
Oxidative stress induced by cerium oxide nanoparticles in cultured BEAS-2B cells
11
Available online at www.sciencedirect.com Toxicology 245 (2008) 90–100 Oxidative stress induced by cerium oxide nanoparticles in cultured BEAS-2B cells Eun-Jung Park a , Jinhee Choi b , Young-Kwon Park b , Kwangsik Park a,∗ a College of Pharmacy, Dongduk Women’s University, 23-1, Wolgok-dong, Seongbuk-gu, Seoul 136-714, Republic of Korea b Faculty of Environmental Engineering, College of Urban Science, University of Seoul, 90 Jeonnong-dong, Dongdaemun-gu, Seoul 130-743, Republic of Korea Received 18 August 2007; received in revised form 17 December 2007; accepted 17 December 2007 Available online 3 January 2008 Abstract Cerium oxide nanoparticles of different sizes (15, 25, 30, 45 nm) were prepared by the supercritical synthesis method, and cytotoxicity was evaluated using cultured human lung epithelial cells (BEAS-2B). Exposure of the cultured cells to nanoparticles (5, 10, 20, 40 g/ml) led to cell death, ROS increase, GSH decrease, and the inductions of oxidative stress-related genes such as heme oxygenase-1, catalase, glutathione S-transferase, and thioredoxin reductase. The increased ROS by cerium oxide nanoparticles triggered the activation of cytosolic caspase-3 and chromatin condensation, which means that cerium oxide nanoparticles exert cytotoxicity by an apoptotic process. Uptake of the nanoparticles to the cultured cells was also tested. It was observed that cerium oxide nanoparticles penetrated into the cytoplasm and located in the peri-region of the nucleus as aggregated particles, which may induce the direct interaction between nanoparticles and cellular molecules to cause adverse cellular responses. © 2007 Elsevier Ireland Ltd. All rights reserved. Keywords: Ceria oxide nanoparticles; Cytotoxicity; Oxidative stress; BEAS-2B cells 1. Introduction Due to their unique physical and chemical characteristic, nanoparticles have been the focus of much research such as in industrial applications, environmental toxicity studies and human health impacts (Oberdorster et al., 2005; Priestly et al., 2007; Warheit et al., 2007). Various industrial nanomaterials made from titanium oxide, silver, gold, cadmium selenide and other carbon nanoparticles have been widely used for indus- trial purposes, and, cerium oxide nanoparticles may be the most important materials with wide ranges of applications for solar cells, fuel cells, gas sensors, oxygen pumps, and glass/ceramic applications (Gao et al., 2006). With the increased applications of cerium oxide nanoparticles, the concerns about their poten- tial human toxicity effects and their environmental impact were also increased. However, toxicity data for cerium oxide nanopar- ticles relating to human health are limited and only a few results ∗ Corresponding author. Tel.: +82 29404522; fax: +82 29404159. E-mail address: [email protected] (K. Park). from basic studies have been published. Furthermore, the toxic- ity of cerium oxide nanoparticles is still controversial, especially regarding the oxidant/antioxidant effect (Thill et al., 2006; Lin et al., 2006; Schubert et al., 2006). When bacteria were treated with cerium oxide nanoparti- cles (7 nm CeO 2 ), Gram-negative bacteria (Escherichia coli) adsorbed the positively charged particles in the pH 7 growth media. The adsorption of cerium oxide nanoparticles to bacteria may induce oxidative stress and cause cell death (Thill et al., 2006). In A549 cells treated with 20 nm CeO 2 nanoparticles, reactive oxygen species (ROS) were induced, and cell viabil- ity was decreased. The elevated oxidative stress was found to increase the production of MDA and LDH, indicators of lipid peroxidation and membrane damage, respectively (Lin et al., 2006). However, there are a few papers that show antioxidative and cell protective effects of ceria nanoparticles using nervous cell systems. Ceria nanoparticles decreased the endogenous ROS induced by glutamate and showed cell protective effect in cultured HT22 cells which are derived from the rodent nervous (Schubert et al., 2006). In the publication, authors wrote that cerium oxide act 0300-483X/$ – see front matter © 2007 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.tox.2007.12.022
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Oxidative stress induced by cerium oxide nanoparticles in cultured BEAS-2B cells

A
ecScttr©
K
1
nih2moticaotat
0d
Available online at www.sciencedirect.com
Toxicology 245 (2008) 90–100
Oxidative stress induced by cerium oxide nanoparticlesin cultured BEAS-2B cells
Eun-Jung Park a, Jinhee Choi b, Young-Kwon Park b, Kwangsik Park a,∗a College of Pharmacy, Dongduk Women’s University, 23-1, Wolgok-dong, Seongbuk-gu, Seoul 136-714, Republic of Korea
b Faculty of Environmental Engineering, College of Urban Science, University of Seoul, 90 Jeonnong-dong,Dongdaemun-gu, Seoul 130-743, Republic of Korea
Received 18 August 2007; received in revised form 17 December 2007; accepted 17 December 2007Available online 3 January 2008
bstract
Cerium oxide nanoparticles of different sizes (15, 25, 30, 45 nm) were prepared by the supercritical synthesis method, and cytotoxicity wasvaluated using cultured human lung epithelial cells (BEAS-2B). Exposure of the cultured cells to nanoparticles (5, 10, 20, 40 �g/ml) led toell death, ROS increase, GSH decrease, and the inductions of oxidative stress-related genes such as heme oxygenase-1, catalase, glutathione-transferase, and thioredoxin reductase. The increased ROS by cerium oxide nanoparticles triggered the activation of cytosolic caspase-3 andhromatin condensation, which means that cerium oxide nanoparticles exert cytotoxicity by an apoptotic process. Uptake of the nanoparticles to
he cultured cells was also tested. It was observed that cerium oxide nanoparticles penetrated into the cytoplasm and located in the peri-region ofhe nucleus as aggregated particles, which may induce the direct interaction between nanoparticles and cellular molecules to cause adverse cellularesponses.2007 Elsevier Ireland Ltd. All rights reserved.
cells
fire
camm2riip
eywords: Ceria oxide nanoparticles; Cytotoxicity; Oxidative stress; BEAS-2B
. Introduction
Due to their unique physical and chemical characteristic,anoparticles have been the focus of much research such asn industrial applications, environmental toxicity studies anduman health impacts (Oberdorster et al., 2005; Priestly et al.,007; Warheit et al., 2007). Various industrial nanomaterialsade from titanium oxide, silver, gold, cadmium selenide and
ther carbon nanoparticles have been widely used for indus-rial purposes, and, cerium oxide nanoparticles may be the mostmportant materials with wide ranges of applications for solarells, fuel cells, gas sensors, oxygen pumps, and glass/ceramicpplications (Gao et al., 2006). With the increased applicationsf cerium oxide nanoparticles, the concerns about their poten-
ial human toxicity effects and their environmental impact werelso increased. However, toxicity data for cerium oxide nanopar-icles relating to human health are limited and only a few results∗ Corresponding author. Tel.: +82 29404522; fax: +82 29404159.E-mail address: [email protected] (K. Park).
2ac
bc2
300-483X/$ – see front matter © 2007 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.tox.2007.12.022
rom basic studies have been published. Furthermore, the toxic-ty of cerium oxide nanoparticles is still controversial, especiallyegarding the oxidant/antioxidant effect (Thill et al., 2006; Lint al., 2006; Schubert et al., 2006).
When bacteria were treated with cerium oxide nanoparti-les (7 nm CeO2), Gram-negative bacteria (Escherichia coli)dsorbed the positively charged particles in the pH 7 growthedia. The adsorption of cerium oxide nanoparticles to bacteriaay induce oxidative stress and cause cell death (Thill et al.,
006). In A549 cells treated with 20 nm CeO2 nanoparticles,eactive oxygen species (ROS) were induced, and cell viabil-ty was decreased. The elevated oxidative stress was found toncrease the production of MDA and LDH, indicators of lipideroxidation and membrane damage, respectively (Lin et al.,006). However, there are a few papers that show antioxidativend cell protective effects of ceria nanoparticles using nervousell systems.
Ceria nanoparticles decreased the endogenous ROS inducedy glutamate and showed cell protective effect in cultured HT22ells which are derived from the rodent nervous (Schubert et al.,006). In the publication, authors wrote that cerium oxide act

colog
acntHwvgtago
oftHcsBt
2
2
ttuestsr
bctfiKsui5waso
2
wVtfNt2(ff(Tc
2
E.-J. Park et al. / Toxi
s a free radical scavenger. Das et al. (2007) also showed thateria nanoparticles offer neuroprotection to adult rat spinal cordeurons. Spinal cord neurons and other CNS neurons are proneo be damaged due to oxidative stress, both in vitro and in vivo.owever, when cerium nanoparticles were added to the culturesith hydrogen peroxide, a significantly higher number of sur-iving cells were shown as compared to the non-treated controlroup. These results contrast with results that show the induc-ion of oxidative stress and cell death by nanoparticles. It waslso published that CeO2 nanoparticles protect against the pro-ression of cardiac dysfunction and remodeling by attenuationf myocardial oxidative stress (Niu et al., 2007).
In the present work, we evaluated the cytotoxic effect of ceriaxide nanoparticles using BEAS-2B cells which are derivedrom human bronchial epithelia normal cells and comparedhe cytotoxicity using different cell lines, such as T98G and
9C2. Furthermore, ROS generation, intracellular GSH levels,aspase-3 activity, chromosomal condensation, and gene expres-ions as markers of oxidative stress were also investigated inEAS-2B cells, to reveal possible mechanisms of cell death by
he cerium oxide nanoparticles.
. Materials and methods
.1. Preparation of cerium oxide nanoparticles
The cerium oxide was prepared by the supercritical synthesis method, andhe schematic diagram for the synthesis apparatus is shown in Fig. 1. The syn-hesis process is outlined as follows. Distilled water, pumped into the systemsing a high-pressure pump, was heated to a moderate temperature by a heat
xchanger and heated further to a desired temperature by a pre-heater. Theolution of the metal precursor Ce(NO3)3 and ammonia water for pH con-rol, was pumped into the system using high pressure pumps, and mixed withupercritical water at the mixing point located above the reactor. A sufficienteaction time for the formation of cerium oxide nanoparticles was provided(12co
Fig. 1. Schematic diagram for the continuous hydrothermal synthesis of ceri
y 245 (2008) 90–100 91
y the reactor in order to maintain a supercritical condition. The synthesizederium oxide was collected by filter, and this was followed by spray dryingo obtain the final product. The crystal structure of cerium oxide was con-rmed by powder X-ray diffraction (XRD) patterns using monochromic Cu� radiation (Rigaku, D/MAX III). The average crystallite size was also mea-
ured by the X-ray line broadening technique employing the Scherrer formulasing the profiles of the (1 1 1) peak. SEM (Scanning Electron Microscope)mages of cerium oxide nanoparticles were obtained by a Phillips SEM-35M microscope operating at an acceleration voltage of 20 kV. The samplesere prepared by sprinkling the powder oxides onto double-sided sticky tape
nd mounting them on a microscope stub. The samples were coated withputtered gold. Fig. 2 shows the morphology and crystallite size of ceriumxide.
.2. Materials
BEAS-2B cells, which are derived from human bronchus epithelial cells,ere purchased from the American type culture collection (ATCC) (Manassas,A, USA). H9C2 cells, which are rat cardiomyocytes, were purchased from
he Korean cell line bank (Seoul, Korea) and T98G cells which are derivedrom human brain fibroblast were kindly donated by S.J. Kim working for theational Institute of Health, USA. Growth properties of all cell lines used in
his study are as adherent cells. DMEM/F12 media, 3-(4,5-dimethylthiazol--yl)-2,5-diphenyltetrazolium bromide (MTT), dichlorofluorescein-diacetateDCFH-DA), reduced glutathione and orthophthaldialdehyde were purchasedrom Sigma (St. Louis, MO, USA). The caspase-3 assay kit was purchasedrom R&D systems Inc. (Minneapolis, MN, USA). The RNA isolation kitRNAgents®) was purchased from the Promega Company (Madison, WI, USA).he AccuPower RT/PCR Premix for the amplification of nucleic acid was pur-hased from Bioneer (Daejeon, Korea).
.3. Cell culture and nanoparticles treatment
BEAS-2B cells were maintained in Dulbecco’s modified Eagle’s medium
DMEM)/F12 containing 10% heat-inactivated fetal bovine serum, penicillin00 IU/ml, and streptomycin 10 �g/ml. Cells were grown and maintained in8 cm2 cell culture flasks at 37 ◦C in a 5% CO2 humidified incubator. T98Gells and H9C2 cells were grown and maintained as above but DMEM insteadf DMEM/F12 was used.um oxide nanoparticles in supercritical water (supercritical synthesis).

92 E.-J. Park et al. / Toxicology 245 (2008) 90–100
F rticles ow difn
mUcss
2
d2ct4(ttnito
2
dUgtaDwoun
riawe1
ad1t1pLrm4oga
2
(cficamrc
ig. 2. Morphological characterization of different sized cerium oxide nanopacanning electron microscope. Four different sizes of ceriaum nanoparticles shanoparticles.
The test solution of cerium oxide nanoparticles was prepared in the cultureedia and dispersed for 20 min by using a sonicator (Branson Inc., Danbury, CT,SA) to prevent aggregation. During the test periods, cerium oxide nanoparti-
les were not aggregated at the highest concentration of 40 �g/ml used in thistudy. The cells were treated with various concentrations of particles, and a timechedule is designated in the following section of each toxicological study.
.4. Cell viability test
Cell viability was measured by the MTT (3-(4-5-dimethylthiazol-2-yl)-2,5-iphenyltetrazolium bromide, Sigma, St. Louis, MO) assay (Park and Park,006). Cells were seeded on 96-well tissue culture plates with 5 × 103–2 × 104
ells in 100 �l media per well. After a 24 h stabilization of the cells, they werereated with 10, 20, and 40 �g/ml concentrations of 30 nm-sized particles for 24,8, 72, and 96 h, respectively. At the end of exposure, 40 �l of MTT solution2 mg/ml) was added and the cells were incubated for 4 h at 37 ◦C. Cells werereated with 150 �l of DMSO and absorbance was quantified in 540 nm usinghe microplate spectrophotometer system (VersaMax, Molecular Devices, Sun-yvale, CA, USA). The cells were treated with 15, 25, 30, and 45 nm-particlesn order to compare the cytotoxicities of different sized nanoparticles on thereated cells. The viability of the treated group was expressed as the percentagef control group that was assumed to be 100%.
.5. Measurement of ROS and GSH
To measure ROS generation, a fluorometric assay using intracellular oxi-ation of 2,7-dichlorofluoroscein diacetate (DCFH-DA, Sigma, St. Louis, MO,SA) was performed (Elbekai and El-Kadi, 2005; Fotakis et al., 2005). Cellsrown to confluence at 24 h after seeding were pretreated with different concen-rations (5, 10, 20, 40 �g/ml) of 30 nm-nanoparticles for 24 h, washed with PBS,nd then incubated with 40 �M DCFH-DA for 30 min. At the end of DCFH-
A incubation, cells were washed with PBS, lysed with NaOH, and aliquotsere transferred to the black well plate. Then the fluorescence of dichloroflu-roscein (DCF), which is the oxidized product of DCFH-DA, was measuredsing the microplate spectrofluorometer (GeminiXPS, Molecular Devices, Sun-yvale, CA, USA) with excitation and emission wavelengths of 485 and 530 nm,(stn
s. Structures of cerium oxide nanoparticles used in this study were shown byferent morphologies: (A) 15 nm; (B) 25 nm; (C) 35 nm; and (D) 40 nm size of
espectively. For the visual image of ROS generation, the cells were culturedn the presence or absence of cerium oxide nanoparticles, washed with PBS,nd then loaded with 40 �M DCFH-DA for 30 min. The fluorescence in cellsas visualized using a fluorescent microscope (Nicon, Tokyo, Japan) with an
xcitation of 485 nm and an emission of 530 nm, and an exposure time of/100–1/130 s.
To investigate the relationship between the increased ROS and the level ofntioxidant materials in cells, the intracellular GSH level (reduced form) wasetermined. The cells treated with 30 nm-nanoparticles (5, 10, 20, 40 �g/ml) in2-well plates for 24 h were washed with PBS, and 1% perchloric acid was addedo the cell pellet and left for 10 min on ice. The cell lysates were centrifuged at3,000 rpm at 4 ◦C for 5 min prior to analysis in order to remove precipitatedrotein. Cell lysates, KH2PO4/EDTA buffer, and o-phthaldialdehyde (Sigma, St.ouis, MO, USA) were put in 96-black well plates and incubated in the dark at
oom temperature for 30 min. Fluorescence was measured using a fluorescenceulti-well plate reader with excitation and emission wavelengths of 350 and
20 nm, respectively (Hissin and Hilf, 1976). Results were calculated as nmolf glutathione per mg of protein and presented as a percentage of the controlroup. Protein assays in the cell lysate were performed using a BCA proteinssay reagent (Pierce, Rockfold, IL, USA).
.6. Caspase-3 activity and chromosome condensation
The activity of caspase-3 was determined using a colorimetric assay kitR&D systems Inc, MN, USA). Briefly, cells were incubated with different con-entrations of 30 nm-nanoparticles (5, 10, 20, 40 �g/ml) for 24 h. The cells wererst lysed by the solution provided in the assay kit to collect their intracellularontents. The cell lysates could then be tested for their enzyme activity by theddition of a caspase-specific peptide that is conjugated to the color reporterolecules p nitroanaline (pNA). The cleavage of the peptide by the caspase,
eleased the chromophore pNA, which can be quantitated spectrophotometri-ally at a 450 nm.
To evaluate chromosome condensation, 4′,6-diamidino-2-phenylindoleDAPI, Sigma, St. Louis, MO, USA) staining was also performed. The DAPIolution was applied to cultured cells in chamber slides (10, 20, 40 �g/ml), andhe slides were incubated for 10 min in the dark at 37 ◦C, and the images of theucleus were made by a fluorescent microscope (Nicon, Tokyo, Japan) with an

E.-J. Park et al. / Toxicology 245 (2008) 90–100 93
Table 1Primer sequences of oxidative stress-related genes used in this study
Gene name GB No. Primer sequences
Glutathione S-transferase NM146421.1 F: 5′-CCATCTTTGAGAACACAGGT-3′R: 5′-GAGAAGATTCGTGTGGACAT-3′
Heme oxygenase-1 NM002133.1 F: 5′-CTCTGAAGTTTAGGCCATTG-3′R: 5′-AGTTGCTGTAGGGCTTTATG-3′
Thioredoxin reductase NM182742.1 F: 5′-AAACCAATACCAGCAAGAAA-3′R: 5′-CTATGAGAATGCTTATGGGC-3′
Catalase NM001752.2 F: 5′-TCATGACATTTAATCAGGCA-3′
A
e1
2
fRWt2twPfspa
2
n3b
2
ietTw
3
3
B(dAwHbn
cuwda
cTnw
3
theott4gsle
3
twrobug1a
3
ctin NM001101.2
xcitation of 330–380 nm and an emission of 420 nm, and an exposure time of/100–1/130 s (Dhar-Mascareno et al., 2005).
.7. Gene expression analysis
Cells were incubated with 40 �g/ml concentration of 30 nm-nanoparticlesor the designated times (0.5, 1, 2, 4, and 8 h). For the preparation of totalNA, the RNAgent total RNA isolation system (Promega Corporation, Madison,I, USA) was used according to the manufacturer’s instructions. Reverse-
ranscription (RT)-PCR was performed using oligo deoxythymidine primer in0 �l volumes at 42 ◦C for 60 min. The RT-PCR reaction was done with 1 �g ofotal RNA, 1 �l of 20 �M oligo dT primer, and 18 �l of reaction mixture whichas provided by AccuPower RT/PCR PreMix (Bioneer, Daejeon, Korea). ThenCR was performed in a 20 �l total mixture volume for 25–28 cycles at 95 ◦Cor 1 min, 55 ◦C for 1 min, and 72 ◦C for 1 min. Amplified cDNA products wereeparated on 1.5% agarose gel by electrophoresis (Park and Park, 2007). Therimer sequences of amplified genes are shown in Table 1. Actin mRNA waslso amplified and shown to be a loading control.
.8. Nanoparticles uptake test
Cells were treated with a 40 �g/ml concentration of 30 nm-cerium oxideanoparticles and observed using a phase-contrast microscope (200×) after 1.5,, and 12 h, respectively. Photos of particle treated cells were further magnifiedy a computer imaging system.
.9. Statistical analysis
The results of cell viability, ROS generation, GSH level, and caspase-3 activ-ty are presented as the mean ± standard deviation (S.D) of three or four separatexperiments. The results of the chemically-treated groups were compared tohose of the control group and represented as the percentage of the control value.he values were compared using the Student’s t-test, and levels of significanceere represented for each result.
. Results
.1. Cytotoxicity
To examine the toxic effects of cerium oxide nanoparticles,EAS-2B cells were incubated with different concentrations
10, 20, 40 ppm) of 30 nm-nanoparticles and viability wasetermined 24, 48, 72, and 96 h after treatment, respectively.s shown in Fig. 3A, cell viability was decreased by treatment
ith nanoparticles in a time- and dose-dependent manner.owever, the dose-dependency was not so impressive. Via-ility of the treated group with a 20 �g/ml concentration ofanoparticles was down to 40–50% of the control group, andc
w
R: 5′-GTGTCAGGATAGGCAAAAAG-3′F: 5′- GGCGGACTATGACTTAGTTG-3′R: 5′-AAACAACAATGTGCAATCAA-3′
ytotoxicity was not intensified by the increase in concentrationp to 40 �g/ml. Different sizes of cerium oxide nanoparticlesere also compared in this toxicity test but no statisticallyifferences were found among groups treated with 15, 25, 30,nd 40 nm-particles (Fig. 3B).
The cytotoxicity of cerium oxide nanoparticles to differentell lines was compared, and the results are shown in Fig. 4.he cytotoxicity of cerium oxide nanoparticles (5 �g/ml) wasot significant in both T98G cells and H9C2 cells. Cell viabilityas not decreased during the 98 h exposure time.
.2. ROS generation and GSH reduction
The fluorescence intensity was increased in BEAS-2B cellsreated with nanoparticles at the concentrations of 5 �g/ml andigher, as shown in Fig. 5. This means that there is a gen-ration of ROS by cerium oxide nanoparticles. Visualizationf ROS production under the fluorescent microscope showedhat oxidized DCF fluorescence was increased in the cells cul-ured in the presence of cerium oxide nanoparticles (10, 20,0 �g/ml), while fluorescence was insignificant in the controlroup (Fig. 6). In Fig. 7, decreased levels of GSH were alsohown in the nanoparticles-treated group. The level of intracel-ular GSH seemed to be about 75% of the control group at 24 hxposure of 40 �g/ml of 30 nm-sized particles.
.3. Molecular biomarkers
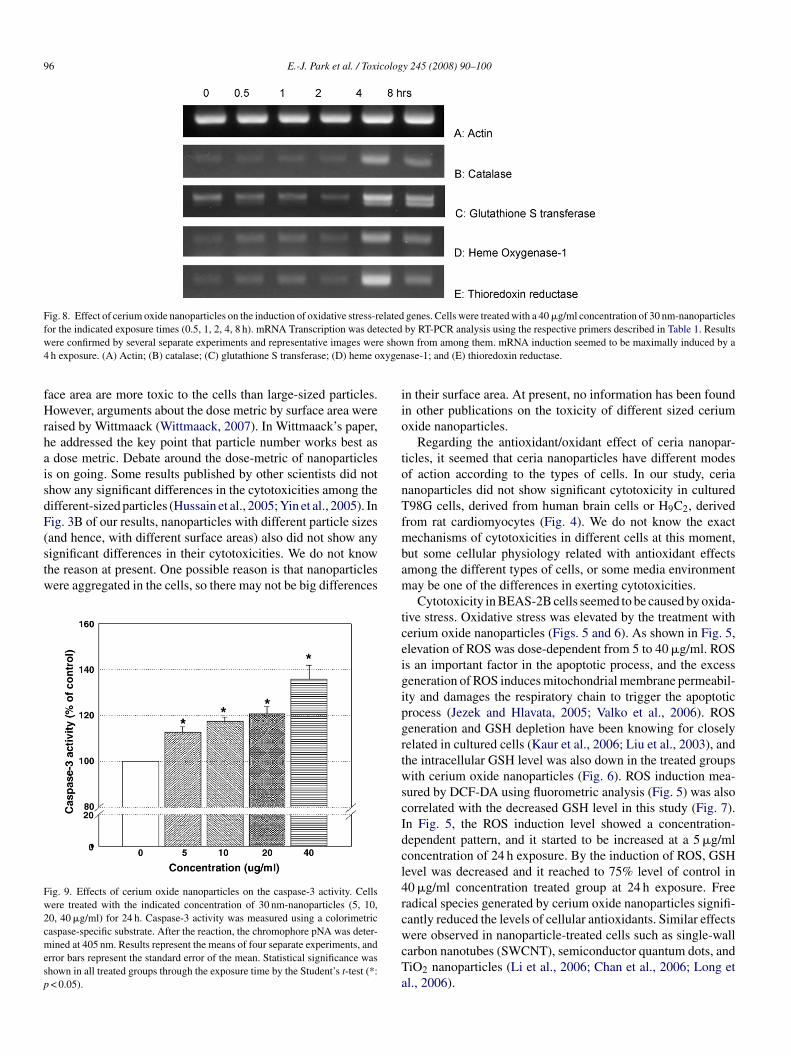
To investigate the expression level of each mRNA relatedo the oxidative stress responses in BEAS-2B cells, RT-PCRas performed. As shown in Fig. 8, various oxidative stress-
elated genes including catalase, glutathione S-transferase, hemexygenase-1, and thioredoxin reductase shown to be inducedy 40 �g/ml concentrations of 30 nm-nanoparticles in the cellssing agarose gel electrophoresis. Maximal inductions of theenes, catalase, glutathione S-transferase, and heme oxygenase-occurred at the exposure time of 4 h. The house keeping gene,
ctin, was not changed by nanoparticles.
.4. Induction of caspase-3 activity and chromosome
ondensationCaspase-3, which plays a key role in the apoptotic path-ay of cells, was increased following treatment with cerium

94 E.-J. Park et al. / Toxicology 245 (2008) 90–100
Fig. 3. Effects of cerium oxide nanoparticles on the viability of BEAS-2B celllines. Cell viability was assessed by MTT assays and results are presented asthe percentage of control group viability. (A) Cells (5 × 103–2 × 104 cells) weretreated with the indicated concentrations of cerium oxide nanoparticles (30 nmsize) for 24, 48, 72, and 96 h for dose-dependent tests. Cell viability was greatlyreduced in a concentration-dependent and time-dependent manner by nanoparti-cle exposure. All the points showed statistically significant differences from thecontrol group by the Student’s t-test (p < 0.05). (B) Cells (5 × 103–2 × 104 cells)were treated with 40 �g/ml of different sized-cerium oxide nanoparticles (15, 25,30 and 45 nm) for 24, 48, 72, and 96 h for the size-difference test. Cell viabilitywas greatly reduced in all the groups treated with different sized nanoparticles.However, no significant differences in cytotoxicity among the different-sizedn(b
o5a(masccc
Fig. 4. Effects of cerium oxide nanoparticles on the cytotoxicity in differentcultured cells, BEAS-2B, T98G and H9C2. Cell viability was assessed by MTTassays and results are presented as a percentage of control group viability. Cells(5 × 103–2 × 104 cells) were treated with 5 ppm of cerium oxide nanoparticles(is
cct
3
observed at the time of 1.5, 3, and 12 h, respectively. Particle-images appeared in the cytoplasm about 1.5 h after treatmentand they were located and accumulated around the nuclearmembrane (3 and 12 h).
Fig. 5. Effects of cerium oxide nanoparticles on ROS induction in BEAS-2Bcells. Cells grown in confluence were treated with 30 nm-nanoparticles of desig-nated concentrations for 24 h, washed with phosphate-buffered saline, and thenincubated with 40 �M DCFH-DA. At the end of DCFH-DA incubation, the cells
anoparticles were shown using the statistical analysis of the Student’s t-testp > 0.05). Results represent the means of three separate experiments, and errorars represent the standard error of the mean.
xide nanoparticles (Fig. 9). When the cells were treated with, 10, 20, and 40 �g/ml concentrations of particles for 24 h, thectivity of caspase-3 was increased in the lowest concentration5 �g/ml) group. The activity was gradually increased by theaximal 40 �g/ml concentration. In addition to the caspase-3
ctivity, chromatin condensation was also evaluated by DAPI
taining. When cells were treated with 10, 20, and 40 �g/mloncentrations of cerium oxide nanoparticles for 24 h, chromatinondensation was observed using a fluorescent microscope. Theondensations were increased relative to the increase of exposurewrsdr
30 nm size) for 24, 48, 72, and 96 h using three different cell lines. Cell viabil-ty was greatly reduced in a time-dependent manner in BEAS-2B cells but noignificant cytotoxicity was shown in eather T98G cells or H9C2 cells.
oncentration (Fig. 10). The caspase-3 activation and chromatinondensation in BEAS-2B suggested that cerium oxide nanopar-icles caused cell death by apoptotic process.
.5. Uptake of cerium oxide nanoparticle
After treatment of cells with nanoparticles, cells were
ere lysed with NaOH and the fluorescence of aliquot was measured. Resultsepresent the means of three separate experiments, and error bars represent thetandard error of the mean. All treated groups showed statistically significantifferences from the control group by the Student’s t-test (**: p < 0.01). Data isepresented as the percentage of the ROS level in the control group.
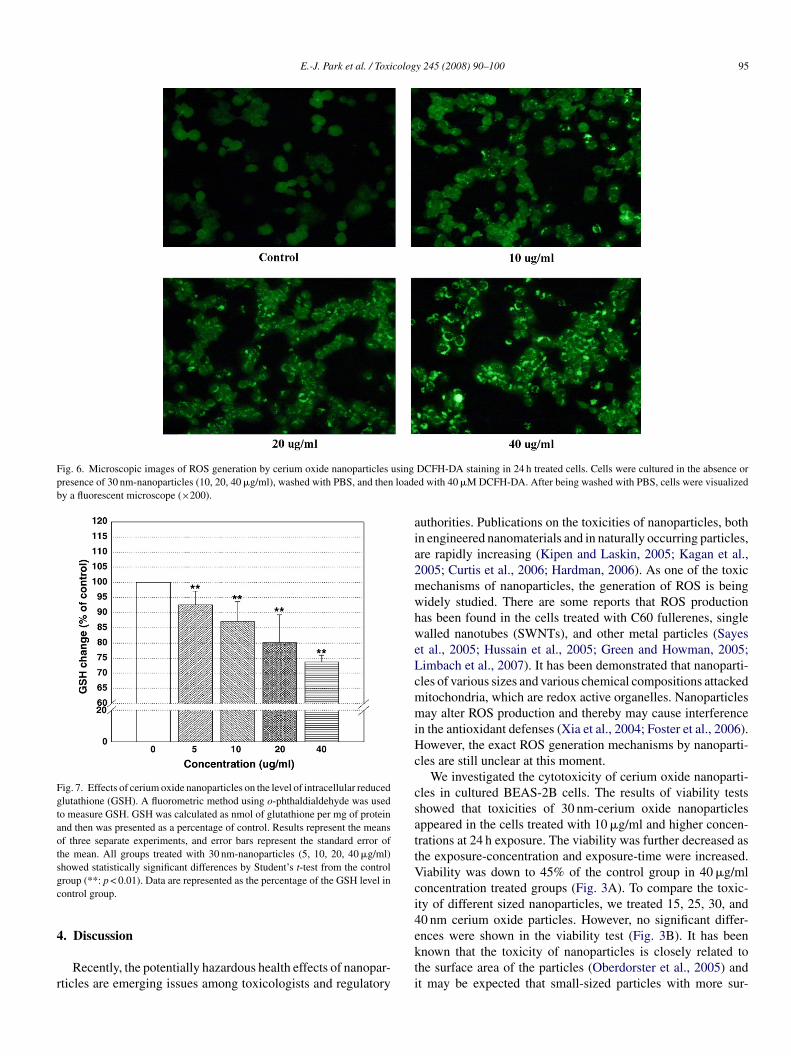
E.-J. Park et al. / Toxicology 245 (2008) 90–100 95
Fig. 6. Microscopic images of ROS generation by cerium oxide nanoparticles usingpresence of 30 nm-nanoparticles (10, 20, 40 �g/ml), washed with PBS, and then loadeby a fluorescent microscope (×200).
Fig. 7. Effects of cerium oxide nanoparticles on the level of intracellular reducedglutathione (GSH). A fluorometric method using o-phthaldialdehyde was usedto measure GSH. GSH was calculated as nmol of glutathione per mg of proteinand then was presented as a percentage of control. Results represent the meansof three separate experiments, and error bars represent the standard error ofthe mean. All groups treated with 30 nm-nanoparticles (5, 10, 20, 40 �g/ml)showed statistically significant differences by Student’s t-test from the controlgc
4
r
aia2mwhweLcmmiHc
csattVci4ences were shown in the viability test (Fig. 3B). It has been
roup (**: p < 0.01). Data are represented as the percentage of the GSH level inontrol group.
. Discussion
Recently, the potentially hazardous health effects of nanopar-ticles are emerging issues among toxicologists and regulatory
kti
DCFH-DA staining in 24 h treated cells. Cells were cultured in the absence ord with 40 �M DCFH-DA. After being washed with PBS, cells were visualized
uthorities. Publications on the toxicities of nanoparticles, bothn engineered nanomaterials and in naturally occurring particles,re rapidly increasing (Kipen and Laskin, 2005; Kagan et al.,005; Curtis et al., 2006; Hardman, 2006). As one of the toxicechanisms of nanoparticles, the generation of ROS is beingidely studied. There are some reports that ROS productionas been found in the cells treated with C60 fullerenes, singlealled nanotubes (SWNTs), and other metal particles (Sayes
t al., 2005; Hussain et al., 2005; Green and Howman, 2005;imbach et al., 2007). It has been demonstrated that nanoparti-les of various sizes and various chemical compositions attackeditochondria, which are redox active organelles. Nanoparticlesay alter ROS production and thereby may cause interference
n the antioxidant defenses (Xia et al., 2004; Foster et al., 2006).owever, the exact ROS generation mechanisms by nanoparti-
les are still unclear at this moment.We investigated the cytotoxicity of cerium oxide nanoparti-
les in cultured BEAS-2B cells. The results of viability testshowed that toxicities of 30 nm-cerium oxide nanoparticlesppeared in the cells treated with 10 �g/ml and higher concen-rations at 24 h exposure. The viability was further decreased ashe exposure-concentration and exposure-time were increased.iability was down to 45% of the control group in 40 �g/mloncentration treated groups (Fig. 3A). To compare the toxic-ty of different sized nanoparticles, we treated 15, 25, 30, and0 nm cerium oxide particles. However, no significant differ-
nown that the toxicity of nanoparticles is closely related tohe surface area of the particles (Oberdorster et al., 2005) andt may be expected that small-sized particles with more sur-
96 E.-J. Park et al. / Toxicology 245 (2008) 90–100
Fig. 8. Effect of cerium oxide nanoparticles on the induction of oxidative stress-related genes. Cells were treated with a 40 �g/ml concentration of 30 nm-nanoparticlesf ectedw e show4 xygen
fHrhaisdF(stw
Fw2cmesp
iio
tonTf
or the indicated exposure times (0.5, 1, 2, 4, 8 h). mRNA Transcription was detere confirmed by several separate experiments and representative images werh exposure. (A) Actin; (B) catalase; (C) glutathione S transferase; (D) heme o
ace area are more toxic to the cells than large-sized particles.owever, arguments about the dose metric by surface area were
aised by Wittmaack (Wittmaack, 2007). In Wittmaack’s paper,e addressed the key point that particle number works best asdose metric. Debate around the dose-metric of nanoparticles
s on going. Some results published by other scientists did nothow any significant differences in the cytotoxicities among theifferent-sized particles (Hussain et al., 2005; Yin et al., 2005). Inig. 3B of our results, nanoparticles with different particle sizes
and hence, with different surface areas) also did not show anyignificant differences in their cytotoxicities. We do not knowhe reason at present. One possible reason is that nanoparticlesere aggregated in the cells, so there may not be big differencesig. 9. Effects of cerium oxide nanoparticles on the caspase-3 activity. Cellsere treated with the indicated concentration of 30 nm-nanoparticles (5, 10,0, 40 �g/ml) for 24 h. Caspase-3 activity was measured using a colorimetricaspase-specific substrate. After the reaction, the chromophore pNA was deter-ined at 405 nm. Results represent the means of four separate experiments, and
rror bars represent the standard error of the mean. Statistical significance washown in all treated groups through the exposure time by the Student’s t-test (*:< 0.05).
mbam
tceigipgrtwscIdcl4rcwcTa
by RT-PCR analysis using the respective primers described in Table 1. Resultsn from among them. mRNA induction seemed to be maximally induced by a
ase-1; and (E) thioredoxin reductase.
n their surface area. At present, no information has been foundn other publications on the toxicity of different sized ceriumxide nanoparticles.
Regarding the antioxidant/oxidant effect of ceria nanopar-icles, it seemed that ceria nanoparticles have different modesf action according to the types of cells. In our study, ceriaanoparticles did not show significant cytotoxicity in cultured98G cells, derived from human brain cells or H9C2, derived
rom rat cardiomyocytes (Fig. 4). We do not know the exactechanisms of cytotoxicities in different cells at this moment,
ut some cellular physiology related with antioxidant effectsmong the different types of cells, or some media environmentay be one of the differences in exerting cytotoxicities.Cytotoxicity in BEAS-2B cells seemed to be caused by oxida-
ive stress. Oxidative stress was elevated by the treatment witherium oxide nanoparticles (Figs. 5 and 6). As shown in Fig. 5,levation of ROS was dose-dependent from 5 to 40 �g/ml. ROSs an important factor in the apoptotic process, and the excesseneration of ROS induces mitochondrial membrane permeabil-ty and damages the respiratory chain to trigger the apoptoticrocess (Jezek and Hlavata, 2005; Valko et al., 2006). ROSeneration and GSH depletion have been knowing for closelyelated in cultured cells (Kaur et al., 2006; Liu et al., 2003), andhe intracellular GSH level was also down in the treated groupsith cerium oxide nanoparticles (Fig. 6). ROS induction mea-
ured by DCF-DA using fluorometric analysis (Fig. 5) was alsoorrelated with the decreased GSH level in this study (Fig. 7).n Fig. 5, the ROS induction level showed a concentration-ependent pattern, and it started to be increased at a 5 �g/mloncentration of 24 h exposure. By the induction of ROS, GSHevel was decreased and it reached to 75% level of control in0 �g/ml concentration treated group at 24 h exposure. Freeadical species generated by cerium oxide nanoparticles signifi-antly reduced the levels of cellular antioxidants. Similar effects
ere observed in nanoparticle-treated cells such as single-wallarbon nanotubes (SWCNT), semiconductor quantum dots, andiO2 nanoparticles (Li et al., 2006; Chan et al., 2006; Long etl., 2006).
E.-J. Park et al. / Toxicology 245 (2008) 90–100 97
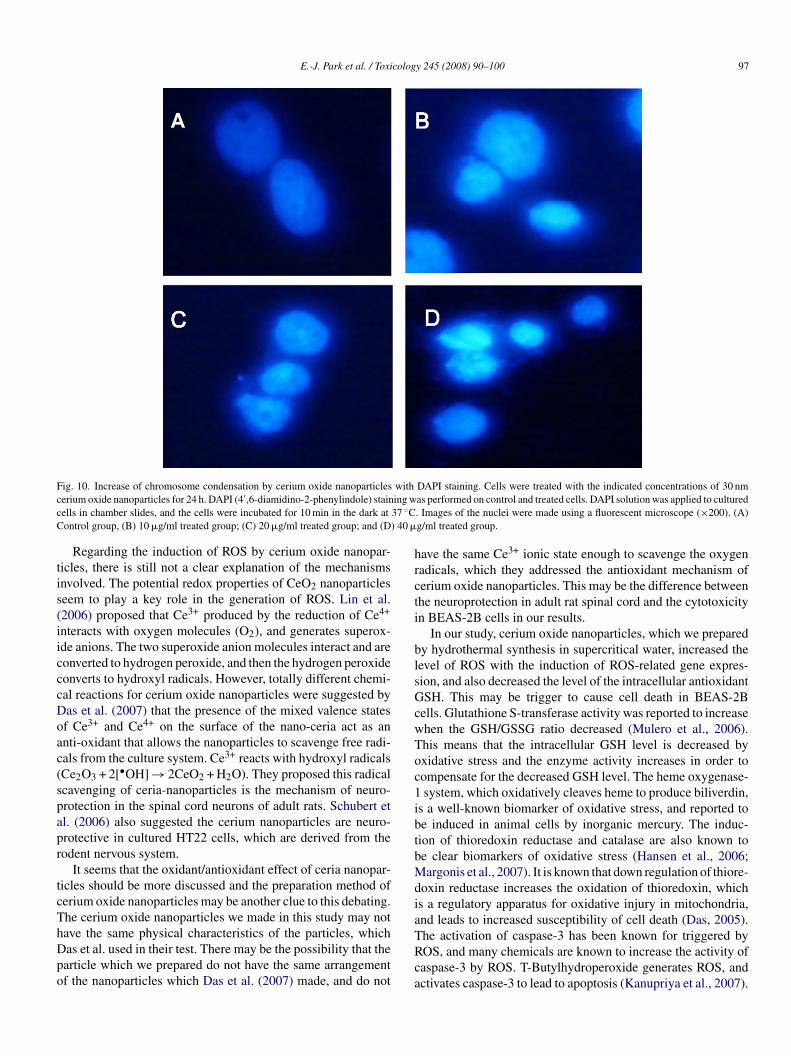
F withc ing wc 37 ◦CC ) 40 �
tis(iicccDoac(spapr
tcThDpo
hrcti
blsGcwToc1ibtbMdia
ig. 10. Increase of chromosome condensation by cerium oxide nanoparticleserium oxide nanoparticles for 24 h. DAPI (4′,6-diamidino-2-phenylindole) stainells in chamber slides, and the cells were incubated for 10 min in the dark atontrol group, (B) 10 �g/ml treated group; (C) 20 �g/ml treated group; and (D
Regarding the induction of ROS by cerium oxide nanopar-icles, there is still not a clear explanation of the mechanismsnvolved. The potential redox properties of CeO2 nanoparticleseem to play a key role in the generation of ROS. Lin et al.2006) proposed that Ce3+ produced by the reduction of Ce4+
nteracts with oxygen molecules (O2), and generates superox-de anions. The two superoxide anion molecules interact and areonverted to hydrogen peroxide, and then the hydrogen peroxideonverts to hydroxyl radicals. However, totally different chemi-al reactions for cerium oxide nanoparticles were suggested byas et al. (2007) that the presence of the mixed valence statesf Ce3+ and Ce4+ on the surface of the nano-ceria act as annti-oxidant that allows the nanoparticles to scavenge free radi-als from the culture system. Ce3+ reacts with hydroxyl radicalsCe2O3 + 2[•OH] → 2CeO2 + H2O). They proposed this radicalcavenging of ceria-nanoparticles is the mechanism of neuro-rotection in the spinal cord neurons of adult rats. Schubert etl. (2006) also suggested the cerium nanoparticles are neuro-rotective in cultured HT22 cells, which are derived from theodent nervous system.
It seems that the oxidant/antioxidant effect of ceria nanopar-icles should be more discussed and the preparation method oferium oxide nanoparticles may be another clue to this debating.he cerium oxide nanoparticles we made in this study may not
ave the same physical characteristics of the particles, whichas et al. used in their test. There may be the possibility that thearticle which we prepared do not have the same arrangementf the nanoparticles which Das et al. (2007) made, and do notTRca
DAPI staining. Cells were treated with the indicated concentrations of 30 nmas performed on control and treated cells. DAPI solution was applied to cultured. Images of the nuclei were made using a fluorescent microscope (×200). (A)g/ml treated group.
ave the same Ce3+ ionic state enough to scavenge the oxygenadicals, which they addressed the antioxidant mechanism oferium oxide nanoparticles. This may be the difference betweenhe neuroprotection in adult rat spinal cord and the cytotoxicityn BEAS-2B cells in our results.
In our study, cerium oxide nanoparticles, which we preparedy hydrothermal synthesis in supercritical water, increased theevel of ROS with the induction of ROS-related gene expres-ion, and also decreased the level of the intracellular antioxidantSH. This may be trigger to cause cell death in BEAS-2B
ells. Glutathione S-transferase activity was reported to increasehen the GSH/GSSG ratio decreased (Mulero et al., 2006).his means that the intracellular GSH level is decreased byxidative stress and the enzyme activity increases in order toompensate for the decreased GSH level. The heme oxygenase-system, which oxidatively cleaves heme to produce biliverdin,
s a well-known biomarker of oxidative stress, and reported toe induced in animal cells by inorganic mercury. The induc-ion of thioredoxin reductase and catalase are also known toe clear biomarkers of oxidative stress (Hansen et al., 2006;argonis et al., 2007). It is known that down regulation of thiore-
oxin reductase increases the oxidation of thioredoxin, whichs a regulatory apparatus for oxidative injury in mitochondria,nd leads to increased susceptibility of cell death (Das, 2005).
he activation of caspase-3 has been known for triggered byOS, and many chemicals are known to increase the activity ofaspase-3 by ROS. T-Butylhydroperoxide generates ROS, andctivates caspase-3 to lead to apoptosis (Kanupriya et al., 2007).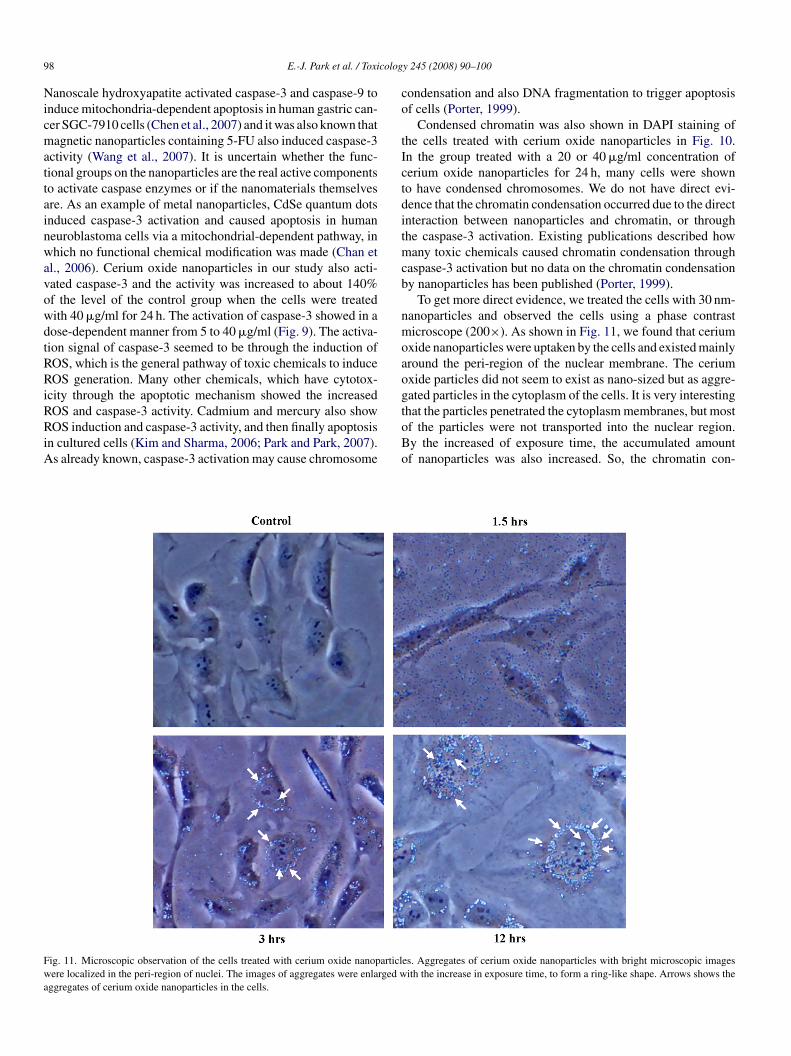
9 colog
NicmattainwavowdtRRiRRiA
co
tIctditmcb
nmoaogt
Fwa
8 E.-J. Park et al. / Toxi
anoscale hydroxyapatite activated caspase-3 and caspase-9 tonduce mitochondria-dependent apoptosis in human gastric can-er SGC-7910 cells (Chen et al., 2007) and it was also known thatagnetic nanoparticles containing 5-FU also induced caspase-3
ctivity (Wang et al., 2007). It is uncertain whether the func-ional groups on the nanoparticles are the real active componentso activate caspase enzymes or if the nanomaterials themselvesre. As an example of metal nanoparticles, CdSe quantum dotsnduced caspase-3 activation and caused apoptosis in humaneuroblastoma cells via a mitochondrial-dependent pathway, inhich no functional chemical modification was made (Chan et
l., 2006). Cerium oxide nanoparticles in our study also acti-ated caspase-3 and the activity was increased to about 140%f the level of the control group when the cells were treatedith 40 �g/ml for 24 h. The activation of caspase-3 showed in aose-dependent manner from 5 to 40 �g/ml (Fig. 9). The activa-ion signal of caspase-3 seemed to be through the induction ofOS, which is the general pathway of toxic chemicals to induceOS generation. Many other chemicals, which have cytotox-
city through the apoptotic mechanism showed the increasedOS and caspase-3 activity. Cadmium and mercury also show
OS induction and caspase-3 activity, and then finally apoptosisn cultured cells (Kim and Sharma, 2006; Park and Park, 2007).s already known, caspase-3 activation may cause chromosome
oBo
ig. 11. Microscopic observation of the cells treated with cerium oxide nanoparticlere localized in the peri-region of nuclei. The images of aggregates were enlarged w
ggregates of cerium oxide nanoparticles in the cells.
y 245 (2008) 90–100
ondensation and also DNA fragmentation to trigger apoptosisf cells (Porter, 1999).
Condensed chromatin was also shown in DAPI staining ofhe cells treated with cerium oxide nanoparticles in Fig. 10.n the group treated with a 20 or 40 �g/ml concentration oferium oxide nanoparticles for 24 h, many cells were showno have condensed chromosomes. We do not have direct evi-ence that the chromatin condensation occurred due to the directnteraction between nanoparticles and chromatin, or throughhe caspase-3 activation. Existing publications described how
any toxic chemicals caused chromatin condensation throughaspase-3 activation but no data on the chromatin condensationy nanoparticles has been published (Porter, 1999).
To get more direct evidence, we treated the cells with 30 nm-anoparticles and observed the cells using a phase contrasticroscope (200×). As shown in Fig. 11, we found that cerium
xide nanoparticles were uptaken by the cells and existed mainlyround the peri-region of the nuclear membrane. The ceriumxide particles did not seem to exist as nano-sized but as aggre-ated particles in the cytoplasm of the cells. It is very interestinghat the particles penetrated the cytoplasm membranes, but most
f the particles were not transported into the nuclear region.y the increased of exposure time, the accumulated amountf nanoparticles was also increased. So, the chromatin con-es. Aggregates of cerium oxide nanoparticles with bright microscopic imagesith the increase in exposure time, to form a ring-like shape. Arrows shows the

colog
dab
otctpamm
A
G2b
R
C
C
C
D
D
D
E
F
F
G
G
H
H
H
H
J
K
K
K
K
K
L
L
L
L
L
M
M
N
O
P
P
P
P
S
E.-J. Park et al. / Toxi
ensation seemed to be caused by the activation of caspase-3nd further cascade processes, and not by the direct interactionetween the nanoparticles and chromatin.
In conclusion, the present study revealed the cytotoxicityf cerium oxide nanoparticles with the induction of ROS andhe decreased level of intracellular GSH in cultured BEAS-2Bells. Cytosolic caspase-3 activation and chromatin condensa-ion were also shown in the cells, which suggested the apoptoticrocess. The nanoparticles penetrated into the plasma membranend located in the peri-region of nuclear membranes, whicheans nanoparticles may have direct interactions with cellularolecules to cause adverse responses in cells.
cknowledgement
This work was supported by the Korea Research Foundationrant funded by the Korean Government (MOEHRD) (KRF-006-311-D00126) and the Eco-technopia 21 project supportedy the Korea Ministry of the Environment.
eferences
han, W., Shiao, N., Lu, P., 2006. CdSe quantum dots induce apoptosis in humanneuroblastoma cells via mitochondrial-dependent pathways and inhibitionof survival signals. Toxicol. Lett. 167, 191–200.
hen, X., Deng, C., Tang, S., Zhang, M., 2007. Mitochondria-dependent apopto-sis induced by nanoscale hydroxyapatite in human gastric cancer SGC-7901cells. Biol. Pharm. Bull. 30 (1), 128–132.
urtis, J., Greenberg, M., Kester, J., Phillips, S., Krieger, G., 2006. Nanotech-nology and nanotoxicology: a primer for clinicians. Toxicol. Rev. 25 (4),245–260.
as, K.C., 2005. Thioredoxin and its role in premature newborn biology.Antioxid. Redox. Signal. 7, 1740–1743.
har-Mascareno, M., Carcamo, J.M., Golde, D.W., 2005. Hypoxia-reoxygenation-induced mitochondrial damage and apoptosis in humanendothelial cells are inhibited by vitamin C. Free Radical Biol. Med. 38,1311–1322.
as, M., Patil, S., Bhargava, N., Kang, J., Riedel, L.M., Seal, S., Hickman, J.J.,2007. Auto-catalytic ceria nanoparticles offer neuroprotection to adult ratspinal cord neurons. Biomaterials 28, 1918–1925.
lbekai, R.H., El-Kadi, A.O.S., 2005. The role of oxidative stress in the mod-ulation of aryl hydrocarbon receptor-regulated genes by As3+, Cd2+, andCr6+. Free Radical Biol. Med. 39, 1499–1511.
oster, K.A., Galeffi, F., Gerich, F.J., Turner, D.A., Muller, M., 2006. Opticaland pharmacological tools to investigate the role of mitochondria duringoxidative stress and neurodegeneratation. Prog. Neurobiol. 79, 136–171.
otakis, G., Cemeli, E., Anderson, D., Timbrell, J.A., 2005. Cadmium chloride-induced DNA and lysosomal damage in a hepatoma cell line. Toxicol. InVitro 19, 481–489.
ao, F., Lu, Q., Komarneni, S., 2006. Fast synthesis of cerium oxide nanopar-ticles and nanorods. J. Nanosci. Nanotechnol. 6 (12), 3812–3819.
reen, M., Howman, E., 2005. Semiconductor quantum dots and free radicalinduced DNA nicking. Chem. Commun. 121, 121–123.
ansen, J.M., Zhang, H., Hones, D.P., 2006. Differential oxidation ofthioredoxin-1, thioredoxin-2, and glutathione by metal ions. Free RadicalBiol. Med. 40, 138–145.
ardman, R., 2006. A Toxicological review of quantum dots: toxicity dependson physicochemical and environmental factors. Environ. Health Perspect.114 (2), 165–172.
issin, P.J., Hilf, R., 1976. Fluorometric method for determination of oxidizedand reduced glutathione in tissues. Anal. Biochem. 74, 214–226.
ussain, S.M., Hess, K.L., Gearhart, J.M., Geiss, K.T., Schlager, J.J., 2005. Invitro toxicity of nanoparticles in BRL 3A rat liver cells. Toxicol. In Vitro 19,975–983.
S
T
y 245 (2008) 90–100 99
ezek, P., Hlavata, L., 2005. Mitochondria in homeostasis of reactive oxy-gen species in cell, tissues, and organism. Int. J. Biochem. Cell Biol. 37,2478–2503.
agan, V.E., Bayer, H., Shvedova, A.A., 2005. Nanomedicine and nanotoxicol-ogy: two sides of the same coin. Nanomedicine 1 (4), 313–316.
anupriya, Prasad, D., Ram, M.S., Sawhney, R.C., Ilavazhagan, G., Banerjee,P.K., 2007. Mechanism of tert-butylhydroperoxide induced cytotoxicity inU-937 macrophages by alteration of mitochondrial function and generationof ROS. Toxicol. In Vitro 21, 846–854.
aur, P., Aschner, M., Syversen, T., 2006. Glutathione modulation influencesmethyl mercury induced neurotoxicity in primary cell cultures of neuronsand astrocytes. Neurotoxicology 27, 492–500.
im, J., Sharma, R.P., 2006. Cadmium-induced apoptosis in murinemacrophages is antagonized by antioxidants and caspase inhibitors. J. Tox-icol. Environ. Health A 69 (12), 1182–1201.
ipen, H.M., Laskin, D.L., 2005. Smaller is not always better: nanotechnol-ogy yields nanotoxicology. Am. J. Physiol.-Lung Cell Mol. Physiol. 289,L696–L697.
i, Z., Hulderman, T., Salmen, R., Chapman, R., Leonard, S.S., Young, S.,Shvedova, A., Luster, M.I., Simeonova, P.P., 2006. Cardiovascular effectsof pulmonary exposure to single-wall carbon nanotubes. Environ. HealthPerspect. 115, 377–382.
in, W., Huang, Y., ZAhou, X., Ma, Y., 2006. Toxicity of cerium oxide nanopar-ticles in human lung cancer cells. Int. J. Toxicol. 25, 451–457.
imbach, L.K., Wick, P., Manser, P., Grass, R.N., Bruinink, A., Stark, W.J.,2007. Exposure of engineered nanoparticles to human lung epithelial cells:influence of chemical composition and catalytic activity on oxidative stress.Environ. Sci. Technol. 41 (11), 4158–4163.
iu, J., Lei, D., Waalkes, M.P., Beliles, R.P., Morgan, D.L., 2003. Genomicanalysis of the rat lung following elemental mercury vapor exposure. Toxicol.Sci. 74, 174–181.
ong, T.C., Saleh, N., Tilton, R.D., Lowry, G.V., Veronesi, B., 2006. Titat-nium dioxide (P25) produces reactive oxygen species in immortalized brainmicroglia (BV2): implications for nanoparticle neurotoxicity. Environ. Sci.Technol. 40 (14), 4346–4352.
argonis, K., Fatouros, I.G., Jamurtas, A.Z., Kikolaidis, M.G., Douroudos, I.,Chatzinikolaou, A., Mitrakou, A., Mastorakos, G., Papassotiriou, I., Tax-ildaris, K., Kouretas, D., 2007. Oxidative stress biomarkers responses tophysical overtraining: implications for diagnosis. Free Radical Biol. Med.43, 901–910.
ulero, M., Romeu, M., Giralt, M., Folch, J., Nogues, M.R., Fortuno, A.,Sureda, F.X., Linares, V., Cabre, M., Paternain, J.L., Mallol, J., 2006. Oxida-tive stress-related markers and langerhans cells in a hairless rat modelexposed to UV radiation. J. Toxicol. Environ. Health A 69 (14), 1371–1385.
iu, J., Axfer, A., Rogers, L.M., Wang, X., Kolattukudy, P.E., 2007. Cardiopro-tective effects of cerium oxide nanoparticles in a t ransgenic murine modelof cardiomyopathy. Cardiovasc. Res. 73 (3), 549–559.
berdorster, G., Oberdoster, E., Oberdorster, J., 2005. Nanotoxicology: anemerging discipline evolving from studies of ultrafine particles. Environ.Health Perspect. 113 (7), 823–829.
ark, E.J., Park, K., 2006. Gene expression profiles of cultured rat cardiomy-ocytes (H9C2 cells) in response to arsenic trioxide at subcytotoxic level andoxidative stress. J. Health Sci. 52, 512–521.
ark, E.J., Park, K., 2007. Induction of reactive oxygen species and apoptosisin BEAS-2B cells by mercuric chloride. Toxicol. In Vitro 21, 789–794.
orter, A.G., 1999. Protein translocation in apoptosis. Trend. Cell Biol. 9,394–401.
riestly, B.G., Harford, A.J., Rsim, M.R., 2007. Nanotechnology: a promisingnew technology-but how safe? Med. J. Aust. 186, 187–188.
ayes, C.M., Gobin, A.M., Ausman, K.D., Mendez, J., West, J.L., Colvin, V.L.,2005. Nano-C60 cytotoxicity is due to lipid peroxidation. Biomaterials 26,7587–7595.
chubert, D., Dargusch, R., Raitano, J., Chan, S., 2006. Cerium and yttriumoxide nanoparticles are neuroprotective. Biochem. Biophys. Res. Commun.32, 86–91.
hill, A., Zeyons, O., Spalla, O., Chauvat, F., Rose, J., Auffan, M., Flank, A.M.,2006. Cytotoxicity of CeO2 nanoparticles for Escherichia coli, physico-

1 colog
V
W
W
W
X
induce mitochondrial dysfunction: implications for ultrafine particle toxicity.
00 E.-J. Park et al. / Toxi
chemical insight of the cytotoxicity mechanism. Environ. Sci. Technol. 40,6151–6156.
alko, M., Rhodes, C.J., Moncol, J., Izakovic, M., Mazur, M., 2006. Free radi-cals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol.Interact. 160, 1–40.
ang, J., Xiao, B., Zheng, J., Chen, H., Zou, S., 2007. Effect of targeted magneticnanoparticles containing 5-FU on expression of bcl-2, bax and caspase 3 in
nude mice with transplanted human liver cancer. World J. Gastroenterol. 13(23), 3171–3175.arheit, D.B., Borm, P.J., Hennes, C., Lademann, J., 2007. Testing strateges toestablish the safety of nanomaterials: conclusion of an ECETOC workshop.Inhal. Toxicol. 19 (8), 631–643.
Y
y 245 (2008) 90–100
ittmaack, K., 2007. In search of the most relevant parameter for quan-tifying lung inflammatory response to nanoparticle exposure: particlenumber, surface area, or what? Environ. Health Perspect. 115 (2), 187–194.
ia, T., Korge, P., Weiss, J.N., Li, N., Venkatesen, M.I., Sioutas, C., Nel, A.,2004. Quinones and aromatic chemicals compounds inparticulate matter
Environ Health Perspect. 112, 1347–1358.in, H., Too, H.P., Chow, G.M., 2005. The effects of particle size and sur-
face coating on the cytotoxicity of nickel ferrite. Biomaterials 26, 5818–5826.





![FINANCIAL MANAGEMENT 2B [BSR2B01, FNM02B2] LAST ...](https://static.fdokumen.com/doc/165x107/633c134708bfd70fe20afb65/financial-management-2b-bsr2b01-fnm02b2-last-.jpg)














