
Biotransformation of Ni-Substituted Hydrous Ferric Oxide by an Fe(III)-Reducing Bacterium
Upload
independentCategory
view
0download
0
lable at ScienceDirect
Biomaterials 32 (2011) 3988e3999
Contents lists avai
Biomaterials
journal homepage: www.elsevier .com/locate/biomateria ls
Long term in vivo biotransformation of iron oxide nanoparticles
Michael Levy a, Nathalie Luciani a, Damien Alloyeau b, Dan Elgrabli c, Vanessa Deveaux d,Christine Pechoux e, Sophie Chat e, Guillaume Wang b, Nidhi Vats a, François Gendron f,Cécile Factor g, Sophie Lotersztajn d, Alain Luciani d,h, Claire Wilhelm a, Florence Gazeau a,*
a Laboratoire Matières et Systèmes Complexes (MSC), UMR 7057 CNRS/Université Paris - Diderot, 10 rue Alice Domon et Léonie Duquet, 75205 Paris cedex 13, Franceb Laboratoire Matériaux et Phénomènes Quantiques (MPQ), UMR 7162 CNRS/Université Paris - Diderot, 10 rue Alice Domon et Léonie Duquet, 75205 Paris cedex 13, Francec INERIS, BP2, 60550 Verneuil en Halatte, Franced Inserm U955, Equipe 17, CHU Henri Mondor, 51 Avenue du Maréchal de Lattre de Tassigny, 94010 Créteil Cedex, FranceeMIMA2- UR1196 Génomique et Physiologie de la Lactation, INRA, Plateau de Microscopie Electronique 78352 Jouy-en-Josas, Francef Institut des Nanosciences de Paris (INSP), UMR 7588 CNRS/Université Pierre et Marie Curie, 4 place Jussieu, 75205 Paris, FrancegGUERBET, BP57400, 95943 Roissy CdG Cedex, FrancehAP-HP, Groupe Henri Mondor Albert Chenevier, Imagerie Médicale, 51 Avenue du Maréchal de Lattre de Tassigny, 94010 Créteil Cedex, France
a r t i c l e i n f o
Article history:Received 3 February 2011Accepted 15 February 2011Available online 9 March 2011
Keywords:NanoparticlesNanomagnetismNanotoxicityBiodegradationIron oxideContrast agent
* Corresponding author. Tel.: þ33 01 57 27 62 03;E-mail address: florence.gazeau@univ-paris-didero
0142-9612/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.biomaterials.2011.02.031
a b s t r a c t
The long term outcome of nanoparticles in the organism is one of the most important concerns raised bythe development of nanotechnology and nanomedicine. Little is known on the way taken by cells toprocess and degrade nanoparticles over time. In this context, iron oxide superparamagnetic nano-particles benefit from a privileged status, because they show a very good tolerance profile, allowing theirclinical use for MRI diagnosis. It is generally assumed that the specialized metabolism which regulatesiron in the organism can also handle iron oxide nanoparticles. However the biotransformation of ironoxide nanoparticles is still not elucidated. Here we propose a multiscale approach to study the fate ofnanomagnets in the organism. Ferromagnetic resonance and SQUID magnetization measurements areused to quantify iron oxide nanoparticles and follow the evolution of their magnetic properties.A nanoscale structural analysis by electron microscopy complements the magnetic follow-up of nano-particles injected to mice. We evidence the biotransformation of superparamagnetic maghemite nano-particles into poorly-magnetic iron species probably stored into ferritin proteins over a period of threemonths. A putative mechanism is proposed for the biotransformation of iron-oxide nanoparticles.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
The long term outcome of nanoparticles (NPs) in the body isa major concern, which has been raised by the recent developmentof nanotechnology and nanomedicine. How nanomaterials interactwith living constituents and are processed by the organism arejustifiably the most-asked questions whenever an application ofnanomaterials is proposed [1]. Safety issues are becoming prom-inent in the development of new nanodevices, especially in thebiomedical field. Inorganic nanoparticles that exhibit unique opticalor magnetic properties are of special interest for diagnosis andtherapeutic applications exploiting their response to physical,remotely activated, stimuli [2,3]. However, while tremendousadvances have beenmade in the tuning of the chemical and physicalhallmarks of NPs, their fate once injected in the organism is still not
fax: þ33 01 57 27 62 11.t.fr (F. Gazeau).
All rights reserved.
controlled. In particular, the transformations induced by cellularactivity on inorganic nanoparticles are largely unknown yet.
Most studies carry out in vitro assessments of NPs toxicity usingcellular models [1,4e6]. They point out some critical parametersgoverning the biological response to nanomaterials, such as theirsize, composition, surface reactivity, architecture, stability withrespect to the biological medium and intracellular distribution.However, such studies always require validation in vivo, wheremost of these parameters are no longer controlled. Hence reliablemethods are eminently desirable to monitor the fate of inorganicnanoparticles directly in the organism.
One approach to assess the in vivo behaviour of inorganic nano-particles is to rely on their most salient properties, their physicalproperties, which precisely make their interest for applications. Forexample, optical properties of plasmonic or luminescent nano-particles can be exploited to study their biodistribution [7]. Similarly,magnetic properties of iron oxide NPs enable their follow-up byMRI,a particularly attractive non-invasive imaging technique, paving the

M. Levy et al. / Biomaterials 32 (2011) 3988e3999 3989
way of many diagnostic applications [8]. However a quantitativerelationship between the physical signal and the amount of nano-particles is often compromised, because the physical properties maybe impacted by the biological environment or by the peculiar orga-nization adopted by nanomaterials when immerged in the bloodflow or processed by cells [9]. It is particularly true for MRI whichrelies on the indirect effect of the magnetic field produced by nano-particles on the dynamics of surrounding protons [10]. Thereforea reliable monitoring of nanoparticles should be based on theirspecific physical properties, independently of their environment.
Herewepropose to refer on the superparamagnetic properties ofnanomagnets to monitor their fate in vivo. We perform comple-mentary techniques of nanomagnetism in order to specificallyquantify iron oxide NPs based on their magnetic moment and toassess their degradation and biotransformation over time. Theprogressive modification and loss of their characteristic magneticproperties are interpreted as the signatures of the transformationsoperated by cells on nanoparticles. Because the degradation obvi-ously occurs at a nanometer scale, the magnetic follow-up is herecomplemented by a nanoscale structural characterization of theintracellular outcome of nanoparticles. Our multiscale approach, toevaluate biologically-induced changes of the nanoparticle state,applies here to iron oxide NPs used as MRI contrast agent. Althoughiron-based NPs have never shown acute or sub-acute toxicities, theway they are handled by the iron metabolism is still not elucidated[11e14]. The present results are thus directly relevant to currentsafety considerations in clinical diagnostic and therapeutic uses ofnanomagnets.
2. Materials and methods
2.1. Design of the study
In this study, we aimed to assess the biodistribution and outcome of P904nanoparticles (Guerbet SA) administrated intravenously tomice. P904 nanoparticlesconsist in 8 nm superparamagnetic cores of maghemite coated by hydrophilicderivatives of glucose. These nanoparticles are currently developed by Guerbet SA,as a contrast agent for non-invasive imaging of inflammation by MRI. The diagnosisand therapeutic follow-up of various pathologies involving inflammatory processes,such as atherosclerosis or obesity, may benefit from the development of P904 ironoxide nanoparticles. P904 NPs are expected to be uptaken by cells of the reticulo-endothelial system, especially the macrophages which are present or recruited ininflammatory areas, allowing their non-invasive detection [15e17].
In our study, we tested two doses to evaluate the long term fate of nanoparticles inmice: 1000 mmol of iron/kg, the dose previously proposed for the detection ofinflammation in atherosclerotic plaques [16] and 50 mmol of iron/kg, the potentially-useddose inhumanclinic. P904nanoparticleswere followedduring threemonthsafterinjection at low dose in normal healthy mice (C57-B6 mice) and during 44 days afterinjection at high dose in obese mice (ObOb leptin-deficient mice). We choose obesemice to assess if inflammatory macrophages located within adipose tissue can uptakeP904 nanoparticles and thus allow quantification of obesity-related inflammation.
Besides their imaging applications, P904 nanoparticles are representative ofa class of sugar-coated iron oxide ferrimagnetic nanoparticles, with a typical sizearound 8 nm, which have found applications for magnetic drug targeting (whenassociated with a drug carrier) [18], cell targeting (when internalized in cells)[19,20], magnetic hyperthermia [21] or magnetically-assisted drug delivery [22]. Inall these applications, a precise knowledge of the long term in vivo outcome of NPs ismandatory.
2.2. Magnetic nanoparticles
Superparamagnetic P904 NPs were provided by Guerbet (Roissy, France). Theyare synthesized by coprecipitation of ferric and ferrous ions in alkaline mediumfollowing a classical procedure [23]. They consist in a ferrimagnetic core of maghe-mite coated by a hydrophilic derivative of glucose. The magnetic size distributions,deduced from the field-dependentmagnetization curve, fit a log-normal distributionwith characteristic diameter d0 ¼ 7:2 nm and polydispersity index s ¼ 0.24.
2.3. Animal model
All animal experiments were conducted in keeping with French AgricultureMinistry guidelines. A total of 62 mice were used in the study: 30 of themwere male8-weeks-old C57/Bl6 mice (Janvier, France) (mean weight 25 � 3 g), the other 32
mice were 8-weeks old leptin-deficient “ob/ob” mice (B6.V-Lepob, Janvier, France)showing obesity (weight range 47 � 5 g). 25 C57/Bl6 mice and 27 obob mice wereinjected with P904 nanoparticles respectively at a dose of 50 mmol/kg (low dose) and1000 mmol/kg (high dose) via a retro-orbitary route. 5 C57/Bl6 mice and 5 obob miceserved as control. Five injected C57/Bl6 mice were sacrificed at each different time-points after P904 injection (Days, D1, D7, D30, D60 and D90). Six injected obob micewere sacrificed at D3, D10, D31 and D44 and only 2 mice at D24. Liver, spleen andinflammatory adipose tissue (only for obob mice) were excised and shared for ICP-OES, FMR, SQUID and TEM characterizations.
2.4. Iron quantification by FMR and ICP-OES
Ferromagnetic Resonance was performed with a Varian E102 EPR (ElectronParamagnetic Resonance) spectrometer operating at Q band (9.26 GHz). The firstderivativeof thepowerabsorptiondW(B)/dBwas recordedas a functionof theappliedfield B in the range 0e6000 Gauss. The modulation field had a frequency of 100 kHzandanamplitudeof 10G. Theareaof the FMRabsorptioncurve (calculatedbyadoubleintegration of the spectrum dW(B)/dB) was proportional to the amount of P904nanoparticles in the sample. Anabsolute calibrationwasperformedusing suspensionsof P904 nanoparticles (2 ml sample) at different concentrations quantified by ICP-OES[24]. Before FMR measurements, the excised pieces of organs were first weighed,rinsed, finely sliced and let dry for 3 days in an oven at 80 �C. The dried organ sampleswere weighed again and introduced into a quartz tube suitable for FMR measure-ments. For adipose tissue, the dehydrated tissue was sucked in a PTFEmicrotube andthenpushedaway in thebottomof thequartz tube. Thenetmass of dehydratedorgansin the quartz tube was then weighed and FMR spectra were recorded.
Total iron content in organs was quantified by inductively coupled plasma -optical emission spectrometry (ICP-OES). After excision, samples were conservedat�80 �C until preparation for ICP-OESmeasurements. The organs weremineralizedin 3.5 ml of nitric acid and 1.5 ml of chloridric acid for 4 h at 100 �C and then treatedwith 1.5ml of H2O2 for 1 h at 95 �C on a heat block. The volumewas adjusted to 10mlwith distilled water and samples were analyzed for iron content by ICP-OES (Ultima,HORIBA Jobin Yvon, Edison, NJ).
2.5. SQUID magnetization measurements
Magnetization measurements were carried out on a Quantum Design MPMS-5SSQUID magnetometer. Field-dependent magnetization curves were measured at300 K as a function of the external magnetic field in the range 0e3�104 Gauss.Temperature-dependent magnetization was measured at low field (50 Gauss) fortemperatures increasing from 5 K to 300 K for field cooled (FC) and zero-field cooled(ZFC) samples. Magnetic field during cooling was 50 Gauss. Dried organs wereprepared as for FMR quantification and introduced in an SQUID gelatin capsule formeasurements. For spleen and adipose tissue, the samples stemming from thedifferent mice sacrificed at each time point (n ¼ 5) were pooled. For liver samples,SQUID measurements were performed for different individual mouse to demon-strate reproducibility among animals. In all cases, the (temperature-independent)diamagnetic signal coming from the capsule and tissue was subtracted from themeasured magnetization.
2.6. Transmission electron microscopy (TEM)
For electron microscopic analysis, organs were cut into 1 mm3 pieces afterexcision and fixedwith 2% glutaraldehyde in 0.1 M Na pH 7.2 cacodylate buffer for 4 hat room temperature and then postfixed with 1% osmium tetroxide containing 1.5%potassium cyanoferrate, contrasted with uranyl acetate 2% in water, graduallydehydrated in ethanol (30%e100%) and embedded in Epon. For a previsualization(specially to locate macrophages in adipose tissue), 1 micron sections were collectedonto glass slides, conterstained with methylen blue-Azur II and imaged on an epi-fluorescent microscope (DMRB e Leica - France) with a 63x plan apochromat oilimmersion lens. Thin sections (70 nm) of selected zones were collected onto 200mesh cooper grids, and conterstained with lead citrate before examination withZeiss EM902 electron microscope operated at 80 kVe (MIMA2- UR1196 Génomiqueet Physiologie de la Lactation, INRA, Plateau de Microscopie Electronique 78352Jouy-en-Josas, France).
High resolutionmicrographswere acquiredwith a charge-coupled device cameraMegaView III CCD camera and analysed with ITEM software (Eloïse e SARL e RoissyCDG e France). High magnification TEM, electron diffraction and X-ray energy-dispersive spectroscopy (XEDS) experiments were carried out on a Technai G2F20field-emission electron microscope operating at 200 kV and equipped with anenergy-dispersive X-ray analyzer. XEDS spectrum imaging combines the imaging andanalytical capabilities of electron microscopy. A dark field image in scanning mode(STEMmode) of the area of interest is first collected and this image is divided into anarray of pixelswith a userdefined resolution (here 100 by 100). The nanometric STEMprobe is then scanned over the same area, but stopped for a pre-determined timeinterval (here 500 ms), at each pixel. During this time interval, an entire X-ray spec-trum from 0 to 20 keV is acquired. The resulting array of spectra is known as a spec-trum image data cube. Chemical maps were reconstructed from the data cubes with

M. Levy et al. / Biomaterials 32 (2011) 3988e39993990
theTechnai ImagingAnalysis (TIA) softwaredevelopedby FEI.Weused theFeKaX-rayline at 6.4 keV to generate the Fe chemical map.
3. Results
3.1. Specific quantification of superparamagnetic nanoparticlesby ferromagnetic resonance
We first evaluate Ferromagnetic Resonance as a quantitativemethod to monitor iron oxide nanoparticles in tissues. Ferromag-netic Resonance (FMR) is a spectroscopic method to probe thedynamics of magnetization of unpaired electrons collectivelycoupled in a ferro- or ferrimagnetic lattice. For single domain NPs,resonance arises from the precessional motion of their globalmagnetic moment, which experiences the external magnetic fieldB0, plus the internal local fields due to anisotropy energy barriers.FMR behaviour of 4e20 nm maghemite nanoparticles is thus finelytuned by the particle size, appearing as a signature of the super-paramagnetic state of nanoparticles [25]. FMR provides a quantita-tive and sensitive tool tomeasure thequantityof superparamagneticNPs in a small volume sample (2 ml for a liquid suspension or a fewmg of dried sample) [26]. The area of the FMR absorption curve(calculated by a double integration of the spectrum dW/dB) isproportional to the concentration of superparamagnetic NPs [10].We show here that FMR spectra of dried organs from mice injectedwith P904 display a similar wide line as the non-injected NPs incolloidal suspension (Fig.1A). Following cellular uptake of P904, theresonance line is only slightlyenlarged, due tomagnetic interactionsexperienced byNPs sequestered into intracellular compartments. Bycontrast, this characteristicwide line centered at a resonancefield ofHres¼ 3000G is not observed in organs of control non-injectedmice.In control liver, a sharp narrow line at Hres ¼ 3300 G (g ¼ 2) onlyappears when the gain is amplified, presumably attributed to thepresence of free radicals [27] (Inset of Fig. 1A). However, as soon asP904 NPs are present, the endogenous sharp line is no more visibleand has a negligible contribution to the FMR absorption power. Weconfirm that FMR quantification is purely specific to the presence ofiron organized in a superparamagnetic nanocrystal (i.e. Super-Paramagnetic (SP) iron), with a detection sensitivity of 10�2 mg or1.8 � 10�4 mmol of iron (corresponding to an FMR signal of 4). Bycontrast, diamagnetic and paramagnetic free iron ions (stemmingfrom Fe2þ ions of desoxihemoglobin or from transferrin-bond non-hem Fe3þ) do not contribute to the FMR absorption power at roomtemperature and the antiferromagnetic contribution of ferritin, themajor form of iron storage, is shown to be negligible [27].
FMR quantification is used here to monitor P904 NPs (i.e. Super-Paramagnetic (SP) iron) in the major sites of nanoparticle uptake.Over the course of threemonths after the low dose administration ofP904 (injected dose: 61 mg), liver and spleen show a significantdecrease of their SP iron content (Fig. 1B). At day 1, 49% of the totalinjected iron (low dose) is sequestered in the liver and 5.2% in thespleen. Note that the SP iron concentration per gram of organ ishigher in the spleen than in the liver (46.2 mg of iron/g of spleenversus 22.9 mg/g of liver)). SP iron in the liver fall to 21% of totalinjected dose at day 7, 2.3% at day 30 and only 0.25% after threemonths. In the spleen, a slight increase of SP iron concentration isobserved at day 7 and the further decrease is less steep than in theliver. Three months after injection, the amounts of SP iron stillpresent in the liver and in the spleen represent respectively 0.5% and6.6% of the content at day 1.
The time decrease of SP iron concentration is also demonstratedafter injection of high dose of P904 (injected dose: 2630 mg), butwith a slower kinetics. Liver and spleen concentrate 29% and 2% ofthe dose injected by day 3 post-injection. By day 44, SP ironconcentration still represents more than 40% of the concentrations
at day 3 in the liver and in the spleen. Compared to the low doseinjection, liver and spleen are unable to process the high dose ofinjected NPs, probably because of a saturation effect of the degra-dation or elimination mechanisms. After high dose injection, theadipose tissue of obese mice show a measurable content of SP iron,representing about 0.2% of the injected dose (1.2 mg of iron/g ofadipose tissue by day 3). Hence FMR quantifies a significant uptakeof P904 nanoparticles into the targeted inflammation sites, likelyperformed by recruited inflammatory macrophages. Over time,the SP iron content decreases in adipose tissue, but with a largevariability.
3.2. Total iron quantification
Inductively coupled plasma - optical emission spectrometry(ICP-OES) measures the total amount of elemental iron in a sample.Despite its sensitivity, this spectroscopic method suffers from a lackof specificity, because it is unable to separate iron contained inthe injected exogenous NPs from the endogenous iron pool of theorganism. ICP-OES measurements are carried out to compare thetotal iron concentration in organs to the concentration of exoge-nous P904 NPs (or SP iron) quantified by FMR. As shown in Fig. 2A,the endogenous total iron detected by ICP in control organs is muchlarger than the exogenous SP iron injected at clinical dose (50 mmolof iron/kg). Actually, the quantity of SP iron in liver and spleen is ofthe same order of magnitude as the standard deviation of the totaliron detected by ICP. For example, the total amount of injected SPiron is less than 60% of the endogeneous iron in liver. As a conse-quence, ICP is unable to detect the time decrease of P904 SP ironconcentration which is clearly observed with FMR quantification.For the high dose injected in obese mice (1000 mmol iron/kg), thetotal iron measured by ICP is significantly enhanced in comparisonto the control in liver and spleen. However in tissue with macro-phages infiltration, such as adipose tissue of obese mice, theendogenous iron detected by ICP is still much larger than thecontent of SP iron detected by FMR. Thus, in the tissue of interest forMRI detection of inflammation, ICP quantification fails to detectexogenous nanoparticles, whereas FMR technique demonstratesboth specificity and sensitivity.
3.3. Quantitative evidence of the transformationof superparamagnetic nanoparticles
Comparing FMR and ICP results is particularly instructive toascertain the long term degradation and biotransformation ofnanoparticles. At each time point, the difference between ICP(representing total iron) and FMR (representing SP iron) directlyyields the amount of non-SP iron, which comprises the controlendogenous iron, but also the non-magnetic products of degrada-tion of the exogenous nanoparticles. This difference is shown inFig. 2B in comparison with control endogenous iron. Interestingly,non-SP iron increases in the spleenwith time course after P904 NPsinjection, while it is nearly constant in the liver. This tendency issignificant both for the low and high doses. At low dose, it isnoteworthy that the augmentation of non-SP iron in the spleenrepresents about 40% of the total injected iron (25 mg) and by farexceeds the amount of SP iron (i.e. P904 NPs) sequestered in thisorgan at day 1 (5.2% of total injected iron). At high dose, a similarincrease in non-SP iron, in the range of 40 mg, is detected in thespleen. This result suggests a redistribution of the products ofdegradation of nanoparticles from the liver to the spleen. A possiblescenario is that a majority of P904 NPs are transformed in the liver.It is consistent e first with the large uptake capacity of the livercompared to the spleen and - second with the gradual disappear-ance of SP iron from the liver. Subsequently, the non-SP products of

Fig. 1. Ferromagnetic resonance to quantify superparamagnetic nanoparticles. Ae FMR spectra of P904 nanoparticles in colloidal suspension (blue line) and of dried liver samples atdays 1, 7, 30, 60 and 90 after low dose injection of P904 (shaded red lines). P904 NPs with a mean diameter of 8 nm display a characteristic FMR spectrum with a resonance field of3050 Gauss and a linewidth of 600 Gauss. A similar slightly enlarged spectrum is observed in organs with an absorption signal decreasing over time after P904 administration. FMRabsorption signal is proportional to the quantity of superparamagnetic P904 NPs (denoted as SP iron). In contrast, the liver spectrum of control non-injected mouse (inset) showsa narrow line centered at 3330 Gauss revealing the presence of free radicals. B- Amount of superparamagnetic (SP) nanoparticles in organs as a function of time afterP904 administration at low (50 mmol/kg) and high dose (1000 mmol/kg). The mass of iron in the form of SP nanoparticles (SP iron) is expressed as a percentage of injected irondose � standard deviation (n ¼ 6 mice per time point). (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
M. Levy et al. / Biomaterials 32 (2011) 3988e3999 3991
degradation are not stocked in the liver but mostly transferred tothe spleen to increase the iron storage tank, probably in the non-SPform of ferritin or hemosiderin. Remarkably we found the sameamount of non-SP iron at low and high doses in the spleen,demonstrating that the storage capacity of the spleen is finite andcould limit the in vivo degradation rate of nanoparticles.
3.4. Evolution of the magnetic properties of nanoparticles
Some issues are now raised from the in vivo processes of NPsdegradation. How long persists the magnetic response of nano-particles? Once processed by the biological activity, do the NPsgradually decrease in size while keeping a magnetic structure or do

Fig. 2. A- Total iron content in organs measured by ICP-OES as a function of time after P904 administration at low and high dose. Total iron is expressed as a percentage of injectediron dose � standard deviation (n ¼ 6). B- Change in the quantity of non-superparamagnetic (non-SP) iron in organs following injection of P904 SP NPs. Non-SP iron is given by thedifference between total iron measured by ICP-AES and SP iron measured by FMR.
M. Levy et al. / Biomaterials 32 (2011) 3988e39993992
they loss their magnetic order? Going on with the nanomagnetismapproach, we thought to gain insight into such issues, by moni-toring the evolution of magnetic properties in tissues over time.
To this purpose, we performed static magnetization measure-ments bymeans of a superconducting quantum interference device(SQUID) magnetometer. The magnetizations of liver and spleenweremeasured at 300 K as a function of themagnetic field strength(up to 3 � 104 Gauss) and as a function of temperature from 5 K to
300 K for a fixed magnetic field of 50 Gauss. Both the field andthermal dependence of the magnetization give insight into thesuperparamagnetic behaviour of NPs in tissue. First the measure-ment of the saturation magnetization at high field (here 3 � 104
Gauss) provides a quantitative assessment of the concentration ofmagnetically-responsive SP iron in the sample (Fig. 3A). Consis-tently with FMR results of Fig. 1B, the magnetization per gram ofliver or spleen decreases over time after the administration of P904
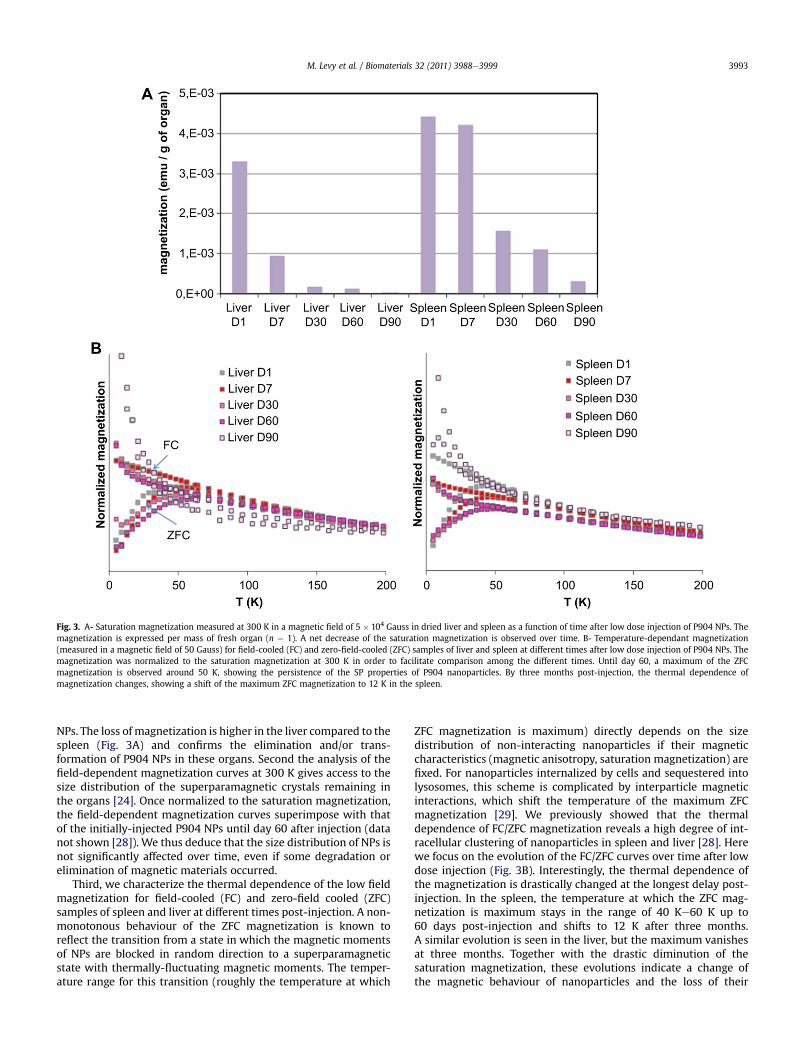
Fig. 3. A- Saturation magnetization measured at 300 K in a magnetic field of 5 � 104 Gauss in dried liver and spleen as a function of time after low dose injection of P904 NPs. Themagnetization is expressed per mass of fresh organ (n ¼ 1). A net decrease of the saturation magnetization is observed over time. B- Temperature-dependant magnetization(measured in a magnetic field of 50 Gauss) for field-cooled (FC) and zero-field-cooled (ZFC) samples of liver and spleen at different times after low dose injection of P904 NPs. Themagnetization was normalized to the saturation magnetization at 300 K in order to facilitate comparison among the different times. Until day 60, a maximum of the ZFCmagnetization is observed around 50 K, showing the persistence of the SP properties of P904 nanoparticles. By three months post-injection, the thermal dependence ofmagnetization changes, showing a shift of the maximum ZFC magnetization to 12 K in the spleen.
M. Levy et al. / Biomaterials 32 (2011) 3988e3999 3993
NPs. The loss of magnetization is higher in the liver compared to thespleen (Fig. 3A) and confirms the elimination and/or trans-formation of P904 NPs in these organs. Second the analysis of thefield-dependent magnetization curves at 300 K gives access to thesize distribution of the superparamagnetic crystals remaining inthe organs [24]. Once normalized to the saturation magnetization,the field-dependent magnetization curves superimpose with thatof the initially-injected P904 NPs until day 60 after injection (datanot shown [28]). We thus deduce that the size distribution of NPs isnot significantly affected over time, even if some degradation orelimination of magnetic materials occurred.
Third, we characterize the thermal dependence of the low fieldmagnetization for field-cooled (FC) and zero-field cooled (ZFC)samples of spleen and liver at different times post-injection. A non-monotonous behaviour of the ZFC magnetization is known toreflect the transition from a state in which the magnetic momentsof NPs are blocked in random direction to a superparamagneticstate with thermally-fluctuating magnetic moments. The temper-ature range for this transition (roughly the temperature at which
ZFC magnetization is maximum) directly depends on the sizedistribution of non-interacting nanoparticles if their magneticcharacteristics (magnetic anisotropy, saturation magnetization) arefixed. For nanoparticles internalized by cells and sequestered intolysosomes, this scheme is complicated by interparticle magneticinteractions, which shift the temperature of the maximum ZFCmagnetization [29]. We previously showed that the thermaldependence of FC/ZFC magnetization reveals a high degree of int-racellular clustering of nanoparticles in spleen and liver [28]. Herewe focus on the evolution of the FC/ZFC curves over time after lowdose injection (Fig. 3B). Interestingly, the thermal dependence ofthe magnetization is drastically changed at the longest delay post-injection. In the spleen, the temperature at which the ZFC mag-netization is maximum stays in the range of 40 Ke60 K up to60 days post-injection and shifts to 12 K after three months.A similar evolution is seen in the liver, but the maximum vanishesat three months. Together with the drastic diminution of thesaturation magnetization, these evolutions indicate a change ofthe magnetic behaviour of nanoparticles and the loss of their

M. Levy et al. / Biomaterials 32 (2011) 3988e39993994
superparamagnetic properties. It must be noted that ZFC curveswith a maximum around 10 K, similar to the one found here inspleen at three months, have been previously observed in patientswith genetic iron overload [27,30,31]. They were attributed to themagnetic behaviour of iron-rich ferritin. The ferritin is a proteincapable of storing iron in a non-toxic form, while maintaining ironavailability [32e34]. It is synthesized both in physiological condi-tions of iron balance and in pathological iron overload as a defencemechanism against the potentially-toxic labile iron surplus. Theprotein shell of apoferritin (12.5 nm external diameter) canaccommodate various amounts of iron (up to 4500 iron atoms)within its internal 7 nm cavity in the non-toxic form of a growingFe(III) crystal usually of ferrihydrite. As ferrihydrite is antiferro-magnetic at low temperature, the crystalline cores of ferritinexhibit a magnetic moment - due to uncompensated magneticsublattices - which is much smaller than that of maghemite parti-cles of the same size [31]. They show a maximum of ZFC magne-tization around 10 K [30]. As a consequence, as long as maghemitestructures are present in the tissue, they mask the magneticcontribution of iron-rich ferritin molecules. However, by threemonths post-injection, we observe the only magnetic response offerritin, pointing out the complete transformation of P904 nano-particles. It must be noticed that, in the time interval up to com-plete transformation, there is almost no evolution of the FC/ZFCmagnetic behaviour. A progressive diminution of the crystal size ofP904 nanoparticles while conserving a maghemite structure,would have led to a continuous shift of ZFCmaximum toward lowertemperatures. Instead of that, it suggests that some intact nano-particles with the initial size distribution and structure of P904 NPsare still present in the spleen and liver up to three months. Thisfinding is consistent with the persistence of the same shape offield-dependent magnetization curves.
3.5. Nanostructural follow-up of nanoparticles biotransformation
To gain insight into the subcellular and cellular changesfollowing iron oxide injection and the biotransformation of P904NPs leading to the loss of their magnetic properties, an ultrastruc-tural investigation was carried out using electron microscopy.Spleen and liver (high and low dose injection) and adipose tissuefromobesemice (high dose)were examined at different times post-administration. Large views of tissues reveal that nanoparticleswiththe typical size and shape of P904 NPs [24] are conspicuous in onlyfew cells, likely of macrophage type, with remarkably high intra-cellular concentration from one day post-injection (Fig. 4A). Incontrast, other cell types of the tissue (e.g. hepatocytes and endo-thelial cells in the liver, lymphocytes in the spleen and adipocytes inadipose tissue) are free of electron-dense nanoparticles of the samesize. This first observation confirms the exclusive and very efficientuptake of nanoparticles by specialized macrophages of the organ(liver Kupffer cells, spleen macrophages or adipose tissue macro-phages), which shows the capacity to sequester huge amounts ofexogenous nanoparticles. The intracellular sequestration which isachieved in vivo is remarkably high compared to cells labelled invitro with same nanoparticles [35].
At the subcellular scale, large clusters ofNPs are seen sequesteredinto few intracellular lysosomes with a well defined membrane(Fig. 4B). Cytosol and other intracellular compartments (mitochon-dria, nucleus, reticulum.) are free of nanoparticles. Elementalanalysis by EDX-Spectrum Imaging confirms the colocalization ofiron-rich punctuated nano-objects with electron-dense nano-particles within vesicles (Fig. 4C). Moreover electron diffractioncarried out on the iron-rich zones definitely points out structuralparameters consistent with maghemite nanocrystals (Fig. 4D).Hence P904 NPs are found within macrophages with their initial
structure and size at day 1 post-injection (day 3 for the high dosecondition). Their spatial distribution within lysosomes is quiteheterogenous, showingdense homogenousfillingormoredispersedNPs, with some area devoid of any NPs and with some osmiophiliclipid-associated droplets (Fig. 4A).
From day 7 post-injection (low dose), the pattern of NPs local-ization is unchanged at the tissue scale (NPs are still sequesteredinto scarce macrophages) (Fig. 5A), but the intralysosomal distri-bution displays qualitative changes. Within the same lysosome,intact P904 NPs are seen in the vicinity with electron-dense areasshowing “fingerprint like” circular arrays or whorls (Fig. 5B).Alternatively, some osmiophilic patches appear close to NPs clus-ters, some of them presenting nanometric holes. Other NPs-filledlysosomes (Fig. 5C) are observed close to a cytosolic agglomerate ofmonodisperse nanoparticles with facetted shape (5e7 nm indiameter), showing quasi-organized distribution and a bright areaaround individual nanoparticles (Fig. 5D). Remarkably, such typesof organizations and large clusters of monodisperse nanoparticleswere previously observed in patients with genetic iron overload[36] and attributed to the presence of iron storage proteins offerritin and hemosiderin. Iron-containing ferritin molecules arereadily detected because of the electron-density of the core, theirdiameter depending on the degree of iron deposition. Typically,ferritin particles have an octahedral appearance, surrounded bya bright area occupied by the apoferritin protein shell. Underphysiological conditions (iron equilibrium), ferritinmolecules is themajor iron storage form and aremainly seen randomly dispersed inthe cytosol. However, in diseases of iron overload like idiopathichemochromatosis, iron-rich ferritin molecules tend to be clus-terized and segregated into lysosomal structures, named side-rosomes, where they are potentially transformed into hemosiderin[32]. Ferritin proteins can thus appear as large clusters of iron-richregular 5e7 nm nanoparticles similar to the one observe in Fig. 5Dor alternatively they organize into paracrystalline hexagonal orcircular arrays within siderosomes, resembling the structuresobserved in Fig. 5B. Such features, characteristic of iron storageproteins [36,37], are observed here for the first time coexisting withexogenously-injected nanoparticles. It suggests an ongoing trans-formation process, by which surplus iron imported by exogenousmaghemite nanoparticles could be stored into endogenous storageproteins, as early proposed in [12].
It must be noted that the ferritin nanoparticles can be clearlydistinguished from the exogenous maghemite P904 NPs (at least atshort times post-injection (Fig. 5E)) thanks to their smaller size(5e7 nm) and lower size dispersion [38]. Both types of particlescontain iron (as confirmed by EDX-SI), but ferritin cores displaya distinct electron diffraction pattern, demonstrating a poorly-crystalline structure (Fig. 5E).
Further evolution of structural features reinforces our hypothesisof a local continuous transformation of NPswithin lysosomes. At day30 after low dose injection (day 44 for the high dose), concentratedassemblies of electron-dense nanoparticles are still observed withinlysosomes (Fig. 6A), but the individual particles are less resolved(Fig. 6B). They show poorly-defined contours and smaller sizes thanthe initially-injected NPs. Moreover, while EDX-SI proves that theycontain iron, electron diffraction demonstrates a loss of crystallinity(Fig. 6C). This pool of weakly-defined NPs could possibly arise fromthe degradation of the injected P904 NPs, which have lost theiroriginal structure and magnetic order. Concomitantly, organized“fingerprint-like” structures and assemblies of monodisperse iron-rich proteins are more frequently observed than at shorter time.Observations of tissues by threemonths post-injection show similarfeaturesbut accentuated (Fig. 6D). Individual degradednanoparticlesare hardly resolved and the frequency of fingerprint-like structuresis further enhanced (Fig. 6D). Interestingly, these structural clues of
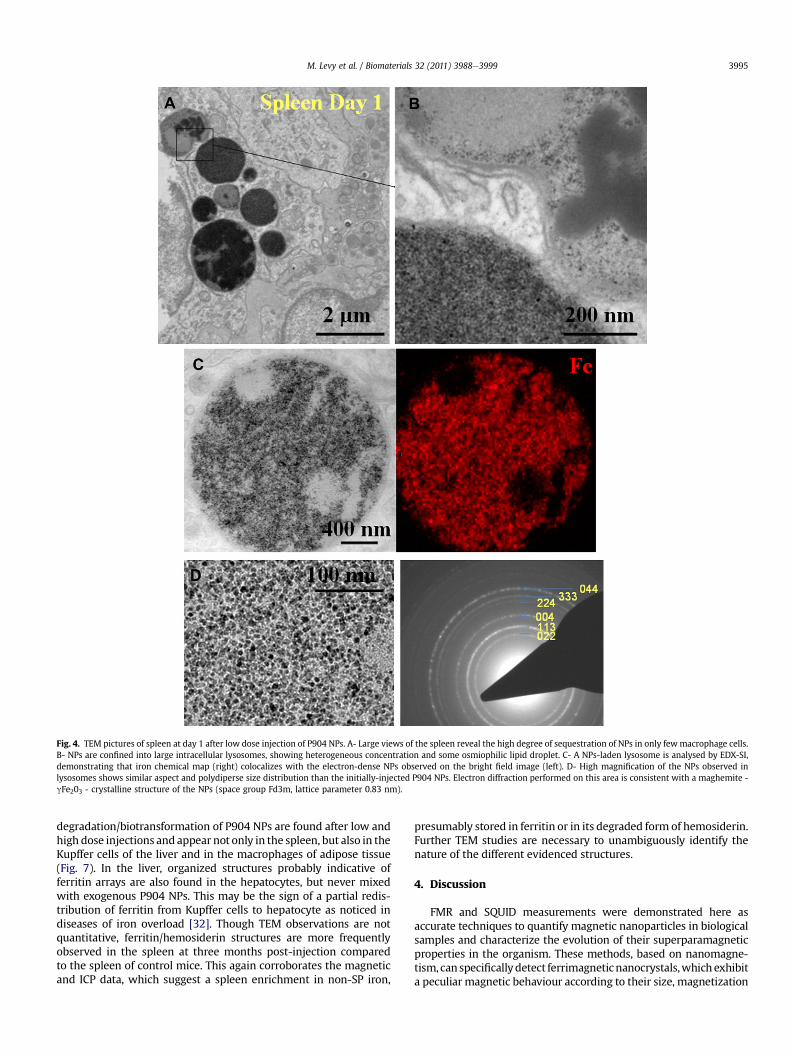
Fig. 4. TEM pictures of spleen at day 1 after low dose injection of P904 NPs. A- Large views of the spleen reveal the high degree of sequestration of NPs in only fewmacrophage cells.B- NPs are confined into large intracellular lysosomes, showing heterogeneous concentration and some osmiophilic lipid droplet. C- A NPs-laden lysosome is analysed by EDX-SI,demonstrating that iron chemical map (right) colocalizes with the electron-dense NPs observed on the bright field image (left). D- High magnification of the NPs observed inlysosomes shows similar aspect and polydiperse size distribution than the initially-injected P904 NPs. Electron diffraction performed on this area is consistent with a maghemite -gFe203 - crystalline structure of the NPs (space group Fd3m, lattice parameter 0.83 nm).
M. Levy et al. / Biomaterials 32 (2011) 3988e3999 3995
degradation/biotransformation of P904 NPs are found after low andhigh dose injections and appear not only in the spleen, but also in theKupffer cells of the liver and in the macrophages of adipose tissue(Fig. 7). In the liver, organized structures probably indicative offerritin arrays are also found in the hepatocytes, but never mixedwith exogenous P904 NPs. This may be the sign of a partial redis-tribution of ferritin from Kupffer cells to hepatocyte as noticed indiseases of iron overload [32]. Though TEM observations are notquantitative, ferritin/hemosiderin structures are more frequentlyobserved in the spleen at three months post-injection comparedto the spleen of control mice. This again corroborates the magneticand ICP data, which suggest a spleen enrichment in non-SP iron,
presumably stored in ferritin or in its degraded form of hemosiderin.Further TEM studies are necessary to unambiguously identify thenature of the different evidenced structures.
4. Discussion
FMR and SQUID measurements were demonstrated here asaccurate techniques to quantify magnetic nanoparticles in biologicalsamples and characterize the evolution of their superparamagneticproperties in the organism. These methods, based on nanomagne-tism, can specificallydetect ferrimagneticnanocrystals,whichexhibita peculiar magnetic behaviour according to their size, magnetization

Fig. 5. TEM pictures of spleen at day 7 after low dose injection of P904 NPs. A- Large views show that the electron dense areas are still confined within lysosomal structures ofmacrophages. B- Inside a lysosome, some assemblies of intact P904 NPs (red arrows) are observed in the vicinity of fingerprint-like structures characteristic of an array of ferritinproteins (yellow arrows). C- Another lysosome shows a high concentration of NPs together with osmiophilic lipid droplets in the periphery. Close to this lysosome (D), a cytosolicassembly of monodisperse 5e7 nm nanoparticules (dotted yellow arrow) is indicative of the presence of iron-rich ferritin. E� High magnification of this assembly shows that thismonodisperse NPs have a very different aspect compared to P904 NPs. Electron diffraction demonstrates their poorly-crystalline structure. The proximity of intact maghemite NPswith some assemblies of iron storage proteins suggests a relationship between these nanostructures. (For interpretation of the references to colour in this figure legend, the reader isreferred to the web version of this article.)
M. Levy et al. / Biomaterials 32 (2011) 3988e39993996
andmagnetic anisotropy. The high sensitivity of FMRmeasurementsallows a quantification of superparamagnetic iron in organswith lowNPs uptake (such as targeted inflammation sites like adipose tissue)andat long termafter injection,whenmostNPshavebeeneliminatedor degraded. The nanoparticles content in liver and spleen can beprecisely monitored and the combined detections of SP iron only(FMR) and total iron (ICP) give an insight in the transformation ofnanoparticles into other forms of iron compounds and in theirrespective distribution within liver and spleen. Note that ICP alone
failed to detect NPs injected at low dose, due to the high level ofendogeneous non-magnetic iron. The kinetics of degradation/elimi-nation of P904 nanoparticles is shown here to be dose- and organ-dependent. Low dose of P904 are more rapidly assimilated by theorganism compared to high dose. Of note, a transfer of non-magneticiron from the liver to the spleen isnoticedwhatever thedose injected.However the iron concentration in the spleen tends to a value, whichis identical for both doses. This could be explained by a saturation ofiron storage capacity of the spleen.

Fig. 6. A-B-C TEM micrographs of spleen at day 30 after low dose injection of P904 NPs. A- Large view shows the persistence of lysosomes with electron dense areas. B- Insidea lysosome, the conspicuous electron-dense NPs have smaller sizes and less-defined contours compared to NPs observed at shorter times. C- High magnification and electrondiffraction on these NPs show that they have lost their crystalline structure. D e Spleen at day 90 after low dose injection of P904 NPs shows numerous lysosomes with electron-dense patches (siderosomes). No more NPs can be resolved, but many fingerprint-like arrays are seen, suggesting the complete degradation of P904 NPs and the potential transfer ofreleased iron into storage ferritin/hemosiderin proteins.
M. Levy et al. / Biomaterials 32 (2011) 3988e3999 3997
The magnetic analysis, in complement to ICP measurements,provides a sensitive assessment of NPs degradation in the organism.Actually, thenon-degradedNPs,whichkeep their superparamagneticproperties, can be separated from their products of degradation,which have lost ferrimagnetic order. Thereby iron stored in ferritin/hemosiderin and ionic forms of iron (heme, transferring-bound ironetc.) are distinguished from the still magnetic nanoparticles. Takingadvantage of FMR and SQUID specificity, we demonstrate the long
termdecrease of P904NPs concentration in spleen and liver. A part ofthese NPs are transformed into poorly- or non-magnetic iron (likelystored in ferritin and hemosiderin proteins), while the rest of NPsconserve their original magnetic properties.
Interestingly, a similar degradation profile was observed for NPsexposed to an acidic medium mimicking intracellular environment[24]. At pH ¼ 4.7 in presence of a citrate chelating agent, P904NPs were progressively degraded and transformed into ionic iron

Fig. 7. TEM pictures of spleen, liver and adipose tissue at day 10 and day 44 after high dose injection of P904 NPs. P904 NPs are conspicuous in the lysosomes of the macrophages ofeach organ (Kupffer cells in the liver and inflammatory macrophages in adipose tissue). The clues of NPs biotransformation can be seen whatever the organ: Intact P904 NPs(red arrows) coexist with monodisperse 5e7 nm iron-rich ferritin proteins (dotted yellow arrows) or fingerprint-like structures (yellow arrows), showing the ongoing trans-formation process within membrane-bound compartments. The intact P904 NPs seem to form a reservoir, with some outlying connections with ferritin proteins. (For interpretationof the references to colour in this figure legend, the reader is referred to the web version of this article.)
M. Levy et al. / Biomaterials 32 (2011) 3988e39993998
species. While the global magnetization and FMR signal decreasedwith time due to the dissolution of some NPs and the release of freeiron, the size distribution andmagnetic properties of the remainingparticles stayed unchanged in the course of degradation.
The combination of nanoscale observations and magnetic anal-ysis prompts us to propose a possible scenario for the biotransfor-mation of NPs. P904 nanoparticles are efficiently sequestered intolysosomes of macrophages, where some of them remain intact,conserving their size and magnetic order up to two months. In thesame lysosomes, the intact NPs coexist with putative iron-richproteins and with degraded NPs having lost their crystalline struc-tures. Over time, the storage iron forms become more prominent.We thus hypothesize that the transformation of NPs takes place inlysosomes [39] and that NPs are processed contingent on theavailability of iron storage proteins. Thus NP degradation should belimited by the availability and affinity of iron chelating agents
produced intracellularly and the capacity of ferritin or hemosiderinprotein to store the released iron. The remaining superparamagneticnanocrystals form a reservoir of non-toxic Fe(III) iron organized intheir ferrimagnetic lattice, with poor availability as long as they arenot degraded by intracellular iron chelates. On the other side, theferritin protein will bind the labile iron species resulting from thedissolution of the nanoparticles and store them into another non-toxic form. Such a mechanism may offer a chance for the cell toregulate the labile iron pool generated by the degradation productsof NPs and thus tominimize toxicity [4,5], consistentlywith the verygood tolerance profile of iron oxide NPs. How iron transfer takesplace from exogenous nanoparticles to endogeneous iron storageprotein is not elucidated. A biological analysis, investigating theresponse of iron metabolism [40] to the accumulation of ironoxide NPs and a deeper ultrastructural study is underway to unravelthesemechanisms of nanoparticles biotransformation. It would also

M. Levy et al. / Biomaterials 32 (2011) 3988e3999 3999
important to assess if NPs can visit several cells before beingdegraded, via the shedding of vesicles which cargo nanoparticlesfrom cell to cell, as previously observed in vitro [41].
5. Conclusion
Amultiscale approach was used here to delineate the way takenby cells in vivo to process and degrade iron oxide nanoparticles overtime. The three-month magnetic follow-up of nanoparticles, usingferromagnetic resonance and SQUID measurements, gave evidenceof the degradation of magnetic nanocrystals through a loss of theirpeculiar superparamagnetic properties. At the organism level, weobserved the relocation of iron species from the liver to the spleen.The nanoscale investigation using electronmicroscopy supports theview of a continuous biotransformation of the injected nano-particles into iron storage species within intracellular lysosomes.Further investigations are needed to decipher the intracellularpathwayof iron transfer fromnanoparticles to endogenousproteins.
Acknowledgement
We acknowledge Guerbet for providing us with the nano-particles. We are grateful to Anne d’Encausse, Eric Lancelot,Sébastien Ballet, Emmanuelle Canet-Soulas, Martin Devaud andChristian Ricolleau for fruitful discussions and to Achraf Al Faraj,Sylvie Manin and Thomas Decaen for technical assistance. Thiswork has been supported by the ANR (Agence Nationale de laRecherche) TecSan “Inflam” and by the European project Magnifyco(Contract NMP4-SL-2009-228622).
References
[1] Rivera Gil P, Oberdorster G, Elder A, Puntes V, Parak WJ. Correlating physico-chemical with toxicological properties of nanoparticles: the present and thefuture. ACS Nano 2010;4(10):5527e31.
[2] Pankhurst QA, Thanh NKT, Jones SK, Dobson J. Progress in applications ofmagnetic nanoparticles in biomedicine. J Phys D Appl Phys 2009;22:224001.
[3] Halas NJ. The photonic nanomedicine revolution: let the human side ofnanotechnology emerge. Nanomedicine (Lond) 2009;4(4):369e71.
[4] Pawelczyk E, Arbab AS, Pandit S, Hu E, Frank JA. Expression of transferrinreceptor and ferritin following ferumoxides-protamine sulfate labeling ofcells: implications for cellular magnetic resonance imaging. NMR Biomed2006;19(5):581e92.
[5] Kedziorek DA, Muja N, Walczak P, Ruiz-Cabello J, Gilad AA, Jie CC, et al. Geneexpression profiling reveals early cellular responses to intracellular magneticlabeling with superparamagnetic iron oxide nanoparticles. Magn Reson Med2010;63(4):1031e43.
[6] Lehmann AD, Parak WJ, Zhang F, Ali Z, Rocker C, Nienhaus GU, et al. Hybridnanoparticles: fluorescent-magnetic hybrid nanoparticles induce a dose-dependent increase in proinflammatory response in kung cells in vitrocorrelated with intracellular localization. Small 2010;6(6).
[7] Le Masne de Chermont Q, Chaneac C, Seguin J, Pelle F, Maitrejean S, Jolivet JP,et al. Nanoprobes with near-infrared persistent luminescence for in vivoimaging. Proc Natl Acad Sci U S A 2007;104(22):9266e71.
[8] Bulte JW. In vivo MRI cell tracking: clinical studies. AJR Am J Roentgenol2009;193(2):314e25.
[9] Liu W, Frank JA. Detection and quantification of magnetically labeled cells bycellular MRI. Eur J Radiol 2009;70(2):258e64.
[10] Billotey C, Wilhelm C, Devaud M, Bacri JC, Bittoun J, Gazeau F. Cell internali-zation of anionic maghemite nanoparticles: quantitative effect on magneticresonance imaging. Magn Reson Med 2003;49(4):646e54.
[11] Weissleder R, Stark DD, Engelstad BL, Bacon BR, Compton CC, White DL, et al.Superparamagnetic iron oxide: pharmacokinetics and toxicity. AJR Am JRoentgenol 1989;152(1):167e73.
[12] Okon E, Pouliquen D, Okon P, Kovaleva ZV, Stepanova TP, Lavit SG, et al.Biodegradation of magnetite dextran nanoparticles in the rat. A histologic andbiophysical study. Lab Invest 1994;71(6):895e903.
[13] Corot C, Robert P, Idee JM, Port M. Recent advances in iron oxide nanocrystaltechnology for medical imaging. Adv Drug Deliv Rev 2006;58(14):1471e504.
[14] Briley-Saebo K, Bjornerud A, Grant D, Ahlstrom H, Berg T, Kindberg GM.Hepatic cellular distribution and degradation of iron oxide nanoparticlesfollowing single intravenous injection in rats: implications for magneticresonance imaging. Cell Tissue Res 2004;316(3):315e23.
[15] Sigovan M, Bessaad A, Alsaid H, Lancelot E, Corot C, Neyran B, et al. Assess-ment of age modulated vascular inflammation in ApoE-/- mice by USPIO-enhanced magnetic resonance imaging. Invest Radiol 2010;45(11):702e7.
[16] Sigovan M, Boussel L, Sulaiman A, Sappey-Marinier D, Alsaid H, Desbleds-Mansard C, et al. Rapid-clearance iron nanoparticles for inflammation imagingof atherosclerotic plaque: initial experience in animal model. Radiology2009;252(2):401e9.
[17] Meier R, Henning TD, Boddington S, Tavri S, Arora S, Piontek G, et al. Breastcancers: MR imaging of folate-receptor expression with the folate-specificnanoparticle P1133. Radiology 2010;255(2):527e35.
[18] Fortin-Ripoche JP, Martina MS, Gazeau F, Menager C, Wilhelm C, Bacri JC, et al.Magnetic targeting of magnetoliposomes to solid tumors with MR imagingmonitoring in mice: feasibility. Radiology 2006;239(2):415e24.
[19] Cheng K, Li TS, Malliaras K, Davis DR, Zhang Y, Marban E. Magnetic targetingenhances engraftment and functional benefit of iron-labeled cardiosphere-derived cells in myocardial infarction. Circ Res 2010;106(10):1570e81.
[20] Gazeau F, Wilhelm C. Magnetic labeling, imaging and manipulation of endo-thelial progenitor cells using iron oxide nanoparticles. Future Med Chem2010;2(3):397e408.
[21] Fortin JP, Wilhelm C, Servais J, Menager C, Bacri JC, Gazeau F. Size-sortedanionic iron oxide nanomagnets as colloidal mediators for magnetic hyper-thermia. J Am Chem Soc 2007;129(9):2628e35.
[22] Brule S, Levy M, Wilhelm C, Letourneur D, Gazeau F, Menager C, et al.Doxorubicin release triggered by alginate embedded magnetic nanoheaters:A Combined Therapy. Adv Mater 2011.
[23] Massart R. Preparation of aqueous magnetic liquids in alkaline and acidicmedia. IEEE Trans Magn 1981;17:1247e8.
[24] Levy M, Lagarde F, Maraloiu VA, Blanchin MG, Gendron F, Wilhelm C, et al.Degradability of superparamagnetic nanoparticles in a model of intracellularenvironment: follow-up of magnetic, structural and chemical properties.Nanotechnology 2010;21(39):395103.
[25] Gazeau F, Shilov V, Bacri J-C, Dubois E, Gendron F, Perzynski R, et al. Magneticresonance of nanoparticles in a ferrofluid: evidence of thermofluctuationaleffects. J Magn Magn Mat 1999;202:535e46.
[26] Wilhelm C, Gazeau F, Bacri JC. Magnetophoresis and ferromagnetic resonanceof magnetically labeled cells. Eur Biophys J 2002;31(2):118e25.
[27] Slawska-Waniewska A, Mosiniewics-Szablewska E, Nedelko N, Galazka-Friedman J, Friedman A. Magnetic studies of iron-entities in human tissues.J Magn Magn Mat 2004;272-276:2417e9.
[28] Levy M, Devaud M, Luciani N, Devaux V, Elgrabli D, Pechoux C, et al. Nano-magnetism reveals the intracellular fate of nanoparticles in the organism.submitted.
[29] Lopez A, Gutierrez L, Lazaro FJ. The role of dipolar interaction in the quanti-tative determination of particulate magnetic carriers in biological tissues.Phys Med Biol 2007;52(16):5043e56.
[30] Allen PD, St Pierre TG, Chua-anusorn W, Strom V, Rao KV. Low-frequency low-field magnetic susceptibility of ferritin and hemosiderin. Biochim BiophysActa 2000;1500(2):186e96.
[31] Brem F, Stamm G, Hirt AM. Modeling the magnetic behavior of horse spleenferritin with a two-phase core structure. J Appl Phys 2006;99(12):123906.
[32] Harrison PM, Arosio P. The ferritins:molecular properties, iron storage functionand cellular regulation. Biochim Biophys Acta 1996;1275(3):161e203.
[33] MacKenzie EL, Iwasaki K, Tsuji Y. Intracellular iron transport and storage:from molecular mechanisms to health implications. Antioxid Redox Signal2008;10(6):997e1030.
[34] Arosio P, Ingrassia R, Cavadini P. Ferritins: a family of molecules for ironstorage, antioxidation and more. Biochim Biophys Acta 2009;1790(7):589e99.
[35] Wilhelm C, Gazeau F. Universal cell labelling with anionic magnetic nano-particles. Biomaterials 2008;29(22):3161e74.
[36] Iancu TC. Ferritin and hemosiderin in pathological tissues. Electron MicroscRev 1992;5(2):209e29.
[37] Lazaro FJ, Abadia AR, Romero MS, Gutierrez L, Lazaro J, Morales MP. Magneticcharacterisation of rat muscle tissues after subcutaneous iron dextran injec-tion. Biochim Biophys Acta 2005;1740(3):434e45.
[38] Quintana C, Cowley JM, Marhic C. Electron nanodiffraction and high-resolu-tion electron microscopy studies of the structure and composition of physi-ological and pathological ferritin. J Struct Biol 2004;147(2):166e78.
[39] Kidane TZ, Sauble E, Linder MC. Release of iron from ferritin requires lyso-somal activity. Am J Physiol Cell Physiol 2006;291(3):C445e55.
[40] Beaumont C, Delaby C. Recycling iron in normal and pathological states.Semin Hematol 2009;46(4):328e38.
[41] Luciani N, Wilhelm C, Gazeau F. The role of cell-released microvesicles in theintercellular transfer of magnetic nanoparticles in the monocyte/macrophagesystem. Biomaterials 2010;31(27):7061e9.
Copyright © 2022 FDOKUMEN



















