
Sonographic structure of isolation-induced ultrasonic calls of rat pups
10
q 1999 John Wiley & Sons, Inc CCC 1012-1630/99/030195-10 Stefan M. Brudzynski Department of Psychology, Brock University, St. Catharines, Ontario, L2S 3A1 Canada Priscilla Kehoe Megan Callahan Neuroscience Program, Trinity College, Hartford, CT 06106 Sonographic Structure of Isolation-Induced Ultrasonic Calls of Rat Pups Received 15 April 1997; accepted 8 September 1998 ABSTRACT: Sonographic analysis of isolation-induced calls of 10- to 17-day-old Sprague- Dawley rat pups showed that average acoustic parameters of calls changed with pups’ age. Average call duration increased with age from 80 ms to over 140 ms in 15-day-old pups. The peak frequency increased from approximately 50 kHz to an average of 64 kHz in 17-day-old pups, and the average bandwidth increased from 34 to 45 kHz in 17-day-old pups. Analysis of the sonographic structure of pup calls additionally revealed a tendency to produce two or more alternating sweeps of sound frequency in each call. Development of sweeps is the most typical feature of pup calls with a dominant call type resembling “U” or inverted “U” shape in the sonogram. Number of “U” or inverted “U” call types significantly increased with pups’ age. It is concluded that pups developed and strengthened those acoustic features of distress calls which play a role in intraspecific communication and maximize pup survival. q 1999 John Wiley & Sons, Inc. Dev Psychobiol 34: 195– 204, 1999 Keywords: ultrasonic vocalization; isolation calls; distress calls; rat pups; vocal communi- cation; sonographic analysis Infant rats emit ultrasonic vocalization in a number of distress situations such as separation of the neonates from the mother, acute isolation from the nest, unfa- miliar environment or odors, thermal or tactile stimuli, etc. (Allin & Banks, 1971, 1972; Conely & Bell, 1978; Gardner, 1985; Ha ˚rd & Engel, 1988; Hofer & Shair, 1978, 1980; Insel, Hill, & Mayor, 1986; Insel, Miller, Gelhard, & Hill, 1988; Kehoe & Blass, 1986a, 1986b; Miczek, Tornatzky, & Vivian, 1991; Noirot, 1968, 1972; Okon, 1971; Oswalt & Meier, 1975; Sales & Pye, 1974). Behavioral and physiological studies of the mother – pup relationship clearly indicate that Correspondence to: S. M. Brudzynski Contract grant sponsor: Natural Sciences and Engineering Re- search Council of Canada Contract grant number: RGPIN 98 emission of ultrasonic calls by pups in dangerous or potentially dangerous situations plays an adaptive role in activating maternal response directed to the endan- gered pup(s). Pup ultrasonic isolation calls elicit searching and retrieval behaviors by the dam and shorten the response latency to transport litters from an endangered nest (Allin & Banks, 1972; Brewster & Leon, 1980; Smotherman, Bell, Starzec, Elias, & Zachman, 1974). The calls elicit not only maternal search behavior but have been shown to act as direc- tional cues for the dams (Brunelli, Shair, & Hofer, 1994). The isolation calls also contribute to an increase in anogenital licking by the dam following retrieval and induce prolactin release in lactating females (Brouette-Lahlou, Vernet-Maury, & Vigouroux, 1992; Terkel, Damassa, & Sawer, 1979; Voloschin & Tra- mezzani, 1984). There is also evidence that the rat pup
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Sonographic structure of isolation-induced ultrasonic calls of rat pups

q 1999 John Wiley & Sons, Inc CCC 1012-1630/99/030195-10
DEV (WILEJ) RIGHT INTERACTIVE
shortstandardlong
Stefan M. BrudzynskiDepartment of Psychology, Brock
University, St. Catharines, Ontario,L2S 3A1 Canada
Priscilla KehoeMegan Callahan
Neuroscience Program, TrinityCollege, Hartford, CT 06106
Sonographic Structure of
Isolation-Induced Ultrasonic
Calls of Rat Pups
Received 15 April 1997; accepted 8 September 1998
ABSTRACT: Sonographic analysis of isolation-induced calls of 10- to 17-day-old Sprague-Dawley rat pups showed that average acoustic parameters of calls changed with pups’ age.Average call duration increased with age from 80 ms to over 140 ms in 15-day-old pups. Thepeak frequency increased from approximately 50 kHz to an average of 64 kHz in 17-day-oldpups, and the average bandwidth increased from 34 to 45 kHz in 17-day-old pups. Analysis ofthe sonographic structure of pup calls additionally revealed a tendency to produce two or morealternating sweeps of sound frequency in each call. Development of sweeps is the most typicalfeature of pup calls with a dominant call type resembling “U” or inverted “U” shape in thesonogram. Number of “U” or inverted “U” call types significantly increased with pups’ age.It is concluded that pups developed and strengthened those acoustic features of distress callswhich play a role in intraspecific communication and maximize pup survival. q 1999 JohnWiley & Sons, Inc. Dev Psychobiol 34: 195–204, 1999
Keywords: ultrasonic vocalization; isolation calls; distress calls; rat pups; vocal communi-cation; sonographic analysis
Infant rats emit ultrasonic vocalization in a numberof distress situations such as separation of the neonatesfrom the mother, acute isolation from the nest, unfa-miliar environment or odors, thermal or tactile stimuli,etc. (Allin & Banks, 1971, 1972; Conely & Bell, 1978;Gardner, 1985; Hard & Engel, 1988; Hofer & Shair,1978, 1980; Insel, Hill, & Mayor, 1986; Insel, Miller,Gelhard, & Hill, 1988; Kehoe & Blass, 1986a, 1986b;Miczek, Tornatzky, & Vivian, 1991; Noirot, 1968,1972; Okon, 1971; Oswalt & Meier, 1975; Sales &Pye, 1974). Behavioral and physiological studies ofthe mother–pup relationship clearly indicate that
Correspondence to: S. M. BrudzynskiContract grant sponsor: Natural Sciences and Engineering Re-
search Council of CanadaContract grant number: RGPIN 98
emission of ultrasonic calls by pups in dangerous orpotentially dangerous situations plays an adaptive rolein activating maternal response directed to the endan-gered pup(s). Pup ultrasonic isolation calls elicitsearching and retrieval behaviors by the dam andshorten the response latency to transport litters froman endangered nest (Allin & Banks, 1972; Brewster &Leon, 1980; Smotherman, Bell, Starzec, Elias, &Zachman, 1974). The calls elicit not only maternalsearch behavior but have been shown to act as direc-tional cues for the dams (Brunelli, Shair, & Hofer,1994). The isolation calls also contribute to an increasein anogenital licking by the dam following retrievaland induce prolactin release in lactating females(Brouette-Lahlou, Vernet-Maury, & Vigouroux, 1992;Terkel, Damassa, & Sawer, 1979; Voloschin & Tra-mezzani, 1984). There is also evidence that the rat pup

196 Brudzynski, Kehoe, and Callahan
DEV (WILEJ) LEFT INTERACTIVE
shortstandardlong
calls are specifically addressed to their mother and thatthe number of calls of 12- to 14-day-old pups wassignificantly decreased in the presence of an unfamil-iar adult male (Takahashi, 1992a, 1992b, 1994). Sim-ilar maternal responses of locating and retrieving thevocalizing pup and increased nest-building activitiesin response to pup calls have been reported in houseand field mice (Noirot, 1972; Sewell, 1970).
These findings support the hypothesis that pup callsconvey biologically significant information for themother and trigger appropriate maternal responseswhich increase the chances of pup survival (Brunelliet al., 1994; Hofer, 1994a, 1994b). The pups’ distresscalls may serve as a general signal which producesmother’s arousal and caregiving behavior (Bell, 1974,1979) or the calls may be characteristic for a particularsituation, e.g., thermal versus tactile stimuli, and ini-tiate a specific type of maternal response such assearching for pups versus preventing a possible ag-gression of the young (Brunelli et al., 1994; Noirot,1972).
The pup calls are characterized by a high degree ofvariability, particularly in their production rate andrange of sound frequencies. Little is known about du-ration of these calls. In 5-day-old pups, for example,duration of single calls varies from less than 20 tomore than 160 ms (Elsner, Suter, & Alder, 1990). Anaverage sound frequency ranges from 30 to 65 kHz,and call repetition rate varies greatly depending on theparadigm but usually does not exceed 80–90 calls/min(Brunelli, Keating, Hamilton, & Hofer, 1996; Carden& Hofer, 1992; Elsner et al., 1990; Goodwin, Molina,& Spear, 1994; Kehoe & Harris, 1989; Nyby & Whit-ney, 1978). Assessment of the sound frequency of adistress call is difficult because of high variability ofsonographic shape (Callahan, Kehoe, & Brudzynski,1996; Elsner et al., 1990; Miczek et al., 1991), i.e.,rapid changes of sound frequency during emission ofa single call. Because of these varying frequencysweeps, there are rarely two identical calls emitted bythe same pup over a long period of observation. Be-cause the calls are so variable, the question arises asto what are the acoustic features of these calls whichconvey information for the dam and help to localizethe pup. The goal of the present study is to analyzesonographic structure and other acoustic features ofisolation calls induced by isolation in 10- to 17-day-old rat pups. Although ultrasonic calls could be ana-lyzed in an automated manner (Brudzynski & Bihari,1990; Brudzynski, Ociepa, & Bihari, 1991; Elsner etal., 1990), each call has been analyzed individually inthe present study. This approach guaranteed that noneof the short calls were overlooked and that the longercalls were not broken into smaller components. Thistype of analysis allows for recognition of changes in
the temporal structure of sound frequency for eachcall.
METHODS
Subjects
Primiparous and multiparous females of Sprague-Dawley rats were mated with males of the same strain(Charles River Laboratories, Wilmington, MA) in theTrinity College animal colony and their litters wereused as subjects. The pregnant females were housedindividually in plastic cages with water and Purina RatChow available ad libitum. The colony room wasmaintained at 227C and with a 12:12 hr light:dark cy-cle (7 a.m. to 7 p.m. light). All recorded newborn pupsup to 5 p.m. were considered to be born on that day(Day 0). The litters were culled to 12 pups on Day 1.A total of 235 pups from 24 litters were studied; 78pups (seven litters) were 10 days of age, 86 pups (10litters) were 15 days of age, and 71 pups (seven litters)were 17 days of age.
Isolation Procedure
On Day 10, the dam was placed in a separate cage andpups remained in a huddle maintained at 307C by aheating pad placed under the cage. Pups were thenindividually removed from the nest at random andgently placed in a 22 3 22 cm black wooden box witha wire-mesh bottom. Because the number of calls de-creases with age from approximately Day 10, a coolerambient temperature was selected for older rats in or-der to maintain sufficient number of emitted calls foranalysis. Acoustic parameters of calls were analyzedbut the number of calls per time unit was not takeninto consideration. As tested in a group of eight 10-day-old pups, the change of the ambient temperatureby 67C did not influence the acoustic parameters ofpup calls, t(7) 5 0.8 to 1.8, n.s. The box temperaturewas 27 6 27C, for 10-day-old pups and 20 6 27C for15 and 17-day-old pups. Each pup was recorded for 5min and returned to the huddle.
Recording and Analysis of Ultrasonic Calls
An ultrasonic microphone (Ultrasound Advice, Lon-don, England, Model SM1, working range 10–180kHz) was suspended 8 cm above the bottom of therecording cage and was connected to a bat detector(QMC Instruments, Ltd, London, England, ModelS200, frequency range 10–180 kHz). Using the broad-band output of the bat detector with the frequency di-vision of 1:16, all ultrasonic calls were recorded on acassette recorder (Realistic, VSC-2001) for furtheranalysis.

Rat Pup Isolation Calls 197
DEV (WILEJ) RIGHT INTERACTIVE
shortstandardlong
FIGURE 1 Explanation of sonographic analysis of an ex-emplary isolation call obtained from a 10-day-old rat pup.Sonographic structures of the call are shown in the lowergraph as changes of sound frequency (kHz) in time (ms).The duration of the call was measured from the sonogram(68.75 ms in this example) and power spectrum was ob-tained for all sounds between the beginning and end of thecall. The power spectrum is shown in the upper graph aschanges of relative power (2dB) across sound frequency(kHz). Relative power is calculated from the waveform dis-play, referenced from the top of the scale, and expressed innegative decibels. The lowest and the highest point of thepower spectrum refer to the highest and lowest frequency inthe sonogram (22.1 kHz and 63.5 kHz). Difference betweenthese frequencies, calculated from the power spectrum, wasregarded as bandwidth (41.4 kHz). The highest peak in thepower spectrum (44.48 kHz in this example) was recordedas the peak frequency. Its location is also indicated on thesonogram.
Signals from the tape recorder were analyzed onthe DSP Sona-Graph signal analysis work station(Model 5500, Kay Elemetrics Corp., Pine Brook, NJ).All calls recorded during a 5-min test or the first 60successive calls per pup were analyzed sonographi-cally, whichever came sooner. Sonograms and powerspectra were obtained for individual calls. Sound fre-quency and duration of each call were measured onthe monitor by movable cursors. Single call durationwas measured between the cursor displays positionedat the beginning and the end of a given call. The powerspectrum was then obtained for that call and its peakfrequency measured, i.e., the strongest sound fre-quency indicated by the highest peak in the powerspectrum (Figure 1). The power spectra showed a rel-ative power of calls as an average of many transformsbetween cursors with a 30% overlap to ensure that alldata points were included. The distance between twocursors positioned 17 dB above the noise level, at thebeginning and the end of each power spectrum, wasregarded as bandwidth. Figure 1 illustrates the wayacoustic parameters were measured for an exemplarycall. For the range of input frequency and transformsize selected, frequency and time resolution was 59Hz and 25 ms for power spectra, and 20 Hz and 6.25ms for sonograms. Selected sonograms and powerspectra were printed on a Gray Scale Printer (Model5509, Kay Elemetric Corp.).
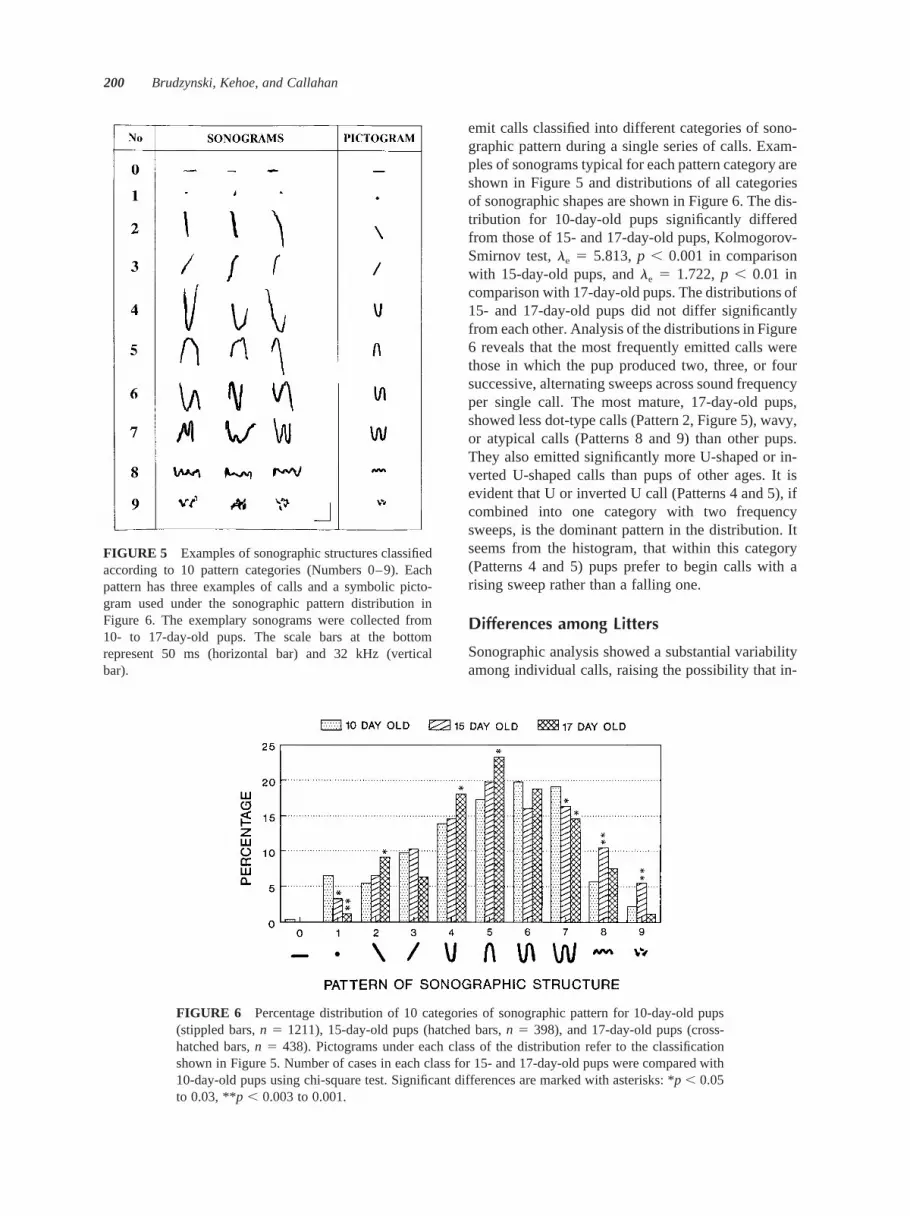
Finally, sonograms of all analyzed calls were as-sessed to identify their structural features. Based onprevious experience, 10 categories of call patternswere distinguished and numbered from 0 to 9. Theywere as follows:
0 - constant sound frequency maintained through-out the duration of the call
1 - very brief call resembling a dot or short commaon the sonogram
2 - continuously rising frequency (rising sweep) orrising tendency (i.e., consistent frequency rise witheventual small breaks or deflections)
3 - continuously falling frequency (falling sweep)or falling tendency
4 - fall and a subsequent rise in sound frequency ina shape approximating the letter “U”
5 - rise and a subsequent fall in sound frequency ina shape approximating inverted “U”
6 - three sweeps of sound frequency within a singlecall in the sequence fall– rise– fall or rise– fall– rise
7 - four sweeps of sound frequency within a singlecall in a shape of the letter “M” or “W”
8 - multiple sweeps of sound frequency appearingin the sonogram as a wavy line without a clear overalltendency to rise or fall
9 - complex sonogram not resembling any of thepatterns mentioned above
Two researchers independently assessed each of thecall shape and classified it according to the nine patterncategories. The results of the assessments were com-pared. There was no statistical difference between dis-tributions obtained by the two assessments, Kolmo-gorov-Smirnov test, n 5 1211, le 5 0.425, n.s. A totalof 2047 calls from 235 pups of 10–17 days of agewere measured and classified.
Statistics
Differences between distributions were tested usingthe Kolmogorov-Smirnov test (Dayton & Stunkard,

198 Brudzynski, Kehoe, and Callahan
DEV (WILEJ) LEFT INTERACTIVE
shortstandardlong
FIGURE 2 Changes of average call duration (stippled bars), average peak frequency (hatchedbars), and average bandwidth (cross-hatched bars) for calls obtained from 10-day-old pups (n 51211), 15-day-old pups (n 5 398), and 17-day-old pups (n 5 438). Pup age is expressed in daysfrom Day 0, and sound frequency and bandwidth in kHz. The vertical lines represent SEMs. Thefirst three bars relate to the scale on the left and the remaining bars to the scale on the right side.Using a t test, averages of the parameters were compared to those of the 10-day-old pups: *p ,0.002, **p , 0.0001.
1971) by comparison of cumulative proportions fortwo populations, and were followed by a chi-squaretest for each class. Differences in other parametersacross ages were tested by the Student t test for in-dependent comparisons. Differences in duration, fre-quency, and bandwidth across litters were tested witha one-way ANOVA for nonpaired comparisons andwith a Kurskal-Wallis one-way ANOVA for compar-ison of the sonographic shape.
RESULTS
Call Duration
Average call duration of 80.89 6 1.1 ms (Figure 2,stippled bars) emitted by 10-day-old pups almost dou-bled from Day 10 to Day 15 and significantly in-creased to 144.03 6 4.8 ms in 15-day-old pups,t(438) 5 212.6, p , 0.0001. It remained at that levelin 17-day-old pups, t(504) 5 212.8, p , 0.0001, incomparison to 10-day-olds). There was no significantdifference in average call duration between 15- and17-day-old pups, t(792) 5 21.49, n.s. The minimalcall duration was 9.37 ms for a 10-day-old pup andthe maximal was 615.6 ms for a 15-day-old pup. Dis-tributions of call duration for 10- to 17-day-old pupsis shown in Figure 3a. The distribution for 10-day-oldpups significantly differed from that for 15- and 17-day-old pups, Kolmogorov-Smirnov test, le 5 6.28and 6.54, p , 0.001. There was no difference between
distributions for 15- and 17-day-old pups. The distri-butions indicate that call duration for 10-day-olds wasmostly contained between 10–200 ms while calls of15- and 17-day-old pups were contained between 10and over 450 ms. The histogram (Figure 3a) showsthat calls lasting approximately 200–400 ms were al-most exclusively emitted by 15- to 17-day-old pups.
Sound Frequency
Average peak frequency of distress calls showed asteady and significant increase across ages (Figure 2,hatched bars). Average peak frequency of 49.5 6 0.73kHz for 10-day- old pups increased to 53.24 6 0.94kHz in 15-day-old pups, t(910) 5 23.13, p , 0.002,and further increased to 64.2 6 1.4 kHz in 17-day-oldpups, t(754) 5 6.47, p , 0.0001 in comparison to 15-day-old pups, and t(689) 5 29.28, p , 0.0001, incomparison to 10-day-old pups. The lowest peak fre-quency of calls was in the audible range (1.92 kHz) insome 10-day-old pups and the highest peak frequencyreached 124.48 kHz in a 17- day-old pup. Distribu-tions of the peak frequency are shown in Figure 3b.The distributions for all ages significantly differed fromeach other, Kolmogorov-Smirnov test, le 5 2.285 to4.322, p , 0.001 for all comparisons. The distributionsindicate the largest number of calls with frequenciesbetween approximately 40–70 kHz for all ages. Therewere, however, significantly more calls with peakfrequency between 10–25 kHz in 10-day-old pups

Rat Pup Isolation Calls 199
DEV (WILEJ) RIGHT INTERACTIVE
shortstandardlong
FIGURE 3 Histograms of percentage distribution of callduration (A), peak sound frequency (B), and bandwidth (C)for 10-day-old pups (black columns), 15-day-old pups(hatched columns), and 17-day-old pups (cross-hatched col-umns). The class width for call duration was 30 ms, forsound frequency 11 kHz, and for bandwidth 15 kHz. Num-ber of cases in each class for 15- and 17-day-old pups werecompared with 10-day-old pups using chi-square test. Sig-nificant differences are marked with dots: • p , 0.03 to 0.01,• • p , 0.003 to 0.0001.
than in older pups, significantly more calls with peakfrequency between 50–70 kHz in 15-day-old pupsthan in other ages, and significantly more calls withpeak frequency between 85–120 kHz in 17-day-oldpups than in pups of other ages (see Figure 3b).
Bandwidth
Average bandwidth also showed a steady and signifi-cant increase across ages (Figure 2, cross-hatchedbars). Average bandwidth of 33.59 6 0.56 kHz for10-day-old pups increased to 40.13 6 1.23 kHz in 15-day-old pups, t(570) 5 24.84, p , 0.0001, and furtherincreased to 45.13 6 1.18 kHz in 17-day-old pups,t(830) 5 2.12, p , 0.004 compared to 15-day-oldpups, and t(641) 5 28.79, p , 0.0001 compared to10-day-old pups. The minimal recorded bandwidthwas 2.56 kHz in a 10-day-old pup and the maximalone of 182.24 kHz in a 15-day-old pup. Distributionsof bandwidth are shown in Figure 3c. The distributionsfor all ages significantly differed from each other, Kol-mogorov-Smirnov test, le 5 1.52 to 3.085, p , 0.02to 0.001. There were significantly more calls withbandwidth between 5–15 kHz in 10-day-old pups thanpups of other ages, and significantly more calls withbandwidth between 65–110 kHz in 17-day-old pupsthan in pups of other ages.
Pattern of Sonographic Structure
Pups emitted single calls or calls in short series. In anycase, the sonographic structure of individual calls wasvariable and frequently changed, even during a singleseries of calls (Figure 4). Thus, the same animal could
FIGURE 4 Sonogram of a series of distress calls obtained from a 10-day-old pup. The seriescontains 10 calls with varying sonographic structure (U shape, W shape, N shape, rising sweep, dot).The intercall intervals represent true time spaces between emission of successive calls.
200 Brudzynski, Kehoe, and Callahan
DEV (WILEJ) LEFT INTERACTIVE
shortstandardlong
FIGURE 5 Examples of sonographic structures classifiedaccording to 10 pattern categories (Numbers 0–9). Eachpattern has three examples of calls and a symbolic picto-gram used under the sonographic pattern distribution inFigure 6. The exemplary sonograms were collected from10- to 17-day-old pups. The scale bars at the bottomrepresent 50 ms (horizontal bar) and 32 kHz (verticalbar).
emit calls classified into different categories of sono-graphic pattern during a single series of calls. Exam-ples of sonograms typical for each pattern category areshown in Figure 5 and distributions of all categoriesof sonographic shapes are shown in Figure 6. The dis-tribution for 10-day-old pups significantly differedfrom those of 15- and 17-day-old pups, Kolmogorov-Smirnov test, le 5 5.813, p , 0.001 in comparisonwith 15-day-old pups, and le 5 1.722, p , 0.01 incomparison with 17-day-old pups. The distributions of15- and 17-day-old pups did not differ significantlyfrom each other. Analysis of the distributions in Figure6 reveals that the most frequently emitted calls werethose in which the pup produced two, three, or foursuccessive, alternating sweeps across sound frequencyper single call. The most mature, 17-day-old pups,showed less dot-type calls (Pattern 2, Figure 5), wavy,or atypical calls (Patterns 8 and 9) than other pups.They also emitted significantly more U-shaped or in-verted U-shaped calls than pups of other ages. It isevident that U or inverted U call (Patterns 4 and 5), ifcombined into one category with two frequencysweeps, is the dominant pattern in the distribution. Itseems from the histogram, that within this category(Patterns 4 and 5) pups prefer to begin calls with arising sweep rather than a falling one.
Differences among Litters
Sonographic analysis showed a substantial variabilityamong individual calls, raising the possibility that in-
FIGURE 6 Percentage distribution of 10 categories of sonographic pattern for 10-day-old pups(stippled bars, n 5 1211), 15-day-old pups (hatched bars, n 5 398), and 17-day-old pups (cross-hatched bars, n 5 438). Pictograms under each class of the distribution refer to the classificationshown in Figure 5. Number of cases in each class for 15- and 17-day-old pups were compared with10-day-old pups using chi-square test. Significant differences are marked with asterisks: *p , 0.05to 0.03, **p , 0.003 to 0.001.

Rat Pup Isolation Calls 201
DEV (WILEJ) RIGHT INTERACTIVE
shortstandardlong
stages of pup development (Days 15–17), may reflectdevelopmental emphasis on those acoustic parameterswhich are important for communication. While 17-day-old pups did not increase duration of their callscompared to 15-day-olds, they significantly raised thepeak frequency. Pups of all ages studied were capableof emitting sounds within the range of 90–130 kHz,but only 17-day-old rats used this band as a peak com-ponent of their calls (see Figure 3b).
Development of communication with age in ratpups may be further corroborated by the observationthat 12- to 14-day-old pups but not 7-day-old pupsrespond to the presence of an unfamiliar adult male(Takahashi, 1992a, 1992b). The 12- to 14-day-oldpups showed freezing or avoided the unfamiliar malebut not their mother and emitted less ultrasonic callsin the presence of the male compared to the numberof calls emitted in response to their mother. The abilityof rat pups to inhibit emission of ultrasounds whenthreatened is not present at birth and develops by theend of the 2nd postnatal week (Takahashi, 1992b). Inparallel with these changes, some acoustic parametersof pup calls, which play a role in communication, maybe also strengthened.
Our results also showed that the bandwidth steadilyincreased from Day 10 to Day 17. Pups producesweeps across a larger span of frequencies with age.Although pups do not leave their burrows until about28 days of age (Boice, 1977) and, most probably, theircalls do not travel beyond the burrow, this type ofsound production increases the chances of being no-ticed, localized, and taken care of by mother and islikely to play a critical role in pup survival. The findingthat the mean sound frequency decreased with age(Elsner et al., 1990) while the bandwidth increasedindicates that pups extended their frequency sweeps atthe lower end of sound frequency rather than thehigher end. This particular direction may be dictatedby developmental changes in physical properties of thelarynx (e.g., larger diameter or length of the vocaltract). Disturbances in rat pup development by prenatalexposure to carbon monoxide produced just a selectivedecrease in the minimum frequency of distress calls of5-day-old pups (Di Giovanni et al., 1993). However,it is highly possible that changes of other acoustic fea-tures of distress calls do not reflect development-de-pendent physical changes in the respiratory and laryn-geal structures.
The dynamics of changes in call duration, peak fre-quency, and bandwidth from Day 15 to Day 17 arenot parallel. Therefore, the changes are unlikely to bea simple function of maturation of the respiratory cav-ities and sound-producing structures. The develop-mental increase in the size and length of the larynx
dividual litters may also differ in the acoustic profile.Thus, differences in acoustic parameters were ana-lyzed across litters. The analysis was done for Day-10pups only because they emitted the highest number ofcalls and allowed an individual analysis of n 5 1211calls (i.e., from n 5 33 to 286 calls per litter). Allacoustic parameters differed significantly among lit-ters. Call duration, peak sound frequency, and band-width differed highly significantly among the sevenlitters studied, Figures 7a and 7b, ANOVA, F(6, 1204)5 12.03 to 8.38, p , 0.0001. The sonographic struc-ture also differed highly significantly among litters,Figure 7b, Kurskal-Wallis ANOVA, X2 5 26.34, df5 6, p , 0.0002. The highest range of variability wasfound for call duration and the smallest range forsonographic structure.
DISCUSSION
Analysis of distress calls of 10- to 17-day-old rat pupsshowed that average acoustic parameters changed withpups’ age. Average call duration, peak frequency, andaverage bandwidth increased over 7 days of pup de-velopment from Day 10 to Day 17. Analysis of thesonographic structure of pup calls additionally re-vealed a tendency to produce two or more sweepsacross sound frequency in each call. Production ofsweeps is the most typical feature of pup calls with adominant call type resembling U or inverted U shapein the sonogram. Number of U or inverted U-type callssignificantly increased with pups’ age. Sonographicshape without apparent frequency sweeps decreased incalls of older pups.
Some developmental changes in acoustic parame-ters of distress calls have been reported in rat pups.After an initial increase up to Day 9, calling rate de-clined whereas intercall interval steadily decreasedwith age (Elsner et al., 1990; Insel et al., 1988; Kehoe& Harris, 1989; Noirot, 1968). Call duration increasedfrom Day 5 to Day 13 (Elsner et al., 1990) and thisincrease was further confirmed and extended from Day10 to Day 15 in the present study. In 17-day-old pups,call duration remained at a steady level. While themean sound frequency was reported to decrease withage from Day 5 to Day 15 (Elsner et al., 1990), ourresults showed a growing increase in peak frequencyin all three ages tested. These two results are not con-tradictory because mean frequency and peak fre-quency component represent different measures. Peakfrequency, i.e., the strongest frequency component ofthe call, is more likely to play a role in the intraspecificcommunication rather than the mean frequency. Thus,the increase in the peak frequency, particularly at later

202 Brudzynski, Kehoe, and Callahan
DEV (WILEJ) LEFT INTERACTIVE
shortstandardlong
FIGURE 7 Differences in acoustic parameters across lit-ters (A-G). Changes in call duration and sound frequencyare shown in A and changes in bandwidth and median of thesonographic structure in B. Particular sonographic structuresare numbered according to the scale explained in Figure 5.There are highly significant differences across litters in callduration, ANOVA, F(6, 1204) 5 12.02, p , 0.0001, insound frequency, F(6, 1204) 5 8.67, p , 0.0001, in band-width, F(6, 1204) 5 8.38, p , 0.0001, and in sonographicstructure (X2 5 26.34, df 5 6, p , 0.0002.
and the vocal tract would be expected to lower thefundamental frequency of the sound produced. Such adecrease in the mean frequency with age has been re-ported for rat pups (Elsner et al., 1990) and a decreasein the fundamental frequency with age has been re-cently shown in humans (Stathopoulos & Sapienza,1997), although the mechanism of sound productionin rats is different from that in the human larynx (Rob-erts, 1975). In light of these findings, the increase inthe peak sound frequency with age found in the presentstudy, therefore, is likely to reflect the development ofan active communication rather than a passive physi-cal change caused by maturation.
In addition to the changes of the peripheral respi-ratory system, the central generators of the respiratoryand vocal patterns are subject to robust developmentalchanges. It has recently been found in mice that anage-dependent change in the motor pattern of the res-
piratory generator occurs during the first 15 days ofpup life (Paton & Richter, 1995). The central patterngenerators for production of calls would also be sub-jected to a similar rapid change.
Finally, sonographic analysis of call structure pro-vides further evidence that the developmental changesshow trends emphasizing those acoustic featureswhich are likely to play a role in pup survival. A cleartendency to produce calls with two or more alternatingfrequency sweeps has been observed in this study. Al-ternating frequency sweeps work on the same princi-ple as an ambulance siren and can be more easily de-tected and localized by mothers than a steady sound.An abrupt beginning and end of each call, alternatingfrequency sweeps, and succession of many calls arewell-known features maximizing the speed and accu-racy of sound localization (Marler, 1959; Marler &Hamilton, 1966). The results indicate that two to threesweeps per call represent the most economical patternwhich increased with pup age.
Accordingly, 17-day-old pups produced signifi-cantly more U- or inverted-U-shaped calls than pupsof other ages and significantly decreased the numberof calls with three sweeps per call (see Figure 6). Callswith other sonographic structures may be regarded asfragments of the principal two-sweep pattern, e.g., therising or falling sweep itself, or ill-elaborated soundsparticularly in younger (10- to 15-day-old) pups, e.g.,dot-type (Pattern 1). Fifteen-day-old pups producedsignificantly more complex calls (Patterns 8 and 9)than 10-day-old pups, which may suggest that they aremore prone to produce incomplete or aborted calls,probably by “practicing” and trying different combi-nations of sound patterns.
The basic features of call repetitiveness and fre-quency sweep in rat pup calls, which permit localiza-tion of the signal, have also been observed in otherspecies. Infant bats, for example, produce greater fre-quency sweeps and higher repetition rates than rodents(Gould, 1979), most probably because they live in aconsiderably more “noisy” environment of the com-munal roost of the colony and sonar calls of multipleadults (Gould, 1970). In spite of the noisy acousticenvironment, many bats recognize and nurse only theirown infant (Gould, 1970, 1979). Highly significantdifferences in acoustic parameters among rat littersfound in the present study suggest, therefore, that thedam can recognize calls from her own litter and mayeven be able to recognize calls of individual pups.
Although our data were obtained from the Sprague-Dawley rat strain and may differ from those from otherstrains (Nitschke, Bell, Bell, & Zachman, 1975; Sales,1979), it seems reasonable to postulate in summarythat over the 7 days of development studied, rat pupsgenerated and strengthened those acoustic features of

Rat Pup Isolation Calls 203
DEV (WILEJ) RIGHT INTERACTIVE
shortstandardlong
calls which play a role in intraspecific communicationand maximize pup survival.
NOTES
The technical assistance of Ms. Anna Pniak is greatlyacknowledged.
REFERENCES
Allin, J. T., & Banks, E. M. (1971). Effects of temperatureon ultrasound production by infant albino rats. Develop-mental Psychobiology, 4, 149–156.
Allin, J. T., & Banks, E. M. (1972). Functional aspects ofultrasound production by infant albino rat (Rattus norv-egicus). Animal Behaviour, 20, 175–185.
Bell, R. W. (1974). Ultrasounds in small rodents: Arousal-produced and arousal-producing. Developmental Psycho-biology, 7, 39–42.
Bell, R. W. (1979). Ultrasonic control of maternal behavior:Developmental implications. American Zoologist, 19,413–418.
Boice, R. (1977). Burrows of wild and albino rats: Effectsof domestication, outdoor raising, age, experience, andmaternal state. Journal of Comparative and PhysiologicalPsychology, 91, 649–661.
Brewster, J., & Leon, M. (1980). Relocation of the site ofmother–young contact: Maternal transport behavior inNorway rats. Journal of Comparative and PhysiologicalPsychology, 94, 69–79.
Brouette-Lahlou, I., Vernet-Maury, E., & Vigouroux, M.(1992). Role of pups’ ultrasonic calls in a particular ma-ternal behavior in Wistar rat: Pups’ anogenital licking.Behavioural Brain Research, 50, 147–154.
Brudzynski, S. M., & Bihari, F. (1990). Ultrasonic vocali-zation in rats produced by cholinergic stimulation of thebrain. Neuroscience Letters, 109, 222–226.
Brudzynski, S. M., Ociepa, D., & Bihari, F. (1991). Com-parison between cholinergically and naturally induced ul-trasonic vocalization in the rat. Journal of Psychiatry andNeuroscience, 16, 221–226.
Brunelli, S. A., Shair, H. N., & Hofer, M. A. (1994). Hy-pothermic vocalizations of rat pups (Rattus norvegicus)elicit and direct maternal search behavior. Journal ofComparative Psychology, 108, 298–303.
Brunelli, S. A., Keating, C. C., Hamilton, N. A., & Hofer,M. A. (1996). Development of ultrasonic vocalization re-sponses in genetically heterogenous National Institute ofHealth (N:NIH) rats: I. Influence of age, testing experi-ence, and associated factors. Developmental Psychobiol-ogy, 29, 507–516.
Callahan, M., Kehoe, P., & Brudzynski, S. M. (1996). Theeffect of cholinergic stimulation on rat pup vocalizations.Developmental Psychobiology, 29, 281.
Carden, S. E., & Hofer, M. A. (1992). Effect of social com-
panion on the ultrasonic vocalizations and contact re-sponses of 3-day-old rat pups. Behavioral Neuroscience,106, 421–426.
Conely, L., & Bell, R. W. (1978). Neonatal ultrasounds elic-ited by odor cues. Developmental Psychobiology, 11,193–197.
Dayton, C. M., & Stunkard, C. L. (1971). Statistics for prob-lem solving, New York: McGraw-Hill.
Di Giovanni, V., Cagiano, R., De Salvia, M. A., Giustino,A., Lacomba, C., Renna, G., & Cuomo, V. (1993). Neu-robehavioral changes produced in rats by prenatal expo-sure to carbon monoxide. Brain Research, 616, 126–131.
Elsner, J., Suter, D., & Alder, S. (1990). Microanalysis ofultrasound vocalizations of young rats: Assessment of thebehavioral teratogenicity of methylmercury. Neurotoxi-cology & Teratology, 12, 7–14.
Gardner, C. R. (1985). Distress vocalization in rat pups. Asimple screening method for anxiolytic drugs. Journal ofPharmacological Methods, 14, 181–187.
Goodwin, G. A., Molina, V. A., & Spear, L. P. (1994). Re-peated exposure of rat pups to isolation attenuates isola-tion-induced ultrasonic vocalization rates: Reversal withnaltrexone. Developmental Psychobiology, 27, 53–64.
Gould, E. (1970). Echolocation and communication in bats.In B. Slaughter & D. Walton, (Eds.), About bats (pp. 144–161). Dallas: Southern Methodist University Press.
Gould, E. (1979). Neonatal vocalizations of ten species ofMalaysian bats (Megachiroptera and Microchiroptera).American Zoologist, 19, 481–491.
Hard, E., & Engel, J. (1988). Effects of 8-OH-DPAT onultrasonic vocalization and audiogenic immobility reac-tion in preweanling rats. Neuropharmacology, 27, 981–986.
Hofer, M. A. (1994a). Hidden regulators in attachment, sep-aration, and loss. Monographs of the Society for Researchin Child Development, 59, 192–207.
Hofer, M. A. (1994b). Early relationships as regulators ofinfant physiology and behavior. Acta Paediatrica Scan-dinavica (Suppl. 397), 9–18.
Hofer, M. A., & Shair, H. (1978). Ultrasonic vocalizationduring social interaction and isolation in 2-week-old rats.Developmental Psychobiology, 11, 495–504.
Hofer, M. A., & Shair, H. (1980). Sensory processes in thecontrol of isolation-induced ultrasonic vocalization by 2-week-old-rats. Journal of Comparative and PhysiologicalPsychology, 94, 271–279.
Insel, T. R., Hill, J. L., & Mayor, R. B. (1986). Rat pupultrasonic isolation calls: Possible mediation by the ben-zodiazepine receptor complex. Pharmacology Biochem-istry and Behavior, 24, 1263–1267.
Insel, T. R., Miller, L. P., Gelhard, R. G., & Hill, J. (1988).Rat pup ultrasonic isolation calls and the benzodiazepinereceptor. In J. D. Newman (Ed.), The physiology of mam-malian vocalization (pp. 331–342). New York: PlenumPress.
Kehoe, P., & Blass, E. M. (1986a). Behaviorally functionalopioid system in infant rats. II. Evidence for pharmaco-logical, physiological, and psychological mediation ofpain and stress. Behavioral Neuroscience, 100, 624–630.

204 Brudzynski, Kehoe, and Callahan
DEV (WILEJ) LEFT INTERACTIVE
shortstandardlong
Kehoe, P., & Blass, E. M. (1986b). Opioid mediation ofseparation distress in 10-day-old rats: Reversal of stresswith maternal stimuli. Developmental Psychobiology, 19,389–398.
Kehoe, P., & Harris, J. C. (1989). Ontogeny of noradrenergiceffects on ultrasonic vocalizations in rat pups. BehavioralNeuroscience, 103, 1099–1107.
Marler, P. (1959). Developments in the study of animal com-munication. In P. R. Bell (Ed.), Darwin’s biological work.Some aspects reconsidered (pp. 150&206). Cambridge:University of Cambridge Press.
Marler, P., & Hamilton, W. J., III. (1966). Mechanisms ofanimal behavior. New York: Wiley.
Miczek, K. A., Tornatzky, W., & Vivian, J. (1991). Ethologyand neuropharmacology: Rodent ultrasounds. In B. Oliv-ier, J. Mos, & J. L. Slangen (Eds.), Animal models inpsychopharmacology [Advances in Pharmacological Sci-ences (pp. 409–427)]. Basel: Birkhauser Verlag.
Nitschke, W., Bell, R. W., Bell, N. J., & Zachman, T. (1975).The ontogeny of ultrasounds in two strains of Rattus norv-egicus. Experimental Aging Research, 1, 229–242.
Noirot, E. (1968). Ultrasounds in young rodents. II. Changeswith age in albino rats. Animal Behaviour, 16, 129–134.
Noirot, E. (1972). Ultrasounds and maternal behavior insmall rodents. Developmental Psychobiology, 5, 371–387.
Nyby, J., & Whitney, G. (1978). Ultrasonic communicationof adult myomorph rodents. Neuroscience and Biobehav-ioral Reviews, 2, 1–14.
Okon, E. E. (1971). The temperature relations of vocaliza-tion in infant Golden hamsters and Wistar rats. Journal ofZoology (London), 164, 227–237.
Oswalt, G. L., & Meier, G. W. (1975). Olfactory, thermal,and tactual influences on infantile ultrasonic vocalizationin rats. Developmental Psychobiology, 8, 129–135.
Paton, J. F., & Richter, D. W. (1995). Maturational changes
in the respiratory rhythm generator of the mouse. PflugersArchives, 430, 115–124.
Roberts, L. H. (1975). The rodent ultrasound productionmechanism. Ultrasonics, 13, 83–88.
Sales, G. D. (1979). Strain differences in the ultrasonic be-havior of rats (Rattus norvegicus). American Zoologist,19, 513–527.
Sales, G., & Pye, D. (1974). Ultrasonic communication byanimals. New York: Wiley.
Sewell, G. D. (1970). Ultrasonic communication in rodents.Nature, 227, 410.
Smotherman, W. P., Bell, R. W., Starzec, I., Elias, J., &Zachman, T. A. (1974). Maternal responses to infant vo-calizations and olfactory cues in rats and mice. BehavioralBiology, 12, 55–66.
Stathopoulos, E. T., & Sapienza, C. M. (1997). Develop-mental changes in laryngeal and respiratory function withvariations in sound pressure level. Journal of Speech, Lan-guage, and Hearing Research, 40, 595–614.
Takahashi, L. K. (1992a). Developmental expression of de-fensive responses during exposure to conspecific adults inpreweanling rats (Rattus norvegicus). Journal of Compar-ative Psychology, 106, 69–77.
Takahashi, L. K. (1992b). Ontogeny of behavioral inhibitioninduced by unfamiliar adult male conspecifics in prew-eanling rats. Physiology & Behavior, 52, 493–498.
Takahashi, L. K. (1994). Stimulus control of behavioral in-hibition in the preweanling rat. Physiology & Behavior,55, 717–721.
Terkel, J., Damassa, D., & Sawer, C. (1979). Ultrasonic criesfrom infant rats stimulate prolactin release in lactatingmothers. Hormones and Behavior, 12, 95–102.
Voloschin, L., & Tramezzani, J. (1984). Relationship of pro-lactin release in lactating rats to milk ejection, sleep state,and ultrasonic vocalization by the pups. Endocrinology,114, 618–623.




















