
Population dynamics in a cyclic environment: consequences of cyclic food abundance on tawny owl...
13
Population dynamics in a cyclic environment: consequences of cyclic food abundance on tawny owl reproduction and survival Patrik Karell 1 *, Kari Ahola 2 , Teuvo Karstinen 3 , Aniko Zolei 4 and Jon E. Brommer 1 1 Bird Ecology Unit, Department of Biological and Environmental Sciences, P.O. Box 65 (Viikinkaari 1), FI-00014 University of Helsinki, Finland; 2 Tornihaukantie 8D 72, FI-02620 Espoo, Finland; 3 Juusinkuja 1, FI-02700 Kauniainen, Finland; and 4 Duna-Ipoly National Park Directorate, 1021 Budapest, Hu ´ ¢vo ¨svo ¨ lgyi u ´t52 Summary 1. Understanding which factors regulate population dynamics may help us to understand how a population would respond to environmental change, and why some populations are declining. 2. In southern Finland, vole abundance shows a three-phased cycle of low, increase and decrease phases, but these have been fading out in recent years. During five such cycles (1981–1995), all tawny owls Strix aluco were censused in a 250-km 2 study area, and their reproduction and survival were monitored. 3. Males and females showed similar dynamics, but experienced breeders recruited more offspring and had higher survival than first breeders. Offspring recruitment, but not survival of breeding individuals varied in accordance with vole abundance. 4. The population’s numerical response to prey abundance was primarily due to first-breeding individuals entering the population in the increase phase when immigration was the highest. First- breeding birds were younger, but experienced breeders were older in more favourable vole years. 5. A stage-specific matrix population model integrating survival and fecundity showed that, despite obvious variation in fecundity between vole cycle phases, this variation had limited impor- tance for overall tawny owl population dynamics, but that the survival of experienced breeders during the low phase is most important for population growth. 6. Model and data agreed that the vole cycle drives the dynamics of this avian predator by limiting the recruitment of new breeders during the low phase. Population dynamics hence differ not only from the classic example of the species in a more temperate region in the UK where the number of territories is stable across years, but also from the dynamics of other avian vole predators in Fen- noscandia where the recurring crash in vole abundance drastically lowers adult survival thereby creating vacancies. Key-words: Clethrionomys glareolus, life stage, Microtus agrestis, population cycle, predator- prey interaction Introduction Since the seminal work of Charles Elton (1927), population cycles of predators and their prey have been a focal topic of animal ecology and population dynamics (Southern 1970; Hanski, Hansson & Henttonen 1991; Hanski & Korpima¨ ki 1995; Krebs et al. 1995; Lambin, Petty & MacKinnon 2000; Lindstro¨ m et al. 2001; Gilg, Hanski & Sittler 2003; Sundell et al. 2004; Korpima¨ ki et al. 2005a, b). Cyclic fluctuations in the abundance of herbivores are commonly found in popula- tions on high latitudes and ⁄ or high altitudes (Lindstro¨ m et al. 2001). Because these herbivores typically are basal to the ecosystem, the effect of the cycles in their abundance reverberates across the food web (Ims, Henden & Killengreen 2008). The consequences of herbivore cycles are thus appar- ent also on higher trophic levels, even when these predators do not directly drive the cycle. This perspective is in contrast to the classic view of predator–prey dynamics as a Lotka– Volterra type of dynamics, where the predator drives the fluc- tuations in the abundance of the prey and shows similar cycles as its prey but lagging in time. A classic example of predator-prey population dynamics where the predator’s *Correspondence author. E-mail: patrik.karell@helsinki.fi Journal of Animal Ecology 2009, 78, 1050–1062 doi: 10.1111/j.1365-2656.2009.01563.x ȑ 2009 The Authors. Journal compilation ȑ 2009 British Ecological Society
Transcript of Population dynamics in a cyclic environment: consequences of cyclic food abundance on tawny owl...

Population dynamics in a cyclic environment:
consequences of cyclic food abundance on tawny
owl reproduction and survival
Patrik Karell1*, Kari Ahola2, TeuvoKarstinen3, Aniko Zolei4 and Jon E. Brommer1
1Bird Ecology Unit, Department of Biological and Environmental Sciences, P.O. Box 65 (Viikinkaari 1), FI-00014 University
of Helsinki, Finland; 2Tornihaukantie 8D 72, FI-02620 Espoo, Finland; 3Juusinkuja 1, FI-02700 Kauniainen, Finland; and4Duna-Ipoly National Park Directorate, 1021 Budapest, Hu¢vosvolgyi ut 52
Summary
1. Understanding which factors regulate population dynamics may help us to understand how a
population would respond to environmental change, and why some populations are declining.
2. In southern Finland, vole abundance shows a three-phased cycle of low, increase and decrease
phases, but these have been fading out in recent years. During five such cycles (1981–1995), all
tawny owls Strix alucowere censused in a 250-km2 study area, and their reproduction and survival
were monitored.
3. Males and females showed similar dynamics, but experienced breeders recruited more offspring
and had higher survival than first breeders. Offspring recruitment, but not survival of breeding
individuals varied in accordance with vole abundance.
4. The population’s numerical response to prey abundance was primarily due to first-breeding
individuals entering the population in the increase phase when immigration was the highest. First-
breeding birds were younger, but experienced breeders were older in more favourable vole years.
5. A stage-specific matrix population model integrating survival and fecundity showed that,
despite obvious variation in fecundity between vole cycle phases, this variation had limited impor-
tance for overall tawny owl population dynamics, but that the survival of experienced breeders
during the low phase is most important for population growth.
6. Model and data agreed that the vole cycle drives the dynamics of this avian predator by limiting
the recruitment of new breeders during the low phase. Population dynamics hence differ not only
from the classic example of the species in a more temperate region in the UK where the number of
territories is stable across years, but also from the dynamics of other avian vole predators in Fen-
noscandia where the recurring crash in vole abundance drastically lowers adult survival thereby
creating vacancies.
Key-words: Clethrionomys glareolus, life stage, Microtus agrestis, population cycle, predator-
prey interaction
Introduction
Since the seminal work of Charles Elton (1927), population
cycles of predators and their prey have been a focal topic of
animal ecology and population dynamics (Southern 1970;
Hanski, Hansson & Henttonen 1991; Hanski & Korpimaki
1995; Krebs et al. 1995; Lambin, Petty & MacKinnon 2000;
Lindstrom et al. 2001; Gilg, Hanski & Sittler 2003; Sundell
et al. 2004; Korpimaki et al. 2005a, b). Cyclic fluctuations in
the abundance of herbivores are commonly found in popula-
tions on high latitudes and ⁄or high altitudes (Lindstrom
et al. 2001). Because these herbivores typically are basal to
the ecosystem, the effect of the cycles in their abundance
reverberates across the foodweb (Ims,Henden&Killengreen
2008). The consequences of herbivore cycles are thus appar-
ent also on higher trophic levels, even when these predators
do not directly drive the cycle. This perspective is in contrast
to the classic view of predator–prey dynamics as a Lotka–
Volterra type of dynamics, where the predator drives the fluc-
tuations in the abundance of the prey and shows similar
cycles as its prey but lagging in time. A classic example of
predator-prey population dynamics where the predator’s*Correspondence author. E-mail: [email protected]
Journal of Animal Ecology 2009, 78, 1050–1062 doi: 10.1111/j.1365-2656.2009.01563.x
� 2009TheAuthors. Journal compilation� 2009 British Ecological Society

population size does not track the fluctuations in the abun-
dance of its main prey, voles, is provided by Southern’s
(1970) study of tawny owls Strix aluco Lin. in southern Eng-
land.
In Northern boreal environments in Fennoscandia, tawny
owls and other birds of prey occur in such low densities that
they do not have the potential to impose sufficient predation
pressure to make a serious impact on the vole dynamics and
hence are, themselves, not driving the cyclic fluctuations in
their main prey (Korpimaki et al. 2002; Norrdahl et al. 2004).
Resident owl species (Ural owlStrix uralensisPall. and tawny
owl) respond to fluctuations in food abundance by adjusting
their reproduction, but – once they have occupied a territory
– do not disperse to breed where there are plenty of voles as
other (semi-) nomadic species do (Andersson 1980).
By refraining from breeding when food is scarce, the pro-
portion of breeding site-tenacious owls can increase rapidly
with increasing numbers of voles, without any delay (South-
ern 1970; Brommer, Pietiainen & Kolunen 2002; see also
Korpimaki & Norrdahl 1989, 1991; Rohner 1996). On the
other hand, the mortality of territorials (and their offspring)
is drastically increased when the voles crash in abundance
every third year (Brommer et al. 2002). This recurring ‘bottle-
neck’ creates opportunities for prebreeding individuals (float-
ers; Rohner 1996) to start breeding when food abundance
increases again. As a consequence, fluctuations in food abun-
dance generate changes in the population’s age distribution,
as the proportion of young, first-breeding individuals in the
population increases when food abundance becomes more
favourable (Brommer, Pietiainen &Kolunen 1998).
For a variety of reasons, young and ⁄or inexperienced indi-
viduals may respond differently to environmental fluctua-
tions than older and experienced ones (Metcalf & Pavard
2007). The change in age structure over a cycle therefore
potentially creates marked variation in the population’s
reproductive output and survival. One powerful way to
incorporate such individual differences in performance is to
group the individuals in relevant stages. In general, such
grouping has important consequences for the understanding
of population growth and dynamics (Caswell 2001). In case
of a population living in a cyclic environment, changes in
population structure across the cycle need to be incorporated
and the consequences of a variable population structure for
reproduction and survival need to be understood when mod-
elling cyclic population dynamics.
In many places, and particularly in Fennoscandia, herbi-
vore cycles are fading out (Ims et al. 2008), which is expected
to present a major change in the environment for many other
species that are (partly) dependent on these herbivores. Avian
predators of voles are prime candidates for species likely to
be negatively affected by changes in the vole dynamics
(Hornfeldt, Hipkiss & Eklund 2005). Although the tawny
owl is a generalist predator in Northerly populations, it
almost non-exclusively uses voles as a prey when vole abun-
dance is high (Petty 1999) and it is highly dependent on voles
for reproduction (Kekkonen et al. 2008). In this study, we
determine how reproduction and survival, which together
define population growth, of different stages of tawny owls
respond to variation in food supply during 15 years of persis-
tent cycles in vole abundance. Our aim was to provide a
benchmark for understanding the cyclic tawny owl – vole sys-
tem to evaluate changes in this system when vole cycles fade
out. In particular, we aim to assess the relative importance of
variation in reproduction and survival for the dynamics of a
predator population subsisting on prey that shows periodic
fluctuations in its abundance. Previous studies have shown
that reproduction and survival vary in such an environment
(e.g. Brommer et al. 1998), but no study has – to our knowl-
edge – quantitatively compared the importance of such varia-
tion for the population dynamics. It is not obvious how
variation in reproduction can be compared with variation in
survival without formal consideration in a population
dynamical model. We therefore construct a matrix popula-
tion model based on our study of fecundity and survival to
perform an elasticity analysis, in which we quantitatively
resolve how a change in survival and fecundity rate of differ-
ent stages across the vole cycle would alter population
growth rate.
Materials andmethods
STUDY AREA
Tawny owls were studied in a study area of c. 250 km2 in southern
Finland (60�15¢ N, 24�15¢ E). Between 1975 and 1980, the study area
was established by setting up boxes. Pairs nested almost exclusively
in nest boxes, which were provided in high abundance (c. 125 were
available from 1980 onwards). Tawny owls were also ringed and con-
trolled by ornithologists in regions surrounding the study area. We
here consider data collected from the study area during 1981–1995,
when vole dynamics were cyclic.
FIELD PROTOCOL
The census and handling of all tawny owls were carried out by KA
and TK. Territorial tawny owls are rather vocal, especially prior to
the breeding season. In early spring (March–April), recordings of
hooting tawny owls were played at regular locations along roads
transecting the study area. The location where tawny owls responded
was marked on a map. Starting at the end of April, all boxes and
other possible breeding sites were checked. Considerable effort was
put into finding the nests of all tawny owls by searching for natural
nest sites and using new boxes set up by private individuals in the
approximate area where a hooting owl was recorded and where a
breeding thus was expected.We here distinguish between the number
of territories and the number of pairs that are breeding, where the
former refers to the number of sites where tawny owls responded to
the playback, and the latter to the number of pairs observed breed-
ing.
Practically all females and males were trapped when the offspring
were 1–2 weeks old. Brooding females were taken from their nest
boxes in the evening by netting them at the opening of the nest box.
After handling, the female was put back into the nest box and a
swing-door trap for the male was mounted in front of it and left over
night. In the following, morning traps were checked and the males
were handled. When the oldest chick was c. 25 days old, all offspring
in the brood were ringed with a unique aluminium ring to allow life-
Tawny owl population dynamics and the vole cycle 1051
� 2009 TheAuthors. Journal compilation� 2009 British Ecological Society, Journal of Animal Ecology, 78, 1050–1062

long individual identification. Adult birds were categorized into three
age classes at ringing )1, 2 and +2 years old – using characteristic
patterns on juvenile and adult primary and secondary feathers (Ah-
ola & Niiranen 1986). Individuals were considered as first breeders if
they were unringed when first caught, or if they had been ringed as
pulli and did not have a previous breeding record. Because consider-
able effort has been put into finding all nests and ringing all nestlings,
we considered all individuals that were unringed when first caught as
immigrants.
PREY ABUNDANCE
Spring and autumn snap trapping of small mammals was carried out
by KA and TK within the study area from 1981 onwards to estimate
the abundance of prey. Snap trapping was conducted in two localities
on each trapping event: one in the eastern part of the study area and
one in the western part of the study area. Each trapping locality con-
sists of open (field ⁄ clear-cut) habitat and forest habitat. Traps were
set as a transect of 16 trapping spots (15 m between) with three traps
each, giving a total of 48 traps per habitat (96 traps per replicate). All
traps were triggered for two consecutive nights (192 traps for two
nights = 384 trap nights in total per trapping). Not only field voles
Microtus agrestis and bank voles Clethrionomys glareolus were
caught in the traps, but also, to some extent, wood mice Apodemus
flavicollis and shrews Sorex araneus. We include all species in the
analysis of prey abundance (number of individuals caught per 100
trap nights). Based on these trap indices, we categorized years into
low, increase and decrease phases of the vole cycle (see, e.g. Brommer
et al. 2002).
L IFE-H ISTORY STAGES
We here distinguish owls according to their sex and whether they are
first breeders or have bred before (termed experienced breeders). Dif-
ferences in reproductive performance between the stages of first and
experienced breeders are expected (Metcalf & Pavard 2007). Our
main motivation for classifying individuals into first vs. experienced
breeders is that such a classification is possible in a non-arbitrary way
for any individual in a population, whereas ageing all individuals is
not always possible. For example, age classes in tawny owls older
than 2 years cannot be distinguished (Ahola & Niiranen 1986), such
that the exact ages of only a subset of individuals is known, and simi-
lar constraints apply to most avian populations. We here consider
the sex of an individual as a second-stage variable. An individual’s
sex may be particularly important in birds of prey, where males are
smaller and mainly provide food (almost exclusively so prior and
during incubation), whereas the larger females incubate and defend
the brood. Such sex differences may conceivably lead to differences
in their sensitivity to changes in the environmental conditions that
occur when food supply fluctuates. Sex differences in reproduction
and survival in birds of prey have rarely been examined (but see Altw-
egg, Schaub &Roulin 2007), mainly due to the difficulty of capturing
males.
STATIST ICAL ANALYSES
Statistical analyses were carried out using r 2.5.0 (R Development
Core Team 2007). To investigate associations between vole and owl
numbers, we assumed a saturating nonlinear response and used linear
regression with square root of the prey trapping indices in the preced-
ing autumn to explain the number of breeding individual owls. A sat-
urating response is expected and biologically motivated as follows: (i)
voles are the only available prey of tawny owls in Finland prior to
breeding (most birds migrate); and (ii) a previous study of Finnish
tawny owls has shown that breeding parameters correlate with voles
but not with other prey items (Kekkonen et al. 2008). In the other
analyses (on reproduction and survival), vole phase was entered as a
three-level categorical variable (low, increase and decrease phases).
We grouped years according to vole cycle phases rather than describ-
ing variation across years due to vole abundance (i.e. as a continuous
function of prey abundance), because the phase-based approach cap-
tures not only the present, but also the future dynamics of the vole
cycle. For example, in March–April when the breeding season for
tawny owls starts in Finland, the vole abundance is reasonably high
both in vole increase and in vole decrease phases, but the autumn
density of voles is high in the former whereas voles are almost absent
in the latter (Karell et al. 2009). Because we study five vole cycles, the
phase-based approach constitutes a replicated design (each phase
occurs five times). Hence, to explicitly explore whether such categori-
zation indeed captures an intrinsic part of the dynamics, we specifi-
cally modelled the effect of years (nested in phase) in all analyses
[denoted ‘year(phase)’ in the results]. In case phases are irrelevant to
the dynamics and variation across years is in fact the main driver,
analyses should show that phase is insignificant.
The effects of breeding experience on breeding activity and recruit
production were analysed separately for males and females, and we
thus onlymade a qualitative comparison of the sex-related differences
inbreedingperformance.Dataonboth sexeswerenot combined in the
sameanalysesbecause thiswouldrequireanalysing thedataperpair.
Survival of first and experienced breeders in relation to the vole
cycle was estimated using capture–mark–recapture (CMR) method-
ology on live encounters data (Cormack–Jolly–Seber model, CJS)
using the program MARK (White & Burnham 1999). With the CJS
model, one can separate survival probability (F) from recapture
probability (P) using a maximum likelihood approach. We used data
on individual encounters from 1981 to 1996 in order to also get an
estimate of survival for the last year of the study (1995). Individuals
were categorized into males and females to test for sex-specific differ-
ences in survival. We built a full model coding for sex and including
two experience categories (first breeder denoted by ‘first’ and experi-
enced breeder denoted by ‘exp’) and full-time dependence in survival
and recapture [F(sex · timefirst ⁄ timeexp)p(sex · timefirst ⁄ timeexp)].
To correct for overdispersion, we calculated the parameter c as the
ratio of the observed deviance of the full model over the mean devi-
ance achieved from 500 bootstrap simulations of the same model.
Time dependence was replaced by vole cycle phase to test if F and p
of the different age and sex categories were affected by the vole cycle.
Models were built by entering the vole cycle phases directly into the
Parameter IndexMatrix in programMARK as a three-class variable
coding for low, increase and decrease phase. Therefore, F and p
could be either time dependent (time), constant (c) or vole phase
dependent (phase) for each experience class (first or experienced bree-
der) and sex. Models were ranked on the basis of their quasi-likeli-
hood Akaike information criterion (QAIC) calculated as
)2 ln(L) ⁄ c + 2K + (2K(K + 1) ⁄ (n – K – 1), where L is the likeli-
hood of the model, K the number of parameters and n the effective
sample size. We used the following approach in model selection: for
‘F’ we first tested all candidate models nested under the full model
(n = 22) while keeping ‘p’ constant at ‘sex · time ⁄ time’. Models
including ‘sex’ as a factor did not receive high support (QAIC), and
therefore two-way interactions of ‘sex · experience’ and ‘sex · -
phase’ were not tested. The models with the ighest QAIC among
these 22 models (QAIC weight more than 5%, n = 5) were then
tested with all the different ‘p’ possibilities (n = 22) giving a total of
1052 P. Karell et al.
� 2009 TheAuthors. Journal compilation� 2009 British Ecological Society, Journal of Animal Ecology, 78, 1050–1062

127 models [n = 22 + (5 · 21)]. The five models with the highest
QAIC were ranked in an equal order for all the different recapture
possibilities. Because we did not find strong support for a single
model (see Results), we obtained estimates of survival and recapture
probabilities through model averaging. Model averaging calculates
an average value over all models in the candidatemodel set with com-
mon elements in the parameter structure, weighted by normalized
QAIC model weights [exp()DQAIC ⁄ 2) ⁄P
(exp()DQAIC ⁄ 2))]. Formore information on the modelling approach, see Burnham &
Anderson (1998) andCooch&White (2001).
Proportional data on population composition were analysed using
generalized linear models (GLM) with binomial errors and logit
links.We estimated reproductive success in the population as the pro-
portion of recruits per number of eggs produced in a given year. To
allow qualitative comparison between sexes recruitment probability
was analysed separately for males and females in (nested) GLMswith
binomial errors emphasizing vole cycle and experience-dependent
effects. We constructed the minimal adequate models by stepwise
backwards modelling in which we compared the models using likeli-
hood ratio tests (LRT, v2; Crawley 2002). We also compared the
models using AIC. Both the AIC and LRT produced the same mini-
mal adequate models for the data. Although our data on reproduc-
tion is individual-based, we chose to analyse it on an annual basis.
Our aim with the analysis of recruitment was to evaluate the popula-
tion-level productivity of first and experienced breeders, and hence,
to deliver a general picture of reproduction in relation to the vole
cycle as a baseline for the population dynamical model. Although
generalized linear mixed models (LMMs) can account for individual
characteristics, we chose not to use such a model because the focus is
on the population-level contribution of first and experienced breed-
ers.We thus accepted some level of pseudo-replication (120 ⁄ 258 indi-viduals are included both as a first breeder and as an experienced
breeder) as the preference of a straightforward and robust method of
analysis. We report in the results the minimal adequate models
achieved through the stepwise backwards procedure.
Age composition was based on individuals that were ringed as nes-
tlings and whose age was therefore known exactly. The data were
analysed with LMMs with ‘individual identity’ as a random effect,
because certain individuals occurred repeatedly in the analysis. We
constructed the minimal adequate model in the same manner as for
GLMs by stepwise backwards modelling in which we compared
LMMs solved under maximum likelihood using LRT between mod-
els. In an LRT, )2 times the difference in the likelihood of a model
with and without an effect was tested as a chi-squared value with the
effect’s degrees of freedom (Pinheiro & Bates 2000). The significance
of the random effect was also based on an LRT. Significance of fixed
effects in LMMswas based on F-tests.
Results
Atotal of 351 clutchesweremonitored in the studypopulation
between1981and1995.Oftheseclutches,300producedhatch-
lings and 274 succeeded to fledge offspring. In 294 breeding
attempts, the femaleparentswere caughtand identifiedand, in
278cases,also themaleparentswerecaughtandidentified.
PAIR FORMATION
Pairs were formed assortatively with respect to breeding
experience as most (68%, 125 of 184) pair formations were
between either two inexperienced breeders or two experi-
enced breeders (Table 1). After first pair formation mate
change was rare: in total 32 of 122 (26%)males changedmate
during the study, of which 13 changed more than once [in
total 51 mate changes of which 43 changes (84%) was due to
death of the partner]. Even fewer females (29 ⁄ 129, 16%)
changed mate, with only six females changing more than
once (in total 37 mate changes of which 28 changes (76%)
was due to death of the partner).
TERRITORY OCCUPANCY AND BREEDING ACTIV ITY IN
RELATION TO THE VOLE CYCLE
Territories were considered occupied when an owl responded
to play-back calls prior to the breeding season. Once estab-
lished on a territory, most owls remained faithful to their ter-
ritory. Only 23 ⁄ 122 (19%) males and 29 ⁄ 136 (21%) females
changed territory during the study. Territory change
occurred after the first breeding event in 15 ⁄ 23 cases (65%)
for males and in 19 ⁄ 29 (66%) of the cases for females.
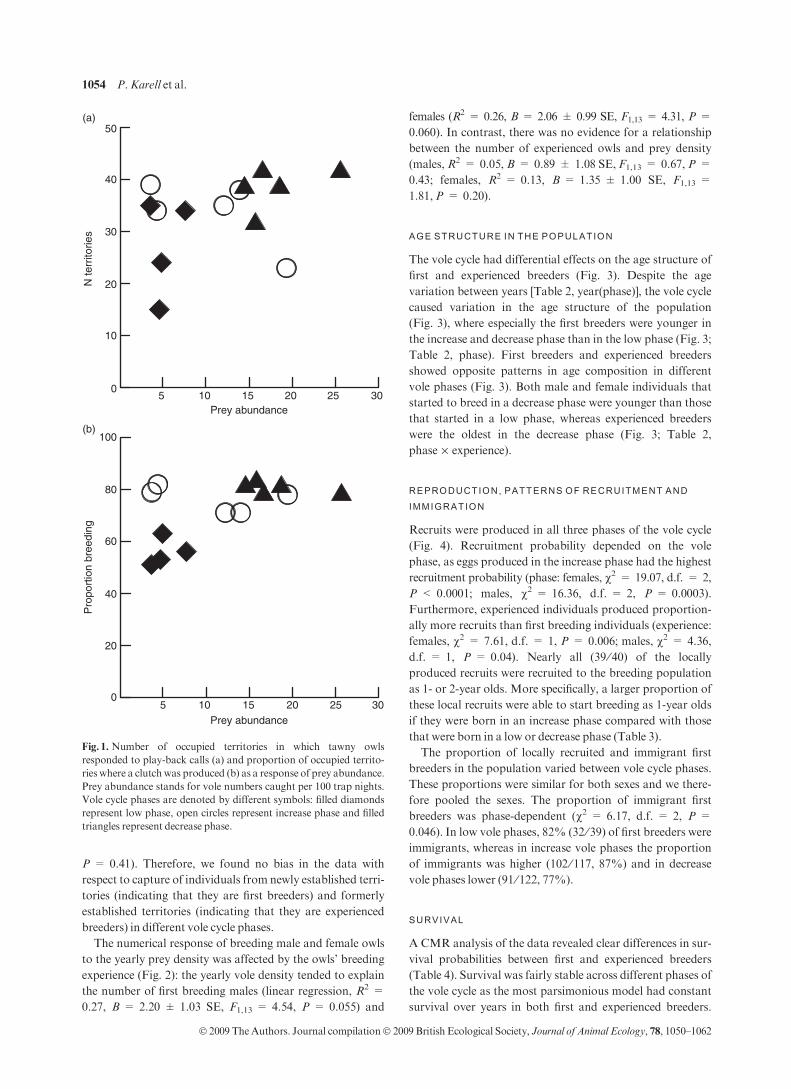
There was variation in the number of occupied territories
between vole cycle phases (Fig. 1a): in low phases 28 ± 4 ter-
ritories were occupied and the number increased to 34 ± 3
territories in increase phases and further to 39 ± 2 territories
in decrease phases [anova: phase, F2,12 = 3.76, P = 0.05;
decrease phase vs. low phase, t12 = 2.74, P = 0.02; year
(phase) dropped]. This difference in territory occupancy
between years was less clear when prey abundance in the pre-
ceding autumn was entered as an explanatory variable
(Fig. 1a; linear regression: R2 = 0.14, B = 2.70 ± 1.96,
F1,12 = 1.89,P = 0.19).
The proportion of occupied territories where the residing
owl pair attempted to breed increased as vole abundance
increased (Fig. 1b; logistic regression, B = 0.27 ± 0.10,
v2 = 7.39, d.f. = 1, P = 0.007). Also when categorizing
vole abundance into vole cycle phases, a substantially higher
proportion of territorial pairs bred in the increase and
decrease phase years than in the low phase years [Fig. 1b;
GLM: phase, v2 = 9.64, d.f. = 2, P < 0.0001; year(phase)
dropped].
Some nests failed at an early stage before parents could be
caught and identified (16.2%, 57 ⁄ 351). There was no differ-
ence in the proportion of failed nests between vole cycle
phases (v2 = 1.09, d.f. = 2, P = 0.58). Among the failed
nests, 48% (31 ⁄ 65) were in newly established territories
and this proportion of failed newly established territories
did not vary between vole cycle phases (v2 = 1.80, d.f. = 2,
Table 1. Pair formation in the tawny owl population
##
$$
First
breeder (%)
Experienced
breeder (%)
First breeder 96 (52) 26 (14)
Experienced breeder 33 (18) 29 (16)
There were in total 184 new pair formations of whichmost were
between two first breeders.
Tawny owl population dynamics and the vole cycle 1053
� 2009 TheAuthors. Journal compilation� 2009 British Ecological Society, Journal of Animal Ecology, 78, 1050–1062
P = 0.41). Therefore, we found no bias in the data with
respect to capture of individuals from newly established terri-
tories (indicating that they are first breeders) and formerly
established territories (indicating that they are experienced
breeders) in different vole cycle phases.
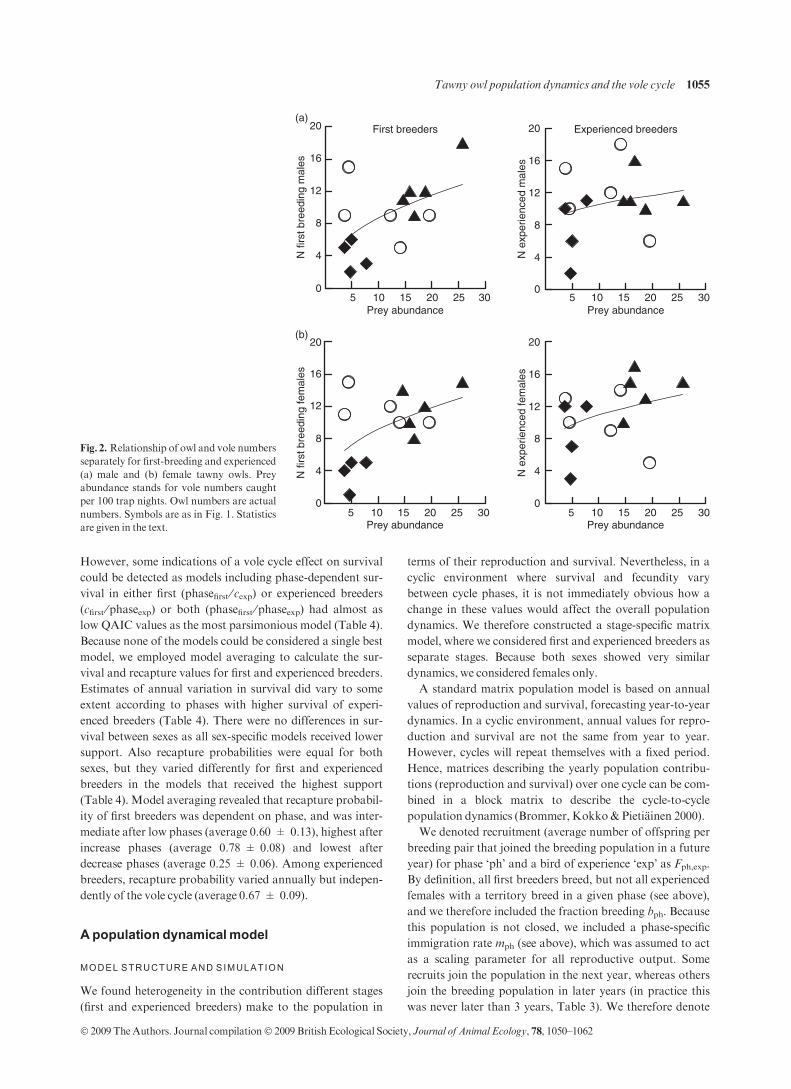
The numerical response of breeding male and female owls
to the yearly prey density was affected by the owls’ breeding
experience (Fig. 2): the yearly vole density tended to explain
the number of first breeding males (linear regression, R2 =
0.27, B = 2.20 ± 1.03 SE, F1,13 = 4.54, P = 0.055) and
females (R2 = 0.26, B = 2.06 ± 0.99 SE, F1,13 = 4.31, P =
0.060). In contrast, there was no evidence for a relationship
between the number of experienced owls and prey density
(males,R2 = 0.05, B = 0.89 ± 1.08 SE, F1,13 = 0.67, P =
0.43; females, R2 = 0.13, B = 1.35 ± 1.00 SE, F1,13 =
1.81,P = 0.20).
AGE STRUCTURE IN THE POPULATION
The vole cycle had differential effects on the age structure of
first and experienced breeders (Fig. 3). Despite the age
variation between years [Table 2, year(phase)], the vole cycle
caused variation in the age structure of the population
(Fig. 3), where especially the first breeders were younger in
the increase and decrease phase than in the low phase (Fig. 3;
Table 2, phase). First breeders and experienced breeders
showed opposite patterns in age composition in different
vole phases (Fig. 3). Both male and female individuals that
started to breed in a decrease phase were younger than those
that started in a low phase, whereas experienced breeders
were the oldest in the decrease phase (Fig. 3; Table 2,
phase · experience).
REPRODUCTION, PATTERNS OF RECRUITMENT AND
IMMIGRATION
Recruits were produced in all three phases of the vole cycle
(Fig. 4). Recruitment probability depended on the vole
phase, as eggs produced in the increase phase had the highest
recruitment probability (phase: females, v2 = 19.07, d.f. = 2,
P < 0.0001; males, v2 = 16.36, d.f. = 2, P = 0.0003).
Furthermore, experienced individuals produced proportion-
ally more recruits than first breeding individuals (experience:
females, v2 = 7.61, d.f. = 1, P = 0.006; males, v2 = 4.36,
d.f. = 1, P = 0.04). Nearly all (39 ⁄ 40) of the locally
produced recruits were recruited to the breeding population
as 1- or 2-year olds. More specifically, a larger proportion of
these local recruits were able to start breeding as 1-year olds
if they were born in an increase phase compared with those
that were born in a low or decrease phase (Table 3).
The proportion of locally recruited and immigrant first
breeders in the population varied between vole cycle phases.
These proportions were similar for both sexes and we there-
fore pooled the sexes. The proportion of immigrant first
breeders was phase-dependent (v2 = 6.17, d.f. = 2, P =
0.046). In low vole phases, 82% (32 ⁄ 39) of first breeders wereimmigrants, whereas in increase vole phases the proportion
of immigrants was higher (102 ⁄ 117, 87%) and in decrease
vole phases lower (91 ⁄ 122, 77%).
SURVIVAL
A CMR analysis of the data revealed clear differences in sur-
vival probabilities between first and experienced breeders
(Table 4). Survival was fairly stable across different phases of
the vole cycle as the most parsimonious model had constant
survival over years in both first and experienced breeders.
Prey abundance
0
10
20
30
40
50
N te
rrito
ries
5 10 15 20 25 30
5 10 15 20 25 30
Prey abundance
0
20
40
60
80
100
Pro
port
ion
bree
ding
(a)
(b)
Fig. 1. Number of occupied territories in which tawny owls
responded to play-back calls (a) and proportion of occupied territo-
ries where a clutchwas produced (b) as a response of prey abundance.
Prey abundance stands for vole numbers caught per 100 trap nights.
Vole cycle phases are denoted by different symbols: filled diamonds
represent low phase, open circles represent increase phase and filled
triangles represent decrease phase.
1054 P. Karell et al.
� 2009 TheAuthors. Journal compilation� 2009 British Ecological Society, Journal of Animal Ecology, 78, 1050–1062
However, some indications of a vole cycle effect on survival
could be detected as models including phase-dependent sur-
vival in either first (phasefirst ⁄ cexp) or experienced breeders
(cfirst ⁄phaseexp) or both (phasefirst ⁄phaseexp) had almost as
low QAIC values as the most parsimonious model (Table 4).
Because none of the models could be considered a single best
model, we employed model averaging to calculate the sur-
vival and recapture values for first and experienced breeders.
Estimates of annual variation in survival did vary to some
extent according to phases with higher survival of experi-
enced breeders (Table 4). There were no differences in sur-
vival between sexes as all sex-specific models received lower
support. Also recapture probabilities were equal for both
sexes, but they varied differently for first and experienced
breeders in the models that received the highest support
(Table 4). Model averaging revealed that recapture probabil-
ity of first breeders was dependent on phase, and was inter-
mediate after low phases (average 0.60 ± 0.13), highest after
increase phases (average 0.78 ± 0.08) and lowest after
decrease phases (average 0.25 ± 0.06). Among experienced
breeders, recapture probability varied annually but indepen-
dently of the vole cycle (average 0.67 ± 0.09).
Apopulation dynamical model
MODEL STRUCTURE AND SIMULATION
We found heterogeneity in the contribution different stages
(first and experienced breeders) make to the population in
terms of their reproduction and survival. Nevertheless, in a
cyclic environment where survival and fecundity vary
between cycle phases, it is not immediately obvious how a
change in these values would affect the overall population
dynamics. We therefore constructed a stage-specific matrix
model, where we considered first and experienced breeders as
separate stages. Because both sexes showed very similar
dynamics, we considered females only.
A standard matrix population model is based on annual
values of reproduction and survival, forecasting year-to-year
dynamics. In a cyclic environment, annual values for repro-
duction and survival are not the same from year to year.
However, cycles will repeat themselves with a fixed period.
Hence, matrices describing the yearly population contribu-
tions (reproduction and survival) over one cycle can be com-
bined in a block matrix to describe the cycle-to-cycle
population dynamics (Brommer, Kokko& Pietiainen 2000).
We denoted recruitment (average number of offspring per
breeding pair that joined the breeding population in a future
year) for phase ‘ph’ and a bird of experience ‘exp’ as Fph,exp.
By definition, all first breeders breed, but not all experienced
females with a territory breed in a given phase (see above),
and we therefore included the fraction breeding bph. Because
this population is not closed, we included a phase-specific
immigration rate mph (see above), which was assumed to act
as a scaling parameter for all reproductive output. Some
recruits join the population in the next year, whereas others
join the breeding population in later years (in practice this
was never later than 3 years, Table 3). We therefore denote
Prey abundance
0
4
8
12
16
20
N e
xper
ienc
ed m
ales
Experienced breeders
Prey abundance
0
4
8
12
16
20
N fi
rst b
reed
ing
mal
es
First breeders
5 10 15 20 25 30
5 10 15 20 25 30 5 10 15 20 25 30
5 10 15 20 25 300
4
8
12
16
20
N e
xper
ienc
ed fe
mal
esPrey abundance Prey abundance
0
4
8
12
16
20
N fi
rst b
reed
ing
fem
ales
(a)
(b)
Fig. 2. Relationship of owl and vole numbers
separately for first-breeding and experienced
(a) male and (b) female tawny owls. Prey
abundance stands for vole numbers caught
per 100 trap nights. Owl numbers are actual
numbers. Symbols are as in Fig. 1. Statistics
are given in the text.
Tawny owl population dynamics and the vole cycle 1055
� 2009 TheAuthors. Journal compilation� 2009 British Ecological Society, Journal of Animal Ecology, 78, 1050–1062

the fraction of recruits born in phase ‘ph’ and recruiting ‘y’
(1, 2 or 3) years later as rph,y.
Survival for phase ‘ph’ to the next phase for a bird of expe-
rience ‘exp’ was denoted by Pph,exp. The cycle-to-cycle popu-
lation dynamics of a three-phase cycle are then given by the
blockmatrix
We parameterized the model with the observed data. The
fractions (r) of offspring that were born in a low, increase or
decrease phase and recruited 1, 2 or 3 years later are given in
Table 3. The values for the reproductive parametersm, b and
F and their rationale are outlined in Table 5. Stage- and
phase-specific survival values are reported in Table 4.
The dominant eigenvalue k1 of B [k1(B)] denotes the
change in population size (i.e. the number of females with a
territory) across cycles. Elasticity analysis is a prospective
analysis in which one can explore how much k would change
in response to changes in reproduction and survival (Caswell
2000). Elasticity can be calculated for each stage ⁄phase com-
bination by using standard methods (Caswell 2001) to calcu-
late the elasticities of B. Each elasticity value gives a
proportional contribution to k1(B) as the sum of elasticities
always equals 1. In a stable cyclic population k1(B) = 1. The
right eigenvector of B that corresponds with the dominant
eigenvalue [w1(B)] gives the stable stage distribution, i.e. the
proportion of first and experienced breeders and non-breed-
ers in each phase during a complete cycle. Again, this vector
can be calculated using standardmethods (Caswell 2001).
The elasticities of matrix B indicate the proportional con-
tribution of each matrix element to population dynamics.
The sum of all elasticities equals 1 by definition, and the elas-
ticity values therefore provide a ranking of importance. This
procedure models both demographic stochasticity and uncer-
tainty in the estimates of fecundity and survival. To gain
insight into the variability of our findings with respect to sto-
chasticity in fecundity and survival, we calculated k1(B) andthe elasticities for matrixB based on 10 000 simulations vary-
ing Fph,exp and Pph,exp. We treated all other parameters as
constants as we have no prior expectation of their uncer-
tainty. Hence, the variability addressed by our simulations is
conservative.
Simulated reproductive output was the average of a ran-
domly drawn Poisson distribution over the number of phase-
and stage-specific territorieswithmeanFph,exp (Table5 forval-
ues).We used the estimated error of the CMR survival values
(Table 4) to vary the survival values. The logit of the simulated
survival value was logit �Pph;exp
� �þN 0; r2 �Pph;exp
� �� �, where
�Pph;exp is the estimated mean survival (Table 5), and
N 0; r2 �Pph;exp
� �� �is a random draw of a normal distribution
with zero mean and variance r2ð �Pph;expÞ (i.e. the square of thestandard error inTable 5).For eachof the simulations,we cal-
culated thedominant eigenvalueand the elasticities.
We used the simulated dominant eigenvalues to calculate
the 95% confidence interval (CI) around k1(B), and tested
whether the elasticities showed a significant difference using
the distribution of the simulated elasticities. Pairwise com-
parisons were made between all simulated fecundity elastici-
ties and between all simulated survival elasticities. We first
calculated the proportion p of the differences between the
10 000 simulated values that was either smaller or larger than
zero (whichever was smallest). This gives a pairwise test value
(e.g. Manly 1997) for whether two elasticities are equal after
taking into account demographic uncertainty in the estimates
of fecundity and survival. Overall significance of the multiple
comparison of all pairwise P-values was based on the Holm–
Bonferroni method (Holm 1979) retaining the overall signifi-
cance level a = 0.05. The elasticity values can then be
grouped into groups that differ from each other significantly.
POPULATION DYNAMICS: MODEL RESULTS
The dominant eigenvalue of the population dynamical model
was0.981 (95%CI:0.867–1.100).Weconsidered thisa reason-
ablereflectionoftheobserveddynamics,becausetheCIclearly
overlaps with 1 (stable cycle-to-cycle population dynamics).
The expected stage ⁄phase distribution (given by the right
eigenvector) produced a satisfactory fit (v2 = 9.60, d.f. = 5,
P = 0.09) to the observed distribution (Fig. 5b), although the
model predicted a relatively large proportion of first breeders
in the increase phase. In the field data, the number of experi-
encedbreeders stayedfairlyconstantacrossphases (Figs5b,c,f
and 2). The matrix model showed that the survival of experi-
enced breeders was more important for population growth
than that of inexperienced breeders and that the main differ-
ence stemmed from the survival elasticity of an experienced
breeder in the low phase (Fig. 5a). The elasticity values of
reproduction showed only small differences, with none of the
non-zeroelasticitiesdifferingsignificantly (Fig.5a).
Discussion
NUMERICAL RESPONSE OF TAWNY OWLS
We have analysed 15 years (five 3-year vole cycles) of data on
both sexes of tawny owls and their small mammal prey in a
B ¼
rlow;3mlowFlow;f rlow;3mlowblowFlow;e rinc;2mlowFinc;f rinc;2mlowbincFinc;e rdec;1mlowFdec;f rdec;1mlowbdecFdec;e
0 0 0 0 Pdec;f Pdec;e
rlow;1mincFlow;f rlow;1mincblowFlow;e rinc;3mincFinc;f rinc;3mincbincFinc;e rdec;2mincFdec;f rdec;2mincbdecFdec;e
Plow;f Plow;e 0 0 0 0rlow;2mdecFlow;f rlow;2mdecblowFlow;e rinc;1mdecFinc;f rinc;1mdecbincFinc;f rdec;3mdecFdec;f rdec;3mdecbdecFdec;e
0 0 Pinc;f Pinc;e 0 0
26666664
37777775:
1056 P. Karell et al.
� 2009 TheAuthors. Journal compilation� 2009 British Ecological Society, Journal of Animal Ecology, 78, 1050–1062

population in southern Finland. Based on responses to play-
back, the number of active territories has been censused in
this population, allowing us to investigate the response of the
total population size to fluctuations in vole abundance. We
find that territorial activity (potential breeders) in the tawny
owl population in southern Finland varies between vole cycle
phases in response to fluctuations in the abundance of their
main prey. Thus, our result differs from studies of tawny owls
in temperate regions without multi-annual fluctuations in the
abundance of their main prey. In particular, territory num-
bers in temperate regions are stable between years (Southern
1970; Hirons 1985; Jedrzejewski et al. 1996; Sunde & Bølstad
2004; Desfor, Boomsma& Sunde 2007).We find that the var-
iation in territory number between vole cycle phases is not
driven by extensive mortality after the decrease phase as in
the closely related Ural owl living in the same environment
(Brommer et al. 1998, 2002), because tawny owl survival is
fairly stable between vole phases. Instead, recruitment of new
breeders to the population (including immigration) drives the
variation in territory numbers between vole phases and con-
stitutes the main part of the observed numerical response of
the species. Because first breeders survive less well than expe-
rienced breeders, they are less important for population
growth, and a population dynamical model emphasizes that
the survival of experienced breeders is most important for
population growth above other components.
STAGE-DEPENDENT REPRODUCTION AND SURVIVAL
A numerical response of the number of breeding pairs to the
abundance of their main prey has been documented in several
owl studies (Tengmalm’s owl Aegolius funereus and voles:
Korpimaki & Norrdahl 1991; barn owl Tyto alba and voles:
Taylor 1994; tawny owl and voles: Jedrzejewski et al. 1996;
Ural owl and voles: Brommer et al. 2002; Northern spotted
owl Strix occidentalis caurina and deer mice: Rosenberg,
Swindle & Anthony 2003). These studies have, however, not
accounted for any sex- or stage-related differences. We find
that fluctuating prey abundance has distinctive consequences
for the age composition of first breeding and experienced
individuals. In particular, the majority of tawny owl off-
spring hatched in an increase phase starts to breed already as
1-year olds in the following decrease phase when conditions
(at least at the onset of reproduction) are favourable (see also
Brommer et al. 1998). This flexible and potentially young age
at first breeding allows for a rapid tracking of the vole cycle.
As a further consequence, one stage in the population (first
breeders) becomes younger as voles increase in abundance,
whereas already established territorial individuals get older
when voles become more abundant. Hence, over the course
of a cycle, the structure of the population changes, in terms
of both the proportion of first vs. experienced breeders and
their age distributions.
Bird populations consist of two sexes, but most studies on
birds of prey consider one sex only. In birds of prey, males
and females are sexually dimorphic in size and havemarkedly
different parental roles. Males and females may therefore
0
2
4
6
8
14
17
17
28
28
26
Experienced breeders
Low Incr Decr
Low Incr Decr
0
2
4
6
8A
ge in
yea
rsA
ge in
yea
rs
9
6
22 17
27
27
First breeders
Fig. 3.Mean age (±SE) of first-breeding and experienced females
(open circles) and males (filled diamonds) in different phases of the
vole cycle. Included are individuals ringed as nestlings, which were
born either within or outside the study area, but which bred within
the study area are included. See Table 2 for statistics.
Tawny owl population dynamics and the vole cycle 1057
� 2009 TheAuthors. Journal compilation� 2009 British Ecological Society, Journal of Animal Ecology, 78, 1050–1062

respond differently to variations in prey availability. The
smaller males are the main hunters during courtship, prior
and during incubation, and they do most of the hunting dur-
ing the nestling stage. The larger female incubates and con-
centrates on defending the brood (e.g. Lundberg 1986;
Wallin 1987; Newton 1989; Meijer, Daan & Hall 1990; Sun-
de, Bølstad &Moller 2003). However, in spite of these differ-
ent sex roles, both males and females largely respond
similarly to variations in prey availability in terms of their
numerical response, reproduction and survival. Our results
thus correspond with those of Kruger (2002), who found that
the lifetime reproductive success of male and female common
buzzardButeo buteowere affected by similar factors.
We find strong contrasts between first and experienced
breeders also in terms of their reproduction. First-breeding
tawny owls have a much lower production of recruits in all
phases of the vole cycle. The future prospects of tawny owl
offspring thus depend to a large extent on whether their par-
ents have reproduced before or not. Again, this effect may be
due to the parents gaining experience (individually or
together, as most pairs breeding the first time consist of two
inexperienced parents and parents rarely divorce). Alterna-
tively, poor parents are selected out of the population after
their first breeding attempt. In addition to parental experi-
ence, the vole cycle itself creates different future prospects for
the offspring. Over 80% of the local recruits produced in the
increase phase are able to start breeding as 1-year olds,
whereas the majority of recruits born in low or decrease
phases start breeding as 2-year olds. This aspect of tawny owl
reproduction is similar to Tengmalm’s owls (Korpimaki
1988, 1992) and Ural owls (Brommer et al. 1998). Intrigu-
ingly, experienced tawny owls are rather successful in pro-
ducing recruits even in a low phase as recruitment probability
of offspring hatched in a low phase is almost equal to that of
offspring hatched in the decrease phase. This latter finding is
in contrast to Tengmalm’s owls, which rarely breed during
low phases (Korpimaki & Norrdahl 1989; Laaksonen, Kor-
pimaki & Hakkarainen 2002), and is also in contrast to Ural
owls, which do breed in low numbers, but rarely produce
recruits in low phases (Brommer et al. 1998). This difference
is likely caused by the fact that tawny owl reproduction is less
dependent on small voles than the reproduction of the other
two owl species. In particular, water voles Arvicola terrestris,
which do not follow the 3-year vole cycle, are an important
prey item affecting tawny owl reproduction in Finland (Ke-
kkonen et al. 2008).
We find only marginal effects of the vole cycle on tawny
owl survival. This is in contrast to findings in Finnish Ural
owls, where CMR analysis revealed a reduced survival prob-
ability after a decrease phase (Brommer et al. 2002). Simi-
larly, in a recapture and recovery analysis of long-term
Table 2. Minimal adequate model of factors that explain the
variation in age in the tawny owl population
Variable F d.f. P
Fixed effects
Phase 28.24 2, 122 <0.0001
Experience 3059.32 1, 122 <0.0001
Year (phase) 856.90 3, 122 <0.0001
Phase · experience 21.73 2, 122 <0.0001
Variance (95%CI) % v2 P
Random effect
Individual 0.98 (0.84–1.13) 99.4 268.29 <0.0001
The results are from a linear mixed effects model with normal errors
and individual ID as the random effect. The significance of the ran-
dom effect is tested with a likelihood ratio test (v2).
Low Incr Decr
Low Incr Decr
0
0·05
0·1
0·15
0·2
Pro
port
ion
recr
uite
d
54
178
231
35
240
233
First breeders
0
0·05
0·1
0·15
0·2
Pro
port
ion
recr
uite
d
108
252203
107
250291
Experienced breeders
Fig. 4. Proportion of offspring that recruited to the breeding popula-
tion in low, increase and decrease vole phases produced by first
breeders and experienced breeders. Error bars (mean ± SE,
n = 5 years for each phase) are clustered by the sex of parent (open
circles: females; filled diamonds: males). Total number of eggs pro-
duced is given above bars.
1058 P. Karell et al.
� 2009 TheAuthors. Journal compilation� 2009 British Ecological Society, Journal of Animal Ecology, 78, 1050–1062

nationwide data on Finnish tawny owls, Francis & Saurola
(2004) found that most (more than 50%) of the variation in
adult survival is explained by winter temperature, with only
9% of variation explained by the vole cycle. In addition to
winter temperature, which increases energy expenditure in
owls (Mosher & Henny 1976), also the depth of the snow
cover may affect mortality in tawny owls. A deep snow cover
makes vole prey less accessible during winter. For example,
in Swiss barn owls Tyto alba, the number of days with snow
cover during winter explained substantial variation in annual
survival (Altwegg et al. 2003, 2006). We find that first-breed-
ing tawny owls of both sexes have lower survival than experi-
enced individuals. It is therefore possible that, in Northern
tawny owl populations such as the one studied here, winter
severity has prominent effects on survival, especially in first
breeders. We do not have sufficient data to distinguish
whether the higher survival of experienced breeders is due to
their older age or because they have gained more experience
than first breeders. In addition, experienced birds represent,
by definition, a subset of owls that have already undergone
survival selection as a first breeder and may therefore,
because of intrinsic or extrinsic factors, enjoy a higher sur-
vival than the unselected group of first breeders. Whatever
the reason for the observed differences in survival, our pri-
mary interest here is to understand its consequences for pop-
ulation dynamics.
Our CMR analysis also revealed that recapture probability
varied between vole cycle phases for first breeders whereas it
varied annually for experienced breeders. Previously recap-
ture rate has been found to vary in tawny owls due to individ-
ual characteristics (Roulin et al. 2003). We find here that
recapture rate of first breeders is the lowest in a low vole
phase. This is unlikely to be due to movement out of the pop-
ulation as we find that both first and experienced breeders are
highly site tenacious. Instead, we find it most likely that these
individuals refrain from breeding under such poor food con-
ditions (see also Southern 1970; Brommer et al. 2002) and
not that theymove out of the population, because we find sig-
nificantly lower breeding activity in the population in low
phases compared with other vole phases. Therefore, we sug-
gest that refraining from breeding is more common after the
first breeding event than at later stages and explains the vole
cycle effect on recapture rate of first breeders.
THE VOLE CYCLE AND TAWNY OWL POPULATION
DYNAMICS
Individuals of different stages of a population are often
affected differently in an environment where food availability
fluctuates annually or in a cyclic manner (Pietiainen 1988;
Ratcliffe, Furness & Hamer 1998; Cam & Monnat 2000;
Kruger & Lindstrom 2001; Laaksonen et al. 2002). To draw
conclusions on the persistence of a population, it is therefore
crucial to evaluate how these environmental fluctuations
affect reproduction and survival of different stages, and how
Table 3. Age at recruitment of offspring born in different phases of
the vole cycle
Phase at birth (%)
Low Increase Decrease
Age at recruitment
1 1 ⁄ 3 (33) 24 ⁄ 28 (85.7) 3 ⁄ 9 (33)2 2 ⁄ 3 (67) 3 ⁄ 28 (10.7) 6 ⁄ 9 (67)3 – 1 ⁄ 28 (3.6) –
Included are local recruits that were born in the study area between
1981 and 1995 (n = 40, 21males and 19 females). Data from both
sexes are pooled as there was no difference in age at recruitment
between sexes (Fisher’s exact test,P = 0.59). The age distribution of
recruits differ between phases of the vole cycle (Fisher’s exact test,
P = 0.002).
Table 4. Survival and recapture probabilities of tawny owl males and females between 1981 and 1995
Model QAIC QAICweight N par Likelihood
F(cfirst ⁄ cexp)p(phasefirst ⁄ timeexp) 993.3 0.344 19 356.60
F(cfirst ⁄ phaseexp)p(phasefirst ⁄ timeexp) 994.0 0.246 21 352.97
F(phasefirst ⁄ cexp)p(phasefirst ⁄ timeexp) 994.6 0.181 21 353.58
F(phasefirst ⁄ phaseexp)p(phasefirst ⁄ timeexp) 995.4 0.123 23 350.02
First breeder F Experienced breeder F
Model averaging
Low 0.56 ± 0.07 (0.427–0.688) 0.73 ± 0.05 (0.620–0.819)
Increase 0.55 ± 0.06 (0.437–0.635) 0.78 ± 0.07 (0.598–0.896)
Decrease 0.60 ± 0.08 (0.440–0.739) 0.71 ± 0.06 (0.578–0.823)
Themodels separate between survival (F) and recapture (p) and have two age categories (reported as ‘first breeder ⁄ experienced breeder’) thatcan be either constant (c), phase dependent (phase) or time dependent (time).We tested all possible model permutations for both experience
categories, and for both sexes including their interactions. Shown here are the four best models as judged by their QAIC values, which together
provide 89.4% support for the data (models including sex-specific effects received little support). Statistics given are quasi-likelihoodAkaike
information criterion (QAIC; the AIC corrected for overdispersion, c = 1.15), proportional support for themodel (QAICweight), number of
parameters (N par) andmodel likelihood. The lower part of the table shows the estimates of survival (F)±SE (95%CI) achieved by averaging
the estimates over all candidate models.
Tawny owl population dynamics and the vole cycle 1059
� 2009 TheAuthors. Journal compilation� 2009 British Ecological Society, Journal of Animal Ecology, 78, 1050–1062
these differences translate into population dynamics (Caswell
2001). We have here taken a pragmatic view, and rather than
considering age-dependent performance (on which we have
incomplete information), we distinguish stages according to
their breeding experience (first time vs. experienced breeders).
Our cycle-to-cycle population dynamical matrix model,
which was parameterized with stage- and phase-specific esti-
mates of reproduction, survival and immigration, produced a
reasonable estimate of population growth and the numerical
distribution of the two stages across the vole cycle’s phases.
First breeders have a strong numerical response as prey
abundance increases, whereas the number of territories of
experienced breeders remain fairly stable between vole cycle
phases. Hence, our population dynamical model captures the
essentials of the tawny owl population dynamics.
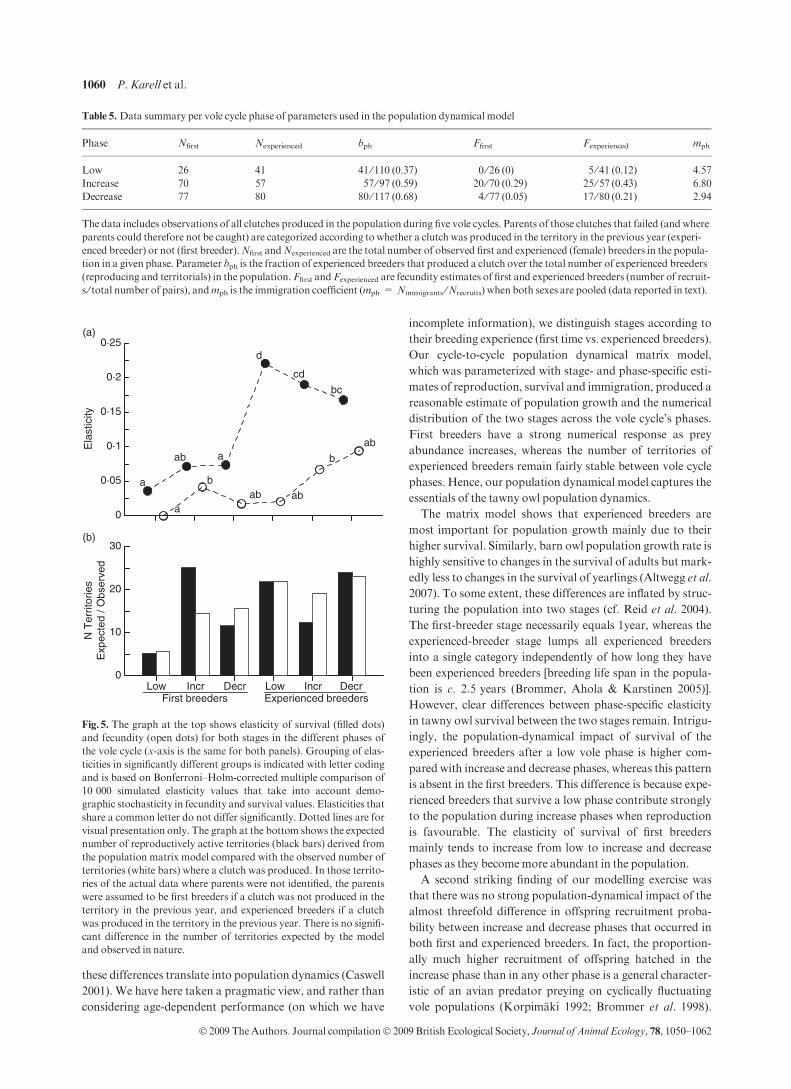
The matrix model shows that experienced breeders are
most important for population growth mainly due to their
higher survival. Similarly, barn owl population growth rate is
highly sensitive to changes in the survival of adults but mark-
edly less to changes in the survival of yearlings (Altwegg et al.
2007). To some extent, these differences are inflated by struc-
turing the population into two stages (cf. Reid et al. 2004).
The first-breeder stage necessarily equals 1year, whereas the
experienced-breeder stage lumps all experienced breeders
into a single category independently of how long they have
been experienced breeders [breeding life span in the popula-
tion is c. 2.5 years (Brommer, Ahola & Karstinen 2005)].
However, clear differences between phase-specific elasticity
in tawny owl survival between the two stages remain. Intrigu-
ingly, the population-dynamical impact of survival of the
experienced breeders after a low vole phase is higher com-
pared with increase and decrease phases, whereas this pattern
is absent in the first breeders. This difference is because expe-
rienced breeders that survive a low phase contribute strongly
to the population during increase phases when reproduction
is favourable. The elasticity of survival of first breeders
mainly tends to increase from low to increase and decrease
phases as they becomemore abundant in the population.
A second striking finding of our modelling exercise was
that there was no strong population-dynamical impact of the
almost threefold difference in offspring recruitment proba-
bility between increase and decrease phases that occurred in
both first and experienced breeders. In fact, the proportion-
ally much higher recruitment of offspring hatched in the
increase phase than in any other phase is a general character-
istic of an avian predator preying on cyclically fluctuating
vole populations (Korpimaki 1992; Brommer et al. 1998).
Table 5. Data summary per vole cycle phase of parameters used in the population dynamical model
Phase Nfirst Nexperienced bph Ffirst Fexperienced mph
Low 26 41 41 ⁄ 110 (0.37) 0 ⁄ 26 (0) 5 ⁄ 41 (0.12) 4.57
Increase 70 57 57 ⁄ 97 (0.59) 20 ⁄ 70 (0.29) 25 ⁄ 57 (0.43) 6.80
Decrease 77 80 80 ⁄ 117 (0.68) 4 ⁄ 77 (0.05) 17 ⁄ 80 (0.21) 2.94
The data includes observations of all clutches produced in the population during five vole cycles. Parents of those clutches that failed (andwhere
parents could therefore not be caught) are categorized according to whether a clutch was produced in the territory in the previous year (experi-
enced breeder) or not (first breeder).Nfirst andNexperienced are the total number of observed first and experienced (female) breeders in the popula-
tion in a given phase. Parameter bph is the fraction of experienced breeders that produced a clutch over the total number of experienced breeders
(reproducing and territorials) in the population. Ffirst and Fexperienced are fecundity estimates of first and experienced breeders (number of recruit-
s ⁄ total number of pairs), andmph is the immigration coefficient (mph = Nimmigrants ⁄Nrecruits) when both sexes are pooled (data reported in text).
Low Incr Decr Low Incr DecrFirst breeders Experienced breeders
0
10
20
30
N T
errit
orie
sE
xpec
ted
/ Obs
erve
d
0
0·05
0·1
0·15
0·2
0·25
Ela
stic
ity
a
ab a
d
cd
bc
a
bab ab
b
ab
(a)
(b)
Fig. 5. The graph at the top shows elasticity of survival (filled dots)
and fecundity (open dots) for both stages in the different phases of
the vole cycle (x-axis is the same for both panels). Grouping of elas-
ticities in significantly different groups is indicated with letter coding
and is based on Bonferroni–Holm-corrected multiple comparison of
10 000 simulated elasticity values that take into account demo-
graphic stochasticity in fecundity and survival values. Elasticities that
share a common letter do not differ significantly. Dotted lines are for
visual presentation only. The graph at the bottom shows the expected
number of reproductively active territories (black bars) derived from
the population matrix model compared with the observed number of
territories (white bars) where a clutch was produced. In those territo-
ries of the actual data where parents were not identified, the parents
were assumed to be first breeders if a clutch was not produced in the
territory in the previous year, and experienced breeders if a clutch
was produced in the territory in the previous year. There is no signifi-
cant difference in the number of territories expected by the model
and observed in nature.
1060 P. Karell et al.
� 2009 TheAuthors. Journal compilation� 2009 British Ecological Society, Journal of Animal Ecology, 78, 1050–1062

Increase vole phases have therefore been considered to be of
major importance for population persistence and viability.
For example, offspring hatched in the increase phase starts
to breed at an earlier age (this study; Brommer et al. 1998;
Laaksonen et al. 2002). However, no study has – to our
knowledge – investigated the actual population dynamical
impact of this difference in recruitment probability by inte-
grating fecundity and survival differences in a proper model.
We find that the elasticity of reproduction in the increase
phase was not any higher than the value for the decrease
phase in both first and experienced breeders. Hence, this
finding shows that explicit modelling of the dynamics can
provide non-intuitive insights.
Conclusions
Experience-dependent reproductive performance and sur-
vival are key aspects to be taken into account whenmodelling
population dynamics in variable environments. This is
because changes in food supply will have a clear impact on
the proportion of first breeders in the population. Whenever
first breeders have a markedly lower fecundity and survival
than experienced breeders, such changes in the composition
of the population will have population dynamical conse-
quences, especially when the population dynamics are pri-
marily driven by the recruitment of new breeders in years of
abundant food supply. Our findings further caution for inter-
preting possibly large differences in fecundity across years of
varying quality as indicative of having population dynamical
impact, and we emphasize the need to integrate fecundity
and survival in a model for making robust conclusions on the
population dynamics.
Acknowledgements
This is report number 6 of Kimpari Bird Projects. We thank the other
members of KBP – Juhani Ahola, Pentti Ahola, Bo Ekstam, Arto
Laesvuori and Martti Virolainen – for the many hours spent con-
ducting field work.We thankHannu Pietiainen, Peter Sunde and two
anonymous referees for insightful comments. Jari Valkama and Sep-
po Niiranen from the Finnish Ringing Bureau kindly provided data
on tawny owl recruits. Author contributions: all data were collected
byKA andTK; initial analyses were carried out byAZ; final analyses
and writing by PK and JEB. AZwas supported by CIMO, PK by the
Academy of Finland (project 1118484 to JEB) and JEB was
employed as anAcademyResearcher.
References
Ahola, K. & Niiranen, S. (1986) Lehtopollon ian maarittaminen (age determi-
nation of the tawny owl).Lintumies, 21, 81–85.
Altwegg, R., Roulin, A., Kestenholz, M. & Jenni, L. (2003) Variation and
covariation in survival, dispersal, and population size in barn owlsTyto alba.
Journal of Animal Ecology, 72, 391–399.
Altwegg, R., Roulin, A., Kestenholz, M. & Jenni, L. (2006) Demographic
effects of extreme winter weather in the barn owl.Oecologia, 149, 44–51.
Altwegg, R., Schaub,M. &Roulin, A. (2007) Age-specific fitness components and
their temporal variation in the barn owl.AmericanNaturalist, 169, 47–61.
Andersson, M. (1980) Site tenacity in birds. Journal of Animal Ecology, 49,
175–184.
Brommer, J.E., Pietiainen, H. & Kolunen, H. (1998) The effect of age at first
breeding on Ural owl lifetime reproductive success and fitness under cyclic
food conditions. Journal of Animal Ecology, 67, 359–369.
Brommer, J.E., Kokko, H. & Pietiainen, H. (2000) Reproductive effort and
reproductive values in periodic environments. American Naturalist, 155,
454–472.
Brommer, J.E., Pietiainen, H.&Kolunen,H. (2002) Reproduction and survival
in a variable environment: the Ural owl and the three-year vole cycle. Auk,
119, 194–201.
Brommer, J.E., Ahola, P. &Karstinen, T. (2005) The colour of fitness: plumage
coloration and lifetime reproductive success in the tawny owl.Proceedings of
the Royal Society of London. Series B, Biological Sciences, 272, 935–940.
Burnham, K.P. & Anderson, D.R. (1998) Model Selection and Inference – A
Practical Information Theoretical Approach. Springer, NewYork.
Cam, E. & Monnat, J.Y. (2000) Apparent inferiority in first-time breeders in
the kittiwake: the role of heterogeneity among age-classes. Journal of Animal
Ecology, 69, 380–394.
Caswell, H. (2000) Prospective and retrospective perturbation analyses: their
role in conservation biology.Ecology, 81, 619–627.
Caswell, H. (2001) Matrix Population Models: Construction, Analysis, and
Interpretation, 2nd edn. Sinauer Associates, Sunderland,MA.
Cooch, E. &White, G. (2001)ProgramMARK.AGentle Introduction, 2nd edn.
Available at: http://www.phidot.org/software/mark/docs/book/, accessed
21 January 2009.
Crawley, M.J. (2002) Statistical Computing. An Introduction to Data Analysis
Using S-Plus. JohnWiley and Sons, NewYork.
Desfor, K.B., Boomsma, J.J. & Sunde, P. (2007) Tawny owls Strix aluco with
reliable food supply producemale-biased broods. Ibis, 149, 98–105.
Elton, C. (1927)Animal Ecology. University of Chicago Press, Chicago.
Francis, C.M. & Saurola, P. (2004) Estimating components of variance in
demographic parameters of tawny owls, Strix aluco. Animal Biodiversity and
Conservation, 27, 489–502.
Gilg, O., Hanski, I. & Sittler, B. (2003) Cyclic dynamics in a simple vertebrate
predator-prey community. Science, 302, 866–868.
Hanski, I. &Korpimaki, E. (1995)Microtine rodent dynamics in northern Eur-
ope: parameterized models for the predator-prey interaction. Ecology, 76,
840–850.
Hanski, I., Hansson, L. & Henttonen, H. (1991) Specialist predators, generalist
predators, and the microtine rodent cycle. Journal of Animal Ecology, 60,
353–367.
Hirons, G.J.M. (1985) The effects of territorial behaviour on the stability and
dispersion of Tawny owl (Strix aluco) populations. Journal of Zoology, Lon-
don (B), 1, 21–48.
Holm, S. (1979) A simple sequentially rejective multiple test procedure. Scandi-
navian Journal of Statistics, 6, 65–70.
Hornfeldt, B., Hipkiss, T. & Eklund, U. (2005) Fading out of vole and predator
cycles? Proceedings of the Royal Society of London. Series B, Biological Sci-
ences, 272, 2045–2049.
Ims, R.A., Henden, J.-A. & Killengreen, S.T. (2008) Collapsing population
cycles.Trends in Ecology and Evolution, 23, 79–86.
Jedrzejewski, W., Jedrzejewska, B., Szymura, A. & Zub, K. (1996) Tawny owl
(Strix aluco) predation in a pristine deciduous forest (Bialowieza National
Park, Poland). Journal of Animal Ecology, 65, 105–120.
Karell, P., Pietiainen, H., Siitari, H., Pihlaja, T., Kontiainen, P. & Brommer,
J.E. (2009) Parental allocation of additional food to own health and off-
spring growth in a variable environment. Canadian Journal of Zoology, 87,
8–19.
Kekkonen, J., Kolunen, H., Pietiainen, H., Karell, P. & Brommer, J.E. (2008)
Tawny owl reproduction and offspring sex ratio in variable food conditions.
Journal of Ornithology, 149, 59–66.
Korpimaki, E. (1988) Effects of age on breeding performance of Tengmalm’s
owlAegolius funereus in western Finland.Ornis Scandinavica, 19, 21–26.
Korpimaki, E. (1992) Fluctuating food abundance determines the lifetime
reproductive success in male Tengmalm’s owls. Journal of Animal Ecology,
61, 103–111.
Korpimaki, E. & Norrdahl, K. (1989) Predation of Tengmalm’s owls: numeri-
cal responses, functional responses and dampening impact on population
fluctuations ofmicrotines.Oikos, 54, 154–164.
Korpimaki, E. & Norrdahl, K. (1991) Numerical and functional responses of
kestrels, short-eared owls, and long-eared owls to vole densities.Ecology, 72,
814–826.
Korpimaki, E., Norrdahl, K., Klemola, T., Pettersen, T. & Stenseth, N.C.
(2002) Dynamic effects of predators on cyclic voles: field experimentation
and model extrapolation. Proceedings of the Royal Society of London. Series
B, Biological Sciences, 269, 991–997.
Tawny owl population dynamics and the vole cycle 1061
� 2009 TheAuthors. Journal compilation� 2009 British Ecological Society, Journal of Animal Ecology, 78, 1050–1062

Korpimaki, E., Norrdahl, K., Huitu, O. & Klemola, T. (2005a) Predator-
induced synchrony in population oscillations of coexisting small mammal
species. Proceedings of the Royal Society of London. Series B, Biological Sci-
ences, 272, 193–202.
Korpimaki, E., Oksanen, L., Oksanen, T., Klemola, T., Norrdahl, K. & Banks,
P.B. (2005b) Vole cycles and predation in temperate and boreal zones of Eur-
ope. Journal of Animal Ecology, 74, 1150–1159.
Krebs, C.J., Boutin, S., Boonstra, R., Sinclair, A.R.E., Smith, J.N.M., Dale,
M.R.T., Martin, K. & Turkington, R. (1995) Impact of food and predation
on the snowshoe hare cycle. Science, 269, 1112–1115.
Kruger, O. (2002) Dissecting common buzzard lifespan and lifetime reproduc-
tive success: the relative importance of food, competition, weather, habitat
and individual attributes.Oecologia, 133, 474–482.
Kruger, O. & Lindstrom, J. (2001) Lifetime reproductive success in common
buzzard, Buteo buteo: from individual variation to population demography.
Oikos, 93, 260–273.
Laaksonen, T., Korpimaki, E. & Hakkarainen, H. (2002) Interactive effects of
parental age and environmental variation on the breeding performance of
Tengmalm’s owls. Journal of Animal Ecology, 71, 23–31.
Lambin, X., Petty, S.J. &MacKinnon, J.L. (2000) Cyclic dynamics in field vole
populations and generalist predation. Journal of Animal Ecology, 69, 106–
118.
Lindstrom, J., Ranta, E., Kokko, H., Lundberg, P. & Kaitala, V. (2001) From
arctic lemmings to adaptive dynamics: Charles Elton’s legacy in population
ecology. Biological Reviews of the Cambridge Philosophical Society, 76, 129–
158.
Lundberg, A. (1986) Adaptive advantages of reversed sexual size dimorphism
in European owls.Ornis Scandinavica, 17, 133–140.
Manly, B.F.J. (1997) Randomization, Bootstrap and Monte Carlo Methods in
Biology. Chapman&Hall, London.
Meijer, T., Daan, S. &Hall,M. (1990) Family planning in the kestrel (Falco tin-
nunculus): the proximate control of covariation of laying date and clutch size.
Behaviour, 114, 117–136.
Metcalf, C.J.E. & Pavard, S. (2007) Why evolutionary biologists should be
demographers.Trends in Ecology and Evolution, 22, 205–212.
Mosher, J.A. & Henny, C.J. (1976) Thermal adaptiveness of plumage color in
screech owls.Auk, 93, 614–619.
Newton, I. (1989)Lifetime Reproduction in Birds. Academic Press, London.
Norrdahl, K., Heinila, H., Klemola, T. & Korpimaki, E. (2004) Predator-
induced changes in population structure and individual quality of Microtus
voles: a large-scale field experiment.Oikos, 105, 312–324.
Petty, S.J. (1999) Diet of tawny owls (Strix aluco) in relation to field vole (Mi-
crotus agrestis) abundance in a conifer forest in northern England. Journal of
Zoology, London, 248, 451–465.
Pietiainen, H. (1988) Breeding season quality, age, and the effect of experience
on the reproductive success of the Ural owl (Strix uralensis). Auk, 105, 316–
324.
Pinheiro, J.C. & Bates, D.M. (2000) Mixed-Effects Models in S and S-PLUS.
Springer, NewYork BerlinHeidelberg.
Ratcliffe, N., Furness, R.W. & Hamer, K.C. (1998) The interactive effects of
age and food supply on the breeding ecology of great skuas. Journal of Ani-
mal Ecology, 67, 853–862.
Reid, J.M., Bignal, E.M., Bignal, S., McCracken, D.I. & Monaghan, P. (2004)
Identifying the demographic determinants of population growth rate: a case
study of red-billed choughs Pyrrhocorax pyrrhocorax. Journal of Animal
Ecology, 73, 777–788.
Rohner, C. (1996) The numerical response of great horned owls to the snow-
shoe hare cycle: consequences of non-territorial ‘floaters’ on demography.
Journal of Animal Ecology, 65, 359–370.
Rosenberg, D.K., Swindle, K.A. & Anthony, R.G. (2003) Influence of prey
abundance on northern spotted owl reproductive success in western Oregon.
Canadian Journal of Zoology, 81, 1715–1725.
Roulin, A., Ducret, B., Ravussin, P.-A. & Altwegg, R. (2003) Female colour
polymorphism covaries with reproductive strategies in the tawny owl Strix
aluco. Journal of Avian Biology, 34, 393–401.
Southern, H.N. (1970) The natural control of a population of Tawny owls
(Strix aluco). Journal of Zoology, 162, 197–285.
Sunde, P. & Bølstad, M.S. (2004) A telemetry study of the social organi-
zation of a tawny owl (Strix aluco) population. Journal of Zoology,
263, 65–76.
Sunde, P., Bølstad, M.S. &Moller, J.D. (2003) Reversed sexual dimorphism in
tawny owls, Strix aluco, correlates with duty division in breeding effort. Oi-
kos, 101, 265–278.
Sundell, J., Huitu, O., Henttonen, H., Kaikusalo, A., Korpimaki, E., Pietiai-
nen, H., Saurola, P. & Hanski, I. (2004) Large-scale spatial dynamics of vole
populations in Finland revealed by the breeding success of vole-eating preda-
tors. Journal of Animal Ecology, 73, 167–178.
Taylor, I. (1994) Barn Owls: Predator-Prey Relationships and Conservation.
CambridgeUniversity Press, Cambridge.
Wallin, K. (1987) Defence as parental care in tawny owls (Strix aluco). Behav-
iour, 102, 213–230.
White, G.C. & Burnham, K.P. (1999) Program MARK: survival estimation
from populations ofmarked animals.Bird Study, 46, S120–S138.
Received 9 June 2008; accepted 17 April 2009
Handling Editor: AlexRoulin
1062 P. Karell et al.
� 2009 TheAuthors. Journal compilation� 2009 British Ecological Society, Journal of Animal Ecology, 78, 1050–1062




















