
Postnatal modulation of hepatic biotransformation system enzymes via translactational exposure of F...
7
ELSEVIER CANCER LETTERS Cancer Letters 96 (1995) 87-93 Postnatal modulation of hepatic biotransformation system enzymes via translactational exposure of F, mouse pups to turmeric and curcumin Anjali Singh, S.P. Singh, R. Bamezai* Human Genetics Laboratory, School of Life Sciences, Jawahalal Nehru University, New Delhi 110067, India Received 26 April 1995;revision received 12 July 1995; accepted 13 July 1995 Abstract The potential for the transfer of active principle(s) of turmeric (Curcuma longa L.) and curcumin (major pigment in tur- meric) via translactational route and its modulatory influence on the hepatic biotransformation system enzymes in the lactat- ing dams and their suckling offspring was assessed. Turmeric (4 g/kg b.w. per day) and curcumin (0.4 g/kg b.w. per day) induced significant (P < 0.01) increase in the hepatic levels of glutathione S-transferase (GST) and acid soluble sulfhydryl (-SH) after 14 or 21 days treatment in lactating dams and translactationally exposed F1 pups. However the lower dose of curcumin (0.2 g/kg b.w. per day) could modulate hepatic GST activity (P < 0.05) and -SH (P < 0.01) only after 21 days of treatment in dams and pups. Cytochrome b5 and cytochrome P450 levels were significantly elevated (P < 0.05) in the dams as well as their suckling pups of both 14 and 21 days age groups at the selected dose levels of turmeric (4 g/kg b.w.) and cur- cumin (0.4 g/kg b.w.). The induction in hepatic biotransformation system enzymes in lactating dams and Ft progeny suggests the passage of active constituents and/or metabolites of turmeric and curcumin via the translactational route. Keywords: Biotransformation system; Translactational; Turmeric; Curcumin 1. Introduction A number of micronutrients [ 181, macronutrients [ 171, and non-nutrients [25] have been repotted to block the different stages of the carcinogenic process in animal models and a number of them are on trial in high risk human populations [6,13]. Turmeric, the ground rhizome of Curcuma longa L., has been widely used as a spice and credited with therapeutic properties [2]. The beneficial effects of turmeric are postulated to be due to curcumin * Corresponding author. (diferuloyl methane), the major yellow pigment in turmeric, which possesses antioxidant [15] and anti- carcinogenic [3] effects. Application of curcumin to mouse skin strongly inhibited 12-0-tertadecanoyl phorbol-13-acetate-induced inflammation, epidermal ornithine decarboxylase activity, hyperplasia and formation of hydrogen peroxide [ll]. Dietary ad- ministration of curcumin and turmeric has also been reported to modulate the levels of drug metabolizing system enzymes [5] besides inhibiting lipid peroxi- dation [24]. These findings are particularly important because of the widespread dietary ingestion of tur- meric, and hence its constituent curcumin, in varying proportions and amounts by the human population. 0304-3835/95/$09.50 0 1995 Elsevier Science Ireland Ltd. All rights reserved SSDI 0304-3835(95)03909-G
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Postnatal modulation of hepatic biotransformation system enzymes via translactational exposure of F...

ELSEVIER
CANCER LETTERS
Cancer Letters 96 (1995) 87-93
Postnatal modulation of hepatic biotransformation system enzymes via translactational exposure of F, mouse pups to turmeric and curcumin
Anjali Singh, S.P. Singh, R. Bamezai* Human Genetics Laboratory, School of Life Sciences, Jawahalal Nehru University, New Delhi 110067, India
Received 26 April 1995; revision received 12 July 1995; accepted 13 July 1995
Abstract
The potential for the transfer of active principle(s) of turmeric (Curcuma longa L.) and curcumin (major pigment in tur- meric) via translactational route and its modulatory influence on the hepatic biotransformation system enzymes in the lactat- ing dams and their suckling offspring was assessed. Turmeric (4 g/kg b.w. per day) and curcumin (0.4 g/kg b.w. per day) induced significant (P < 0.01) increase in the hepatic levels of glutathione S-transferase (GST) and acid soluble sulfhydryl (-SH) after 14 or 21 days treatment in lactating dams and translactationally exposed F1 pups. However the lower dose of curcumin (0.2 g/kg b.w. per day) could modulate hepatic GST activity (P < 0.05) and -SH (P < 0.01) only after 21 days of treatment in dams and pups. Cytochrome b5 and cytochrome P450 levels were significantly elevated (P < 0.05) in the dams as well as their suckling pups of both 14 and 21 days age groups at the selected dose levels of turmeric (4 g/kg b.w.) and cur- cumin (0.4 g/kg b.w.). The induction in hepatic biotransformation system enzymes in lactating dams and Ft progeny suggests the passage of active constituents and/or metabolites of turmeric and curcumin via the translactational route.
Keywords: Biotransformation system; Translactational; Turmeric; Curcumin
1. Introduction
A number of micronutrients [ 181, macronutrients [ 171, and non-nutrients [25] have been repotted to block the different stages of the carcinogenic process in animal models and a number of them are on trial in high risk human populations [6,13].
Turmeric, the ground rhizome of Curcuma longa L., has been widely used as a spice and credited with therapeutic properties [2]. The beneficial effects of turmeric are postulated to be due to curcumin
* Corresponding author.
(diferuloyl methane), the major yellow pigment in turmeric, which possesses antioxidant [15] and anti- carcinogenic [3] effects. Application of curcumin to mouse skin strongly inhibited 12-0-tertadecanoyl phorbol-13-acetate-induced inflammation, epidermal ornithine decarboxylase activity, hyperplasia and formation of hydrogen peroxide [ll]. Dietary ad- ministration of curcumin and turmeric has also been reported to modulate the levels of drug metabolizing system enzymes [5] besides inhibiting lipid peroxi- dation [24]. These findings are particularly important because of the widespread dietary ingestion of tur- meric, and hence its constituent curcumin, in varying proportions and amounts by the human population.
0304-3835/95/$09.50 0 1995 Elsevier Science Ireland Ltd. All rights reserved SSDI 0304-3835(95)03909-G

88 A. Singh et al. I Cancer Letters 96 (1995) 87-93
As the postnatal period of development represents the potential pathway for a number of xenobiotic modulators, in their proximate/metabolized forms, to be transferred to the Fi generation via P generation female [ 11, the modulation in competing pathways of biotransformation system enzymes in lactating dams may affect the rate and extent of maternal detoxica- tion and, thus, influence the translactational passage of metabolites of an administered compound. In the present study the potential for the transfer of active principle(s)/metabolites of turmeric and curcumin via the translactational route was assessed following the modulatory influence on the hepatic biotransforma- tion system enzymes in dams and their suckling neo- nates.
2. Materials and methods
2.1. Animals Random-bred Swiss albino female mice (28-30 g)
with their first born litters were housed in separate cages and maintained in an air-conditioned room of J.N.U. Animal Facility, New Delhi. The mice were provided with (unless otherwise stated) standard food pellets (Hindustan Lever Ltd., India) and tap water ad libitum.
2.2. Chemicals Bovine serum albumin (BSA), 1-chloro-2,4-dini-
trobenzene (CDNB), Coomassie brilliant blue (G- 250), curcumin, 5,5’-dithiobis(2-nitrobenzoic acid) (DTNB), glutathione reduced (GSH), nicotinamide adenine dinucleotide reduced (NADH) and 5-sulfo- salicylic acid were obtained from Sigma Chemical Co., St. Louis, MO, USA. All other chemicals used were of highest available grade of purity obtained from local firms.
Turmeric powder (MDH, a commercially avail- able brand) was obtained from the local market in New Delhi, India.
2.3. Experimental design Fig. 1 depicts the groups maintained and treatment
schedule followed. The lactating dams (n = 6/group) each with suck-
ling pups were assorted into control and experimental groups, maintaining the number of pups per dam as 6 by redistribution before commencing the experiment.
Curcumin (0.2 or 0.4 g in corn oil/kg b.w. per day) or turmeric (4 g in corn oil/kg b.w. per day) was delivered by gavage to dams from day 1 of lactation and continued for 14 or 21 days.
The body weight of the animals was recorded ini- tially and at the time of sacrifice. Dams were starved overnight prior to sacrificing for the biochemical analysis of liver.
2.4. Hepatic studies The animals were sacrificed by cervical disloca-
tion and their livers were excised. Livers from pups of each litter were pooled. After perfusion with chilled normal saline (0.95), a 10% (w/v) homogen- ate was made using buffer containing 0.154 M KC1 and 50 mM Tris-HCl (pH 7.4). A part of the ho- mogenate was used to measure acid soluble sulfhy- dry1 groups. The rest was centrifuged at 10 000 rev./ min (10 086 g) for 20 min. The resultant supernatant was centrifuged at 40 000 rev./min (105 000 g) for
0lO”PS TP.UATNUWT SCBllDULE (DAYS) --
Lactation Period G------------------------------~
A-- 7 14. 21
I CONTROL 4 ~~~~~~c”“-!‘_‘--------,
1 1
Curcumin O.Zg/kg b.u./dsy 2 EXPEPJHENTAL
~--------------------->
I -4
Curcumin O.Og/kg b.v./day
3 EXPgRIHENTAL f---------------------)
-- I
Turmeric 4glkg b.v.,dsy
4 EXPERIMENTAL (---------------------)
I I
corn Oil
5 CONTROL ~-------------------------------)
, ------4
Curcumin O.Zg/kg b.u./day
6 EXPggIIgNTAL (--------------------------------,
1 4
Curcumin 0.4glkg b.v./day
7 EXPERMENTAL c--------------------------------,
I---------- -4
Turmeric 4glkg b.w.iday
8 EXPERIPlENTAL (--------------------------------)
I---------------___,
Fig. 1. Diagrammatic representation of the experimental protocol.
umoh of CDNB-QSH conJgate tormed/mln /mq Drotein
A. Sir@ et al. I Cancer Letters 96 (1995) U-93 89
a 14 Days Treatment b 21 Days Treatment
umoh Of CDNB-QSH conJ~t6 torm6dhnlnlmg protein
1
I L
** P* 0.01 - Ps 0.06 -* PI 0.01
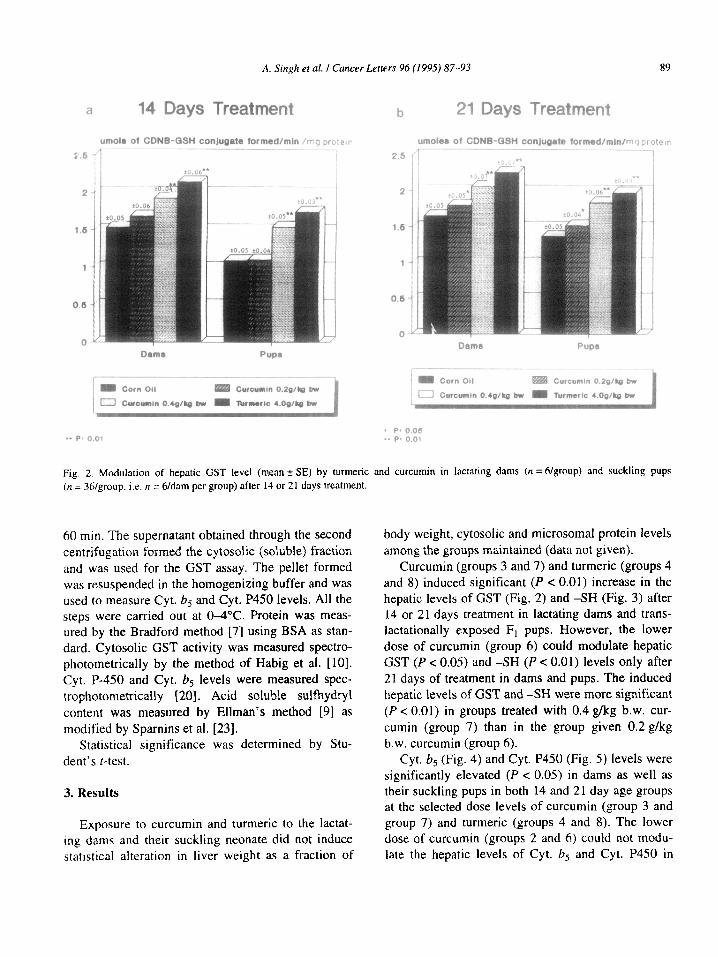
Fia. 2. Modulation of hepatic GST level (mean + SE) by turmeric and curcumin in lactating dams (n =6/group) and suckling pups (n = 36/group. i.e. n = 6/dim per group) after 14 or 21 days treatment.
60 min. The supernatant obtained through the second centrifugation formed the cytosolic (soluble) fraction and was used for the GST assay. The pellet formed was resuspended in the homogenizing buffer and was used to measure Cyt. b5 and Cyt. P450 levels. All the steps were carried out at 0-4”C. Protein was meas- ured by the Bradford method [7] using BSA as stan- dard. Cytosolic GST activity was measured spectro- photometrically by the method of Habig et al. [lo]. Cyt. P-450 and Cyt. b5 levels were measured spec- trophotometricaily (201. Acid soluble sulfhydryl content was measured by Ellman’s method [9] as modified by Sparnins et al. [23].
Statistical significance was determined by Stu- dent’s t-test.
3. Results
Exposure to curcumin and turmeric to the lactat- ing dams and their suckling neonate did not induce statistical alteration in liver weight as a fraction of
body weight, cytosolic and microsomal protein levels among the groups maintained (data not given).
Curcumin (groups 3 and 7) and turmeric (groups 4 and 8) induced significant (P < 0.01) increase in the hepatic levels of GST (Fig. 2) and -SH (Fig. 3) after 14 or 21 days treatment in lactating dams and trans- lactationally exposed F, pups. However, the lower dose of curcumin (group 6) could modulate hepatic GST (P < 0.05) and -SH (P < 0.01) levels only after 21 days of treatment in dams and pups. The induced hepatic levels of GST and -SH were more significant (P < 0.01) in groups treated with 0.4 g/kg b.w. cur- cumin (group 7) than in the group given 0.2 g/kg b.w. curcumin (group 6).
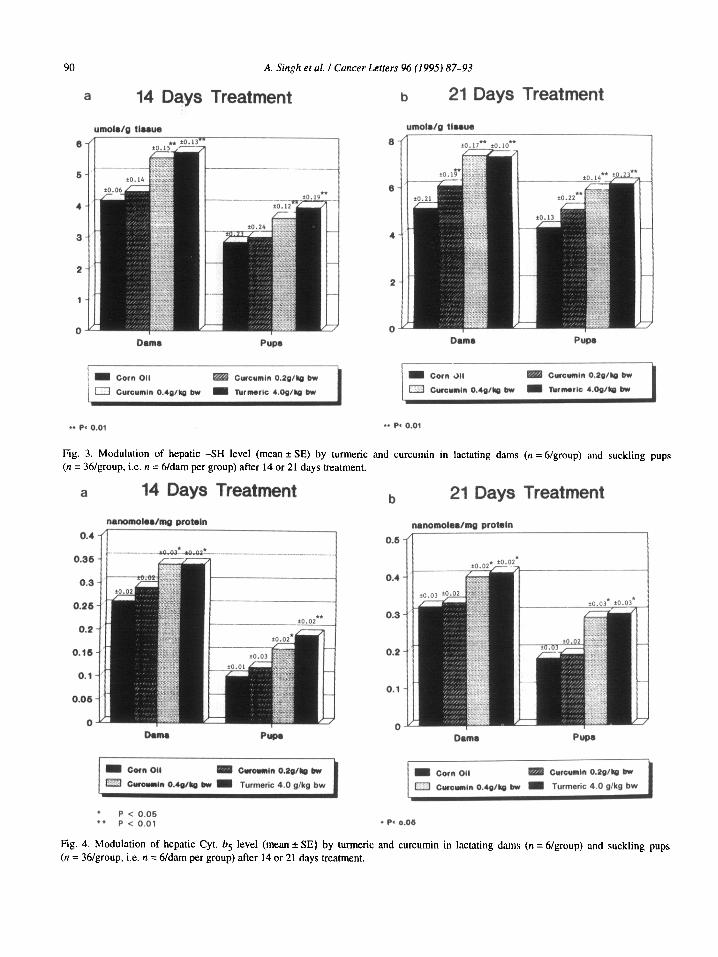
Cyt. b5 (Fig. 4) and Cyt. P450 (Fig, 5) levels were significantly elevated (P < 0.05) in dams as well as their suckling pups in both 14 and 21 day age groups at the selected dose levels of curcumin (group 3 and group 7) and turmeric (groups 4 and 8). The lower dose of curcumin (groups 2 and 6) could not modu- late the hepatic levels of Cyt. b, and Cyt. P450 in
90 A. Singh et ul. I Cuncer Letters 96 (1995) 87-93
a 14 DEI~S Treatment b 21 Days Treatment
umol81a tlNu0
** Pa 0.01 ** Pa 0.01
Fig. 3. Modulation of hepatic -SH level (mean ?T SE) by turmeric and curcumin in lactating dams (n =6/group) and suckling pups (n = 36/group, i.e. n = 6/darn per group) after 14 or 21 days treatment.
a 14 Days Treatment b
0.66
0.3 0.4
0.26 0.3
0.2
0.16 0.2
0.1
0.1 0.06
0 n Dams
”
I
I m Corn 011 I Cwomln 0.2gl@ bw
m CUommln 0.1011p k m Turnwic 4.0 alka bw
21 Days Treatment
~nomolu/mg protein
I- *
Dama
m Corn Oil I Cwowln O.PoIIo b*,
m Cwcullr OAg/tq bw m Turmeric 4.0 g/kg bw I
l P < 0.05 l * P < 0.01 l PI 0.0s
Fig. 4. Modulation of hepatic Cyt. bg level (mean + SE) by turmeric and curcumin in lactating dams (n = 6/group) and suckling pups (n = 36/group, i.e. n = 6/darn per group) after 14 or 21 days treatment,

A. Singh et al. I Cuncer Letters 96 (I 995) 87-93 91
a 14 Days Treatment
Lonolnoloo/nlg protoln b 21 Days Treatment
nM0m0lo0/lag protoln 0.8 l
tn.nPt0.03
* p < 0.05
li* p < 0.01
:. 'p: "0:;:
Fig. S. Modulation of hepatic Cyt. P450 level (mean ? SE) by turmeric and curcumin in lactating dams (n = 6/group) and suckling pups (n = 36/group, i.e. n = B/dam per group) after. 14 or 21 days treatment
lactating dams and translactationally exposed Fi
pups.
4. Discussion
The postnatal induction of biotransformation sys- tem enzymes in neonatal tissues, by chemopreventive agents delivered via translactational route, can pre- vent the process of initiation or promotion brought about by perinatal exposure to a carcinogen [ 1,211. A means of identifying potential chemopreventive agents in dietary constituents, besides their mkcha- nism(s) of action, is their capacity to induce in- creased levels of enzymes that can detoxify chemical carcinogens [19,26]. These enzymes appear in the later part of gestation and early postnatal life as de- velopmental clusters [22], and are inducible in fetal and neonatal tissues by direct or maternal exposure to carcinogens [4] or by certain dietary factors [ 121.
The bifunctional inducing activity of curcumin and turmeric is evident via inductive effects on he-
patic phase I and phase II system enzymes of bio- transformation and -SH content in lactating dams and translactationally exposed Fi pups.
The observed enhancement of hepatic GST level in dams and suckling pups suggests the chemopre- ventive role of curcumin and turmeric through a blocking mechanism, as phase II catalyzed conjuga- tion reactions offer hydrophilicity to xenobiotics for their metabolic disposition [8]. A number of studies have also suggested the significant correlation be- tween elevated GST level and a concurrent reduction in carcinogenic response [ 8,141.
In lactating dams and suckling pups, the aug- mented -SH level by curcumin and turmeric supports the correlation between elevated SH level and anti- carcinogenesis [16], as its potential in protecting the cells against free radicals, besides acting as an accep- tor of reactive electrophilic moieties, has been well documented [14].
As the co-induction of phase I and phase II sys- tem enzymes is projected to be highly protective

92 A. Singh et al. I Cancer Letters 96 (1995) 87-93
[ 19,261, thus, the cjlrcumin and turmeric-induced levels of Cyt. b, an& Cyt. P450 in dams and pups may offer a potential approach for prevention of car- cinogenesis.
In view of the enhanced detoxication ability of bi- functional inducers in most instances [26], the poten- tial chemopreventive role of turmeric and its major pigment-curcumin is predicted which further sug-
gests that dietary manipulation during the critical postnatal period of development may modulate sus- ceptibility to xenobiotics: (a) in the Ft progeny fol- lowing the translactational passage of active constitu- ents and/or metabolites or modulators; (b) in the lac- tating dams by substantially detoxifying the adminis- tered compounds besides reducing the translacta- tional passage of potential toxins to the neonate.
As the translactational route permits effective moieties of chemopreventive agents in the suckling neonate [ 1,2 11, there exists the possibility of preven- tion of the passage of toxic effects of xenobiotics that might enter the progeny. Further studies evaluating the translactational modifying effect of potent car- cinogens on potential chemopreventive efficacy of curcumin in murine model system are in progress.
Acknowledgements
Financial assistance by the Council for Scientific and Industrial Research, India, to Anjali Singh and by University Grants Commission to S.P. Singh as Research Associates is gratefully acknowledged.
References
[l] Alexandrov, V., Aiello, C. and Rossi, L. (1990) Modifying factors in prenatal carcinogenesis. In Vivo, 4, 327-336.
[2] Ammon, H.P.T. and Wahl, M.A. (1990) Pharmacology of Curcuma bnga. Planta Med., 57, l-7.
[3] Ammon, H.P.T. and Wahi, M.A. (1991) Pharmacology of curcumin. Plant Med., 57, l-7.
[4] Anderson, L.M., Jones, A.B., Miller, MS. and Chauhan, D.P. (1989) Metabolism of transplacental carcinogens. In: Perinatal and Multigeneration Carcinogenesis. IARC Scien- tific Publication No. 96, pp 155-188. Editors: N.P. Napal- kov, J.M. Rice, L. Tomatis and H. Yamasaki. IARC Scien- tific, Lyon.
[5] Azuine, M.A. and Bhide, S.V. (1992) Chemopreventive effect of turmeric against stomach and skin tumors induced by chemical carcinogens in Swiss mice. Nutr. Cancer, 17, 77-83.
[6] Blot, W.J., Li, J.-Y., Taylor, P.R., Guo, W., Dawsey, S.,
[71
PI
[91
Ito1
I111
[I21
u31
1141
1151
tt61
[I71
1181
u91
Wang, G.-Q., Yang, C.-S., Zhang, S.-F., Gail, M., Li, G-Y., Yu, Y., Liu, B.-Q., Tangrea, J., Sun, Y.-H., Liu, F., Frau- meni, I.F.J., Zhang, Y.-H. and Li, B. (1993) Nutrition inter- vention trials in Linxian, China: supplementation with spe- cific vitamin/mineral combinations, cancer incidence, and disease-specific mortality in the general population. J. Nab. Cancer Inst., 85, 1483-1491. Bradford, M.M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72, 248- 254. Coles, B. and Ketteter, B. (1990) The role of glutathione and glutathione tmnsferases in chemical carcinogenesis. Crit. Rev. Biochem. Mol. Biol., 25.47-70. Ellmann, G.L. (1959) Tissue sulthydryl groups. Arch. Bio- them. Biophys., 82,70-77. Habig, W.H., Pabst, M.J. and Jakoby, W.B. (1974) Glu- tathione S-transferases. The first enzymatic step in mercap- turic acid formation. J. Biol. Chem., 249.7130-7139. Huang, M.-T., Robertson, F.M., Lysz, T., Ferraro, T., Wang, Z.Y., Georgiadis, CA., La&in, J.D. and Conny, A.H. (1992) Inhibitory effects of curcumin on carcinogenesis in mouse epidermis. In: Phenolic Compounds in Food and Their Ef- fects on Health II: Antioxidants and Cancer Prevention, pp. 339-349. Editors: M.-T. Haung, C-T. Ho and C.Y. Lee. American Chemical Society, Washington, DC. Karnik, H.B., Sonawane, B.R., Adkins, J.S. and Mohla, S. (1990) High dietary fat feeding during perinatal develop- ment of rats alters hepatic drug metabolism of progeny. Dev. Pharmacol. Ther., 14, 135-140. Kelloff, G.J., Boone, C.W., Malone, W.E. and Steele, V.E. (1992) Chemoprevention clinical trials. Mutat. Res., 267, 292-2.95. Ketterer, B. (1988) Protective role of glutathione and glu- tathione transferases in mutagenesis and carcinogenesis. Mutat. Res., 202, 343-361. Kunchandy, E. and Rao, M.N.A. (1990) Oxygen radical scavenging activity of curcumin. Int. J. Pharm., 58, 237- 240. Meister, A. (1994) Glutathione, ascorbate and cellular pro- tection. Cancer Res., 54s. 1969-1975. Micozzi, M.S. and Moon, T.E. (Editors) (1992) Macronutri- ents: Investigating Their Role in Cancer. Marcel Dekker, New York. Moon, T.E. and Micozzi, MS. (Editors) (1988) Nutrition and Cancer Prevention: Investigating the Role of Micronu- trients. Marcel Dekker, New York. Nijhoff, W.A., Groen, G.M. and Peters, W.H.M. (1993) Induction of rat hepatic and intestinal glutathione S-tmns- ferases and glutathione by dietary naturally occurring anti- carcinogenesis. Int. J. Oncol., 3, 1131-1135.
[20] Omura, T. and Sate, R. (1964) The carbon monoxide bind- ing pigment of liver microsomes. J. Biol. Chem., 239,2370- 2378.
[21] Rao, A.R. (1989) 7,12-Dimethylbenz(a)anthracene-induced perinatal carcinogenesis and its modulation by butylated hy- droxyanisole in mice. In: Perinatal Multigeneration Car- cinogenesis. IARC Scientific Publication No. 96, pp. 63-73.

A. Singh et ~11. I Cuncer Letters 96 (1995) 87-93 93
Editors: N.P. Napalkov, J.M. Rice, L. Tom&is and H. Ya- ma&i. IARC Scientific, Lyon.
[22] Snell, K. (1982) Developmental enzyme pathology. In: Developmental Toxicology, pp. 299-399. Editor: K. Snell. Praeger, New York.
[23] Spamins, V.L., Venegas, P.L. and Wattenberg, L.W. (1982) Glutathione S-transferase activity: enhancement by com- pounds inhibiting chemical carcinogenesis and by dietary constituents. J. Natl. Cancer Inst., 68,493496.
[24] Tonnesen, H.H., Smistad, G., Agren, T. and Karlsen, J. (1993) Studies on curcumin curcuminoids. XXIII: Effects of curcumin on liposomal lipid peroxidation. Int. J. Pharm., 90, 221-228.
[25] Wattenberg, L.W. (1990) Inhibition of carcinogenesis by minor anutrient constituents of the diet. Proc. Nutr. Sot., 49, 173-183.
[26] Wattenberg, L.W. (1992) Inhibition of carcinogenesis by minor dietary constituents. Cancer Res., 52s. 20852091.




















