
Population dynamics of the emergent conifer Agathis australis (D. Don) Lindl. (Kauri) in New Zealand...
12
New Zealand Journal of Botany, 1987, Vol. 25: 231-242 0028-825X/87/2502-0231 $2.50/0 © Crown copyright 1987 231 Population dynamics of the emergent conifer Agathis australis (D. Don) Lindl. (kauri) in New Zealand II. Seedling population sizes and gap-phase regeneration JOHN OGDEN GLENDA M. WARDLE MOINUDDIN AHMED Department of Botany, University of Auckland Private Bag, Auckland, New Zealand Keywords Agathis australis; cohort regenera- tion; gap-phase regeneration; half-life; kauri; seed- ling populations; seedling survivorship; Trounson Kauri Park Abstract The view that kauri {Agathis aus- tralis) is a successional species which does not regenerate in mature forest is entrenched in the New Zealand literature. However, seedling and sapling populations ranging from c. 200 to >2000 stems ha" 1 were recorded in 25 mature kauri stands throughout the species' range in the North Island. Higher densities were recorded in gap-phase and successional communities. Gaps created by the fall of mature kauri trees at Trounson Kauri Park aver- aged 0.04 ha, and a few contained abundant kauri seedling and sapling populations. Denser seedling aggregates were sometimes associated with rotting stumps or logs on the forest floor. Trays placed in gaps showed kauri germination and survivorship rates greater than those placed beneath adjacent canopies. Our data support a "cohort regeneration model" in which dense regeneration occurs in successional communities following large-scale disturbance. This leads to self-thinning "ricker" stands in which seedling recruitment is rare, producing a localised "regeneration gap". Continued mortality, increas- ing as the initial survivors begin to senesce, creates a higher frequency of canopy gaps, allowing a second less synchronous wave of recruitment to occur. However, many gaps may be lost to hard- wood species so that succeeding cohorts are less dense. Despite a low efficiency of gap capture, the great longevity of kauri (^ 600 years) implies that the species will survive on any site for 1500 to 2000 years, long enough for large scale stochastic dis- turbance by landslip, storm, or fire to reinitiate the process. Received 2 April 1986; accepted 12 September 1986 INTRODUCTION This paper reviews data on natural regeneration from two recent unpublished works on the ecology of kauri Agathis australis (D. Don) Lindl. (Ahmed 1984; Wardle 1984) and relates them to earlier lit- erature on kauri regeneration. The two studies were independent and different in geographical scope and character. Ahmed (1984) studied 25 stands of mature kauri forest scattered throughout the range of the species. His primary emphasis was on the mature tree population structure (size and age), and the dendrochronological attributes of the species (Ahmed & Ogden 1985). Stand maturity was denned by the presence of trees >60 cm diameter at breast height (d.b.h., c. 1.4 m) in an apparently homogeneous and undisturbed area of at least 2 ha. "Trees" were defined as > 10 cm d.b.h. Wardle (1984) used the same definitions but carried out a more intensive study of stand structure and regen- eration in selected areas at Trounson Kauri Park (Lat. 35°44'S, Long. 173°39'E) and Puketi State Forest (Lat. 35°13'S, Long. 173°45'E), including some preliminary work on seedling recruitment and survivorship in different microsites. Early observations on the ecology of kauri, reviewed below, must be understood within the ecological framework popular at that time. Vege- tation was thought to develop through a series of stages (a succession) until the species composition and structure was in equilibrium with site and cli- matic conditions. In the final, or climax, commu- nity only shade-tolerant species could regenerate, and these were expected to have population struc- tures in which successive size (age) classes were represented by successively fewer individuals (a "reverse-J"-shaped frequency distribution). Light- demanding species were, of necessity, serai, often with single generations occupying temporary hab- itats for time spans dictated by the species longev- ity. This distinction between the climax forest and
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Population dynamics of the emergent conifer Agathis australis (D. Don) Lindl. (Kauri) in New Zealand...

New Zealand Journal of Botany, 1987, Vol. 25: 231-2420028-825X/87/2502-0231 $2.50/0 © Crown copyright 1987
231
Population dynamics of the emergent coniferAgathis australis (D. Don) Lindl. (kauri) in New ZealandII. Seedling population sizes and gap-phase regeneration
JOHN OGDENGLENDA M. WARDLEMOINUDDIN AHMEDDepartment of Botany, University of AucklandPrivate Bag, Auckland, New Zealand
Keywords Agathis australis; cohort regenera-tion; gap-phase regeneration; half-life; kauri; seed-ling populations; seedling survivorship; TrounsonKauri Park
Abstract The view that kauri {Agathis aus-tralis) is a successional species which does notregenerate in mature forest is entrenched in the NewZealand literature. However, seedling and saplingpopulations ranging from c. 200 to >2000 stemsha"1 were recorded in 25 mature kauri standsthroughout the species' range in the North Island.Higher densities were recorded in gap-phase andsuccessional communities. Gaps created by the fallof mature kauri trees at Trounson Kauri Park aver-aged 0.04 ha, and a few contained abundant kauriseedling and sapling populations. Denser seedlingaggregates were sometimes associated with rottingstumps or logs on the forest floor. Trays placed ingaps showed kauri germination and survivorshiprates greater than those placed beneath adjacentcanopies.
Our data support a "cohort regeneration model"in which dense regeneration occurs in successionalcommunities following large-scale disturbance. Thisleads to self-thinning "ricker" stands in whichseedling recruitment is rare, producing a localised"regeneration gap". Continued mortality, increas-ing as the initial survivors begin to senesce, createsa higher frequency of canopy gaps, allowing asecond less synchronous wave of recruitment tooccur. However, many gaps may be lost to hard-wood species so that succeeding cohorts are lessdense. Despite a low efficiency of gap capture, thegreat longevity of kauri (^ 600 years) implies thatthe species will survive on any site for 1500 to 2000years, long enough for large scale stochastic dis-turbance by landslip, storm, or fire to reinitiate theprocess.
Received 2 April 1986; accepted 12 September 1986
INTRODUCTIONThis paper reviews data on natural regenerationfrom two recent unpublished works on the ecologyof kauri Agathis australis (D. Don) Lindl. (Ahmed1984; Wardle 1984) and relates them to earlier lit-erature on kauri regeneration. The two studies wereindependent and different in geographical scope andcharacter. Ahmed (1984) studied 25 stands ofmature kauri forest scattered throughout the rangeof the species. His primary emphasis was on themature tree population structure (size and age), andthe dendrochronological attributes of the species(Ahmed & Ogden 1985). Stand maturity wasdenned by the presence of trees >60 cm diameterat breast height (d.b.h., c. 1.4 m) in an apparentlyhomogeneous and undisturbed area of at least 2 ha."Trees" were defined as > 10 cm d.b.h. Wardle(1984) used the same definitions but carried out amore intensive study of stand structure and regen-eration in selected areas at Trounson Kauri Park(Lat. 35°44'S, Long. 173°39'E) and Puketi StateForest (Lat. 35°13'S, Long. 173°45'E), includingsome preliminary work on seedling recruitment andsurvivorship in different microsites.
Early observations on the ecology of kauri,reviewed below, must be understood within theecological framework popular at that time. Vege-tation was thought to develop through a series ofstages (a succession) until the species compositionand structure was in equilibrium with site and cli-matic conditions. In the final, or climax, commu-nity only shade-tolerant species could regenerate,and these were expected to have population struc-tures in which successive size (age) classes wererepresented by successively fewer individuals (a"reverse-J"-shaped frequency distribution). Light-demanding species were, of necessity, serai, oftenwith single generations occupying temporary hab-itats for time spans dictated by the species longev-ity. This distinction between the climax forest and

232 New Zealand Journal of Botany, 1987, Vol. 25
successional communities has been pervasive in theNew Zealand forest literature (e.g., Holloway 1954)until recently, when gap-phase regeneration hasbeen recognised as an organising paradigm in forestecology (Veblen & Stewart 1982; Ogden 1985;Pickett & White 1985).
Earlier views on the regeneration of kauriWithin the literature the idea has persisted thatregeneration is scarce under groves of mature kauri.Cockayne (1908) commented that he noticed noth-ing between seedlings and mature trees and, later(Cockayne 1928), that "there are no kauri seedlingsin the dense forest". He considered that kauri wasstrongly light-demanding and regarded it as asuccessional species which would eventually bereplaced by the more shade-tolerant hardwoods onany given site. Cheeseman (1914) also noted that"young kauris are not commonly found in grovesof mature trees. Probably the reason is that theamount of light is not sufficient for the growth ofthe very young tree." Although Bieleski (1959a, b,c) demonstrated that reduced light, alone, was notlikely to limit seedling growth in the mature forest— seedlings are shade-tolerant and survive even at2% relative light intensity — he did not doubt thatregeneration was scarce in such situations and sug-gested that soil podzolisation and root competitioncould inhibit seedling establishment. The compar-ative absence of young kauris in mature groves isalso mentioned by Sando (1936), Mirams (1957),Robbins (1957), and Whitmore (1977). Morerecently, Esler (1983) and Halkett (1983) have beenin agreement with these findings.
It is probable that the original observations ofpoor regeneration beneath mature groves werecomparative, contrasting this situation with theabundant and obvious kauri regeneration whichoccurs under a "nurse" of Leptospermum spp.*following extensive forest clearance or fire. Cock-ayne (1928) states that, ".... outside the forest inthe manuka scrub, kauri can be seen coming up inabundance and trees of all ages»are seen". Althoughthe first observation is clearly true, Cockayne'scomment that the trees are of "all ages" may bemisleading. The Leptospermum community per-sists only for c. 150 years (see e.g., Burrows 1973)and kauri seedling establishment is poor in its earlystages, and survivorship poor in its later stages, sothat most of the kauri population falls within arestricted age range (<100 years). In view of theexpected longevity of the dominant individuals in
*Nomenclature follows Allan (1961) except in the case ofAgathis australis (D. Don). Lindl. where Ecroyd (1982)is followed.
such populations (3* 600 years; Ecroyd 1982;Ahmed & Ogden 1987) they should be regarded as"relatively even aged" (Ogden 1985).
The successional process commencing with Lep-tospermum spp. has been described several times(Cockayne 1928; McKinnon 1930; Mirams 1957;Ecroyd 1982; Ogden 1983) and is summarisedbelow (based on Mirams 1957):1. Colonisation by Leptospermum spp.2. Invasion by kauri and/or podocarp seedlings.3. Overtopping of the Leptospermum by the gym-
nosperms ("Ricker" development).4. Elimination of Leptospermum and competitive
thinning of the ricker stand, allowing someinvasion by seedlings of shade-toleranthardwoods.
5. Crown expansion and continued growth of thedominant kauri, with the elimination of smallerindividuals and failure of recruitment creatinga "regeneration gap". The associated podocarpsand hardwoods form a sub-canopy (the matureforest stage).
6. Eventual death of kauri so that the site becomesdominated by podocarp/hardwood forest.Weinmannia silvicola and Nothofagus truncata
may occasionally take the place of Leptospermum,prolonging the early stages (Ecroyd 1982). Thewhole sequence is thought to take 600-1000 years.Although the early stages have been described indetail (Mirams 1957; Ogden 1983) the last twostages, which comprise most of the life span of thecommunity, have received scanty quantitativeattention. Mirams (1957) included "kauri forest"in his study in the Waitakeres, but the total areaof his sampled quadrats in this communityamounted to only 0.14 ha.
From his studies of Puketi forest — then littledisturbed by logging — Latter (1932a, b) suggestedthe possibility that a large proportion of the naturalregeneration of kauri takes placed in small even-aged patches in gaps formed by the fall of old kauris.He found several even-aged patches of pole-sizedkauri which he judged to be of similar size to amature kauri crown. Such patches would eventu-ally give rise to one or two mature trees. He exam-ined seedling abundance in plots and commentedthat the seedlings necessary to fill the gaps causedby the death of old trees are probably present beforethe death occurs. Latter also noted the presence ofhealthy sub-dominant ricker-sized trees in matureforest areas, and suggested that on the fall of thepresent dominants, such trees could expand theircrowns and assume dominance, implying a "steady-state" or "climax" status for the species. Editorialcomment on Latter's paper stated that it wasinteresting because "the generally accepted theory"was that "kauri comes in after manuka (Lepto-

Ogden et al.—Population dynamics of kauri II 233
spermum scoparium, occupies the terrain for a gen-eration, and is then succeeded by a taraire(Beilschmiedia tarairi) association".
Although Latter's (1932a, b) observations do notseem to have weakened the widely held view thatkauri was a successional species, several laterobservations are in agreement with his findings. AtWaipoua Forest, McKinnon (1954) noted that suf-ficient seed was produced to enable some 45 000-62 000 seedlings ha"1 to germinate from a singleseedfall, and observed that seedling survivorshipbeneath the canopy was enhanced by removing thelitter, trenching, and clearing away low under-growth. In the same forest both Cathersides (1972)and Beveridge (1977) noted that occasional saplingsand poles are found in, or close to, mature groves.Beveridge (1977) comments that it is possible forseedlings to persist for long periods in shaded situ-ations, especially on the rotting trunks of large fallentrees where they may eventually reach maturity (R.C. Lloyd pers. comm. in Ecroyd 1982). Bieleski(1959b) conclusively demonstrated that the seed-lings are shade-tolerant and suggested a cyclicregeneration system in which kauri groves werereplaced by hardwoods, which subsequentlyimproved the soil nutrient status and allowed rein-vasion by kauri. Some limited observations on soilprofiles in kauri forest (Gibbs et al. 1968) have beenthought to lend support to this concept (Ecroyd1982). Young kauri have also been reported to growon ridges where podocarps have died, and in slipsamongst broad-leaved shrubs (Ecroyd 1982).
Thus, three main postulates have attempted toaccount for the persistence of mature kauri groves.The "succession theory", generally attributed toCockayne (1928), implies that the kauri populationat any one place is temporary (one generation), sothat the location of mature groves will vary throughtime, tracking the history of major disturbances.Latter's (1932a, b) concepts included group, even-aged regeneration in gaps created by the fall of largetrees, and some more direct replacement of deadkauri trees by subcanopy individuals (the "climax"theory). In contrast to the succession theory, Lat-ter's mechanisms would allow kauri forest to per-petuate itself in situ without major disturbance.Bieleski's (1958c) "cyclic regeneration theory"(based on the work of Watt (1947) and Richards(1952) in forests elsewhere) was intermediate in thesense that it envisaged a landscape mosaic in whichpatches of kauri forest alternated with patches ofpodocarp-hardwood forest in space and time.
The cohort regeneration modelClearly, kauri can act as an early serai species,colonising Leptospermum stands after disturbance,and persisting into a more diverse forest commu-
nity due to its great longevity. It is the ability ofthese old stands to replace themselves in situ whichhas been debated. In a previous paper (Ahmed &Ogden 1987) we presented tree size-frequency dis-tributions (and age data) from mature stands, andargued that these strongly support the view thatsome recruitment occurs in such stands. Our rea-soning was as follows. Relatively even-aged stands,developing after disturbance, would pass througha phase of low recruitment before entering a"senescent" phase in which the individual's sus-ceptibility to death (from a variety of possiblecauses) would increase. During this phase, whichcould extend over one or two centuries, the fre-quency, size, and longevity of canopy gaps wouldbe greater than before, allowing more opportunitiesfor recruitment of kauri seedlings, and giving riseto a second generation of kauris, somewhat lesssynchronous and probably less dense than theoriginal. Our data suggested that two or three suchdepleting generations, or cohorts, could be presentin any area, separated by c. 300 years.
Although the hypothesis that kauri in suchmature forest is able to regenerate by a gap-fillingprocess was proposed over fifty years ago, neitherit, nor the alternative hypothesis that kauri forestin any local site will be succeeded by other foresttypes, has been thoroughly or quantitatively exam-ined. Furthermore, although the shade tolerance ofthe seedlings has been demonstrated, it has not beenestablished what proportion of the sapling andseedling populations must survive in order tomaintain kauri as the physiognomic dominant.
In this paper we present data on seedling andsapling numbers in some mature forest stands, esti-mates of the survivorship of young seedlings in dif-ferent forest environments, and figures on gap sizeand frequency at Trounson Kauri Park. The datawe present are preliminary rather than extensiveor definitive, but taken together they lend supportto the concept of periodic cohort regeneration bygap-filling.
METHODS AND DEFINITIONSMethods for describing the mature tree populationstructure by a point-centered-quarter techniquehave been explained in Ahmed & Ogden (1987). Inaddition, within each stand 20 "seedling" sampleswere taken at 20 m intervals on a paced grid cover-ing about 0.5 ha (80 X 60 m usually). At each sam-pling point a circular plot of 2.5 m radius (19.6 m2)was defined, to give a total sample size of 0.04 haper plot. In most stands "seedlings" were not sub-divided, and this category included all stems< 10 cm d.b.h. In five of the 25 stands, where seed-

234 New Zealand Journal of Botany, 1987, Vol. 25
lings and saplings seemed relatively abundant, thekauri population was classified in greater detail:diameters were recorded to the nearest mm forstems 1-10 cm d.b.h., and smaller seedlings weredivided into "under 1 cm d.b.h." and "cotyledon-ary" (possessing cotyledons only, or with one ortwo additional immature leaves). Stand locationsare given in the notes on Fig. 1.
Wardle (1984) provides total enumerations of thekauri seedling and sapling populations in two 50X 50 m plots (0.25 ha) at Trounson Kauri Park anda similar plot at Puketi Forest (Walnut track). War-dle also studied two "ricker" kauri populationsthought to have arisen following the death of maturetrees, and examined 20 gaps created by the fall ofkauris at Trounson Kauri Park. The sapling speciesconsidered most likely to capture each gap wasdenned on the basis of proximity to the fallen tree,abundance, height, and vigour. "Seedlings" wereclassified either as "cotyledonary", or measuredfrom ground level to apex if <2 cm d.b.h. Stems2-10 cm d.b.h. were called "saplings", and largerindividuals "trees". These definitions are adheredto in this paper.
In a preliminary study of seed dispersal and ger-mination of seeds in canopy gaps, six seed trays(42 X 30 cm; 0.13 m2) containing sterile potting mixwere placed within each of two canopy gaps cre-ated by recent kauri tree falls at Trounson. Detailsof the gaps are given in Table 1. Two additionalsets of six trays were placed beneath the intact kauriforest canopy adjacent to each gap. This samplingregime was set up on February 23 1983, beforeseedfall had begun, and trays were monitored forgerminated seeds (possessing cotyledons) on 28April, 6 July, and 9 September 1983.
Table 1 Details of gaps used in kauri seed germinationstudy at Trounson Kauri Park.
Gap Number1 2
Date of gap creationCauseWater table levelDirection of fall (°)Trunk diameter (cm)Trunk length (m)Crown length (m)Gap length (m)Gap width (m)Diameter of root plate (m)
? 1979 18 Aug 1978? Windthrow Windthrow
high high348 25795 17320 2120 1940 40
c. 15 194.4 7.2
*See map in Wardle 1984, p. 127. Gap 2 is the gap with the view-ing platform on the tourist track. Gap 1 is a more recent gapsituated within the area enclosed by the path, not far from "thefour sisters". Gap 2 data corrected from those given in Wardle(1984).
Seedling survivorship was also monitored overthe same period in two small patches of naturallyestablished seedlings. One of these patches wasassociated with the basal debris mound of a livingkauri tree (120.5 cm d.b.h.) and the other with theremains of a rotting trunk or branch (probably notkauri). The debris mound population was relocatedand counted also on 10 August 1986. Both seedlingpatches were within the area enclosed by the tour-ist track at Trounson Kauri Park.
RESULTS
Seedling and sapling abundance in mature forestTotal stand basal area, and the abundance of kaurisaplings and seedlings (<10 cm d.b.h.) are depictedin Fig. 1, from Ahmed's (1984) survey of 25 maturekauri stands. No relationship was found betweenstand (or kauri) basal area (or density) and theabundance of kauri in the sapling and seedlingclasses. Although seedlings and saplings were rare(<200 ha"1) in a few stands (e.g., Stand 19, ManaiaSanctuary, Coromandel), in others they were abun-dant (>2000 ha^1)- IQ many stands, numbers in thisall-inclusive category ranged from 400 to 800 ha"1.No distinction was made between canopy gaps andclosed forest, so that the results are an average valuefor the stand as a whole. However, it was observedthat seedling distribution was patchy, with areas ofhigher abundance often associated with canopyopenings and/or rotting tree trunks on the forestfloor.
The five stands studied in detail all contained"obvious" seedling and sapling populations. Inmarked contrast to the results obtained by Mirams(1957), and to a lesser extent to those of Wardle(1984), these stands contained more saplings thanlarger trees (Table 2).
Table 2 summarises the results of studies byvarious authors in a comparable format where pos-sible (see note 6). In the mature stands cotyledon-ary seedling populations ranged from 225 to2360 ha"1, established seedlings from 120 to3998 ha"1 and saplings from 0 to 789 ha"'. Thewide variation and the trend for the standarddeviation to increase relative to the mean in olderpopulation categories (cotyledonary < establishedseedlings < saplings) are noteworthy. However,although high seedling numbers were obtained inmature stands by all three authors, Ahmed's (1984)figures are based on a small total area of samplesand may be inflated. Fig. 1 implies that manymature forest stands have lesser seedling numbersthan these means suggest. Wide variation is to beexpected because the seedlings and saplings are dis-tributed in patches rather than at random. Also,

Ogden et al.—Population dynamics of kauri II 235
'a 2500
12000•3
- 1 5 0 0uV
iiooo
500
J3
KEY
Cotyledonary seedlings
Seedlings < 2 cm dbh
Saplings 2 - 1 0 cm dbh
All of the above categories
(0
ia ioo1<0
XI
5 200
Fig. 1 Abundance of small kauri size classes and seedlings (stems <10cm d.b.h.) and stand basal area (m2ha~';stems > 10 cm d.b.h., all species). The upper part of the figure gives the abundance (no. ha ') classified into coty-ledonary seedlings, established seedlings, and saplings in five instances. The lower part of the figure gives basal area.Numbers above the columns are stand numbers as follows (with latitude and longitude in brackets): 1. Te Paki CoastalPark (34°28', 172°46'). 2. Omahuta Sanctuary (35°15\ 173°37'). 3. Te Harua Stream, Puketi S.F. 35°16', 173°44').4. Onekura bluff, Puketi S.F. (35M1', 173°45'). 5. Loop track, Puketi S.F. (35°15', 173°44'). 6. Takapau track, PuketiS.F. (35°13', 173°45')- 7. Manginangina Scenic Reserve (35°12', 173°48'). 8. Russell S.F. (35°23', 174M5'). 9. RidgeWarawara S.F. (35°22\ 173°17'). 10. Plateau Warawara S.F. (35°23', 173°17'). 11. Waipoua Sanctuary (35°29', 173°34').12. Coastal Waipoua S.F. (35°37', 173*29'). 13. Trounson Kauri Park (35°43\ 173°38'). 14. Mt Manaia, Whangarei(35°49', 174°31'). 15. Upper Thumb track, Little Barrier Island (36°12', 175°04'). 16. Lower Thumb track, LittleBarrier Island (36°13', 175°04'). 17. Valley track, Little Barrier Island (3613', 175°04'). 18. Mt Moehau, Coromandel(36°3T, 175°24'). 19. Manaia Sanctuary, Coromandel (36°52', 175°32'). 20. Cascades Kauri Park, Waitakere Range(36°53\ 174°31'). 21. Piha Road, Waitakere Range (36°58\ 174°30'). 22. Huia, Waitakere Range (36°58\ 174°34').23. Konini Forks, Hunua Range (37°04\ 175°08'). 24. Mt William, Hunua Range (37°13\ 175°02'). 25. Katikati S.F.(37°36', 175°52'). For further details of locations see Ahmed (1984). S.F. = State Forest.
the ephemeral nature of the cotyledonary seedlingsmay have led to increased variation produced bysampling at different lengths of time after seedfall.
In all the mature sites, except the kauri forestplot of Mirams (1957), there are between 3 and 14established seedlings or saplings for every mature(^ 10 cm d.b.h.) kauri tree. Despite its pronouncedlack of saplings, the former site has relatively highreplacement potential (34 established seedlings pertree).
The results from both the Trounson stands, and"Gate 8" in particular, are suggestive of decliningpopulations — their densities of big trees are low,as also are numbers of established seedlings andsaplings. However, kauri has by no means relin-quished these sites as there are over 1000 seed-lings ha"1 of various ages potentially available tocolonise gaps should they occur, and a small pro-portion of well established saplings. It is notewor-thy also that the Trounson gap study plot, with

Table 2 Kauri population structures in some mature and "ricker" communities at various sites, and in gap regen-eration patches at Trounson Kauri Park. All figures expressed as no. ha~'.
Site location(and samplingmonth whereknown)
Mature stands"Gate 8"Trounson KauriPark (April)"Camp ground"Kauri Park (Sept)Stand 11.Waipoua ForestStand 6.Takapau track,Puketi ForestStand 3. Te Harua Stream,Puketi ForestKauri ForestWaitakeres(l)Stand 25.Katikati S.F.Walnut trackPuketi (May)Stand 1. Te PakiCoastal Park
Mean for maturestandsStandard deviations
Ricker standsSubmature ricker,Waitakeres(2)Leptospermum-Agathis communityWaitakeres(3)Swanson Waitakeres(Feb)Huapai Waitakeres(4)Oratia Waitakeres(Oct)Leigh (Feb)Gap regeneration^)Puketi Forest(May)Trounson KauriPark (Sept)Gap 4 TrounsonKauri Park(9)
Means forsubmature standsStandard deviations
Cotyledonary(1 yr seedlings)
828
2360
1527
1324
1273
1245
662
2272
225
1302699
190
1406
nrnr
nrnr
1114
4265
440
14831631
Individuals
Seedlings(6)<2 cm d.b.h.
208
120
611
1069
560
3998
1349
632
1095
10711170
7317
13780
nrnr
nrnr
14799
4529
5120
91094855
h a '
Saplings(6)2-10 cm
4
8
305
178
393
0
789
16
458
239273
524
1379
14663832
14301383
314
353
100
11981134
Trees(6)5= 10 cm d.b.h.
16(8)
20
71
103
114
118
149
208
180
10966
812
451
636873
360900
57
176
20
473347
Kauribasalarea
m2ha-'(7)
40
33
49
65
39
-
38
102
23
4925
-
_
4637
2128
6
7
-
2416
References
Wardle 1984
Wardle 1984
Ahmed 1984
Ahmed 1984
Ahmed 1984
Mirams 1957
Ahmed 1984
Wardle 1984
Ahmed 1984
Mirams 1957
Mirams 1957
Ogden 1983Ogden 1983
Ogden 1983Ogden 1983
Wardle 1984
Wardle 1984
Wardle 1984
Notes: (l)Regarded by Mirams (1957) as "mature", but probably younger than the other stands included as mature here; his community5. All Mirams' data are means derived from two sets presented in his Table 2 (p. 665). (2)Mirams (1957) community 4. (3)Mirams(1957) community 3. This community is roughly equivalent to those of Ogden (1983). (4)Mean of two samples Feb 1980 and Sept1981. (5)The gaps studied at Puketi and Trounson were respectively 0.035, 0.034, and 0.05 ha. (6)In the instance of the data fromMirams (1957) "seedlings" includes his Classes II to IV and thus includes a few up to 5 cm d.b.h. Likewise Mirams' "saplings" rangefrom 5-15 cm and "trees" are > 15 cm d.b.h. (7)Trees > 10 cm only. (8)There were no trees in the 10-30 cm d.b.h. range. (9)Incorrectlyreferred to as gap 13 in Wardle (1984) p. 123.
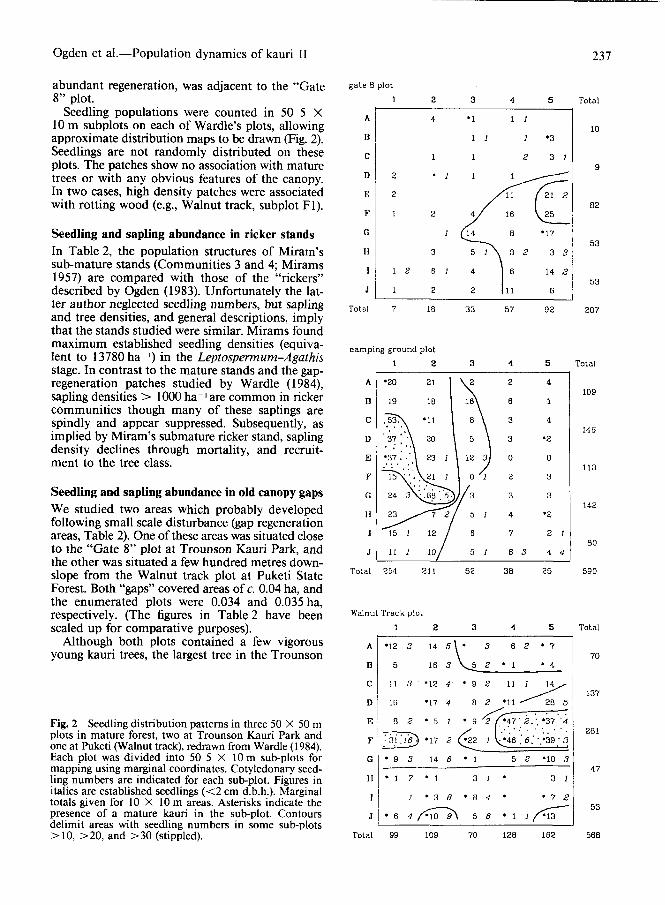
Ogden et al.—Population dynamics of kauri II 237
abundant regeneration, was adjacent to the "Gate 8ale B P'°1
8" plot.Seedling populations were counted in 50 5 X
10 m subplots on each of Wardle's plots, allowingapproximate distribution maps to be drawn (Fig. 2).Seedlings are not randomly distributed on theseplots. The patches show no association with maturetrees or with any obvious features of the canopy.In two cases, high density patches were associatedwith rotting wood (e.g., Walnut track, subplot Fl).
Seedling and sapling abundance in ricker standsIn Table 2, the population structures of Miram'ssub-mature stands (Communities 3 and 4; Mirams1957) are compared with those of the "rickers"described by Ogden (1983). Unfortunately the lat-ter author neglected seedling numbers, but sapling Totaland tree densities, and general descriptions, implythat the stands studied were similar. Mirams foundmaximum established seedling densities (equiva-lent to 13780 ha"1) in the Leptospermum-Agathisstage. In contrast to the mature stands and the gap-regeneration patches studied by Wardle (1984),sapling densities > 1000 ha~' are common in rickercommunities though many of these saplings arespindly and appear suppressed. Subsequently, asimplied by Miram's submature ricker stand, saplingdensity declines through mortality, and recruit-ment to the tree class.
Seedling and sapling abundance in old canopy gapsWe studied two areas which probably developedfollowing small scale disturbance (gap regenerationareas, Table 2). One of these areas was situated closeto the "Gate 8" plot at Trounson Kauri Park, andthe other was situated a few hundred metres down-slope from the Walnut track plot at Puketi StateForest. Both "gaps" covered areas of c. 0.04 ha, andthe enumerated plots were 0.034 and 0.035 ha,respectively. (The figures in Table 2 have been Walnul Track Plot
scaled up for comparative purposes).Although both plots contained a few vigorous
young kauri trees, the largest tree in the Trounson
Fig. 2 Seedling distribution patterns in three 50 X 50 mplots in mature forest, two at Trounson Kauri Park andone at Puketi (Walnut track), redrawn from Wardle (1984).Each plot was divided into 50 5 X 10 m sub-plots formapping using marginal coordinates. Cotyledonary seed-ling numbers are indicated for each sub-plot. Figures initalics are established seedlings (<2 cm d.b.h.). Marginaltotals given for 10 X 10 m areas. Asterisks indicate thepresence of a mature kauri in the sub-plot. Contoursdelimit areas with seedling numbers in some sub-plots> 10, > 20, and > 30 (stippled). Total 99
A
B
C
D
E
F
G
H
I
J
1
2
2
1
1 2
1
2
4
1
•
2
3
6
1
1
1
I
3
•1
1 1
1
1
V
5 l\
4
2
4
1
1
'll
16
8
3
6
11
1
1
2
_
2
5
•3
3
--—
•17
3
14
6
1
_~~
2
3
2
18
camping ground plot1 2
A
B
C
D
E
F
G
H
I
J
Total 254 211 82 38 25
Total
10
92
•20
19
."53\
•37 . •' |
24 3>
23 ^
15 J
11 1
21
18
•11
20
23
12
10
V 2
8
I 5
1 \ 12
I 0
/ 3
2 5
/ 5
\
\3j
1
1
1
2
8
3
3
0
2
3
4
7
8 3
4
1
4
•2
0
3
3
•2
2 1
4 4
B2
53
53
207
Total
109
146
113
B0
A
B
C
D
E
F
G
H
I
J
1
•12
5
11
IB
3
: 31.'
• 9
• 1
• 6
3
3
2
3)3
7
1
4 1
Z
14
16
•12
•17
• 5
•17
14
* 1
• 3
'no
5\3 ̂
4
4
1
s <6
8
9\
3
•
• 9
8
• 9/
• 1
3
• 8
5
3
2
2
2
Ai
4
6
4
6
• 1
11
•11
/~^~~
-46
5
•
•
• 1
2
1
' 2.\
.'6.'
2
1 {
b
• 7
• 4
14
28
•37
.•39
•10
3
• 7
•^13
5
'4
• 3
3
1
2
109 70 128 163
Total
70
137
281
47
53
568

238 New Zealand Journal of Botany, 1987, Vol. 25
plot was 35.2 cm d.b.h., and the largest in the Puketiplot was 44.8 cm d.b.h. The total (tree) basal areaat Trounson was 43m2ha~' with Beilschmiediatarairi, Cyathea dealbata, and Podocarpus hallii allhaving greater basal areas than kauri (c. 6 m2 ha~')-At Puketi the total (tree) basal area was 52 m2 ha"1
of which kauri comprised c. 7 m2 ha~'. The mainassociates were Leptospermum ericoides (domi-nant), Cyathea dealbata, Weinmannia silvicola, andPhyllocladus trichomanoides.
The species composition and location of the plotsare consistent with the view that the Trounson gapregeneration area originated following collapse ofcanopy trees in the area while the Puketi study areaarose after a localised fire. In both instances, thedisturbance must have occurred over a century ago,judged from the presence of kauri, podocarp, andB. tarairi stems c. 35 cm d.b.h. at Trounson, andLeptospermum ericoides of similar size at Puketi.However, most of the kauri regeneration at bothsites is in the seedling classes, implying that con-ditions for kauri seedling establishment did notdevelop immediately, but were delayed until a"nurse" canopy of other species had developed. Thelarge number of established seedlings in the Puketiplot suggests that it is highly favourable for regen-eration, as expected from the abundance of Lep-tospermum on the plot (cf. Mirams 1957,Leptospermum-Agathis community).
The significant points are that both areas are ona scale consistent with formation by the fall of oneor two large kauri trees, both are situated withinlarger areas of mature forest, and both containabundant populations of well established seedlings(Table 2).
In addition to the studies described above, 20gaps created by the fall of (normally) single kauritrees were surveyed at Trounson Kauri Park. Thegaps varied in age from <5 years to >100 years,but difficulties of ageing them with certainty pre-vented them from being ranked to depict the gapclosure process. Established kauri seedlings wererecorded in seven of the 20 gaps, and in threeinstances were regarded as potential replacementsfor the fallen tree. However, in the majority ofinstances (85-95%), gaps appeared most likely tobe colonised by Beilschmiedia tarairi, Weinman-nia silvicola, Ixerba brexioides, Quintinia serrata,or Podocarpus ferrugineus. In ten instances, gapdimensions were recorded. Gap length ranged from18 to 41 m with a mean of 30 m, and width rangedfrom 7 to 20 m with a mean of 15 m. These figuresimply average gap areas of c. 0.04 ha. The trunkdiameter of the trees creating these gaps rangedfrom 85 to 293 cm with a mean of 155 cm. Windwas clearly the main cause of mortality, with mosttrees uprooted and orientated between north-west
and south-west, implicating cyclonic storms fromthe north-east (Conway 1959). Examination of themore recent gaps showed that the fall of a largekauri normally destroys at least three and some-times as many as five associated canopy trees (B.tarairi, W. silvicola, etc.) as well as numerousunderstorey trees and saplings.
In one gap, created by the fall of two adjacent,large kauri trees (c. 3 m d.b.h.) and containing theirdecomposing stumps and prostrate trunks, the kauriseedling and sapling population was counted(Gap 4, Table 2). This gap contained 252 estab-lished kauri seedlings, especially on the rotting woodaround the bases of the fallen kauris, six seedlings>1 m tall, four saplings (2.5, 4.7, 5.8, and 6.7 cmd.b.h.) and one young kauri tree (28 cm d.b.h.).Although such dense gap seedling populations maybe unusual, Gap 4 illustrates the potential for atleast a small proportion of gaps to be colonised bykauri, probably some decades after their originalformation. One of the kauri saplings (5.8 cm d.b.h.and 9 m tall) was growing directly on a fallen kauritrunk, along with a miro {Podocarpus ferrugineus)of 20 cm d.b.h. and an Ixerba brexioides of 24 cmd.b.h.
Seedling survivorshipTable 3 summarises the results of the seed-tray ger-mination and survivorship study. The actual num-bers germinating may be high because of the "ideal"substrate provided in the trays, although they aresimilar to figures recorded by McKinnon (1954),and an order of magnitude less than the numberof sound seeds per hectare recorded by Mirams(1957). Mortality accounted for about 30% of theseedlings over the six months (winter) period. Bothgermination and survivorship were greatest in traysplaced in canopy gaps.
The half-life of a population is the time requiredfor its numbers to become reduced to half theirformer value. Its calculation assumes that mortal-ity removes a constant proportion of the survivorseach year (Harper 1967). Because of the short timeover which observations were made, the half-lifevalues are crude approximations only but they doreflect the differential survivorship between the twomicrosites, and allow comparisons with other data.The preliminary results suggest higher recruitmentrates in gaps.
Survivorships of two small groups of naturallyestablished seedlings, situated on the debris moundof a large, living kauri tree, and in association witha decomposing subcanopy log, are given in Table 4.Mortality accounted for 40-50% of both popula-tions during the period of observation, and theirestimated half-lives were similar, and longer thanthose of first year seedlings also situated beneath

Ogden et al.—Population dynamics of kauri II 239
Table 3 Germination and survivorship of first-year kauri seedlings on sterilepotting mix in 12 0.126 m2 trays placed beneath the canopy, and 12 0.126 m2
trays in adjacent canopy gaps at Trounson Kauri Park in the autumn and winterof 1983.
Mean number germinated per tray1
Feb 23-July 6, 1983Total survivorship %28 April-9 Sept 1983Approximate half-life (yrs)2
PositionBeneath canopy
0.88 (33 X 103)
670.9
of traysIn gap
2.26 (63 X 103)
761.3
'Figures in brackets are expressed on a per hectare basis. Means are significantly different(t=2.708, n=24, P<0.05).Calculated on the assumption that measured mortality rates continue.
Table 4 Survivorship of naturally established kauriseedlings on a basal trunk mound and adjacent to a rot-ting log beneath a closed canopy at Trounson Kauri Park1983.
SiteMound Log
Initial sample size (24 Feb 1983) 21 12No. of leaves, mean, and range1 3.4(2-6) 4.0(2-12)Survivorship % (24 Feb-13 Oct) 62 50Approximate half-life (years) 1.3 1.2
•Age range probably 2 to 6 years; see Mirams (1957): 664-678.
the canopy and growing in favourable substrates(Table 3). Survivorship probably increases as theseedlings get bigger. In a re-examination of themound population (only) on 18 August 1986, fiveof the seedlings tagged in February 1983 were cer-tainly identified. The expected number, on the basisof the previously calculated half-life (1.3 years;Table 4) was three. These seedlings had gained fromzero to six new leaves (mean = 2.8). A further threeuntagged seedlings might have been "survivors"and there were eight new recruits, so that the totalpopulation had declined from 21 to 16 over Vkyears.
DISCUSSIONThe results presented suggest a new emphasis onthe regeneration of kauri. Gap-phase regenerationis envisaged as a mechanism whereby kauri popu-lations are maintained on any site between periodsof larger-scale destruction initiating dense seraipopulations. This regeneration mechanism is
reflected in the physiology of the seedlings. Peri-odic large-scale regeneration after disturbancefollowed by gap-phase regeneration may be com-mon in long-lived conifers in New Zealand (Ogden1985), and its recognition largely eliminates theproblem of climatic climax species with "regenera-tion gaps" which has troubled earlier authors.
For most of the stands depicted in Fig. 1, no dis-tinction was made between ephemeral seedlings afew years old and larger saplings which may havebeen up to 100 years old (<10cm d.b.h.). Con-sequently, Fig. 1 must be regarded as suggestive orindicative only, and be interpreted in the contextof the total age span in the mature stands studied,usually >600 years (Ahmed & Ogden 1987). Takenalong with independent data, reviewed in Table 2,it shows that some mature kauri stands have estab-lished seedling populations ranging from a fewhundred to a few thousand per hectare. Much higherdensities occur in ricker stands and in old canopygaps. Saplings and small trees are sometimes rareor even absent (in small samples), but number sev-eral hundred per hectare in some stands. The pre-viously reported scarcity of individuals in this sizeclass (e.g., Mirams 1957) probably arose, in part,through selective sampling of high density groves,in which self-thinning was still occurring andrecruitment consequently rare. Conversely it mightbe argued that our samples included more juveniletrees because they often extended beyond the mar-gins of dense groves and/or included areas of hard-wood-dominated, low density kauri forest. Botharguments imply spatial and temporal heteroge-neity in regeneration and recruitment to larger sizeclasses. This is consistent with our hypothesisedregeneration model, involving periodic large scaledisturbance ("synchronising events") giving an ini-tially high density population which persists throughseveral generations by gap-phase regeneration. To

240 New Zealand Journal of Botany, 1987, Vol. 25
argue that our results may apply to one "type" ofkauri forest (in the sense of Nicholls 1976) but notto another, would ascribe an unrealistic discrete-ness to such types. Kauri regeneration can beunderstood only by examining it as a populationphenomenon crossing arbitrary community bound-aries. Indeed, it seems likely that some of the 22kauri forest types recognised by Nicholls (1976) dif-fer mainly in the age structure of their kauripopulations.
Our observations of regeneration in maturestands are not new, rather they confirm and extendthe earlier work of Latter (1932a, b) (in particular)and Beveridge (1977). Demonstration of the shadetolerance of kauri seedlings (Bieleski 1959a) andobservations on the considerable ages achieved bykauri seedlings in shaded situations (Ogden 1983)are consistent with the ability of the species tomaintain subcanopy seedling populations. Indi-viduals are "released" from such populations bycanopy opening following the fall of a large tree.Recruitment following release is implied also byobservations of increment cores and cross sections,which frequently show a central core of very nar-row rings.
Gaps may also become suitable for seedling ger-mination and survival, once a suitable microcli-mate has developed beneath shorter-lived gap-fillingspecies. Within such old gaps rotting wood pro-vides a favourable rooting medium, leading to twoscales of patchiness in the seedling population: dif-fuse patches associated with old light gaps, andsuperimposed local aggregations on suitable sub-strates, such as rotting logs. The maps (Fig. 2) sug-gest other intermediate scales for which no generalexplanation can be suggested. In contrast to the slowseedling growth rates noted by Ogden (1983) inricker stands, 17 vigorous-looking seedlings 2-3.5 mtall selected from gaps in mature forest had a meanage of 16 + 2 years (mean and standard error),showing that seedling growth rates vary widely, andcan be high.
Enumeration of 20 gaps of various ages atTrounson Kauri Park suggests that only 5-15% ofkauri tree fall gaps are recolonised by kauri, theremainder being lost (at least temporarily) toshorter-lived, hardwood species or miro (Podocar-pus ferrugineus). If these figures are even approxi-mately correct they imply that each succeedingcohort is much reduced; indeed a populationdeveloping from a dense ricker stand would bealmost eliminated by the third cohort regeneration.However, three cohorts imply a time span of atleast 1000 years, during which time massive dis-turbance by cyclone or fire might be expected toreinitiate the successional process.
Past emphasis on regeneration in successionalcommunities following disturbance may havecaused the relatively sparse and patchy regenera-tion in mature forest to have been largely over-looked. Another problem lies in the definition of"seedlings", a term which has been used differentlyby different authors. In forest ecology, crops ofcotyledonary seedlings are often regarded asephemeral phenomena and disregarded, only"established" seedlings being counted. While thereis some justification for this in so far as mortalityis clearly high in very young seedlings, it is never-theless from these annual crops that the "estab-lished seedling pool" is recruited, so that knowledgeof their survivorship is important. Cotyledonaryseedling populations ranged from c. 200 to>2000 ha"'in the mature stands reported here. Theestimated survivorship of these seedlings suggestspopulation half-lives of c. 1 year. Assuming thatsurvivorship increases (rather than decreases) as theseedlings get bigger, these figures alone imply acontinuous, if small, population of older seedlings.For example, based on the seed trays, the esti-mated seedling population germinating in average(0.04 ha) gaps in Trounson in 1983 was c. 2500 pergap. Even assuming (arbitrarily) that this figure isan order of magnitude greater than the real ger-mination rate on the forest floor, some of theseindividuals can be expected to survive for a dec-ade. The half-life estimates obtained for thesepopulations, albeit based on very small samples,are similar to those obtained for cotyledonary seed-ling populations of red beech (Nothofagus fused) infavourable microsites (June & Ogden 1975). Wehave calculated higher half-lives for kauri seedlingpopulations in Mirams' Leptospermum-Agathisstand (Table 5), emphasising that this communityis highly favourable for kauri regeneration.
Despite the work of Latter (1932a) the impor-tance of gap-phase regeneration in mature kauriforest has not been appreciated. Our results, pre-sented in Table 2, show patches of relatively densekauri regeneration on a scale similar to that of can-opy gaps in areas of otherwise mature forest. In oneinstance (Gap 4) a relatively dense seedling popu-lation and some well-established saplings wereoccupying an old tree-fall gap.
The average gap size recorded at Trounson(0.04 ha) is large relative to those reported in podo-carp-hardwood forest (0.023 ha) by Smale & Kim-berley (1983). In red beech {Nothofagus fused) forestJune & Ogden (1978) reported gaps up to 40 m wideand 0.07 ha in area.
Some simple calculations serve to illustrate that,even in the absence of catastrophic destruction,random mortality will result in every part of theforest floor entering a "gap phase" condition over

Ogden et al.—Population dynamics of kauri II 241
Table 5 Survivorship of naturally established first-year kauri seedlings in a varietyof different community types in the Waitakeres (data from Mirams 1957).
Community type1
1 3 4 5
Average no. lst-year seedlings ha~l
% Survivorship year 1 to 2Approximate half-life (yrs)
'Type 1, young Leptospermum; type 3, Leptospermum-Agathis; type 4, kauri ricker <munity; type 5, mature kauri.
14034.10.6
109088.
6.25
252038.30.7
2360not given
a time span less than the longevity of the maturetrees. Assume that only trees ^ 1 m d.b.h. creategaps (of 0.04 ha) and that such trees occur at a den-sity of 10 per hectare and have a mortality rate of0.5% per annum. These figures imply that gaps willoccur with a frequency of one per hectare every 20years, and that all parts of the hectare will haveexperienced a gap phase over a 500-year period. Ifwe further assume that gaps remain suitable forcolonisation by kauri for at least 50 years then c.three gaps per hectare (0.12 ha) are continuouslyavailable for colonisation.
These figures are of theoretical interest, but thecohort regeneration model suggested by our dataimplies that recruitment is not continuous or ran-dom in time, but rather it is periodic. Dense regen-eration following disturbance leads to self-thinningricker stands in which seedling recruitment is low,perhaps for one or two centuries. As such standsreach maturity and begin to senesce, mortality willcreate a higher frequency of larger gaps. At thisstage, which may also be prolonged, a second waveof recruitment can occur. The data presented here,albeit preliminary and of limited extent, indicatethat gaps in mature forest are indeed favourablesites for kauri regeneration.
ACKNOWLEDGMENTSPersonnel of the New Zealand Forest Service, in the HeadOffice of Auckland Conservancy, in the Kauri Manage-ment Unit at Kaikohe, and in the district office at Thameshelped with site location and field logistics. Research atTrounson Kauri Park was facilitated by a grant from theDepartment of Lands and Survey, and by the cooperationof the ranger, Mr Ken Hamilton. Martin and Jo Bridge,Elizabeth Brown, Ian Horner, Colin Macdonald, ShonaMyers, Marion Steel, Nigel Taylor, Mark Wardle, andCarol West helped with stand enumeration at Trounsonand Puketi. The manuscript was prepared while one ofus (J. O.) held a Bullard Fellowship at Harvard Univer-sity and was typed by Ms Dottie Smith.
REFERENCESAhmed, A. 1984: Ecological and dendrochronological
studies on Agathis australis Salisb. (kauri). Unpub-lished Ph.D. thesis, University of Auckland, NewZealand.
Ahmed, M.; Ogden, J. 1985: Modern New Zealand tree-ring chronologies. 3. Agathis australis (Salisb.) —kauri. Tree ring bulletin 45 : 11-24.
1987: Population dynamics of the emergentconifer Agathis australis (D. Don) Lindl. (kauri) inNew Zealand. I. Population structures and treegrowth rates in mature stands. New Zealand jour-nal of botany 25: 217-229.
Allan, H. H. 1961: Flora of New Zealand, Vol. 1. Wel-lington, Government Printer.
Beveridge, A. E. 1977: Notes on silviculture of kauri. In:Chavasse, C. G. R. ed.. Forestry Handbook. Roto-rua, New Zealand Institute of Foresters (Inc.). Pp.125-294.
Bieleski, R. L. 1959a: Factors affecting growth and dis-tribution of kauri (Agathis australis Salisb.). 1. Effectof light on the establishment of kauri and Phyl-locladus trichomanoides D. Don. Australian jour-nal of botany 7: 252-267.
1959b: Factors affecting growth and distribu-tion of kauri (Agathis australis Salisb.). 2. Effect oflight intensity of seedling growth. Australian jour-nal of botany 7: 268-278.
1959c: Factors affecting growth and distribu-tion of kauri (Agathis australis Salisb.). 3. Effect oftemperature and soil conditions. Australian jour-nal of botany 7: 279-294.
Burrows, C. J. 1973: The ecological niches of Leptosper-mum scoparium and L. ericoides (Angiospermae:Myrtaceae). Mauri Ora 1: 5-12.
Cathersides, P. S. 1972: A geographical appraisal of thedistribution of Agathis australis, Beilschmiedia tawaand Dacrydium cupressinum communities inselected areas of the North Island of New Zealand.Unpublished M.A. thesis. University of Auckland,New Zealand.
Cheeseman, T. F. 1914: The age and growth of the kauriAgathis australis). Transactions and proceedings ofthe New Zealand Institute 46 : 9-19.

242 New Zealand Journal of Botany, 1987, Vol. 25
Cockayne, L. 1908: Report on a botanical survey of Wai-poua kauri forest. Appendix to the Journals of theHouse of Representatives (New Zealand). C-14. 44P-
1928: The vegetation of New Zealand. (Ed. 2)Leipzig, Engelmann.
Conway, M. J. 1959: Hurricane damage in Northland.New Zealand journal of forestry 8 : 151.
Ecroyd, C. E. 1982: Biological flora of New Zealand. 8.Agathis australis (D. Don) Lindl. (Araucariaceae)Kauri. New Zealand journal of botany 20 : 17-36.
Esler, A. E. 1983: Forest and scrubland zones of the Wai-takere Range, Auckland. Tone 29: 109-117.
Gibbs, H. S.; Cowie, J. D.; Pullar, W. A. 1968: Soils ofnorth Auckland. In: Soils of New Zealand. NewZealand Soil Bureau bulletin 26 : 48-52.
Halkett, J. C. 1983: A basis for the management of NewZealand kauri (Agathis australis (D. Don) Lindl.)forest. New Zealand journal of forestry 28 : 15-23.
Harper, J. L. 1967: A Darwinian approach to plant ecol-ogy. Journal of ecology 55 : 247-270.
Holloway, J. T. 1954: Forests and climates in the SouthIsland of New Zealand. Transactions of the RoyalSociety of New Zealand 82: 329-410.
June, S. R.; Ogden, J. 1975: Studies on the vegetation ofMount Colenso, New Zealand. 3. The populationdynamics of red beech seedlings. Proceedings of theNew Zealand Ecological Society 22 : 61-66.
1978: Studies on the vegetation of Mount Col-enso, New Zealand. 4. An assessment of the pro-cess of canopy maintenance and regenerationstrategy in a red beech (Nothofagus fusca) forest.New Zealand journal of ecology 1: 7-15.
Latter, H. B. 1932a: The Puketi kauri forest. An attemptto apply the quadrat method to the ecological studyof the kauri forest. Unpublished B. For. Sci. thesis.Canterbury University College, New Zealand.
1932b: An ecological study of kauri forest (the-sis abstract). Te kura ngahere 3 : 88-92.
McKinnon, A. D. 1930: Vegetation survey of the Auck-land Forest School Reserve, Swanson, and theHuia, Nihotapu and Waitakere Catchment areas.
Unpublished B. For. Sci. thesis, University ofAuckland, New Zealand. (In Mirams 1957).
1954: Natural regeneration of kauri. NewZealand journal of forestry 5 : 133-137.
Mirams, R. V. 1957: Aspects of the natural regenerationof kauri (Agathis australis Salisb.) Transactions ofthe Royal Society of New Zealand 84 : 661-680.
Nicholls, J. L. 1976: A revised classification of the NorthIsland indigenous forests. New Zealand journal offorestry 21: 105-132.
Ogden, J. 1983: The scientific reserves of Auckland Uni-versity. II. Quantitative vegetation studies. Tane29: 163-180.
1985: An introduction to plant demographywith special reference to New Zealand trees. NewZealand journal of botany 23 : 751-772.
Pickett, S. T. A.; White, P. S. (ed.). 1985: The ecology ofnatural disturbance and patch dynamics. London,Academic Press Ltd.
Richards, P. W. 1952: The tropical rain forest: an eco-logical study. Cambridge, Cambridge UniversityPress.
Robbins, R. G. 1957: The status and classification of NewZealand forest vegetation. Unpublished Ph.D. the-sis, University of Auckland, New Zealand.
Sando, C. T. 1936: Notes on Agathis australis. NewZealand journal of forestry 4 : 16-21.
Smale, M. C; Kimberley, M. O. 1983: Regeneration pat-terns in Beilschmiedia tawa forest at Rotoehu. NewZealand journal of forestry science 13 : 58-71.
Veblen, T. T.; Stewart, G. H. 1982: On the conifer regen-eration gap in New Zealand: the dynamics of Libo-cedrus bidwillii stands on South Island. Journal ofecology 70: 413-436.
Wardle, G. M. 1984: Observations on the structure anddynamics of kauri (Agathis australis) forest.Unpublished M.Sc. thesis, University of Auck-land, New Zealand.
Watt, A. S. 1947: Pattern and process in the plant com-munity. Journal of ecology 35 : 1-22.
Whitmore, T. C. 1977: A first look at Agathis. Tropicalforestry papers No. 11. Department of Forestry,Oxford University.




















