
Prenatal exposure to valproic acid leads to reduced expression of synaptic adhesion molecule...
10
PRENATAL EXPOSURE TO VALPROIC ACID LEADS TO REDUCED EXPRESSION OF SYNAPTIC ADHESION MOLECULE NEUROLIGIN 3 IN MICE E. KOLOZSI, a,b R. N. MACKENZIE, a,b F. I. ROULLET, a,b D. DECATANZARO c AND J. A. FOSTER a,b * a Department of Psychiatry and Behavioural Neurosciences, McMaster University, Hamilton, ON, Canada b Brain-Body Institute, St. Joseph’s Healthcare, Hamilton, ON, Canada c Department of Psychology, Neuroscience and Behaviour, McMaster University, Hamilton, ON, Canada Abstract—In rodents, a single administration of valproic acid (VPA) in utero leads to developmental delays and lifelong deficits in motor performance, social behavior, and anxiety- like behavior in the offspring. Recently, we have demon- strated that VPA mice show alterations in postnatal growth and development, and deficits in olfactory discrimination and social behavior early in development. Based on behavioral and molecular parallels between VPA rodents and individuals with autism, maternal challenge with VPA has been sug- gested to be a good animal model of autism. Neuroligins (NLGN) are a family of postsynaptic cell-adhesion molecules that play a role in synaptic maturation through association with their presynaptic partners, the neurexins (NRXN). Both NLGNs and NRXN members have been implicated in genetic studies of autism. In the present study, we examined changes at the level of expression of NLGN and NRXN mRNAs in the adult brain from mice exposed in utero to VPA. Mouse brain tissue was processed using in situ hybridization and ana- lyzed with densitometry to examine expression of three NLGN genes (NLGN1, NLGN2, and NLGN3) and three NRXN genes (NRXN1, NRXN2, and NRXN3). Expression levels of NLGN1, NLGN2, NRXN1, NRXN2, and NRXN3 were observed to be similar in VPA and control mice. NLGN3 mRNA expres- sion was found to be significantly lower in the VPA mice relative to control animals in hippocampal subregions, cornu ammonis (CA1) and dentate gyrus, and somatosensory cor- tex. This lowered expression may be linked to autistic-like behavioral phenotype observed in the VPA mice. © 2009 IBRO. Published by Elsevier Ltd. All rights reserved. Key words: valproate, neuroligin, neurexin, genetics of au- tism, animal model, in utero challenge. The importance of the intrauterine environment to normal, healthy brain development is well established. Environ- mental challenges during the prenatal period alter the tra- jectory of CNS development and may confer vulnerability to a variety of diseases later in life (Christianson et al., 1994; Rasalam et al., 2005; Bromley et al., 2008). Animal models provide an opportunity to study how environmental challenges lead to alterations in behavior and to examine the underlying molecular changes that contribute to deficits in the offspring. In rats and in mice, a single exposure to valproic acid (VPA), a known teratogen, leads to alter- ations in behavior that are persistent throughout life (Vo- rhees, 1987; Schneider and Przewlocki, 2005; Schneider et al., 2006, 2008; Wagner et al., 2006; Tsujino et al., 2007; Markram et al., 2008). In addition, neuroanatomical and molecular deficits have been described in VPA-treated rodents (Rodier et al., 1996, 1997; Ingram et al., 2000; Tsujino et al., 2007; Nakasato et al., 2008). Most behavioral studies examining the impact of pre- natal VPA exposure have been completed in rats. These studies reported that in comparison to normal adult rats, VPA rats showed delays in postnatal growth (body mass) and development (eye opening) (Schneider and Prze- wlocki, 2005), deficits in olfactory discrimination (Schnei- der and Przewlocki, 2005), less reaction to pain (Markram et al., 2008; Schneider et al., 2008), increased activity in a novel open field (Schneider et al., 2007; Tsujino et al., 2007), increased activity and feeding during the light pe- riod (Tsujino et al., 2007), increased stereotypic behavior (Schneider et al., 2008), reduced exploratory behavior (Schneider and Przewlocki, 2005), impaired sensorimotor gating (Schneider and Przewlocki, 2005; Markram et al., 2008), increased anxiety-like behavior (Vorhees, 1987; Schneider et al., 2007, 2008; Markram et al., 2008), in- creased conditioned fear memory to both tone and context (Markram et al., 2008), and social interaction deficits (Schneider and Przewlocki, 2005; Markram et al., 2008; Schneider et al., 2008). Recent work in our laboratory has demonstrated deficits in early postnatal behavior in mouse pups exposed in utero to VPA that are comparable to those observed in rats, including deficits in olfactory discrimina- tion at postnatal day (P)-9 and reduced sociability at P22. VPA-treated pups showed significant developmental de- lays in eye opening and reduced weight at the time of weaning (Roullet et al., 2007). Wagner et al. (2006) also showed motor deficits in preweaning and adolescent VPA mice, and increased activity in adolescent VPA mice as was previously demonstrated in rats. Together, these re- ports demonstrate that maternal exposure to VPA leads to developmental delays and lifelong deficits in motor perfor- mance, social behavior, and anxiety-like behavior in the offspring. *Correspondence to: J. A. Foster, Brain–Body St. Joseph’s Health- care, 50 Charlton Avenue, Tower T3308, Hamilton, Ontario, Canada L8N 4A6. Tel: 1-905-522-115535993; fax: 1-905-540-6593. E-mail address: [email protected] (J. A. Foster). Abbreviations: ASD, autism spectrum disorder; CA, cornu ammonis; CB, cerebellum; CNV, copy number variation; CON, control (mice); DG, dentate gyrus; DPM, disintegrations per minute; MOB, main ol- factory bulb; M1, motor cortex; NLGN, neuroligin; NRXN, neurexin; P, postnatal day; S1, somatosensory cortex; TEA, triethanolamine–HCl; VPA, valproic acid. Neuroscience 163 (2009) 1201–1210 0306-4522/09 $ - see front matter © 2009 IBRO. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.neuroscience.2009.07.021 1201
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Prenatal exposure to valproic acid leads to reduced expression of synaptic adhesion molecule...

PEN
EDa
Ub
c
U
A(dlsasawg(twNsaatlNgNtsratbI
Kt
Thm
*cLEACDfpV
Neuroscience 163 (2009) 1201–1210
0d
RENATAL EXPOSURE TO VALPROIC ACID LEADS TO REDUCEDXPRESSION OF SYNAPTIC ADHESION MOLECULE
EUROLIGIN 3 IN MICEjt1mctivareMmrT
nsVawden2r((g2Sc((SdpotVlwsmwpdm
. KOLOZSI,a,b R. N. MACKENZIE,a,b F. I. ROULLET,a,b
. DECATANZAROc AND J. A. FOSTERa,b*
Department of Psychiatry and Behavioural Neurosciences, McMasterniversity, Hamilton, ON, Canada
Brain-Body Institute, St. Joseph’s Healthcare, Hamilton, ON, Canada
Department of Psychology, Neuroscience and Behaviour, McMasterniversity, Hamilton, ON, Canada
bstract—In rodents, a single administration of valproic acidVPA) in utero leads to developmental delays and lifelongeficits in motor performance, social behavior, and anxiety-
ike behavior in the offspring. Recently, we have demon-trated that VPA mice show alterations in postnatal growthnd development, and deficits in olfactory discrimination andocial behavior early in development. Based on behavioralnd molecular parallels between VPA rodents and individualsith autism, maternal challenge with VPA has been sug-ested to be a good animal model of autism. NeuroliginsNLGN) are a family of postsynaptic cell-adhesion moleculeshat play a role in synaptic maturation through associationith their presynaptic partners, the neurexins (NRXN). BothLGNs and NRXN members have been implicated in genetictudies of autism. In the present study, we examined changest the level of expression of NLGN and NRXN mRNAs in thedult brain from mice exposed in utero to VPA. Mouse brainissue was processed using in situ hybridization and ana-yzed with densitometry to examine expression of threeLGN genes (NLGN1, NLGN2, and NLGN3) and three NRXNenes (NRXN1, NRXN2, and NRXN3). Expression levels ofLGN1, NLGN2, NRXN1, NRXN2, and NRXN3 were observed
o be similar in VPA and control mice. NLGN3 mRNA expres-ion was found to be significantly lower in the VPA miceelative to control animals in hippocampal subregions, cornummonis (CA1) and dentate gyrus, and somatosensory cor-ex. This lowered expression may be linked to autistic-likeehavioral phenotype observed in the VPA mice. © 2009
BRO. Published by Elsevier Ltd. All rights reserved.
ey words: valproate, neuroligin, neurexin, genetics of au-ism, animal model, in utero challenge.
he importance of the intrauterine environment to normal,ealthy brain development is well established. Environ-ental challenges during the prenatal period alter the tra-
Correspondence to: J. A. Foster, Brain–Body St. Joseph’s Health-are, 50 Charlton Avenue, Tower T3308, Hamilton, Ontario, Canada8N 4A6. Tel: �1-905-522-1155�35993; fax: �1-905-540-6593.-mail address: [email protected] (J. A. Foster).bbreviations: ASD, autism spectrum disorder; CA, cornu ammonis;B, cerebellum; CNV, copy number variation; CON, control (mice);G, dentate gyrus; DPM, disintegrations per minute; MOB, main ol-
actory bulb; M1, motor cortex; NLGN, neuroligin; NRXN, neurexin; P,
oostnatal day; S1, somatosensory cortex; TEA, triethanolamine–HCl;PA, valproic acid.
306-4522/09 $ - see front matter © 2009 IBRO. Published by Elsevier Ltd. All rightoi:10.1016/j.neuroscience.2009.07.021
1201
ectory of CNS development and may confer vulnerabilityo a variety of diseases later in life (Christianson et al.,994; Rasalam et al., 2005; Bromley et al., 2008). Animalodels provide an opportunity to study how environmental
hallenges lead to alterations in behavior and to examinehe underlying molecular changes that contribute to deficitsn the offspring. In rats and in mice, a single exposure toalproic acid (VPA), a known teratogen, leads to alter-tions in behavior that are persistent throughout life (Vo-hees, 1987; Schneider and Przewlocki, 2005; Schneidert al., 2006, 2008; Wagner et al., 2006; Tsujino et al., 2007;arkram et al., 2008). In addition, neuroanatomical andolecular deficits have been described in VPA-treated
odents (Rodier et al., 1996, 1997; Ingram et al., 2000;sujino et al., 2007; Nakasato et al., 2008).
Most behavioral studies examining the impact of pre-atal VPA exposure have been completed in rats. Thesetudies reported that in comparison to normal adult rats,PA rats showed delays in postnatal growth (body mass)nd development (eye opening) (Schneider and Prze-locki, 2005), deficits in olfactory discrimination (Schnei-er and Przewlocki, 2005), less reaction to pain (Markramt al., 2008; Schneider et al., 2008), increased activity in aovel open field (Schneider et al., 2007; Tsujino et al.,007), increased activity and feeding during the light pe-iod (Tsujino et al., 2007), increased stereotypic behaviorSchneider et al., 2008), reduced exploratory behaviorSchneider and Przewlocki, 2005), impaired sensorimotorating (Schneider and Przewlocki, 2005; Markram et al.,008), increased anxiety-like behavior (Vorhees, 1987;chneider et al., 2007, 2008; Markram et al., 2008), in-reased conditioned fear memory to both tone and contextMarkram et al., 2008), and social interaction deficitsSchneider and Przewlocki, 2005; Markram et al., 2008;chneider et al., 2008). Recent work in our laboratory hasemonstrated deficits in early postnatal behavior in mouseups exposed in utero to VPA that are comparable to thosebserved in rats, including deficits in olfactory discrimina-ion at postnatal day (P)-9 and reduced sociability at P22.PA-treated pups showed significant developmental de-
ays in eye opening and reduced weight at the time ofeaning (Roullet et al., 2007). Wagner et al. (2006) alsohowed motor deficits in preweaning and adolescent VPAice, and increased activity in adolescent VPA mice asas previously demonstrated in rats. Together, these re-orts demonstrate that maternal exposure to VPA leads toevelopmental delays and lifelong deficits in motor perfor-ance, social behavior, and anxiety-like behavior in the
ffspring.s reserved.

iaeBrwoMaapHsio2tcdeTtmb
p(mmatitsa2CotodvidMpaw
crtam3eV
A
TRCnh9fmtw(o1Viect
T
Awswu
R
CScPNqbNIwMpPb
T
m
NNNNNN
E. Kolozsi et al. / Neuroscience 163 (2009) 1201–12101202
There are also molecular changes in offspring exposedn utero to VPA, including cerebellar pathology (Rodier etl., 1996, 1997) and altered levels of monoamines (Naritat al., 2002; Tsujino et al., 2007; Nakasato et al., 2008).ased on behavioral and molecular parallels between VPA
odents and individuals with autism, maternal challengeith VPA has been suggested to be a good animal modelf autism (Rodier et al., 1997; Schneider et al., 2006;arkram et al., 2008). Clinically, the link between VPA andutism was originally based on the increased frequency ofutistic symptoms in children diagnosed with fetal val-roate syndrome (Christianson et al., 1994; Williams andersh, 1997; Williams et al., 2001). Recent work demon-trated that VPA monotherapy during pregnancy resultedn seven times greater incidence of autism spectrum dis-rder (ASD) or key symptoms of ASD (Bromley et al.,008). While there may be limits in the construct validity ofhe VPA model to the wide spectrum of etiologies that likelyontribute to autism, there is significant face validity (Ro-ier et al., 1997; Crawley, 2004; Moy et al., 2006) andvidence of predictive validity (Schneider et al., 2006).herefore, studies using the VPA mouse model are likely
o contribute to our understanding of the neurobiologicalechanisms underlying the development of autistic-likeehavior.
In the present study, we examined gene expressionatterns of synaptic adhesion molecules, the neuroliginsNLGN) and neurexins (NRXN), in adult VPA and controlice. NLGNs are a family of postsynaptic cell-adhesionolecules that play a role in synaptic maturation throughssociation with their presynaptic partners, the NRXN (Ich-chenko et al., 1995, 1996). NLGN–NRXN interactions aremportant to synaptic maturation and stabilization and con-ribute to the formation of both inhibitory and excitatoryynapses (Dean et al., 2003; Graf et al., 2004; Chubykin etl., 2005; Levinson et al., 2005; Lise and El-Husseini,006; Budreck and Scheiffele, 2007; Comoletti et al., 2007;hen et al., 2008). We selected these gene targets basedn recent clinical studies in autistic patients suggestinghat alterations in synaptic function contribute to the devel-pment of autism. Szatmari and colleagues (2007) con-ucted whole genome linkage and gene copy numberariation (CNV) analysis of over 1400 families with autismn an attempt to identify genetic risk loci; mutations andeletions in NLGN and NRXN genes were implicated.utations in NLGN3 and NLGN4 have been reported inatients with autism (Jamain et al., 2003; Laumonnier etl., 2004; Yan et al., 2005). Structural variants in NRXN1ere detected in patients with autism but not in healthy
able 1. Gene-specific primers used for probe preparation
RNA Accession number Forward primer
LGN1 NM_138666 AGGGATTCAGCCCTTALGN2 NM_198862 GAGGAAAGGGGGAATCLGN3 NM_172932 CGTGTATAGCTCTATCCRXN1 NM_020252 TTACTCTGGCTGCTGCRXN2 NM_020253 ACGAAGAGGAAACGCA
RXN3 NM_172544 GCGGCCATAAGAAACAGAAAontrols (Yan et al., 2008). These “private” mutations areare and some studies have not detected mutations inhese target genes in individuals with autism (Vincent etl., 2004; Gauthier et al., 2005). In our laboratory, we haveade riboprobes for NLGN1, 2, and 3 and NRXN1, 2, andmRNAs, and in the current study we examined adult
xpression of these mRNAs in mice exposed in utero toPA.
EXPERIMENTAL PROCEDURES
nimals
his research was approved by the McMaster University Animalesearch Ethics Board, conforming to standards of the Canadianouncil on Animal Care. All efforts were made to minimize theumber of animals used and their suffering. All animals wereoused under a reversed light/dark cycle (lights on from 7 PM untilAM). Fifteen pairs of HS mice (heterogeneous strain, previously
ormed by interbreeding C57, DBA, Swiss and CF-1 strains) wereated. Sperm plugs were visually detected and the day of detec-
ion was labeled gestational day 0. On gestational day 11, femalesere fed 1.5 g of peanut butter (Kraft smooth) mixed with VPA
VPA, sodium salt, Sigma P4543, Oakville, ON, Canada) basedn animal’s weight on day of administration (VPA, 800 mg/kg) or.5 g of peanut butter alone. Dams that ate the entire mixturePA/PB (plate found clean, without a visible trace of PB) were
ncluded (n�3) and their pups (VPA group) were used for thexperiment. Two pregnant dams ate PB only and their pupsonstituted the CON group. After birth, the litter was housed withhe mother until weaning at P28.
issue collection
dult mice were euthanized by decapitation on P60 or -61. Brainsere removed and frozen in 2-methylbutane at �60 °C and thentored at �70 °C until sectioned. Sagittal sections 10 �m thickere thaw-mounted to gelatin-coated slides and stored at �35 °Cntil processed for in situ hybridization.
iboprobes
ortical cDNA was generated by reverse transcriptase usingtratagene’s 5.0 Multi-Temperature RT Protocol from mouseortical total RNA. Gene-specific primers were used to amplifyCR cDNA products for probe preparation for NLGN1, NLGN2,LGN3, NRXN1, NRXN2, and NRXN3 (Table 1). Primer se-uences for NLGN1 and NLGN3 were obtained from the Allanrain atlas (Lein et al., 2007). Primer sequences for NLGN2,RXN1, NRXN2, and NRXN3 were designed using Primer 3
nput 4.0 software (Rozen and Skaletsky, 2000). PCR productsere purified using QIA quick PCR purification kit (Qiagen,ississauga, ON, Canada) and then ligated into PromegaGEM-T Easy vector (VWR Int., Mississauga, ON, Canada).robe sequences were confirmed using DNA sequencing (Mo-ix Laboratory, McMaster University). Antisense and sense ribo-
Reverse primer cDNA size (bp)
TGCCATTAACAGCATAGGTGTC 619AGACACAAGTGGGAGGGTTG 514ATTGCTAGCCTCTTCCTCCTCT 644GTCTGAAATCCAGGCCACAT 528GAAACCCAAAGGAAGGGAAG 459
CATACATCTGCGGAG
AATGAAAA
TCCAAGGTCATGTGGAGTCA 349
ptP
mp
I
PawMw7ca(sRamun
Fdel
Fwm
E. Kolozsi et al. / Neuroscience 163 (2009) 1201–1210 1203
robes were transcribed from linearized plasmid DNA by in vitroranscription with �-35S-UTP (specific activity �1000 Ci/mmol;erkinElmer, Boston, MA, USA) using appropriate RNA poly-
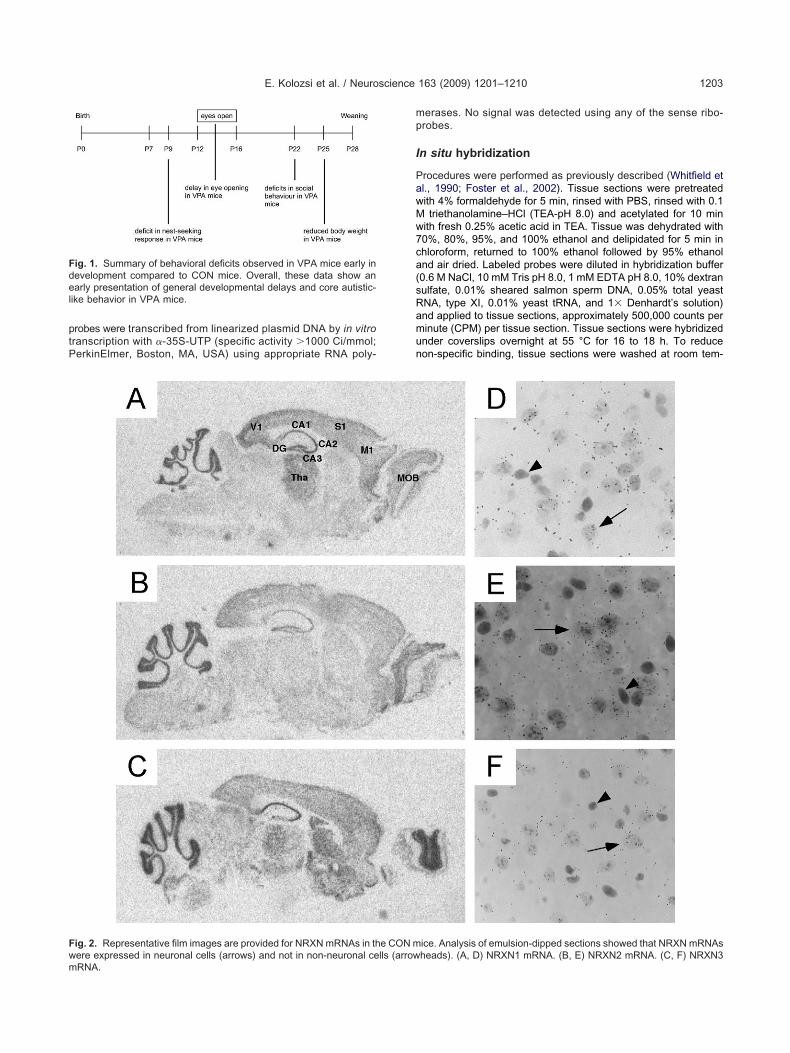
ig. 1. Summary of behavioral deficits observed in VPA mice early inevelopment compared to CON mice. Overall, these data show anarly presentation of general developmental delays and core autistic-
ike behavior in VPA mice.
ig. 2. Representative film images are provided for NRXN mRNAs in th
ere expressed in neuronal cells (arrows) and not in non-neuronal cells (arrowRNA.erases. No signal was detected using any of the sense ribo-robes.
n situ hybridization
rocedures were performed as previously described (Whitfield etl., 1990; Foster et al., 2002). Tissue sections were pretreatedith 4% formaldehyde for 5 min, rinsed with PBS, rinsed with 0.1triethanolamine–HCl (TEA-pH 8.0) and acetylated for 10 min
ith fresh 0.25% acetic acid in TEA. Tissue was dehydrated with0%, 80%, 95%, and 100% ethanol and delipidated for 5 min inhloroform, returned to 100% ethanol followed by 95% ethanolnd air dried. Labeled probes were diluted in hybridization buffer0.6 M NaCl, 10 mM Tris pH 8.0, 1 mM EDTA pH 8.0, 10% dextranulfate, 0.01% sheared salmon sperm DNA, 0.05% total yeastNA, type XI, 0.01% yeast tRNA, and 1� Denhardt’s solution)nd applied to tissue sections, approximately 500,000 counts perinute (CPM) per tissue section. Tissue sections were hybridizednder coverslips overnight at 55 °C for 16 to 18 h. To reduceon-specific binding, tissue sections were washed at room tem-
ice. Analysis of emulsion-dipped sections showed that NRXN mRNAs
e CON m heads). (A, D) NRXN1 mRNA. (B, E) NRXN2 mRNA. (C, F) NRXN3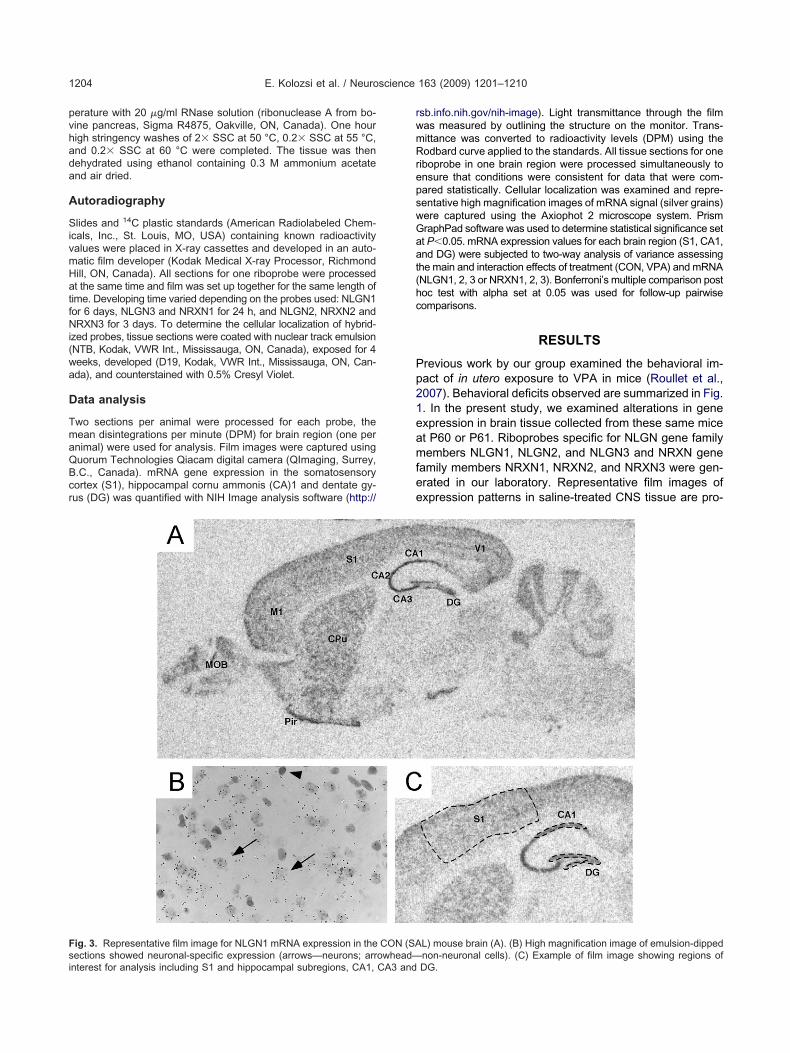
pvhada
A
SivmHatfNi(wa
D
TmaQBcr
rwmRrepswGaat(hc
Pp21eamfee
Fsi
E. Kolozsi et al. / Neuroscience 163 (2009) 1201–12101204
erature with 20 �g/ml RNase solution (ribonuclease A from bo-ine pancreas, Sigma R4875, Oakville, ON, Canada). One hourigh stringency washes of 2� SSC at 50 °C, 0.2� SSC at 55 °C,nd 0.2� SSC at 60 °C were completed. The tissue was thenehydrated using ethanol containing 0.3 M ammonium acetatend air dried.
utoradiography
lides and 14C plastic standards (American Radiolabeled Chem-cals, Inc., St. Louis, MO, USA) containing known radioactivityalues were placed in X-ray cassettes and developed in an auto-atic film developer (Kodak Medical X-ray Processor, Richmondill, ON, Canada). All sections for one riboprobe were processedt the same time and film was set up together for the same length ofime. Developing time varied depending on the probes used: NLGN1or 6 days, NLGN3 and NRXN1 for 24 h, and NLGN2, NRXN2 andRXN3 for 3 days. To determine the cellular localization of hybrid-
zed probes, tissue sections were coated with nuclear track emulsionNTB, Kodak, VWR Int., Mississauga, ON, Canada), exposed for 4eeks, developed (D19, Kodak, VWR Int., Mississauga, ON, Can-da), and counterstained with 0.5% Cresyl Violet.
ata analysis
wo sections per animal were processed for each probe, theean disintegrations per minute (DPM) for brain region (one pernimal) were used for analysis. Film images were captured usinguorum Technologies Qiacam digital camera (QImaging, Surrey,.C., Canada). mRNA gene expression in the somatosensoryortex (S1), hippocampal cornu ammonis (CA)1 and dentate gy-us (DG) was quantified with NIH Image analysis software (http://
ig. 3. Representative film image for NLGN1 mRNA expression in the
ections showed neuronal-specific expression (arrows—neurons; arrowhead—nterest for analysis including S1 and hippocampal subregions, CA1, CA3 andsb.info.nih.gov/nih-image). Light transmittance through the filmas measured by outlining the structure on the monitor. Trans-ittance was converted to radioactivity levels (DPM) using theodbard curve applied to the standards. All tissue sections for one
iboprobe in one brain region were processed simultaneously tonsure that conditions were consistent for data that were com-ared statistically. Cellular localization was examined and repre-entative high magnification images of mRNA signal (silver grains)ere captured using the Axiophot 2 microscope system. PrismraphPad software was used to determine statistical significance sett P�0.05. mRNA expression values for each brain region (S1, CA1,nd DG) were subjected to two-way analysis of variance assessinghe main and interaction effects of treatment (CON, VPA) and mRNANLGN1, 2, 3 or NRXN1, 2, 3). Bonferroni’s multiple comparison postoc test with alpha set at 0.05 was used for follow-up pairwiseomparisons.
RESULTS
revious work by our group examined the behavioral im-act of in utero exposure to VPA in mice (Roullet et al.,007). Behavioral deficits observed are summarized in Fig.. In the present study, we examined alterations in genexpression in brain tissue collected from these same micet P60 or P61. Riboprobes specific for NLGN gene familyembers NLGN1, NLGN2, and NLGN3 and NRXN gene
amily members NRXN1, NRXN2, and NRXN3 were gen-rated in our laboratory. Representative film images ofxpression patterns in saline-treated CNS tissue are pro-
L) mouse brain (A). (B) High magnification image of emulsion-dipped
CON (SA non-neuronal cells). (C) Example of film image showing regions ofDG.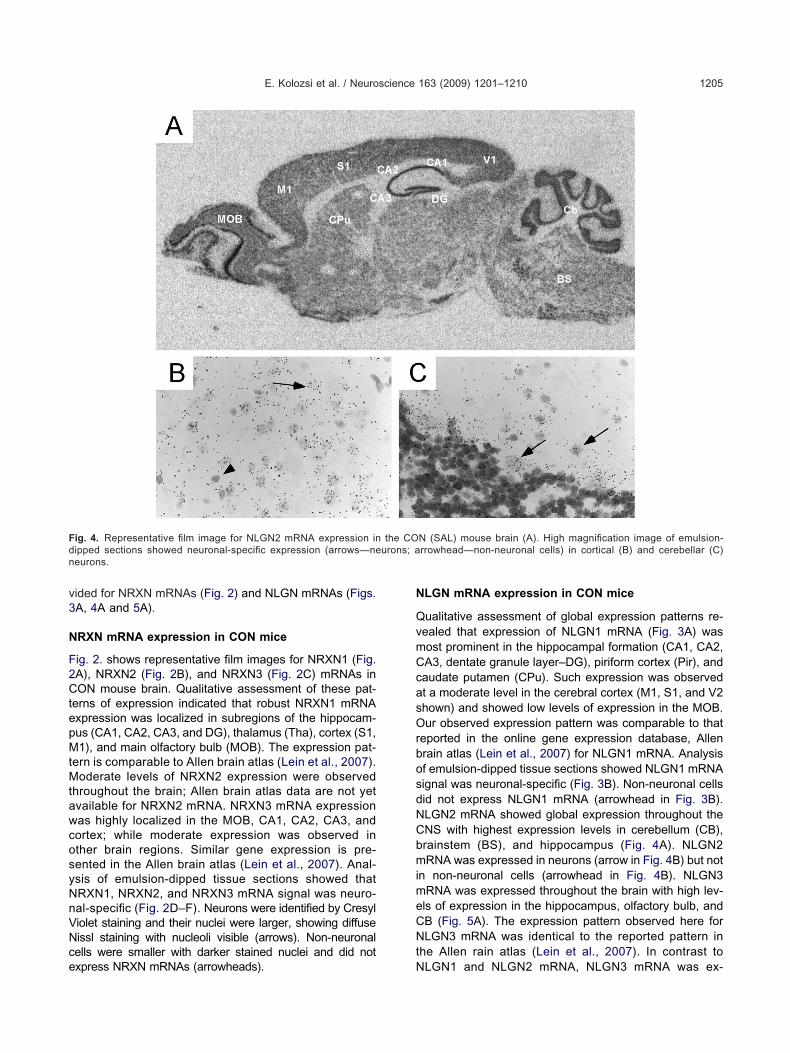
v3
N
F2CtepMtMtawcosyNnVNce
N
QvmCcasOrbosdNCbmimeCNt
Fd urons; an
E. Kolozsi et al. / Neuroscience 163 (2009) 1201–1210 1205
ided for NRXN mRNAs (Fig. 2) and NLGN mRNAs (Figs.A, 4A and 5A).
RXN mRNA expression in CON mice
ig. 2. shows representative film images for NRXN1 (Fig.A), NRXN2 (Fig. 2B), and NRXN3 (Fig. 2C) mRNAs inON mouse brain. Qualitative assessment of these pat-
erns of expression indicated that robust NRXN1 mRNAxpression was localized in subregions of the hippocam-us (CA1, CA2, CA3, and DG), thalamus (Tha), cortex (S1,1), and main olfactory bulb (MOB). The expression pat-
ern is comparable to Allen brain atlas (Lein et al., 2007).oderate levels of NRXN2 expression were observed
hroughout the brain; Allen brain atlas data are not yetvailable for NRXN2 mRNA. NRXN3 mRNA expressionas highly localized in the MOB, CA1, CA2, CA3, andortex; while moderate expression was observed inther brain regions. Similar gene expression is pre-ented in the Allen brain atlas (Lein et al., 2007). Anal-sis of emulsion-dipped tissue sections showed thatRXN1, NRXN2, and NRXN3 mRNA signal was neuro-al-specific (Fig. 2D–F). Neurons were identified by Cresyliolet staining and their nuclei were larger, showing diffuseissl staining with nucleoli visible (arrows). Non-neuronalells were smaller with darker stained nuclei and did not
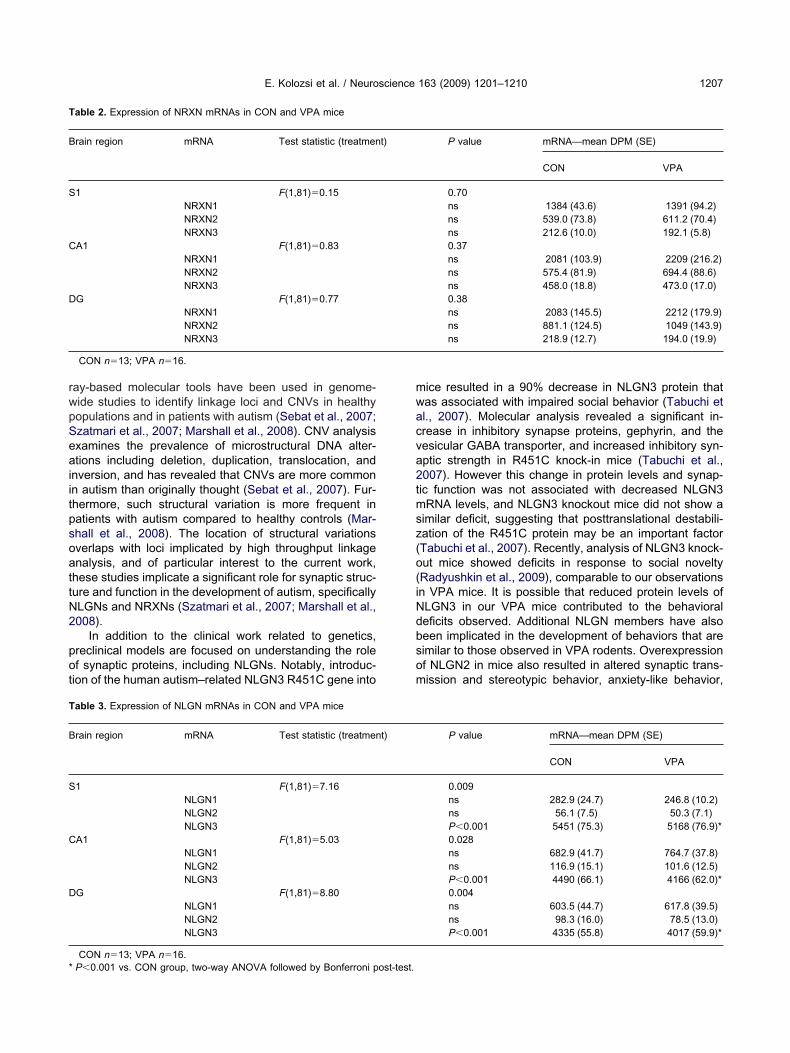
ig. 4. Representative film image for NLGN2 mRNA expression inipped sections showed neuronal-specific expression (arrows—neeurons.
xpress NRXN mRNAs (arrowheads). N
LGN mRNA expression in CON mice
ualitative assessment of global expression patterns re-ealed that expression of NLGN1 mRNA (Fig. 3A) wasost prominent in the hippocampal formation (CA1, CA2,A3, dentate granule layer–DG), piriform cortex (Pir), andaudate putamen (CPu). Such expression was observedt a moderate level in the cerebral cortex (M1, S1, and V2hown) and showed low levels of expression in the MOB.ur observed expression pattern was comparable to that
eported in the online gene expression database, Allenrain atlas (Lein et al., 2007) for NLGN1 mRNA. Analysisf emulsion-dipped tissue sections showed NLGN1 mRNAignal was neuronal-specific (Fig. 3B). Non-neuronal cellsid not express NLGN1 mRNA (arrowhead in Fig. 3B).LGN2 mRNA showed global expression throughout theNS with highest expression levels in cerebellum (CB),rainstem (BS), and hippocampus (Fig. 4A). NLGN2RNA was expressed in neurons (arrow in Fig. 4B) but not
n non-neuronal cells (arrowhead in Fig. 4B). NLGN3RNA was expressed throughout the brain with high lev-ls of expression in the hippocampus, olfactory bulb, andB (Fig. 5A). The expression pattern observed here forLGN3 mRNA was identical to the reported pattern in
he Allen rain atlas (Lein et al., 2007). In contrast to
N (SAL) mouse brain (A). High magnification image of emulsion-rrowhead—non-neuronal cells) in cortical (B) and cerebellar (C)
the CO
LGN1 and NLGN2 mRNA, NLGN3 mRNA was ex-

pbcrmi
V
TNsFDHaVTdNpwN
Wm
trlwRfsfY
atimrSP2antiumgt
Fs rtical rege
E. Kolozsi et al. / Neuroscience 163 (2009) 1201–12101206
ressed in neuronal and non-neuronal cells. In mostrain regions, NLGN3 mRNA was localized to neuronalells (shown in Fig. 5B for S1). However, in fiber tractegions such as the fimbria (shown in Fig. 5C), NLGN3RNA was localized to non-neuronal cells (arrowheads
n Fig. 5C).
PA mice showed a global decrease in NLGN3 mRNA
o assess the impact of in utero exposure to VPA onLGN and NRXN mRNAs, gene expression was mea-ured at l.80 mm lateral from the midline (Paxinos andranklin, 2001) in hippocampal subregions CA1, CA3,G and S1. Regions of interest are shown in Fig. 2C.ippocampal and S1 expression of NRXN1, NRXN2,nd NRXN3 mRNAs did not differ between CON andPA mice. Data are presented for NRXN mRNAs inable 2. Similarly, expression of NLGN1 and NLGN2id not differ between CON and VPA mice; however,LGN3 mRNA was significantly reduced in all hip-ocampal regions measured and in S1 in VPA micehen compared to CON mice. Data are presented forLGN mRNAs in Table 3.
DISCUSSION
e show that exposure to VPA on gestational day 11 in
ig. 5. Representative film image for NLGN3 mRNA expression in thections showed neuronal-specific expression (arrows—neurons) in coxpression (arrowhead) of NLGN3 mRNA was detected (C).
ice results in a long-term, global reduction of NLGN3 at a
he level of gene expression. Previous work in mice andats has demonstrated behavioral validity of VPA chal-enge as an animal model of autism (Schneider and Prze-locki, 2005; Schneider et al., 2006; Wagner et al., 2006;oullet et al., 2007; Markram et al., 2008). Our results
urther validate this model at the molecular level, showingpecific downregulation of NLGN3 mRNA, an autism riskactor gene in the clinical population (Jamain et al., 2003;an et al., 2005).
Our decision to study NRXNs and NLGNs in the VPAnimal model was motivated directly by clinical literatureargeted at understanding the genetic basis of ASDs. Themportance of genetics in autism has been recognized for
ore than 20 years (Smalley et al., 1988). Genetic-basedesearch began with family and twin studies (Jones andzatmari, 1988; Ritvo et al., 1989; Bailey et al., 1995;ickles et al., 1995; Risch et al., 1999; Hallmayer et al.,002) that demonstrated both a genetic component inutism and the involvement of multiple genetic loci. Ge-ome-wide linkage studies have identified several suscep-ibility loci for ASD (review by Freitag, 2007). Efforts todentify single gene mutations linked to autism have beennsuccessful. Instead, a picture emerges that supportsultiple susceptibility genes influenced both by gene–ene interactions and gene–environment interactions con-ributing to multiple etiologies of ASD (Freitag, 2007; Rapin
SAL) mouse brain (A). High magnification image of emulsion-dippedions (B), however, in fiber tract regions (fimbria shown) non-neuronal
e CON (
nd Tuchman, 2008). Recently, high throughput microar-
rwpSeaiitpsoattN2
pot
mwacva2tmsz(o(iNdbsom
T
B
S
C
D
T
B
S
C
D
*
E. Kolozsi et al. / Neuroscience 163 (2009) 1201–1210 1207
ay-based molecular tools have been used in genome-ide studies to identify linkage loci and CNVs in healthyopulations and in patients with autism (Sebat et al., 2007;zatmari et al., 2007; Marshall et al., 2008). CNV analysisxamines the prevalence of microstructural DNA alter-tions including deletion, duplication, translocation, and
nversion, and has revealed that CNVs are more commonn autism than originally thought (Sebat et al., 2007). Fur-hermore, such structural variation is more frequent inatients with autism compared to healthy controls (Mar-hall et al., 2008). The location of structural variationsverlaps with loci implicated by high throughput linkagenalysis, and of particular interest to the current work,hese studies implicate a significant role for synaptic struc-ure and function in the development of autism, specificallyLGNs and NRXNs (Szatmari et al., 2007; Marshall et al.,008).
In addition to the clinical work related to genetics,reclinical models are focused on understanding the rolef synaptic proteins, including NLGNs. Notably, introduc-ion of the human autism–related NLGN3 R451C gene into
able 2. Expression of NRXN mRNAs in CON and VPA mice
rain region mRNA Test statistic (treatmen
1 F(1,81)�0.15NRXN1NRXN2NRXN3
A1 F(1,81)�0.83NRXN1NRXN2NRXN3
G F(1,81)�0.77NRXN1NRXN2NRXN3
CON n�13; VPA n�16.
able 3. Expression of NLGN mRNAs in CON and VPA mice
rain region mRNA Test statistic (treatmen
1 F(1,81)�7.16NLGN1NLGN2NLGN3
A1 F(1,81)�5.03NLGN1NLGN2NLGN3
G F(1,81)�8.80NLGN1NLGN2NLGN3
CON n�13; VPA n�16.
P�0.001 vs. CON group, two-way ANOVA followed by Bonferroni post-test.ice resulted in a 90% decrease in NLGN3 protein thatas associated with impaired social behavior (Tabuchi etl., 2007). Molecular analysis revealed a significant in-rease in inhibitory synapse proteins, gephyrin, and theesicular GABA transporter, and increased inhibitory syn-ptic strength in R451C knock-in mice (Tabuchi et al.,007). However this change in protein levels and synap-ic function was not associated with decreased NLGN3RNA levels, and NLGN3 knockout mice did not show a
imilar deficit, suggesting that posttranslational destabili-ation of the R451C protein may be an important factorTabuchi et al., 2007). Recently, analysis of NLGN3 knock-ut mice showed deficits in response to social noveltyRadyushkin et al., 2009), comparable to our observationsn VPA mice. It is possible that reduced protein levels ofLGN3 in our VPA mice contributed to the behavioraleficits observed. Additional NLGN members have alsoeen implicated in the development of behaviors that areimilar to those observed in VPA rodents. Overexpressionf NLGN2 in mice also resulted in altered synaptic trans-ission and stereotypic behavior, anxiety-like behavior,
P value mRNA—mean DPM (SE)
CON VPA
0.70ns 1384 (43.6) 1391 (94.2)ns 539.0 (73.8) 611.2 (70.4)ns 212.6 (10.0) 192.1 (5.8)0.37ns 2081 (103.9) 2209 (216.2)ns 575.4 (81.9) 694.4 (88.6)ns 458.0 (18.8) 473.0 (17.0)0.38ns 2083 (145.5) 2212 (179.9)ns 881.1 (124.5) 1049 (143.9)ns 218.9 (12.7) 194.0 (19.9)
P value mRNA—mean DPM (SE)
CON VPA
0.009ns 282.9 (24.7) 246.8 (10.2)ns 56.1 (7.5) 50.3 (7.1)P�0.001 5451 (75.3) 5168 (76.9)*0.028ns 682.9 (41.7) 764.7 (37.8)ns 116.9 (15.1) 101.6 (12.5)P�0.001 4490 (66.1) 4166 (62.0)*0.004ns 603.5 (44.7) 617.8 (39.5)ns 98.3 (16.0) 78.5 (13.0)P�0.001 4335 (55.8) 4017 (59.9)*
t)
t)

acaiiAwakto2b2sioawpmb(tosso
AtatrsT
B
B
B
B
C
C
C
C
C
D
D
F
F
G
G
H
H
H
I
I
I
J
J
J
L
E. Kolozsi et al. / Neuroscience 163 (2009) 1201–12101208
nd deficits in social response (Hines et al., 2008). Re-ently, the NLGN4 mouse gene was identified (Bolliger etl., 2008), and loss of function NLGN4 mice showed def-
cits in social behavior and reduced ultrasonic vocalizationn response to maternal separation (Jamain et al., 2008).dditional synaptic proteins are implicated in autism andarrant investigation in animal models of autism. For ex-mple, shank proteins are synaptic scaffolding proteinsnown to play a critical role in the structural organization ofhe post-synaptic density and to be important in the devel-pment of functional dendritic spines (Roussignol et al.,005; Sala et al., 2005). Mutations in human shank 3 haveeen described in patients with autism (Durand et al.,007; Moessner et al., 2007). Shank proteins have beenhown to bind NLGNs (Meyer et al., 2004), and alterationsn either Shank proteins or NLGNs could impact the devel-pment of synapses and contribute to the development ofutistic-like behavior. Recently, the Shank1 knockout miceere characterized and shown to have altered synapticroteins, dendritic morphology, and weaker synaptic trans-ission. These connectivity alterations were accompaniedy increased anxiety-like behavior and learning deficitsHung et al., 2008). These studies demonstrate the impor-ance of animal models in understanding the neurobiologyf autism; the VPA model is an excellent system to con-ider the complex protein–protein interactions of the manyynaptic proteins that have been implicated in the devel-pment of autistic-like behavior.
cknowledgments—This research was supported by grants fromhe Natural Sciences and Engineering Research Council of Can-da awarded to Jane A. Foster and Denys deCatanzaro, and from
he Canadian Foundation for Innovation to Jane A. Foster. Flo-ence I. Roullet is a recipient of the Father Sean O’Sullivan Re-earch Centre post-doctoral fellowship and the Autism Researchraining program.
REFERENCES
ailey A, Le Couteur A, Gottesman I, Bolton P, Simonoff E, Yuzda E,Rutter M (1995) Autism as a strongly genetic disorder: evidencefrom a British twin study. Psychol Med 25:63–77.
olliger MF, Pei J, Maxeiner S, Boucard AA, Grishin NV, Sudhof TC(2008) Unusually rapid evolution of Neuroligin-4 in mice. Proc NatlAcad Sci U S A 105:6421–6426.
romley RL, Mawer G, Clayton-Smith J, Baker GA (2008) Autismspectrum disorders following in utero exposure to antiepilepticdrugs. Neurology 71:1923–1924.
udreck EC, Scheiffele P (2007) Neuroligin-3 is a neuronal adhesionprotein at GABAergic and glutamatergic synapses. Eur J Neurosci26:1738–1748.
hen X, Liu H, Shim AH, Focia PJ, He X (2008) Structural basis forsynaptic adhesion mediated by neuroligin-neurexin interactions.Nat Struct Mol Biol 15:50–56.
hristianson AL, Chesler N, Kromberg JG (1994) Fetal valproatesyndrome: clinical and neuro-developmental features in two siblingpairs. Dev Med Child Neurol 36:361–369.
hubykin AA, Liu X, Comoletti D, Tsigelny I, Taylor P, Sudhof TC(2005) Dissection of synapse induction by neuroligins: effect of aneuroligin mutation associated with autism. J Biol Chem 280:22365–22374.
omoletti D, Grishaev A, Whitten AE, Tsigelny I, Taylor P, Trewhella
J (2007) Synaptic arrangement of the neuroligin/beta-neurexincomplex revealed by X-ray and neutron scattering. Structure15:693–705.
rawley JN (2004) Designing mouse behavioral tasks relevant toautistic-like behaviors. Ment Retard Dev Disabil Res Rev 10:248–258.
ean C, Scholl FG, Choih J, DeMaria S, Berger J, Isacoff E, ScheiffeleP (2003) Neurexin mediates the assembly of presynaptic termi-nals. Nat Neurosci 6:708–716.
urand CM, Betancur C, Boeckers TM, Bockmann J, Chaste P, Fau-chereau F, Nygren G, Rastam M, Gillberg IC, Anckarsater H,Sponheim E, Goubran-Botros H, Delorme R, Chabane N, Mouren-Simeoni MC, de Mas P, Bieth E, Roge B, Heron D, Burglen L,Gillberg C, Leboyer M, Bourgeron T (2007) Mutations in the geneencoding the synaptic scaffolding protein SHANK3 are associatedwith autism spectrum disorders. Nat Genet 39:25–27.
oster JA, Quan N, Stern EL, Kristensson K, Herkenham M (2002)Induced neuronal expression of class I major histocompatibilitycomplex mRNA in acute and chronic inflammation models. J Neu-roimmunol 131:83–91.
reitag CM (2007) The genetics of autistic disorders and its clinicalrelevance: a review of the literature. Mol Psychiatry 12:2–22.
authier J, Bonnel A, St-Onge J, Karemera L, Laurent S, Mottron L,Fombonne E, Joober R, Rouleau GA (2005) NLGN3/NLGN4 genemutations are not responsible for autism in the Quebec population.Am J Med Genet B Neuropsychiatr Genet 132B:74–75.
raf ER, Zhang X, Jin SX, Linhoff MW, Craig AM (2004) Neurexinsinduce differentiation of GABA and glutamate postsynaptic spe-cializations via neuroligins. Cell 119:1013–1026.
allmayer J, Glasson EJ, Bower C, Petterson B, Croen L, Grether J,Risch N (2002) On the twin risk in autism. Am J Hum Genet 71:941–946.
ines RM, Wu L, Hines DJ, Steenland H, Mansour S, Dahlhaus R,Singaraja RR, Cao X, Sammler E, Hormuzdi SG, Zhuo M, El-Husseini A (2008) Synaptic imbalance, stereotypies, and impairedsocial interactions in mice with altered neuroligin 2 expression.J Neurosci 28:6055–6067.
ung AY, Futai K, Sala C, Valtschanoff JG, Ryu J, Woodworth MA,Kidd FL, Sung CC, Miyakawa T, Bear MF, Weinberg RJ, Sheng M(2008) Smaller dendritic spines, weaker synaptic transmission, butenhanced spatial learning in mice lacking Shank1. J Neurosci28:1697–1708.
chtchenko K, Hata Y, Nguyen T, Ullrich B, Missler M, Moomaw C,Sudhof TC (1995) Neuroligin 1: a splice site-specific ligand forbeta-neurexins. Cell 81:435–443.
chtchenko K, Nguyen T, Sudhof TC (1996) Structures, alternativesplicing, and neurexin binding of multiple neuroligins. J Biol Chem271:2676–2682.
ngram JL, Peckham SM, Tisdale B, Rodier PM (2000) Prenatal ex-posure of rats to valproic acid reproduces the cerebellar anomaliesassociated with autism. Neurotoxicol Teratol 22:319–324.
amain S, Quach H, Betancur C, Rastam M, Colineaux C, Gillberg IC,Soderstrom H, Giros B, Leboyer M, Gillberg C, Bourgeron T (2003)Mutations of the X-linked genes encoding neuroligins NLGN3 andNLGN4 are associated with autism. Nat Genet 34:27–29.
amain S, Radyushkin K, Hammerschmidt K, Granon S, Boretius S,Varoqueaux F, Ramanantsoa N, Gallego J, Ronnenberg A, WinterD, Frahm J, Fischer J, Bourgeron T, Ehrenreich H, Brose N (2008)Reduced social interaction and ultrasonic communication in amouse model of monogenic heritable autism. Proc Natl Acad SciU S A 105:1710–1715.
ones MB, Szatmari P (1988) Stoppage rules and genetic studies ofautism. J Autism Dev Disord 18:31–40.
aumonnier F, Bonnet-Brilhault F, Gomot M, Blanc R, David A,Moizard MP, Raynaud M, Ronce N, Lemonnier E, Calvas P,Laudier B, Chelly J, Fryns JP, Ropers HH, Hamel BC, Andres C,Barthelemy C, Moraine C, Briault S (2004) X-linked mental retar-
dation and autism are associated with a mutation in the NLGN4
L
L
L
M
M
M
M
M
N
N
P
P
R
R
R
R
R
R
R
R
R
R
S
S
S
S
S
S
E. Kolozsi et al. / Neuroscience 163 (2009) 1201–1210 1209
gene, a member of the neuroligin family. Am J Hum Genet 74:552–557.
ein ES, Hawrylycz MJ, Ao N, Ayres M, Bensinger A, Bernard A, BoeAF, Boguski MS, Brockway KS, Byrnes EJ, Chen L, Chen L, ChenTM, Chin MC, Chong J, Crook BE, Czaplinska A, Dang CN, DattaS, Dee NR, Desaki AL, Desta T, Diep E, Dolbeare TA, Donelan MJ,Dong HW, Dougherty JG, Duncan BJ, Ebbert AJ, Eichele G, EstinLK, Faber C, Facer BA, Fields R, Fischer SR, Fliss TP, Frensley C,Gates SN, Glattfelder KJ, Halverson KR, Hart MR, Hohmann JG,Howell MP, Jeung DP, Johnson RA, Karr PT, Kawal R, Kidney JM,Knapik RH, Kuan CL, Lake JH, Laramee AR, Larsen KD, Lau C,Lemon TA, Liang AJ, Liu Y, Luong LT, Michaels J, Morgan JJ,Morgan RJ, Mortrud MT, Mosqueda NF, Ng LL, Ng R, Orta GJ,Overly CC, Pak TH, Parry SE, Pathak SD, Pearson OC, PuchalskiRB, Riley ZL, Rockett HR, Rowland SA, Royall JJ, Ruiz MJ, SarnoNR, Schaffnit K, Shapovalova NV, Sivisay T, Slaughterbeck CR,Smith SC, Smith KA, Smith BI, Sodt AJ, Stewart NN, Stumpf KR,Sunkin SM, Sutram M, Tam A, Teemer CD, Thaller C, ThompsonCL, Varnam LR, Visel A, Whitlock RM, Wohnoutka PE, Wolkey CK,Wong VY, Wood M, Yaylaoglu MB, Young RC, Youngstrom BL,Yuan XF, Zhang B, Zwingman TA, Jones AR (2007) Genome-wideatlas of gene expression in the adult mouse brain. Nature 445:168–176.
evinson JN, Chery N, Huang K, Wong TP, Gerrow K, Kang R, PrangeO, Wang YT, El-Husseini A (2005) Neuroligins mediate excitatoryand inhibitory synapse formation: involvement of PSD-95 andneurexin-1beta in neuroligin-induced synaptic specificity. J BiolChem 280:17312–17319.
ise MF, El-Husseini A (2006) The neuroligin and neurexin families:from structure to function at the synapse. Cell Mol Life Sci 63:1833–1849.
arkram K, Rinaldi T, La Mendola D, Sandi C, Markram H (2008)Abnormal fear conditioning and amygdala processing in an animalmodel of autism. Neuropsychopharmacology 33:901–912.
arshall CR, Noor A, Vincent JB, Lionel AC, Feuk L, Skaug J, ShagoM, Moessner R, Pinto D, Ren Y, Thiruvahindrapduram B, Fiebig A,Schreiber S, Friedman J, Ketelaars CE, Vos YJ, Ficicioglu C,Kirkpatrick S, Nicolson R, Sloman L, Summers A, Gibbons CA,Teebi A, Chitayat D, Weksberg R, Thompson A, Vardy C, CrosbieV, Luscombe S, Baatjes R, Zwaigenbaum L, Roberts W, Fernan-dez B, Szatmari P, Scherer SW (2008) Structural variation ofchromosomes in autism spectrum disorder. Am J Hum Genet 82:477–488.
eyer G, Varoqueaux F, Neeb A, Oschlies M, Brose N (2004) Thecomplexity of PDZ domain-mediated interactions at glutamatergicsynapses: a case study on neuroligin. Neuropharmacology 47:724–733.
oessner R, Marshall CR, Sutcliffe JS, Skaug J, Pinto D, Vincent J,Zwaigenbaum L, Fernandez B, Roberts W, Szatmari P, SchererSW (2007) Contribution of SHANK3 mutations to autism spectrumdisorder. Am J Hum Genet 81:1289–1297.
oy SS, Nadler JJ, Magnuson TR, Crawley JN (2006) Mouse modelsof autism spectrum disorders: the challenge for behavioral genet-ics. Am J Med Genet C Semin Med Genet 142C:40–51.
akasato A, Nakatani Y, Seki Y, Tsujino N, Umino M, Arita H (2008)Swim stress exaggerates the hyperactive mesocortical dopaminesystem in a rodent model of autism. Brain Res 1193:128–135.
arita N, Kato M, Tazoe M, Miyazaki K, Narita M, Okado N (2002)Increased monoamine concentration in the brain and blood of fetalthalidomide- and valproic acid-exposed rat: putative animal modelsfor autism. Pediatr Res 52:576–579.
axinos G, Franklin KBJ (2001) The mouse brain in stereotaxic coor-dinates. New York: Academic Press.
ickles A, Bolton P, Macdonald H, Bailey A, Le Couteur A, Sim CH,Rutter M (1995) Latent-class analysis of recurrence risks for com-plex phenotypes with selection and measurement error: a twin and
family history study of autism. Am J Hum Genet 57:717–726.adyushkin K, Hammerschmidt K, Boretius S, Varoqueaux F, El-KordiA, Ronnenberg A, Winter D, Frahm J, Fischer J, Brose N, Ehren-reich H (2009) Neuroligin-3 deficient mice: model of a monogenicheritable form of autism with an olfactory deficit. Genes BrainBehav 8:416–425.
apin I, Tuchman RF (2008) What is new in autism? Curr Opin Neurol21:143–149.
asalam AD, Hailey H, Williams JH, Moore SJ, Turnpenny PD, LloydDJ, Dean JC (2005) Characteristics of fetal anticonvulsant syn-drome associated autistic disorder. Dev Med Child Neurol 47:551–555.
isch N, Spiker D, Lotspeich L, Nouri N, Hinds D, Hallmayer J,Kalaydjieva L, McCague P, Dimiceli S, Pitts T, Nguyen L, Yang J,Harper C, Thorpe D, Vermeer S, Young H, Hebert J, Lin A, Fer-guson J, Chiotti C, Wiese-Slater S, Rogers T, Salmon B, NicholasP, Petersen PB, Pingree C, McMahon W, Wong DL, Cavalli-SforzaLL, Kraemer HC, Myers RM (1999) A genomic screen of autism:evidence for a multilocus etiology. Am J Hum Genet 65:493–507.
itvo ER, Jorde LB, Mason-Brothers A, Freeman BJ, Pingree C, JonesMB, McMahon WM, Petersen PB, Jenson WR, Mo A (1989) TheUCLA-University of Utah epidemiologic survey of autism: recur-rence risk estimates and genetic counseling. Am J Psychiatry146:1032–1036.
odier PM, Ingram JL, Tisdale B, Croog VJ (1997) Linking etiologiesin humans and animal models: studies of autism. Reprod Toxicol11:417–422.
odier PM, Ingram JL, Tisdale B, Nelson S, Romano J (1996) Embry-ological origin for autism: developmental anomalies of the cranialnerve motor nuclei. J Comp Neurol 370:247–261.
oullet FI, Wollaston L, deCatanzaro D, Foster JA (2007) Environ-mental challenge during pregnancy: analysis of behaviour andgene expression in a mouse model to study autism. Program No.61.13. 2007 Neuroscience Meeting Planner. San Diego, CA: So-ciety for Neuroscience, 2007.
oussignol G, Ango F, Romorini S, Tu JC, Sala C, Worley PF, Boc-kaert J, Fagni L (2005) Shank expression is sufficient to inducefunctional dendritic spine synapses in aspiny neurons. J Neurosci25:3560–3570.
ozen S, Skaletsky H (2000) Primer3 on the WWW for general usersand for biologist programmers. Methods Mol Biol 132:365–386.
ala C, Roussignol G, Meldolesi J, Fagni L (2005) Key role of thepostsynaptic density scaffold proteins Shank and Homer in thefunctional architecture of Ca2� homeostasis at dendritic spines inhippocampal neurons. J Neurosci 25:4587–4592.
chneider T, Przewlocki R (2005) Behavioral alterations in rats pre-natally exposed to valproic acid: animal model of autism. Neuro-psychopharmacology 30:80–89.
chneider T, Roman A, Basta-Kaim A, Kubera M, Budziszewska B,Schneider K, Przewlocki R (2008) Gender-specific behavioral andimmunological alterations in an animal model of autism induced byprenatal exposure to valproic acid. Psychoneuroendocrinology33:728–740.
chneider T, Turczak J, Przewlocki R (2006) Environmental enrich-ment reverses behavioral alterations in rats prenatally exposed tovalproic acid: issues for a therapeutic approach in autism. Neuro-psychopharmacology 31:36–46.
chneider T, Ziolkowska B, Gieryk A, Tyminska A, Przewlocki R(2007) Prenatal exposure to valproic acid disturbs the enkephalin-ergic system functioning, basal hedonic tone, and emotional re-sponses in an animal model of autism. Psychopharmacology (Berl)193:547–555.
ebat J, Lakshmi B, Malhotra D, Troge J, Lese-Martin C, Walsh T,Yamrom B, Yoon S, Krasnitz A, Kendall J, Leotta A, Pai D, ZhangR, Lee YH, Hicks J, Spence SJ, Lee AT, Puura K, Lehtimaki T,Ledbetter D, Gregersen PK, Bregman J, Sutcliffe JS, JobanputraV, Chung W, Warburton D, King MC, Skuse D, Geschwind DH,Gilliam TC, Ye K, Wigler M (2007) Strong association of de novo
copy number mutations with autism. Science 316:445–449.
S
S
T
T
V
V
W
W
W
W
Y
Y
E. Kolozsi et al. / Neuroscience 163 (2009) 1201–12101210
malley SL, Asarnow RF, Spence MA (1988) Autism and genetics. Adecade of research. Arch Gen Psychiatry 45:953–961.
zatmari P, Paterson AD, Zwaigenbaum L, Roberts W, Brian J, LiuXQ, Vincent JB, Skaug JL, Thompson AP, Senman L, Feuk L, QianC, Bryson SE, Jones MB, Marshall CR, Scherer SW, Vieland VJ,Bartlett C, Mangin LV, Goedken R, Segre A, Pericak-Vance MA,Cuccaro ML, Gilbert JR, Wright HH, Abramson RK, Betancur C,Bourgeron T, Gillberg C, Leboyer M, Buxbaum JD, Davis KL,Hollander E, Silverman JM, Hallmayer J, Lotspeich L, Sutcliffe JS,Haines JL, Folstein SE, Piven J, Wassink TH, Sheffield V, Ge-schwind DH, Bucan M, Brown WT, Cantor RM, Constantino JN,Gilliam TC, Herbert M, Lajonchere C, Ledbetter DH, Lese-MartinC, Miller J, Nelson S, Samango-Sprouse CA, Spence S, State M,Tanzi RE, Coon H, Dawson G, Devlin B, Estes A, Flodman P, KleiL, McMahon WM, Minshew N, Munson J, Korvatska E, Rodier PM,Schellenberg GD, Smith M, Spence MA, Stodgell C, Tepper PG,Wijsman EM, Yu CE, Roge B, Mantoulan C, Wittemeyer K, Pou-stka A, Felder B, Klauck SM, Schuster C, Poustka F, Bolte S,Feineis-Matthews S, Herbrecht E, Schmotzer G, Tsiantis J, Papa-nikolaou K, Maestrini E, Bacchelli E, Blasi F, Carone S, Toma C,Van Engeland H, de Jonge M, Kemner C, Koop F, Langemeijer M,Hijmans C, Staal WG, Baird G, Bolton PF, Rutter ML, Weisblatt E,Green J, Aldred C, Wilkinson JA, Pickles A, Le Couteur A, BerneyT, McConachie H, Bailey AJ, Francis K, Honeyman G, HutchinsonA, Parr JR, Wallace S, Monaco AP, Barnby G, Kobayashi K, LambJA, Sousa I, Sykes N, Cook EH, Guter SJ, Leventhal BL, Salt J,Lord C, Corsello C, Hus V, Weeks DE, Volkmar F, Tauber M,Fombonne E, Shih A, Meyer KJ (2007) Mapping autism risk lociusing genetic linkage and chromosomal rearrangements. NatGenet 39:319–328.
abuchi K, Blundell J, Etherton MR, Hammer RE, Liu X, Powell CM,
Sudhof TC (2007) A neuroligin-3 mutation implicated in autismincreases inhibitory synaptic transmission in mice. Science 318:71–76.
sujino N, Nakatani Y, Seki Y, Nakasato A, Nakamura M, SugawaraM, Arita H (2007) Abnormality of circadian rhythm accompanied byan increase in frontal cortex serotonin in animal model of autism.Neurosci Res 57:289–295.
incent JB, Kolozsvari D, Roberts WS, Bolton PF, Gurling HM,Scherer SW (2004) Mutation screening of X-chromosomal neuroli-gin genes: no mutations in 196 autism probands. Am J Med GenetB Neuropsychiatr Genet 129B:82–84.
orhees CV (1987) Behavioral teratogenicity of valproic acid: selectiveeffects on behavior after prenatal exposure to rats. Psychophar-macology (Berl) 92:173–179.
agner GC, Reuhl KR, Cheh M, McRae P, Halladay AK (2006) A newneurobehavioral model of autism in mice: pre- and postnatal ex-posure to sodium valproate. J Autism Dev Disord 36:779–793.
hitfield HJ Jr, Brady LS, Smith MA, Mamalaki E, Fox RJ, HerkenhamM (1990) Optimization of cRNA probe in situ hybridization meth-odology for localization of glucocorticoid receptor mRNA in ratbrain: a detailed protocol. Cell Mol Neurobiol 10:145–157.
illiams G, King J, Cunningham M, Stephan M, Kerr B, Hersh JH(2001) Fetal valproate syndrome and autism: additional evidenceof an association. Dev Med Child Neurol 43:202–206.
illiams PG, Hersh JH (1997) A male with fetal valproate syndromeand autism. Dev Med Child Neurol 39:632–634.
an J, Noltner K, Feng J, Li W, Schroer R, Skinner C, Zeng W,Schwartz CE, Sommer SS (2008) Neurexin 1alpha structural vari-ants associated with autism. Neurosci Lett 438:368–370.
an J, Oliveira G, Coutinho A, Yang C, Feng J, Katz C, Sram J,Bockholt A, Jones IR, Craddock N, Cook EH Jr, Vicente A, Som-mer SS (2005) Analysis of the neuroligin 3 and 4 genes in autism
and other neuropsychiatric patients. Mol Psychiatry 10:329–332.(Accepted 9 July 2009)(Available online 14 July 2009)




















