
Polar lipid ester-linked fatty acid composition of Lake Vechten seston: an ecological application of...
16
FEMS Microbiology Ecology 38 (1986) 381-396 381 Published by Elsevier FEC 00094 Polar lipid ester-linked fatty acid composition of Lake Vechten seston: an ecological application of lipid analysis (Fatty acid; mesotrophic monomictic lake; organic geochemistry; capillary GC-MS) H.L. Fredrickson a,*, T.E. Cappenberg a and J.W. de Leeuw b a Limnological Institute, Vijverhof Laboratory, Rijl~straatweg 6, 3631 A C Nieuwersluis, and b Delft University of Technology, Department of Chemistry and Chemical Engineering, Organic Geochemistry Unit, De Vries van Heystplantsoen 2, 2628 RZ Delft, The Netherlands Received 4 June 1986 Revision received 7 October 1986 Accepted 8 October 1986 1. SUMMARY In order to relate the benthic lipid composition to possible sources in the water column, the sestonic communities of a monomictic lake were profiled using their saponifiable polar lipid fatty acids, which were identified by capillary gas chro- matography-mass spectrometry (GC-MS). The epilimnion, dominated by the dinoflagellate alga Ceratium hirundella, was characterized by C20:5 and C22:6 polyunsaturated fatty acids. The photic anoxic metalimnion supported a radically differ- ent community, dominated by photosynthetic sulfur-oxidizing bacteria ( Chromatium and Chloro- nema spp.) and a Synechococcus-like cyano- bacterium, and was characterized by high con- centrations of C16 and Cls monounsaturated fatty acids. The fatty acid compositions of the hypo- limnetic seston and the sediment were qualita- tively similar to that of the metalimnion. Methyl- branched acids, commonly found in eubacteria, increased with depth in the water column. The concentrations of several unusual fatty acids found * To whom correspondence should be addressed. in Desulfovibrio spp. Desulfobacter spp. and De- sulfotomaculum spp. were inversely related to sulfate concentration in the metalimnion. After the water column mixed in the winter, steep gradi- ents of respiratory terminal electron acceptors de- veloped in the surface sediment and were reflected in the polar lipid fatty acid compositions. The results show that fatty acids derived from the membranes of epilimnetic phytoplankton were ef- ficiently metabolized in the oxic portion of the water column. The fatty acids synthesized by pro- karyotic microorganisms at and below the oxy- cline dominated the sediment. The polar lipid fatty acid composition of the sediment showed seasonal changes which can be associated with concentrations of terminal electron acceptors of microbial respiration, and thus with physiologi- cally distinct bacterial groups. 2. INTRODUCTION Organic geochemical studies have been per- formed on a number of oligotrophic [1] and eutrophic lakes [2,3] and current research into the organic compounds extractable from lake sedi- 0168-6496/87/$03.50 © 1987 Federation of European Microbiological Societies
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Polar lipid ester-linked fatty acid composition of Lake Vechten seston: an ecological application of...

FEMS Microbiology Ecology 38 (1986) 381-396 381 Published by Elsevier
FEC 00094
Polar lipid ester-linked fatty acid composition of Lake Vechten seston: an ecological application of lipid analysis
(Fatty acid; mesotrophic monomictic lake; organic geochemistry; capillary GC-MS)
H.L. Fredrickson a,*, T.E. Cappenberg a and J.W. de Leeuw b
a Limnological Institute, Vijverhof Laboratory, Rijl~straatweg 6, 3631 A C Nieuwersluis, and b Delft University of Technology, Department of Chemistry and Chemical Engineering, Organic Geochemistry Unit, De Vries van Heystplantsoen 2, 2628 RZ Delft, The
Netherlands
Received 4 June 1986 Revision received 7 October 1986
Accepted 8 October 1986
1. SUMMARY
In order to relate the benthic lipid composition to possible sources in the water column, the sestonic communities of a monomictic lake were profiled using their saponifiable polar lipid fatty acids, which were identified by capillary gas chro- matography-mass spectrometry (GC-MS). The epilimnion, dominated by the dinoflagellate alga Ceratium hirundella, was characterized by C20:5 and C22:6 polyunsaturated fatty acids. The photic anoxic metalimnion supported a radically differ- ent community, dominated by photosynthetic sulfur-oxidizing bacteria ( Chromatium and Chloro- nema spp.) and a Synechococcus-like cyano- bacterium, and was characterized by high con- centrations of C16 and Cls monounsaturated fatty acids. The fatty acid compositions of the hypo- limnetic seston and the sediment were qualita- tively similar to that of the metalimnion. Methyl- branched acids, commonly found in eubacteria, increased with depth in the water column. The concentrations of several unusual fatty acids found
* To whom correspondence should be addressed.
in Desulfovibrio spp. Desulfobacter spp. and De- sulfotomaculum spp. were inversely related to sulfate concentration in the metalimnion. After the water column mixed in the winter, steep gradi- ents of respiratory terminal electron acceptors de- veloped in the surface sediment and were reflected in the polar lipid fatty acid compositions. The results show that fatty acids derived from the membranes of epilimnetic phytoplankton were ef- ficiently metabolized in the oxic portion of the water column. The fatty acids synthesized by pro- karyotic microorganisms at and below the oxy- cline dominated the sediment. The polar lipid fatty acid composition of the sediment showed seasonal changes which can be associated with concentrations of terminal electron acceptors of microbial respiration, and thus with physiologi- cally distinct bacterial groups.
2. INTRODUCTION
Organic geochemical studies have been per- formed on a number of oligotrophic [1] and eutrophic lakes [2,3] and current research into the organic compounds extractable from lake sedi-
0168-6496/87/$03.50 © 1987 Federation of European Microbiological Societies

382
ments has been summarized [4,5]. Organic com- pounds extracted from lake sediments can provide information on the trophic status [6-8] as well as the source of sedimentary organic carbon [9-12]. However, detailed ecological interpretation of bio- geochemical data is limited by a lack of informa- tion on the changes lipids undergo between the time of cell death and their possible incorporation into the sediment [5]. Recent studies have shown that microbial succession [13] and biological par- ticle reworking [14] can greatly affect the total lipid composition of particles as they sink to the benthos.
In order to relate lipids present in the benthos to their possible sources in the overlying water column, we have determined the polar lipid fatty acid compositions of sestonic particles and surface sediments in a much studied lacustrine environ- ment.
Lake Vechten is a small (4.7 ha) slightly eutrophic monomictic lake, 10 m deep, located in The Netherlands (52°04'N and 5°05'E). A com- plete physical description of the lake can be found elsewhere [15]. The lake has been the subject of intensive limnological study for the past 20 years, and this research has been summarized recently [161.
Summer thermal stratification prevents the water column from mixing and effectively parti- tions it into two physically distinct compartments. Most of the lake's carbon is fixed by epilimnetic phytoplarukton [17]. This primary production is efficiently cropped by zooplankters, principally Diaphanosoma brachyrum and Ceriodaphnia pul- chella [18] and undergoes aerobic mineralization by heterotrophic bacteria [19]. Depletion of re- spiratory terminal electron acceptors (oxygen, nitrate and sulfate) in the deeper water layers results in a chemocline in the photic zone of the lake and limits microbial physiological activities. Water in the region of the thermocline/chemocline is colored red. Pigment analysis has shown that this is due to the presence of the photosynthetic purple sulfur-oxidizing bacteria Chromatium and Thiocapsa spp., which coexist with the green sulfur bacteria Chlorobium spp. and a Synechococcus-like cyanobacterium [20].
Although physically distinct from one another,
these microbial communities are related by energy flow and nutrient recycling. Consumption of epi- limnetic phytoplankton results in a rain of par- ticles through the water column to the sediment surface. For example, during the 151-day stratifi- cation period in 1979, 5.9 kg dry weight of material sedimented to the 8 m horizon of the lake provid- ing energy and nutrients to the benthos [21]. The sediment in turn generates carbon dioxide, meth- ane, ammonia and reduced sulfur compounds which re-enter the water column via diffusion and bubble formation [22,23].
In this study, the sestonic microbial communi- ties which mediate the majority of carbon cycling in Lake Vechten were characterized by the ester- linked fatty acids of their polar lipid. This ap- proach was chosen because it can provide quanti- tative data on extant in situ microbial communi- ties free of the biases which can be introduced by selective growth of microbes on nutritive media. Membrane lipids with restricted phylogenetic dis- tributions and fast environmental turnover times can be used as measures of microbial community biomass and community structure [24]. Phos- pholipids are the principal molecular component of most biological membranes. Their synthesis is genetically regulated, and upon cell death, hydro, lysis of their phosphodiester linkage is rapid [25]. Fatty acid biosynthetic pathways reflect microbial evolution, and thus fatty acid compositions are of taxonomic value. Surveys of the fatty acid com- positions of bacteria [27], photosynthetic bacteria [27,28] and eukaryotic plankton [29-34] have been published. Thus, examination of phospholipids provides a means of examining living biota. The utility of polar lipid fatty acid analysis in profiling sedimentary microbial communities has been demonstrated recently using sediments enriched with selected nutrients [35].
Here we show that the polar lipid fatty acid profile of Lake Vechten surface sediment: (1) dif- fers from the total lipid fatty acid profile of the same sample; (2) strongly resembles the polar lipid profile of the metalimnetic photosynthetic prokaryotic community when the lake is stratified; and (3) changes rapidly at holomixus when oxygen, nitrate and sulfate are made available to the sedi- ment. We interpret these results in light of lipid

analyses of axenic microbial cultures of genera which inhabit the lake and discuss these results in relation to pertinent physico-chemical parameters of the water column and sediment, and biological processes which influence the lipid composition of the benthos.
3. MATERIALS AND METHODS
3.1. Materials Glass-distilled solvents were obtained from
Merck (Amsterdam, The Netherlands). Fatty acid methyl ester standards (FAME, 99% pure) were obtained from Applied Science (State College, PA, U.S.A.).
The purple and green photosynthetic sulfur- oxidizing bacteria Thiocapsa roseopersicina (Chro- matiaceae) and Chlorobium phaeobacterioides (Chlorobiaceae) were obtained from Dr. H. van Gemerden (Groningen, The Netherlands) and grown in batch culture [36]. Stephanodiscus hantzschii (Bacillariophyceae), Chlorella oulgaris (Chlorophyta), and a Synechococcus-type micro- organism (Cyanophyceae) were isolated from Lake Vechten and grown in continuous culture under conditions which simulated those found in the lake [20]. Formalin-killed cultures were harvested by continuous centrifugation and extracted as de- scribed below. D. desulfuricans ssp. aestuarii (ATCC17900) was grown in a mineral salts marine medium containing lactate and reduced with sodium sulfide. A freshwater sulfate-reducing bacterium was isolated from Lake Vechten sedi- ment using a freshwater mineral salts medium containing lactate and reduced with sodium thio- sulfate.
3.2. Sample handling Water samples (10 1) were collected from Lake
Vechten at 9:00 a.m. on September 15, 1984, using a Friedinger water sampler. The samples were stored on ice in tightly capped 10-1 carboys and returned to the laboratory within 2 h of collection. The use of this method precluded the sampling of a water layer less than 1 m thick. The water was centrifuged (20000 × g) at 2°C using a continu-
383
ous flow rotor (M.S.E., Sussex, U.K.) at a flow rate of 80 ml- min-1.
Sediment cores were collected from the eastern basin of the lake at a depth of 10 m with a modified Jenkin mud sampler. Cores were re- turned to the laboratory within 2 h of collection, subcored with a 30-mm-diameter plexiglass tube, extruded, sectioned into 2-cm slices, and im- mediately extracted. Dry weight per unit volume was determined on replicate samples, which were lyophilized.
3.3. Lipid analysis Lipids were extracted from samples by a mod-
ified Bligh and Dyer [37] procedure (Fig. 1). The total lipid fraction thus obtained was thoroughly dried by 3 consecutive chloroform evaporations and weighed. It was dissolved in chloroform and quantitatively loaded onto a 1-g Unisil column 1 cm in diameter (100-200 mesh, Clarkson Chem- ical, Williamsport, PA, U.S.A.). Sequential 10-ml elutions with chloroform, acetone and methanol
I I Chtoroform phase t
{$i021 '
J Chtomform etuate
I Wet SampLe I Btigh - Dyer e x t r a c t i o n
I Aqueous Hethanol ]
1 Hethanol,
etuate I 1) Saponification 2) Extraction
~ P a r t i t i o n ,
I Ne trat I !./MeOH
2) TLC ,
I °H-F"NE I
GC GC-NS
Fig. 1. Row diagram of the analytical procedure used to recover lipids from cultures and environmental samples (CK~, gas liquid chromatography; TLC, thin-layer chromatography).

384
produced 3 lipid fractions with increasing polar- ity; non-polar lipid (NL), polar lipid fraction I (PLI) and polar lipid fraction II (PLII), respec- tively. Solvents were removed under a stream of nitrogen and the separate lipid fractions were weighed.
The PLII fraction was dissolved in 0.2 ml of chloroform and hydrolyzed for 24 h at 20°C in 0.5 ml of a solution composed of 1 M aqueous potassium hydroxide and methanol (1:4, v/v) . Lipids were recovered in chloroform after the pH of the aqueous phase was lowered to 6 with 1 M HCI and dried under N 2. Non-saponifiable lipid was removed by dissolving this dry lipid residue in 1 ml of petroleum ether, vigorously mixing with 2 ml of aqueous 1 M potassium hydroxide, and centrifuging. The aqueous base was extracted sequentially with 2 additional volumes of petro- leum ether. The fatty acids were recovered from the aqueous base by lowering the pH to 6 with HC1, extracting them into chloroform and drying them under N 2. The acids were methylated for 2 h at 100°C in 0.5 ml of a solution containing chlo- roform, concentrated HC1 and methanol (1 : 1 : 10, v / v / v ) . After cooling, 2 ml of chloroform and 2 ml of distilled water were added, the solution was mixed and the FAME were recovered in chloro- form. FAME (R F = 0.6) were separated from hyd- roxylated fatty acid methyl esters (OHFAME, R F = 0.2) by thin-layer chromatography on Whatman K6 silica gel plates developed in petroleum ether, diethyl ether and glacial acetic acid (80 : 20 : 1).
The recovery efficiency of phospholipid fatty acids through the analytical procedure was de- termined by standard additions of L-a-phos- phatidylcholine dimyristoyl (1 /tg) to 5 sediment samples. It was found to be 93% with a standard error of 5% (N = 5). It is therefore assumed that ester-linked fatty acids from lipids with polarities similar to phosphatidylcholine extractable from environmental samples will be efficiently re- Covered in the PLII fraction with this procedure and precisely measured.
Capillary gas chromatography (GC) analyses were performed on a Carlo Erba Model 4160 equipped with an on-column injector, a 25-m CP- Sil 5 fused silica capillary column (Chrompack, Middelburg, The Netherlands) and a flame ioniza-
tion detector. Helium was used as a carrier gas at an average linear velocity of 40 cm. s -1. The initial oven temperature was 40°C. At injection the oven temperature rose ballistically to 130°C and was then programmed to increase at 3°C • m i n -1 until a final temperature of 320°C. All gas chromatographic analyses were repeated using the same chromatographic system except that a 50 m CP-Sil 88 fused silica column was used with hy- drogen (33 cm. s -1) serving as a carrier gas.
Peaks eluting from the capillary column were tentatively identified by co-injection with stan- dards and by calculating retention indices as equivalent carbon length (ECL) and by compari- son with published values. Polyunsaturated fatty acid standards were prepared from cod liver oil using methods previously described [38]. Reten- tion times (RT) of individual peaks in a trace were corrected so that the RT of 16 :0 was constant from trace to trace, and relative RT (RRT) were thus generated. The RRT of the straight-chain saturated acids containing 13-20 carbons were plotted against carbon number and fitted with a linear function (r = 0.999974, A = -31.492, B = 2.968). The ECL value for 16 : 0 was set at 16.00, and all other values were calculated from their respective RRT through this function. All re- sponse factors for relating integration area to mg of FAME were assumed to be equal to Cxs:0.
Mass spectral (MS) analyses were performed on a Hewlett-Packard Mass Selective Detector (HP 5970B) directly coupled to an HP 5890 capillary gas chromatograph without an interface. The chromatographic conditions were the same except a splitless injection was used with an isolation time of 2 min.
3.4. Sulfate analysis Sulfate was separated from other anions by
high pressure liquid chromatography on an ion-ex- change column and quantified by indirect photo- metric detection [39].
3.5. Dissolved oxygen and temperature Dissolved oxygen and temperature were de-
terrnined by lowering a calibrated sensor (Orbi- sphere, Switzerland) into the lake on a graduated cable.

3.6. Particulate organic carbon The organic carbon content of lyophilized ses-
ton and sediment samples was determined on a Carlo Erba Elemental Analyzer (model 1106).
4. RESULTS
The points of collection of the 10-1 seston sam- ples are related to the temperature, oxygen and sulfate profiles of the lake in Fig. 2. The thermo- cÀine occurred at 5.5 m and effectively prevented E water column mixing. Terminal electron acceptors ~ 5- for respiration, i.e., oxygen, nitrate (data not shown) and sulfate, are rapidly consumed at the thermocline and result in the diffusion gradients shown. The depletion of these terminal electron acceptors limits microbial physiological activities and hence the microbial community composition.
The lipid content of Lake Vechten seston and sediment is related to bulk carbon measures in 10 Table 1. The results are expressed on a volume basis so that comparison between the water col- umn and sediment can be made simply. However,
385
0 10 20 . . . . . . . . . . . . . . . . . . . . . . . . TEMP.(*C) . - - .
? . . . . ,5 . . . . 1 , o . .
0 10 20 30 40 O i . . . . K . . . . , . . . . i . . . , l . . . .
[
zt / i
DO (rag/t) . - - .
SO 4 {mglt) o--,.
- S
- S
-S
- S
-S
Fig. 2. Depth profiles of some pertinent physicochemical parameters of Lake Vechten during the autumn stratification in relation to the depths of the 10-1 seston samples (DO, dissolved oxygen; S, seston sample).
Table 1
Bulk carbon measures of Lake Vechten seston and sediment during stratification
Depth Dry weight Particulate a
organic carbon (% dry weight)
Lipid (% particulate organic carbon) b
Total Nonpolar PLII Total PLII fatty acids ( × 1 0 -3 )
Water column ( pg . c m -3) 2.0 m 41.55 27 7.8 4.5 0.9 92 4.0 m 47.58 39 1.6 0.5 0.3 24 6.0 m 58.97 36 3.4 1.0 0.5 45 8.0 m 187.53 13 5.7 2.8 0.8 77
10.0 m 283.01 10 6.2 1.9 0.7 71 Sediment c (rag.era-3)
0 -2 cm 136.22 6.9 11 7.4 1.2 142 2 -4 cm 201.78 6.3 7.6 4.6 0.6 38 4 -6 cm 235.90 5.8 6.6 4.2 0.1 5
a Particulate organic carbon expressed as percentage of the total dry weight of the sample. b Lipid content expressed as a percentage of the absolute concentration of particulate organic carbon. c Repficate analyses (N ffi 5) were performed on the sediment. The values represent means. The standard error of the mean never
exceeded 15% (1-way analysis of variance).

386
notice must be taken of the 1000-fold change in concentration between the water and sediment (/xg. cm -3 to m g - c m - 3 ) . The concentrations of seston and total organic carbon increased with depth through the water column and these trends continued into the first 6 cm of the sediment. The total lipid content of the sestonic particles, as well as that of all constituent lipid fractions, reached a minimum at 4 m. The total lipid concentration of the 10-m water sample was twice that of the 2-m sample. The concentration of NL relative to the total lipid was highest in the epilimnion. This NL fraction contained alkanes, storage lipids (i.e., wax esters and triacylglycerols), degradation products of polar lipids and photosynthetic pigments. The N L fraction of seston collected from below the epilimnion contained large amounts of elemental sulfur. The PLI lipid fraction showed concentra- tion maxima at 2 and 6 m. This lipid fraction contained glycolipids, which are abundant in the photosynthetic apparatus of prokaryotic and eukaryotic phytoplankton. However, these lipids lack the phosphodiester linkage, and their en- vironmental turnover rates have not been de- termined. Therefore, the result of the detailed analysis of this lipid fraction is beyond the scope of this paper.
The sediment collected from the 10 m depth of the lake during autumnal stratification proved to be horizontally homogeneous. However, the mass of sediment per unit volume almost doubled through the first 6 cm as a result of compaction. The total organic carbon content per volume in- creased by 30% but the total lipid content re- mained almost constant. The concentration of the PLII fraction decreased by a factor of 10, while the mass of material remaining unrecovered on the silica column doubled.
The PLII ester-linked fatty acid composition of the epilimnetic seston was markedly different from that of the seston collected at greater depths (Fig. 3 and Table 2). The epilimnetic microbial com- munity was characterized by C18, C20, and C22 mono-, di- and polyunsaturated fatty acids. The most abundant acids were C22:6 (Table 2) (51), C16:0 (20), C18:1~, 9 (34) and C20:5 (41). The compo- nents which elute between C26:0 and C27:0 on the non-polar capillary column (63,64) elute between
C30:0 and C32:0 on the polar capillary column, suggesting that they are C28 polyunsaturated fatty acids, although their precise structures have not yet been determined. The water sample taken at 4 m contained essentially the same fatty acids as in the 2-m sample, although at approximately half the concentrations. However, low concentrations of iC15:0 and a C15:0 became apparent at 4 m.
The PLII fatty acid composition of seston from the red-colored water at 6 m was profoundly different from that of the overlying water. The concentration of the C22:6 fatty acid (51) was reduced to 15% of that found at 2 m. Palmitoleic acid (C16:1~7c) (35) was the major fatty acid com- prising 35% of the total epilimnetic PLII fatty acids. Iso and anteiso methyl-branched acids in- creased in concentration, and C18:1,~ 7 (35) became more abundant than Cx8:l,~ 9 (34). This fatty acid composition showed little qualitative change with depth in the water column below the thermocline, but the total concentrations of fatty acids in- creased to a maximum at the sediment-water in- terface. The fatty acid composition of the sedi- ment (Fig. 4A) strongly resembled that of the anoxic water column (Fig. 3C) and did not change qualitatively with depth (0-6 cm) during the period when the lake was stratified. However, the total concentration of these fatty acids in the deeper sediments was only 5% of that of the surface sediment.
A portion of the total lipid collected from the top 2 cm of lake sediment in September was saponified before being fractionated into the polarity classes (Table 3). The total lipid fatty acid profile was characterized by saturated straight even-carbon-numbered chains ranging from C14 to C34 (Table 3). When compared to the PLII fatty acid profile of the same extract, the monoun- saturated acid C16:1 in the total lipid profile de- creased in abundance relative to C16:0 , and C18:1w9 (34) was more abundant than C18:1,o 7 (35). C18:2o~ 6 (32), C18:3,o 3 (33) and several polyunsaturated C2o, C22 , C24 and C26 acids were present but C2o:5 (41) and C22:6 (51) were detectable only in low con- centrations.
The PLII fatty acid compositions of surface sediments collected when the lake was undergoing holomixus were markedly different from those

A
20 34
41
~5 ~8
Seston 2 m
_1
63
387
B
L
20
35
Seston 6 m
C ~6 Seston 8 m
4 7 8
20
35
ECL 14 15 16 1~ 1~ 19 20 21 22 2"3 2~ 25 26
Fig. 3. Capillary GC traces of polar lipid ester-linked fatty acids of Lake Vechten seston from 2 m (A), 6 m (B) and 8 m (C).

388
Table 2
The major ester-linked polar lipid fatty acid contents of Lake Vechten seston during the summer stratification
FAME, fatty acid methyl esters, are abbreviated as follows; 16 : l~7t represents a 16 carbon fatty acid with 1 double bond located between the 7th and 8th carbon from the aliphatic end of the molecule. ' t ' indicates the double bond is in the trans configuration. ' i ' represents an iso methyl branch, 'a ' represents anteiso branching and 'b ' indicates that the point of methyl branching is not certain. 'c' indicates the presence of a cyclopropane ring. Structures were assigned based on ECL values on 2 columns and mass fragmentography. ECL, equivalent carbon lengths, were calculated using the retention times (25 m CP Sil 5) of the saturated straight-chain fatty acids to derive a linear function (A = - 31.492, B = 2.9683, r = 0.99997). The ECL of 16 : 0 was set to 16.00 and all other ECL values were calculated through the equation. ND, not detected; TR, trace.
Component Content (pg-cm-3) at indicated water depths (m)
No. a FAME ECL 2.0 4.0 6.0 8.0 10.0
1 i14:0 13.68 ND ND N D 16 52 2 14 : 1 13.81 ND ND N D ND 14 3 14 : 1 13.89 ND ND 8 25 56 4 14:0 13.99 ND 41 110 413 687
5 i15 : 1 14.34 ND ND ND 24 56 6 a15 : 1 14.39 ND ND ND 19 31 7 i15 : 0 14.68 ND 13 57 399 699 8 a15 : 0 14.74 ND 12 39 285 570 9 15 : 1 14.81 ND N D 16 139 290
10 15 : 0 14.99 10 20 78 294 344
11 15.40 ND N D ND 14 34 12 15.48 ND N D ND 19 27 14 i16 : 0 15.66 13 8 7 96 108 16 16 : 1~o7 15.70 166 322 3434 7026 7617 17 16 : loa7t 15.76 16 8 180 ND ND 18 16 : 1 15.84 20 42 183 936 1162 20 16 : 0 16.00 2160 1040 1617 2643 2 889
21 i17 : 1 16.35 ND N D 102 586 707 22 a17 : 1 16.40 ND N D 8 32 60 23 b17 : 0 16.42 ND N D 21 101 223 24 i17 : 0 16.68 8 14 47 125 131 25 al,7 : 0 16.74 ND 27 88 263 318 27 17 : 1 16.84 ND 5 59 166 197 28 17 : 0 17.02 10 17 79 144 138
29 18 : 3co6 17.39 241 91 97 60 54 30 18 : 4~03 17.44 581 198 128 130 154 31 17.50 11 7 104 77 50 32 18 : 2aJ6 17.59 204 105 117 263 268 33 18 : 3~3 17.63 81 63 115 272 279 34 18 : 1~9 17.72 1467 431 377 387 418 35 18 : lw7 17.76 221 198 1089 1290 1363 36 18 : l~07t 17.81 68 9 7 6 10 38 18 : 0 18.02 180 166 179 296 255
40 19.17 256 130 121 264 226 41 20 : 5 19.21 703 344 313 297 276 42 19.40 ND N D ND 23 30 43 19.42 10 20 38 45 16 44 19.54 ND ND 72 64 5 45 20 : 1 19.89 150 31 19 5 8 46 20 : 0 19.99 39 14 11 12 10
47 20.26 ND ND 74 57 34

Table 2 (continued)
389
Component Content (pg. c m - 3 ) at indicated water depths (m)
No. a F A M E ECL 2.0 4.0 6.0 8.0 10.0
51 22 : 6 21.00 2 390 962 350 101 21 52 22 : 4 21.09 116 31 21 6 N D 53 21.18 32 14 11 N D N D 54 21.22 37 11 7 N D N D 56 21.83 35 48 34 42 25 57 22 : 0 21.91 25 13 16 20 15
58 22.40 59 26 7 N D N D 60 23.32 26 8 7 2 N D 61 24 : 0 23.58 73 16 16 13 11 63 25.64 532 52 27 N D 5 64 25.67 193 21 13 N D N D
a Reference numbers of major components refer to Figs. 3 and 4.
collected when the lake was stratified (Table 2; Fig. 4 A and B). The latter resembled the micro- bial community at the thermocline but the former contained much less Ct6:t. The oxic sediments of April contained increased concentrations of C14:t, Ct5:1 and straight chain even carbon numbered acids. The April sediment showed a strong vertical stratification pattern which corresponded to steep oxygen, ell, nitrate and sulfate gradients in the top 2 cm. The interfacial sediment (approx. 0-1 cm) was enriched in C t 4 : t , i C t s : 0 , i C t 5 : t , aCts:t , iCt7:0 and aC17:1 relative to the fatty acid com- position of the entire 0-2 cm sediment sample.
To relate the saponifiable PLII fatty acid com- position of Lake Vechten seston to lipids of mi- croorganisms which inhabit the lake, axenic cul- tures of pertinent microorganisms were analyzed using the same methods. Algal species which had been previously isolated from the lake, S. hantzschii and C. vulgaris, contained unsaturated fatty acids (Table 4) characteristic of eukaryotic algae. However, the absence from the epilimnetic seston of the predominant fatty acids in these algae, C22 polyunsaturated fatty acids of Stepha- nodiscus (tentatively identified as ECL 20.7) and the unusual C16:t,0t3 t and polyunsaturated C18 acids of Chlorella indicate that these algae were not abundant in the epilimnion at the time of sampling. Microscopic observations showed that Ceratium hirundinella was the most abundant alga during the sampling period. Unfortunately, this
alga is not easily cultured in the laboratory and as a result there are few reports of its lipid composi- tion.
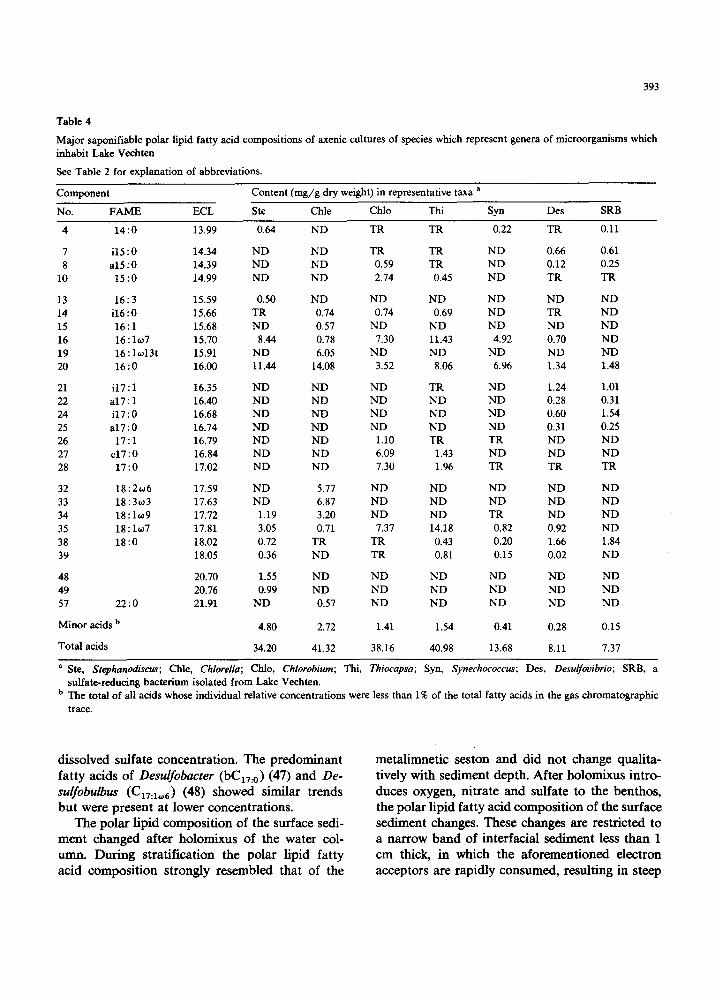
The photosynthetic prokaryotes examined did not Contain polyunsaturated fatty acids (Table 4). The Synechococcus-like cyanobacterium isolated from the lake lacked iso and anteiso methyl- branched fatty acids and contained almost exclu- sively C16:x and Cx6:0 (86%). The purple sulfur bacterium Thiocapsa roseopersicina also contained CI6:I and C16:0 as major fatty acids but in ad- dition contained C18:10, 7 (35%). Chlorobium phaeobacterioides, isolated from a marine environ- ment, differed from the other photosynthetic pro- karyotes examined in that it contained C15 (9%) and CI7 (38%) carbon acids. Both of the colored sulfur bacteria contained significant amounts of methyl branched acids. The sulfate-reducing bacteria examined, D. desulfuricans ssp. aestuarii and a lactate-oxidizing sulfate-reducing bacterium isolated from the lake, were similar to one another in their high content of iso and anteiso methyl- branched C15 and C17 carbon saturated and un- saturated fatty acids.
5. DISCUSSION
Analysis of the organic carbon composition of Lake Vechten seston provides insights into the

390
Table 3
The major ester-linked total and polar lipid fatty acid content of Lake Vechten surface sedimentary layers during summer stratification and Spring holomixus
Abbreviations are explained in Table 2
Component Mean fatty acid content in indicated sediment layer (ng. c m - 3)
No. FAME ECL September 1984
Total Polar lipid lipid
0-2 cm 0-2 cm 2-4 cm 4-6 cm
April 1985
Polar lipid
0-2 cm 0-1 cm
1 i14 : 0 13.68 140 96 40 ND 67 ND 3 14 : 1 13.89 305 14 ND ND 130 29 4 14 : 0 13.99 1843 407 210 25 586 47
5 i15 : 1 14.34 97 39 8 TR 11 TR 6 a15 : 1 14.39 100 49 17 TR 18 TR 7 i15 : 0 14.68 660 356 156 17 340 173 8 a15 : 0 14.74 690 482 269 189 428 15 9 15 : 1 14.81 65 46 14 ND 106 21
10 15 : 0 14.99 396 99 37 4 146 10
11 15.48 146 31 4 ND TR TR 12 15.55 330 66 13 6 TR TR 14 i16 : 0 15.66 602 82 57 5 98 23 16 16 : 1~7 15.70 6 982 4423 953 92 88 TR 17 16 : 1 15.76 618 152 28 10 202 56 20 16 : 0 16.00 8 284 2115 718 101 2120 734
21 i17 : 1 16.35 75 26 18 10 142 127 22 a17 : 1 16.40 70 23 15 TR 112 46 24 i17 : 0 16.68 304 63 72 7 287 177 25 a17 : 0 16.74 338 136 96 12 110 32 27 c17 : 0 16.84 83 96 44 TR 41 26 28 17 : 0 17.02 229 55 33 6 86 8
32 18 : 2~6 17.59 839 316 136 21 46 ND 33 18 : 3~3 17.63 456 244 90 12 ND ND 34 18 : 1~9 17.72 3 943 633 357 58 200 ND 35 18 : 1~7 17.76 1773 1050 375 51 200 ND 36 phytanic 17.86 149 N D ND ND 36 15 38 18 : 0 18.02 1522 164 110 40 268 240
44 19.54 110 15 4 ND ND ND 45 20:1 19.67 120 20 10 TR 36 10 46 20 : 0 19.99 844 67 32 17 241 TR
48 20.70 154 44 TR TR 236 13 50 20.80 163 39 TR TR 250 TR 51 22 : 6 21.00 79 32 TR TR ND ND 55 21.65 281 13 16 7 262 ND 57 22 : 0 21.91 844 34 32 4 404 ND
59 23 : 0 22.72 140 5 8 TR 96 ND
60 23.32 2 041 18 3 N D TR TR 61 24 : 0 23.58 2 722 26 27 3 381 TR
62 26 : 0 25.21 2 338 15 18 TR 275 TR

Table 3 (continued)
Component Mean fatty acid content in indicated sediment layer (ng. c m - 3 )
391
No. FAME ECL September 1984 April 1985
Total Polar lipid Polar lipid lipid
0 - 2 cm 0 - 2 cm 2 -4 cm 4 -6 cm 0 - 2 cm 0-1 cm
63 25.64 2700 ND 64 25.67 1313 15
65 28:0 2581 8 66 30:0 921 ND 67 32:0 260 N D 68 34 :0 862 ND
Number of replicate samples analyzed 3
ND N D TR TR 18 TR TR TR
12 TR 206 TR N D N D 67 N D N D N D N D N D N D N D N D N D
3 3 3 3 1
lake's sestonic microbial community compositions, trophic structure and sediment formation. The rapid sedimentation rate during the period in which the lake is stratified is evidenced by the mass of sestonic particles in the water column. Although the total mass of particles increases with depth in the water column, the percentage of organic carbon (extractable and combustible) comprising the sestonic mass decreases with depth. In the epilimnion, this is indicative of biological particle reworking, which recycles readily metabo- lizable organic compounds and results in sestonic particles comprised of recalcitrant substances. In the hypolimnion a large portion of the sestonic mass is probably composed of reduced mineral salts of nitrogen and sulfur which were deposited as the result of anaerobic microbial respiration.
The polar lipid concentration per volume of lake water showed that membrane biomass had maxima in the epilimnion and sediment-water in- terface and a minimum at 4 m. Saponification of this lipid and analysis of the resulting fatty acid methyl esters revealed that the microbial commun- ity above the thermocline was markedly different from that found in the deeper water and sediment. The fatty acid composition of the epilimnetic ses- ton is indicative of phototrophic eukaryotes. Even-carbon-number fatty acids were predomi- nant, with modes at C16 and C22. Half the acids were polyunsaturated, with C20:s and C22:6 com- prising 30% of the total polar fatty acids. The
C20:5 acid is widely distributed through photo- eukaryotic taxa, but C22:6 has a far more limited distribution and is usually found as the major phospholipid fatty acid in the unicellular flagel- lated algae, division Pyrrophyta, class Dinophyce- ae [32]. Previous observations have revealed that Ceratium hirundinella, a member of the Di- nophyceae, was the principal alga in the epilimn- ion at the time of sampling (3.5 × 105 cells, l -x) [40]. No information on the fatty acid composition of pure cultures of Ceratium spp. could be found in the literature. However, the fatty acid composi- tion of the epilimnetic seston strongly resembles the fatty acid compositions reported for other members of the Dinophyceae [28,41] and is similar to the fatty acids identified in analysis of a dense bloom of this organism collected from the British Lake District [42]. The last peaks to elute from the capillary column (Fig. 3A) comprise a consider- able portion of the PLII fatty acids, and have been tentatively identified as C2s polyunsaturated acids. Long-chain polyunsaturated fatty acids have previously been reported in species of Cryp- tomonas and Chlorella, but the origin of the acids reported in this paper remains to be determined. Some polyunsaturated fatty acids were conspicu- ous by their absence. Polyunsaturated C16 fatty acids which are found in most members of the Bacillariophyceae (diatoms) were not detectable in the epilimnetic seston. The unusual acid Cxr:x~,x3t, which is abundant in Chlorella vulgaris isolated

392
S e d i m e n t O-2cm
S t r a t i f i e d Lake
2O
2o
4
l iT . . 3.
Sediment 0-2cm Nonstr atified Lake
C ~o ~i
7 24
2
S e d i m e n t 0 - 1 cm
N o n s t r a t i f i e d Lake
L .
'iL 38
D. desulfuricans
ECL I~. 15 16 17 18 19 2'0 2'I
Fig. 4. Capillary GC traces of polar lipid ester-linked fatty acids of (A), lake sediment (0-2 cm) 9/84; (B) lake sediment (0-2 cm) 4/85; (C) lake sediment (0-1 cm) 4/85; (D) and D. desulfuricans ssp. aestuarii.
from Lake Vechten and most other Chlorophyceae species examined, was also absent. This is indica- tive of the absence of these species from the epilimnion at the time of sampling.
The water above the thermocline freely mixes but the concentration of polyunsaturated algal fatty acids was greatly reduced in the seston col- lected at 4 m. This is probably the result of the phototactic migration of the phytoplankters up to the shallow water layers where sufficient insola- tion can be intercepted for photosynthetic produc- tion, and to where they are followed by zooplank- ton. Efficient transfer of photoeukaryotic carbon to subsequent trophic levels is mediated by zoo- plankton [40] and heterotrophic bacteria [19], and is evidenced in these results by the lack of polyun- saturated fatty acids in the polar lipids in the deeper water layers and sediment.
The concentrations of polar lipids at the photic chemocline in Lake Vechten are as high as those in the epilimnion, but the large differences in their fatty acid compositions indicate that their mic- robial community structures are very different. Previous microscopic observations and pigment analysis have shown that a Synechococcus-like cyanobacterium was the most populous member of this anoxic phototropic community and that it coexisted with the colored sulfur-oxidizing bacteria Chromatium, Thiocapasa and Chlorobium spp. [20]. The principal fatty acids of the metalimnetic ses- ton, palmitoleic acid (C16:1~7), C16:0 and C18:1to 7 are major fatty acids in these microbial taxa. Precise estimates of the biomass contributed by the metalimnetic species to the community can be made by measuring complex polar lipids which can be associated unambiguously with individual species [43] within the microbial consortium. This can be readily achieved by further chromato- graphic fractionation of the complex polar lipid components before saponification and fractiona- tion of the fatty acids.
Several unusual fatty acids have been isolated from sulfate-reducing bacteria and these acids may be used as markers for this microbial guild in the lake. The concentration of iC17:1, the principal fatty acid of Desulfooibrio [44], also present in Desulfotomaculum [45,46], increased with depth below the chemocline and was inversely related to
393
Table 4
Major saponifiable polar lipid fatty acid compositions of axenic cultures of species which represent genera of microorganisms which inhabit Lake Vechten
See Table 2 for explanation of abbreviations.
Component Content (mg/g dry weight) in representative taxa a
No. FAME ECL Ste Chle Chlo Thi Syn Des SRB
4 14: 0 13.99 0.64 ND TR TR 0.22 TR 0.11
7 i15 : 0 14.34 ND ND TR TR ND 0.66 0.61 8 a15 : 0 14.39 ND ND 0.59 TR ND 0.12 0.25
10 15 : 0 14.99 ND ND 2.74 0.45 ND TR TR
13 16 : 3 15.59 0.50 ND N D ND ND ND ND 14 i16 : 0 15.66 TR 0.74 0.74 0.69 ND TR ND 15 16 : 1 15.68 ND 0.57 ND ND ND ND ND 16 16 : lo~7 15.70 8.44 0.78 7.30 11.43 4.92 0.70 ND 19 16 : l~13t 15.91 ND 6.05 ND ND ND ND ND 20 16 : 0 16.00 11.44 14.08 3.52 8.06 6.96 1.34 1.48
21 i17 : 1 16.35 ND ND ND TR ND 1.24 1.01 22 a17 : 1 16.40 ND ND ND N D ND 0.28 0.31 24 i17 : 0 16.68 ND ND ND ND ND 0.60 1.54 25 a17 : 0 16.74 ND ND ND ND ND 0.31 0.25 26 17 : 1 16.79 ND ND 1.10 TR TR ND ND 27 c17 : 0 16.84 ND ND 6.09 1.43 ND ND ND 28 17 : 0 17.02 ND ND 7.30 1.96 TR TR TR
32 18 : 2~6 17.59 ND 5.77 ND ND N D ND ND 33 18 : 3 t~ 3 17.63 ND 6.87 ND ND ND ND N D 34 18 : 1~9 17.72 1.19 3.20 N D ND TR ND ND 35 18 : loJ7 17.81 3.05 0.71 7.37 14.18 0.82 0.92 ND 38 18 : 0 18.02 0.72 TR TR 0.43 0.20 1.66 1.84 39 18.05 0.36 ND TR 0.81 0.15 0.02 ND
48 20.70 1.55 ND ND N D ND ND ND 49 20.76 0.99 ND ND ND N D ND ND 57 22 : 0 21.91 ND 0.57 ND ND ND ND ND
Minor acids b 4.80 2.72 1.41 1.54 0.41 0.28 0.15
Total acids 34.20 41.32 38.16 40.98 13.68 8.11 7.37
a Ste, Stephanodiscus; Chle, Chlorella; Chlo, Chlorobium; Thi, Thiocapsa; Syn, Synechococcus; Des, Desulfovibrio; SRB, a sulfate-reducing bacterium isolated from Lake Vechten.
b The total of all acids whose individual relative concentrations were less than 1% of the total fatty acids in the gas chromatographic trace.
dissolved sulfate concentration. The predominant fatty acids of Desulfobacter (bE17:0) (47) and De- sulfobulbus (C17:10,6) (48) showed similar trends but were present at lower concentrations.
The polar lipid composition of the surface sedi- ment changed after holomixus of the water col- unto. During stratification the polar lipid fatty acid composition strongly resembled that of the
metalimnetic seston and did not change qualita- tively with sediment depth. After holomixus intro- duces oxygen, nitrate and sulfate to the benthos, the polar lipid fatty acid composition of the surface sediment changes. These changes are restricted to a narrow band of interracial sediment less than 1 cm thick, in which the aforementioned electron acceptors are rapidly consumed, resulting in steep

394
concen t ra t ion grad ien ts [49]. The fa t ty acid com- pos i t ion of the u p p e r m o s t layer s t rongly resembles that of D. desulfuricans, especial ly in the relat ive abundances of the unusua l iC17:a and aCl7:l acids, which have been found in a n u m b e r of lacta te- ox id iz ing su l fa te - reduc ing bac t e r i a [45]. The su l fa te- reducing bac te r ium which was isola ted f rom Lake Vechten sed iment also con ta ined these unusua l b ranched m o n o u n s a t u r a t e d acids.
Saponi f ica t ion of the total l ip id ex t rac ted f rom sed iment p r o d u c e d a fa t ty acid prof i le which dif- fered f rom that of the PLI I alone. The po la r l ip id fa t ty acids are a var iable subset of a larger relat ive cons tan t poo l of fa t ty acids in the sediment . In a subsequent pub l i ca t ion we will show that the ma jo r i ty of ex t rac tab le sed imen ta ry fa t ty acids are e i ther free or esterif ied to less po l a r molecules.
The results of this s tudy demons t r a t e the ut i l i ty of l ip id analysis in ecological studies. Conversely , we emphas ize the need for an ecological basis to p rope r ly recons t ruc t the c i rcumstances in the pas t under which sed iments have been deposi ted . The presen t s tudy has focused on fa t ty acid analysis of the to ta l saponi f ied complex po la r l ipids. How- ever, more de ta i led in fo rmat ion on microb ia l com- m u n i t y s t ructure m a y be ob ta ined by the struct- ura l de t e rmina t ion of in tac t complex po la r l ip id which can be unambiguous ly associa ted with specific microbia l taxa. This would be pa r t i cu la r ly useful in the examina t ion of a rchaebacter ia , which lack es ter- l inked fa t ty acids. Me thanogens obvi- ously p lay a ma jo r role in ca rbon cycl ing in the lake, bu t have escaped examina t ion in the current s tudy.
A C K N O W L E D G E M E N T S
The authors wish to thank H. van G e m e r d e n (Dept . of Microbio logy , The Univers i ty of G r o - n ingen) and C.L.M. Steenbergen (The Limnologi - cal Inst i tute, Nieuwers luis) for bac te r ia l cultures, P.J .D. Sakkers (Hewle t t -Packard , Amste lveen) for mass spec t romet r ic analysis , H. Goossens and P.H. Best for their cri t ical view of the manuscr ip t , W.I .C. Ri jps t ra for d rawing the figures and C.C.C. K r o o n for typ ing the manuscr ip t .
R E F E R E N C E S
[1] Cranwell, P.A. (1981) Diagenesis of free and bound lipids in the terrestrial detritus deposited in a lacustrine sedi- ment. Org. Geochem. 3, 79-81.
[2] Nishimura, M. (1977) The geochemical significance in e~irly sedimentation of geolipids obtained by saponifica- tion of lacustrine sediment. Geochim. Cosmochim. Acta 41, 1817-1823.
[3] Matsuda, H. and Koyama, T. (1977) Positional isomer composition of monounsaturated fatty acids from a lacustrine sediment. Geochlm. Cosmochim. Acta 41, 341-345.
[4] Barnes, M. and Barnes, W. (1978) Organic compounds in lake sediments, in Lakes: Chemistry, Geology, Physics (A. Lerman, Ed.) pp. 127-152. Springer-Verlag, New York.
[5] Cranwell, P.A. (1982) Lipids of aquatic sediments and sedimenting particles. Progr. Lipid Res. 21,271-308.
[6] Matsuda, H. and Koyama, T. (1977) Early diagenesis of fatty acids in lacustrine sediments, I. Identification and distribution of fatty acids in recent sediments from a freshwater lake. Geochim. Cosmochim. Acta 41,777-783.
[7] Cranwell, P.A. (1984) Lipid biogeochemistry of the sedi- ments from Upton Broad, a small productive lake. Org. Geochem. 7, 25-37.
[8] Cranwell, P.A. (1974) Monocarboxylic acids in lake sedi- ments: indicators, derived from terrestrial and aquatic biota, of paleoenvironmental trophic levels. Chem. Geol. 14, 1-14.
[9] Ishiwatari, R., Ogura, K. and Horie, S. (1980) Organic geochemistry of a lacustrine sediment (Lake Haruna, Japan). Chem. Geol. 29, 261-280.
[10] Pottz, V.J. (1972) Investigation on the occurrence and decomposition of fats and fatty acids in lakes. Arch. Hydrobiol., Suppl., 40, 315-399.
[11] Matsuda, H. (1978) Early diagenesis of fatty acids in a lactustrine sediment, III. Changes in fatty acid composi- tion in the sediments from a brackish water lake. Geo- chim. Cosmochim. Acta 42, 1027-1034.
[12] Cardosa, J., Gaskell, S., Quirk, M. and Eglinton, G. (1983) Hydrocarbon and fatty acid distributions in the Rostherne Lake sediment (England). Chem. Geol. 38, 107-128.
[13] Robinson, N., Cranwell, P.A., Finlay, B. and Eglinton, G. (1984) The lipids of aquatic organisms as potential con- tributors to lacustrine sediments. Org. Geochem. 6, 143-152.
[14] Prahl, F.G., Eglinton, G., Corner, E.D.S. and O'Hara, S.C.M. (1984) Changes in plant lipids during passage through the gut of Calanus. J. Mar. Biol. Assoc. 64, 317-334.
[15] Steenbergen, C.LM. and Verdouw, H. (1982) Lake Vech- ten: aspects of its morphology, climate, hydrology and physicochemical characteristics. Hydrobiologia 95, 11-23.
[16] Gulati, R.D. and Parma, S. (Eds.) (1982) Studies of Lakes Vechten and Tjeukemeer, The Netherlands. Developments in Hydrobiology, Vol. 95. W. Junk, The Hague.
[17] Gons, H.J. (1981) The carbon cycle in Lake Vechten, in

Progress Report 1980 (Parma, S. and Gulati, R.D., Eds.) Institute of the Royal Netherlands Academy of Arts and Sciences.
[18] Gulati, R.D., Siewertsen, K. and Postema, G. (1982) The zooplankton: its community structure, food and feeding, and role in the ecosystem of Lake Vechten. Hydrobiologia 95, 127-164.
[19] Olie, J.J. and Cappenberg, T.E. (1984) Kinetics of aerobic carbon mineralization: the use of a diffusion chamber. Arch. Hydrobiol. Beih. 19, 175-182.
[20] Steenhergen, C.L.M. and Korthals, H.J. (1982) Distribu- tion of phototrophic microorganisms in the anaerobic and microaerophilic strata of Lake Vechten (The Netherlands). Pigment analysis and role in primary production. Limnol. Oceanogr. 27, 883-895.
[21] Verdouw, H. and Dekkers, E.M.J. (1982) Nitrogen cycle of Lake Vechten (The Netherlands); role of sedimenta- tion. Arch. Hydrobiol. 94, 251-263.
[22] Cappenberg, T.E. and Verdouw, H. (1982) Sedimentation and breakdown kinetics of organic matter in the anaerobic zone of Lake Vechten. Hydrobiologia 95, 165-179.
[23] Cappenberg, T.E., Hordijk, K.A., Jonkheer, G.J. and Lauwen, J.P.M. (1982) Carbon flow across the sediment water interface in Lake Vechten, The Netherlands. Hydro- biologia 91, 161-168.
[24] White, D.C. (1986) Quantitative physical-chemical char- acterization of bacterial habitates, in Bacteria in Nature, Vol. 2 (Poindexter, J. and Leadbetter, E., Eds.) pp. 177-203. Plenum Press, New York.
[25] White, D.C., Davis, W.M., Nichols, J.N., King, J.D. and Bobbie, R.J. (1979) Determination of the sedimentary microbial biomass by extractable lipid phosphate. Oeco- logia 40, 51-62.
[26] Lechevalier, M.P. (1976) Lipids in bacterial taxonomy: a taxonomist's view. Crit. Rev. Microbiol. 7, 109-210.
[27] Kenyon, C.N. (1978) Complex lipids and fatty acids of photosynthetic bacteria, in The Photosynthetic Bacteria (Clayton, R.K. and Sistron, W.R., Ed.) pp. 281-313. Plenum Press, New York.
[28] Harrington, G.W., Beach, D.H., Dunhan, J.E. and Holz, G.G. (1970) The polyunsaturated fatty acids of marine dinoflagellates. J. Protozool. 17, 213-219.
[29] Ackman, R.G. and Tocher, C.S. (1968) Marine phyto- plankter fatty acids. J. Fish. Res. Brd. Canada 25, 1603-1620.
[30] Beach, D.H., Harrington, G.W. and Holz, G.G. (1970) The polyunsaturated fatty acids of marine and freshwater cryptomonads. J. Protozool. 17, 501-510.
[31] Cheucas, L. and Riley, J.P. (1969) Component fatty acids of the total lipids of some marine phytoplankton. J. Mar. Biol. Assoc. 49, 97-116.
[32] Erwin, J. (1973) Comparative biochemistry of fatty acids in eukaryotic microorganisms, in Lipids and Biotnem- branes of Eukaryotic Microorganisms (J.A. Erwin, Ed.) pp. 41-144. Academic Press, New York.
[33] Kates, M. and Volcani, B.E. (1966) Lipid components of diatoms. Biochim. Biophys. Acta 116, 264-278.
395
[34] Wood, B.J.B. (1974) Fatty acids and saponifiable lipids, in Algal Physiology and Biochemistry (Stewart, W.D.P., Ed.) pp. 236-265. Blackwell, Oxford.
[35] Guckert, J.B., Antworth, C.B., Nichols, P.D. and White, D.C. (1985) Phospholipid ester-linked fatty acid profiles as reproducible assays for changes in prokaryotic com- munity structure of estuarine sediments. FEMS Microbiol. Ecol. 31,147-158.
[36] Van Gemerden, H. and Beeftink, N.H. (1978) Specific rates of substrate oxidation and product formation in autotrophically growing Chroraatium vinosum cultures. Arch. Microbiol. 199, 135-143.
[37] Bligh, E. and Dyer, W. (1952) A rapid method of lipid extraction and purification. Can. J. Biochem. Physiol. 35, 911-917.
[38] Ackman, R.G. and Burgher, R.D. (1965) Cod liver oil as secondary reference standard in the GLC of the polyun- saturated fatty acids of animal origin: analysis of the dermal oil of the Atlantic leatherback turtle. J. Am. Oil Chem. Soc. 42, 38-42.
[39] Hordijk, C.A., Hagenaars, C.P.C.M. and Cappenberg, T.E. (1984) Analysis of sulfate at the mud-water interface of freshwater sediment using indirect photometric chro- matography. J. Microbiol. Methods 2, 49-56.
[40] Blaauboer, M.C.I. (1982) The phytoplankton species com- position and the seasonal periodicity in Lake Vechten from 1956 to 1979. Hydrobioiogia 95, 25-36.
[41] Nichols, P.D., Jones, G.J., de Leeuw, J.W. and Jones, R.B. (1984) The fatty acids and sterol composition of two marine dinoflagellates. Phytochem. 23, 1043-1047.
[42] Cranweli, P.A. (1976) Decomposition of aquatic biota and sediment formation: organic compounds in detritus result- ing from microbial attack on the alga Ceratium hirundinella. Freshwater Biol. 6, 41-48.
[43] Cruden, D.L. and Startler, R.Y. (1970) The characteriza- tion of chiorobium vesicles and membranes isolated from green bacteria. Arch. Microbiol. 72, 115-134.
[44] Boon, J.J., de Leeuw, J.W., van der Hock, G.J. and Vosjan, J.H. (1976) Significance and taxonomic value of iso and anteiso monoenoic fatty acids and branched fl- hydroxy acids in Desulfovibrio desulfuricans. J. Bacteriol. 129, 1183-1191.
[45] Ueki, A. and Suto, T. (1979) Cellular fatty acid composi- tion of sulfate-reducing bacteria. J. Gen. Appl. Microbiol. 25, 185-196.
[46] Fredrickson, H. (1984) Fatty acid composition of selected sulfate-reducing bacteria and sedimentary communities. Ph.D. Dissertation, Florida State University, Tallahassee, FL, U.S.A.
[47] Taylor, J. and Parkes, R.J. (1983) The cellular fatty acids of sulfate-reducing bacteria, Desulfobacter sp., Desulfo- bulbus sp. and Desulfovibrio desulfuricans. J. Gen. Micro- biol. 129, 3303-3309.
[48] Taylor, J. and Parkes, R.J. (1985) The monoenoic fatty acid composition of a marine species of Desulfobulbus grown of lactate. Lipids 40, 393-397.
[49] Hordijk, K.A., Hagenaars, C.P. and Cappenherg, T.E.

396
(1985) Kinetic studies of bacterial sulfate reduction in freshwater sediments by high pressure liquid chromatogra- phy and microdistillation. Appl. Environ. Microbiol. 49, 434-440.
[50] Edlund, A., Nichols, P.A., Roffey, R. and White, D.C. (1985) Extractable and lipopolysaccharide fatty acid and hydroxy acid profiles from Desulfovibrio species. J. Lipid Res. 26, 982-988.




















