
Population genetics of the Antarctic heteronemertean Parbolasia corrugatus from the South Orkney...
13
A. D. Rogers Æ A. Clarke Æ L. S. Peck Population genetics of the Antarctic heteronemertean Parbolasia corrugatus from the South Orkney Islands Received: 24 July 1997 / Accepted: 31 October 1997 Abstract An allozyme survey, using starch-gel electro- phoresis, was carried out on eight populations of the Antarctic nemertean worm Parborlasia corrugatus (McIntosh, 1876) collected from locations around the South Orkney Islands, Antarctica. These populations were separated by distances in the order of tens of kilometres. Genetic variation was estimated over 22 enzyme loci for all populations examined, giving an observed heterozygosity of 0.142. This was much lower than the expected heterozygosity (H e 0.201), and it was found that there was a significant deficiency of het- erozygotes across four enzyme loci ( p £ 0.01). A more detailed examination of this deficiency of heterozygotes was undertaken for the six populations and six variable enzyme loci for which the most complete data sets ex- isted. A significant deficiency of heterozygotes was found at the enzyme locus Odh-1 for four of the six populations examined ( p £ 0.01). Mean F is (0.240) in- dicated a significant ( p £ 0.01) within-population com- ponent of the heterozygote deficiency estimated for the six populations sampled, and this was mainly due to the Ap-1, Odh-1 and Pgm-1 loci. The mean F st value (0.036) was also significant ( p £ 0.01), indicating a de- gree of genetic dierentiation between populations. The observed levels of genetic dierentiation between popu- lations of P. corrugatus and the significant heterozygote deficiencies were unexpected, because this species has been reported to have a long-lived planktotrophic larva. It is hypothesised that recruitment of P. corrugatus in the South Orkney Islands originates from genetically distinct populations located in the Weddell Sea and to the west of the Antarctic Peninsula. Shifts in the relative position of the Weddell Sea Front, Weddell–Scotia Confluence and Scotia Front, relative to the South Orkney Islands, provide a mechanism for variation in the origin of recruits over time. Introduction Nemerteans are common animals in marine benthic habitats all over the world, from the littoral fringe (e.g. Roe 1993) to abyssal depths (e.g. Rogers et al. 1996), although they are often overlooked because of their cryptic habits, or ignored because of diculties in identifying them (Gibson 1994). Interestingly, nemerte- ans appear to have been reported with unusual fre- quency in southerly latitudes, although this may be a consequence of the presence of one especially conspic- uous species, Parborlasia corrugatus (McIntosh, 1876) (see Dearborn 1965). P. corrugatus has a circumpolar distribution in antarctic and subantarctic waters, and its vertical range extends from the intertidal to 3000 m depth (Gibson 1983). It is a large nemertean, reaching 2 m in length (Knox 1970) and over 140 g in wet mass (Heine et al. 1991). Like many species of nemerteans, it is a voracious predator and scavenger (Gibson 1994) and has been recorded as ingesting diatoms, sponges, anemones, polychaetes, amphipods, isopods, gastro- pods, fish, seal skin and blubber (Dearborn 1965). De- spite being conspicuous and common, there are no recorded observations of predation on P. corrugatus, and it has been reported as being distasteful to fish (Heine et al. 1991). Parborlasia corrugatus is a dioecious broadcast spawner that probably reproduces aseasonally (Shreeve and Peck 1995; Stanwell-Smith et al. 1997). The eggs Marine Biology (1998) 131: 1–13 Ó Springer-Verlag 1998 Communicated by J.P. Thorpe, Port Erin A.D. Rogers (&) 1 The Marine Biological Association of the United Kingdom, The Laboratory, Citadel Hill, Plymouth PL1 2PB, Devon, England A. Clarke Æ L.S. Peck British Antarctic Survey, High Cross, Madingley Road, Cambridge CB3 OET, England Present address: 1 The Division of Biodiversity and Ecology, School of Biological Sciences, University of Southampton, Biomedical Sciences Building, Bassett Crescent East, Southampton SO16 7PX, Hants, England
Transcript of Population genetics of the Antarctic heteronemertean Parbolasia corrugatus from the South Orkney...

A. D. Rogers á A. Clarke á L. S. Peck
Population genetics of the Antarctic heteronemerteanParbolasia corrugatus from the South Orkney Islands
Received: 24 July 1997 /Accepted: 31 October 1997
Abstract An allozyme survey, using starch-gel electro-phoresis, was carried out on eight populations of theAntarctic nemertean worm Parborlasia corrugatus(McIntosh, 1876) collected from locations around theSouth Orkney Islands, Antarctica. These populationswere separated by distances in the order of tens ofkilometres. Genetic variation was estimated over 22enzyme loci for all populations examined, giving anobserved heterozygosity of 0.142. This was much lowerthan the expected heterozygosity (He � 0.201), and itwas found that there was a signi®cant de®ciency of het-erozygotes across four enzyme loci ( p £ 0.01). A moredetailed examination of this de®ciency of heterozygoteswas undertaken for the six populations and six variableenzyme loci for which the most complete data sets ex-isted. A signi®cant de®ciency of heterozygotes wasfound at the enzyme locus Odh-1 for four of the sixpopulations examined ( p £ 0.01). Mean Fis (0.240) in-dicated a signi®cant ( p £ 0.01) within-population com-ponent of the heterozygote de®ciency estimated for thesix populations sampled, and this was mainly due tothe Ap-1, Odh-1 and Pgm-1 loci. The mean Fst value(0.036) was also signi®cant ( p £ 0.01), indicating a de-gree of genetic di�erentiation between populations. Theobserved levels of genetic di�erentiation between popu-lations of P. corrugatus and the signi®cant heterozygote
de®ciencies were unexpected, because this species hasbeen reported to have a long-lived planktotrophic larva.It is hypothesised that recruitment of P. corrugatus inthe South Orkney Islands originates from geneticallydistinct populations located in the Weddell Sea and tothe west of the Antarctic Peninsula. Shifts in the relativeposition of the Weddell Sea Front, Weddell±ScotiaCon¯uence and Scotia Front, relative to the SouthOrkney Islands, provide a mechanism for variation inthe origin of recruits over time.
Introduction
Nemerteans are common animals in marine benthichabitats all over the world, from the littoral fringe (e.g.Roe 1993) to abyssal depths (e.g. Rogers et al. 1996),although they are often overlooked because of theircryptic habits, or ignored because of di�culties inidentifying them (Gibson 1994). Interestingly, nemerte-ans appear to have been reported with unusual fre-quency in southerly latitudes, although this may be aconsequence of the presence of one especially conspic-uous species, Parborlasia corrugatus (McIntosh, 1876)(see Dearborn 1965). P. corrugatus has a circumpolardistribution in antarctic and subantarctic waters, and itsvertical range extends from the intertidal to 3000 mdepth (Gibson 1983). It is a large nemertean, reaching2 m in length (Knox 1970) and over 140 g in wet mass(Heine et al. 1991). Like many species of nemerteans, itis a voracious predator and scavenger (Gibson 1994) andhas been recorded as ingesting diatoms, sponges,anemones, polychaetes, amphipods, isopods, gastro-pods, ®sh, seal skin and blubber (Dearborn 1965). De-spite being conspicuous and common, there are norecorded observations of predation on P. corrugatus,and it has been reported as being distasteful to ®sh(Heine et al. 1991).
Parborlasia corrugatus is a dioecious broadcastspawner that probably reproduces aseasonally (Shreeveand Peck 1995; Stanwell-Smith et al. 1997). The eggs
Marine Biology (1998) 131: 1±13 Ó Springer-Verlag 1998
Communicated by J.P. Thorpe, Port Erin
A.D. Rogers (&)1
The Marine Biological Association of the United Kingdom,The Laboratory, Citadel Hill, Plymouth PL1 2PB,Devon, England
A. Clarke á L.S. PeckBritish Antarctic Survey, High Cross, Madingley Road,Cambridge CB3 OET, England
Present address:1 The Division of Biodiversity and Ecology,School of Biological Sciences, University of Southampton,Biomedical Sciences Building, Bassett Crescent East,Southampton SO16 7PX, Hants, England

develop into pilidia larvae which have been observed tosurvive for up to 50 to 56 d in the laboratory (Peck1993). However these larvae were only partially devel-oped, since they contained no developing juveniles in thepilidial envelope and showed no signs of settlement be-haviour. Sampling of natural populations of nemerteanlarvae in the Antarctic has suggested that the pilidiumstage may last for up to 150 d or more if growth ratesare similar to those measured in the laboratory (see Peck1993). Pilidia are present in the water column all yearround (Pearse et al. 1991; Stanwell-Smith et al. 1998),and it is likely that they not only feed on phytoplankton(present at very low concentrations during winter, seeClarke and Leakey 1996) but may be able to directlyassimilate nutrients or ingest small particulate materialsuch as bacteria (Peck 1993). After the pilidia meta-morphose, the juveniles are also pelagic for an unknownperiod of time and have been found deeper and furthero�shore than the pilidia (Shreeve and Peck 1995). Larvalduration in P. corrugatus is therefore extremely long andmigration between even geographically distant popula-tions may be possible. Adult P. corrugatus have alsobeen found between the fronds and in the holdfasts oflarge brown algae such as Himantothallus sp. (Gibson1983), so the possibility of passive dispersal by raftingalso exists (e.g. Edgar 1987).
The major current systems in the upper layers of thesouthern ocean are conducive to long-distance trans-port of larvae and planktonic organisms. Near thecontinent, the Antarctic Coastal Current moves waterwestwards but is broken up into a series of gyres.Further north, the Antarctic Circumpolar Current is amore or less continuous ¯ow of water in the oppositedirection. These circumpolar currents facilitate thetransport of holoplanktonic and meroplanktonic or-ganisms from region to region around the Antarcticcontinent and may allow a high level of gene ¯ow be-tween populations located in these regions (Fevoldenand Schneppenheim 1989). In Antarctic krill (Euphausiasuperba) for example, populations sampled from theAtlantic sector of the Southern Ocean, the Belling-shausen Sea, the Ross Sea and from the Indian Oceansector of the southern ocean have been shown to begenetically homogenous using allozyme data (seeFevolden and Schneppenheim 1989). For benthic or-ganisms which have a planktonic larval phase in theirlife history, genetic homogeneity would also be ex-pected to occur if larval duration was su�ciently longto enable migration between geographically distant lo-cations. However, it has been shown that the larvae ofcertain benthic organisms located in nearshore envi-ronments may generally be restricted to inshore waters(Shreeve and Peck 1995). If this is the case, then on aregional scale benthic organisms maybe subjected torestricted gene ¯ow by the formation of gyres or frontalsystems in the Antarctic Coastal Current.
There are few genetic studies of Antarctic organisms,and most of these have been of planktonic or pelagicanimals such as the amphipod Themisto gaudichaudii
(e.g. Schneppenheim and Weigmann-Haass 1986), krillof the genus Euphausia (e.g. KuÈ hl and Schneppenheim1986; Fevolden and Schneppenheim 1989), squid (e.g.Carvalho and Pitcher 1989; Brierley et al. 1993) and ®sh(e.g. Duhamel et al. 1995). In an allozyme survey of theAntarctic limpet Nacella concinna, di�erences in allelefrequency were detected between populations located inSouth Georgia and the South Orkney Islands (Beau-mont and Wei 1991). Previous work had shown thisspecies to be a broadcast spawner with a planktoniclarva (Picken 1980). However larval duration and type(lecithotrophic or planktotrophic) are uncertain (seePicken 1980). More recently, a study of the populationgenetics of the Antarctic octopus Pareledone turquetidetected striking levels of genetic di�erentiation betweenpopulations located around Shag Rocks and those lo-cated around South Georgia (a distance of <150 km)(Allcock et al. 1997). This species, which occurs between50 and 400 m, has a non-pelagic larva and it has beensuggested that such larvae are not capable of traversingthe deep-water channel between Shag Rocks and SouthGeorgia. There is, therefore, a geographic barrier todispersal between these populations (Allcock et al.1997). In this study, we present evidence for geneticdi�erentiation over a small geographic scale betweenpopulations of the nemertean worm Parborlasiacorrugatus located around Signy Island, South Orkneys.
Materials and methods
Sample collection
Specimens of Parborlasia corrugatus (McIntosh, 1876) were hand-collected from the South Orkney Islands by British AntarcticSurvey SCUBA-divers. Specimens were collected from a variety oflocations around Signy Island, from Lynch Island and OlivinePoint on Coronation Island (see Figs. 1 and 2). Signy Island andthe Coronation Islands are separated by the Normanna Strait,through which a strong current ¯ows. Collections took place overthe antarctic summer in the years 1992/1993 (Paal Harbour, Fac-tory Cove, Stygian Cove, Flensing Islands, Fyr Channel, LynchIsland, North Point, Olivine) and 1993/1994 (Fyr Channel, NorthPoint). Specimens from the 1992/1993 collections were frozen aswhole individuals together in bags. This led to problems duringspecimen preparation for electrophoresis, and in the followingseason (1993/1994) tissue samples were therefore taken from indi-vidual specimens in Antarctica and frozen in individual cryotubes.This method had the additional advantage that it was not de-structive to the worms, which were subsequently released. Allspecimens were frozen at )70 °C and transported back to the UKby ship.
Electrophoresis
A piece of tissue, '3 to 4 mm, cubed was excised from frozen tissuefor electrophoresis. Tissue was homogenised over ice in 100 ll of0.06 M Tris-HCl pH 8.0 using a glass rod. The homogenate wasabsorbed onto ®lter-paper wicks (Whatman No. 1) and was appliedto a starch gel. Horizontal starch-gel electrophoresis was performedby standard methods (see Harris and Hopkinson 1978; Ferguson1980; Richardson et al. 1986; Murphy et al. 1990) using 10 mm-thick 12.5% starch gels (Sigma Chemical Co. Ltd., Poole, Dorset,England) as standard; 6 mm thick 10% starch gels were used forsome enzyme loci that showed a slow rate of migration. Four bu�er
2

systems were used: Bu�er System I, Tris-citrate, continuous,pH 8.0 (Ward and Beardmore 1977); Bu�er System II, Tris-ver-sene-borate, continuous, pH 8.0 (Shaw and Prasad 1970; Bu�erNo. 3); Bu�er System III, Tris-citrate, boric-sodium hydroxide,discontinuous, pH 8.3 (Poulik 1957); Bu�er System IV, Tris-ci-trate, continuous, pH 7.1 (Williams et al. 1983). Bu�er Systems I,II and IV were run at 150 V, 40 mA for 7 to 8 h. Bu�er System IIIwas run at 130 V, 30 mA for 7 h.
Eighteen enzymes coding for a total of 22 loci were visualisedusing enzyme-speci®c stains modi®ed from Harris and Hopkinson(1978) and Manchenko (1994): aminopeptidase (AP substrateGLY-LEU (staining for Ap-1), AP substrate LEU-TYR (stainingfor Ap-1/2), AP substrate LEU-GLY-GLY (staining for Ap-3/4),(Sigma Chemical Co. Ltd., Poole, Dorset) E.C. 3.4.11 or 13 (Bu�er
System I,II,III); carbonic anhydrase (CA) E.C. 4.2.1.1 (Bu�erSystem III); non-speci®c esterase (EST) E.C. 3.1.1 (Bu�er SystemII); fumarate hydratase (FUM) E.C. 4.2.1.2 (Bu�er System I);glutamate oxaloacetate transaminase (GOT) E.C. 2.6.1.1 (Bu�erSystem II); glycerol-3-phosphate dehydrogenase (GPD) E.C.1.1.1.8 (Bu�er System I); hexokinase (HEX) E.C. 2.7.1.1 (Bu�erSystem I, III); isocitrate dehydrogenase (ICD) E.C. 1.1.1.42 (Buf-fer System II,III); lactate dehydrogenase (LDH) E.C. 1.1.1.27(Bu�er System IV); malate dehydrogenase (MDH) E.C. 1.1.1.37(Bu�er System I, II, III); mannose phosphate isomerase (MPI) E.C.5.3.1.8 (Bu�er System III); octopine dehydrogenase (ODH) E.C.1.5.1.11 (Bu�er System I, III) 6-phosphogluconate dehydrogenase(PGD) E.C. 1.1.4.4 (Bu�er System I); phosphoglucose isomerase(PGI) E.C. 5.3.1.9 (Bu�er System I); phosphoglucomutase (PGM)
Fig. 1 Map showing geograph-ic location of South OrkneyIslands with approximate posi-tions of local currents andfronts (major currents indicatedby thick arrow heads)
Fig. 2 Map showing six samplesites on Signy and CoronationIslands for which detailed ge-netic population analyses werecarried out. Redrawn fromThomson (1974)
3

E.C. 5.4.2.2 (Bu�er System I, III); superoxide dismutase (SOD)E.C. 1.15.1.1 (Bu�er System I, counterstain on ODH).
Data analysis
Mobility of the most common allele for each locus was designated1.000. Alleles were labelled in ascending order of migration rate,and mobilities were calculated relative to the most common allele.Allele frequencies were calculated for 22 loci for all specimenspooled together for all sites. In addition, allele frequencies werecalculated for six polymorphic loci which had stained most con-sistently in specimens for six localities (Factory Cove, FlensingIslands, Fyr Channel, Lynch Island, North Point, Olivine). Threespecimens representing two putative unidenti®ed species whichexternally resembled Parborlasia corrugatus were identi®ed duringelectrophoresis. In each case, ®ve enzyme loci were scored for thesespecimens and allele frequencies were calculated separately.
Genetic variability for all loci and specimens was estimatedusing a set of variables including the mean number of alleles perlocus, the percentage of polymorphic loci (i.e. loci with frequencyof commonest allele below 0.99), the observed heterozygosity (i.e.the proportion of sampled individuals heterozygous under the 0.99criterion), and the expected heterozygosity based on Hardy±Weinberg assumptions.
Polymorphic loci for pooled samples and for six populationstreated separately were tested for conformity to Hardy±Weinbergexpectations using Fisher's (1935) exact test.
Wright's (1951, 1965) ®xation index (F ) was calculated forvariable loci for pooled populations of Parborlasia corrugatus andfor the six populations taken separately. F values indicated the levelof heterozygote de®ciency (positive F value) or excess (negative Fvalue) in these populations.
Genetic di�erentiation among the six populations was esti-mated using FST for all variable loci. Signi®cance of departure ofFST from zero was calculated using the formula of Workman andNiswander (1970) given in Waples (1987):
v2 � 2NFST �k ÿ 1�; df � �k ÿ 1��sÿ 1� ; �1�where N � the number of individuals sampled, k � number ofalleles at a locus, and s � number of populations. The number ofmigrants per deme per generation (Nem) was calculated from FSTaccording to the formula:
Nem � f�1=FST � ÿ 1g=4 �Wright 1931; see Waples 1987� : �2�
Deviations from random mating expectations within the sixindividual populations was estimated using FIS. Signi®cance ofdeparture of FIS from zero was calculated using the chi-squared testof Li and Horvitz (1953) given in Waples (1987):
v2 � F 2ISN�k ÿ 1�; df � k�k ÿ 1�=2 ; �3�
where N � number of individuals sampled and k � number ofalleles at a locus.
Overall genetic di�erentiation between Parborlasia corrugatusand the two other taxa which were detected in the present studywere estimated using Nei's (1972) genetic identity (I ) and geneticdistance (D). These measures were used as a guide to the taxonomicrelationships of these species. Overall genetic di�erentiation wasalso calculated between each of the six populations of P. corrugatustreated separately, as above. Cluster analysis was also performedon these populations using the unweighted pair-group methodusing arithmetic averages (UPGMA).
All tests of signi®cance were adjusted using the sequentialBonferroni technique given in Rice (1989). All calculations werecarried out using BIOSYS Release 1.7 (Swo�ord and Selander1989).
Results
From a comparison of results between a pilot study runwith live tissue of Parborlasia corrugatus and resultsfrom frozen tissue (all subsequent electrophoresis runs),it was apparent that a reduction of enzyme activity oc-curred with freezing of tissue. This has been observed inother organisms for starch-gel electrophoresis (e.g.Scozzani et al. 1980). Partly because of this variableactivity, sample size varied for di�erent enzyme locisince enzyme activity was not always su�cient to enableaccurate scoring of genotypes. For scorable runs, en-zymes generally produced well-resolved staining patternsconsistent with known enzyme subunit-structure. Forthe locus Pgm-1, a second locus very close to the ®rstoccasionally showed partial staining, which led to threebanded staining patterns not consistent with a mono-meric enzyme.
Table 1 Parborlasia corrugatus. Estimates of allele frequency for all loci for all specimens from all sample sites combined (N numberallele = 1.00
Allele Ap-1 Ap-2 Ap-3 Ap-4 Ca-1 Ca-2 Est-1 Est-2 Fum-1 Fum-2 Got-1(N = 336) (N = 34) (N = 132) (N = 31) (N = 132) (N = 132) (N = 82) (N = 56) (N = 20) (N = 70) (N = 39)
A 0.006 1.000 1.000 0.129 1.000 0.977 0.012 0.080 1.000 1.000 0.256 1.000 0(0.90) (1.00) (1.00) (0.92) (1.00) (1.00) (0.91) (0.87) (1.00) (1.00) (0.48) (1.00) (
B 0.933 ± ± 0.806 ± 0.023 0.988 0.857 ± ± 0.654 ± 0(1.00) (1.00) (1.04) (1.00) (1.00) (1.00) (
C 0.058 ± ± 0.065 ± ± ± 0.063 ± ± 0.090 ± ±(1.16) (1.07) (1.06) (1.65)
D 0.001 ± ± ± ± ± ± ± ± ± ± ± ±(1.25)
E 0.001 ± ± ± ± ± ± ± ± ± ± ± ±(1.42)
F ± ± ± ± ± ± ± ± ± ± ± ± ±
G ± ± ± ± ± ± ± ± ± ± ± ± ±
H ± ± ± ± ± ± ± ± ± ± ± ± ±
4

Allele frequencies for 22 enzyme loci are given forpooled populations in Table 1. Of these loci, 14 arepolymorphic at the 0.99 level. Allele frequencies for thesix populations taken separately for the variable lociAp-1, Icd-1, Mdh-1, Odh-1, Pgi-1, Pgm-1 are broadlysimilar, but do show some di�erences, with rare allelesbeing unique to one or a few localities (Table 2). For thetwo unknown species designated SP1 and SP2, allelefrequencies are very di�erent from comparable loci forParborlasia corrugatus. SP2 does share some commonalleles with P. corrugatus (Tables 1, 2 and 3).
Nei's (1972) genetic identity (I � 0.955±0.873) be-tween separate samples of Parborlasia corrugatus is high,and conversely, genetic-distance values (D � 0.045 to0.127) are low (Table 4). Comparison of I values showthat they are consistently lower between the Lynch Is-land population (0.873 to 0.922) and other populationsthan between all other such comparisons (0.914 to0.955). UPGMA cluster-analysis (Fig. 3) also shows theLynch Island population to be an outlier to ®ve otherpopulations which are extremely similar (but note thatsimilarity does not fall below a clustering level of 0.981).
Genetic identity values between Parborlasiacorrugatus and SP1 (I � 0.000) and SP2 (I � 0.304)are very low (Table 4).
Observed heterozygosity (Ho � 0.142) for pooledpopulations of Parborlasia corrugatus was markedlylower than the expected heterozygosity (He � 0.201) ifgenotype frequencies of the pooled sample populationswere in Hardy±Weinberg equilibrium (Table 5). In allindividual populations for which six enzyme loci (Ap-1,Icd-1, Mdh-1, Odh-1, Pgi-1, Pgm-1) were analysed sep-arately there was a similar heterozygote de®ciency.
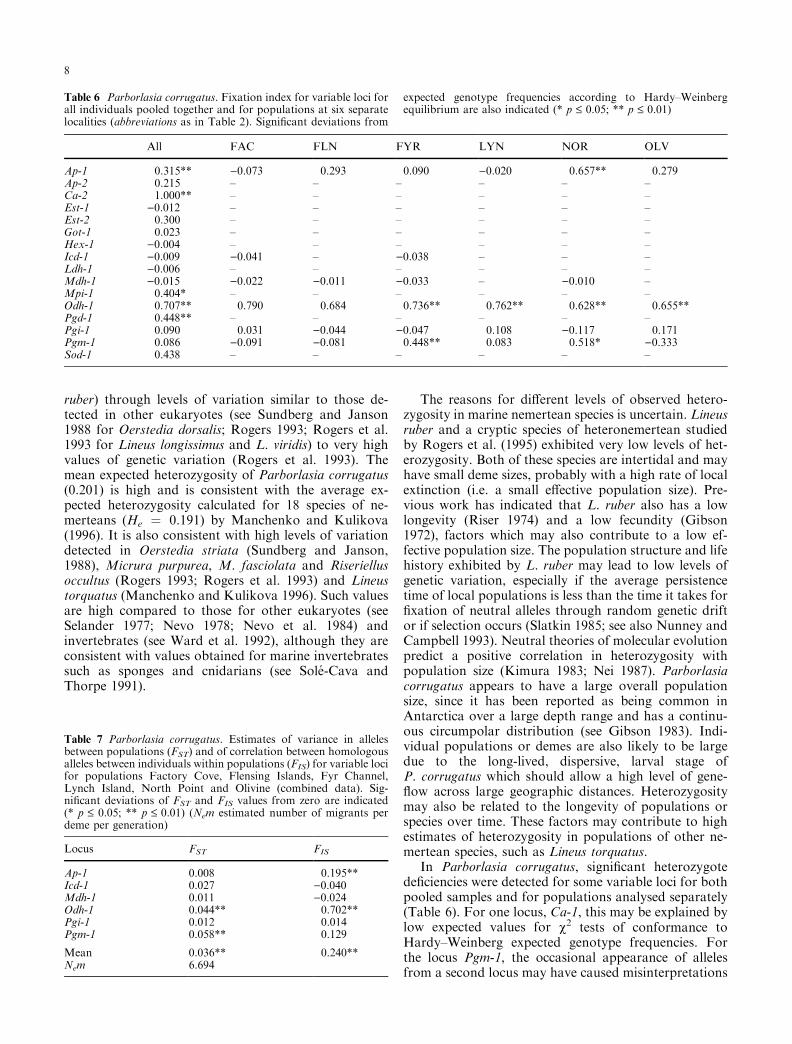
Wright's (1951, 1965) ®xation index (F ) indicates thatheterozygote de®ciencies are not uniform across all loci(Table 6). The loci Ap-1, Ca-2, Odh-1 and Pgd-1 showsigni®cant heterozygote de®ciencies ( p £ 0.01) forpooled samples, whilst Ap-1, Odh-1 and Pgm-1 show
signi®cant heterozygote de®ciencies for individual sam-ple sites (Table 6). In the case of Ca-2, single-locusheterozygosity is very low and signi®cant heterozygotede®ciencies are due to rare alleles occurring as homo-zygotes. This is not the case for other loci, which showsigni®cant heterozygote de®ciencies. In Odh-1, there aretwo common alleles, B and C which rarely occur asheterozygotes (Tables 1 and 2).
Heterozygote de®ciencies are further re¯ected by thewithin-population genetic structuring represented by FISvalues. FIS values are signi®cantly di�erent from zero( p £ 0.01) for two loci, Ap-1 and Odh-1 and for themean FIS value (Table 7). Between-population geneticstructuring represented by FST values also show signi®-cant di�erences from zero for Odh-1 and Pgm-1. Thisindicates that not only do individual populations ofParborlasia corrugatus show a heterozygote de®ciency inthese loci which is signi®cant in some cases, but thatallele frequencies for some loci are also signi®cantlydi�erent between populations (Table 7).
The estimated number of migrants per deme pergeneration (Nem) calculated from FST (Nem � 6.694) ismoderate compared to data for other marine inverte-brates. Nem for Parborlasia corrugatus is in excess of theestimated 1 individual per deme per generation neces-sary to prevent divergence between populations due torandom genetic drift (Wright 1940; Kimura 1955; Slat-kin 1985; Maynard Smith 1989).
Discussion
Analyses of existing data on levels of genetic di�erenti-ation compared to levels of taxonomic separation indi-cate that there is a low level of overlap in I values betweenconspeci®c populations and between species; 97% of Ivalues between congeneric species fall below 0.85 whilst98% of I values between conspeci®c populations exceed
of individuals sampled for given locus; ± allele absent). Relative mobility of alleles given in parentheses; mobility of most common
Gpd-1 Hex-1 Icd-1 Ldh-1 Mdh-1 Mpi-1 Odh-1 Pgd-1 Pgi-1 Pgm-1 Sod-1(N = 19) (N = 112) (N = 176) (N = 78) (N = 368) (N = 65) (N = 243) (N = 88) (N = 198) (N = 96) (N = 18)
0.256 1.000 0.996 0.000 0.006 0.985 0.115 0.021 0.153 0.013 0.016 0.889(0.48) (1.00) (1.00) (0.28) (0.83) (1.00) (0.72) (0.35) (0.69) (0.58) (0.60) (1.00)0.654 ± 0.004 0.000 0.994 0.000 0.154 0.667 0.744 0.169 0.052 0.111(1.00) (1.04) (0.46) (1.00) (1.04) (0.83) (1.00) (1.00) (0.74) (0.78) (1.85)0.090 ± ± 0.000 ± 0.000 0.315 0.245 0.102 0.000 0.552 ±(1.65) (0.69) (1.10) (1.00) (2.12) (2.26) (0.91) (1.00)± ± ± 0.003 ± 0.000 0.246 0.060 0.000 0.614 0.000 ±
(0.93) (1.19) (1.18) (3.52) (3.17) (1.00) (1.05)± ± ± 0.989 ± 0.015 0.169 0.008 ± 0.005 0.016 ±
(1.00) (1.20) (1.34) (4.08) (1.09) (1.14)± ± ± 0.009 ± ± ± ± ± 0.184 0.286 ±
(1.09) (1.28) (1.24)± ± ± ± ± ± ± ± ± 0.010 0.078 ±
(1.49) (1.38)± ± ± ± ± ± ± ± ± 0.005 0.000 ±
(1.71) (1.52)
5

Table 3 Unidenti®ed nemertean Species 1 (SP1) and Species 2 (SP2). Estimated allele frequencies for all detectable loci (N number ofspecimens sampled)
Locus Allele
A B C D E F G H
SP1 (N = 2)Icd-1 0.000 0.500 0.500 0.000 0.000 0.000 ± ±Mdh-1 0.000 0.000 0.000 1.000 0.000 ± ± ±Pgd-1 0.000 0.000 0.000 1.000 ± ± ± ±Pgi-1 0.000 0.000 1.000 0.000 0.000 0.000 0.000 0.000Pgm-1 0.000 0.000 0.000 1.000 0.000 0.000 0.000 0.000
SP2 (N = 1)Icd-1 1.000 0.000 0.000 0.000 0.000 0.000 ± ±Mdh-1 0.000 1.000 0.000 0.000 0.000 ± ± ±Odh-1 0.000 1.000 0.000 0.000 0.000 ± ± ±Pgi-1 0.000 0.000 0.000 0.000 0.000 0.000 0.500 0.500Pgm-1 0.000 0.000 0.000 0.000 0.000 0.000 0.500 0.500
Table 2 Parborlasia corrugatus. Estimates of allele frequency for the six variable loci with most complete data sets for Factory Cove(FAC ), Flensing Islands (FLN ), Fyr Channel (FYR), Lynch Island (LYN ), North Point (NOR) and Olivine (OLV )
Locus,allele
FAC FLN FYR LYN NOR OLV
Ap-1 (N = 22) (N = 45) (N = 99) (N = 25) (N = 83) (N = 40)A 0.000 0.000 0.010 0.000 0.012 0.000B 0.932 0.933 0.929 0.980 0.964 0.925C 0.068 0.056 0.056 0.020 0.024 0.075D 0.000 0.011 0.000 0.000 0.000 0.000E 0.000 0.000 0.005 0.000 0.000 0.000
Icd-1 (N = 19) (N = 45) (N = 27) (N = 25) (N = 20) (N = 30)A 0.000 0.000 0.000 0.000 0.000 0.000B 0.000 0.000 0.000 0.000 0.000 0.000C 0.000 0.000 0.000 0.000 0.000 0.000D 0.026 0.000 0.000 0.000 0.000 0.000E 0.947 1.000 0.963 1.000 1.000 1.000F 0.026 0.000 0.037 0.000 0.000 0.000
Mdh-1 (N = 23) (N = 45) (N = 109) (N = 25) (N = 101) (N = 41)A 0.978 0.989 0.968 1.000 0.990 1.000B 0.000 0.000 0.000 0.000 0.000 0.000C 0.000 0.000 0.000 0.000 0.000 0.000D 0.000 0.000 0.000 0.000 0.000 0.000E 0.022 0.011 0.032 0.000 0.010 0.000
Odh-1 (N = 11) (N = 17) (N = 95) (N = 25) (N = 58) (N = 37)A 0.000 0.029 0.000 0.000 0.017 0.095B 0.682 0.765 0.726 0.800 0.526 0.595C 0.318 0.206 0.153 0.140 0.405 0.297D 0.000 0.000 0.100 0.060 0.052 0.014E 0.000 0.000 0.021 0.000 0.000 0.000
Pgi-1 (N = 22) (N = 28) (N = 38) (N = 26) (N = 23) (N = 40)A 0.023 0.018 0.000 0.000 0.022 0.025B 0.091 0.179 0.197 0.231 0.217 0.112C 0.000 0.000 0.000 0.000 0.000 0.000D 0.705 0.589 0.618 0.538 0.587 0.663E 0.000 0.018 0.000 0.000 0.000 0.000F 0.159 0.196 0.158 0.231 0.152 0.188G 0.023 0.000 0.026 0.000 0.022 0.013H 0.000 0.000 0.000 0.000 0.000 0.000
Pgm-1 (N = 19) (N = 28) (N = 53) (N = 11) (N = 28) (N = 2)A 0.026 0.036 0.066 0.000 0.000 0.000B 0.053 0.054 0.085 0.045 0.054 0.000C 0.605 0.607 0.538 0.318 0.482 0.750D 0.000 0.000 0.000 0.000 0.000 0.000E 0.000 0.000 0.028 0.000 0.000 0.000F 0.316 0.250 0.208 0.318 0.411 0.250G 0.000 0.054 0.075 0.318 0.054 0.000H 0.000 0.000 0.000 0.000 0.000 0.000
6

0.85 (see Ayala 1975, 1983; Thorpe 1982, 1983; Thorpeand Sole -Cava 1994). In previous studies on nemerteans Ivalues between conspeci®c populations have also beenshown to be high, whilst genetic identities between dif-ferent species have been shown to be unusually low(Rogers et al. 1993, 1995). The high genetic identities be-tween populations of Parborlasia corrugatus (I � 0.873to 0.946) indicate that they are likely to be conspeci®c.
Genetic-identity values between Parborlasia co-rrugatus and SP1 (I � 0.00) and SP2 (I � 0.302) aresubject to a large degree of sampling error due to the lownumber of specimens of these species obtained and thelow number of loci over which genetic identity wascalculated (see Gorman and Renzi 1979). However, Ivalues obtained between these species are still a usefulguide to their systematic relationships in the absence ofany other data: 97% of I values between congenericspecies fall below 0.85 while 85% of I values betweencongeneric species are above 0.35 (see Ayala 1975, 1983;Thorpe 1982, 1983; Thorpe and Sole -Cava 1994). SP1shows no degree of genetic relationship to P. corrugatus,while SP2 may represent a congeneric species or a spe-cies from a closely related genus. Two other species ofParborlasia are found in Antarctica ± P. fueguina andP. landrumae (Gibson 1985). P. fueguina is found aroundthe Antarctic Peninsula and islands of the Scotia Arc(Gibson 1985), and SP2 may represent an example ofthis species. As SP1 and SP2 both super®cially resembleP. corrugatus, there is potential for confusion whenidentifying these nemerteans.
Nemerteans show a range of genetic variation fromextremely low expected and observed heterozygosities(see Williams et al. 1983; Rogers et al. 1995 for Lineus
Table 4 Parborlasia corrugatus. Pairwise comparisons of Nei's(1972) genetic identity (above diagonal ) and Nei's (1972) geneticdistance (below diagonal ). Data from ®ve loci for all populations vs
two other species (SP1, SP2) detected during electrophoresis andfor six loci for comparisons of populations at six localities (ab-breviations as in Table 2)
All FAC FLN FYR LYN NOR OLV SP1 SP2
All ± ± ± ± ± ± 0.000 0.304FAC ± 0.944 0.934 0.877 0.922 0.946 ± ±FLN ± 0.056 0.955 0.922 0.925 0.940 ± ±FYR ± 0.066 0.045 0.915 0.914 0.916 ± ±LYN ± 0.123 0.078 0.085 0.901 0.873 ± ±NOR ± 0.078 0.075 0.086 0.099 0.919 ± ±OLV ± 0.054 0.060 0.084 0.127 0.081 ± ±SP1 1 ± ± ± ± ± ± ±SP2 1.189 ± ± ± ± ± ± ±
Table 5 Parborlasia corrugatus.Genetic variability for all locifor all specimens and for the sixvariable loci with the mostcomplete data sets for six sepa-rate localities; standard errorsin parentheses (Ho mean ob-served heterozygosity; He meanexpected heterozygosity)
Population Mean samplesize/locus
Mean no.alleles/locus
% loci polymorphic(0.99 criteria)
Ho He
All 114.8 2.8 72.7 0.142 0.201(20.8) (0.4) (0.041) (0.053)
Factory Cove 19.3 3.0 100.00 0.235 0.293(1.8) (0.5) (0.091) (0.091)
Flensing Islands 34.7 3.2 83.3 0.241 0.283(4.9) (0.7) (0.117) (0.110)
Fyr Channel 70.2 3.7 100.0 0.219 0.321(14.3) (0.6) (0.084) (0.108)
Lynch Island 22.8 2.3 66.7 0.216 0.288(2.4) (0.5) (0.119) (0.133)
North Point 52.2 3.2 83.3 0.198 0.309(14.0) (0.6) (0.103) (0.125)
Olivine 31.7 2.5 66.7 0.202 0.286(6.2) (0.7) (0.088) (0.109)
Fig. 3 Parborlasia corrugatus. UPGMA dendrogram of Nei's (1978)genetic identity between six populations, calculated using six enzymeloci
7
ruber) through levels of variation similar to those de-tected in other eukaryotes (see Sundberg and Janson1988 for Oerstedia dorsalis; Rogers 1993; Rogers et al.1993 for Lineus longissimus and L. viridis) to very highvalues of genetic variation (Rogers et al. 1993). Themean expected heterozygosity of Parborlasia corrugatus(0.201) is high and is consistent with the average ex-pected heterozygosity calculated for 18 species of ne-merteans (He � 0.191) by Manchenko and Kulikova(1996). It is also consistent with high levels of variationdetected in Oerstedia striata (Sundberg and Janson,1988), Micrura purpurea, M. fasciolata and Riseriellusoccultus (Rogers 1993; Rogers et al. 1993) and Lineustorquatus (Manchenko and Kulikova 1996). Such valuesare high compared to those for other eukaryotes (seeSelander 1977; Nevo 1978; Nevo et al. 1984) andinvertebrates (see Ward et al. 1992), although they areconsistent with values obtained for marine invertebratessuch as sponges and cnidarians (see Sole -Cava andThorpe 1991).
The reasons for di�erent levels of observed hetero-zygosity in marine nemertean species is uncertain. Lineusruber and a cryptic species of heteronemertean studiedby Rogers et al. (1995) exhibited very low levels of het-erozygosity. Both of these species are intertidal and mayhave small deme sizes, probably with a high rate of localextinction (i.e. a small e�ective population size). Pre-vious work has indicated that L. ruber also has a lowlongevity (Riser 1974) and a low fecundity (Gibson1972), factors which may also contribute to a low ef-fective population size. The population structure and lifehistory exhibited by L. ruber may lead to low levels ofgenetic variation, especially if the average persistencetime of local populations is less than the time it takes for®xation of neutral alleles through random genetic driftor if selection occurs (Slatkin 1985; see also Nunney andCampbell 1993). Neutral theories of molecular evolutionpredict a positive correlation in heterozygosity withpopulation size (Kimura 1983; Nei 1987). Parborlasiacorrugatus appears to have a large overall populationsize, since it has been reported as being common inAntarctica over a large depth range and has a continu-ous circumpolar distribution (see Gibson 1983). Indi-vidual populations or demes are also likely to be largedue to the long-lived, dispersive, larval stage ofP. corrugatus which should allow a high level of gene-¯ow across large geographic distances. Heterozygositymay also be related to the longevity of populations orspecies over time. These factors may contribute to highestimates of heterozygosity in populations of other ne-mertean species, such as Lineus torquatus.
In Parborlasia corrugatus, signi®cant heterozygotede®ciencies were detected for some variable loci for bothpooled samples and for populations analysed separately(Table 6). For one locus, Ca-1, this may be explained bylow expected values for v2 tests of conformance toHardy±Weinberg expected genotype frequencies. Forthe locus Pgm-1, the occasional appearance of allelesfrom a second locus may have caused misinterpretations
Table 7 Parborlasia corrugatus. Estimates of variance in allelesbetween populations (FST) and of correlation between homologousalleles between individuals within populations (FIS) for variable locifor populations Factory Cove, Flensing Islands, Fyr Channel,Lynch Island, North Point and Olivine (combined data). Sig-ni®cant deviations of FST and FIS values from zero are indicated(* p £ 0.05; ** p £ 0.01) (Nem estimated number of migrants perdeme per generation)
Locus FST FIS
Ap-1 0.008 0.195**Icd-1 0.027 )0.040Mdh-1 0.011 )0.024Odh-1 0.044** 0.702**Pgi-1 0.012 0.014Pgm-1 0.058** 0.129
Mean 0.036** 0.240**Nem 6.694
Table 6 Parborlasia corrugatus. Fixation index for variable loci forall individuals pooled together and for populations at six separatelocalities (abbreviations as in Table 2). Signi®cant deviations from
expected genotype frequencies according to Hardy±Weinbergequilibrium are also indicated (* p £ 0.05; ** p £ 0.01)
All FAC FLN FYR LYN NOR OLV
Ap-1 0.315** )0.073 0.293 0.090 )0.020 0.657** 0.279Ap-2 0.215 ± ± ± ± ± ±Ca-2 1.000** ± ± ± ± ± ±Est-1 )0.012 ± ± ± ± ± ±Est-2 0.300 ± ± ± ± ± ±Got-1 0.023 ± ± ± ± ± ±Hex-1 )0.004 ± ± ± ± ± ±Icd-1 )0.009 )0.041 ± )0.038 ± ± ±Ldh-1 )0.006 ± ± ± ± ± ±Mdh-1 )0.015 )0.022 )0.011 )0.033 ± )0.010 ±Mpi-1 0.404* ± ± ± ± ± ±Odh-1 0.707** 0.790 0.684 0.736** 0.762** 0.628** 0.655**Pgd-1 0.448** ± ± ± ± ± ±Pgi-1 0.090 0.031 )0.044 )0.047 0.108 )0.117 0.171Pgm-1 0.086 )0.091 )0.081 0.448** 0.083 0.518* )0.333Sod-1 0.438 ± ± ± ± ± ±
8

of gels and may explain signi®cant heterozygote de®-ciencies which occurred for two populations. Ap-1 andOdh-1, however, are both monomeric enzymes for whichgood activity and resolution were observed and forwhich strong heterozygote de®ciencies were obtained. Atthe Odh-1 locus the de®ciency was observed for almostevery population. In Odh-1 heterozygote de®ciency oc-curred because of a lack of individuals heterozygous forthe two most common alleles rather than because of theoccurrence of rare alleles as homozygotes, as in Ap-1 inthe present study and in other studies of allozyme vari-ation in marine invertebrates (see Creasey et al. 1996).
Heterozygote de®ciency is a common phenomenon inallozyme studies of genetic variation in marine inverte-brates (see Zouros and Foltz 1984; Singh and Green1985; Volckaert and Zouros 1989; Borsa et al. 1991),including nemerteans (see Rogers et al. 1995). Expla-nations for heterozygote de®ciency fall into four cate-gories (i) the presence of null alleles or the loss ofchromosomes causing incorrect estimates of genotypefrequency (Zouros and Mallet 1989); (ii) as a result ofinbreeding; (iii) as a result of sampling from two or moregenetically distinct populations (Wahlund e�ect); (iv)selection.
Null alleles occur very rarely in studies of allozymevariation in marine invertebrates, and it is consideredunlikely that such a phenomena could occur over severalloci in a single study on one species. Previous work hasfound the frequency of individuals in which null allelesare present to be <1 in 1000 at a dimeric PGI locus forthe deep-sea crustaceans Rimicaris exoculata and Mu-nidopsis scobina (Creasey and Rogers unpublished data).There may be a possibility that strong heterozygote de-®ciencies in Odh-1 were due to two alleles with a verysimilar mobility (i.e. a failure to resolve two alleles),although there was no evidence of this. Chromosomeloss, as with the occurrence of null alleles, is rare.
Parborlasia corrugatus is a dioecious, free-spawningspecies, with a planktotrophic larva which may be highlydispersive (see Peck 1993; Shreeve and Peck 1995). In-breeding seems to be an unlikely cause of heterozygotede®ciency in such a case. It is possible that the formationof trapped eddies in the vicinity of Signy Island mayprovide a mechanism for larval retention (see Boehlertand Mundy 1993 for discussion of this phenomenon).However, the possibility that such features would besu�ciently permanent to cause inbreeding in P. co-rrugatus populations are unlikely, especially in the faceof immigration from the eastward current-¯ow pastSigny Island that originates from the Drake Passage andthe Weddell Sea (see below). Furthermore, observationsalso indicate that the larvae of P. corrugatus are presentin the water around Signy Island all year round irre-spective of local oceanographic conditions (Stanwell-Smith personal communication).
The sampling of two or more genetically distinctpopulations is a likely explanation for the strong he-terozygote de®ciencies detected in the present study. Thequestion arises, however, as to whether the genetically
distinct populations sampled in the present study areconspeci®c. Cryptic species have been detected in othernemertean taxa (see Sundberg and Janson 1988; Rogerset al. 1993, 1995; Manchenko and Kulikova 1996). Ge-netic distances between cryptic nemertean species havebeen higher than mean genetic distance values detectedbetween congeneric species in other organisms, and ithas been suggested that nemerteans exhibit morpho-logical stasis (Rogers et al. 1995; Manchenko andKulikova 1996). Because of the high genetic identitybetween populations of Parborlasia corrugatus, thepossibility that these populations represent two or moreclosely related species is unlikely. However, in view ofthe high genetic identities that have been found betweenconfamilial species of Antarctic ®sh (Anderson 1982;McDonald et al. 1992) the possibility that the presentstudy sampled two closely related cryptic species must beconsidered. This would have considerable rami®cationsfor previous and future studies on the ecology andphysiology of this important Antarctic species.
Signy Island is located within the extremely complexfrontal system that exists in the Drake Passage and theScotia Sea region (Muench 1990; see present Fig. 1). Inthis region strong eastwardly ¯owing currents from theDrake Passage converge with northward ¯owing cur-rents from the Weddell Sea Gyre. Water from the Drakepassage originates from three geographic regions, theAntarctic Circumpolar Current, the Bellingshausen Seaand the Brans®eld Strait, and it is di�erent in terms ofphysical characteristics, such as temperature and sali-nity, from Weddell Sea water (Foster and Middleton1984). In addition to the Polar Front, the waters of theScotia and Weddell Seas are separated by the Weddell±Scotia Con¯uence, a region of relatively homogenouscold water. The boundary of this con¯uence is de®nedby the Scotia Front to the north and the Weddell Frontto the south (see Gordon et al. 1977; Muench 1990).Current ¯ow in this region is eastwards and is intensi®edwithin the fronts themselves (Nowlin and Cli�ord 1982).The mesoscale oceanography within and around theScotia and Weddell Sea Fronts is extremely complex.Records of the duration of sea-ice around Signy Islandindicate interannual variability on a seven-year cycle,which is probably related to variability in large-scalecircumpolar oceanographic and atmospheric processesincluding the El NinÄ o Southern Oscillation (ENSO)(Murphy et al. 1995). Interannual variation in phyto-plankton bloom size and in the abundance of krill andother species including seabirds and mammals in theregion around the South Orkney Islands and SouthGeorgia have also been related to these large-scaleoceanographic processes (Croxall et al. 1988; Clarke andLeakey 1996; Fedulov et al. 1996; Whitehouse et al.1996). In the case of krill, variation in the origin of re-cruits (i.e. from the Brans®eld Strait/Antarctic Peninsularegion or from the Weddell Sea) has been suggested as acause of strong interannual variations in abundancearound South Georgia (Heywood et al. 1985; Brierleyet al. 1997).
9

It is conceivable that a signi®cant proportion of re-cruitment to the Signy Island population of Parborlasiacorrugatus does not occur locally. The larvae are su�-ciently long-lived that unless behavioural mechanismsprevent dispersal, which data from plankton surveystends to suggest is not the case (Shreeve and Peck 1995),the majority of larvae produced around Signy Island arelikely to drift away from it in the prevailing eastwardcurrent ¯ow. Signy Island populations of P. corrugatusmay be dependent populations which rely on a supply ofrecruits from currents impinging on the South OrkneyIslands from the Drake Passage and Weddell Sea. De-pendent populations have been observed on isolatedtopographic features in the past. For example, a popu-lation of the rock lobster Jasus tristani on the VemaSeamount, South Atlantic Ocean, has been shown toprobably recruit, on an episodic basis, from larvaeoriginating from the distant island of Tristan Da Cunha(Heydorn 1969; Lutjeharms and Heydorn 1981a,b; seealso Boehlert et al. 1994).
The Fine-Resolution Antarctic Model (FRAM) sug-gests that particles would take in the order of 100 to150 d to reach the South Orkney Islands from thenorthern Antarctic Peninsula (Stevens and Thompson1994). This period of time is within the expected lengthof the pelagic larval/juvenile phase of Parborlasiacorrugatus, so recruitment of larvae to the South OrkneyIslands from the sea around the Antarctic Peninsula ispossible. Fluctuations in the relative positions of thefrontal systems across Signy Island, because of meso-scale and larger-scale oceanographic and atmosphericvariation (see Foster and Middleton 1984; Muench1990; Stein and Heywood 1994), provide a mechanismfor recruitment of larvae from two alternative sources-populations in the Weddell Sea and populations fromthe Bellingshausen Sea/Brans®eld Strait/Drake Passageregions. The similarities between this hypothetical modelof recruitment for P. corrugatus in the South Orkneysand that proposed for krill from South Georgia arestriking.
Genetic di�erences in source populations for recruitsto the South Orkney Island Parborlasia corrugatuspopulations would explain the heterozygote de®cienciesfound at several loci in the present study. Di�erentproportions of adult worms recruited from these popu-lations would also explain di�erences in allele frequen-cies between sample localities around Signy Island whichled to some signi®cant FST values for variable loci. It isnotable that the Lynch Island population is a geneticoutlier to other sample sites for P. corrugatus (seeFig. 3). This suggests genetic isolation by geographicdistance, as the Lynch Island population is removedfrom most other populations, although such an expla-nation would imply that the population located atOlivine should also show a similar level of genetic dif-ferentiation to those on Signy Island. These geneticdi�erences may be due to the hydrographic situation ofthe Lynch Island population, which lies in a bay which isadjacent to the Normanna Strait (see Fig. 2). A strong
eastwardly ¯owing current through this strait (BritishAntarctic Survey unpublished data) may result in trap-ping of water within this bay, although there are nodirect observations at present to support this. An alter-native explanation is the low sample size for the LynchIsland population leading to a high sampling error forestimates of genotype frequencies.
Recruitment from or mixing of genetically distinctpopulations has been suggested for other species ofmarine animals. In the New Zealand ling Genypterusblacoides for example, signi®cant heterozygote de®cien-cies have been detected for a population sampled from ageographic area associated with a convergence zonebetween two di�erent water masses (Smith 1979). Eachof these water masses was occupied by a geneticallydistinct stock which were separated by a hydrographicbarrier (the Subtropical Convergence). In a populationof the Atlantic haddock Melanogrammus aegle®nus lo-cated in the Georges Bank, North Atlantic Ocean, sig-ni®cant heterogeneity in the frequency of mitochondrialDNA haplotypes between samples taken in di�erentyears has been detected (Purcell et al. 1996). The ex-planation for these data was that recruitment to theGeorges Bank population occurs episodically from anumber of other geographic regions (i.e. from a numberof genetically distinct stocks). The mixing of geneticallydistinct populations has also been put forward as anexplanation for signi®cant heterozygote de®ciencies de-tected in the deep-sea spider crab Encephaloides arm-strongi, which is found in the northern Indian Ocean(Creasey et al. 1997). In this case, the mixing of geneti-cally distinct populations may be related to breedingmigrations undertaken by these crabs when spawning.
The maintenance of genetic di�erences between pu-tative source populations of Parborlasia corrugatus re-cruiting to Signy Island is an interesting question whichcannot be answered by the present study. Genetic dif-ferences between source populations could have arisenand been maintained by selection for di�erent alleleswhich are favoured by the prevailing physical conditionsin the Weddell Sea and Bellingshausen Sea regions.Octopine dehydrogenase, the enzyme in which the moststriking heterozygote de®ciencies occur, may be involvedin vigorous muscular activity and anaerobiosis (seeDando et al. 1981). There is evidence for selection inother enzymes which are important in metabolic pro-cesses for other marine invertebrates (e.g. Jollivet et al.1995). The occurrence of signi®cant heterozygote de®-ciencies in some, but not all, variable enzyme loci fa-vours selection as a cause of heterozygote de®ciencies inthe present study, but this may be an artefact of single-locus sample size and overall sample size for severalpopulations. Alternatively, di�erences may have arisenby genetic drift of populations between which there is apresent or historical barrier to gene-¯ow, by e�ectivegeographic distance, or by the hydrography of this re-gion. This becomes more likely if the length of thepelagic larval/juvenile phase in P. corrugatus is actuallyshorter than has been predicted from laboratory culture
10

experiments (Peck 1993) and observations of plankton(Shreeve and Peck 1995), or if behavioural mechanismsexist that restrict the dispersal of larvae. There areseveral cases where the dispersal potential of larvae asestimated from laboratory observations have not beenrealised in naturally occurring populations (e.g. Toddet al. 1988). If this is also the case for P. corrugatus, thena lack of gene-¯ow between the populations sampled inthe present study also becomes a plausible explanationfor the genetic di�erences between them ± but note thatin such a case larval dispersal would have to be highlyrestricted.
The open nature of the marine environment is notoften considered in studies of population genetics, es-pecially where populations are structured. The commonoccurrence of heterozygote de®ciencies in populationsmay be related to occasional recruitment from geo-graphically isolated and genetically di�erent populationsinto study populations (e.g. Smith 1979; Creasey et al.1996). Recent evidence indicates that temporal hetero-geneity of genotypes of recruits over several cohorts mayalso be an indicator of recruitment from two or moregenetically distinct populations (e.g. Purcell et al. 1996).The role of such occasional recruitment from geo-graphically distant and genetically distinct populationson gene ¯ow, genetic population structure and ulti-mately on evolutionary processes in marine populationsis not generally appreciated at the present time. In asimilar way, the loss of planktotrophic larvae frommarine populations subject to prevailing unidirectionalcurrent systems, and their subsequent reliance on re-cruitment from other geographically removed popula-tions, which may be sporadic, has also generally beenoverlooked.
Acknowledgements Support by the Natural Environment ResearchCouncil (Grant No. GR3/7168 to Professor R. Gibson, LiverpoolJohn Moores University, and Dr J. Thorpe, The University ofLiverpool) and by the Marine Biological Association of the U.K. isacknowledged by the authors. Special thanks are due to BritishAntarctic Survey personnel who collected material for the presentstudy. Thanks are also due to S. Creasey, N. Bryars and S. Laidler,who assisted in electrophoresis and to Dr M. Whit®eld for the useof laboratory facilities at the Marine Biological Association,Plymouth.
References
Allcock AL, Brierley AS, Thorpe JP, Rodhouse PG (1997) Re-stricted gene ¯ow and evolutionary divergence between geo-graphically separated populations of the Antarctic octopusPareledone turqueti. Mar Biol 129: 97±102
Anderson RC (1982) Electrophoretic analysis of Antarctic ®shfrom South Georgia. Anim Blood Grps biochem Genet 13:11±18
Ayala FJ (1975) Genetic di�erentiation during the speciation pro-cess. Evolutionary Biol 8: 1±78
Ayala FJ (1983) Enzymes as taxonomic characters. In: Oxford GS,Rollinson D (eds) Protein polymorphism: adaptive and taxo-nomic signi®cance. Academic Press, London, pp 3±26
Beaumont AR, Wei HC (1991) Morphological and genetic varia-tion in the Antarctic limpet Nacella concinna (Strebel, 1908).J mollusc Stud 57: 443±450
Boehlert GW, Mundy BC (1993) Ichthyoplankton assemblages atseamounts and oceanic islands. Bull mar Sci 53: 336±361
Boehlert GW, Wilson CD, Mizuno K (1994) Populations of thesternoptychid ®sh Maurolicus muelleri on seamounts in theCentral North Paci®c. Pacif Sci 48: 57±69
Borsa P, Zainuri M, Delay B (1991) Heterozygote de®ciency andpopulation structure in the bivalve Ruditapes decussatus. He-redity, Lond 66: 1±8
Brierley AS, Rodhouse PG, Thorpe JP, Clarke MR (1993) Geneticevidence of population heterogeneity and cryptic speciation inthe ommastrephid squidMartialia hyadesi from the PatagonianShelf and Antarctic Polar Frontal Zone. Mar Biol 116: 593±602
Brierley AS, Watkins JL, Murray AWA (1997) Interannual vari-ability in krill abundance at South Georgia. Mar Ecol Prog Ser150: 87±98
Carvalho GR, Pitcher TJ (1989) Biochemical genetic studies on thePatagonian squid Loligo gahi d'Orbigny. II. Population struc-ture in Falkland waters using isozymes, morphometrics and lifehistory data. J exp mar Biol Ecol 126: 243±258
Clarke A, Leakey RJG (1996) The seasonal cycle of phytoplank-ton, macronutrients and the microbial community in anearshore Antarctic marine ecosystem. Limnol Oceanogr 41:1281±1294
Creasey S, Rogers AD, Tyler PA (1996) Genetic comparison of twopopulations of the deep-sea vent shrimp Rimicaris exoculata(Decapoda: Bresiliidae) from the Mid-Atlantic Ridge. Mar Biol125: 473±482
Creasey S, Rogers AD, Tyler PA, Gage JD (1997) The populationbiology and genetics of the deep-sea spider crab, Encephaloidesarmstrongi Wood-Mason 1891 (Decapoda: Majidae). PhilTrans R Soc (Ser B) 352: 365±379
Croxall JP, McCann TS, Prince PA, Rothery P (1988) Reproduc-tive perfomance of seabirds and seals at South Georgia andSigny Island, South Orkney Island, 1976±1987: implications forSouthern Ocean monitoring studies. In: Sahrage D (ed) Ant-arctic ocean and resources variability. Springer-Verlag, Berlin,pp 261±285
Dando PR, Storey KB, Hochachka PW, Storey JM (1981) Multipledehydrogenases in marine molluscs: electrophoretic analysis ofalanopine dehydrogenase, strombine dehydrogenase, octopinedehydrogenase and lactate dehydrogenase. Mar Biol Lett 2:249±257
Dearborn JH (1965) Ecological and faunistic investigations of themarine benthos at McMurdo Sound, Antarctica. Ph.D. thesis.Stanford University, Stanford, California
Duhamel G, Ozouf-Costaz C, Cattaneo-Berrebi G, Berrebi P(1995) Interpopulation relationships of Antarctic ®sh Not-othenia rossii and Champsocephalus gunneri from the KerguelenIslands: an allozyme study. Antarct Sci 7: 351±356
Edgar GJ (1987) Dispersal of faunal and ¯oral propagules associ-ated with drifting Macrocystis pyrifera plants. Mar Biol 95:599±610
Fedulov PP, Murphy EJ, Shulgovsky KE (1996) Environment±krillrelations in the South Georgia marine ecosystem. Sel scient Papscient Comm Conserv antarct mar living Resour 1996(3): 13±30
Ferguson A (1980) Biochemical systematics and evolution. Blackie& Son, Glasgow
Fevolden SE, Schneppenheim R (1989) Genetic homogeneity ofkrill (Euphausia superba Dana) in the Southern Ocean. PolarBiol 9: 533±539
Fisher RA (1935) The logic of inductive inference. Jl R statist Soc98: 39±54
Foster TD, Middleton JH (1984) The oceanographic structure ofthe eastern Scotia Sea. I. Physical oceanography. Deep-Sea Res31: 529±550
Gibson R (1972) Nemerteans. Hutchinson University Library.Hutchinson & Co. (Publishers) Ltd., London
Gibson R (1983) Antarctic nemerteans: the anatomy, distribution,and biology of Parborlasia corrugatus (McIntosh, 1876)(Heteronemertea, Lineidae). Biology of the Antarctic seas. XIV.Antarctic Res Ser 39: 289±316
11

Gibson R (1985) Antarctic nemerteans: Heteronemertea ± de-scriptions of new taxa, reappraisals of the systematic status ofexisting species and a key to the heteronemerteans recordedsouth of Latitude 50°S. Zool J Linn Soc 83: 95±277
Gibson R (1994) Nemerteans. Synopses Br Fauna (New Ser) 24:1±224
Gordon AL, Georgi DT, Taylor HW (1977) Antarctic Polar Frontzone in the western Scotia Sea ± summer 1975. J phys Oceanogr89: 641±648
Gorman GC, Renzi J (1979) Genetic distance and heterozygosityestimates in electrophoretic studies: e�ects of sample size.Copeia 2: 242±249
Harris H, Hopkinson DA (1978; and subsequent supplements)Handbook of enzyme electrophoresis in human genetics. NorthHolland Publishing Co., Amsterdam
Heine JN, McClintock JB, Slattery M, Weston J (1991) Energeticcomposition, biomass, and chemical defense in the commonAntarctic nemertean Parborlasia corrugatus McIntosh. J expmar Biol Ecol 153: 15±25
Heydorn SE (1969) The South Atlantic rock lobster Jasus tristaniat Vema Seamount, Gough Island and Tristan Da Cunha.Investl Rep Div Sea Fish Repub S Afr 73: 1±20
Heywood RB, Everson I, Priddle J (1985) The absence of krillfrom the South Georgia zone, winter 1983. Deep-Sea Res 32:369±378
Jollivet D, DesbruyeÁ res D, Ladrat C, Laubier L (1995) Evidencefor di�erences in the allozyme thermostability of deep-sea hy-drothermal vent polychaetes Alvinellidae: a possible selectionby habitat. Mar Ecol Prog Ser 123: 125±136
Kimura M (1955) Solution of a process of random genetic driftwith a continuous model. Proc natn Acad Sci USA 41: 144±150
Kimura M (1983) The neutral theory of molecular evolution.Cambridge University Press, Cambridge
Knox GA (1970) Antarctic marine ecosystems. In: Holgate MW(ed.) Antarctic ecology. Vol. 1. Academic Press, New York,pp 69±96
KuÈ hl S, Schneppenheim R (1986) Electrophoretic investigation ofgenetic variation in two krill species Euphausia superba andE. crystallorophias (Euphausiidae). Polar Biol 6: 17±23
Li CC, Horvitz DG (1953) Some methods of estimating the in-breeding coe�cient. Am J hum Genet 5: 107±117
Lutjeharms JRE, Heydorn AEF (1981a) The rock lobster Jasustristani on Vema Seamount: drifting buoys suggest a possiblerecruiting mechanism. Deep-Sea Res 28A: 631±636
Lutjeharms JRE, Heydorn AEF (1981b) Recruitment of rocklobster on Vema Seamount from the islands of Tristan DaCunha. Deep-Sea Res 28A: p. 1237
Manchenko GP (1994) A handbook of detection of enzymes onelectrophoretic gels. CRC Press, London
Manchenko GP, Kulikova VI (1996) Allozyme and colour di�er-ences between two sibling species of the heteronemertean Lineustorquatus from the Sea of Japan. Mar Biol 125: 687±691
Maynard Smith J (1989) Evolutionary genetics. Oxford UniversityPress, Oxford
McDonald MA, Smith MH, Smith MW, Novak JM, Johns PE,Devries AL (1992) Biochemical systematics of notothenioid®shes from Antarctica. Biochem Syst Ecol 20: 233±241
McIntosh WC (1876) Descriptions of some new species of Annelidafrom Kerguelen's Island. Ann Mag nat Hist (Ser 4) 17: 318±323
Muench RD (1990) Mesoscale phenomena in the polar oceans. In:Smith WO (ed) Polar oceanography. Part A. Physical science.Academic Press Ltd., London, pp 223±285
Murphy EJ, Clarke A, Symon C, Priddle J (1995) Temporalvariation in Antarctic sea-ice: analysis of a long term fast-icerecord from the South Orkney Islands. Deep-Sea Res 42: 1045±1062
Murphy RW, Sites JW Jr, Buth DG, Hau¯er CH (1990) Proteins. I.Isozyme electrophoresis. In: Hillis D, Moritz C (eds) Molecularsystematics, Sinauer Associates Inc., Sunderland, Mass, pp 45±126
Nei M (1972) Genetic distance between populations. Am Nat 106:283±292
Nei M (1978) Estimation of average heterozygosity and geneticdistance from a small number of individuals. Genetics, Austin,Tex 89: 583±590
Nei M (1987) Molecular evolutionary genetics. Columbia Univer-sity Press, New York
Nevo E (1978) Genetic variation in natural populations: patternsand theory. Theor Popul Biol 13: 121±177
Nevo E, Beiles A, Ben-Schlomo R (1984) The evolutionarysigni®cance of genetic diversity: ecological, demographic andlife-history correlates. In: Mani GS (ed) Evolutionary dy-namics of genetic diversity. Springer-Verlag, Heidelberg, pp13±212
Nowlin WD, Cli�ord M (1982) The kinematic and thermohalinezonation of the Antarctic Circumpolar Current at Drake Pas-sage. J mar Res 40: 481±507
Nunney L, Campbell KA (1993) Assessing minimum viable pop-ulation size: demography meets population genetics. TrendsEcol Evolut 8: 234±239
Pearse JS, McClintock JB, Bosch I (1991) Reproduction of Ant-arctic benthic marine invertebrates: tempos, modes and timing.Am Zool 31: 65±80
Peck LS (1993) Larval development of the Antarctic nemerteanParborlasia corrugatus (Heteronemertea: Lineidae). Mar Biol116: 301±310
Picken GB (1980) The distribution, growth, and reproduction ofthe antarctic limpet Nacella (Patinigera) concinna (Strebel,1908). J exp mar Biol Ecol 42: 71±85
Poulik MD (1957) Starch gel electrophoresis in a discontinuoussystem of bu�ers. Nature, Lond 180: 1477±1479
Purcell MK, Korn®eld I, Fogarty M, Parker A (1996) Interdecadalheterogeneity in mitochondrial DNA of Atlantic haddock(Melanogrammus aegle®nus) from Georges Bank. Molec marBiol Biotechnol 5: 185±192
Rice WR (1989) Analyzing tables of statistical tests. Evolution 43:223±225
Richardson BJ, Baverstock PM, Adams M (1986) Allozyme elec-trophoresis ± a handbook for animal systematics and popula-tion studies. Academic Press, Sydney
Riser NW (1974) Nemertinea. In: Giese AC, Pearse JS (eds) Re-production of marine invertebrates. Vol. 1. Acoelomate andpseudocoelomate metazoans. Academic Press, New York andLondon, pp 359±389
Roe P (1993) Aspects of the biology of Pantinonemertescaliforniensis, a high intertidal nemertean. Hydrobiologia 266:29±44
Rogers AD (1993) Aspects of the genetics and taxonomy of marinenemerteans. Ph.D. thesis. University of Liverpool, Liverpool
Rogers AD, Gibson R, Tunnicli�e V (1996) A new genus andspecies of monostiliferous hoplonemertean colonizing an in-choate hydrothermal ®eld on Juan de Fuca Ridge. Deep-SeaRes 43: 1581±1599
Rogers AD, Junoy J, Gibson R, Thorpe JP (1993) Enzyme elec-trophoresis, genetic identity and description of a new genusand species of heteronemertean (Nemertea, Anopla) fromnorthwestern Spain and North Wales. Hydrobiologia 266:219±238
Rogers AD, Thorpe JP, Gibson R (1995) Genetic evidence for theoccurrence of a cryptic species with the littoral nemerteansLineus ruber and L. viridis (Nemertea: Anopla). Mar Biol 122:305±316
Schneppenheim R, Weigmann-Haass R (1986) Morphological andelectrophoretic studies on the genus Themisto (Amphipoda,Hyperiidae) from the South and North Atlantic. Polar Biol 6:215±225
Scozzani R, Ulizzi A, Ferracin A, Lunadei M (1980) An electro-phoretic polymorphism in Triturus cristatus carnifex (Am-phibia, Urodela). Experientia 36: p. 645
Selander RK (1977) Genic variation in natural populations. In:Ayala FJ (ed) Molecular evolution. Sinauer Associates Incor-porated, Sunderland, Mass, pp 21±45
Shaw CR, Prasad R (1970) Starch gel electrophoresis ± a compi-lation of recipes. Biochem Genet 4: 297±320
12

Shreeve RS, Peck LS (1995) Distribution of pelagic larvae ofbenthic marine invertebrates in the Bellingshausen Sea. PolarBiol 15: 369±374
Singh SM, Green RH (1985) Excess of allozyme homozygosityin marine molluscs and its possible biological signi®cance.Malacalogia 25: 569±581
Slatkin M (1985) Gene ¯ow in natural populations. A Rev EcolSyst 16: 393±430
Smith PJ (1979) Glucosephosphate isomerase and phosphogluco-mutase polymorphisms in the New Zealand ling Genypterusblacodes. Comp Biochem Physiol 62B: 573±577
Sole -Cava AM, Thorpe JP (1991) High levels of genetic variation innatural populations of marine lower invertebrates. Biol J LinnSoc 44: 65±80
Stanwell-Smith D, Hood A, Peck L (1997) A ®eld guide to thepelagic invertebrate larvae of the maritime Antarctic. BritishAntarctic Survey, Cambridge, England
Stanwell-Smith D, Peck LS, Clarke A, Murray A, Todd CD (1998)The distribution, abundance and seasonality of pelagic marineinvertebrate larvae in the maritime Antarctic. Phil Trans R Soc(Ser B) (In press)
Stein M, Heywood RB (1994) Antarctic environment ± physicaloceanography: the Antarctic Peninsula and Southwest Atlanticregion of the Southern Ocean. In: El-Sayed SZ (ed) Southernocean ecology: the BIOMASS perspective, Cambridge Univer-sity Press, Cambridge, pp 11±24
Stevens DP, Thompson SR (1994) The South Atlantic in the ®ne-resolution Antarctic model. Annls Geophysicae 12: 826±839
Sundberg P, Janson K (1988) Polymorphism in Oerstedia dorsalis(Abilgaard, 1806) revisited. Electrophoretic evidence for aspecies complex. Hydrobiologia 156: 93±98
Swo�ord DL, Selander RB (1989) Biosys-1. A computer pro-gramme for the analysis of allelic variation in population ge-netics and biochemical systematics. Release 1.7. Illinois NaturalHistory Survey, Champaign, Illinois
Thomson JW (1974) The Geology of the South Orkney Islands. III.Coronation Island. Scient Rep Br Antarct Surv 86: 1±39 + 6plates
Thorpe JP (1982) The molecular clock hypothesis: biochemicalevolution, genetic di�erentiation and systematics. A Rev EcolSyst 13: 139±168
Thorpe JP (1983) Enzyme variation, genetic distance and evolu-tionary divergence in relation to levels of taxonomic separation.In: Oxford GS, Rollinson D (eds) Protein polymorphism:
adaptive and taxonomic signi®cance. Academic Press, London,pp 131±152
Thorpe JP, Sole -Cava AM (1994) The use of allozyme electro-phoresis in invertebrate systematics. Zoologica Scr 23: 3±18
Todd CD, Havenhand JN, Thorpe JP (1988) Genetic di�erentia-tion, pelagic larval transport and gene ¯ow between localpopulations of the intertidal marine mollusc Adalaria proxima(Alder & Hancock). Funct Ecol 2: 441±451
Volckaert F, Zouros E (1989) Allozyme and physiological variationin the scallop Placopecten magellanicus and a general model forthe e�ects of heterozygosity on ®tness in marine molluscs. MarBiol 103: 51±61
Waples RS (1987) A multispecies approach to the analysis of gene¯ow in marine shore ®shes. Evolution 41: 385±400
Ward RD, Beardmore JA (1977) Protein variation in the plaice(Pleuronectes platessa). Genet Res 30: 45±62
Ward RD, Skibinski DOF, Woodwark M (1992) Protein hetero-zygosity, protein structure, and taxonomic di�erentiation.Evolutionary Biol 26: 73±159
Whitehouse MJ, Priddle J, Symon C (1996) Seasonal and annualchange in seawater temperature, salinity, nutrient and chloro-phyll a distributions around South Georgia, South-Atlantic.Deep-Sea Res 43: 425±443
Williams KA, Skibinski DOF, Gibson R (1983) Isoenzyme di�er-ences between three closely related species of Lineus(Heteronemertea). J exp mar Biol Ecol 66: 207±211
Workman PL, Niswander JD (1970) Population studies on south-western Indian tribes. II. Local genetic di�erentiation in thePapago. Am J hum Genet 22: 24±29
Wright S (1931) Evolution in Mendelian populations. Genetics,Austin, Tex 16: 97±159
Wright S (1940) Breeding structure of populations in relation tospeciation. Am Nat 74: 232±248
Wright S (1951) The genetical structure of populations. Ann Eugen15: 323±354
Wright S (1965) The interpretation of population structure byF-statistics with special regard to systems of mating. Evolution19: 395±420
Zouros E, Foltz DW (1984) Possible explanations of heterozygotede®ciency in bivalve molluscs. Malacologia 25: 583±591
Zouros E, Mallet AL (1989) Genetic explanations of the growth/heterozygosity correlation in marine mollusks. Proc 23rd Eurmar Biol Symp 317±324 [Ryland JS, Tyler PA (eds) Olsen &Olsen, Fredensborg, Denmark]
13




















