system (Promega,Charbonnières, France). In brief, PCR fragments covering three dis-tinct regions of the rNEPP1 promoter were obtained using the fol-lowing primer pairs 5#AAGCTTGACCGAGAGC3#/5#CGACACATCC

Neuron552
CGACC3# raising a 295 bp probe, 5#GGAACCTCCCCCAAGTCC3#/ JS5#CCTTTCTCCCTCAGC3# (219 bp probe), and 5#GGTCGGGATGTG
TCG3#/5#CAGTAGCGGCTCCTTCC3# (301 bp probe); then the frag- Jments were end labeled using [32P]ATP (6000 Ci/mmol, ICN Bio- Bmedicals, Orsay, France). For the preparation of nuclear extracts, RHEK293 cells were cultivated in 100 mm diameter dishes and tran- tsiently transfected with 12 �g cDNA of either empty pcDNA3 vector Cor a mix of myc-tagged-AICDC59 (6 �g), TIP60 (3 �g), and Fe65 (3
B�g) by means of DAC30 reactive as previously described. Forty-
Height hours after transfection, cells were harvested and nuclear ex-
btracts were prepared according to the Current Protocols in Molecu-
alar Biology (Ausubel et al., 2002). Binding reactions containing nuclear
Cextracts (10 �g) were performed at 37°C, using the shorter 219 bpoprobe according to the manufacturer’s directions. Then protein-1DNA complexes were resolved by electrophoresis through 7% na-
tive polyacrylamide gels in buffer containing 5mM Tris (pH, 8.3) and C38 mM Glycine for 3 hr at 300V. Gels were dried and autoradio- fgraphed on a BAS-1500 phosphorimager (Fujifilm, Tokyo, Japan). 8The specificity of the above described reactions were verified by CSupershift gel assay corresponding to the preincubation of nuclear iextracts with either anti-myc (specific) or anti-rabbit (nonspecific) gantibodies before allowing the binding reactions and subsequent Sincorporation of the labeled probe.
Ct
Normal and Pathological Human Brain TissuesCAll brain samples correspond to frontal cortices. Samples fromWRouen include a control brain (female, 74 years) and one FamilialiAD brain (female, 51 years, Leu392Val-PS1 mutation). Samples�from la Pitié Salpêtrière (Paris) correspond to two control brainsC(males, 72 and 55 years), two sporadic AD brains (females, 75 andD82 years), and two Familial AD brains (female, 37 years, Leu235Pro-gPS1 mutation; male, 44 years, Phe386Ser-PS1 mutation).
DStatistical Analysis GStatistical analyses were performed with PRISM Software (Graph- pPad Software, San Diego) by using the unpaired Student’s t test for tpairwise comparisons. D
Me
Acknowledgments m5
We sincerely thank Drs. B. De Strooper, P. Saftig, F. Chen, Q. Hu, DG. Martin, C. Dumanchin, D. Campion, N. Girardot, and C. Duyc- Pkaerts for providing cell lines and brain tissues. We are grateful to sDrs. T.C. Sudhof, R. Kopan, P. Marambaud, J.P. Borg, F. Authier, G. rBoileau, L. Mercken, and W. Araki for generously providing us with 1cDNAs and antibodies. This work was supported by the EU con-
Etract LSHM-CT-2003-503330 (APOPIS), by the Association FrancemAlzheimer (RPP) and by the Fondation pour la Recherche Médicalec(C.S.). Drs. P. St. George-Hyslop and Toshitaka Kawarai are sup-Cported by the Canadian Institutes of Health Research, HowardFHughes Medical Institute, and Alzheimer Society of Ontario. ThisFwork is dedicated to our friend and colleague Nicole Boyer, whoSdied suddenly on April 19, 2005.ad
Received: August 24, 2004FRevised: November 17, 2004JAccepted: April 12, 2005aPublished: May 18, 2005c8
ReferencesG(Alves da Costa, C., Paitel, E., Mattson, M.P., Amson, R., Telerman,sA., Ancolio, K., and Checler, F. (2002). Wild-type and mutated pre-Bsenilin-2 trigger p53-dependent apoptosis and down-regulate pre-Hsenilin-1 expression in HEK293 human cells and in murine neurons.eProc. Natl. Acad. Sci. USA 99, 4043–4048.
Armogida, M., Petit, A., Vincent, B., Scarzello, S., Alves da Costa, HpC., and Checler, F. (2001). Endogenous β-amyloid production in
presenilin-deficient embryonic mouse fibroblasts. Nat. Cell Biol. 3, 11030–1033. H
HAusubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman,
.G., Smith, J.A., and Struhl, K. (2002). DNA-protein interactions. Inhort Protocols in Molecular Biology, Fifth Edition, Vol. 2 (Boston:ohn Wiley and Sons), pp. 12.1.1–12.1.3.
org, J.-P., Yang, Y., De Taddeo-Borg, M., Margolis, B., and Turner,.S. (1998). The X11α protein slows cellular amyloid precursor pro-
ein processing and reduces Aβ40 and Aβ42 secretion. J. Biol.hem. 273, 14761–14766.
urdick, D., Soreghan, B., Kwon, M., Kosmoski, J., Knauer, M.,enschen, A., Yates, J., Cotman, C., and Glabe, C. (1992). Assem-ly and aggregation properties of synthetic Alzheimer’s A4/betamyloid peptide analogs. J. Biol. Chem. 267, 546–554.
ao, X., and Südhof, T.C. (2001). A transcriptively active complexf APP with Fe65 and histone acetyltransferase tip60. Science 293,15–120.
arson, J.A., and Turner, A.J. (2002). β-amyloid catabolism: rolesor neprilysin (NEP) and other metallopeptidases. J. Neurochem.1, 1–8.
hecler, F. (1993). Neuropeptide-degrading peptidases. In Methodsn Neurotransmitters and Neuropeptides Research. Part 2., T. Na-atsu, H. Parvez, M. Naoi, and S. Parvez, eds. (Amsterdam: Elseviercience Publishers), pp. 375–418.
hecler, F. (1999). Presenilins: Structural aspects and post-transla-ional events. Mol. Neurobiol. 19, 255–265.
hen, F., Gu, Y., Hasegawa, H., Ruan, X., Arawaka, S., Fraser, P.,estaway, D., Mount, H., and St. George-Hyslop, P. (2002). Presen-
lin 1 mutations activate γ-42-secretase but reciprocally inhibit-secretase cleavage of APP and S3-cleavage of Notch. J. Biol.hem. 277, 36521–36526.
e Strooper, B. (2003). Aph-1, Pen-2, and nicastrin with presenilinenerate an active γ-secretase complex. Neuron 38, 9–12.
e Strooper, B., Saftig, P., Craessaerts, K., Vanderstichele, H.,uhde, G., Von Figura, K., and Van Leuven, F. (1998). Deficiency ofresenilin 1 inhibits the normal cleavage of amyloid precursor pro-
ein. Nature 391, 387–390.
e Strooper, B., Annaert, W., Cupers, P., Saftig, P., Craessaerts, K.,umm, J.S., Schroeter, E.H., Schrijvers, V., Wolfe, M.S., Ray, W.J.,
t al. (1999). A presenilin-1-dependent γ-secretase-like proteaseediates release of Notch intracellular domain. Nature 398, 518–
22.
ovey, H.F., John, V., Anderson, J.P., Chen, L.Z., de Saint Andrieu,., Fang, L.Y., Freedman, S.B., Folmer, B., Goldbach, E., Holsztyn-ka, E.J., et al. (2001). Functional gamma-secretase inhibitorseduce beta-amyloid peptide levels in brain. J. Neurochem. 76,73–181.
ckman, E.A., Watson, M., Marlow, L., Sambamurti, K., and Eck-an, C.B. (2003). Alzheimer’s disease β-amyloid peptide is in-
reased in mice deficient in endothelin-converting enzyme. J. Biol.hem. 278, 2081–2084.
arris, W., Mansourian, S., Chang, Y., Lindsley, L., Eckman, E.A.,rosch, M.P., Eckman, C.B., Tanzi, R.E., Selkoe, D.J., and Guénette,. (2003). Insulin-degrading enzyme regulates the levels of insulin,myloid β-protein, and the β-amyloid precursor protein intracellularomain in vivo. Proc. Natl. Acad. Sci. USA 100, 4162–4167.
rancis, R., McGrath, G., Zhang, J., Ruddy, D.A., Sym, M., Apfeld,., Nicoll, M., Maxwell, M., Hai, B., Ellis, M.C., et al. (2002). aph-1nd pen-2 are required for notch pathway signaling, γ-secretaseleavage of βAPP, and presenilin protein accumulation. Dev. Cell 3,5–97.
u, Y., Misonou, H., Sato, T., Dohmae, N., Takio, K., and Ihara, Y.2001). Distinct intramembrane cleavages of the β-amyloid precur-or protein family resembling γ-secretase-like cleavage of Notch. J.iol. Chem. 276, 35235–35238.
aass, C., and De Strooper, B. (1999). The presenilins in Alzheim-r’s disease—proteolysis holds the key. Science 286, 916–919.
aass, C., and Selkoe, D.J. (1993). Cellular processing of β-amyloidrecursor protein and the genesis of amyloid β-peptide. Cell 75,039–1042.
ama, E., Shirotani, K., Masumoto, H., Sekine-Aizawa, Y., Aizawa,., and Saido, T.C. (2001). Clearance of extracellular and cell-asso-

Presenilins Regulate Neprilysin Transcription553
ciated amyloid β peptide through viral expression of neprilysin inprimary neurons. J. Biochem. (Tokyo) 130, 721–726.
Hauss-Wegrzyniak, B., and Wenk, G.L. (2002). Beta-amyloid depo-sition in the brains of rats chronically infused with thiorphan or lipo-polysaccharide: the role of ascorbic acid in the vehicle. Neurosci.Lett. 322, 75–78.
Heber, S., Herms, J., Gajic, V., Hainfellner, J.A., Aguzzi, A., Rülicke,T., Kretzschmar, H., von Koch, C., Sisodia, S.S., Tremml, P., et al.(2000). Mice with combined gene knock-outs reveal essential andpartially redundant functions of amyloid precursor protein familymembers. J. Neurosci. 20, 7951–7963.
Herreman, A., Hartmann, D., Annaert, W., Saftig, P., Craessaerts, K.,Serneels, L., Umans, L., Schrijvers, V., Checler, F., Vanderstichele,H., et al. (1999). Presenilin 2 deficiency causes a mild pulmonaryphenotype and no changes in amyloid precursor protein process-ing but enhances the embryonic lethal phenotype of presenilin 1deficiency. Proc. Natl. Acad. Sci. USA 96, 11872–11877.
Herreman, A., Serneels, L., Annaert, W., Collen, D., Schoonjans, L.,and De Strooper, B. (2000). Total inactivation of γ-secretase activityin presenilin-deficient embryonic stem cells. Nat. Cell Biol. 2, 461–462.
Ishimaru, F., Mari, B., and Shipp, M.A. (1997). The type 2 CD10/neutral endopeptidase 24.11 promoter: functional characterizationand tissue-specific regulation by CBF/NF-Y isoforms. Blood 89,4136–4145.
Iwata, N., Tsubuki, S., Takaki, Y., Shirotani, K., Lu, B., Gerard, N.P.,Gerard, C., Hama, E., Lee, H.-J., and Saido, T.C. (2001). Metabolicregulation of brain Aβ by neprilysin. Science 292, 1550–1552.
Kimberly, W.T., Zheng, J.B., Guénette, S., and Selkoe, D.J. (2001).The intracellular domain of the β-amyloid precursor protein is stabi-lized by Fe65 and translocates to the nucleus in a Notch-like man-ner. J. Biol. Chem. 276, 40288–40292.
King, G.D., and Turner, R.S. (2004). Adaptor protein interactions:modulators of amyloid precursor protein metabolism and Alzheim-er’s disease risk? Exp. Neurol. 185, 208–219.
Kinoshita, A., Whelan, C.M., Smith, C.J., Berezovska, O., and Hy-man, B.T. (2002). Direct visualization of the gamma secretase-gen-erated carboxy-terminal domain of the amyloid precursor protein:association with Fe65 and translocation to the nucleus. J. Neu-rochem. 82, 839–847.
Kopan, R., and Goate, A. (2000). A common enzyme connectsNotch signaling and Alzheimer’s disease. Genes Dev. 14, 2799–2806.
Kopan, R., Schroeter, E.H., Weintraub, H., and Nye, J.S. (1996). Sig-nal transduction by activated mNotch: importance of proteolyticprocessing and its regulation by the extracellular domain. Proc.Natl. Acad. Sci. USA 93, 1683–1688.
Lee, J.-H., Lau, K.-F., Perkinton, M.S., Standen, C.L., Shemilt,S.J.A., Mercken, L., Cooper, J.D., McLoughlin, D.M., and Miller,C.C.J. (2003). The neuronal adaptor protein X11α reduces Aβ levelsin the brains of Alzheimer’s APPswe Tg2576 transgenic mice. J.Biol. Chem. 278, 47025–47029.
Leissring, M.A., Murphy, M.P., Mead, T.R., Akbari, Y., Sugarman,M.C., Jannatipour, M., Anliker, B., Müller, U., Saftig, P., de Strooper,B., et al. (2002). A physiologic signaling role for the γ-secretase-derived intracellular fragment of APP. Proc. Natl. Acad. Sci. USA99, 4697–4702.
Leissring, M.A., Farris, W., Chang, A.Y., Walsh, D.M., Wu, X., Sun,X., Frosch, M.P., and Selkoe, D.J. (2003). Enhanced proteolysis ofβ-amyloid in APP transgenic mice prevents plaque formation,secondary pathology and premature death. Neuron 40, 1087–1093.
Luo, W.J., Wang, H., Li, H., Kim, B.S., Shah, S., Lee, H.-J., Thina-karan, G., Kim, T.-W., Yu, G., and Xu, H. (2003). PEN-2 and APH-1coordinately regulate proteolytic processing of presenilin 1. J. Biol.Chem. 278, 7850–7854.
Marambaud, P., Shioi, J., Serban, G., Georgakopoulos, A., Sarner,S., Nagy, V., Baki, L., Wen, P., Efthimiopoulos, S., Shao, Z., et al.(2002). A presenilin-1/γ-secretase cleavage releases the E-cadherinintracellular domain and regulates disassembly of adherens junc-tions. EMBO J. 21, 1948–1956.
Marambaud, P., Wen, P.H., Dutt, A., Takashima, A., Siman, R., andRobakis, N. (2003). A CPB binding transcriptional repressor pro-duced by the PS1/�-cleavage of N-cadherin is inhibited by PS1 FADmutations. Cell 114, 635–645.
Marr, R.A., Rockenstein, E., Mukherjee, A., Kindy, M.S., Hersh, L.B.,Gage, F.H., Verma, I.M., and Masliah, E. (2003). Neprilysin genetransfer reduces human amyloid pathology in transgenic mice. J.Neurosci. 23, 1992–1996.
Paitel, E., Sunyach, C., Alves da Costa, C., Bourdon, J.C., Vincent,B., and Checler, F. (2004). Primary cultured neurons devoid of cellu-lar prion display lower responsiveness to staurosporine through thecontrol of p53 at both transcriptional and post-transcriptionallevels. J. Biol. Chem. 279, 612–618.
Passer, B., Pellegrini, L., Russo, C., Siegel, R.M., Lenardo, M.J.,Schettini, G, Bachmann, M., Tabaton, M., and D’Adamio, L. (2000).Generation of an apoptotic intracellular peptide by γ-secretasecleavage of Alzheimer’s amyloid β protein precursor. J. AlzheimersDis. 2, 289–301.
Petit, A., Bihel, F., Alves da Costa, C., Pourquié, O., Kraus, J.L.,and Checler, F. (2001). New protease inhibitors prevent γ-secretase-mediated Aβ 40/42 production without affecting Notch cleavage.Nat. Cell Biol. 3, 507–511.
Pollack, S.J., and Lewis, H. (2005). γ-secretase inhibitors for Alz-heimer’s disease: challenges of a promiscuous protease.. Curr.Opin. Investig. Drugs 6, 35–47.
Roques, B.P., Noble, F., Dauge, V., Fournie-Zaluski, M.C., and Beau-mont, A. (1993). Neutral endopeptidase 24.11: structure, inhibitionand experimental and clinical pharmacology. Pharmacol. Rev. 45,87–146.
Sastre, M., Turner, R.S., and Levy, E. (1998). X11 interaction withβ-amyloid precursor protein modulates its cellular stabilization andreduces amyloid β-protein secretion. J. Biol. Chem. 273, 22351–22357.
Sastre, M., Steiner, H., Fuchs, K., Capell, A., Multhaup, G., Condron,M.M., Teplow, D.B., and Haass, C. (2001). Presenilin dependentγ-secretase processing of β-amyloid precursor protein at a site cor-responding to the S3 cleavage of Notch. EMBO Rep. 2, 835–841.
Saura, C.A., Choi, S.-Y., Beglopoulos, V., Malkani, S., Zhang, D.,Rao, B.S.S., Chattarji, S., Kelleher, R.J. III, Kandel, E.R., et al. (2004).Loss of presenilin function causes impairments of memory andsynaptic plasticity followed by age-dependent neurodegeneration.Neuron 42, 23–36.
Shearman, M.S., Beher, D., Clarke, E.E., Lewis, H.D., Harrison, T.,Hunt, P., Nadin, A., Smith, A.L., Stevenson, G., and Castro, J.L.(2000). L-685,458, an aspartyl protease transition state mimic, is apotent inhibitor of amyloid β-protein precursor γ-secretase activity.Biochemistry 39, 8698–8704.
Shen, J., Bronson, R.T., Chen, D.F., Xia, W., Selkoe, D., and Tone-gawa, S. (1997). Skeletal and CNS defects in presenilin-1-deficientmice. Cell 89, 629–639.
Shirotani, K., Tsubuki, S., Iwata, N., Takaki, Y., Harigaya, W., Maru-yama, K., Kiryu-Seo, S., Kiyama, H., Iwata, H., Tomita, T., et al.(2001). Neprilysin degrades both amyloid β peptides 1–40 and1–42 most rapidly and efficiently among thiorphan- and phosphor-amidon-sensitive endopeptidases. J. Biol. Chem. 276, 21895–21901.
Sisodia, S.S., and St. George-Hyslop, P. (2002). γ-secretase, Notch,Aβ and Alzheimer’s disease: where do presenilins fit in? Nat. Rev.Neurosci. 3, 281–290.
Suda, H., Aoyagi, T., Takeuchi, T., and Umezawa, H. (1973). Letter:A thermolysin inhibitor produced by actinomycetes: phosphorami-don. J. Antibiot. (Tokyo) 26, 621–623.
Vassar, R., and Citron, M. (2000). Aβ-generating enzymes: recentadvances in β- and γ-secretases research. Neuron 27, 419–422.
Vincent, B., Beaudet, A., Dauch, P., Vincent, J.P., and Checler, F.(1996). Distinct properties of neuronal and astrocytic endopepti-dase 3.4.24.16: a study on differentiation, subcellular distributionand secretion processes. J. Neurosci. 16, 5049–5059.
Wang, B., Hu, Q., Hearn, M.G., Shimizu, K., Ware, C.B., Liggitt, D.H.,

Neuron554
Jin, L.-W., Cool, B.H., Storm, D.R., and Martin, G.M. (2004). Iso-form-specific knockout of Fe65 leads to impaired learning andmemory. J. Neurosci. Res. 75, 12–24.
Weidemann, A., Eggert, S., Reinhard, F.B.M., Vogel, M., Paliga, K.,Baier, G., Masters, C.L., Beyreuther, K., and Evin, G. (2002). A novel�-cleavage within the transmembrane domain of the Alzheimer am-yloid precursor protein demonstrates homology with the Notchprocessing. Biochemistry 41, 2825–2835.
Wolfe, M.S., Citron, M., Diehl, T.S., Xia, W., Donkor, I.O., and Selkoe,D.J. (1998). A substrate-based difluoro ketone selectively inhibitsAlzheimer’s γ-secretase activity. J. Med. Chem. 41, 6–9.
Wolfe, M.S., Xia, W., Ostaszewski, B.L., Diehl, T.S., Kimberly, W.T.,and Selkoe, D.J. (1999). Two transmembrane aspartates in prese-nilin-1 required for presenilin endoproteolysis and γ-secretase ac-tivity. Nature 398, 513–517.
Yu, G., Nishimura, M., Arawaka, S., Levitan, D., Zhang, L., Tandon,A., Song, Y.-Q., Rogaeva, E., Chen, F., Kawarai, T., et al. (2000).Nicastrin modulates presenilin-mediated notch/glp1 signal trans-duction and βAPP processing. Nature 407, 48–54.
Zhang, Z., Nadeau, P., Song, W., Donoviel, D., Yuan, M., Bernstein,A., and Yankner, B.A. (2000). Presenilins are required for γ-secre-tase cleavage of βAPP and transmembrane cleavage of Notch. Nat.Cell Biol. 2, 463–465.
Copyright © 2022 FDOKUMEN




















