
LIPID-BASED OXIDATIVE PROTEIN MODIFICATIONS IN ...
302
LIPID-BASED OXIDATIVE PROTEIN MODIFICATIONS IN GLAUCOMA by ANNANGUDI PALANI SURESH BABU Submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy Thesis Advisor: Dr. Robert G. Salomon Co-Advisors: Dr. John W. Crabb Dr. Sanjoy K. Bhattacharya Department of Chemistry CASE WESTERN RESERVE UNIVERSITY January 2006
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of LIPID-BASED OXIDATIVE PROTEIN MODIFICATIONS IN ...

LIPID-BASED OXIDATIVE PROTEIN MODIFICATIONS IN GLAUCOMA
by
ANNANGUDI PALANI SURESH BABU
Submitted in partial fulfillment of the requirements for the degree of
Doctor of Philosophy
Thesis Advisor: Dr. Robert G. Salomon
Co-Advisors: Dr. John W. Crabb
Dr. Sanjoy K. Bhattacharya
Department of Chemistry
CASE WESTERN RESERVE UNIVERSITY
January 2006

CASE WESTERN RESERVE UNIVERSITY
SCHOOL OF GRADUATE STUDIES
We hereby approve the dissertation of
______________________________________________________
candidate for the Ph.D. degree *.
(signed)_______________________________________________ (chair of the committee) ________________________________________________ ________________________________________________ ________________________________________________ ________________________________________________ ________________________________________________ (date) _______________________ *We also certify that written approval has been obtained for any proprietary material contained therein.

iv
This thesis is dedicated to my Mom and Dad
Banumathi & Palani

v
TABLE OF CONTENTS
Table of Contents v
List of Schemes xii
List of Tables xvi
List of Figures xvii
Index of Appendix xxiii
Acknowledgements xxvii
List of Abbreviations and Acronyms xxviii
Abstract xxxii
Lipid-Based Oxidative Protein Modification in Glaucoma
Chapter 1. Introduction 1
1.1 Background 2
1.2 HNE as an important lipid peroxidation product 3
1.3 Levuglandins – discovery, formation and pathology 8
1.4 Anatomy of anterior section of the eye 13
1.5 Glaucoma 15
1.5.1 Primary open angle glaucoma (POAG) 16
1.5.2 Secondary glaucomas 16
1.6 Animal models for glaucoma. 17
1.7 Glaucoma and oxidative stress 18
1.8 References 22

vi
Chapter 2. A Short and Efficient Synthesis of 4-Oxo-2-alkenoic
Acids from 2-Alkylfurans 25
2.1. Background 26
2.1.1. Previous synthesis of γ-keto α,β-unsaturated alkenoates 27
2.2. Results and discussion 33
2.3. Conclusions 40
2.4. Experimental procedures 41
2.5. References 51
Chapter 3. Detection and Characterization of Multiple 4-
Hydroxynonenal Adducted Amino acids Using
Deuterium Labeled HNE and Mass Spectrometry 54
3.1. Background 55
3.1.1. Previous syntheses of deuterated HNE 58
3.2. Results 62
3.2.1. Synthesis of [8,8,9,9-2H4]-HNE 62
3.2.2. Reactions of N-acetyl amino acids with HNE 64
3.2.3. Mass spectrometric analysis of the HNE-amino acid adducts 64
3.3. Discussion 85
3.4. Conclusion 101

vii
3.5. Experimental procedures 103
3.5.1. General methods 103
3.6. References 115
Chapter 4. Oxidative Protein Modifications in the Pathogenesis
of Primary Open Angle Glaucoma 118
4.1 Background 119
4.1.1. Oxidative stress and glaucoma 119
4.1.2. Levuglandins and isolevuglandins: formation and pathobiology 122
4.1.3. 4-Hydroxynonenal and its role in pathobiology 124
4.1.4. Advanced glycation end products and their pathobiology 125
4.1.5. Oxidative products of tryptophan and α-hydroxykynurenine 127
4.2 Results 129
4.2.1. Immunoblot analysis of TM tissues for analyzing the levels of
oxidative protein modifications in glaucomatous TM compared to
the controls 129
4.2.2. Localization of modified proteins by immunohistochemical
analysis 132
4.2.3. Immunoprecipitation of modified proteins 133
4.2.4. Identification of immunoprecipitated proteins 134
4.2.6. Identification of modified proteins by two-dimensional gel
electrophoresis 136

viii
4.3 Discussion 139
4.4 Experimental procedures 148
4.4.1. General methods 148
4.4.2. Tissue procurement 148
4.4.3. Protein extraction 149
4.4.4. Western analysis 149
4.4.5. Histochemical analysis 150
4.4.6. Immunoprecipitation 151
4.4.7. 2D Gel electrophoresis 151
4.4.8. LC MS/MS analysis and protein identification 152
4.5 References 154
Chapter 5. Iso[4]LGE2 Modified Proteins in Trabecular
Meshwork of Glaucomatous DBA/2J mice 159
5.1. Background 160
5.1.1 Oxidative stress and inflammation pathways 160
5.1.2. Different forms of secondary glaucoma 162
5.1.3. DBA/2J, mouse model for pigmentary glaucoma 163
5.1.4. Iso[4]Levuglandin E2 – formation and pathology 164
5.2. Results 167
5.2.1. Clinical examination of DBA/2J mice 167

ix
5.2.2. Increase in levels of iso[4]LGE2 modified proteins in DBA/2J
with age 169
5.2.3. Immunohistochemical localization of iso[4]LGE2 modified
proteins 171
5.2.4. Immunoprecipitation of iso[4]LGE2-modified proteins 172
5.2.5. Identification of modified proteins using LC-MS/MS 174
5.3. Discussion 176
5.4. Experimental procedures 180
5.4.1. Tissue procurement 180
5.4.2. Protein extraction 180
5.4.3. Western analysis 180
5.4.4. Immunohistochemistry 181
5.4.5. Immunoprecipitation 182
5.4.6. LC MS/MS analysis 182
5.5. References 184
Chapter 6.
Part A: Pilot Studies Towards Identification of Levuglandin
Modified Proteins in Macrophages
Part B: Initial Studies Towards Developing a Model System
to Differentiate Enzyme Mediated and Free- Radical
Mediated Formation of Levuglandins 187

x
6.1. Background 188
6.1.1. Macrophages, inflammation and atherosclerosis 188
6.1.2. Macrophages and COX-2 188
6.1.3. Lipopolysaccharides (LPS) and cyclooxygenase (COX)
expression 190
6.1.4. Distinguishing levuglandins and isolevuglandins 191
6.2. Results 194
PART A
6.2.1. LPS stimulates formation of LGE2 and iso[4]LGE2 modified
proteins in mouse peritoneal macrophage cell cultures
194
6.2.2. Separation and identification of modified protein by 2D PAGE
and mass spectrometry 196
6.2.3. Immunohistochemical analysis of macrophages 197
PART B
6.2.4. Protein modifications in LPS treated cornea – 1D SDS PAGE 200
6.2.5. Immunohistochemical analysis 201
6.3. Discussion PART A 203
6.3. Discussion PART B 205
6.4. Experimental Procedures 206
6.4.1. General methods 206
6.4.2. Protein extraction 206
6.4.3. Western analysis 207

xi
6.4.4. Histochemical analysis 207
6.4.5. 2D gel electrophoresis 208
6.4.6. LC MS/MS analysis and protein identification 208
6.5. References 210
Appendix (See p. xxiii for Index of Appendix) 213
Thesis conclusion 243
Bibliography 245

xii
LIST OF SCHEMES
Chapter 1
Scheme 1.1: Proposed mechanisms for the formation of HNE from LA. 4Scheme 1.2: Adducts formed by HNE with amino acid side chain
residues. 5Scheme 1.3: Proposed mechanism for the formation of Michael adduct,
Schiff base, amine-HNE (2:1) crosslink adduct and pyrrole adduct. 6
Scheme 1.4: The free radical mediated oxidation of AA-PC and LA-PC leading to the formation of HOOA-PC and HODA-PC that are analogous to HNE and their subsequent reaction with lysine side chain residues to form the corresponding pyrrole adducts, 2-(ω-carboxypropyl) pyrrole (CPP) and 2-(ω-carboxyheptyl) pyrrole (CHP). 7
Scheme 1.5: Formation of prostaglandin endoperoxides from arachidonic acid. 9
Scheme 1.6: Formation of levuglandins from prostaglandin endoperoxides. 9
Scheme 1.7: Representative pathways showing a difference in enzyme and free radical mediated pathways generating LGs from AA-PC. 10
Scheme 1.8: Formation of levuglandins from arachidonyl phosphatidylcholine esters by free radical mediated processes. 11
Scheme 1.9: Formation of isolevuglandin and levuglandin based protein adducts. 12
Scheme 1.10: Daughter ion generated by LG-lysine lactam in the mass spectrometric ionization process. This feature was used in tandem MS/MS to identify the LG modified proteins. 13
Chapter 2
Scheme 2.1: Suggested mechanism for generation of KOdiA-PC and
KDdiA-PC. 26Scheme 2.2: Natural products incorporating 4-oxo-2-alkenoate
functionality. 27Scheme 2.3: 2-Alkoxyfuran as precursor for 4-oxo-2-alkenoate. 28Scheme 2.4: 2-Acylfuran as precursor for 4-oxo-2-alkenoate. 28Scheme 2.5: 2-Siloxyfuran as precursor for 4-oxo-2-alkenoate. 29Scheme 2.6: 2-Silylfuran and 2-siloxyfuran oxidation using dimethyl
dioxirane. 29

xiii
Scheme 2.7: Oxidation of a 2-alkylfuran using pyridinium chlorochromate. 30
Scheme 2.8: Oxidation of 2-alkylfuran using N-bromo succinimide. 30Scheme 2.9: Methods reported for synthesis of a pyrenophorin
precursor: a) using Jones reagent with furyl precursor; b) using (3+2) cyclo-addition of primary nitro group and methyl acrylate, c) using NBS to oxidize a furyl precursor. 31
Scheme 2.10: A patented process for generation of 4-oxo-2-alkenoic acids by Takeya. 32
Scheme 2.11: Synthetic method for generating 2-alkylfurans from furan. 33
Scheme 2.12: Synthesis of the 4-oxo-2-alkenoic acid functional array. 34Scheme 2.13: Ring-chain tautomers of 4-oxo-2-pentenoic acid. 34Scheme 2.14: Comparison of synthetic methods used to generate 4-oxo-
2(E)-alkenoic acids from 2-alkylfurans. 35Scheme 2.15: Synthesis of 4-oxo-2-alkenoic acids from 2-alkylfurans. 36 Scheme 2.16: Synthesis of pyrenophorin precursor from 2-alkylfuran
2.20. 37Scheme 2.17: Synthesis of KOdiA-PC and KDdiA-PC by furan
oxidation protocols employing two different solvent conditions. 38
Scheme 2.18: Synthesis of 1,4-enediones from 2,5-dialkylfurans. 39
Chapter 3
Scheme 3.1. Synthesis of [2H1]-HNE reported by Sugamoto’s group. 59Scheme 3.2. Synthesis of [2H3]-HNE and [2H11]-HNE from
fumaraldehyde dimethylacetal. 60Scheme 3.3. Synthesis of [8,8,9,9-2H4]-HNE. 63Scheme 3.4. Synthesis of HNE. 63Scheme 3.5. Proposed mass spectrometric fragmentation of a 1:1 N-
acetyl histidine - HNE Michael adduct parent ion at m/z 354. See Scheme 3.2S in appendix for fragment structures. 66
Scheme 3.6: Possible Michael adduct HNE - (N-acetyl-Gly-Lys-OMe) fragmentation. See appendix Scheme 3.4S for fragment structures. 72
Scheme 3.7: Possible Schiff base HNE- (gly-lys) Schiff base fragmentation. See appendix Scheme 3.5S for fragment structures. 73
Scheme 3.8: Possible fragmentation of the molecular ion m/z 380 corresponding to an HNE-(N-acetyl-Gly-Lys-OMe) pyrrole adduct. See appendix Scheme 3.6S for fragment structures. 75
Scheme 3.9: Structure of (N-acetyl-Gly-Lys-OMe)–HNE (2:1) 76

xiv
crosslink proposed by Xu et. al. Scheme 3.10: Suggested mode of fragmentations for N-acetyl cysteine
dimer m/z 324. See Scheme 3.7S in appendix for fragment structures. 80
Scheme 3.11: Fragmentation of the molecular ion m/z 302 arising from the dehydration of N-acetyl-Cys-HNE Michael adduct. See Scheme 3.8S in appendix for fragment structures. 81
Scheme 3.12: Possible isomers of the m/z 510 (2:1) HNE-N-acetyl-Cys adduct. 89
Scheme 3.13: Possible fragmentation of the putative parent ion m/z 510 to generate a unique daughter ion at m/z 310 and 408. 90
Scheme 3.14: Possible mass spectrometric cleavage sites of adduct at 510 m/z with a structure 3.19. See Scheme 3.9S in appendix for fragment structures. 91
Scheme 3.15a: Possible fragmentations of the m/z 666 3:1 (HNE/N-acetyl-His) adduct. Inset (tandem MS/MS of the parent ions m/z A) 510, B) 674, C) 670 and D) 666 in the range m/z 394-402). See Scheme 3.10S in appendix for fragment structures. 93
Scheme 3.15b: Possible structures of daughter ions formed by retro-Michael cleavage of the 3:1 parent ion m/z 666. 93
Scheme 3.16: One of the possible structures and the mass spectrometric fragmentation for the molecular ion at m/z 728 (3:1, HNE/(N-acetyl-Gly-Lys-OMe) adduct). See Figure 3.10 for mass spectra. 96
Scheme 3.17: Daughter ions formed from the tandem MS/MS of 2:1 N-acetyl-Cys: HNE adduct A) d0-HNE-2N-acetyl-Cys adduct B) d4-HNE-2N-acetyl-Cys adduct. 98
Scheme 3.18: Daughter ions formed from the tandem MS/MS of 2:2 (N-acetyl cysteine: HNE) adduct m/z 603, 607 and 611.
99Chapter 4
Scheme 4.1: Enzymatic (cyclooxygenase) and non enzymatic (free
radical) routes for the formation of LGE2 and iso[4]LGE2 and their protein adducts. 123
Scheme 4.2: Some of the commonly reported adducts formed by 4-hydroxynonenal reaction with histidine, lysine and cysteine residues. 124
Scheme 4.3: Mechanistic pathway for the formation of AGE’s and argpyrimidine. 126
Scheme 4.4: Catabolic pathway of tryptophan under oxidative conditions. 128
Chapter 5

xv
Scheme 5.1: Diagrammatic representation of activation and expression of proteins involved in inflammation pathway by oxidative stress. Details of the scheme are discussed in the text. (ROS – reactive oxygen species, COX-2 – cyclooxygenase-2, LO – lipooxygenase, NF-кB – nuclear factor- kappa B, IL-18 – interleukin 18, TNF – tumor necrosis factor). 161
Scheme 5.2: Generation of levuglandins from arachidonic acid by enzyme mediated and free radical mechanisms (Detailed scheme presented in Scheme 4.1.2, page 4-4). 165

xvi
LIST OF TABLES
Chapter 2
Table 2.1: Yield and reaction time for alcohol protected alkyl furans in Scheme
2.15. 36Table 2.2: Synthesis of 2-ene-1,4-diones from 2,5-dialkyl furans (Scheme 2.16) 39
Chapter 3
Table 3.1: Nucleophilic functional groups in LDL 57Table 3.2: Isotopic mass abundances of some common elements 61Table 3.3: Optimized parameters for mass spectrometer. 111
Chapter 4
Table 4.1: Total number of tissue samples used for Western blots for each of
the antibodies probed. Some of the tissues were used in more than one Western blot analysis. 129
Table 4.2: Identification of TM proteins immunoprecipitated using iso[4]LGE2 and HNE pAbs. A. Proteins identified by in-gel digestion of bands from lane A-2 (Figure 4.9) using mass spectrometry (LC-MS/MS). B. Proteins identified by in-gel digestion of bands from lane B-2 (Figure 4.9) using mass spectrometry (LC-MS/MS). 136
Table 4.3: Iso[4]LGE2 and HNE immunoreactive proteins identified by 2D PAGE and mass spectrometry. Calculated MW and pI refers to value derived from Swissprot protein database; Measured MW and pI refers to value from the 2D PAGE. 138
Table 4.4: List of crosslinked proteins identified by 2D PAGE and mass spectrometry of glaucomatous TM. 145
Chapter 5
Table 5.1: Proteins immunoprecipitated from TM extract of DBA/2J mice
using iso[4]LGE2-pAb. Gel slices from top to the bottom of lane 4 of the gel in Figure 5.6 were digested using trypsin and the proteins identified using LC-MS/MS and MassLynx™ software with the Swissprot database. 174
Chapter 6
Table 6.1: List of LGE2 – modified proteins electrophoretically separated by 2D gel electrophoresis and identified by mass spectrometry. 197

xvii
LIST OF FIGURES
Chapter 1
Figure 1.1: Anatomy of human eye showing the anterior and posterior regions (Copyright© www.nie.nih.gov). 14
Figure 1.2: Artistic rendering of anterior section of the eye, showing pathway of aqueous flow through the trabecular meshwork (arrow). (Copyright© www.nie.nih.gov) 14
Figure 1.3: Flow chart illustration of the components involved in the pathogenesis of POAG induced by decrease of vitamin E in the ciliary body. MMP – matrix metalloproteinases, TIMP – tissue inhibitor of metalloproteinases, ON – optic nerve, ECM – extracellular matrix. (Copyright obtained from Br. J. Nutrition, CABI Publishing)62 20
Chapter 2
Figure 2.1: UV-spectrograph of 4-oxo-2-pentenoic acid at varing pH. 35
Chapter 3
Figure 3.1: ESI-TOF-MS of reaction mixture containing HNE (d4:d0, 1:1) with amino acids A) N-acetyl-gly-lys-OMe, B) N-acetyl-cysteine and C) N-acetyl-histidine. 65
Figure 3.2: ESI-TOF-MS/MS analysis of Michael adducts parent ions at m/z 354 and 358 from the infusion of reaction mixture from HNE and N-acetyl-His (1:1). 66
Figure 3.3: ESI-MS/MS of molecular ions at A) m/z 518, B) m/z 514 and C) m/z 510, from the infusion of reaction mixture containing HNE and N-acetyl-His. Peaks labeled with an asterix (*) or a solid rectangle (■) denote fragments with or without the C5-C9 alkyl chain of HNE. 67
Figure 3.4: ESI-MS/MS of molecular ions A) m/z 674, B) m/z 670 and C) m/z 666 from infusion of reaction mixture HNE and N-acetyl-His (1:1). 68
Figure 3.5: ESI-LC-SIR for the reaction mixture from N-acetyl-His and HNE with 13 channels of which 7 channels are shown here. A) m/z 670, B) m/z 666, C) m/z 518, D) m/z 514, E) m/z 510, F) m/z 358 G) m/z 354 and H) TIC (for 13 channels).(for chromatograms of channels not shown here, see appendix figure 3.1S) 70
Figure 3.6: ESI-MS/MS analysis of m/z 260 molecular ion. Tandem MS/MS analysis of a solution of N-acetyl-glycine-lysine-OMe was injected into a Q-TOF by infusion at a flow rate of 0.5 µL/min. 71
Figure 3.7: ESI-MS/MS of molecular ions at A) m/z 416 and B) m/z 420 corresponding to a 1:1 HNE-(N-acetyl-Gly-Lys-OMe) Michael adduct. 72

xviii
Figure 3.8: ESI-MS/MS analysis of molecular ions A) m/z 402 and B) m/z 398 corresponding to an HNE- (N-acetyl-Gly-Lys-OMe) Schiff base adduct. 73
Figure 3.9: ESI-MS/MS analysis of molecular ions at A) m/z 384 and B) m/z 380 corresponding to the HNE- (N-acetyl-Gly-Lys-OMe) pyrrole adduct 74
Figure 3.10: ESI-MS/MS analysis of molecular ions A) m/z 656 and B) m/z 652 corresponding to the 1:2 (HNE:(N-acetyl-Gly-Lys-OMe)) crosslink. 76
Figure 3.11: ESI-MS/MS analysis of molecular ions corresponding to the 2:1 (HNE:(N-acetyl-Gly-Lys-OMe)) adducts A) m/z 580, B) m/z 576, C) m/z 572 and the 3:1 (HNE:(N-acetyl-Gly-Lys-OMe)) adducts D) m/z 736, E) m/z 732 and F) m/z 728 G) m/z 740. 77
Figure 3.12: ESI-LC-SIR for the (N-acetyl-Gly-Lys-OMe) and HNE reaction mixture monitored through 17 channels of which the spectrum of 7 channels are shown here (Chromatograms for other channels in presented in appendix). LC-SIR chromatograms for molecular ions at A) m/z 732 (M+4 of 3:1 adduct), B) m/z 728 (M of 3:1 adduct). C) m/z 576 (M+4 of 2:1 adduct), D) m/z 572(M of 2:1 adduct); E) m/z 416 (M of 1:1 Michael adduct); F) m/z 398 (Schiff base adduct); G) m/z 380 (M of 1:1 pyrrole adduct); and H) total ion chromatogram (TIC). (See appendix figure 3.3S for M+8 and M+12 of 3:1 adducts and M+4 of Michael, Schiff base and pyrrole 1:1 adducts) 78
Figure 3.13: ESI-MS/MS analysis for N-acetyl cysteine. 79Figure 3.14: ESI-MS/MS analysis for N-acetyl cysteine dimer at m/z 325. 79Figure 3.15: ESI-MS/MS analysis of the dehydrated N-acetyl-Cys–HNE Michael
adduct. 81Figure 3.16: ESI-MS/MS analysis for molecular ion at m/z A) 469 and B) 465
corresponding to HNE-N-acetyl-Cys (1:2) adduct. 82Figure 3.17: ESI-MS/MS analysis of the ions with A) m/z 611, B) m/z 607 and C)
m/z 603, which correspond to 2:2 N-acetyl-Cys-HNE adducts. 83Figure 3.18: ESI-LC-SIR analysis of the molecular ions observed in N-acetyl-Cys-
HNE reaction mixture through 8 channels. (Chromatograms shown for 8 channels: A) m/z 611, B) m/z 607, C) m/z 603, D) m/z 469, E) m/z 465, F) m/z 346, G) m/z 342 H) m/z 324 m/z and I) TIC. 84
Figure 3.19: Tandem MS/MS of the parent ions A) m/z 358, B) m/z 354, C) m/z 518, D) m/z 514 and E) m/z 510 in the m/z range 207-212 and 238-244. Only the (2:1) HNE/N-acetyl-His adducts C, D and E shows unique daughter ions at m/z 209, 213 and m/z 239, 243. 90
Figure 3.20: Tandem MS/MS of the parent ions at A) m/z 358, B) m/z 354, C) m/z 518, D) m/z 514 and E) m/z 510 in the mass range m/z 309-316. 91
Figure 3.21: Fragmentation of parent ions at A) m/z 518, B) m/z 514 and C) m/z 510 showing the presence of daughter ions m/z 408 and/or m/z 412. 92
Figure 3.22: Relative amounts of adducts formed by N-acetyl-His/HNE reaction, calculated using LC-SIR. The amounts reflect the relative amount (of the adducts monitored) of each of the adduct present in the reaction mixture. Values are average of 2 independent experiments and the

xix
error bars indicate half the difference between the experimental values. 94
Figure 3.23: Relative amounts of adducts formed by (N-acetyl-Gly-Lys-OMe)/HNE reaction, calculated using LC-SIR. The ‘y’ scale reflects percentage of the each adduct present in the reaction mixture (with respect to the adducts monitored). Values are average of 2 independent experiments and the error bars indicate half the difference between the experimental values. 97
Figure 3.24: Relative amounts of adducts formed by N-acetyl-Cys and HNE reaction, calculated using LC-SIR. The ‘y’ scale reflects percentage of the each adduct present in the reaction mixture (with respect to all the adducts monitored). Values are average of 2 independent experiments and the error bars indicate half the difference between the experimental values. 100
Figure 3.25: A) ESI-MS/MS of CHD derived HNE B) ESI-MS/MS of CHD derived d4-HNE. 110
Chapter 4
Figure 4.1: Diagrammatic representation of the cross section of an eye showing the two forms of glaucoma defined according to the difference in the angle of the anterior chamber formed by lines drawn parallel to the iris and cornea (iridial angle). A. normal open angle (~40o) associated with POAG and B. a closed angle (~15o) associated with angle closure glaucoma. 120
Figure 4.2: Diagrammatic representation of the cross section of the human eye showing the aqueous humor outflow pathway. 121
Figure 4.3: Western analysis of POAG and normal trabecular meshwork using anti-iso[4]LGE2 pAb antibodies. TM extracts (5 µg) were subjected to SDS-PAGE, blotted to PVDF membrane and probed with anti-iso[4]LGE2 antibodies. A,C,E . Coomassie blue stained gels. B,D,E. Western blot; Age, race and gender of the tissue samples are indicated (M-Male; F-Female; W-Caucasian; B-African American). 130
Figure 4.4: Western analysis of POAG and normal trabecular meshwork using anti-HNE pAb antibodies. TM extracts (5 µg) were subjected to SDS-PAGE, blotted to PVDF membrane and probed with anti- anti-HNE antibodies. A,C,E. Coomassie blue stained gels. B,D,E. Western blot. M-Male; F-Female; W-Caucasian; B-African American). 130
Figure 4.5: Western analysis of POAG and normal trabecular Meshwork using anti-argpyrimidine mAb. TM extracts (5 µg) were subjected to SDS-PAGE, blotted to PVDF membrane and probed with anti-argpyrimidine antibodies. A,C,E. Coomassie blue stained gels. B,D,E. Western blot. Age, race and gender of the tissue samples are indicated (M-Male; F-Female; W-Caucasian; B-African American). 131
Figure 4.6: Western Analysis of POAG and Normal Trabecular Meshwork using

xx
anti-OHKYN mAb. TM extracts (5 µg) were subjected to SDS-PAGE, blotted to PVDF membrane and probed with anti-OHKYN antibodies. Age, race and gender of the tissue samples are indicated (M-Male; F-Female; W-Caucasian; B-African American). 131
Figure 4.7: Localization of iso[4]LGE2 modified proteins in glaucomatous TM. Anterior segment histochemical sections through the trabecular meshwork are shown. A, B, C – Probed with pre-immune serum; D, E, F,G, H, I – Probed with anti-iso[4]LGE2 antibody; Red Channel – iso[4]LGE2 specific immunofluorescence ; Green Channel – autofluorescence 132
Figure 4.8: Localization of HNE modified proteins in glaucomatous TM. Anterior segment histochemical sections through the trabecular meshwork are shown. A, B, C – Probed with pre-immune serum; D, E, F,G, H, I – Probed with anti-HNE antibody; Red Channel – HNE specific immuno-fluorescence; Green Channel – auto-fluorescence 133
Figure 4.9: Immunoprecipitation of antibody modified protein from TM extract. 10µg of antibody coupled beads and 30µg of TM proteins were used for immunoprecipitations. A. Anti-iso[4]LGE2 antibody B. Anti-HNE antibody .Lane 1 – Protein extract, Lane 2 – IP, Lane 3 – Ab coupled beads, Lane 4 – control-protein A beads, Lane 5 – Western blot of protein extract. 134
Figure 4.10: 2D PAGE analysis of TM extract. ~80 µg of extracted TM proteins was used for the 2D PAGE. The gel was partially transferred onto a PVDF membrane for immunochemical analysis. A. Coomassie stained gel B. Western analysis using Iso[4]LGE2 pAb. C. Western analysis using HNE pAb. D. Merged image showing Coomassie stain in black and Western blots probed with iso[4]LGE2 in blue; HNE in orange. 137
Figure 4.11: Quantification of immunoreactive bands in 1D Western blots. Intensity for each of the lanes detected (all the bands in the lane included) in the Western blots was normalized with respect to the total of all the bands detected in that blot (details of calculations in the experimental section). The minimum intensity for band detection was fixed for each of the blots. ◊ – Relative intensity of a control sample on the blot; □ – OD of a POAG tissue on the blot. A. Levels of iso[4]LGE2 modified proteins (tissue donors – 25 POAGs, 25 controls), B. Levels of HNE modified proteins (tissue donors – 24 POAGs, 25 controls). ‘p’ values calculated by students t-test using Microsoft ® Excel 2003. 141
Figure 4.12: Quantification of immunoreactive bands in 1D Western blots. Relative intensity of optical density (OD) for each of the lanes (all the bands in each lane included) in the Western blots was normalized for each of the blots (8 lanes each). The minimum intensity for band detection was fixed for each of the blots. ◊ – Relative intensity of a control sample on the blot; □ – relative intensity of a POAG tissue on the blot. A. Levels of argpyrimidine modified proteins (tissue donors – 25 POAGs, 25 controls). B. Levels of OHKYN modified proteins (tissue donors – 8

xxi
POAGs, 8 controls). ‘p’ values calculated by students t-test using Microsoft ® Excel 2003. 142
Chapter 5 Figure 5.1: Diagrammatic illustration of sagital and transverse sections of the eye
(not drawn to scale). The sagital section passes through the longitudinal axis of the eye from the anterior to the posterior region. The transverse section passes perpendicularly axis to the sagital axis. 167
Figure 5.2. Anterior segment assessment of DBA/2J mice. The anterior segment pictures were taken with a Leica MZ stereomicroscope (Leica Microsystems Inc., Bannockburn, IL) equipped with a SpotCam RT KE digital camera (Sterling Heights, MI). A. 14-month-old, B. 8-month-old and C. 6-month-old DBA/2J mouse. 168
Figure 5.3. Histochemical assessment of mouse optic nerve. The mouse eye sections (10 µm) passing though optic nerve were stained with hematoxylin. A. 8 months old DBA/2J mouse (x 20) and C. a section through the optic nerve (x 100) B. age matched C57BL6J control mice (x 20) and D. a section through the optic nerve (x 100). A,B – Sagital sections; C,D – Transverse sections. 169
Figure 5.4. Western analysis of TM proteins. TM proteins (~5 µg/lane) were subjected to 1D SDS/PAGE, electroblotted onto a poly(vinylidene difluoride) membrane, and probed with anti-iso[4]LGE2 antibodies. Bovine serum albumin modified with iso[4]LGE2 was used as a positive control. A. Immunoblot of DBA/2J and C57BL6J TM proteins B. Coomassie stained immunoblot.C. Densitometric quantification of the Western blot. 170
Figure 5.5. Histochemical localization of isoLGE2 modified proteins in TM. Representative immunohistochemical analyses with a rabbit polyclonal anti-isoLGE2 are shown. Rhodamine conjugated secondary antibody was used for detection. Immunofluoresence. A. DBA/2J mice TM, 8 months (green channel) B. DBA/2J mice TM, 8 months(red channel) C. C57BL6J mice TM, 8months (green channel). D. C57BL6J mice TM, 8 months (red channel). 172
Figure 5.6: Immunoprecipitation of oxidatively modified TM proteins from 8 month old DBA/2J mice. Immunoprecipitations (Ips) utilized antibody coupled beads (10 µg) and TM protein extracts (10 µg). They were analyzed by SDS-PAGE and mass spectrometry. Coomassie blue stained gels are shown: lane 1, low molecular weight marker; lane 2, antibody coupled beads without TM; lane 3, TM protein extract (5 µg); lane 4, IP products; lane 5, wash containing proteins post IP. 173
Figure 5.7. Proteasome crosslinked in 8-month-old DBA/2J mice. A. Coomassie stained SDS PAGE (truncated Figure 5.4). B. Immunoblot probed with anti-rabbit proteasome polyclonal antibody (arrow shows the position of high molecular weight band immunoreactive for the proteasome antibody). 179

xxii
Chapter 6
Figure 6.1: Schematic description of one of the factors contributing to foam cell f ormation through endocytosis of oxLDL by macrophages. 189
Figure 6.2: LGE2-modified proteins in LPS stimulated macrophages. A. Coomassie stained SDS PAGE proteins from macrophages. B. Western blot analysis for LGE2 immunoreactivity in the electroblotted PVDF membrane. C. Densitometric quantification of the total immunoreactive bands in each of the lanes (as relative intensity). 1- control macrophages, 2 – LPS (10 µg/mL) treated macrophages. 194
Figure 6.3: Iso[4]LGE2-modified proteins in LPS stimulated macrophages. A. Coomassie stained SDS PAGE of proteins from macrophages. B. Iso[4]LGE2 immunoreactivity of the electro-blotted PVDF membrane. C. Densitometric quantification of the total immunoreactive bands in each of the lanes (as relative intensity). 195
Figure 6.4: Separation of proteins extracted from macrophages treated with LPS. A. Western blot using anti-LGE2-KLH polyclonal antibody. B. Coomassie blue stained 2D PAGE. 196
Figure 6.5: Immunohistochemical analysis of macrophages for LGE2 modified proteins. macrophages grown to confluency were plated on to a glass slide and treated with or without LPS. A. Cells treated with LPS and stained with pre-immune serum. B. Controls cells stained with anti-LGE2-KLH pAb. C. Cells treated with LPS stained with anti-LGE2-KLH pAb D. Cells treated with LPS along with indomethacin for 24 h and stained with anti-LGE2-KLH pAb. 198
Figure 6.6: Western blot using anti-LGE2 pAb of C57BL/6 mouse corneal protein extract (2 µg) treated with PBS or LPS. Lane 1. Naïve cornea. Lane 2. PBS treated cornea. Lane 3. LPS (1 µg/mL) treated cornea. 200
Figure 6.7: Relative amount of LGE2 immunoreactivity. Densitometric quantification of immunoreactive bands of the Western blots probed with LGE2-pAb. Naïve – corneal proteins of untreated C57BL/6 mouse, PBS – corneal proteins of mice treated with PBS, LPS - corneal proteins of mice treated with LPS (10 µg/mL). Error bars indicate standard deviation for three experiments. 201
Figure 6.8: Cornea of C57BL/6 mouse was treated with LPS or PBS in vivo and preserved after 6 h and 24 h. Sections (5 µm) of cornea was stained with LGE2 and iso[4]LGE2 pAb. Naïve corneal sections were used as additional controls. A,B,C – Naïve cornea; D,E,F – PBS treated (24 h); G,H,I – LPS treated (6 h); J,K,L – LPS treated (24 h). Panels treated with preimmune, LGE2 and iso[4]LGE2 pAb’s are indicated in the figure
202

xxiii
Index of Appendix
List of Figures
Figure 2.1S: a) 13C NMR and b) 1H NMR of 2.14 214Figure 2.2S: 1H NMR of 2.38 (crude) 215Figure 2.3S: a) 13C NMR and b) 1H NMR of 2.37 215Figure 2.4S: a) 13C NMR and b) 1H NMR of 2.17 216Figure 2.5S: a) 13C NMR and b) 1H NMR of 2.38 217Figure 2.6S: 1H NMR of 2.21 218Figure 2.7S: a) 13C NMR and b) 1H NMR of 2.22 219Figure 2.8S: 1H NMR of 2.39 (crude) 220Figure 2.9S: a) 13C (APT) NMR and b) 1H NMR of sec-butyl ester of 2.40 221Figure 2.10S: 1H NMR of 2.27 (crude) 222Figure 2.11S : 1H NMR of 2.26 223 Figure 3.1S : a) 1H NMR and b) 13C NMR of 3.3 224Figure 3.2S : 2H NMR of 3.4 226Figure 3.3S : a)1H NMR and b) 13C NMR of 3.6 225Figure 3.4S: 2H NMR of 3.7 227Figure 3.5S : a) 1H NMR and b) 13C NMR of 3.7 228Figure 3.6S: How to read a mass spectra? 229Figure 3.7S: ESI-TOF-SIR of N-acetyl-His:HNE reaction mixture.
Channels shown here m/z 696, m/z 692, m/z 688, m/z 684, m/z 678 and m/z 674. 232
Figure 3.8S: ESI-TOF-SIR of N-acetyl-Gly-Lys-OMe:HNE reaction mixture. Channels shown here are from m/z 384, 402, 420, 580, 640, 644, 652, 656, 736 and 740. 233
Figure 3.9S: ESI-TOF-SIR of N-acetyl-His:HNE reaction mixture. Channels shown here are from channels monitored at m/z 320, m/z 326, m/z 352, m/z 356 and m/z 360. 235
Figure 4.1S: Western blot analyses of normal and POAG trabecular
meshwork using anti-iso[4]LGE2 antibodies as described in Section 4.4.4. 238
Figure 4.2S: Western blot analyses of normal and POAG trabecular meshwork using anti-HNE antibodies as described in Section 4.4.4. 238

xxiv
Figure 4.3S: Western blot analyses of normal and POAG trabecular meshwork using anti-argpyrimidine antibodies as described in Section 4.4.4. 239
Figure 4.3S: Immunoprecipitation of POAG trabecular meshwork using anti-iso[4]LGE2 pAb and preimmune serum antibodies as described in Section 4.4.6 239
Figure 5.1S: Western blot of TM proteins from DBA/2J mouse of different
age groups. 5 µg of protein was electrophoretically separated on a SDS PAGE and electroblotted on a PVDF membrane and probed with iso[4]LGE2 pAb. Time course of DBA/2J mice. A. Western blot. B. Coomassie stained SDS PAGE. 240
Figure 5.2S: Immunoprecipitation of trabecular meshwork proteins from a 8 month old DBA/2J mouse using anti-iso[4]LGE2 pAb and preimmune serum antibodies as described in Section 5.4.6. 241
Figure 6.1S: LGE2-modified proteins in LPS stimulated macrophages.
Western blot analysis for LGE2 immunoreactivity in the electroblotted PVDF membrane. 242

xxv
List of Tables
Table 3.1: Summary of tandem MS/MS fragments incorporating the C5
alkyl chain of HNE derived from the precursor ions 354, 358, 510, 514, and 510 thus showing the 4 Da difference. 237

xxvi
List of Schemes
Scheme 3.1S: Fragmentation of N-acetyl-Histidine 229Scheme 3.2S: Fragmentation of (N-acetyl-His)-HNE Michael adduct 230Scheme 3.3S: Suggested fragmentation of (N-acetyl-His)-HNE Michael
(1:1) adduct. 230Scheme 3.4S: Suggested fragmentation of (N-acetyl-Gly-Lys-OMe)-HNE
Schiff base adduct 231Scheme 3.5S: Fragmentation of (N-acetyl-Gly-Lys-OMe)-HNE pyrrole
adduct 231Scheme 3.6S: Fragmentation of (N-acetyl-Cys)-(N-acetyl-Cys) disulphide
adduct 234Scheme 3.7S: Fragmentation of (N-acetyl-Cys)-HNE Michael adduct 234Scheme 3.8S: Suggested fragmentation of (N-acetyl-His)-HNE 1:2 adduct 236Scheme 3.9S: Suggested fragmentation of (N-acetyl-His)-HNE 1:2 adduct 236

xxvii
Acknowledgements
I wish to express my deepest sense of respect and gratitude to Dr. Robert G.
Salomon for his expert guidance, constant encouragement, and advice that helped me to
become a professional researcher over the years.
I also wish to express my deepest thanks to Dr. John W. Crabb, Cole Eye
Institute, Cleveland Clinic Foundation, for giving me a chance to work in his lab and
helped me in learning basic protein chemistry and mass spectrometry. I am indebted to
Dr. Sanjoy K. Bhattacharya for his expert guidance, many insightful discussions and
importantly friendship. He taught me basic molecular biology and related biological
techniques that has enormously helped in completing two projects that are in chapter 4 &
5. Special thanks also to Dr. Podrez (CCF) and Dr. Eric Carlson for their collaboration as
well as friendship.
I thank all my people in the lab, Nathan (my buddy), Xiaorong Gu, Jaiyin Gu,
Bharathi, Jack, Karen and Bogdan for for their friendship and enormous help.
I would like to thank my labmates at CASE - Jim laird, Wujuan, Liang Lu, Xi
Chen, Wei Li, Xiaodong Gu and Jaewoo for their help on many situations and for their
friendship.
I would like to dedicate this thesis to my parents as this could be virtually
impossible without their hard work and sacrifices.

xxviii
LIST OF ABBREVIATIONS AND ACRONYMS
Abbreviations and Acronyms Equivalent AA arachidonic acid AA-PC arachidonyl phosphatidylcholine AcOH acetic acid AD Alzheimer’s disease AGE advanced glycoxidation end products ALS amyotrophic lateral sclerosis AMD age-related macular degeneration Apo B apolipoprotein b APT attached proton test AS atherosclerosis BHT butylated hydroxytoluene BSA bovine serum albumin CapLC capillary liquid chromatography CCD charge coupled device CDCl3 deutrated chloroform CHCA α-cyano-4-hydroxy-cinnamic acid CHCl3 chloroform CHD cyclohexa-1,4-dione CHP 2-(ω-carboxyheptyl) pyrrole Cys cysteine d4-HNE [8,8,9,9-2h4]-4-hydroxynonenal DAPI 4', 6-diaminidophenylindole DCC dicyclohexylcarbodiimide DHA-PC 1-palmitoyl-2-docosahexaenoyl-sn-glycero-3-
phosphatidylcholine DMAP 4-dimethylaminopyridine DMF N,N-dimethyl formamide DMP dimethyl pimelimidate DTPA diethylenetriaminepentaacetate DTT dithiotheoritol EDTA ethylenediaminetetraacetate EI electrospray ionization ESI-MS electrospray ionization mass EtOAc ethyl acetate

xxix
FAB fast atom bombardment GFAP glial fibrillary acidic protein GSH glutathione HHE 4-hydroxy-2-hexenal HNE (E)-4-hydroxy-2-nonenal HPLC high performance liquid chromatography HRMS high resolution mass spectrometry HSA human serum albumin HSP heat shock protein Hz hertz IACUC institutional animal care and use committees IEF isoelectric focusing IgG immunoglobin g TGF- βIGH3 transforming growth factor β inducing gene h3 IL-18 interleukin-18 IOP intraocular pressure IP immunoprecipitation IPD iris pigmentary dispersion IPG iris pigmentary glaucoma ISA iris stromal atrophy iso[4]LGE2 iso[4]levuglandin E2
iso[n]LGs iso[n]levuglandin(s) isoLG(s) isolevuglandin(s) isoLGE2 isolevuglandin E2
ITIC integrated total ion current J hyperfine coupling constant kDa kilo Dalton KHdiA-PC 1-palmitoyl-2-(7-carboxy-4-oxohex-5-enoyl)-sn-
glycero-3-phosphatidylcholine KLH keyhole limpet hemocyanin KODA-PC 1-palmitoyl-2-(9-oxo-12-oxododec-10-enoyl)-sn-
glycero-3-phosphatidylcholine KYN kyneurinin LA linoleic acid LC liquid chromatography LC-ESI-SIR liquid choromatograpy electrospray ionization selected
ion recording LC-MS liquid choromatograpy mass LDL low-density lipoprotein LG levuglandins MALDI matrix assisted laser desorbtion ionization MALDI-TOF matrix assisted laser desorption ionization-time of flight

xxx
MDA malondialdehyde Me methyl MeOH methanol MG methylglyoxal MHz megahertz MMP-1 matrix metalloproteinases MRM multiple reaction monitoring MS/MS tandem mass spectrometry MW molecular weight NAC N-acetyl cysteine NAH N-acetyl histidine NAL N-acetyl lysine NBS N-bromosuccinimide NDRI National Disease Research Interchange NF-kB necrosis factor –kb NGL N-acetyl glycine-lysine methyl ester NL nonlinear NMR nuclear magnetic resonance OCT optimal cutting temperature OHKYN 3-hydroxykynurenine ON optic nerve oxLDL oxidized low density lipoprotein oxPC oxidized phosphatidylcholines p probability pAb polyclonal antibody PAF platelet activation factor PBS phosphate buffered saline PC phosphatidylcholine PCC pyridinium chlorochromate PDC pyridinium dichromate PDI-A3 protein disulphide isomerase a3 PG prostaglandins PGH2 prostaglandin H2
PI4K phosphoinisitol-4-kinase POAG primary open angle glaucoma ppm parts per million PPTS pyridinium p-toluenesulfonate PUFAs polyunsaturated fatty acid(s) PVDF polyvinylidene fluoride Q-TOF quadrapole time-of-flight Rf retention factor

xxxi
ROS reactive oxygen species RP-HPLC reverse phase high performance liquid SDS-PAGE sodium dodecyl sulfate-polyacrylamide gel SIR selected ion recording SOD superoxide dismutase SPE solid phase extraction TBAF tetrabutylammonium fluoride TBARS thiobarbituric acid-reacting substance TBDMS tert-butyldimethylsilyl TBDMSCl tert-butyldimethylsilyl chloride t-BHT tert-butylated hydroxytoluene TBS tris buffered saline TBST 20 mM tris, 150 mm NaCl, ph 7.5, 0.5% tween 20 t-Bu t-butyl TGF transforming growth factor THF tetrahydrofuran TIC total ion chromatogram TLC thin layer chromatography TM trabecular meshwork TNF tumor necrosis factor UV ultraviolet

xxxii
Lipid-Based Oxidative Protein Modifications in Glaucoma
Abstract
by
Annangudi Palani Suresh Babu
We and others have postulated that oxidative protein modifications, including
covalent crosslinks, may accumulate in trabecular meshwork (TM) and contribute to
impaired aqueous outflow and primary open angle glaucoma (POAG). To test this
hypothesis, human TM from normal and POAG donors was probed with antibodies for
oxidative protein modifications. Studies using SDS PAGE and Western blotting showed
elevated levels of covalent protein modifications derived from lipid oxidation products
(iso[4]Levuglandin E2 (isoLGE2) and 4-hydroxy-nonenal (HNE)), an advanced glycation
end product (methylglyoxal), and a tryptophan oxidation product (3-hydroxy-
kynurenine), in POAG compared to age-and-gender matched controls.
Immunoprecipitation (IP) using iso[4]LGE2 and HNE antibodies showed the presence of
several apparently crosslinked proteins in POAG donor TM. 2D PAGE Western analyses,
also showed proteins with altered molecular weights and pIs, implying modified and/or
crosslinked proteins. Immunohistochemistry showed that iso[4]LGE2 and HNE
modifications were localized to the TM. Thus, the study provides direct evidence for
lipid-based oxidative modification of TM proteins in POAG. Additionally, increased
levels of iso[4]LGE2 in the TM of the DBA/2J mouse model of glaucoma was also

xxxiii
established and several putative crosslinked proteins were identified using IP and mass
spectrometry. These results support a role for levuglandins (LGs) in POAG pathology.
LGs are known to avidly bind with and crosslink proteins.
The inability of macrophages to processes modified proteins contributes to “foam
cell” formation and atherosclerosis (AS). To identify the modified proteins in inflamed
macrophages, lipopolysaccharide (LPS) stimulated macrophages were studied. LPS
stimulation increased the levels of LGE2 modified proteins in macrophages.The modified
proteins were identified by 2D PAGE and mass spectrometry. Many of the modified
proteins are involved in cholesterol trafficking, gene expression, and/or lipid efflux. This
suggests a role for LGE2 in AS. These observations inspired a pilot study to explore the
possible role of LGs in LPS-induced inflammation of cornea in vivo. We found that LPS
promotes the generation of LGs in cornea.
An efficient synthesis of the γ-keto-α,β-unsaturated alkenoic acid functional array
present in some of the oxidatively truncated phospholipids, was achieved by oxidation of
furyl precursors using NaClO2. Additionally, detection and structural characterization of
multiple HNE adducts onto lysine and histidine side-chain residues were accomplished
using deuterated HNE and mass spectrometry.

Chapter 1
Introduction
1

1.1. Background
Aerobic organisms use oxygen in processes involving energy metabolism, which
makes them prone to radical induced in vivo oxidative damage. This problem can be
averted by antioxidants and antioxidant enzymes in addition to systems that recycle the
oxidatively damaged molecules (proteins). In conditions that compromise one or more
factors of the antioxidant machinery, the free radical mediated damage may lead to a vast
array of diseased conditions. Aging is one of the important phenomenona that have been
attributed to damage by these radicals in situ.1
Biological membranes consist of lipids that provide structural integrity to cells
and cellular structures. These lipids are also involved in signaling processes across the
membrane, through lipid mediators.2 For example, enzyme mediated peroxidation of
PUFAs is involved in signaling normal physiological functions by generating
prostaglandins and thromboxanes.3,4 Additionally, analogous free radical induced
peroxidation can also generate prostaglandin isomers (isoprostanes) and highly reactive
lipid by-products (levuglandins). These highly reactive lipid by-products are known to
form adducts with side chains of proteins and DNA bases, that are involved in various
pathological processes viz, atherosclerosis,5,6 age related macular degeneration,7
rheumatoid arthritis,8 multiple sclerosis,9 Alzheimer’s disease,10 Parkinson’s disease11
and Alexander’s disease.12
2

Ancient scriptures in Sanskrit have identified the potential for pathological
involvements of lipids in the body.
“Those who eat heavy, cold, and excessively oily foods in excessive quantity and do excessive mental work, suffer from disease of the vessels that nourish the heart”
- The Charaka Samhita (400-200 BC)13 (Translated from Sanskrit)
1.2. HNE as an important lipid peroxidation product. 4-Hydroxy-2-nonenal is one of
the most highly investigated lipid peroxidation products. It has been implicated in a vast
array of disease conditions.14 HNE is considered as one of the established markers for
oxidative stress.15 The ready availability of HNE synthetically and its relatively stable
nature has made the handling and processing easier than other lipid oxidation products,
e.g., LGs and oxidized phospholipids.
HNE is a product of oxidative fragmentation of arachidonyl (AA) or linolenyl
(LA) phospholipid esters. Dr. Herman Esterbauer, who discovered HNE, proposed a
mechanism for the formation of HNE from LA through a dioxetane intermediate from
13-HPODE (Scheme 1.1).16 Fragmentation through the intermediacy of 9-PODE can also
be explained based on the peroxy dioxetane intermediate that is formed as depicted in
Scheme 1.1.17,18 A similar mechanism may be proposed for the formation of HNE from
AA-PC, a ω-6 fatty acid. However, there is no evidence to support the actual
involvement of such a mechanism in the generation of HNE through auto-oxidation of
PUFAs. Numerous other mechanisms, also untested, have been proposed for the
formation of HNE in vivo.19-22
3

HOOCC5H11
C5H11HOOC HOOC C5H11
HOOC C5H11
OHO
13-HPODE
HOOC C5H11
OHO
O O
HOOCO
C5H11
OHO
O
C5H11
OH
O
HOOCC5H11
911
13
LA
H
+
L
LH
O2
O2
dioxetene fragmentation
Oxononanoic acid HPNE
HNE
C5H11HOOCO O
9-PODE
C5H11HOOCO O
9,10-dioxetane
O2
O2
LLH
Scheme 1.1: Proposed mechanisms for the formation of HNE from LA.
The pathological activities of HNE are believed to stem from its bifunctional
nature, i.e., the α,β-unsaturated aldehyde and the alcohol. The β carbon of the α,β-
unsaturated aldehyde group acts as an electrophilic center and thus a potential target for
the nucleophilic amino acid residues of proteins. Nucleophilic side chain residues of
4

lysine, histidine and cysteine are known to form Michael adducts with HNE (Scheme
1.2).23 Apart from the Michael adducts, the ε-amino group of lysine can react with the
aldehyde of HNE to form a Schiff base adduct. Both the Michael adduct and the Schiff
base adduct are formed reversibly with lysyl residues.
OOH
4-HydroxynonenalN
OOH
NH
OOH
N
Lysine Michael adduct
Histidine Michael adduct
Histidine
Lysine
S
OOH
Cysteine Michael adduct
Cysteine
OHN
Lysine
NLysine
LysineN
OHLysine N
Lysine-Lysine crosslink
Lysine Schiff base adduct
Lysine pyrrole adduct
Scheme 1.2: Adducts formed by HNE with amino acid side chain residues.
In the case of the Schiff base, the adduct can undergo enol-keto tautomerization
followed by nucleophilic attack of the imine on the carbonyl carbon, resulting in a
cyclized product that undergoes dehydration and concomitant aromatization to form a
pyrrole (Scheme 1.3, next page).24 The HNE-pyrrole adduct is a relatively stable “end
product” of oxidative protein modification mediated by HNE. Antibodies developed
5

against HNE-pyrrole have been useful for analyzing the levels of proteins modified by
HNE in human plasma.25
RNH2
-H2O
RNH2
n-C5H11O
OH
+ n-C5H11NR
OH
n-C5H11NHR
OH
n-C5H11NHR
O
n-C5H11O
OH
NHR
O OHn-C5H11
NHR
NR
n-C5H11
RN
OHn-C5H11
[O]
n-C5H11NHR
O
n-C5H11NHR
O
NHR
n-C5H11NHR
OH
NHR Pyrrole adduct
Michael Adduct
Schiff base Adduct
[O]n-C5H11
NHRO
NH2R
N
NHR
OHR
R
N
NH2R
OHR
R
Amine-HNE (2:1) crosslink
Scheme 1.3: Proposed mechanism for the formation of Michael adduct, Schiff base,
amine-HNE (2:1) crosslink adduct and pyrrole adduct.24
Lipid oxidation products incorporating a phospholipid group, that are analogous
to HNE, with a γ-hydroxy α,β-unsaturated aldehyde core structure, can be formed from
AA-PC and LA-PC. Oxidative fragmentation of AA-PC generates HOOA-PC and LA-
PC generates HODA-PC (Scheme 1.4). These phospholipids occur intact in vivo.
HOOA-PC and HODA-PC can react with lysine residues as described for HNE and form
2-(ω-carboxypropyl)pyrrole (CPP) and 2-(ω-carboxyheptyl)pyrrole (CHP) respectively
(Scheme 1.4) which have lost the PC ester functionality.
6

ROOCC5H11
ROOC
C5H11OO
9-PODE-PC
HOOCOH
ROOCC5H11
9 1113
LA-PC
H
O2
R = 3-lysophosphatidylcholine
O
HODA-PC
PLA2
ROOC
C5H11
9
AA-PC
ROOCO
O
HOOC OOH
HOOA-PC
PLA2
COOH
NH2ProteinNH2Protein
NProteinNProtein
COOH
Carboxypropylpyrrole (CPP) Carboxyheptylpyrrole (CHP)
ROOC
O2
5-PETE-PC
Scheme 1.4: Free radical mediated oxidation of AA-PC and LA-PC leads to the
formation of HOOA-PC and HODA-PC. These lipids are analogous to HNE. They
subsequently react with lysine side chain residues to form the corresponding pyrrole
adducts, 2-(ω-carboxypropyl) pyrrole (CPP) and 2-(ω-carboxyheptyl) pyrrole (CHP).
Antibodies developed against the CPP modification were useful for
demonstrating a considerable increase in the amounts of CPP in patients with renal
failure and atherosclerosis.26 Analogous oxidative cleavage of docosohexanoic acid
7

(DHA) generates the 2-(ω-carboxyethyl)pyrrole (CEP) that has been implicated in age
related macular degeneration.27,28
Since lysine residues can reversibly form a Schiff base with the aldehyde group
of HNE or a Michael adduct at the β-carbon of HNE, HNE can mediate the formation of
lysine-lysine crosslinks. Such crosslinks were identified and characterized through model
studies.24,29,30 Additionally, monoclonal antibodies were generated against the crosslinks
by Dr. Uchida’s group. These antibodies were used to detect the presence of the adducts
in atherosclerotic lesions from human aorta.31 A suggested mechanism for the formation
of the crosslink adducts is depicted in Scheme 1.3 (see page 5).24
1.3. Levuglandins – discovery, formation and pathology. Arachidonyl phospholipids
(AA-PC), ω-6 fatty acid esters, are precursors to a vast array of biologically active
oxidized by-products through enzyme (cyclooxygenase (COX) or lipoxygenase) or free
radical mediated processes. The enzyme-mediated peroxidation requires the free fatty
acid form, while the free radical pathway operates on esters as well.32 Spontaneous
rearrangements of the prostaglandin endoperoxide, PGH2, generate PGD2 and PGE2.
8

COOH COOH
C5H11
OO
OOH
COX
AA-PC Prostaglandin endoperoxide,PGG2
COOH
C5H11
OO
OHPGH2
hydroperoxidase
Scheme 1.5: Formation of prostaglandin endoperoxides from arachidonic acid.
Dr. Salomon discovered an alternative pathway through which these highly
reactive endoperoxides could also rearrange to levulinaldehydes (γ-ketoaldehydes) with
prostanoid side chains. The rearrangement is triggered by abstraction of the electron rich
bridgehead hydrogen of the endoperoxide by the incipient electron deficient methylene
group and a subsequent rearrangement of bonds (as shown in Scheme 1.6) results in
levuglandins (LGs).
OOO
O
OO
H HOH
H
R1
R2
R1
R2
H
R1
R2δ−
δ+
Scheme 1.6: Formation of levuglandins from prostaglandin endoperoxides.
LGs isomers that are generated by free radical mediated pathways are referred to as
isolevuglandins.
9

C5H11
(CH2)3COOH
C5H11
(CH2)3COOPC
OO
(CH2)3COOPC
OH
C5H11
OHC
(CH2)3COOH
OH
C5H11
O
AA-PC
C5H11
(CH2)3COOHOO
OH
C5H11
(CH2)3COOH
OHCOH
O
Levuglandin E2 Isolevuglandin E2
Cyclooxygenase
Phospholipase A2
Free radical-induced
Scheme 1.7: Representative pathways showing a difference in enzyme and free radical
mediated pathways generating LGs from AA-PC.
OO C15H31
O
OP
ON(CH3)3
O OOC5H11
710
13
OR
OC5H11
710
13
OR
OC5H11
710
13
OR
OC5H11
710
13
(CH2)3COOR
C5H11
710
13
OO
(CH2)3COOR
C5H11
710
13
OO
OO
(CH2)3COORC5H11
OO
10 128
(CH2)3COOR8O
O1012 (CH2)3COOR7
10
C5H11O
O 13
(CH2)3COOR710
C5H11OO
(CH2)3COORC5H11
OO
108
OO
(CH2)3COOROO C5H11
OO
OO
(CH2)3COOR
C5H11
OH
OO
8
9
[H][H]
(CH2)3COOR
OH
OO
OO
OH
(CH2)3COORC5H11
iso[4]-PGH2-PC iso[10]-PGH2-PCiso[7]-PGH2-PC
OO
(CH2)3COOR
OH
C5H11
8-epi-LGE2-PC
[H][H]
Arachidonyl phosphatidylcholine ester
(CH2)3COOR
OHOHC
OHCOHC
OH
(CH2)3COOR(CH2)3COOR
OH
C5H11
OOO
C5H11
iso[4]-LGE2-PC iso[10]-LGE2-PCiso[7]-LGE2-PC
OHC
(CH2)3COOR
C5H11
OH
O
12
8-epi-LGE2-PC
8
9
Scheme 1.8: Formation of levuglandins from arachidonyl phosphatidylcholine esters by
free radical mediated processes.
10

The γ-ketoaldehyde functionality of the LGs makes them highly reactive towards
nucleophiles present in the biological system. Crosslinking of proteins is one of the
important features that have been implicated in pathological conditions, for e.g.,
Alzheimer’s disease (AD), which is also associated with increase in the expression of
COX enzymes. LGs are also known to crosslink proteins orders of magnitude greater
than other products of AA-PC oxidation (MDA and HNE) and additionally, the possible
generation of LGs through the COX pathway makes them a possible suspect in AD. A
possible mechanism of crosslink formation by LGs with proteins is depicted in Scheme
1.9. The initial Schiff base adduct formed by the LGs can cyclize and undergo
dehydration and form pyrroles. These pyrrole can also be oxidized to electrophilic
intermediates33 which can undergo crosslinking with other nucleophilic side chain
residues of the protein or the pyrrole itself to form bis-pyrrole adducts.34 Alternatively, an
aminal crosslinking may be generated.
11

O
OHC C5H11
(CH2)3COOH
OH
NH
H2O
Protein
NH2Protein
Protein
NProtein
NR2
R1O
R1
R2
ProteinNH
R2
R1O
NR1
R2
OH
Protein
Protein
NH2Protein
Iso[4]levuglandin E2
H2O
IsoLGE - pyrrole adductProtein-protein aminal crosslink
NH
NProteinR1
R2
N
R1
R2Protein
Pyrrole-pyrrole crosslink
NProteinR1
R2
N
R2
R1Protein
OR
Scheme 1.9: Formation of isolevuglandin and levuglandin based protein adducts.
LGs react with DNA and form DNA-protein crosslinks. These repair-resistant
DNA-protein crosslinks were shown to cause cell death in Chinese hamster fibroblasts.35
Recently, mass spectrometric identification of adducts formed by LGs that were present
12

in brain tissues of patients with AD was reported.36 In this study proteins from brain
tissue were digested to individual amino acids and analyzed by tandem LC-MS/MS and
selective reaction monitoring (479.3 332.1) (Scheme 1.10) as demonstrated earlier in
cell culture studies.37
C5H11
(CH2)3COOH
OH
H3N
OHO
N
O
(CH2)3COOHH2N
O
m/z 479.4
m/z 332.1
(CH2)3COOHHN
HO m/z 332.1
Scheme 1.10: Daughter ion generated by LG-lysine lactam in the mass spectrometric
ionization process. This feature was used in tandem MS/MS to identify the LG modified
proteins.36,37
1.4. Anatomy of anterior section of the eye. A portion of this thesis involves lipid-
derived oxidative modifications of the trabecular meshwork (TM) of the eye. The
anatomy of the eye can be best described by dividing transversely through the lens into
anterior and posterior segments. The anterior segment consists of the cornea, iris and the
anterior chamber. The posterior segment contains the retina, optic nerve and the vitreous
chamber (Figure 1.1, see next page). As background for our study involving the TM, the
anterior chamber is described here in detail.
13

Figure 1.1: Anatomy of human eye showing the anterior and posterior regions
(Copyright© permission obtained from www.nei.nih.gov).
Figure 1.2: Artistic rendering of anterior section of the eye, showing pathway of
aqueous flow through the trabecular meshwork (arrow). (Copyright© permission
obtained from www.nei.nih.gov)
The aqueous humor is actively secreted by the ciliary body and flows into the
anterior chamber through the pupil. The aqueous humor leaves the anterior chamber
14

through an extensive meshwork of tissues leading to the episcleral veins. Aqueous humor
serves two important functions viz. nourishing the cornea and maintaining its shape. The
amount of aqueous humor entering the eye should be equal to the amount flowing out in
order to maintain the pressure in the eye (intraocular pressure). Any decrease in the
outflow will cause an increase in the intraocular pressure (IOP) leading to a serious
blinding disease called glaucoma (vide infra). There are two main pathways of aqueous
drainage. The major pathway is through the trabecular meshwork into the episcleral veins
through the canal of Schlemm. The minor pathway is the uveoscleral pathway, in which
the aqueous passes through the ciliary muscles into the spaces between the cilary body
and sclera leading to the veins. Another pathway through the iris is described in rabbits,
which lack the canal of Schlemm. This pathway is absent in humans.
1.5. Glaucoma. Glaucoma refers to a group of eye diseases that often occur with increase
in the intraocular pressure leading to irreversible blindness and optic neuropathy.38
Glaucoma is a leading cause of blindness.39 A common system of classification and
definition of glaucomas has not yet been defined. But broadly, glaucomas can be divided
into two broad categories, primary and secondary. Primary glaucoma is a diagnosis of
exclusion, which refers to those conditions where no known potential cause(s) can be
attributed to the disease. Secondary glaucoma are often associated with an injury,
inflammation or a previous illness. These two classes can be further classified as open
angle (OAG) and angle closure glaucoma. This classification is based on the iridial angle,
defined by the angle formed by the tangents of iris and cornea, i.e., around 40o (normal
angle) for open angle glaucoma and around 15o for angle closure glaucoma.
15

1.5.1. Primary open angle glaucoma (POAG). Affecting around 3 million Americans,
this form of glaucoma has no symptoms or early warning signs. POAG is often, but not
always, associated with an increase in the fluid pressure inside the eye (intraocular
pressure, IOP) and involves optic nerve damage leading to irreversible blindness. As the
iridial angle is normal, the aqueous pathway is clear up to the trabecular meshwork (TM),
and it is thought that there could be a clogging of the aqueous drainage in the TM region
or beyond. This type of glaucoma develops slowly and sometimes without noticeable
sight loss for many years. It usually responds well to medication, especially if caught
early and treated.40
1.5.2. Secondary glaucomas. There are several forms of secondary glaucoma, classified
either as open angle or closed angle as described above.
Pseudoexfoliative glaucoma. In this condition, thin flaky dandruff-like material peels off
the outer layer of the lens within the eye and collects in the iridial angle and can clog the
drainage system of the eye, causing eye pressure (IOP) to rise. Treatments include
medication or surgery.
Pigmentary glaucoma. This form of open angle glaucoma has midperipheral iris
transillumination defects and is characterized by heavy pigmentation of the TM. The TM
is pigmented due to the release of pigment granules from the heavily pigmented iris into
the aqueous humor, which then can clog the outflow pathway causing the pressure (IOP)
to rise. The etiology of the disease is poorly understood and the primary causes are still
unknown.41
Traumatic glaucoma. Injury to the eye may cause this type of glaucoma known as
secondary glaucoma, which may also be caused by post injury trauma years later.
16

Neovascular glaucoma. This form of glaucoma is caused by formation of abnormal blood
vessels in the iris or in the drainage canals of the anterior chamber resulting in increased
IOP. Neovascular glaucoma is often associated with other abnormalities, e.g., diabetes.
This form of glaucoma is difficult to treat.38
Iridocorneal endothelial syndrome. This rare form of glaucoma usually appears in only
one eye, rather than both. Cells on the back surface of the cornea spread over the eye’s
drainage tissue and across the surface of the iris, increasing IOP and damaging the optic
nerve. These corneal cells also form adhesions that bind the iris to the cornea, further
blocking the drainage channels.38
1.6. Animal models for glaucoma. Use of animal models to study the molecular and
genetic basis of disease processes provides economical and functional feasibility.42 For
example, differences in the anatomical and physiological features across different species
of animals can broaden the understanding of an experimental outcome, which is not
possible using cell culture or in vitro experiments. A part of this thesis involves use of the
DBA/2J mouse, model of glaucoma, to assess the oxidative damage to the TM in
glaucoma. The pathological mechanisms causing glaucoma in the DBA/2J mouse are
incompletely understood but may be associated with pigmentary dispersion.
In glaucomas, increase in IOP is often implicated in optic neuropathy and
progression of vision loss. The biochemical changes that follow the increase in IOP have
been studied by inducing an increase in the IOP by several mechanisms.42 One of the
commonly used procedures is treating rodent models with hypertonic saline injections
into episcleral veins. This causes an increase in the IOP and subsequent damage to the
17

optic nerve head.43 Use of adrenalin, which induces β-adrenoreceptors, in the
development of experimental glaucoma in rabbits has been reported.44 Adrenalin triggers
a cascade of cellular signaling that causes dystrophic changes in ocular blood vessels and
tissues (including those in the drainage zone), mainly due to excessive entry of Ca2+ into
cells, which are typical symptoms of primary open angle glaucoma in humans.44 IOP
increases have been induced in Rhesus monkeys by cauterization, using an argon laser, in
the anterior chamber angle leading to clogging of the aqueous flow.45,46 α-Chymotrypsin
induced ocular hypertension models in various animals viz, monkey, rabbit and dog, have
been described.42,47 The effects in this model are ascribed to TM blockage by
inflammatory exudates and by debris resulting from enzymatic hydrolysis of the zonules
(sensory ligaments holding the lens).
1.7. Glaucoma and oxidative stress. Increase in oxidative stress has been suggested to
be an important factor in glaucoma pathogenesis. Increased levels of lipid peroxidation
products in the TM of primary open angle glaucoma (POAG) were noted as early as 1989
by Babizhayev et al., through experiments that spectrophotmetrically quantitate the
amount of dienes present in TM lipid extracts.48 Aging, an important factor that has been
long associated with decrease in antioxidant capacity and increase in aggregation of
protein due to oxidation, has also been associated with glaucoma.49 Clinical evidence
supporting this view was substantiated by reports that show the loss of cellularity of TM
cells50 and loss of superoxide dismutase (SOD) activity,51 an antioxidant enzyme, in TM
tissues with aging. Additional evidence obtained through TM cell culture studies, showed
18

decreased amounts of reduced glutathione (GSH)52 and lowered cell adhesion
characteristics53 when the cells were treated with H2O2.
Protein expression is altered in diseased conditions as a homeostatic mechanism
to prevent any damage to the tissues. In the cases of human and monkey TM tissue, there
is an over expression of alpha B-crystallin in response to stress, to prevent any cellular
damage.54 Extracellular matrix (ECM) remodeling in the TM tissue of glaucoma patients
causes an increase in the levels of collagen deposition, which can in turn cause an
increase in the outflow resistance.55
Reactive oxygen species (ROS) are generated in the anterior chamber
photochemically giving rise to reactive substances, e.g., lipid peroxides.56 A decrease in
antioxidant levels of the lacrimal secretions in patients with POAG have also been
reported as the disease progresses.57 A decrease in the levels of plasma GSH58 and an
increase in the lipid peroxidation product, malonaldehyde59 have been observed in
plasma of POAG. These reactive substances have been known to play a role in cataract, a
disease associated with oxidation of lens proteins.60 The aqueous humor is suggested to
be one of the important routes through which ROS may reach the TM, which can act on
the ECM proteins and the membranes.61 TGF-β, a fibrogenic cytokine involved in
atherosclerotic and pulmonary fibrogenesis, is also fibrogenic in TM cells.62 Increase in
the TGF-β levels in the aqueous humor may influence the cytoskeletal structures and
cellular signaling cascades.63 A review62 of the effects of vitamin E deficiency describes
various components that are involved in the pathology in POAG and how they influence
each other (Figure 1.3, next page) .
19

Figure 1.3: Flow chart illustration of the components involved in the pathogenesis of
POAG induced by decrease of vitamin E in the ciliary body. MMP – matrix
metalloproteinases, TIMP – tissue inhibitor of metalloproteinases, ON – optic nerve,
ECM – extracellular matrix. (Copyright permission obtained from Br. J. Nutrition, CABI
Publishing)62
Elevated levels of 8-hydroxy-2'-deoxyguanosine (8-OH-dG), a marker for
oxidative DNA damage, have been found in TM tissues of POAG patients.64 This study
also revealed that there is an increase in 8-OH-dG levels in glutathione S-transferase
20

(GSTM) null subjects (GSTM is a gene corresponding to the enzyme involved in
antioxidant activity of GSH).64 Increase in oxidative stress in the retinal ganglion cell65
and stress related enzymes in the optic nerve head62,66 was observed in animal glaucoma
models. Rescue of retinal ganglion cells in rats having increased IOP has been successful
by using tropic factors and antioxidants.67
“M: Too many free radicals. That's your problem. James Bond: "Free radicals," sir? M: Yes. They're toxins that destroy the body and the brain, caused by eating too much red meat and white bread and too many dry martinis! James Bond: Then I shall cut out the white bread, sir. M: Oh, you'll do more than THAT, 007. From now on you will suffer a strict regimen of diet and exercise; we shall PURGE those toxins from you! James Bond: Shrublands? M: You got it!”
- From “Never say never again” (1983).
21

1.8. References.
(1) Finkel, T.; Holbrook, N. J. Nature 2000, 408, 239-47. (2) Molecular and Cellular Basis of Inflammation; Serhan, C. N.; Ward, P. A., Eds.;
Humana Press: Totowa, New Jersey, 1999. (3) Chakraborti, T.; Ghosh, S. K.; Michael, J. R.; Batabyal, S. K.; Chakraborti, S. Mol
Cell Biochem 1998, 187, 1-10. (4) Tapiero, H.; Ba, G. N.; Couvreur, P.; Tew, K. D. Biomed Pharmacother 2002, 56,
215-22. (5) Brown, M. S.; Goldstein, J. L. Annu Rev Biochem 1983, 52, 223-61. (6) Arlt, S.; Kontush, A.; Muller-Thomsen, T.; Beisiegel, U. Z Gerontol Geriatr
2001, 34, 461-5. (7) Kopitz, J.; Holz, F. G.; Kaemmerer, E.; Schutt, F. Biochimie 2004, 86, 825-31. (8) Winyard, P. G.; Tatzber, F.; Esterbauer, H.; Kus, M. L.; Blake, D. R.; Morris, C.
J. Ann Rheum Dis 1993, 52, 677-80. (9) Newcombe, J.; Li, H.; Cuzner, M. L. Neuropathol Appl Neurobiol 1994, 20, 152-
62. (10) Sayre, L. M.; Zelasko, D. A.; Harris, P. L.; Perry, G.; Salomon, R. G.; Smith, M.
A. J Neurochem 1997, 68, 2092-7. (11) Yoritaka, A.; Hattori, N.; Uchida, K.; Tanaka, M.; Stadtman, E. R.; Mizuno, Y.
Proc Natl Acad Sci U S A 1996, 93, 2696-701. (12) Castellani, R. J.; Perry, G.; Harris, P. L.; Cohen, M. L.; Sayre, L. M.; Salomon, R.
G.; Smith, M. A. Brain Res 1998, 787, 15-8. (13) http://www.ayurveda-ayurvedic.com/e-zine/health-ezine-404.html, Volume 1,
Issue 4. April 2004. (14) Zarkovic, K. Mol Aspects Med 2003, 24, 293-303. (15) Zarkovic, N. Mol Aspects Med 2003, 24, 281-91. (16) Esterbauer, H.; Schaur, R. J.; Zollner, H. Free Radic Biol Med 1991, 11, 81-128. (17) Gardner, H. W.; Deighton, N. Lipids 2001, 36, 623-8. (18) Schneider, C.; Tallman, K. A.; Porter, N. A.; Brash, A. R. J Biol Chem 2001, 276,
20831-8. (19) Schneider, C.; Tallman, K. A.; Porter, N. A.; Brash, A. R. J. Biol. Chem. 2001,
276, 20831-20838. (20) Lee, S. H.; Blair, I. A. Chem Res Toxicol 2000, 13, 698-702. (21) Noordermeer, M. A.; Feussner, I.; Kolbe, A.; Veldink, G. A.; Vliegenthart, J. F.
Biochem Biophys Res Commun 2000, 277, 112-6. (22) Pryor, W. A.; Porter, N. A. Free Radic Biol Med 1990, 8, 541-3. (23) Uchida, K. Amino Acids 2003, 25, 249-57. (24) Xu, G.; Sayre, L. M. Chem Res Toxicol 1998, 11, 247-51. (25) Kaur, K.; Salomon, R. G.; O'Neil, J.; Hoff, H. F. Chem Res Toxicol 1997, 10,
1387-96. (26) Salomon, R. G.; Kaur, K.; Podrez, E.; Hoff, H. F.; Krushinsky, A. V.; Sayre, L.
M. Chem Res Toxicol 2000, 13, 557-64. (27) Crabb, J. W.; Miyagi, M.; Gu, X.; Shadrach, K.; West, K. A.; Sakaguchi, H.;
Kamei, M.; Hasan, A.; Yan, L.; Rayborn, M. E.; Salomon, R. G.; Hollyfield, J. G. Proc Natl Acad Sci U S A 2002, 99, 14682-7.
22

(28) Gu, X.; Meer, S. G.; Miyagi, M.; Rayborn, M. E.; Hollyfield, J. G.; Crabb, J. W.; Salomon, R. G. J Biol Chem 2003, 278, 42027-35.
(29) Xu, G.; Liu, Y.; Sayre, L. M. Chem Res Toxicol 2000, 13, 406-13. (30) Itakura, K.; Osawa, T.; Uchida, K. J Org Chem 1998, 63, 185-187. (31) Itakura, K.; Oya-Ito, T.; Osawa, T.; Yamada, S.; Toyokuni, S.; Shibata, N.;
Kobayashi, M.; Uchida, K. FEBS Lett 2000, 473, 249-253. (32) Salomon, R. G. Chem Phys Lipids 2005, 134, 1-20. (33) Amarnath, V.; Valentine, W. M.; Amarnath, K.; Eng, M. A.; Graham, D. G. Chem
Res Toxicol 1994, 7, 56-61. (34) DiFranco, E.; Subbanagounder, G.; Kim, S.; Murthi, K.; Taneda, S.; Monnier, V.
M.; Salomon, R. G. Chem Res Toxicol 1995, 8, 61-7. (35) Murthi, K. K.; Friedman, L. R.; Oleinick, N. L.; Salomon, R. G. Biochemistry
1993, 32, 4090-7. (36) Zagol-Ikapitte, I.; Masterson, T. S.; Amarnath, V.; Montine, T. J.; Andreasson, K.
I.; Boutaud, O.; Oates, J. A. J Neurochem 2005, 94, 1140-5. (37) Boutaud, O.; Li, J.; Zagol, I.; Shipp, E. A.; Davies, S. S.; Roberts, L. J., 2nd;
Oates, J. A. J Biol Chem 2003, 278, 16926-8. (38) http://www.glaucoma.org/learn/; Vol. 2005. (39) Quigley, H. A. N Engl J Med 1998, 338, 1063-4. (40) Epstein, D. L. Chandler and Grant's glaucoma; 4th ed.; Williams & Wilkins, co.,:
Baltimore, Md, 1997. (41) Sowka, J. Optometry 2004, 75, 115-22. (42) Ritch, R.; Shields, M.; Krupin, T. The Glaucomas; 2 ed.; Mosby: St. Louis, MO,
1996. (43) Morrison, J. C.; Moore, C. G.; Deppmeier, L. M.; Gold, B. G.; Meshul, C. K.;
Johnson, E. C. Exp Eye Res 1997, 64, 85-96. (44) Mikheytseva, I. N.; Kashintseva, L. T.; Krizhanovsky, G. N.; Kopp, O. P.;
Lipovetskaya, E. M. Int Ophthalmol 2004, 25, 75-9. (45) Gaasterland, D.; Kupfer, C. Invest Ophthalmol 1974, 13, 455-7. (46) Gross, R. L.; Ji, J.; Chang, P.; Pennesi, M. E.; Yang, Z.; Zhang, J.; Wu, S. M.
Trans Am Ophthalmol Soc 2003, 101, 163-9; discussion 169-71. (47) Bucolo, C.; Campana, G.; Di Toro, R.; Cacciaguerra, S.; Spampinato, S. J
Pharmacol Exp Ther 1999, 289, 1362-9. (48) Babizhayev, M. A.; Bunin, A. Acta Ophthalmol (Copenh) 1989, 67, 371-7. (49) Babizhayev, M. A.; Brodskaya, M. W. Mech Ageing Dev 1989, 47, 145-57. (50) Alvarado, J.; Murphy, C.; Polansky, J.; Juster, R. Invest Ophthalmol Vis Sci 1981,
21, 714-27. (51) De La Paz, M. A.; Epstein, D. L. Invest Ophthalmol Vis Sci 1996, 37, 1849-53. (52) Kahn, M. G.; Giblin, F. J.; Epstein, D. L. Invest Ophthalmol Vis Sci 1983, 24,
1283-7. (53) Zhou, L.; Li, Y.; Yue, B. Y. J Cell Physiol 1999, 180, 182-9. (54) Tamm, E.; Russell, P.; Johnson, D.; Piatigorsky, J. Invest Ophthalmol Vis Sci
1996, 37, 2402-13. (55) Gonzalez-Avila, G.; Ginebra, M.; Hayakawa, T.; Vadillo-Ortega, F.; Teran, L.;
Selman, M. Arch Ophthalmol 1995, 113, 1319-23.
23

(56) Kurysheva, N. I.; Vinetskaia, M. I.; Erichev, V. P.; Demchuk, M. L.; Kuryshev, S. I. Vestn Oftalmol 1996, 112, 3-5.
(57) Bunin, A.; Filina, A. A.; Erichev, V. P. Vestn Oftalmol 1992, 108, 13-5. (58) Gherghel, D.; Griffiths, H. R.; Hilton, E. J.; Cunliffe, I. A.; Hosking, S. L. Invest
Ophthalmol Vis Sci 2005, 46, 877-83. (59) Yildirim, O.; Ates, N. A.; Ercan, B.; Muslu, N.; Unlu, A.; Tamer, L.; Atik, U.;
Kanik, A. Eye 2004, 1-4. (60) Srivastata, S. K.; Awasthi, S.; Wang, L.; Bhatnagar, A.; Awasthi, Y. C.; Ansari,
N. H. Curr Eye Res 1996, 15, 749-54. (61) Babizhayev, M. A.; Costa, E. B. Biochim Biophys Acta 1994, 1225, 326-37. (62) Veach, J. Br J Nutr 2004, 91, 809-29. (63) Tripathi, R. C.; Li, J.; Chan, W. F.; Tripathi, B. J. Exp Eye Res 1994, 59, 723-7. (64) Izzotti, A.; Sacca, S. C.; Cartiglia, C.; De Flora, S. Am J Med 2003, 114, 638-46. (65) Tezel, G.; Yang, X.; Cai, J. Invest Ophthalmol Vis Sci 2005, 46, 3177-87. (66) Yan, X.; Tezel, G.; Wax, M. B.; Edward, D. P. Arch Ophthalmol 2000, 118, 666-
73. (67) Ko, M. L.; Hu, D. N.; Ritch, R.; Sharma, S. C. Invest Ophthalmol Vis Sci 2000,
41, 2967-71.
24

Chapter 2
A Short and Efficient Synthesis of 4-Oxo-2-alkenoic Acids from 2-Alkylfurans
25

2.1. Background
Polyunsaturated fatty acids (PUFAs) are especially prone to damage by free
radicals owing to the presence of homoconjugated C=C double bonds. The chemistry of
PUFA peroxidation is particularly harmful because the immediate damage may be easily
amplified by the additional release of reactive substances, radicals or fatty acid-derived
aldehydes that can initiate further modification of cellular structures.1-5 However, they
can also be detoxified.6,7
OO C15H31
O
OP
ON(CH3)3
O OO
C4H9 m n
O
On PC
OO
PA-PC, n=3, m=4 (2.1) LA-PC, n=7, m=2 (2.2)
HPOOA-PC, n=3HPODA-PC, n=7
O
On PC
OHO
HO
HOOA-PC, n=3HODA-PC, n=7
O
On PC
OHO
HOdiA-PC, n=3HDdiA-PC, n=7
OH
O
On PC
OO
KOOA-PC, n=3KODA-PC, n=7
O
On PC
OO
KOdiA-PC, n=3 (2.3)KDdiA-PC, n=7 (2.4)
OH
[H]
O2O2
O2
O2
O2
-H2O
O2
Scheme 2.1: Suggested mechanism for generation of KOdiA-PC and KDdiA-PC.8
26

Oxidative modification of low-density lipoprotein and subsequent uptake by
macrophages is an early event in the formation of atherosclerotic plaques.9 Among the
products that are formed by oxidative fragmentation of arachidonyl 2.1 or linoleyl 2.2
phosphatidylcholine (PC), the γ-keto-α,β-unsaturated alkanoate phosphatidylcholine
esters, KOdiA-PC 2.3 and KDdiA-PC 2.4 (Scheme 2.1), are potent ligands responsible
for recognition and uptake of oxidized low-density lipoprotein.10,11
The functional array containing 4-oxo-2-alkenoic acid is also present in many
pharmaceutically important natural products, e.g., pyrenophorin, patulolide, and
grahamimycin that have antifungal, anti-bacterial and/or anti-inflammatory properties.12-
18 To facilitate studies on the biological properties of these compounds, a short and
efficient synthesis was needed to make them readily available.
OO
OO
OOO
OO
O
O
O
O
OH
O
GrahamimycinPyrenophorin Patulolide
Scheme 2.2: Natural products incorporating 4-oxo-2-alkenoate functionality.
2.1.1. Previous syntheses of γ-keto α,β-unsaturated alkanoates
Most of the previous syntheses of these functional arrays either used elaborately
functionalized precursors19-23 or multiple step procedures involving ring opening and
subsequent oxidation (vide infra).
Synthesis using functionalized furans as precursors. Synthesis of 4-oxo-alkenoates
using 2-alkoxy-5-alkylfurans as precursors was achieved previously by using oxidizing
27

agents like bipyridinium chlorochromate,24 followed by isomerization using I2 of the Z-
to E- isomer (Scheme 2.3). Another method, using the hydrogen peroxide adduct of urea
(UHP) in the presence of a catalytic amount of methyltrioxorhenium (MTO) generated
this functional array in good yields.25
OO
1. PCC,CH2Cl2, 2.5h2. I2, 4h
OO
O< 40%
OOO
OO
UHP -Urea-H2O2 adductMTO -Methyltrioxorhenium
UHP5 mol % MTO
95%
ReO
OO
MTO
Scheme 2.3: 2-Alkoxyfuran as precursor for 4-oxo-2-alkenoate.
Photooxidation of 2-acyl-5-alkylfurans using singlet oxygen followed by
reduction was found to generate such products in fair to poor yields (Scheme 2.4).26
OO
O1.1O2, Rose Bengal2.K2CO3, Me2SO4
< 60%, 2 stepsOOHC
O OO O
Scheme 2.4: 2-Acylfuran as precursor for 4-oxo-2-alkenoate.
Lead(IV) acetate mediated oxidation of 2-siloxy-5-alkylfurans and subsequent
acidification yielded 2-alkyl-2-hydroxybutenolides, ring-form tautomers of cis-4-oxo-2-
alkenoates,27 in good yields (Scheme 2.5).28
28

O OSi
OO
Si
O OHOO
H3O+
76% 86%
Pb(OAc)4
Scheme 2.5: 2-Siloxyfuran as precursor for 4-oxo-2-alkenoate.
Based on an earlier method, that used dimethyl dioxirane to oxidatively convert a
4,5-disubstituted furans to γ-keto-enals,29 the use of 2-siloxy-5-alkylfurans30 or 2-silyl-
5-alkylfurans31 as precursors generates hydroxybutenolides in good to excellent yields
(Scheme 2.6).
O OSi
OO
Si
OOHO
O
O OH3O+
100% 77%
OR Si OHOO
R
O O
H3O+
95%OR SiO
Scheme 2.6: 2-Silylfuran and 2-siloxyfuran oxidation using dimethyl dioxirane.
Synthesis using simple furans. Though the use of simple furans to synthesize the 4-oxo-
2-alkenoic acids have been reported, these procedures involved multi-step reactions. For
example, synthesis of (E)-4-oxonon-2-enoic acid (2.6), a natural antibiotic produced by
Streptomyces olivaceus, was achieved Ballini et al.32 by oxidation of 2-pentylfuran (2.5)
using pyridinium chlorochromate followed by Jones reagent gave the product in good
29

yields. However, the procedure is harsh and has limitations in the kind of substrates that
can be used (Scheme 2.7).
OC5H11 C5H11O
OC5H11
OO
OH
PCCJones
reagent70% 85%
2.5 2.6
Scheme 2.7: Oxidation of a 2-alkylfuran using pyridinium chlorochromate.
Use of bromine to oxidize the furan ring has been reported earlier, but gives very
low yields.33-35 Kobayashi et al. modified this procedure and used N-bromosuccinimide
(NBS) to oxidize the furan ring (Scheme 2.8).36,37 This methodology was used by our
group to synthesize some oxidized phospholipids.38
OC5H11 C5H11O
OC5H11
OO
OH
PCCJones
reagent70% 85%
2.5 2.6
Scheme 2.8: Oxidation of 2-alkylfuran using N-bromosuccinimide.
Synthesis of pyrenophorin precursor. Pyrenophorin, a macrocyclic natural product, is
a lactone a of γ-keto-α,β-unsaturated ester of an alkan-7-ol-1-oic acid (Scheme 1.2).
There are many reported methods to synthesize this precursor.15,20,28,36,39-41 Some of
methods are described in Scheme 2.9.
30

a.
OAc
Oa) Jones reagentb) 2-mercaptobenzimidazole
c) HOCH2CH2OH, p-TsOH
O
OHOAc
OO
< 36%
b.
NO2
OAcN
OAc
O
COOMe
COOMeOAc
O OH
COOMeOAc
O
COOMe
PhNCOEt3N80%
H2, Ni-Raneyacetic acid: H2O.
MeSO2ClEt3N
70%
70%
c.
OAc
O
CHOOAc
O
COOHOAc
O
NBS
pyridine
NaClO2
64%
83%
Scheme 2.9: Methods reported for synthesis of a pyrenophorin precursor: a) using Jones
reagent with furyl precursor; b) using (3+2) cycloaddition of primary nitro group and
methyl acrylate;42 c) using NBS to oxidize a furyl precursor.36
31

A patent application reported a one-pot synthesis of 4-oxo-5-phthalimido-2-
pentenoic acid from N-furfurylphthalimide through singlet oxygenation.43 Though this is
a one-step method, the methodology was not substantially exemplified.
O
HO
O
O
NO
O
O
NO O2
hν76%
Scheme 2.10: A patented process for generation of 4-oxo-2-alkenoic acids by Takeya.43
32

2.2. Results and discussion
4-Oxo-2-alkenoic acids, and the closely related α,β-unsaturated-γ-diketones, are
versatile intermediates for organic synthesis.44-52 An important shortcoming of previous
methods for preparing 4-oxo-2-alkenoic acids is the requirement for more elaborately
functionalized furan intermediates such as 2-alkoxy-5-alkyl,24,25 2-acyl-5-alkyl,53 2-
siloxy-5-alkyl28,30 or 2-silyl-5-alkyl31 as precursors (see section 2.1.1). Generation of 4-
oxo-2-alkenoic acids from 2-alkylfurans has also been accomplished previously through a
variety of multi-step procedures involving treatment with reagents such as bromine41 or
NBS36 followed by oxidation with sodium chlorite; PCC followed by Jones reagent;32 or
nitration followed by acidic oxidation (see section 2.1.2).54
Alkylfurans are easily accessible,44-46,55 robust precursors that are compatible with
a wide variety of reagents and reaction conditions making them higly desirable
substrates. The 2-alkylfuran precursors can be prepared by alkylation of 2-furyllithium
using the corresponding alkyl halide as reported earlier (Scheme 2.11).38
O RO t-BuLi O Li RX
Scheme 2.11: Synthetic method for generating 2-alkylfurans from furan.
This study focuses on a efficient stereocontrolled methodology, that expliots
sodium chlorite as an oxidizing agent, for the conversion of 2-alkylfurans 2.7 into 4-oxo-
2-alkenoic acids 2.10 and for the conversion of 2,5-dialkylfurans into 1,4-enediones. The
oxidation cleanly generates cis isomers 2.9 (butenolides 2.8) stereospecifically and in
33

high yield. If desired, these can be cleanly and quantitatively converted to the
corresponding trans isomers by treatment with pyridine.56
O R
2.7 HO RO O
OR
O
OH2.10
OR
OOH
NaClO2 Pyridine2.8
2.9
Scheme 2.12: Synthesis of the 4-oxo-2-alkenoic acid functional array.
Synthesis of 4-oxo-2-alkenoic acids. Oxidation of 2-alkylfurans 2.7 in the presence of
sodium chlorite in a slightly acidic aqueous solution (~ pH 3.5) at room temperature
generates 2-alkyl-2-hydroxy-butenolides 2.8. The 2-alkyl-2-hydroxybutenolides 2.8 are
the ring tautomers of 4-oxo-2(Z)-alkenoic acids 2.9 (Scheme 2.12).27 To confirm this, 2-
pentyl-2-hydroxybutenolide (2.5, R = C5H11) was synthesized (vide infra) and subjected
to various pH conditions and the UV spectrum recorded (Scheme 2.13). The spectrum
shows the appearance of a peak around 240 nm under basic conditions indicating the
predominance of a keto functionality (Figure 2.1).
OC5H11HO
O C5H11 OHO OpH >7
pH <7
Scheme 2.13: Ring-chain tautomers of 4-oxo-2-pentenoic acid.
34

Figure 2.1: UV-spectra of 4-oxo-2-pentenoic acid at varying pH.
Using this furan oxidation protocol, simple 2-alkyl substituted furans were
converted to 4-oxo-alkenoic acids in excellent yields. Compared to the earlier synthetic
methods,32,57 synthesis of 4-oxo-2-nonenoic acid, a natural antibiotic produced by
Streptomyces olivaceus, was prepared conveniently in excellent yields (Scheme 2.14).
C6H13
COOEt COOEt
C5H11
O
COOH
C5H11
O
OR OR
OHO
OC5H11CHO
C5H11
O
CrO3
PCC
NaClO2
Pyridine
K2CO356%
68%
95%100%
70% 85%Jones
R = -CH3, -C5H11
Scheme 2.14: Comparison of synthetic methods used to generate 4-oxo-2(E)-alkenoic
acids from 2-alkylfurans.
35

The generality of the protocol for substituents with commonly used alcohol
protecting groups, as depicted in Scheme 2.15, was established and the results are shown
in Table 2.1.
OPHOOC
O
O OP OOP
OHOa b
a - NaClO2 (3 eq.), NaH2PO4 (1.5 eq.), tBuOH:H20 (5:1), rtb - Pyridine, THF/Acetone/H2O (5:4:1), rt
2.11 2.12 2.13
Scheme 2.15: Synthesis of 4-oxo-2-alkenoic acids from 2-alkylfurans.
Table 2.1: Yield and reaction time for alcohol protected alkylfurans in Scheme 2.15.
Substrate P Yield 2.12
(%)
Reaction time
2.11 2.12 (h)
Yield 2.13
(%)
Reaction time
2.12 2.13 (h) Product
2.14 Ts 90 4 96 2 2.17
2.15 THP 78 2 90 2 2.18
2.16 TBDMS 77 2 95 2 2.19
36

Pyrenophorin, a natural product having antibiotic and antifungal properties, can
be assembled by lactonization of 2.22 as reported earlier.36 This precursor 2.22 was
synthesized using our new furan oxidation protocol (Scheme 2.16) in good yield,
compared to earlier protocols (Scheme 2.9).36
OOTBDMS
OOTBDMS
OHO
OO
OTBDMS
NaClO2
Pyridine
OH
OO
O
OO
O
77%
95%
2.20 2.21
2.222.23
Scheme 2.16: Synthesis of pyrenophorin precursor from 2-alkylfuran 2.20.
Synthesis of oxidatively truncated phospholipids, KOdiA-PC (2.3) and KDdiA-
PC (2.4), for studying their biological properties,10,11 was our primary motivation to
devise an efficient method for the synthesis of 4-oxo-2-alkenoic acid. Our previous
syntheses of 2.3 and 2.4 from 2-substituted furan phospholipid precursors 2.24 and 2.25,
using oxidation with NBS followed by further oxidation with sodium chlorite, gave 46%
and 36% yields, respectively.38 For these phospholipids, the sodium chlorite oxidation
method using t-BuOH-water (5:1) as solvent afforded poor yields, even after long
reaction times. Total syntheses involving phospholipids present special challenges owing
to their unique solubility properties. The direct oxidation of 2.24 or 2.25 with sodium
chlorite can be accomplished by employing a biphasic solvent system containing
37

phosphate buffer (10 mM, pH 3.5), chloroform and water in the ratio 2:1:1. The biphasic
aqueous sodium chlorite protocol decreased the reaction time from 9 h to 1 h, and
consistently gave 40-80% higher yields than the NBS-NaClO2 procedure.
OO
OPC
n
NaClO2 (3 eq.) ,NaH2PO4 (1.5 eq.),tBuOH:H2O (5:1), rt
NaClO2 (3 eq.) PBS (pH 3.5):CHCl3:H2O(2:1:1), rt
< 10%
>75%
O O
OPC
n
HO
O
OOO
OOHPC
Pyridine, THF/Acetone/H2O (5:4:1), rt
> 90%
n
n=2 (2.24)n=6 (2.25)
n=2, KOdiA-PC (2.3)n=6, KDdiA-PC (2.4)
n=2 (2.26)n=6 (2.27)
Scheme 2.17: Synthesis of KOdiA-PC and KDdiA-PC by furan oxidation protocols
employing two different solvent conditions.
Synthesis of 1,4-enediones. Oxidation of 2,5-dialkylfurans 2.28 using the new furan
oxidation protocol generated the 1,4-alk-2-enediones 2.29 (Scheme 2.18).8
R R'O OOR R'
a. NaClO2 ( 3 equiv.), NaH2PO4 ( 1.5 equiv.), tBuOH:H2O (5:1), 93%
a
2.28 2.29
Scheme 2.18: Synthesis of 1,4-enediones from 2,5-dialkylfurans.
38

Substrates incorporating a vinyl substituent present a special challenge. For
example, under the usual reaction conditions, 2.31 did not give any of the corresponding
enedione 2.34. However, in the presence of 2-methylbut-2-ene, a chlorine scavenger,58
the sodium chlorite oxidation protocol gave enedione 2.34 in good yield (Table 2.2).
Table 2.2: Synthesis of 2-ene-1,4-diones from 2,5-dialkylfurans (Scheme 2.16)
Substrate R R’ Product Yield(%)
2.30 -CH3 -CH3 2.33 93b
2.31 -(CH2)7CHO -CH=CH(E)-C4H9 2.34d 76c
2.32 -CH3 -CH=CH(E)-C4H9 2.35 78 c
a NaClO2 (3 equiv.), NaH2PO4 (1.5 equiv.), t-BuOH:H2O (5:1) b Reaction time 1 h c 2-Methyl-2-butene (0.1 mol equiv.) added to the reaction, reaction time ~20
min, yield based on starting material consumed. d The aldehyde is completely oxidized to the acid.
39

2.3. Conclusions
In summary, this new methodology provides ready access to either cis- or trans-
isomers of 4-oxo-2-alkenoic acids from simple 2-alkylfuran precursors in high yields,
using readily available reagents.59 Compared to our previous syntheses, the new
procedure enables a major improvement in the overall yield and ease of preparing
biologically active phospholipids that contain this functional array, thus providing a ready
access for further studies. This protocol is also efficient for converting 2,5-dialkylfurans
to the corresponding 1,4-enediones.
40

2.4. Experimental Procedures
General Methods. Proton magnetic resonance (1H NMR) spectra were recorded on a
Varian Gemini spectrometer operating at 300 or 200 MHz. Proton chemical shifts are
reported in parts per million (ppm) on the δ scale relative to CDCl3 (δ 7.24). Carbon
magnetic resonance (13C NMR) spectra were recorded on a Varian Gemini spectrometer
operating at 50 MHz. Carbon chemical shifts are reported in parts per million (ppm) on
the δ scale relative to CDCl3 (δ 77.0). Thin layer chromatography (TLC) was performed
on glass plates precoated with silica gel (Kiesegel 60 F254, E. Merck, Darmstadt,
Germany). The plates were visualized by viewing under short wavelength UV light or by
treatment with iodine. Flash column chromatography was performed on silica gel (60 Å,
32-63 µm) supplied by Sorbent Technology (Atlanta, GA). UV-visible spectra were
recorded on a Perkin Elmer Lambda 35 UV-visible spectrometer using Fisher Scientific
10 mm cuvette. MALDI-ToF was performed on a Voyager – DE PRO from Applied
Biosystems®, using α-cyano-4-hydroxycinnamic acid (CHCA) as matrix. Chemical were
purchased from Aldrich Chemical Co. (Milwaukee, WI) unless otherwise stated. Solid
phase extraction columns (SPE) were purchased from Whatman Inc, Alabama.
Typical procedure for furan oxidation. To a magnetically stirred solution of NaH2PO4
(1.5 equiv.) and NaClO2 (3 equiv.) in t-BuOH-H2O (5:1, v/v) was added the 2-alkylfuran.
The resulting mixture was stirred at room temperature until the yellow color disappeared
or until the starting material was consumed as determined by TLC. Then the solvent was
removed on a rotary evaporator. The residue can be directly used for the isomerization.
To purify the intermediate formed, the residue is extracted with ethyl acetate, washed
41

with brine, dried over MgSO4 and passed through a short silica gel column using ethyl
acetate as eluent.
Typical procedure for cis/trans isomerization. A catalytic amount of freshly distilled
pyridine was added to a solution (THF/acetone/water 5:4:1) of hydroxybutenolide. The
mixture was stirred for 2 hours at room temperature. Solvents were removed on a rotary
evaporator and a high vacuum pump.
4-Furan-2-ylbutyl-4-toluenesulphate (2.14).
O OTsO OP TsCl, Et3N, DMAP
CH2Cl2, 0 oC
91% 2.142.11, P = H
To a suspension of 2-furylbutan-4-ol38 (33 mg, 0.23 mmol) in CH2Cl2 (3 mL) was
added triethylamine (100 µL, 0.7 mmol), p-toluenesulfonyl chloride (50 mg, 0.26 mmol),
DMAP (3 mg, 0.023 mmol) with stirring at room temperature. When TLC showed no
starting material, 5 mL of dil. HCl (pH 3) was added and the resulting mixture was
extracted with CH2Cl2 (3 x 20 mL). The combined extract was washed with brine, dried
over MgSO4, solvent was removed in vacuo and the residue was purified by column
chromatography with 15% ethyl acetate in hexanes (Rf = 0.4) to yield an oily liquid (62
mg, 91%).60 1H NMR (CDCl3, 300 MHz) δ 7.77 (d, J = 8.4 Hz, 2H), 7.32 (d, J = 8.4 Hz,
2H), 7.25 (dd, J1 = 1.8 Hz, J2 = 0.9 Hz, 1H), 6.24 (dd, J1 = 3.0 Hz, J2 = 1.8 Hz, 1H), 5.92
(dd, J1 = 3.0 Hz, J2 = 0.9 Hz, 1H), 4.01 (t, J = 6.3 Hz, 2H), 2.56 (t, J = 6.6 Hz, 2H), 2.43
(s, 3H), 1.63-1.64 (4H); 13C NMR (CDCl3, 75 MHz) δ 155.1, 144.6, 140.8, 132.9, 129.7,
127.8, 110.0, 105.0, 70.1, 28.1, 27.0, 23.8, 21.5; HRMS (EI) m/z calcd for
C15H18O4S+(M+) 294.0926 found 294.0924.
42

4-(2-Hydroxy-5-oxo-2,5-dihydro-furan-2-yl)-butyl-4-toluenesulphate (2.36).
O
OTs
OHOO OTs NaClO2, NaH2PO4
tBuOH:H2O(5:1), rt
90%2.14 2.36
To a magnetically stirred solution of NaH2PO4 (12.4 mg, 0.09 mmol) and NaClO2
(16 mg, 0.18 mmol) in 1mL t-BuOH-H2O (5:1, v/v) was added alkylfuran 2.14 (20 mg,
0.06 mmol). The resulting mixture was stirred at room temperature until the starting
material was consumed as determined by TLC. Then the solvent was removed on a rotary
evaporator. The residue was extracted with ethyl acetate, washed with brine, dried over
MgSO4 and passed through a short silica-gel column using ethyl acetate to afford 2.36
(17mg, 90%). 1H NMR (CDCl3, 300 MHz) δ 7.75 (d, J = 8.2 Hz, 2H), 7.33 (d, J = 7.9 Hz,
2H), 7.19 (d, J = 5.5 Hz, 1H), 6.09 (d, J = 5.5 Hz, 1H), 4.03 (m, 2H), 2.43 (bs, 3H), 1.4-
1.8 (6H). This was converted to the trans isomer that was further characterized by 1H and
13C NMR as well as HRMS.
4-Oxo-8-(toluene-4-sulphonyloxy)-oct-2-enoic acid (2e).
O
OTs
OHO
HO
O
O
OTsPyridine
THF:Acetone:H2O(5:4:2) 2h, rt2.36 2.17
To a solution of 2.36 (17 mg, 0.05 mmol) in 1 mL THF/acetone/water (5:4:1)
was added a catalytic amount (10 µL) of freshly distilled pyridine. The mixture was
stirred for 2 h at room temperature. Solvents were removed on a rotary evaporator and a
vacuum pump to yield the product 2.17 quantitatively. (TLC: 30% ethyl acetate in
43

hexanes, Rf = 0.32). 1H NMR (CDCl3, 200 MHz) δ 7.77 (d, J = 8.2 Hz, 2H), 7.33 (d, J =
7.9 Hz, 2H), 7.12 (d, J = 15.9 Hz, 1H), 6.63 (d, J = 16.1 Hz, 1H), 4.03 (m, 2H), 2.62 (m,
2H), 2.43 (bs, 3H), 1.6-1.8 (4H); 13C NMR (CDCl3, 50 MHz) δ 198.1, 168.1, 144.9,
144.8, 140.6, 132.9, 129.8, 127.8, 69.9, 40.5, 28.0, 21.6, 19.4; HRMS (FAB) m/z calcd
for C15H19O6S+(M+H+) 327.0902 found 327.0903.
5-Hydroxy-5-pentyl-5H-furan-2-one (2.37).
OC5H11
OHOO C5H11 NaClO2, NaH2PO4
tBuOH:H2O(5:1), rt95%2.5 2.37
Oxidation of 2-pentylfuran (50 mg, 0.36 mmol) was carried out as mentioned in
the general procedure for oxidation using NaH2PO4 (74 mg, 0.54 mmol) and NaClO2 (98
mg, 1.08 mmol) in 3 mL t-BuOH-H2O (5:1, v/v). 1H NMR (300 MHz, CDCl3) δ 7.17 (d,
J = 5.7 Hz, 1H), 6.11 (d, J = 5.7 Hz, 1H), 1.98 (m, 2H), 1.28-1.41 (7H), 0.87 (t, J = 6.3
Hz, 3H). This compound was further characterized by conversion to the trans isomer.
OC5H11
OHO
Pyridine
THF:Acetone:H2O(5:4:2) 2h, rt
HOC5H11
O
O2.37 2.6
The butenolide 2.37 was converted to 4-oxo-2-nonenoic acid (2.6) by the general
isomerization procedure mentioned above in quantitative yields. The spectral data
matched the earlier report.32
44

8-(tert-Butyldimethylsilanyloxy)-4-oxo-oct-2(Z)-enoic acid (2.38).8
O
OTBDMS
OHOO OTBDMS NaClO2, NaH2PO4
tBuOH:H2O(5:1), rt77%2.16 2.38
The oxidation of 2.16 by the general procedure yielded the corresponding
butenolide 2.38. (TLC: 20% ethyl acetate in hexanes, Rf = 0.37) Yield: 77%. 1H NMR
(200 MHz, CDCl3) δ 7.21 (d, J = 5.5 Hz, 1H), 6.10 (d, J = 5.5 Hz, 1H), 3.64 (t, J = 5.9
Hz, 2H), 2.1-1.9 (2H), 1.7-1.4 (4H), 0.88 (s, 9H), 0.05 (s, 6H); 13C NMR (75 MHz,
CDCl3) δ 170.3, 154.2, 123.1, 108.0, 62.9, 36.9, 31.8, 25.9, 20.2, 18.3, -5.3. HRMS
(FAB) m/z calcd for C14H27O4Si+ (M+H+) 287.1673, found 287.1679.
5-[3-(tert-Butyldimethylsilanyloxy)butyl]-5-hydroxy-5H-furan-2-one (2.21).
HO
OH
I
OTBDMS
1.TsCl, Et3N, DMAP2.TBDMSCl, 3.NaI, K2CO3
OTBDMSO
O1.tBuLi2.2.43
2.42 2.43
2.20
O OHOO NaClO2, NaH2PO4
tBuOH:H2O(5:1), rt
TBDMSOOTBDMS
2.20 2.21
The furyl precursor 2.20 was prepared in 4 steps. The diol 2.42 was selectively
tosylated using p-toluenesulfonyl chloride as reported earlier,61 followed by TBDMS
protection of the secondary alcohol using TBDMSCl 62 and iodination63 to yield the iodo
compound 2.43. Alkylation of furan to generate the 2-alkyl derivative 2.20 using tBuLi
45

and 2.43 was performed as reported earlier.64 To a magnetically stirred solution of furan
2.20 (35 mg, 0.05 mmol) in 2 mL of tBuOH:H2O (5:1) was added NaClO2 (0.15 mmol)
and NaH2PO4 (0.75 mmol). The reaction was continued until the reactant was completely
consumed. The solvent was removed by rotary evaporation. The residue, resuspended in
ethyl acetate, was passed through a short plug of Celite to give the butenolide 2.21 in
70% yield. (TLC: 20% ethyl acetate in hexanes, Rf = 0.3) Yield: 75%. 1H NMR (CDCl3,
300 MHz) δ 7.19 (dd, J1 = 12.9 Hz, J2 = 6 Hz, 1H), 6.05 (m, 1H), 4.01 (m, 1/2H), 3.95
(m, 1/2H), 1.6-2.2 (6H), 1.1-1.2 (6H), 0.8-0.9 (9H), (-0.01)-0.16 (6H). This compound
was further characterized by conversion to the trans isomer.
O OHO
HO
O
O
Pyridine
THF:Acetone:H2O(5:4:2) 2h, rt
OTBDMS OTBDMS
2.222.21
The butenolide 2.21 was isomerized to the trans form using the general procedure
described above to yield 2.22 in 95% yield. The proton and carbon spectra were
comparable to the earlier reports.36
5-Hydroxy-5-[4-(tetrahydropyran-2-yloxy)-butyl]-5H-furan-2-one (2.39).
O
OTHP
OHOO OTHP NaClO2, NaH2PO4
tBuOH:H2O(5:1), rt
2.15 2.39
A magnetically stirred suspension of sodium chlorite (24 mg, 0.27 mmol) and
NaH2PO4 (18 mg, 0.13 mmol) in 1mL of tBuOH/H2O (5:1) was added to 2-(4-furan-2-yl-
butoxy)tetrahydropyran (2.39) (20 mg, 0.09 mmol). The reaction mixture was stirred at
46

room temperature and monitored by TLC. After 2 h, solvent was removed by rotary
evaporation, the NMR of the crude residue showed characteristic peaks of the butenolide
2.39. The residue was used for next step without purification.
O
OTHP
OHO
HO
O
O
OTHPPyridine
THF:Acetone:H2O(5:4:2) 2h, rt2.39 2.18
The crude butenolide 2.39, was converted to the trans derivative 2.18 using the
general procedure. The compound 2.18 is not stable on a silica gel column. The sec-butyl
ester derivative of the acid was characterized as described below.
4-Oxo-8-(tetrahydro-pyran-2-yloxy)-oct-2-enoic acid, sec-butyl ester (2.41).
HO
O
O
OTHPO
O
O
OTHPsec-BuOH, DCC,DMAP
CH2Cl2, rt70%, 3 steps2.18 2.40
To a solution of 2.18 (27 mg, 0.1 mmol) in CH2Cl2 (5 mL) was added DCC (31
mg, 0.15 mmol), DMAP (2.4 mg, 0.02 mmol) and sec-butanol (10 µL, 0.11 mmol). The
solution was stirred over night. The reaction mixture was diluted with CH2Cl2, washed
with water, then brine and dried on MgSO4. The solvent was then removed and the
residue was purified using column chromatography (40% ethyl acetate in hexanes Rf =
0.4) to obtain an oil 2.40 (27 mg, 70%). 1H NMR (300 MHz, CDCl3) δ 7.03 (d, J = 16
Hz, 1H), 6.66 (d, J = 16 Hz, 1H), 4.93 (m, 1H), 4.55 (m, 1H), 3.6-3.8 (m, 2H), 3.3-3.9
(2H), 2.68 (t, J = 6.9 Hz, 2H), 1.4-1.8 (14H), 1.25 (d, J = 6.3 Hz, 2H), 1.18 (d, J = 6.3 Hz,
1H), 13C NMR (APT, 50 MHz) (+)199.7, (+)165.2, (-) 139.1, (-) 131.3, (-) 98.9, (-) 73.5,
47

(+) 67.0, (+) 62.4, (+) 41.0, (+) 30.7, (+) 29.0, (+) 28.7, (+) 25.4, (+) 20.6, (+) 19.6, (-)
19.3, (-) 9.6. HRMS (FAB) m/z calcd for C17H29O5 (M+H+) 313.2015 found 313.2017.
1-Palmityl-2-(1-carboxy-4-(2-hydroxy-5-oxo-2,5-dihydro-furan-2-yl)-butanoyl)-sn-
glycero-3-phosphatidylcholine (2.26).
O O
OPC
2
HO
O
OO
OPC
2
NaClO2 (3 eq.) PBS (pH 3.5):CHCl3:H2O(2:1:1), rt
>75%
2.20
2.24 2.26
To a magnetically stirred solution of 8-furyl butanoyl ester derivative of the
phosphatidylcholine 2.2438 (5 mg, 8 µmol) in a solution of phosphate buffer pH 3.5,
chloroform and water (2:1:1) (1 mL) was added sodium chlorite (3.5 mg, 39 µmol). After
20 min, the solution was concentrated and purified using a 1 mL solid phase extraction
(SPE) column, eluting with a methanol:water gradient (50 –100% methanol) to yield 2.26
(3.5 mg, 66%). 1H NMR (CD3OD, 300 MHz) δ 7.41 (bs, 1H), 6.15 (d, J = 6 Hz, 1H),
5.25 (m, 2H), 4.39 (m, 1H), 4.28 (m, 2H), 4.21 (m, 1H), 4.01 (m, 2H), 3.66 (m, 2H), 3.23
(s, 9H), 2.3-2.4 (4H), 1.5-2.1 (6H), 1.2-1.3 (24H), 0.88 ( t, J = 6 Hz, 3H); HRMS
(MALDI-TOF): m/z calcd for C32H59NO11P+ (MH+) 664.3826 found 664.3820
1-Palmityl-2-(7-carboxyl-5-oxohept-6-enoate)-sn-glycero-3-phosphatidylcholine
(KOdiA-PC, 2.3).
OOO
OOHPC
2
O O
OPC
2
HO
O
Pyridine
THF:Acetone:H2O(5:4:2) 2h, rt 2.32.26
48

The butenolide 2.26 was isomerized by the general method mentioned above to
the trans form 2.3. The 1H NMR spectral data of the product 2.3 is in agreement with that
previously reported.38
1-Palmityl-2-(1-carboxy-8-(2-hydroxy-5-oxo-2,5-dihydro-furan-2-yl)-octanoyl)-sn-
glycero-3-phosphatidylcholine (2.27).
O O
OPC
6
HO
O
OO
OPC
6
NaClO2 (3 eq.) PBS (pH 3.5):CHCl3:H2O(2:1:1), rt
>75%2.25 2.27
To a magnetically stirred solution of the 8-furyl octanoyl ester derivative of
phosphatidylcholine 2.2538 (9 mg, 13 µmol) in a solution of phosphate buffer pH 3.5,
chloroform and water (2:1:1) (1 mL) was added sodium chlorite (3.5 mg, 39 µmol). After
1 h the solution was concentrated, and purified using an SPE column, eluting with a
methanol:water gradient (50 –100% methanol) to yield 2.27 (6 mg, 64%). 1H NMR
(CD3OD, 300 MHz) δ 7.41 (bs, 1H), 6.12 (d, J = 6 Hz, 1H), 5.25 (m, 2H), 4.4 (m, 1H),
4.27 (m, 2H), 4.16 (dd, J1 = 12.6 Hz, J2 = 6.3 Hz, 1H), 4.01 (t, J = 5.1 Hz, 2H), 3.66 (m,
2H), 3.23 (s, 9H), 2.3- 2.4 (4H), 1.5-2.1 (5H), 1.2-1.3 (30H), 0.88 ( t, J = 6 Hz, 3H);
HRMS (MALDI-ToF): m/z calcd for C36H67NO11P+ (MH+) 720.4457 found 720.4424.
1-Palmityl-2-(11-carboxyl-9-oxoundec-6-enoate)-sn-glycero-3-phosphatidylcholine
(KDdiA-PC, 2.4).
OOO
OOHPC
6
O O
OPC
6
HO
O
Pyridine
THF:Acetone:H2O(5:4:2) 2h, rt 2.42.27
49

The butenolide 2.27 was isomerized by the general method described above. The
spectral data of the product 2.4 is in agreement with the previously reported data.38
Ring-Chain Tautomerism:
The product of 2-alkylfuran oxidation by sodium chlorite was confirmed to be a
butenolide under acidic conditions and the corresponding chain isomer at higher pH
through monitoring the λmax. The UV data was collected by dissolving compound 2.37 in
water of varying pH. The graph shows the increase of the peak intensity at 240-250 nm at
higher pH.
50

2.5. References: (1) Witztum, J. L.; Berliner, J. A. Curr Opin Lipidol 1998, 9, 441-8. (2) Girotti, A. W. J. Lipid Res. 1998, 39, 1529-1542. (3) Farooqui, A. A.; Horrocks, L. A. Cellular Mol Neurobiol 1998, 18, 599-608. (4) Balamraju, Y. N.; Sun, M.; Salomon, R. G. J Am Chem Soc 2004, 126, 11522-8. (5) Bergamini, C.; Gambetti, S.; Dondi, A.; Cervellati, C. Curr Pharm Design 2004,
10, 1611-26. (6) McCall, M. R.; Frei, B. Free Radic Biol Med 1999, 26, 1034-53. (7) Finkel, T.; Holbrook, N. J. Nature 2000, 408, 239-47. (8) Sun, M. Ph. D, Case Western Reserve University, 2004. (9) Parthasarathy, S.; Quinn, M. T.; Schwenke, D. C.; Carew, T. E.; Steinberg, D.
Arteriosclerosis 1989, 9, 398-404. (10) Podrez, E. A.; Poliakov, E.; Shen, Z.; Zhang, R.; Deng, Y.; Sun, M.; Finton, P. J.;
Shan, L.; Febbraio, M.; Hajjar, D. P.; Silverstein, R. L.; Hoff, H. F.; Salomon, R. G.; Hazen, S. L. J Biol Chem 2002, 277, 38517-23.
(11) Podrez, E. A.; Poliakov, E.; Shen, Z.; Zhang, R.; Deng, Y.; Sun, M.; Finton, P. J.; Shan, L.; Gugiu, B.; Fox, P. L.; Hoff, H. F.; Salomon, R. G.; Hazen, S. L. J Biol Chem 2002, 277, 38503-16.
(12) Walton, H. M. J Org Chem 1957, 22, 308-12. (13) Sekiguchi, J.; Kuroda, H.; Yamada, Y.; Okada, H. Tetrahedron Lett 1985, 26,
2341-2. (14) Pfefferle, C.; Kempter, C.; Metzger, J. W.; Fiedler, H. P. J Antibiot (Tokyo) 1996,
49, 826-8. (15) Nozoe, S.; Hirai, K.; Tsuda, K.; Ishibashi, K.; Shirasaka, M.; Grove, J. F.
Tetrahedron Lett 1965, 4675-7. (16) Kalita, D.; Khan, A. T.; Barua, N. C.; Bez, G. Tetrahedron 1999, 55, 5177-5184. (17) Kobayashi, Y.; Kumar, G. B.; Kurachi, T.; Acharya, H. P.; Yamazaki, T.;
Kitazume, T. J Org Chem 2001, 66, 2011-8. (18) Ronsheim, M. D.; Zercher, C. K. J Org Chem 2003, 68, 1878-1885. (19) Appleton, D.; Duguid, A. B.; Lee, S.-K.; Ha, Y.-J.; Ha, H.-J.; Leeper, F. J. J
Chem Soc, Perkin Trans 1: Org Bio-Org Chem 1998, 89-101. (20) Obrecht, D.; Weiss, B. Helvetica Chim Acta 1989, 72, 117-22. (21) Kawashima, M.; Sato, T.; Fujisawa, T. Bull Chem Soc Jpn 1988, 61, 3255-64. (22) Pappalardo, P.; Ehlinger, E.; Magnus, P. Tetrahedron Lett 1982, 23, 309-12. (23) D'Auria, M.; Piancatelli, G.; Scettri, A. Tetrahedron 1980, 36, 3071-4. (24) Gunn, B. P.; Brooks, D. W. J Org Chem 1985, 50, 4417-18. (25) Finlay, J. M., M. Anthony; Gunaratne, H. Q. Nimal Tetrahedron Lett 1998, 39,
5651. (26) Cottier, L.; Descotes, G.; Eymard, L.; Rapp, K. Synthesis 1995, 303-6. (27) Franzen, R.; Tanabe, K.; Morita, M. Chemosphere 1998, 38, 973-980. (28) Asaoka, M. Y., N.; Sugimura, N.; Takei, H. Bull Chem Soc Jpn 1980, 53, 1061-4. (29) Oishi, S.; Nelson, S. D. J Org Chem 1992, 57, 2744-2747. (30) Boukouvalas, J.; Lachance, N. Synlett 1998, 31-32. (31) Adger, B. M. B., C.; Brennan, J.; McKervey, M. A.; Murray, R. W. J Chem Soc,
Chem Comm 1991, 21, 1553-4.
51

(32) Ballini, R.; Bosica, G. J Nat Prod 1998, 61, 673-4. (33) Hirsch, J. A.; Szur, A. J. Tetrahedron 1972, 28, 2961-7. (34) Gunn, B. P. Tetrahedron Lett 1985, 26, 2869-72. (35) Hase, T. A.; Nylund, E. L. Tetrahedron Lett 1979, 2633-6. (36) Kobayashi, Y.; Kishihara, K.; Watatani, K. Tetrahedron Lett 1996, 37, 4385-
4388. (37) Kobayashi, Y.; Nakano, M.; Kumar, G. B.; Kishihara, K. J Org Chem 1998, 63,
7505-7515. (38) Sun, M.; Deng, Y.; Batyreva, E.; Sha, W.; Salomon, R. G. J Org Chem 2002, 67,
3575-84. (39) Barco, A.; Benetti, S.; De Risi, C.; Pollini, G.; Zanirato, V. Tetrahedron 1995, 51,
7721. (40) Ngooi, T. J Org Chem 1989, 54, 911. (41) Rao, A. V. R.; Reddy, K. B.; Dhar, T. G. M. Indian J Chem, Section B: Org Chem
Incl Med Chem 1986, 25B, 1014-16. (42) Barco, A.; Benetti, S.; De Risi, C.; Pollini, G. P.; Zanirato, V. Tetrahedron 1995,
51, 7721-7726. (43) Takeya, H. In Eur. Pat. Appl.; (Cosmo Research Institute, Japan; Cosmo Oil Co.,
Ltd.). Ep, 1995. (44) Lukevics, E. Y.; Ignatovich, L. M.; Gol'dberg, Y. S.; Shymanskaya, M. V.
Khimiya Geterotsiklicheskikh Soedinenii 1986, 853-4. (45) Nakamura, I.; Saito, S.; Yamamoto, Y. J Am Chem Soc 2000, 122, 2661-2662. (46) Ochiai, M.; Arimoto, M.; Fujita, E. Tetrahedron Lett 1981, 22, 4491-4. (47) Ballini, R.; Bosica, G.; Fiorini, D.; Gil, M. V.; Petrini, M. Org Lett 2001, 3, 1265-
1267. (48) Yu, J.-Q.; Corey, E. J. J Am Chem Soc 2003, 125, 3232-3233. (49) Carles, L.; Narkunan, K.; Penlou, S.; Rousset, L.; Bouchu, D.; Ciufolini, M. A. J
Org Chem 2002, 67, 4304-4308. (50) Dittami, J. P.; Xu, F.; Qi, H.; Martin, M. W.; Bordner, J.; Decosta, D. L.;
Kiplinger, J.; Reiche, P.; Ware, R. Tetrahedron Lett 1995, 36, 4197-200. (51) Halland, N.; Hazell, R. G.; Jorgensen, K. A. J Org Chem 2002, 67, 8331-8338. (52) Halland, N.; Aburel, P. S.; Jorgensen, K. A. Angwante Chem, Int Edn 2003, 42,
661-665. (53) Gollnick, K.; Griesbeck, A. Tetrahedron 1985, 41, 2057-68. (54) Saldabol, N. O.; Popelis, J.; Slavinska, V. Chem Heterocyclic Compds (New York,
NY, United States)(Translation of Khimiya Geterotsiklicheskikh Soedinenii) 2002, 38, 873-881.
(55) Noyori, R.; Sato, T.; Kobayashi, H. Bull Chem Soc Jpn 1983, 56, 2661-79. (56) Williams, P.; Eugene, L. J Org Chem. 1981, 46(21), 4143-4147. (57) Shet, J.; Desai, V.; Tilve, S. Synthesis 2004, 1859-1863. (58) Bouchain, G.; Leit, S.; Frechette, S.; Khalil, E.; Lavoie, R.; Moradei, O.; Woo, S.;
Fournel, M.; Yan, P.; Kalita, A.; Trachy-Bourget, M.; Beaulieu, C.; Li, Z.; Robert, M.; MacLeod, A.; Besterman, J.; Delorme, D. J Med Chem 2003, 46, 820-30.
(59) Annangudi, S. P.; Sun, M.; Salomon, R. G. Synlett 2005, 2005, 1468-1470.
52

(60) O'Neill, J. A.; Lindell, S. D.; Simpson, T. J.; Willis, Christine L. Tetrahedron: Asymm 1994, 5, 117-118.
(61) O'Neil, J. A.; Lindell, S. D.; Simpson, T. J.; Willis, C. L. Tetrahedron: Asymm 1994, 5, 117-18.
(62) Bonini, C.; Chiummiento, L.; Evidente, A.; Funicello, M. Tetrahedron Lett 1995, 36, 7285-6.
(63) Allingham, M. T.; Howard-Jones, A.; Murphy, P. J.; Thomas, D. A.; Caulkett, P. W. R. Tetrahedron Lett 2003, 44, 8677-8680.
(64) Bracher, F.; Schulte, B. Nat Prod Res 2003, 17, 293-299.
53

Chapter 3
Detection and Characterization of Multiple 4-Hydroxynonenal Adducted Amino
acids Using Deuterium Labeled HNE and Mass Spectrometry
54

3.1. Background
Aerobic organisms use oxygen extensively for anabolic as well as catabolic
processes, which makes them prone to radical induced oxidative damage in vivo.1 These
organisms have developed mechanisms to circumvent this problem primarily by using
antioxidants that can quench the toxic radicals2-8 as well as by evolving efficient
recycling pathways to of dispose of the damaged biomolecules.1 Polyunsaturated fatty
acids 9 (PUFAs), proteins10 and DNA11 are some of the targets of radical induced
damage. Enzyme mediated lipid peroxidation products are known to be involved in
normal physiological functions, e.g., generating prostaglandins and thromboxanes.12,13
Analogous radical induced peroxidation of PUFAs generates prostaglandin isomers
(isoprostanes) and in addition it can form highly reactive levulinaldehyde derivatives
with prostaglandin side chains (isolevuglandins).14 Isolevuglandins form adducts with
side chains of proteins and DNA bases. The adducts so formed are implicated in various
pathological processes such as atherosclerosis,15 age related macular degeneration 16-18
and neurodegeneration.19,20
Arachadonic and linoleic acids phospholipids have four and two conjugated
double bonds respectively, making them highly susceptible to free radical attack followed
by rearrangements and/or peroxidation leading to an array of reactive oxidation
products.21 The staggering number of molecular targets in the intracellular as well as
intercellular environment for these highly reactive lipobolites (products of lipid
degradation) form a complicated mixture of products, making it very difficult to identify
them and assign molecular structures.22 Our group is involved in a systematic and
55

mechanistic approach to predict structures for lipobolites and the stable adducts that they
form with macromolecules.
Hydroxynonenal (HNE), a lipid peroxidation product of arachidonyl and linoleyl
lipids, has been very well studied mainly due to its proclivity to form pathological
adducts23-26 with nucleophilic amino acid residues of proteins. Modification by HNE has
been shown to alter the structure27 and function28,29 of proteins. The adducts are
immunogenic and can elicit an immune response, thus demonstrating that these are not
native to the human body.30,31 The HNE- Michael adduct has been studied extensively.
Lysine, cysteine and histidine residues form Michael adducts readily.24 Besides the
reversible formation of Michael adduct, the ε-amino group of lysine residues can
reversibly form Schiff bases, that may rearrange followed by dehydration to form more
stable pyrroles.32 In contrast, histidine and cysteine amino acid residues only form stable
Michael adducts.33 In vitro, a histidine-lysine crosslink was observed for model
compounds, but there are no reports of their detection in vivo.34
NMR studies in collaboration with Dr. Sayre’s group detected the formation of
adducts that have 2 or more aldehydes per amine group, from the reaction of equimolar
amounts of HNE and primary amines.32 The detection and characterization of these
adducts involved derivatization using strong reducing and/or acidic conditions.
Therefore, only adducts that are stable under these conditions were detected. There is a
need for mild techniques that are compatible with the products, which are stable under
physiological conditions, but readily decompose under harsher conditions. To address
this problem, we are applying isotope labeling and mild conditions to probe adduction
chemistry that occurs under physiological conditions.
56

Studies on HNE modification of LDL by Esterbauer’s group,35 have shown that
there is a steady increase in the amount of HNE bound per mol of LDL with increasing
amount of HNE. The number of nucleophilic amino acid residues in LDL proteins
modified by 13C labeled HNE was estimated to be 0.71 - 1.3 mol/mol of HNE. This
suggests that a single amino acid residue might be modified by more than one HNE.
Furthermore, a high concentration of the HNE (> 10 mM) caused the LDL to precipitate.
This may be because HNE neutralizes the positively charged residues (lysine, histidine,
arginine) on the LDL and/or cause protein aggregation.
Deng et al. performed similar studies with tritium labeled [9-3H]-HNE.36
Incubation of [9-3H]-HNE with LDL, revealed that the number of HNE’s that can
become covalently linked to an LDL particle i.e., 486 molecules per particle, was far
greater than the number of “reactive” lysine, ethanolamine phospholipids, histidine and
cysteine residues i.e., 365 present in each particle (Table 2.1). This indicated that 1:1
Table 3.1. Nucleophilic functional groups in LDLa
Residue Total Reactive
lysine ε-NH2 350 225*
PEs 80 80
histidine 115 115 (?)
cysteine 25 25 (?)
Total 570 365
*TNBS reactive amino groups are 317±29 that would include all reactive lysines and all EPs: which is 225 + 80 = 305. "?" means the amount of actually reactive histidine and cysteine remains unclear. (Adapted from Dr. Deng’s Thesis)
57

adducts e.g. Michael adducts, Schiff bases and pyrroles alone cannot account for the
level of adduction observed. Rather adducts incorporating two or more HNE per
nucleophilic residue must be generated.
To characterize the type of adducts that might be generated in an LDL particle, the
present study focused on qualitative analysis of these adducts formed by HNE with
amino acids (N-acetylated lysine, glycine-lysine methyl histidine and cysteine) using
deuterium labeled HNE and mass spectrometric analyses.
3.1.1. Previous syntheses of deuterated HNE. HNE being one of the most investigated
molecules of the lipid oxidation products, a wide range of synthetic methods have been
reported.36 Synthesis incorporating carbon and hydrogen isotopes have been developed
for use as a probe for investigating metabolic pathways,4 as mass spectrometric
standards37 or to study adducts formed by HNE.38 Among the many procedures
described previously to synthesize deuterium labeled HNE, only two were used
extensively. One involved cobalt (II) porphyrin-catalyzed reductive oxygenation of
commercially available α,β,γ,δ-unsaturated aldehydes. Sugamoto’s group used this
method to synthesize a deuterium labeled HNE from 2(E),4(E)-nonadienal in two steps
(Scheme 3.1). 39,40
58

O
Co II (tdcpp)(0.001 eq.)O2 (1 atm)Et3SiD (1.2 equiv.)
PriOD-DCM (1:1)
OOOH
D
PPh3
OOH
D
54%
Scheme 3.1: Synthesis of [2H1]-HNE reported by Sugamoto’s group.40
Another method involved the use of fumaraldehyde dimethylacetal that is
commercially available, as a precursor for synthesis of HNE. Selective deprotection and
alkylation, of the formaldehyde dimethylacetal using readily available deuterated
alkylating reagents for the deuterium incorporation, followed by deprotection of the
dimethylacetal, generated the required HNE (Scheme 3.2).41,42
59

H3CO
OCH3
OCH3
OCH3H3CO
OCH3
O
Amberlyst 15catalyst
RMgBr
H3COR
OCH3
OH
OR
OH
Amberlyst 15catalyst
R = -(CH2)4-CD3, -(CD2)4-CD3
Scheme 3.2: Synthesis of [2H3]-HNE and [2H11]-HNE from fumaraldehyde
dimethylacetal.
[8,8,9,9-2H4]-HNE is optimal for detecting multiple adducts. Choosing the right kind
of deuterated HNE is important for many practical reasons. Firstly, the deuterium atoms
should be in a position that precludes it from loss or any chemical reactivity. The carbons
1-4 are involved in the adduction chemistry and the protons at carbon 5-7 may be
involved in the McLafferty type rearrangements43 that could occur in the mass
spectrometric ionization process as shown below.
BA
D
HBA D
H
αβ
γ
δ
Secondly, the number of deuteria present should be sufficient to readily
differentiate adducts of labeled and unlabeled HNE from the isotopic peaks that arise
from the various isotopic abundances of some of the elements. Since our study was to be
60

focused on the elements C, H, O, N and S, the highest isotopic peak (> 1%) is at M+1 for
carbon and M+2 for sulphur Table 2.2. Thirdly, the number of deuteria incorporated
should be easily achieved synthetically. The use of multiples of two is readily achieved
by hydrogenation of alkenes. All these considerations indicated that the deuterium atoms
should be located at C8 or C9 of the HNE molecule and that there should be > 2
deuterium atoms. We, therefore, chose to incorporate four deuterium atoms at the C8 and
C9 carbons by reduction of an alkyne bond. Consequently, we prepared a deuterated
alkylating reagent by deuteration of an alkyne triple bond.
Table 3.2. Isotopic mass abundances of some common elements 44
Element Mass No. Isotopic Composition (%) Isotopic mass
1H 2H
1
2
100
0.0115
1.007
2.014 12C 13C
12
13
100
1.08
12.000
13.003 14N 15N
14
15
100
0.369
14.003
15.000 16O 17O 18O
16
17
18
100
0.038
0.205
15.995
16.999
17.999 32S 33S 34S 36S
32
33
34
36
100
0.08
4.52
0.02
31.972
32.971
33.967
35.967
61

3.2. Results
3.2.1. Synthesis of [8,8,9,9-2H4]-HNE. 4-Pentynol (3.1) was used as a precursor for
carbons 5-9 and the protected aldehyde 3.5 as the precursor for carbons 1-4 of the d4-
HNE. 4-Pentynol was converted to its tosyl-derivative 3.2 for two reasons; (i) to decrease
the loss of the alcohol due to its low boiling point and (ii) to be easily converted to the
iodopentane 3.4 for the subsequent alkylation step. The d4-pentanol tosyl ester 3.3 was
obtained in 74% yield by deuteration of pent-4-ynyl tosyl ester (3.2)45 using Wilkinson’s
catalyst, RhCl(PPh3)4, and deuterium.46 The d4-pentyl iodide (3.4) was generated by
refluxing the tosylate 3.3 with NaI and K2CO3 in acetone (Scheme 3.3).36
d4-Pentyllithium was generated by metal halogen exchange of 3.4 using t-BuLi.
Alkylation of aldehyde 3.5 by pentyllithium yielded the stable precursor 3.6 of d4-HNE in
84 % overall yield. Preparation of HNE from this precursor was performed by a one-step
procedure, using NaIO4 and aqueous acetic acid, to give d4-HNE (3.7) in 89 % yield
(Scheme 3.3). An alternative two-step procedure using acetic acid and lead acetate gave
HNE in 78 % yield (Scheme 3.3).47 Commercially available pentylmagnesium bromide
(3.8) was used as alkylating agent to synthesize unlabeled HNE (Scheme 3.4).
62

OR
TsCl
RhCl(PPh3)3, D2HD2C
D2C O
SO
O
HD2C
D2C R
3.3, R=OTs3.4, R=I
NaI,Acetone
+ OOO
OOHD2C
CD2 OH
BuLi
Ethyl ether
NaIO4
Acetic acid:H2O (2:1)
OOHD2C
CD2 OH
OHD2C
CD2 OH
3.3
3.6
3.7
3.1, R= H3.2, R=Ts, 86%
C6H6
3.5
89%
84%
74%
Scheme 3.3: Synthesis of [8,8,9,9-2H4]-HNE.
MgBr + OOO
OO
OH
BuLi
Ethyl ether
NaIO4
Acetic acid:H2O (2:1)
OO
OHO
OH
3.9
3.10
3.5
90%
87%3.8
Scheme 3.4: Synthesis of HNE.
63

3.2.2. Reactions of N-acetyl amino acids with HNE. A solution of HNE and d4-HNE
(1:1) (10 µM) was incubated with various amino acids (N-acetyl glycine-lysine-methyl
ester (N-acetyl-Gly-Lys-OMe), N-acetyl lysine, N-acetyl histidine (N-acetyl-His) and N-
acetyl-cysteine (N-Acetyl-Cys) individually in PBS at 37 ºC for 24 h. The resulting
mixtures were dried on a speed vacuum, extracted with methanol and dried using a
stream of nitrogen. The resulting residue was dissolved in 50% methanol/water with a
trace of formic acid (0.01%) and analyzed by mass spectrometry.
3.2.3. Mass spectrometric analysis of the HNE-amino acid adducts. A preliminary
analysis of the reaction mixtures using MALDI-TOF mass spectrometry revealed the
presence of numerous peaks that appear as unique sets of multiplets with a mass
difference of 4 Da as expected. Further analysis was performed using a Q-TOF mass
spectrometer by ESI-MS (Figure 3.1) and ESI-MS/MS to identify the products formed.
Also, semi-quantitative analyses of the molecular ions identified were done by liquid
chromatography and selected ion recording (LC-SIR) using a Quattro Ultima ™
(Micromass) mass spectrometer.
64

Figure 3.1: ESI-TOF-MS of reaction mixture containing HNE (d4:d0, 1:1) with amino
acids A) N-acetyl-gly-lys-OMe, B) N-acetyl-cysteine and C) N-acetyl-histidine.
(i) N-Acetyl-histidine-HNE adducts. The parent ion for the Michael adduct of N-acetyl-
His with HNE (d4:d0, 1:1) appears as a doublet at m/z 354 and 358 (Figure 3.1, panel C).
Tandem MS/MS analysis of these ions resulted in a specific signature of fragments
corresponding to the labeled or unlabeled HNE. Thus, MS/MS of the m/z 354 ion yields
daughter ions at m/z 336, 312, 308, 266, 198, 180, 177, 156, 152, 139, 110 and 95, while
tandem MS/MS of the ion at m/z 358 yields daughter ions at m/z 340, 316, 312, 270, 198,
180, 177, 156, 152, 143, 110 and 95 (Figure 3.2). Based on mass differences between the
ions from both of the spectra, daughter ions containing the C5-C9 alkyl chain of the HNE
were identified (vide infra). The structures of the ions in the mass spectrum can be
rationalized based on the patterns observed in the fragmentation of N-acetyl-His. Similar
to the fragmentation pattern reported for N-acetyl-His,48 fragmentation of the Michael
65
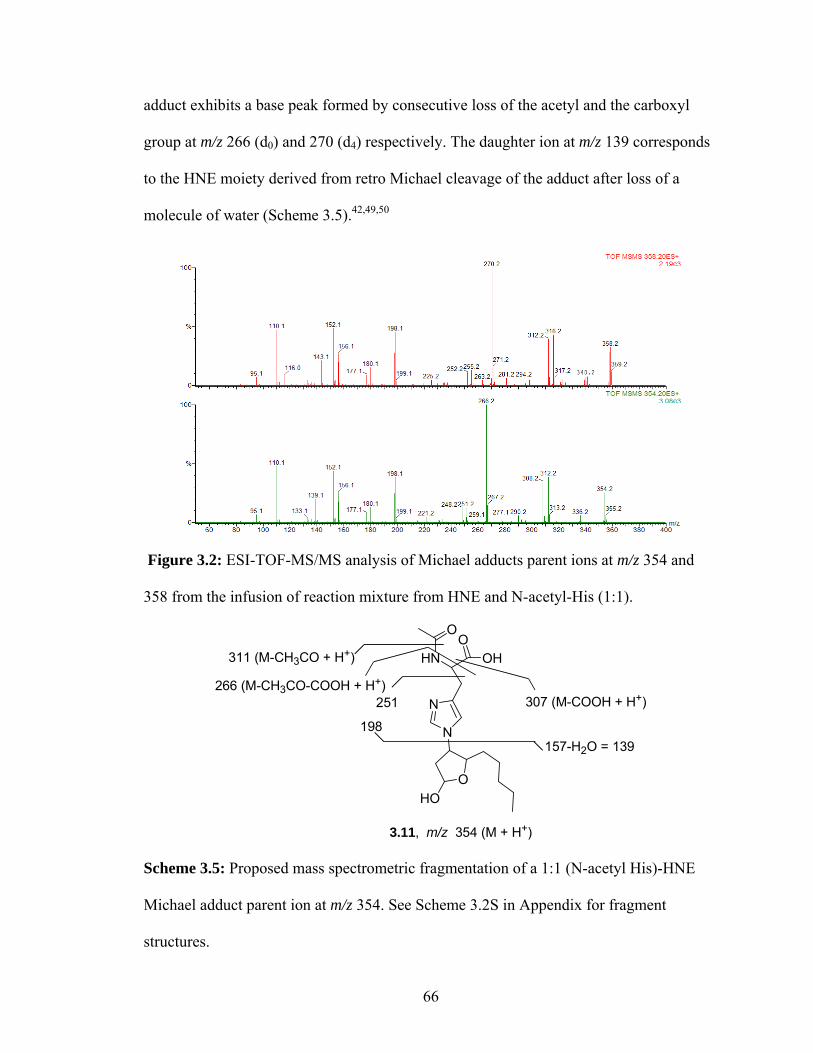
adduct exhibits a base peak formed by consecutive loss of the acetyl and the carboxyl
group at m/z 266 (d0) and 270 (d4) respectively. The daughter ion at m/z 139 corresponds
to the HNE moiety derived from retro Michael cleavage of the adduct after loss of a
molecule of water (Scheme 3.5).42,49,50
Figure 3.2: ESI-TOF-MS/MS analysis of Michael adducts parent ions at m/z 354 and
358 from the infusion of reaction mixture from HNE and N-acetyl-His (1:1).
HN OHO
N
N
O
O
3.11, m/z 354 (M + H+)
HO
311 (M-CH3CO + H+)
307 (M-COOH + H+)266 (M-CH3CO-COOH + H+)
251
198157-H2O = 139
Scheme 3.5: Proposed mass spectrometric fragmentation of a 1:1 (N-acetyl His)-HNE
Michael adduct parent ion at m/z 354. See Scheme 3.2S in Appendix for fragment
structures.
66

The total ion chromatogram of the reaction mixture (Figure 3.1) showed ions with
m/z much greater than 400. These molecular ion peaks can only be formed by reaction of
two or more of the reaction components. A triplet was observed at m/z 510, 514 and 518
(1:2:1) corresponding to an adduct with 2 HNE’s and one N-acetyl-His (Figure 3.1). The
tandem MS/MS of the m/z 510 precursor ion generates some of the prominent fragments
*
*
*
*
*****
*
**
* * * *
***
■■
■
■
■■
■■■
■
■
■
■
■
■
A
B
C
*
*
*
*
*****
*
**
* * * *
***
■■
■
■
■■
■■■
■
■
■
■
■
■
*
*
*
*
*****
*
**
* * * *
***
■■
■
■
■■
■■■
■
■
■
■
■
■
A
B
C
Figure 3.3: ESI-MS/MS of molecular ions at A) m/z 518, B) m/z 514 and C) m/z 510,
from the infusion of reaction mixture containing HNE and N-acetyl-His. Peaks labeled
with an asterix (*) or a solid rectangle (■) denote fragments with or without the C5-C9
alkyl chain of HNE, respectively.
67

corresponding to the loss of carboxyl, acetyl and HNE-d0 at m/z 464, 468 and 354
respectively (Figure 3.3), including ions observed for the fragmentation of the m/z 354
ion (Figure 3.2). The fragmentation of the parent ions at m/z 514 and 518 generated
daughter ions with masses 4 or 8 Da more than those observed for the m/z 510 ion
(Figure 3.3).
A quadruplet of ions (Figure 3.1C) at m/z 666, 670, 674 and 678 is present in the
total ion chromatogram of the reaction mixture representing adduction of three HNE’s
with one N-acetyl-His in an approximate 1:3:3:1 distribution. Tandem MS/MS analysis
of the m/z 666 ion yielded the peaks at m/z 624, 626, 578, 510 and peaks that were
observed in the fragmentation of m/z 510 ions (Figure 3.3). The fragmentation of the m/z
670 and 674 parent ions gave daughter ions that correspond to 4 or 8 Da more compared
to those observed for the m/z 666 ion (Figure 3.4).
Figure 3.4: ESI-MS/MS of molecular ions A) m/z 674, B) m/z 670 and C) m/z 666 from
infusion of reaction mixture HNE and N-acetyl-His (1:1) in Figure 3.1C.
68

HPLC separation of the (N-acetyl-His)-HNE reaction mixture followed by
selected ion recording (LC-ESI-SIR) of the prominent ions mentioned above in 13
channels was monitored on a Quattro ultima® electrospray mass spectrometer
(Micromass). As evident from the chromatograms in Figure 3.5, the peaks derived from
each adduct and its d4 labeled isomer have similar signatures. The 1:1 Michael adduct is
expected to be comprised of four diastereomers, that emerge at 8.6 – 9.4 min. In the case
of 2:1 and 3:1 adducts, much more complex mixtures of diastereomers are anticipated.
The total ion current for 1:1 adducts is greater than for 2:1 adducts, and that for 3:1
adducts is relatively minor. Also, with the increase in the number of HNE molecules
adducted on to N-acetyl-His, the polarity of the adducts change dramatically (1:1 > 2:1 >
3:1), as is evident from the selected ion recording (SIR) chromatograms (Figure 3.5).
69

Figure 3.5: LC-ESI for the reaction mixture from N-acetyl-His and HNE with selected
ion recording for 13 channels of which 7 channels are shown here. A) m/z 670, B) m/z
666, C) m/z 518, D) m/z 514, E) m/z 510, F) m/z 358 G) m/z 354 and H) TIC (for 13
channels).(for chromatograms of channels not shown here, see Appendix Figure 3.1S)
(ii) Lysine HNE adducts. N-acetyl-glycine-lysine methyl ester (N-acetyl-Gly-Lys-OMe)
was used as a lysine residue surrogate for the analysis of the adduction pattern with HNE.
Preliminary analysis using MALDI-TOF indicated that both N-acetyl lysine (NAL) and
N-acetyl-Gly-Lys-OMe showed similar patterns of adduction. Only the (N-acetyl-Gly-
70

Lys-OMe)-HNE reaction mixture was analyzed by electrospray MS to determine the
nature of these adducts. Fragmentation of the parent ion at m/z 260 (Figure 3.1A) from
this dipeptide generates daughter ions at m/z 242, 218, 201, 144 (base peak), 161, 129 and
84 (Figure 3.6).
Figure 3.6: ESI-MS/MS analysis of m/z 260 molecular ion. Tandem MS/MS analysis of
a solution of N-acetyl-Gly-Lys-OMe was injected into a Q-TOF mass spectrometer by
infusion at a flow rate of 0.5 µL/min.
The ESI- MS/MS spectrum of the molecular ion at m/z 416 and the isotope
labeled peak at m/z 420 (Figure 3.1A) corresponds to the Michael (1:1) adduct based on
the molecular weight and pattern of fragmentation. A carbinolamine formed by reaction
of the ε-amino group of N-acetyl-Gly-Lys and the aldehyde group of HNE has the same
molecular weight as the Michael adduct (m/z 354). The possibility of such structural
variations in the molecular ions cannot be ruled out. The major peaks in the spectrum are
formed from the loss of water (-18), an acetyl group (-44) and by retro-Michael
elimination (-156) (Figure 3.7, Scheme 3.6).
71

Figure 3.7: ESI-MS/MS of molecular ions at A) m/z 416 and B) m/z 420) (Figure 3.1A)
corresponding to a 1:1 HNE-(N-acetyl-Gly-Lys-OMe) Michael adduct.
NH O
HN
O
HNO
O
OHO
372 (M - CH3CO)
260 (MH+ - HNE)
139 (M - NGL -H2O)
398(M - H2O + H+)
3.12, m/z 416 (M + H+)
Scheme 3.6: Possible Michael adduct HNE - (N-acetyl-Gly-Lys-OMe) fragmentation.
See Appendix Scheme 3.4S for fragment structures.
The molecular ions corresponding to a Schiff base 3.13, formed from N-acetyl-Gly-Lys-
OMe and HNE, were observed at m/z 398 and 402 (Figure 3.7). Tandem MS analyses of
these ions are shown in Figure 3.8 and possible fragmentations proposed in Scheme 3.7.
72

Figure 3.8: ESI-MS/MS analysis of molecular ions A) m/z 402 and B) m/z 398 (Figure
3.7) corresponding to an HNE- (N-acetyl-Gly-Lys-OMe) Schiff base adduct.
NHO
N
O
NH
O
O
HO
222 (α−cleavage)204 (222 - H2O)
281 (M -H2O - amide cleavage)
354 (M - CH3CHO)
3.13, m/z 398 (M+H+)
Scheme 3.7: Possible Schiff base HNE- (N-acetyl-Gly-Lys-OMe) Schiff base
fragmentation. See Appendix Scheme 3.5S for fragment structures.
73

The tandem MS/MS of molecular ions at m/z 380 and m/z 384, corresponding to
the pyrrole adduct of the HNE with lysine, show a fragmentation consistent with the
structure of these adducts. One prominent feature among these spectra is the absence of
the retro-Michael adduct fragment at m/z 139/143 in the MS/MS of both the Schiff base
adduct 3.13 and the pyrrole adduct 3.14.
Figure 3.9: ESI-MS/MS analysis of molecular ions at A) m/z 384 and B) m/z 380
corresponding to the HNE-(N-acetyl-Gly-Lys-OMe) pyrrole adduct
74

NHO
N
O
NHO
O
338 (M -CH3CO + H+)
281 (amide cleavage)
204 (α -cleavage)
3.14, m/z 380 (MH+)
138
Scheme 3.8: Possible fragmentation of the molecular ion m/z 380 corresponding to an
HNE-(N-acetyl-Gly-Lys-OMe) pyrrole adduct. See Appendix Scheme 3.6S for fragment
structures.
A 1:2 ((N-acetyl-Gly-Lys-OMe):HNE) Schiff base-Michael adduct 3.15 has a
mass of m/z 656 which is same as that of the d4-HNE-(N-acetyl-Gly-Lys-OMe)-(N-
acetyl-Gly-Lys-OMe) crosslink 3.16.51 Since the amounts were low and peaks not
apparent from the TIC (Figure 3.1A), the tandem mass spectra was recorded based on the
structure proposed by Xu et al. This makes ESI-MS/MS of the molecular ions that
correspond to 3.16 at m/z 652 and m/z 656 difficult to analyze (Scheme 3.9). The
fragmentation shows peaks with M /M+ 4 patterns, nevertheless the assignment of
structures to the fragments was not possible due to interferences from 3.15 (Figure 3.10).
75

Figure 3.10: ESI-MS/MS analysis of molecular ions A) m/z 656 and B) m/z 652
corresponding to the 1:2 (HNE:(N-acetyl-Gly-Lys-OMe)) crosslink proposed by Xu et
al.51
NH
O
N
O
HNO
O
OH
HN
O
N
OHN
OO
N
HNO
HN
O
HN
O
O
OH
HN
OO
NH
O
O
Lysine-Lysine Crosslink
3.15, m/z 656 3.16, m/z 652
Scheme 3.9: Structure of (N-acetyl-Gly-Lys-OMe)–HNE (2:1) crosslink proposed by
Xu et. al.51
Ions that correspond to more than one HNE adducted onto N-acetyl-Gly-Lys-OMe
were observed analogous to the (N-acetyl-His)-HNE adducts. The total ion
76

chromatogram showed ions at m/z 572,576,580 and m/z 728,732,736,740 along with the
fragments derived from dehydration of these adducts (Figure 3.1, panel A). The ESI-
MS/MS of these molecular ions generated prominent daughter ions from the loss of
water, an acetyl group, and HNE. A set of MS/MS spectra corresponding to 2:1 adducts
with m/z 572, 576 and 580 (corresponding to M, M+4 and M+8) and those of 3:1
(HNE:(N-acetyl-Gly-Lys-OMe)) adducts at with m/z 728, 732,736 and 740
(corresponding to M, M+4, M+8 and M+12) are shown in Figure 3.11.
Figure 3.11: ESI-MS/MS analysis of molecular ions corresponding to the 2:1 (HNE:(N-
acetyl-Gly-Lys-OMe)) adducts A) m/z 580, B) m/z 576, C) m/z 572 and the 3:1
(HNE:(N-acetyl-Gly-Lys-OMe)) adducts D) m/z 740, E) m/z 736 and F) m/z 732 G) m/z
728.
77

The (N-acetyl-Gly-Lys-OMe)-HNE reaction mixture was separated by HPLC
using a methanol/water gradient and the molecular ions (discussed above) were
monitoring by LC-ESI through SIR of 17 channels (Figure 3.12). The chromatograms of
the adducts which differ by the type of HNE (d0 or d4) adducted, had similar features
(retention times of peaks). Also the complexity of the chromatogram peaks increases with
increasing complexity of the adducts formed.
Michael adduct
Schiff base adduct
Pyrrole adduct
Michael adduct
Schiff base adduct
Pyrrole adduct
Figure 3.12: LC-ESI for the N-acetyl-Gly-Lys-OMe and HNE reaction mixture
monitored through SIR of 17 channels of which the spectrum of 7 channels are shown
here (Chromatograms for other channels are presented in Appendix). LC-SIR
chromatograms for molecular ions at A) m/z 732 (M+4 of 3:1 adduct), B) m/z 728 (M of
3:1 adduct). C) m/z 576 (M+4 of 2:1 adduct), D) m/z 572(M of 2:1 adduct); E) m/z 416
(M of 1:1 Michael adduct); F) m/z 398 (M of 1:1 Schiff base adduct); G) m/z 380 (M of
1:1 pyrrole adduct); and H) total ion chromatogram (TIC). (See Appendix Figure 3.3S for
M+8 and M+12 of 3:1 adducts and M+4 of Michael, Schiff base and pyrrole 1:1 adducts)
78

(iii) Cysteine HNE adducts. ESI-MS analysis of a reaction between N-acetyl-Cys and
HNE (d0 : d4) in PBS buffer at 37 ºC for 24 h, revealed the presence of several multiple
ion sets as observed in the reactions of lysyl residues in N-acetyl-Gly-Lys-OMe and the
histidine derivative, N-acetyl-His. The fragmentation of N-acetyl-Cys was obtained as
reported earlier (Figure 3.13).52
Figure 3.13: ESI-MS/MS analysis for N-acetyl-cysteine.
Cysteine forms a dimer under oxidative conditions through a disulphide bond.53
The molecular ion at m/z 325, corresponding to the dimer of N-acetyl-Cys, generated
daughter ions at m/z 307, 285 and 208 (Figure 3.14 and Scheme 3.10).
Figure 3.14: ESI-MS/MS analysis for N-acetyl-cysteine dimer at m/z 325.
79

HN
HO
O
S
O
NH
OHO
S
O307(M+ - OH)
281 (M+ - CH3CO)
m/z 324 (M+)
208 (α-cleavage)
Scheme 3.10: Suggested mode of fragmentations for N-acetyl-cysteine dimer m/z 324.
See Scheme 3.7S in Appendix for fragment structures.
The Michael adducts of N-acetyl-Cys with HNE (d0:d4) have calculated
molecular masses of m/z 319 and m/z 324. Since both the N-acetyl-Cys dimer and the
isotope labeled (N-acetyl-Cys)-HNE Michael adduct have the same m/z value, the
MS/MS spectra of (N-acetyl-Cys)-HNE adduct ions are complicated. However, the ions
that correspond to the loss of water, i.e., m/z 302 and m/z 306, exhibit fragmentations
without any interference as shown in Figure 3.15. The ions at m/z 139 and 143
correspond to the daughter ions generated by retro-Michael cleavage (vide supra).
Similar fragmentation of a cysteine-HNE Michael adduct was reported earlier using a
deuterated cysteine.54 The molecular ions at m/z 342 and 346 correspond to the M + Na+
ion of the Michael adduct.
80

Figure 3.15: ESI-MS/MS analysis of the dehydrated (N-acetyl-Cys)–HNE Michael
adduct.
NH
OHO
S
O
O
OH
260(M - CH3CO - H2O)
139(M - NAC - H2O)
164 (M -HNE)
m/z 302 (MH+ - H2O)
242 (M - CH3CO - 2H2O)
Scheme 3.11: Fragmentation of the molecular ion m/z 302 arising from the dehydration
of (N-acetyl-Cys)-HNE Michael adduct. See Scheme 3.8S in Appendix for fragment
structures.
The total ion chromatogram of the ESI-TOF-MS (Figure 3.1B) of the (N-acetyl-
Cys)-HNE reaction product mixture shows many peaks that have the signature multiple
ions with m/z incremented by 4 Da. The tandem MS/MS of these ions was recorded.
81

Here, we present some of the tandem MS/MS data of molecular ions that were observed,
but a definite structure could not be assigned based only on the fragmentation.
In the case of the doublet at m/z 465 and 469 ions (Figure 3.1B), may correspond
to an adduct formed by N-acetyl-Cys and HNE in a ratio of 2:1. Tandem MS/MS
analysis of these ions generated daughter ions at m/z 327,302,164 and 327,304,164
respectively (Figure 3.16) (see section 3.3 for discussion, page 45).
Figure 3.16: ESI-MS/MS analysis for molecular ion at A) m/z 469 and B) m/z 465
corresponding to HNE-(N-acetyl-Cys) (1:2) adduct.
A triplet in the TIC at m/z 603, 607 and 611 (Figure 3.1B) correspond to 2HNE+
2(N-acetyl-Cys) - 2H2O (Figure 3.17). ESI-MS/MS spectra of these adducts showed a
fragmentation with 4 set of peaks (m/z [465,327,302,164]; [465,469, 327, 306,302,164];
[469,327,306,164]) (Figure 3.17, see section 3.3 for discussion, page 45). These values
include the daughter ions obtained from the tandem MS/MS 2:1 (N-acetyl-Cys)/HNE
adduct m/z 465 shown above. Other multiplets that had mass difference of 4 Da (see
82

Figure 3.1, panel B) were too complicated to be analyzed based on the mass
fragmentation alone.
Figure 3.17: ESI-MS/MS analysis of the ions with A) m/z 611, B) m/z 607 and C) m/z
603, which correspond to 2:2 (N-acetyl-Cys)-HNE adducts.
LC-ESI analysis of the N-acetyl-Cys and HNE reaction mixture selectively monitored 13
channels accounting for most of the peaks that appear in the total ion chromatogram. ESI
chromatograms shown in Figure 3.18 represent the channels that have similar peak
features for molecular ion sets corresponding to an adduct (M) and its deuterium labeled
isomer(s) (M+4 and/or M+8). For example, molecular ions at m/z 603, 607 and 611 have
peaks with the same features, however, for the parent ions m/z 320 and 324 ((N-acetyl-
Cys)-HNE Michael adduct), the peak features are different. This difference may be due to
interference from molecular ions corresponding to (N-acetyl-Cys)-(N-acetyl-Cys) dimer
83

that also have m/z 324. So, the Michael adduct is monitored as the M + Na+ ion at m/z
342 and 346 (Figure 3.18, for discussions see section 3.3, page 45).
Figure 3.18: LC-ESI-SIR analysis of the molecular ions observed in (N-acetyl-Cys)-
HNE reaction mixture through 8 channels. (Chromatograms shown for 8 channels: A)
m/z 611, B) m/z 607, C) m/z 603, D) m/z 469, E) m/z 465, F) m/z 346, G) m/z 342 H) m/z
324 and I) TIC.
84

3.3. Discussion
HNE is a ubiquitous product of lipid peroxidation.23,55 In the present study, a new
approach to identify and characterize adducts formed by HNE with surrogate amino acid
residues is described. The methodology developed involves mass spectrometric analysis
of the adducts formed using d4-HNE : HNE (1:1) and amino acids, which confers the
spectrum with multiplets characteristic of the number of HNE present in the adduct.
Unlike previous methods, this approach provides an analytical tool for identification of
adducts that are labile under harsh conditions and/or present in low abundance. Adducts
previously cited, i.e., (see section 1.2) were characterized as reference standards for
solving the structures of previously unknown adducts. This study primarily focused on
the adducts formed by the nucleophilic side chains of lysine, histidine and cysteine with
HNE, because earlier reports suggested that other nucleophilic amino acids (arginine,
serine, methionine and tyrosine) required non-physiological conditions (high alkalinity
and temperatures >>37 oC) for adduct formation.56,57
Methodology. Incubation of amino acids individually with a mixture of deuterium
labeled (d4) HNE and unlabeled (d0) HNE in the ratio 1:1 can potentially yield adducts
that can be formed from either the d0 or d4 HNE. Analysis of these reaction mixtures by
mass spectrometry revealed that unique signatures are conferred upon the spectra. Sets of
ions are found with mass differences in multiples of 4 Da owing to the presence of
various combinations of d0 and d4 HNE. Thus, HNE with 4 deuterium atoms will have a
molecular weight of 156 + 4 Da. Therefore, an ion with ‘n’ HNE will have ‘n + 1’ peaks
with successive mass differences of 4 Da. Accordingly, adducts containing more than
85

one molar equivalent of HNE per molar equivalent of N-acetyl-His should appear as
triplets (1:2:1) or multiplets of n + 1 ions, where ‘n’ corresponds to the number of
equivalents of HNE present. Based on this premise, the following conclusions can be
drawn from the pattern of daughter ions derived from tandem MS/MS each of the
molecular ion (P) and their corresponding isotopic isomers ( P + 4 and/or P + 8 ):
a. If the daughter ion appears as M ( from P ) with no M+4 but M ( from P + 4 )
also, then the fragment has no C5-C9 alkyl chain of HNE.
P
M
M
Inte
nsity
m/z
MS/MS of P+ 4
MS/MS of P
P
M
M
Inte
nsity
m/z
MS/MS of P+ 4
MS/MS of P
Parent ion
Daughter ions
b. If the daughter ion appears as both M ( from P ) and M+4 ( from P + 4 ), then the
daughter ion has one C5-C9 alkyl chain from HNE.
P
M+4
M
m/z
Inte
nsity
MS/MS of P+ 4
MS/MS of P
P
M+4
M
m/z
Inte
nsity
MS/MS of P+ 4
MS/MS of P
Parent ion
Daughter ions
c. If the daughter ion appears as M ( from P ), M+4 ( from P + 4 ), and M+8 ( from
P + 8 ), then the daughter ion has two C5-C9 alkyl chains from HNE.
86

Inte
nsity
P
M
M+4
M+8
m/z
MS/MS of P+ 8
MS/MS of P
MS/MS of P+ 4
Parent ion
Daughter ions
Inte
nsity
P
M
M+4
M+8
m/z
MS/MS of P+ 8
MS/MS of P
MS/MS of P+ 4
Inte
nsity
P
M
M+4
M+8
m/z
MS/MS of P+ 8
MS/MS of P
MS/MS of P+ 4
Parent ion
Daughter ions
d. If the daughter ion of a parent ion with two C5-C9 alkyl chains from HNE
appears as [ M ( from P )] , [ M & M+4 ( from P + 4 )] and [ M+4 ( from P + 4 )],
then the daughter ion has one C5-C9 alkyl chain of HNE.
Inte
nsity
P
M
M+4
M+4M
m/z
MS/MS of P+ 8
MS/MS of P
MS/MS of P+ 4
Parent ion
Daughter ions
Inte
nsity
P
M
M+4
M+4M
m/z
MS/MS of P+ 8
MS/MS of P
MS/MS of P+ 4
Parent ion
Daughter ions
(N-acetyl-His)-HNE Adducts
Mass spectrometric analysis of an (N-acetyl-His)/HNE reaction mixture (Figure
3.1C, page 12) showed the presence of a unique triplet at m/z 510, 514 and 518 (1:2:1)
corresponding to an adduct with two HNE’s and one N-acetyl-His, in addition to the
known Michael adduct at m/z 354 and 358. Though, HNE has been suggested to undergo
aldol condensation32 and form adducts with the nucleophilic side chains of amino acids,
87

peaks corresponding to the HNE-HNE aldol (m/z 312) or dehydrated aldol product (m/z
298 ) were not detected under these reaction conditions. The molecular ion m/z 510 (156
+ 156 + 197 + H+) correspond to a molecule formed from two HNEs and one N-acetyl-
His without loss of any neutral molecules. The fragmentation pattern suggests that there
is minimal change in the basic structure of the reactants. The MS/MS of the m/z 510 ion
(Figure 3.1C) shows a strong signal for a m/z 354 daughter ion, which suggests that the
ion at m/z 510 is a simple adduct of d0-HNE with Michael adduct 3.11 (Figure 3.2).
Daughter ions of the 2:1 adduct that are unique compared to the daughter ions
from the Michael 1:1 adduct 3.11 (in the range of m/z 350-150), may be useful for
suggesting a core structure for the m/z 510 adduct. Some of these are shown in Figures
3.19 and 3.20. There are three sets of daughter ions, at a) m/z 209, 213; b) m/z 239, 243;
and c) m/z 310, 314 that are derived from the fragmentation of m/z 510, 514, 518, parent
ions of 2:1 (HNE:(N-acetyl-His)) adducts that are absent in the 1:1 adduct 3.11 (Figures
3.19, 3.20 and 3.21). As discussed earlier (previous page, rule d), these daughter ions are
derived from a parent ion with two HNEs and they are present in an M, (M & M+4) and
M+4 pattern. This implies that these peaks correspond to a fragment that contains one
C5-C9 alkyl chain. Some of the possible structures that have a molecular mass of 510
amu are shown in Scheme 3.12. Also, analysis of the set of daughter ions at m/z 408, 412
from the 2:1 adduct parent ions at m/z 510, 514, 518 are informative (vide infra).
88

C5H11
O OH
HNOH
O
N
N
O
HO
C5H11
OC5H11
OHOH
NH
HOO
N
N
O
OC5H11
O
m/z 510
C5H11
OH
O
HNHO
ON
N
O
O
C5H11
HO
3.21 3.19
3.20
O
OHHO
C5H11
OH
C5H11
HNHO
ON
N
O
3.22
Scheme 3.12: Possible isomers of the m/z 510 (2:1) HNE-(N-acetyl-Cys) adduct.
Among the structures in Scheme 3.12 that may correspond to the parent ion m/z
510, the daughter ions 310, 314 and 408, 412 are expected by α-cleavage in the case of
the acyclic 3.19 or 3.22 or 3.21 (Scheme 3.13, Figures 3.30 and 3.21) but not the bicyclic
3.20. Other daughter ions from the 2:1 adduct that are unique (Figures 3.19 and 3.20)
were also observed, but their structural significance could not be deduced.
89

OH
HNOH
O
N
N
O
O
O
OH
HNOH
O
N
N
O
m/z 408
OHOH
HNOH
O
N
N
O
O
O
m/z 510 m/z 310
Scheme 3.13: Possible fragmentation of the putative parent ion m/z 510 to generate a
unique daughter ion at m/z 310 and 408.
Figure 3.19: Tandem MS/MS in the m/z range 207-212 and 238-244 from the (N-acetyl-
His)-HNE (Figure 3.1C) of the parent ions A) m/z 358, B) m/z 354, C) m/z 518, D) m/z
514 and E) m/z 510. Only the (2:1) HNE/(N-acetyl-His) adducts C, D and E shows
unique daughter ions at m/z 209, 213 and m/z 239, 243.
90

OHOH
HNOH
O
N
N
O
O
O
310 (α−cleavage)
3.21, m/z 510
466 (MH+ - COOH)468 (M - CH3CO-H+)
408 (α−cleavage)
Scheme 3.14: Possible mass spectrometric cleavage sites of adduct at 510 m/z with a
structure 3.19. See Scheme 3.9S in Appendix for fragment structures.
Figure 3.20: Tandem MS/MS in the mass range m/z 309-316 of the parent ions at A) m/z
358, B) m/z 354, C) m/z 518, D) m/z 514 and E) m/z 510 from (N-acetyl-His)-HNE
(Figure 3.1C).
91

Figure 3.21: Fragmentation of parent ions from (N-acetyl-His)-HNE (Figure 3.1C) at A)
m/z 518, B) m/z 514 and C) m/z 510 showing the presence of daughter ions m/z 408
and/or m/z 412.
Each of the molecular ions in the quadruplet m/z 666, 670, 674 and 678 have a
definite fragmentation pattern making them unique molecular ions in contrast to being
simple aggregated molecules or clusters in the MS analyzer, which would have yielded
the corresponding molecular ion peaks. In spite of very low abundance of these adducts
the tandem MS/MS of their molecular ions made the structure determination possible.
The unique fragments from the 3:1 HNE-(N-acetyl-His) adduct, viz, m/z 395, 399 which
were absent in the fragmentation of the 2:1 HNE-(N-acetyl-His) adduct, m/z 510, greatly
facilitated assignment of structure (Scheme 3.15) (additional information in Appendix,
see Scheme 3.9S). In the tandem MS/MS of the diastereomeric 3:1 HNE-(N-acetyl-His)
adducts, daughter ions that correspond to two consecutive retro-Michael cleavages,
92

leading to the formation of the 2:1 and 1:1 adducts is observed (Figure 3.4). Scheme
3.15b (see below) illustrates this fragmentation.
OHOH
HNOH
O
N
N
O
O
HO
O
O
466 (M-HNE+H+)
395
3.22, m/z 666
467
Scheme 3.15a: Possible fragmentations of the m/z 666 3:1 (HNE/(N-acetyl-His)) adduct.
Inset (tandem MS/MS of the parent ions m/z A) 510, B) 674, C) 670 and D) 666 in the
range m/z 394-402). See Scheme 3.10S in Appendix for fragment structures.
OHOH
HNOH
O
N
N
O
O
O
O
HO
m/z 665
O OH
HNOH
O
N
N
O
OH
O
m/z 509 O OH
HNOH
O
N
N
O
m/z 354
Retro-Michael
Retro-Michael
3:1 HNE:NAH
2:1 HNE:NAH 1:1 HNE:NAH
Scheme 3.15b: Possible structures of daughter ions formed by retro-Michael cleavage of
the 3:1 parent ion m/z 666. (NAH = N-acetyl-His)
93

To establish semi-quantitatively (i.e., without internal standard) the amounts of
adducts formed between (N-acetyl-His) and HNE, each of the molecular ion adducts in
the reaction mixture as shown in Figure 3.5 was selectively monitored by LC-ESI.
Multiple peaks are observed for each parent ion m/z. This indicates that the reaction
product contains a complex mixture of isomers, presumably, diastereomers.
The relative amounts of each of the adducts were calculated based on the area
under the curve for each of the peaks in LC-SIR chromatogram (Figure 3.5). The amount
of Michael adduct accounts for about 70% of the adducts formed, while the 1:2 and 1:3
((N-acetyl-Gly-Lys-OMe):HNE) adducts are present in 25% and 6% abundances,
respectively.
0
10
20
30
40
50
60
70
80
Michaeladduct
(1:2)adduct
(1:3)adduct
Rel
ative
amou
nt (%
)
Figure 3.22: Relative amounts of adducts formed by (N-acetyl-His)/HNE reaction,
calculated using LC-ESI. The amounts reflect the relative amount (of the adducts
selectively monitored) of each of the adduct present in the reaction mixture. Values are
average of 2 independent experiments and the error bars indicate the range.
94

Multiple reaction monitoring (MRM) is a widely used technique to quantify
amounts of molecular ion using LC-MS/MS. MRM measures the amount of the
molecular ion of interest, based on its most abundant daughter ion formed during tandem
MS/MS. MRM analyses were not effective compared to selected ion monitoring (SIR)
analyses of molecular ions, probably due to the low abundance of their fragments. LC-
SIR measures the amount of the ion of interest irrespective of its daughter ions.
Lysine HNE adducts
The Michael adduct (Figure 3.7 and Scheme 3.6), Schiff base (Figure 3.8 and
Scheme 3.7), pyrrole (Figure 3.9 and Scheme 3.8) and crosslink (2:1, Lysine:HNE)
(Figure 3.7 and Scheme 3.6) type adducts formed by lysine with HNE were detected. The
fragmentation patterns of the corresponding molecular ions for the unmodified N-acetyl-
glycine-lysine methyl ester (N-acetyl-Gly-Lys-OMe) and the various adducts support the
structures suggested earlier.32,33,51
ESI-MS/MS of the (N-acetyl-Gly-Lys-OMe)-HNE (1:2) molecular ion generated
fragments with loss of H2O, acetyl and HNE’s (see Figure 3.10, panels A, B and C),
which were similar to fragments generated from the (N-acetyl-His)-HNE (1:2) adduct
(see Figure 3.3, panels A, B and C). This suggests that the molecular ions of the
corresponding multiple adducts may share similar structural characteristics. For example,
in the set of of tandem MS/MS spectra of molecular ions at m/z 572, 576, 580, that
correspond to the 1:2 ((N-acetyl-Gly-Lys-OMe):HNE) adducts, as well as in the set of
tandem MS/MS spectra of molecular ions at m/z 728, 732 and 736, that correspond to the
95
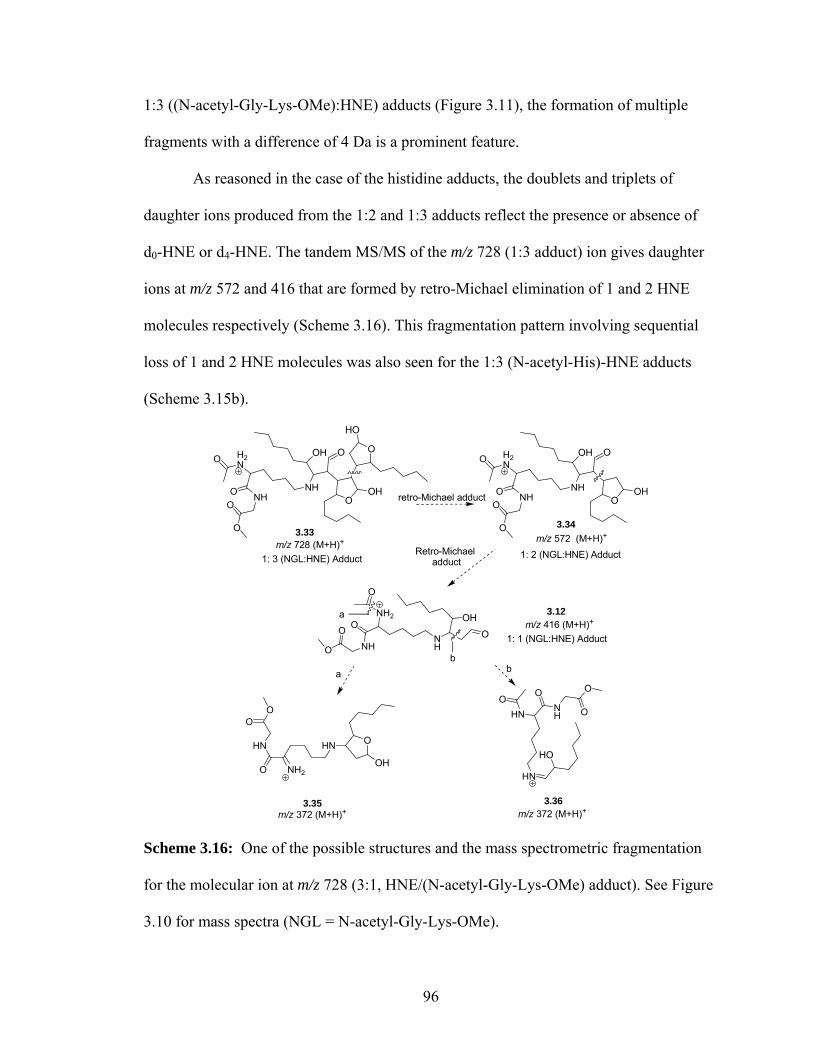
1:3 ((N-acetyl-Gly-Lys-OMe):HNE) adducts (Figure 3.11), the formation of multiple
fragments with a difference of 4 Da is a prominent feature.
As reasoned in the case of the histidine adducts, the doublets and triplets of
daughter ions produced from the 1:2 and 1:3 adducts reflect the presence or absence of
d0-HNE or d4-HNE. The tandem MS/MS of the m/z 728 (1:3 adduct) ion gives daughter
ions at m/z 572 and 416 that are formed by retro-Michael elimination of 1 and 2 HNE
molecules respectively (Scheme 3.16). This fragmentation pattern involving sequential
loss of 1 and 2 HNE molecules was also seen for the 1:3 (N-acetyl-His)-HNE adducts
(Scheme 3.15b).
H2N
O NH
O
NH
O
O
OOH
OHO
HO
O
NH2O
HNHN
OO
OH
O
m/z 372 (M+H)+
HN
O
HN
ONH
O
O
HO
m/z 372 (M+H)+
m/z 728 (M+H)+ m/z 572 (M+H)+3.333.34
3.35 3.36
retro-Michael adduct
Retro-Michael adduct1: 3 (NGL:HNE) Adduct 1: 2 (NGL:HNE) Adduct
NH2O
NH
O
NHO
O O
OHm/z 416 (M+H)+
1: 1 (NGL:HNE) Adduct
3.12a
ba b
H2N
O NH
O
NH
O
O
OOH
OHO
Scheme 3.16: One of the possible structures and the mass spectrometric fragmentation
for the molecular ion at m/z 728 (3:1, HNE/(N-acetyl-Gly-Lys-OMe) adduct). See Figure
3.10 for mass spectra (NGL = N-acetyl-Gly-Lys-OMe).
96

A semi-quantitative analysis of the chromatogram of the adducts detected by LC-
ESI-SIR shows virtually identical mass chromatograms for each of the isotopic molecular
ions (M, M+4, …, M + 4n) of the 3:1 and the 2:1 adducts (Figure 3.12, page 24). In
addition to the fragmentation pattern discussed earlier, the similarity in the SIR
chromatograms suggests that the structure corresponding to the isotopic molecular ions
(d0 and d4) of an adduct are identical.
In addition, the relative amounts of each of the adduct based on the area under the
curve of LC-SIR spectra (Figure 3.12) showed the predominance of the 1:1 adducts,
Schiff base (49%) and Michael adduct (29%), over the multiple HNE-(N-acetyl-Gly-Lys-
OMe) adducts (2:1 (~9%) and 3:1 (9%) adducts) (Figure 3.23). The amounts of
relatively stable adducts, pyrrole and crosslink ((N-acetyl-Gly-Lys-OMe):HNE (2:1)),
were 10% and 1% respectively.
0
5
10
15
20
25
30
35
40
45
50
Pyrrole SchiffBase
Michael Crosslink (1:2)adduct
(1:3)adduct
Rela
tive
amou
nts (
%)
Figure 3.23: Relative amounts of adducts formed by (N-acetyl-Gly-Lys-OMe)/HNE
reaction, calculated using LC-ESI. The ‘y’ scale reflects percentage of the each adduct
present in the reaction mixture (with respect to the adducts selectively monitored). Values
are average of 2 independent experiments and the error bars indicate the range.
97

HNE- Cysteine adducts
Cysteine forms a Michael adduct with HNE and to the best of our knowledge
there have been no reports of any other adducts that can be formed. Here, HNE adducts
with the cysteine were analyzed using N-acetylated cysteine (N-acetyl-Cys). The
molecular ion corresponding Michael adduct with HNE and the (N-acetyl-Cys)-(N-
acetyl-Cys) (disulphide) were analyzed, and the fragmentation pattern was in agreement
with the structure.54
A prominent doublet in the total ion chromatogram of the (N-acetyl-Cys)-HNE
reaction mixture occurs at m/z 465 and 469 (Figure 3.1, panel B). This corresponds to a
2:1 adduct of N-acetyl-Cys and HNE. As shown in Scheme 3.17, the tandem MS/MS of
these ions (Scheme 3.16) results in daughter ions at m/z 164 (corresponding to (N-acetyl-
Cys)+H+) and 302 or 306 (corresponding to (N-acetyl-Cys)+HNE(or d4-HNE)-H2O +H)+.
Another prominent daughter ion from both the parents has m/z 327 that indicates loss of
the d4 labeled portion of the adduct. However, the structure of this daughter ion is not
obvious, and was not assigned.
m/z 465 (2NAC + HNE - H2O+ H+)
m/z 302(NAC + HNE - H2O+ H+)
m/z 327?
m/z 164 (NAC + H+)
m/z 469 (2NAC + (d4-HNE) - H2O+ H+)
m/z 306
(NAC + (d4-HNE) - H2O+ H+)
m/z 327?
m/z 164 (NAC + H+)
A B
Scheme 3.17: Daughter ions formed from the tandem MS/MS of 2:1 (N-acetyl-Cys):
HNE adduct A) d0-HNE-2(N-acetyl-Cys) adduct B) d4-HNE-2(N-acetyl-Cys) adduct,
(NAC = N-acetyl-cysteine).
98

The 2:2 adducts that correspond to the molecular ions at m/z 603, 607 and 611
(Figure 3.1, panel B) may be composed of 2HNE+ 2 (N-acetyl-Cys) - 2H2O. ESI-
MS/MS spectra of these adducts showed a fragmentation with 4 prominent peaks (Figure
3.17) as depicted in Scheme 3.18. A structure was not assigned to this 2:2 adduct. Unlike
(N-acetyl-His) and N-acetyl-Gly-Lys-OMe adducts, peaks corresponding 2:1 or 3:1
adducts of HNE with N-acetyl-Cys were not observed.
302 302 306
327327
164164
302
327
164
(2NAC + 2(HNE) - H2O+ H+) (2NAC + HNE + d4-HNE - H2O+ H+) (2NAC + 2(d4-HNE) - H2O+ H+)
603 (M) 607(M+4) 611(M+8)
465 (M) 465(M) 469(M+4) 469(M+4)
2:2 adducts
2:1 adducts
Scheme 3.18: Daughter ions formed from the tandem MS/MS of 2:2 (N-acetyl-cysteine:
HNE) adduct m/z 603, 607 and 611. (NAC = N-acetyl-cysteine)
99

The relative amounts of each of the adducts with respect to the total number of
adducts monitored showed that the Michael adduct (~50%) was the predominant product
compared to the other adducts (Figure 3.24). Surprisingly, 2:1 and 2:2 adducts were
present in comparatively high levels i.e. ~30 % and ~5% respectively.
0
10
20
30
40
50
60
70
MichaelAdduct
NAC-NACdimer
2:1 adduct 2:2 adduct
Rel
ativ
e am
ount
s (%
)
Figure 3.24: Relative amounts of adducts formed by N-acetyl-Cys and HNE reaction,
calculated using LC-ESI. The ‘y’ scale reflects percentage of the each adduct present in
the reaction mixture (with respect to all the adducts selectively monitored). Values are
average of 2 independent experiments and the error bars indicate the range, (NAC = N-
acetyl-cysteine).
100

3.4. Conclusions
Using a mixture of deuterium labeled HNE and unlabeled HNE in the ratio (1:1)
to study the adduction pattern of HNE on lysine, histidine and cysteine is a robust method
for the characterization of unique adducts. Adducts formed with HNE appear as apparent
multiplets that are indicative of the number of HNE moieties present in each adduct. The
tandem mass spectra of each of the adducts gave sets of unique daughter ions. This
method is valuable for detecting and characterizing different types of adducts that are
formed under mild conditions, including some present in low abundances.
Apart from adduct types reported earlier from the reactions of lysine or histidine
with HNE, the present studies with isotope labeled HNE revealed adducts that
incorporate multiple HNE molecules adducted with a single lysine or histidine residue.
Possible structures for some of the multiple HNE adducts were proposed based on the
unique fragments that were identified. The relative amount of 1:2 plus 1:3 adducts
formed from N-acetyl-Gly-Lys-OMe was about 19%. The amount of 1:1 adducts
(Michael, Schiff base and pyrrole adduct) was about 80% of the total adducts and the 2:1
(N-acetyl-Gly-Lys-OMe)/HNE crosslink comprised about 1% of the total adducts
monitored.
Apart from forming predominantly Michael adduct, incubation of N-acetyl-
cysteine with HNE also generated relatively high amounts of unique adducts with (N-
acetyl-Cys):HNE in the ratio 2:1(~30%) and 2:2 (~5%).
Though high concentrations of HNE might only be possible in certain micro-
domains in situ, e.g. oxidized LDL particles, identification of such multiple HNE adducts
may help in attaining a complete understanding of HNE’s role in physiological processes.
101

The multiple HNE-amine adducts discussed here may constitute an initial response of
proteins towards high levels of HNE. These multiple HNE adducts may rearrange to form
other toxic products or by corollary, this could be one of the in vivo homeostatic
mechanisms to detoxify lipid oxidation products by effectively quenching the HNE’s
reactivity.
102

3.5. Experimental Methods and Procedures
General methods. Proton magnetic resonance (1H NMR) spectra were recorded on a
Varian Gemini spectrometer operating at 200, 300, 400 or 600 MHz. Proton chemical
shifts are reported in parts per million (ppm) on the δ scale relative to CDCl3 (δ 7.25) or
CD3OD (δ 3.30). 1H NMR spectral data are tabulated in terms of multiplicity of proton
absorption (s, singlet; d, doublet; t, triplet; m, multiplet; br, broad), coupling constants
(Hz), number of protons. Carbon magnetic resonance (13C NMR) spectra were recorded
on a Varian Gemini spectrometer operating at 50 MHz and chemical shifts are reported
relative to solvents. Deuterium magnetic resonance (2H NMR) spectra were recorded on a
Varian Gemini spectrometer (400 MHz) and chemical shifts are reported relative to
solvents. All high resolution mass spectra were recorded on a Kratos AEI MS25 RFA
high resolution mass spectrometer at 20 eV. All solvents were distilled under a nitrogen
atmosphere prior to use. Diethyl ether was freshly distilled over lithium aluminum
hydride. Pentane was dried over phosphorus pentoxide. Tetrahydrofuran (THF) was
freshly distilled over potassium and benzophenone. All reaction flasks were flame dried
under argon before use. All materials were obtained from Aldrich (Milwaukee, WI)
unless otherwise specified.
Chromatography was performed with ACS grade solvent (ethyl acetate and
hexanes). Thin layer chromatography (TLC) was performed on glass plates precoated
with silica gel (Kieselgel 60 F254, E. Merck, Darmstadt, West Germany). Rf values are
quoted for plates of thickness 0.25 mm. The plates were visualized with iodine or
phosphomolybdic acid reagent.58 Flash column chromatography was performed on 230 -
400 mesh silica gel supplied by Merck. All other chemicals were obtained from Aldrich
103

(Milwaukee, WI) and Fisher (Chicago, IL). For all reactions performed in an inert
atmosphere, argon was used unless specified.
Synthesis of [8,8,9,9-2H4]-4-hydroxy-2-nonenal.
OSO
OOH TsCl+
3.2Pent-4-yn-1-ol
Et3N
CH2Cl2, 0oC
86%3.1
Toluene-4-sulfonic acid, pent-4-ynyl ester (3.2). A suspension of p-toluenesulfonyl
chloride ( 3.4 g, 17.9 mmol) and triethylamine (6 mL) in CH2Cl2 (25 mL) was added
dropwise to pent-4-yn-1-ol (1 g, 11.9 mmol) at 0 ºC and the reaction mixured was stirred
at 0 ºC. When TLC showed no starting material, the reaction mixture was extracted with
ethyl ether (3 x 20 mL). The extract was washed with saturated sodium bicarbonate,
water, and brine. After drying over MgSO4, the solvent was removed in vacuo and
purified by column chromatography with 5% ethyl acetate in petroleum ether (Rf = 0.7)
to yield an oily liquid 3.2 (2.43 g, 86%).45
OSO
OHD2C
D2C O
SO
O
3.3
RhCl(PPh3)3, D2
C6H6
74%3.2
104

Toluene-4-sulfonic acid, [4,4,5,5-2H4] pent-4-nyl ester (3.3). To a flask containing
benzene (2 mL) flushed with argon was added freshly prepared the Wilkinson catalyst
RhCl(PPh3)3 (12 mg, 12.9 µmol).59 The flask was flushed with argon for 5 min and the
valve turned to deuterium gas. The color of the solution turned ice-tea brown. After 5
min, a solution of 3.2 (336 mg, 1.41 mmol) in degassed benzene (1 mL) was added
dropwise. The reaction was allowed to proceed under a positive pressure of D2 until the
reaction ceased to consume D2. The solvent was removed on a rotatory evaporator. The
product was extracted by trituration with hexanes (3 x 20 mL) and purified by column
chromatography with 5% ethyl acetate in hexanes (Rf = 0.5) to yield a oily liquid 3.3
(260 mg, 74%). 1H NMR (CDCl3, 300 MHz) δ 7.77 (d, J1 = 6.2 Hz, J2 = 1.6 Hz, 2H),
7.33 (d, J = 8.4 Hz, 2H), 4.02 (t, J = 6.5 Hz, 2H), 2.45 (s, 3H), 1.63-1.64 (2H), 1.26 (t, J
= 7.2 Hz, 2H), 0.8031 (bs, 1H); 13C NMR (CDCl3, 75 MHz) δ 144.8, 133.4, 129.9, 128.0,
70.9, 28.6, 27.3, 21.8, 21.1(t), 13.3(q); HRMS (EI) m/z calcd for C12H14D4O3S+(M+)
246.1224 found 247.1292 (M+H)+.46
105

HD2C
D2C IHD2C
D2C O
SO
O
3.3
NaI,Acetone
3.4
HD2C
D2C I
3.4
+O
OOO
OHD2CCD2 OH
tBuLi
Ethyl ether
3.5-78 oC
84% 3.6
[7,7,8,8-2H4]-1-(2,2-Dimethyl-[1,3]dioxolan-4-yl)-oct-1-en-3-ol (3.6). To a stirred
solution of sodium iodide (750mg, 5 mmol) in freshly distilled acetone (15 mL) was
added potassium carbonate (550 mg, 4 mmol) and tosylate 3.3 (500 mg, 2 mmol).36 The
solution was refluxed for 3 h or until the reaction is complete. The solvent was removed
on a rotatory evaporator and the crude product dried on a vacuum pump. This product
was used for the next step without purification. 2H NMR (CDCl3, 90MHz) δ 1.19, 1.12,
0.88, 0.74 1H NMR (CDCl3, 300 MHz) δ 3.19 (t, J = 7 Hz, 2H), 1.78-1.83 (m, 2H), 1.36
(t, J = 7.7 Hz, 2H), 0.86 (s, 1H).
A solution of t-butyllithium (6 mmol) in hexanes was added dropwise to a stirred
solution of 3.4 (~150 mg, 0.74 mmol) in freshly distilled ethyl ether:pentane (2:1, 15 mL)
at -78 ºC under argon. After 10 min at -78 ºC, the reaction mixture was allowed to warm
to room temperature to generate the alkyllithium. The reaction mixture was then cooled
to -78 ºC and 3-(2,2-dimethyl-[1,3]dioxolan-4-yl)-propenal (3.5) 60 (108 mg, 0.7 mmol)
was added dropwise with stirring over 5 min. After stirring the resulting mixture for 30
106

min, saturated NH4Cl was added. The resulting mixture was extracted with ethyl ether (3
x 15 mL). The combined organic extract was washed with brine and solvent removed on
a rotatory evaporator. The crude product was purified on a flash column with 25% ethyl
acetate in hexanes to yield 3.6 (136 mg, 84%). 1H NMR (CDCl3, 300 MHz) δ 5.82 (dd,
J = 15.4, 6.1 Hz, 1H), 5.64 (ddd, J = 16.5, 7.4, 2.3 Hz, 1H), 4.5 (dd, J = 13.7, 7.1 Hz, 1H),
4.06-4.13 (m, 2H), 3.57 and 3.58 (2t, J = 7.9 Hz, J = 7.7 Hz, 1H), 1.25-1.60 (8H), 1.40 (s,
3H), 1.37 (s, 3H), 0.82 (s, 1H); 13C NMR (CDCl3, 300 MHz, some resonances appear as
1:1 apparent doublets owing to the presence of two diastereomers) δ 137.46 and 137.39,
127.81 and 127.76, 109.40 and 109.38, 76.57, 72.04 and 71.98, 69.49 and 69.47, 37.15
and 37.05, 31.48, 26.71, 25.92, 25.05 and 24.99 , 21.62(q), 13.23 (q) HRMS m/z calcd
C13H20D4O3 232.1972 (M)+, found 233.2035 (M+H)+, 231.1872 (M-H)+
[8,8,9,9-2H4]-4-Hydroxy-2(Z)-nonenal (3.7). This product was accessed by two
different methods.
NaIO4
acetic acid:H2O (2:1)
OOHD2C
CD2 OH
OHD2C
CD2 OH
3.6 3.789%40 oC
Method A: A suspension of sodium periodate (24 mg, 0.117 mmol) in acetic acid/water
(2:1, v/v, 2 mL) was added to compound 3.6 (13 mg, 0.056 mmol) and the mixture was
stirred magnetically for 4 h at 40 °C or until the reaction was complete. Then, the solvent
was removed by rotary evaporation under reduced pressure. The last traces of HOAc
were removed by azeotropic distillation with n-heptane (3:1 mL) under high vacuum.
107

The resulting residue was flash chromatographed on a silica gel column (30% ethyl
actetate in hexanes) to give 3.7 (8 mg, 89%).
1. acetic acid:H2O (2:1), 40 o C2. Pb(OAc)4, K2CO3, -78 oC
OOHD2C
CD2 OH
OHD2C
CD2 OH
3.6 3.7
78%
Method B: A solution of the compound 3.6 (13 mg, 0.056 mmol) in acetic acid/water
(2:1, v/v, 1.2 mL) was stirred magnetically for 4 h at 40 °C. Then the solvent was
removed by rotary evaporation under reduced pressure. The last traces of HOAc were
removed by azeotropic distillation with n-heptane (3:1 mL) under high vacuum. Dry
methylene chloride (2 mL) and Na2CO3 (5.7 mg, 0.054 mmol) were added to the residue.
The solution was stirred magnetically at -78 °C under an argon atmosphere, and
Pb(OAc)4 (31 mg, 0.07 mmol) was added. The resulting solution was stirred for 30 min.
The solvent was then removed. The residue was flash chromatographed on a silica gel
column (30% ethyl actetate in hexanes) to give 3.7 (7 mg, 78%). 1H NMR (CD3Cl, 300
MHz) δ 9.57 (d, J = 7.7 Hz, 1H), 6.80 (dd, J1 = 15.7 Hz, J2 = 4.5 Hz, 1H), 6.29 (dd, J1 =
15.7 Hz, J2 = 7.7 Hz, 1H), 4.02 (t, J = 6.5 Hz, 2H), 2.45 (s, 3H), 1.63-1.64 (2H), 1.26 (t,
J = 7.2 Hz, 2H), 0.8031 (bs, 1H); 13C NMR (CD3Cl, 75 MHz) δ 193.1, 158.7, 130.6,
92.3, 71.1, 36.5, 31.3, 29.2, 24.8; HRMS (EI) m/z calcd for (M+) 246.1224 found
247.1292 (M+H)+.
108

Preparation of cyclohexanedione (CHD) reagent. Cyclohexanedione (CHD) was
freshly recrystallized from ethyl acetate. CHD (125 mg), ammonium acetate (5g), and
acetic acid (2.5 mL) were dissolved in deionized water and diluted to 50 mL. The pale
yellow solution was heated at 60 °C for 1 h, cooled with an ice bath, and purified by
passing through a C18 SPE cartridge (1 mL), which was preconditioned with water (5
mL) and methanol (5 mL). The elute was collected and stored at 4 °C.42
Derivatization of 4-hydroxy-2-nonenal by CHD reagent. Stock solutions of HNE, d4-
HNE and benzaldehyde (100 ng/µL) were prepared. Benzaldehyde solution (10 µL) was
mixed individually with 10 µL and 50 µL of HNE and d4-HNE solutions. The solvent
was removed with a gentle steam of dry nitrogen, and the residue was redissolved in 100
µL of methanol. CHD reagent (1 mL) was added, and the mixture was incubated at 60 oC
for 1 h. The resulting solution was cooled, and then passed through a C18 SPE cartridge,
which had been washed sequentially with methanol (5 mL x 2) and water (5 mL x 2). The
cartridge was washed with 5% acetonitrile in water (9 mL) and the product was eluted
with acetonitrile containing 5% water (9 mL). The solvent in the eluant was removed
with a stream of dry nitrogen. Before ESI-MS/MS analysis, the residue was dissolved in
methanol containing 25% water, and diluted to 0.01 ng/µL (final concentration) was
injected into the LC-MS for analysis. ESI-MS/MS analysis of the HNE-CHD derivative
produced characteristic fragments: m/z 326 ([M+H-18]+); m/z 308 ([M+H-18-18]+); m/z
216. ESI-MS/MS analysis of the d4-HNE-CHD derivative produced characteristic
fragments: m/z 330 ([MH-18]+); m/z 312 ([MH-18-18]+); m/z 216. ESI-MS/MS analysis
109

of benzaldehyde-CHD derivative produce characteristic fragments: m/z 294 ([MH]+); m/z
216. (Figure 3.25).
HN
OHC5H7D4
O O
HN
OHC5H11
O O
HN
OHC5H7D4
O O
HN
OHC5H11
O O
A B
Figure 3.25: A) ESI-MS/MS of CHD derived HNE B) ESI-MS/MS of CHD derived d4-
HNE.
Synthesis of HNE adducts of amino acids: A deuterium labeled [8,8,9,9-2H4]-HNE
solution (5 mM in ethyl acetate) was prepared freshly and added to a solution of
unlabeled HNE (5 mM in ethyl acetate) to make a 1:1 stock solution. The concentration
of the stock HNE solution was determined by measurement of UV absorbance at 224 nm
presuming a molar extinction coefficient of 13,750 M-1 for HNE in MeOH.61 To a 1.5
mL glass vial was added 1 mL stock solution of HNE (d4:d0) and the solvent evaporated
using a gentle stream of N2 gas.
To a solution of N-acetyl-lysine (20 µM) in PBS (10 mM, pH 7.4) was added d4-
HNE and HNE in the ratio (1:1) (10 µM). The mixture was incubated at 37 ºC for 24 h.
110

In rare cases longer reaction times were required for the formation of adducts that are
present in low abundances but are of interest for this study, e.g., 36-48 h at room
temperature for the 3:1 (HNE:(N-acetyl-Gly-Lys-OMe)) adduct. Following the reaction,
the reaction mixture is quickly dried using a high-speed vacuum evaporator and stored at
-20 ºC until used. The isolated aliquot was analyzed by either MALDI-TOF or QTOF2
mass spectrometer.
Mass spectrometry. ESI-MS/MS analysis of CHD-d4-HNE derivative was performed
on API-3000 triple quadrapole electrospray mass spectrometer (Applied Biosystems
Inc.). The source temperature was maintained at 100 ºC, and the desolvation temperature
at 200 °C. The drying gas (N2) was maintained at ca. 450 L/Hr, and the cone flow gas at
ca. 50 L/Hr. The multiplier was set at an absolute value of 500. MS scan at m/z 50-800
were obtained for standard compounds.
Table 3.3. Optimized parameters for mass spectrometer.
Ion Mode positive Capillary (KV) 3 Cone (V) 30 Hex 1 (V) 10 Aperture (V) 0 Hex 2 (V) 0.5 LM 1 Resolution 19 HM 1 Resolution 19 Ion Energy 1 1 LM 2 Resolution 19 HM 2 Resolution 19 Ion Energy 2 2
111

Argon was used as collision gas at a pressure of 5 psi for MS/MS analysis. For MS/MS
analysis, the collision energies were optimized for each compound. The instrument was
operated in the positive ion mode.
Analysis of adducts using MALDI-TOF. Initial diagnostic analysis of the samples was
performed by MALDI-TOF mass spectrometry using a PE Biosystems Voyager DE Pro
instrument equipped with a nitrogen laser (337 nm) and operated in the delayed
extraction and reflector mode with a matrix of α-cyano-4-hydroxycinnamic acid (CHCA,
5 mg/ml in acetonitrile/water/3% trifluoroacetic acid, 5:4:1, v/v/v). Internal standards
used for calibration included two synthetic peptides G9I (50 fmol/µL, MH+ 1015.579)
plus L20R (200 fmol/µL, MH+ 2474.6300)29. Each of the reaction aliquot was diluted in
50% aqueous acetonitrile solution to give a final concentration of 100 fmol/µL of the
total amino acid. Each of the solution is properly vortexed before used. One microliter of
sample was mixed with 1 µL of matrix and 0.5 µL of internal standard mix then 1.5 µL
applied to the target and allowed to dry. Each spectrum was accumulated for ~ 50-200
laser shots. The data was processed on Data Explorer 4.0 (Applied Biosystems) software.
Additional sample applications were utilized as needed.
Analysis of adducts using Q-TOF mass spectrometer. The samples were analyzed by a
quadrupole-time of flight (Q-TOF-2) mass spectrometer (Micromass) equipped with
MassLynx™ acquisition and processing software. The samples were infused using a 100
µL Hamilton syringe onto a capillary column (PicoFrit™ 0.050 x 50mm, 5µ tip ID; New
Objective Inc., Woburn, MA) at 0.5 µL/min using a syringe pump (Harvard apparatus,
112

Pump 11) and ionized immediately using an electrosprayer designed in-house. The mass
spectrometer was operated in standard MS and MS/MS switching mode. The MS data
was collected for the mass range of 50-1500 m/z and MS/MS data analyses utilized
Micromass software (MassLynx®) Version 3.5.
The machine was calibrated using a solution of 2 fmol of [Glu1 ]-Fibrinopeptide
B (Sigma) in 50% aqueous acetonitrile (+ 0.1% formic acid) infused using glass
micropipette with 5 µ tip. The intensity of the peak from the MS/MS spectrum at 785.4
m/z was used as a reference for calibration. The final mass measurement accuracy of ≤ 10
ppm was achieved before the use of the machine.
Chromatography. The samples analyzed for selected ion recording were
chromatographically separated on a reverse phase column and recorded in positive mode.
The chromatographic separation was obtained using a 150 × 2.0 mm i.d. Prodigy ODS-2,
5 µ column (Phenomenex, UK), with a binary solvent (water and methanol) gradient.
The solvents were supplemented with 0.2% formic acid. The separation was effected
through a gradient started with 100% water and rose to 100% methanol linearly in 15
min, and elution was continued for 5 min with 100% methanol. Then the gradient was
reversed to 100% water in 2 min, and then held for 5 min at 100% water. The solvents
were delivered at 200 µl/min.
Method to quantify the relative amounts of adducts formed. The selective
monitoring of LC-ESI chromatograms for each of the adduct was integrated using the
Masslynx® software ver 3.5. The total peak area of the chromatogram for each of the
113

adduct (A) was determined and using the formulae below the values of relative amounts
in percentage was calculated. The error bars reflect the results of two independent
experiments.
n – the total number of adducts
A(n) – Total area of the chromatogram for the adduct A(n) in A(n) channel of LC-ESI-
SIR.
LC-ESI. LC-ESI analysis was performed on Quattro Ultima® mass spectrometer
(Micromass,UK). The source temperature was maintained at 100 ºC, and the desolvation
temperature at 200 °C. The drying gas (N2) was maintained at ca. 450 L/Hr, and the cone
flow gas at ca. 50 L/Hr. The multiplier was set at an absolute value of 500. MS scans at
m/z corresponding to selected singly charged molecular ions were obtained for the
adducts. Optimized parameters can be found in Table 3.3.
114

3.6. References.
(1) Hazen, S. L.; Chisolm, G. M. Proc Natl Acad Sci U S A 2002, 99, 12515-7. (2) Hrbac, J.; Kohen, R. Drug Dev Res 2000, 50, 516 - 527. (3) Grandjean, E. M.; Berthet, P.; Ruffmann, R.; Leuenberger, P. Clin Ther 2000, 22,
209-21. (4) Boon, P. J. M.; Marinho, H. S.; Oosting, R.; Mulder, G. J. Toxicol Appl
Pharmacol 1999, 159, 214-223. (5) Rodriguez-Martinez, E.; Rugerio-Vargas, C.; Rodriguez, A. I.; Borgonio-Perez,
G.; Rivas-Arancibia, S. Int J Neurosci 2004, 114, 1133-45. (6) Kondo, Y.; Murakami, S.; Oda, H.; Nagate, T. Adv Exp Med Biol 2000, 483, 193-
202. (7) De Mattia, G.; Bravi, M. C.; Laurenti, O.; Cassone-Faldetta, M.; Proietti, A.; De
Luca, O.; Armiento, A.; Ferri, C. Diabetologia 1998, 41, 1392-6. (8) Wagberg, M.; Jansson, A. H.; Westerlund, C.; Ostlund-Lindqvist, A. M.;
Sarnstrand, B.; Bergstrand, H.; Pettersson, K. J Pharmacol Exp Ther 2001, 299, 76-82.
(9) Marnett, L. J.; Hurd, H. K.; Hollstein, M. C.; Levin, D. E.; Esterbauer, H.; Ames, B. N. Mutat Res 1985, 148, 25-34.
(10) Stadtman, E. R. Science 1992, 257, 1220-4. (11) Fraga, C. G.; Shigenaga, M. K.; Park, J. W.; Degan, P.; Ames, B. N. Proc Natl
Acad Sci U S A 1990, 87, 4533-7. (12) Chakraborti, T.; Ghosh, S. K.; Michael, J. R.; Batabyal, S. K.; Chakraborti, S. Mol
Cell Biochem 1998, 187, 1-10. (13) Tapiero, H.; Ba, G. N.; Couvreur, P.; Tew, K. D. Biomed Pharmacother 2002, 56,
215-22. (14) Salomon, R. G. Antioxid Redox Signal 2005, 7, 185-201. (15) Arlt, S.; Kontush, A.; Muller-Thomsen, T.; Beisiegel, U. Z Gerontol Geriatr
2001, 34, 461-5. (16) Kopitz, J.; Holz, F. G.; Kaemmerer, E.; Schutt, F. Biochimie 2004, 86, 825-31. (17) Crabb, J. W.; Miyagi, M.; Gu, X.; Shadrach, K.; West, K. A.; Sakaguchi, H.;
Kamei, M.; Hasan, A.; Yan, L.; Rayborn, M. E.; Salomon, R. G.; Hollyfield, J. G. Proc Natl Acad Sci U S A 2002, 99, 14682-7.
(18) Gu, X.; Meer, S. G.; Miyagi, M.; Rayborn, M. E.; Hollyfield, J. G.; Crabb, J. W.; Salomon, R. G. J Biol Chem 2003, 278, 42027-35.
(19) Gotz, M. E.; Kunig, G.; Riederer, P.; Youdim, M. B. Pharmacol Ther 1994, 63, 37-122.
(20) Sun, A. Y.; Chen, Y. M. J Biomed Sci 1998, 5, 401-14. (21) Mlakar, A.; Spiteller, G. Chem Phys Lipids 1996, 79, 47-53. (22) Harrison, K. A.; Murphy, R. C. J Biol Chem 1995, 270, 17273-8. (23) Poli, G.; Schaur, R. J. IUBMB Life 2000, 50, 315-21. (24) Uchida, K. Prog Lipid Res 2003, 42, 318-43. (25) Salomon, R. G.; Kaur, K.; Podrez, E.; Hoff, H. F.; Krushinsky, A. V.; Sayre, L.
M. Chem Res Toxicol 2000, 13, 557-64. (26) Sayre, L. M.; Zelasko, D. A.; Harris, P. L.; Perry, G.; Salomon, R. G.; Smith, M.
A. J Neurochem 1997, 68, 2092-7.
115

(27) Macdonald, S.; Dowle, M.; Harrison, L.; Clarke, G.; Inglis, G.; Johnson, M.; Shah, P.; Smith, R.; Amour, A.; Fleetwood, G.; Humphreys, D.; Molloy, C.; Dixon, M.; Godward, R.; Wonacott, A.; Singh, O.; Hodgson, S.; Hardy, G. J Med Chem 2002, 45, 3878-90.
(28) Szweda, L. I.; Uchida, K.; Tsai, L.; Stadtman, E. R. J Biol Chem 1993, 268, 3342-7.
(29) Crabb, J. W.; O'Neil, J.; Miyagi, M.; West, K.; Hoff, H. F. Protein Sci 2002, 11, 831-40.
(30) Sayre, L. M.; Sha, W.; Xu, G.; Kaur, K.; Nadkarni, D.; Subbanagounder, G.; Salomon, R. G. Chem Res Toxicol 1996, 9, 1194-201.
(31) Xu, G.; Liu, Y.; Sayre, L. M. Chem Res Toxicol 2000, 13, 406-13. (32) Sayre, L. M.; Arora, P. K.; Iyer, R. S.; Salomon, R. G. Chem Res Toxicol 1993, 6,
19-22. (33) Nadkarni, D. V.; Sayre, L. M. Chem Res Toxicol 1995, 8, 284-91. (34) Liu, Y.; Sun, G.; David, A.; Sayre, L. M. Chem Res Toxicol 2004, 17, 110-8. (35) Jurgens, G.; Lang, J.; Esterbauer, H. Biochim Biophys Acta 1986, 875, 103-14. (36) Deng, Y. Ph.D, Case Western Reserve University, 2000. (37) van Kuijk, F. J.; Siakotos, A. N.; Fong, L. G.; Stephens, R. J.; Thomas, D. W.
Anal Biochem 1995, 224, 420-4. (38) Amarnath, V.; Valentine, W. M.; Montine, T. J.; Patterson, W. H.; Amarnath, K.;
Bassett, C. N.; Graham, D. G. Chem Res Toxicol 1998, 11, 317-328. (39) Matsushita, Y.-i.; Sugamoto, K.; Matsui, T. Tennen Yuki Kagobutsu Toronkai
Koen Yoshishu 1998, 40th, 613-618. (40) Sugamoto, K.; Matsushita, Y.-i.; Matsui, T. J Chem Soc, Perkin Trans 1: Org
Bio-Org Chem 1998, 3989-3998. (41) Selley, M. L.; Bartlett, M. R.; McGuiness, J. A.; Hapel, A. J.; Ardlie, N. G. J
Chromatogr 1989, 488, 329-40. (42) Gioacchini, A. M.; Calonghi, N.; Boga, C.; Cappadone, C.; Masotti, L.; Roda, A.;
Traldi, P. Rapid Commun Mass Spectrom 1999, 13, 1573-9. (43) Kingston, D. Chem Rev 1974, 74, 215. (44) Gross, J. H. Mass Spectrometry- A Textbook; Springer-Verlag: Heidelberg, 2004. (45) Edwards, G. L.; Muldoon, C. A.; Sinclair, D. J. Tetrahedron 1996, 52, 7779-7788. (46) Felder, S.; Rowan, D. D. J Labelled Compd Radiopharm 1999, 42, 83 - 92. (47) Deng, Y.; Salomon, R. G. J Org Chem 1998, 63, 3504-3507. (48) SDBSWeb : http://www.aist.go.jp/RIODB/SDBS/ (National Institute of
Advanced Industrial Science and Technology; Vol. 2005. (49) Enoiu, M.; Herber, R.; Wennig, R.; Marson, C.; Bodaud, H.; Leroy, P.; Mitrea,
N.; Siest, G.; Wellman, M. Arch Biochem Biophys 2002, 397, 18-27. (50) Aldini, G.; Granata, P.; Carini, M. J Mass Spectrom 2002, 37, 1219-28. (51) Xu, G.; Sayre, L. M. Chem Res Toxicol 1998, 11, 247-51. (52) Steen, H.; Mann, M. J Am Soc Mass Spectrom 2001, 12, 228-32. (53) Doreleijers, J. F., Universiteit Utrecht, 1999. (54) Rathahao, E.; Peiro, G.; Martins, N.; Alary, J.; Gueraud, F.; Debrauwer, L. Anal
Bioanal Chem 2005, 381, 1532-9. (55) Liu, Y.; Jinno, H.; Kurihara, M.; Miyata, N.; Toyo'oka, T. Biomed chromatogr
BMC. 1999, 13, 75-80.
116

(56) Isom, A. L.; Barnes, S.; Wilson, L.; Kirk, M.; Coward, L.; Darley-Usmar, V. J Am Soc Mass Spectrom 2004, 15, 1136-47.
(57) Doorn, J. A.; Petersen, D. R. Chem Biol Interact 2003, 143-144, 93-100. (58) Kates, M. Techniques of lipidology : isolation, analysis, and identification of
lipids; 2nd rev. ed. ed.; Elsevier: Amsterdam,The Netherlands, 1986. (59) Birch, A. J.; Walker, K. A. M. J. Chem. Soc. (C) 1966, 1894. (60) Deng, Y.; Salomon, R. G. J Org Chem 2002, 63, 3504-07. (61) Esterbauer, H.; Schaur, R. J.; Zollner, H. Free Radic Biol Med 1991, 11, 81-128.
117

Chapter 4
Oxidative Protein Modifications in the Pathogenesis of Primary Open Angle
Glaucoma
118

4.1. Background
Free radical induced oxidative damage of polyunsaturated fatty acids generates
highly reactive molecules, which may have deleterious effects on cellular processes by
modifying protein and DNA.1-3 Additionally, these reactive molecules may act as ligands
for receptors 4,5 or they can form adducts on the cellular macromolecules that may be
immunogenic.6,7 Since these modifications are involved in a large variety of disease
processes, a method for early detection or prevention of such damage is important. Our
group has been involved in identification and characterization of the products of lipid
oxidation and their protein adducts.7,8 Antibodies generated against these modifications
and their subsequent use in their immunodetection in vivo were instrumental in
establishing a correlation between their levels in normal and diseased states viz,
atherosclerosis, renal disease6 and age related macular degeneration (AMD).9
4.1.1. Oxidative stress and glaucoma. The lipids present in the eye are prone to light
induced oxidative damage owing to the presence of light in conjunction with high oxygen
concentration.10,11 The products of docosahexaenoate (an ω-3 fatty acid) phospholipid
oxidation have been implicated in AMD.12 Unlike these phospholipids that are present in
high levels only in the posterior region of the eye, the ω-6 fatty acids (arachidonyl and
linolenyl phospholipids) are present in the anterior region, and are implicated in the
pathogenesis of glaucoma.13
Glaucoma, a group of late onset and progressive eye diseases that lead to
irreversible blindness often without any initial symptoms, affects about 70 million people
worldwide.14 The glaucomas are classified as primary, when no known contributory
119

factor is linked to the disease, and as secondary, when a previous disease or injury could
be attributed as a cause. Further, they can be classified as open angle glaucoma, when the
angle between the cornea and iris is normal or open for free flow of aqueous humor and
as angle closure glaucoma, when the angle is more acute than the normal (Figure 4.1).
The angle closure form glaucoma is due to an anatomical condition. However, the open
angle form of glaucoma remains poorly understood. Primary open angle glaucoma
(POAG) occurs most commonly but not always with an increase in the intraocular
pressure (IOP), and leads to progressive optic nerve damage.15
Figure 4.1: Diagrammatic representation of the cross section of an eye showing the two
forms of glaucoma defined according to the difference in the angle of the anterior
chamber formed by lines drawn parallel to the iris and cornea (iridial angle). A. normal
open angle (~40o) associated with POAG and B. a closed angle (~15o) associated with
angle closure glaucoma.
120

The aqueous humor, actively secreted by the ciliary epithelium, flows through the
pupil into the anterior segment of the eye and then passes through the trabecular
meshwork (TM), and drains into the Schlemm’s canal (Figure 4.2). The amount of
aqueous humor flowing into the anterior chamber in comparison with the amount drained
out is important in maintaining the IOP. Also, the TM is the site of maximum resistance
to the flow of aqueous humor, and any change in the TM can adversely affect the
aqueous flow, thus changing the IOP.15
Figure 4.2: Diagrammatic representation of the cross section of the human eye showing
the aqueous humor outflow pathway (Illustration by N. Aravindhan © 2005).
Tissues in the anterior chamber are subjected to chronic oxidative stress due to the
formation of reactive oxygen species (ROS),16,17 and increased ROS may be responsible
for damage to the trabecular meshwork and the Schlemm’s canal.18,19 POAG is
considered as a multifactorial disease and the role of oxidative damage in causing
changes in the trabecular meshwork and blockage of aqueous outflow cannot be
121

ignored.17,20-22 One of the factors contributing to increased oxidative stress may be a
decrease in the “antioxidant potential” of the TM, e.g., owing to inactivation of an
enzyme involved in detoxifying peroxides17,21 or decrease in levels of antioxidants.23-25
Furthermore, obstruction in the flow has also been attributed to the aggregation of
proteins under oxidative stress conditions.26
4.1.2. Levuglandins and isolevuglandins: formation and pathobiology. Levuglandins
(LGs) and isolevuglandins (isoLGs) are arachidonyl phospholipid (AA-PC) ester
metabolites that are formed by cyclooxygenase (COX) or free radical mediated pathways
respectively (Scheme 4.1, next page).27,28 The substrate for COX mediated
peroxidation requires the presence of free arachidonic acid, while the free radical
mediated pathway occurs predominantly in the phospholipid ester form.29 As
depicted in Scheme 4.1, LGE2 can be formed by either of these pathways, while
iso[4]LGE2 can only be formed through the free radical mediated pathway. These
γ-ketoaldehydes are highly reactive and are known to form adducts with lysine residues
followed by dehydration to form pyrroles (Scheme 4.1). LGs have also been shown to
form protein cross links rapidly compared to other lipid peroxidation products viz, 4-
hydroxynonenal (HNE) and malondialdehyde (MDA).30 LG modified LDL was shown
to be recognized and processed by macrophages through a scavenger receptor that
recognizes oxidized low density lipoproteins (oxLDL).31 The LGs and isoLGs-derived
protein adducts as well as free LGs have been shown to hinder the activity of the 20S
proteasome, a component of the protein degradation machinery, in a dose dependent
manner.32 Iso[4]LGE2-protein adducts are generated in blood and vasculature 33
122

AA
(CH2)3COOR
C5H11
OH
OHC
(CH2)3COOR
C5H11
OH
O
OO
O
OO C (CH2)nCH3
OP
ON(CH3)3
O OC5H11
(CH2)3COOR
OH
OO
(CH2)3COOR
OHOHC
O
C5H11
C5H11
(CH2)3COOH
C5H11
OH
N
H3C(CH2)3COOH
OH
C5H11N
H3C
(CH2)3COOR
C5H11
(CH2)3COOR
C5H11
O
(CH2)3COOH
C5H11
OH
OHC
(CH2)3COOH
C5H11
OH
O
OO
(CH2)3COOH
C5H11
NH2
2O2
protein
iso[4]PGH2-PC
protein
12
isoLGE2
AA-PC
iso[4]LGE2
iso[4]LGE2-pyrrole
isoPGH2-PC
LGE2-pyrrole
-2 H2O
-2 H2O
8
13
8
7
710
13
10
7
13
10
2O2
9
1 15
6912
LGE2
PGH2
8
8
7
13
10
2O2
9
1
-2 H2O
PLA2
cyclooxygenase
free radicals
-2-lysoPC-2-lysoPCprotein NH2protein
NH2protein
Scheme 4.1: Enzymatic (cyclooxygenase) and non enzymatic (free radical) routes for the
formation of LGE2 and iso[4]LGE2 and their protein adducts.
and are present in oxidized LDL.34 The presence of these adducts was detected with
antibodies raised against these epitopes using immunodetection techniques viz, ELISA,
Western blots and immunohistochemical analysis.33,34 The detection of isoLGs is
probably indicative of localized oxidative stress within tissues as they form adducts
123

rapidly.30 Accumulation of modified proteins has been identified in various pathological
conditions such as muscular dystrophy,35 rheumatoid arthritis36 and atherosclerosis.37
4.1.3. 4-Hydroxynonenal and its role in pathobiology. HNE is by far the most
investigated of the lipid peroxidation products.38-40 HNE is generated from AA or linoleyl
phospholipids through free radical mediated pathways. Because HNE is a small
bifunctional molecule, it easily transverses the cell membrane and reacts with a wide
variety of substrates.39 HNE is known to form Michael adducts with nucleophilic amino
acid residues viz, lysine, cysteine and histidine (Scheme 4.2).41
OOH
4-Hydroxynonenal
N
OOH
NH
OOH
N
Lysine Michael adduct
Histidine Michael adduct
Histidine
Lysine
S
OOH
Cysteine Michael adduct
Cysteine
OHN
Lysine
NLysine
LysineN
HO
LysineN
Lysine-Lysine crosslink
Lysine Schiff base adduct
Lysine pyrrole adduct
Scheme 4.2: Some of the commonly reported adducts formed by 4-hydroxynonenal
reaction with histidine, lysine and cysteine residues.
124

The presence of three functional groups, viz. aldehyde, C=C double bond and a
hydroxyl group, predisposes HNE to react with more than one residue.42 Consequently,
HNE formation of protein-protein crosslinks has been observed in both in vitro as well as
in vivo model studies.43,44 The HNE derived modifications on proteins have been
implicated in a variety of disease processes including atherosclerosis,45
neurodegeneration46 and proliferative vitreoretinopathies.47 See section 2.1 for a detailed
background.
4.1.4. Advanced glycation end products and their pathobiology. Modification of
proteins by the Maillard reaction is thought to play a role in chemical aging of
proteins.48,49 In the Maillard reaction, the amino side chain residues of a protein are non-
enzymatically glycosylated by reducing sugars to form aminoketoses, also called as
Amadori products.50 This initial adduct can undergo rearrangements, cyclizations and
dehydrations to form products, mostly with fluorescent and chromophoric properties,
collectively called advanced glycation end products (AGEs). At present, there is evidence
that protein glycation is involved in the pathogenesis of several amyloid diseases, such as
Alzheimer's disease51-53 and dialysis-related amyloidosis.54 Some of the cellular effects of
AGEs are mediated by interactions with specific cellular receptors, such as the receptor
for AGEs (RAGE).55
In addition to glucose, methylglyoxal (MG) a product formed by non-enzymatic,
amine catalyzed sugar fragmentation reactions and by spontaneous decomposition of
triose phosphate intermediates of glycolysis, can also modify proteins and contribute to
AGE’s (Scheme 4.3).56 MG may also be formed from lipid oxidation products.57 MG is
125

one of the most powerful glycating reagents that can irreversibly modify lysine and
arginine residues in proteins.
NOH
HOOH
Protein
OHHOH2C
Protein-NH2
OH
O O
N N
OH
HN (CH2)3
NH2
COOH
+ Glucose
Arginine
Argpyrimidine
H
O
O
OH
O
OH
H+
HCOOH
HNOH
HOOH
Protein
OHHOH2C
HNO
HOOH
Protein
OHHOH2C
"Classical"Amadori
rearrangement
O
OHNH
OH
OH
OH
Methylglyoxal
Protein
Intermediates of Glycolysis
Amadori product
Scheme 4.3: Mechanistic pathway for the formation of AGE’s and argpyrimidine.
MG is known to form carboxyethyl lysine (CEL) by adduction with lysine
residues and argpyrimidine [Nd-(5-hydroxy-4,6-dimethylpyrimidin-2-yl)-L-ornithine], a
fluorescent product, by adduction with arginine.56 Recently, argpyrimidine was found in
spinal cord of familial sporadic amyotrophic lateral sclerosis (ALS) patients and mutant
126

SOD-1 (superoxide dismutase 1) mice. Methylglyoxal modification of arginine may
contribute to the pathophysiology associated with aging and other diseases.57,58
4.1.5. Oxidative products of tryptophan and α-hydroxykynurenine. Oxidative
damage of tryptophan by both enzymatic and non-enzymatic pathways generates
products that can modify proteins to form yellow-brown pigments.59 The metabolism of
tryptophan proceeds through the kynurenine pathway.60 The products of this pathway as
well as toxic intermediates are implicated in a range of neurodegenerative, inflammatory,
and immunologic phenomena diseases/disorders, including Huntington’s disease,61
Parkinson’s disease,62 and HIV encephalopathy.63
As outlined in Scheme 4.4, the products formed from the N-formyl kynurenines
include kynurenine (KYN), 3-hydroxykynurenine (OHKYN), 3-hydroxyanthranilic acid,
quinolinic acid, and nicotinic acid.60 OHKYN can form various reactive catabolites,
which can modify proteins and cause them to aggregate.64 One of the products of the
reaction of OHKYN with lens glutathione (GSH) is glutathionyl-3-hydroxykynurenine
glucoside,65 which accumulates during lens aging and accumulates to relatively high
levels in cataractous lenses.66 In addition to the modifications formed by kynurenine and
its auto oxidation products, it can also form adducts by non-oxidative pathways, e.g., α,
β- unsaturated OHKN, 67 formed by deamination of KYN, reacts with nucleophilic amino
acids, such as cysteine, histidine, and lysine in lens proteins.68
Recently, antibodies against the protein modifications formed by OHKYN were
generated and characterized.59 These antibodies were used in this study to assess the
levels of these modifications in TM tissue of POAG subjects.
127

NH2
HOO
NH
NH2
HOO
ONH
O
NH2
OHO
ONH2
NH2
OHO
ONH2HO
O2
Kynurenineformidase
Kynureninehydroxylase
INF-γIndoleamine
2,3-dioxygenase
Proteins
α−Hydroxykynurenine
Protein Adducts
TryptophanN-formyl Kynurenine
Kynurenine
Scheme 4.4: Catabolic pathway of tryptophan under oxidative conditions.
Accumulation of oxidative protein modifications in TM from lipids and sugars69
may contribute to the onset and/or progression of glaucoma. However, their occurrence
in TM tissue in vivo in glaucoma has yet to be demonstrated. In this chapter, a
multifaceted approach was used to assess the levels of oxidative injury/insult in
glaucomatous TM compared to that of a control TM by using antibodies against protein
modifications derived from oxidized lipids, glucose and tryptophan.
128

4.2. Results.
4.2.1. Immunoblot analysis of TM tissues for analyzing the levels of oxidative
protein modifications in glaucomatous TM compared to the controls. To compare the
levels of protein modifications in POAG and normal TM, a western blot analysis of TM
extracts of POAG and age-matched controls was performed. The Western blot analysis
using the anti-iso[4]LGE2 (Figures 4.3 and 4.1S (appendix)), HNE (Figures 4.4 and 4.2S
(appendix)), argpyrimidine (Figures 4.4 and 4.3S (appendix)) and OHKYN antibodies
showed increased immunoreactivity in protein extracts of the POAG TM compared to
that of the control TM extracts. Quantification of this difference is presented in Figures
4.11 and 4.12, see discussion in section 4.3 for details. Totally, 52 POAG samples were
compared with 52 age-matched control samples (Table 4.1). Bovine or human serum
albumin modified by these oxidized lipids was used as a positive control.
Table 4.1: Total number of tissue samples used for Western blots for each of the
antibodies probed. Some of the tissues were used in more than one Western blot analysis.
Number of POAG samples analyzed
Number of Control samples analyzed
Immunoreactivity* Immunoreactivity*
Western Blots
Total + - Total + -
Iso[4]LGE2 23 1 22 22 21 1
HNE 23 2 21 22 12 10
Argpyrimidine 16 2 14 16 16 9
OHKYN 8 1 7 8 4 4
*Immunoreactivity (+) = values above 1 standard deviation (SD) and below mean SD of POAG
*Immunoreactivity (-) = values below 1 SD and below mean SD of POAG
129

88M
W
74 M
W
87 M
W
84 M
W
80 F
W
72 M
W
87 M
W
87 M
W
LMW
c
11488
50
35
28
20
11488
50
35
28
20
Normal TM Glaucomatous TM
D
74 M
W
6 1 F
W
74 M
W
7 0 M
W
78 M
W
64 F
W
7 6 M
W
69 M
W
Normal TM Glaucomatous TM
B
E
42 F
B
44 M
B
74 F
B
79 F
B
42 F
B
42 M
B
74 F
B
81 F
B
Normal TM Glaucomatous TM
C
F
Coo
mas
sie
Wes
tern
Blo
t
A
88M
W
74 M
W
87 M
W
84 M
W
80 F
W
72 M
W
87 M
W
87 M
W
LMW
c
11488
50
35
28
20
11488
50
35
28
20
11488
50
35
28
20
Normal TM Glaucomatous TM
D
74 M
W
6 1 F
W
74 M
W
7 0 M
W
78 M
W
64 F
W
7 6 M
W
69 M
W
Normal TM Glaucomatous TM
B
E
42 F
B
44 M
B
74 F
B
79 F
B
42 F
B
42 M
B
74 F
B
81 F
B
Normal TM Glaucomatous TM
C
F
Coo
mas
sie
Wes
tern
Blo
t
A
Figure 4.3: Western analysis of POAG and normal trabecular meshwork using anti-
iso[4]LGE2 pAb antibodies. TM extracts (5 µg) were subjected to SDS-PAGE, blotted to
PVDF membrane and probed with anti-iso[4]LGE2 antibodies. A,B,C . Coomassie blue
stained gels. D,E,F. Western blot: age, race and gender of the tissue samples are
indicated (M-Male; F-Female; W-Caucasian; B-African American).
6645
2631
14
97
6645
2631
14
97
74 M
W
77 M
W
80 F
W
87 M
W
74 M
W
78M
W
80 F
W
86 M
W
77 M
W
60 F
W
80 F
W
74 M
W
76 M
W
75 F
W
94 F
W
77 M
B
43 M
W
50 M
W
58 M
W
72 M
W
72 M
W
58 F
A
50 M
W
43 M
W
Normal TM Glaucomatous TMNormal TM Glaucomatous TM Normal TM Glaucomatous TM
Coo
mas
sie
Wes
tern
Blo
t
D
B
E
C
F
A
6645
2631
14
97
6645
2631
14
97
74 M
W
77 M
W
80 F
W
87 M
W
74 M
W
78M
W
80 F
W
86 M
W
77 M
W
60 F
W
80 F
W
74 M
W
76 M
W
75 F
W
94 F
W
77 M
B
43 M
W
50 M
W
58 M
W
72 M
W
72 M
W
58 F
A
50 M
W
43 M
W
Normal TM Glaucomatous TMNormal TM Glaucomatous TM Normal TM Glaucomatous TM
Coo
mas
sie
Wes
tern
Blo
t
6645
2631
14
976645
2631
14
97
6645
2631
14
976645
2631
14
97
74 M
W
77 M
W
80 F
W
87 M
W
74 M
W
78M
W
80 F
W
86 M
W
77 M
W
60 F
W
80 F
W
74 M
W
76 M
W
75 F
W
94 F
W
77 M
B
43 M
W
50 M
W
58 M
W
72 M
W
72 M
W
58 F
A
50 M
W
43 M
W
74 M
W
77 M
W
80 F
W
87 M
W
74 M
W
78M
W
80 F
W
86 M
W
77 M
W
60 F
W
80 F
W
74 M
W
76 M
W
75 F
W
94 F
W
77 M
B
43 M
W
50 M
W
58 M
W
72 M
W
72 M
W
58 F
A
50 M
W
43 M
W
Normal TM Glaucomatous TMNormal TM Glaucomatous TM Normal TM Glaucomatous TM
Coo
mas
sie
Wes
tern
Blo
t
D
B
E
C
F
A
Figure 4.4: Western analysis of POAG and normal trabecular meshwork using anti-HNE
pAb antibodies. TM extracts (5 µg) were subjected to SDS-PAGE, blotted to PVDF
membrane and probed with anti- anti-HNE antibodies. A,B,C. Coomassie blue stained
gels. D,E,F. Western blot. M-Male; F-Female; W-Caucasian; B-African American).
130

Normal TM
250150
75
3750
25
100
20
1015
7 4 M
W
61 F
W
7 4 M
W
7 0 M
W
78 M
W
6 4 F
W
7 6 M
W
69 M
W
250150
75
37
50
25
100
20
1015
Glaucomatous TM Normal TM
85 F
W
59 F
W
6 7 F
W
6 1 M
W
81 F
W
5 9 F
W
6 7 F
W
61 M
W
Glaucomatous TM Normal TM
82 F
W
5 9 F
W
8 6 F
W
61 M
W
82 F
W
59 F
W
8 6 F
W
61 M
W
Glaucomatous TM
250150100
Coo
mas
sie
Wes
tern
Blo
t
D
B
E
C
F
A
Normal TM
250150
75
3750
25
100
20
1015
7 4 M
W
61 F
W
7 4 M
W
7 0 M
W
78 M
W
6 4 F
W
7 6 M
W
69 M
W
250150
75
37
50
25
100
20
1015
Glaucomatous TM Normal TM
85 F
W
59 F
W
6 7 F
W
6 1 M
W
81 F
W
5 9 F
W
6 7 F
W
61 M
W
Glaucomatous TM Normal TM
82 F
W
5 9 F
W
8 6 F
W
61 M
W
82 F
W
59 F
W
8 6 F
W
61 M
W
Glaucomatous TM
250150100
Coo
mas
sie
Wes
tern
Blo
t
D
B
E
C
F
A
Figure 4.5: Western analysis of POAG and normal trabecular Meshwork using anti-
argpyrimidine mAb. TM extracts (5 µg) were subjected to SDS-PAGE, blotted to PVDF
membrane and probed with anti-argpyrimidine antibodies. A,B,C. Coomassie blue
stained gels. D,E,F. Western blot: age, race and gender of the tissue samples are
indicated (M-Male; F-Female; W-Caucasian; B-African American).
Coo
mas
sie
Wes
tern
Blo
t
11488
50
35
28
20
Normal TM
85 F
W
5 9 F
W
67 F
W
61 M
W
81 F
W
5 9 F
W
6 7 F
W
61 M
W
Glaucomatous TM
11488
50
35
28
20
Normal TM Glaucomatous TM
82 F
W
59 F
W
8 6 F
W
61 M
W
82 F
W
59 F
W
8 6 F
W
61 M
W
Coo
mas
sie
Wes
tern
Blo
t
11488
50
35
28
20
11488
50
35
28
20
Normal TM
85 F
W
5 9 F
W
67 F
W
61 M
W
81 F
W
5 9 F
W
6 7 F
W
61 M
W
Glaucomatous TM
11488
50
35
28
20
11488
50
35
28
20
Normal TM Glaucomatous TM
82 F
W
59 F
W
8 6 F
W
61 M
W
82 F
W
59 F
W
8 6 F
W
61 M
W
Figure 4.6: Western Analysis of POAG and Normal Trabecular Meshwork using anti-
OHKYN mAb. TM extracts (5 µg) were subjected to SDS-PAGE, blotted to PVDF
membrane and probed with anti-OHKYN antibodies. Age, race and gender of the tissue
samples are indicated (M-Male; F-Female; W-Caucasian; B-African American).
131

4.2.2. Localization of modified proteins by immunohistochemical analysis. To
visualize the localization of modified proteins in the trabecular meshwork, we probed the
tissue sections from POAG and age-matched cadaver tissue with antibodies to these
modifications. Anti-iso[4]LGE2 and anti-HNE antibodies (red fluorescence of the
secondary antibody with Alexa 594 fluorescent tag) showed a strong immunoreactivity in
the TM of POAG compared to the normal TM or the pre-immune stained POAG TM
sections. Adjacent sections were used for immunohistochemical analysis using the
antibody and corresponding pre-immune serum to compare the relative
immunoreactivity.
Figure 4.7: Localization of iso[4]LGE2 modified proteins in glaucomatous TM. Anterior
segment histochemical sections through the trabecular meshwork are shown. A, B, C –
Probed with pre-immune serum; D, E, F,G, H, I - Probed with anti-iso[4]LGE2 antibody;
Red Channel – iso[4]LGE2 specific immunofluorescence ; Green Channel –
autofluorescence ; DAPI - nuclear stain (blue fluorescence)
132

A
B
C
D
E
G
H
I
Glaucomatous TM Glaucomatous TM Normal TM
DAP
IG
reen
Cha
nnel
Red
Cha
nnel
F
A
B
C
D
E
G
H
I
Glaucomatous TM Glaucomatous TM Normal TM
DAP
IG
reen
Cha
nnel
Red
Cha
nnel
F
Figure 4.8: Localization of HNE modified proteins in glaucomatous TM. Anterior
segment histochemical sections through the trabecular meshwork are shown. A, B, C –
Probed with pre-immune serum; D, E, F,G, H, I - Probed with anti-HNE antibody; Red
Channel – HNE specific immuno-fluorescence; Green Channel – auto-fluorescence ;
DAPI – nuclear stain (blue fluorescence)
4.2.3. Immunoprecipitation of modified proteins. To identify the modified proteins
from the TM extracts, we immunoprecipitated the iso[4]LGE2 and HNE modified
proteins using the corresponding antibody-coupled protein A sepharose beads. The
immunoprecipitated proteins were subjected to SDS-PAGE and partially electroblotted
onto a PVDF membrane. The Coomassie stained IP gels clearly show the enrichment of
modified proteins compared to the total protein extracts (Figures 4.9 and 4.3S
(Appendix)). The immunoblot of the IP exhibited protein bands corresponding to those in
133

the total extract (Figure 4.9 and 4.4S (Appendix)). The bands detected at ~55 kDa that is
common to both the lane 2 and 3 (shown with dotted lines in Figure 4.9A & 4.9B)
corresponds to the heavy chain of IgG originating from the cleavage of IgG chains from
the antibody-coupled beads because the extraction was done with Laemmli’s buffers that
contains DTT as reported previously.70 DTT cleaves the disulphide bond that anchor the
IgG heavy chain to the light chain.
114
88
50
35
28
20
250
1 2 3 4 5A
345
2
1
1 2 3 4 5
114
88
50
35
28
20
250
B
IgG(long chain)
12
456
3
114
88
50
35
28
20
250
1 2 3 4 5A
345
2
1
114
88
50
35
28
20
250
1 2 3 4 5A
345
2
1
1 2 3 4 5
114
88
50
35
28
20
250
1 2 3 4 5
114
88
50
35
28
20
250
114
88
50
35
28
20
250
B
IgG(long chain)
12
456
3
Figure 4.9: Immunoprecipitation of antibody modified protein from TM extract. 10µg of
antibody coupled beads and 30µg of TM protein extract were used for
immunoprecipitations. A. Anti-iso[4]LGE2 IP B. Anti-HNE IP .Lane 1 – Protein extract,
Lane 2 – IP products, Lane 3 – Ab coupled beads, Lane 4 – control-protein A beads
without antibody, Lane 5 – Western blot of protein extract. Bands numbered here
corresponds to proteins identified and listed in Table 4.2. Detection was performed by
Coomassie blue staining. Preimmune probed IP is presented in the Appendix Figure 4.4S.
134

4.2.4. Identification of immunoprecipitated proteins. The Coomassie stained 1D gel
bands of the immunoprecipitated proteins in Figure 4.9 were excised and digested in situ
with trypsin. The tryptic peptides were analyzed by liquid chromatography-coupled
electrospray tandem mass spectrometry 71 and the proteins were identified by well
established bioinformatics methods. Two independent immunoprecipitations from two
different POAG TM samples were performed and proteins that are common in both the
samples are listed in Table 4.2. Serum albumin, transforming growth factor β-IGH3,
decorin and HSP-70 are identified to be immunoprecipitated by both anti-Iso[4]LGE2 as
well as anti-HNE antibody. Lumican, prolargin, PDI-A3, pyruvate kinase and vitronectin
were uniquely immunoprecipitated by HNE antibody-coupled beads, while complement -
C3, and GFAP were immunoprecipitated by iso[4]LGE2 antibody-coupled beads.
135

Table 4.2: Identification of TM proteins immunoprecipitated using iso[4]LGE2 and HNE
pAbs. A. Proteins identified by in-gel digestion of bands from lane A-2 (Figure 4.9) using
mass spectrometry (LC-MS/MS). B. Proteins identified by in-gel digestion of bands
from lane B-2 (Figure 4.9) using mass spectrometry (LC-MS/MS). Proteins listed here
were present in three IP experiments.
27
40
42
187
70
74
69
Calculated* MW (kDa)
2,5
1,3
3,5
2
4,5
3,5
3,4,5
BandNo.
70,3301P07585Decorin
ObservedMW (kDa)
Peptide matche
s
Accession NumberProteins Identified
65-701P48677Glial fibrillary acidic protein
70, 2651P48741Heat shock 70 kDa protein 7
2651P01024Complement C3 precursor
65-702P08107Heat shock 70 kDa protein 1
65-703Q15582TGF-β induced protein IG-H3
65-707P02768Serum albumin precursor
27
40
42
187
70
74
69
Calculated* MW (kDa)
2,5
1,3
3,5
2
4,5
3,5
3,4,5
BandNo.
70,3301P07585Decorin
ObservedMW (kDa)
Peptide matche
s
Accession NumberProteins Identified
65-701P48677Glial fibrillary acidic protein
70, 2651P48741Heat shock 70 kDa protein 7
2651P01024Complement C3 precursor
65-702P08107Heat shock 70 kDa protein 1
65-703Q15582TGF-β induced protein IG-H3
65-707P02768Serum albumin precursor
54
75
69
24
58
57
44
38
40
50
Calculated*MW (kDa)
6
3,6
3,5,6
6
2
6
3,6
6
1,4
4,6
BandNo.
63, >2002Q15582TGF-β induced protein IG-H3
ObservedMW (kDa)
Peptide matches
Accession NumberProteins Identified
631Q8TZS0RNase P component 3
63-2001P49822Serum albumin
631P30101Protein disulphide-isomeraseA3
631P04004Vitronectin precursor
2431P14618Pyruvate kinase isozymes
63, >2003P51888Prolargin
632P51884Lumican
67, >2981P07585Decorin
67, 632P30838Aldehyde dehydrogenase
54
75
69
24
58
57
44
38
40
50
Calculated*MW (kDa)
6
3,6
3,5,6
6
2
6
3,6
6
1,4
4,6
BandNo.
63, >2002Q15582TGF-β induced protein IG-H3
ObservedMW (kDa)
Peptide matches
Accession NumberProteins Identified
631Q8TZS0RNase P component 3
63-2001P49822Serum albumin
631P30101Protein disulphide-isomeraseA3
631P04004Vitronectin precursor
2431P14618Pyruvate kinase isozymes
63, >2003P51888Prolargin
632P51884Lumican
67, >2981P07585Decorin
67, 632P30838Aldehyde dehydrogenase
A
B
* for monomeric protein
4.2.6. Identification of modified proteins by two-dimensional gel electrophoresis.
We also identified the iso[4]LGE2- and HNE-modified proteins from glaucomatous
trabecular meshwork by mass spectrometry following two-dimensional immunoblot
analysis. Trabecular meshwork from a 79-year-old caucasian male with primary open
angle glaucoma was used for the 2D SDS PAGE. The mass spectrometry analysis
136

pH
20
28
35
50
88
114
kDa345678910
12
3
54
7
6
8 9 10
15
11 12 13
14-19
20-2425-27
52
42-46 48 49
51
50
37-41
31-35
28-30
54
47
53
1
36
pH
20
28
35
50
88
114
kDa345678910 345678910
12
3
54
7
6
8 9 10
15
11 12 13
14-19
20-2425-27
52
42-46 48 49
51
50
37-41
31-35
28-30
54
47
53
1
36
pH345678910
pH345678910 345678910
345678910pH
20
28
35
50
88114
kDaHNE
345678910 345678910pH
20
28
35
50
88114
20
28
35
50
88114
kDaHNE
A B
C
pH345678910
pH345678910 3456789 345678910
D
pH
20
28
35
50
88
114
kDa345678910
12
3
54
7
6
8 9 10
15
11 12 13
14-19
20-2425-27
52
42-46 48 49
51
50
37-41
31-35
28-30
54
47
53
1
36
pH
20
28
35
50
88
114
kDa345678910 345678910
12
3
54
7
6
8 9 10
15
11 12 13
14-19
20-2425-27
52
42-46 48 49
51
50
37-41
31-35
28-30
54
47
53
1
36
pH345678910
pH345678910 345678910
345678910pH
20
28
35
50
88114
kDaHNE
345678910 345678910pH
20
28
35
50
88114
20
28
35
50
88114
kDaHNE
A B
C
pH345678910
pH345678910 3456789 345678910
D
Figure 4.10: 2D PAGE analysis of TM extract. ~80 µg of extracted TM proteins was
used for the 2D PAGE. The gel was partially transferred onto a PVDF membrane for
immunochemical analysis. A. Coomassie stained gel B. Western analysis using
Iso[4]LGE2 pAb. C. Western analysis using HNE pAb. D. Merged image showing
Coomassie stain in black and Western blots probed with iso[4]LGE2 in blue; HNE in
orange.
revealed the presence of many small proteins present in higher molecular weight region
suggesting an aggregation and a change in their polarity, In corroboration with the 1D
immunoblots.
137

Table 4.3: Iso[4]LGE2 and HNE immunoreactive proteins identified by 2D PAGE and
mass spectrometry. Calculated MW and pI refers to value derived from Swissprot protein
database; Observed MW and pI refers to values from the 2D PAGE.1031
1 homodimer ; 2 multimer * for monomeric protein except where indicated otherwise 95.6 - 4.667, 505.15327,47Vimentin
24.866,1997.96835Transketolase
23--6.269
2,3,5,9,15,16,19-22,24-31,35,36, 38,52,54
Serum albumin precursor
56.1 - 8.4100, 67, 559.643
7-11,16-24,26, 29,30,34,35,38,54
Prolargin
26.1 - 8.492, 676.5387,26-31,33,39, 41,43,53
Lumican
56.47077420,22Lamin A/C
13.3488.64050Trace amine receptor 15
14.5467.84748beta-enolase
14.75064947alpha-enolase
26.3707.27721Transferrin
18.2525124213Transcription factor 8
25.052 - 7287430,43TGF-bIG H3
36.5557.75817,19Pyruvate kinase
16.7556.75218,19Prolineaminopeptidase
28.4927.2407Keratocan
45 - 75586117,18Glutamate dehydrogenase
17.3978.8378,18Cathepsin K
25 - 750 -536.5,5.26017,46ATP synthase
27 - 85585412,17, 18Annexin A11
56.7557.5,6.75418,19, 52Aldehyde dehydrogenase
19.2535.3103111,36a-actinin
pIMWpIMWPeptide match
ObservedCalculated*Spot No.Protein Identifiedz
95.6 - 4.667, 505.15327,47Vimentin
24.866,1997.96835Transketolase
23--6.269
2,3,5,9,15,16,19-22,24-31,35,36, 38,52,54
Serum albumin precursor
56.1 - 8.4100, 67, 559.643
7-11,16-24,26, 29,30,34,35,38,54
Prolargin
26.1 - 8.492, 676.5387,26-31,33,39, 41,43,53
Lumican
56.47077420,22Lamin A/C
13.3488.64050Trace amine receptor 15
14.5467.84748beta-enolase
14.75064947alpha-enolase
26.3707.27721Transferrin
18.2525124213Transcription factor 8
25.052 - 7287430,43TGF-bIG H3
36.5557.75817,19Pyruvate kinase
16.7556.75218,19Prolineaminopeptidase
28.4927.2407Keratocan
45 - 75586117,18Glutamate dehydrogenase
17.3978.8378,18Cathepsin K
25 - 750 -536.5,5.26017,46ATP synthase
27 - 85585412,17, 18Annexin A11
56.7557.5,6.75418,19, 52Aldehyde dehydrogenase
19.2535.3103111,36a-actinin
pIMWpIMWPeptide match
ObservedSpot No.Protein Identifiedz
A. Iso[4]LGE2 immunoreactive proteins
B. HNEimmunoreactive proteins
C. Iso[4]LGE2 HNEimmunoreactive proteins
95.6 - 4.667, 505.15327,47Vimentin
24.866,1997.96835Transketolase
23--6.269
2,3,5,9,15,16,19-22,24-31,35,36, 38,52,54
Serum albumin precursor
56.1 - 8.4100, 67, 559.643
7-11,16-24,26, 29,30,34,35,38,54
Prolargin
26.1 - 8.492, 676.5387,26-31,33,39, 41,43,53
Lumican
56.47077420,22Lamin A/C
13.3488.64050Trace amine receptor 15
14.5467.84748beta-enolase
14.75064947alpha-enolase
26.3707.27721Transferrin
18.2525124213Transcription factor 8
25.052 - 7287430,43TGF-bIG H3
36.5557.75817,19Pyruvate kinase
16.7556.75218,19Prolineaminopeptidase
28.4927.2407Keratocan
45 - 75586117,18Glutamate dehydrogenase
17.3978.8378,18Cathepsin K
25 - 750 -536.5,5.26017,46ATP synthase
27 - 85585412,17, 18Annexin A11
56.7557.5,6.75418,19, 52Aldehyde dehydrogenase
19.2535.3103111,36a-actinin
pIMWpIMWPeptide match
ObservedCalculated*Spot No.Protein Identifiedz
95.6 - 4.667, 505.15327,47Vimentin
24.866,1997.96835Transketolase
23--6.269
2,3,5,9,15,16,19-22,24-31,35,36, 38,52,54
Serum albumin precursor
56.1 - 8.4100, 67, 559.643
7-11,16-24,26, 29,30,34,35,38,54
Prolargin
26.1 - 8.492, 676.5387,26-31,33,39, 41,43,53
Lumican
56.47077420,22Lamin A/C
13.3488.64050Trace amine receptor 15
14.5467.84748beta-enolase
14.75064947alpha-enolase
26.3707.27721Transferrin
18.2525124213Transcription factor 8
25.052 - 7287430,43TGF-bIG H3
36.5557.75817,19Pyruvate kinase
16.7556.75218,19Prolineaminopeptidase
28.4927.2407Keratocan
45 - 75586117,18Glutamate dehydrogenase
17.3978.8378,18Cathepsin K
25 - 750 -536.5,5.26017,46ATP synthase
27 - 85585412,17, 18Annexin A11
56.7557.5,6.75418,19, 52Aldehyde dehydrogenase
19.2535.3103111,36a-actinin
pIMWpIMWPeptide match
ObservedSpot No.Protein Identifiedz
A. Iso[4]LGE2 immunoreactive proteins
B. HNEimmunoreactive proteins
C. Iso[4]LGE2 HNEimmunoreactive proteins
138

4.3. Discussion Oxidative stress is often the final common pathway of cell damage that has been
implicated in pathogenesis in many neurodegenerative diseases such as Alzheimer’s
disease, Parkinson’s disease and glaucoma.72,73 In the eye, oxidative damage has been
implicated in degenerations of the posterior segment, the retina and the optic nerve.74,75 In
glaucoma the increase in the intraocular pressure may be associated with the oxidative
damage in the TM tissue. Increase in oxidative stress and decrease in the total reactive
antioxidant potential have been detected in aqueous humor of glaucoma patients.76
Oxidative DNA damage is increased in the trabecular meshwork of patients with
glaucoma.77,78 Most of the oxidative protein modifications6,7,12,79,80 have been linked to
tissue damage in aging 11 and may be an important component in onset and progression
of POAG.
The present study provided the first direct evidence establishing a correlation
between oxidative stress in the TM and glaucoma. Western blot analysis of protein
extracts from the TM tissue of glaucoma and control subjects was performed using
multiple antibodies against oxidative stress markers derived from lipid and protein
oxidation as well as AGEs. The proteins that may be targets of increased oxidative stress
were isolated either by immunoprecipitation or by 2D PAGE and subsequently identified
by mass spectrometry. The modified proteins were also localized at the tissue level by
immunohistochemical analysis.
Iso[4]LGE2 derived protein modifications are distinctly increased in the
glaucomatous TM compared to that of the controls (p < 2E-11) (Figure 4.11).
Iso[4]LGE2’s are known to irreversibly form adducts that accumulate, and thus reflect the
139

total oxidative stress undergone by the tissue. Thus, the adduct level may be considered
as a dosimeter of oxidative stress. Protein modifications derived from HNE, another lipid
oxidation product, are also elevated in glaucomatous TM compared to that of the controls
(p < 0.0003) (Figure 4.11). Thus, through immunoblots, a strong correlation between
oxidative stress and POAG was established. The vast difference in the ‘p’ values of these
two different lipobolites may suggest that adducts formed from HNE may be formed
reversibly or slowly compared to the iso[4]LGE2 adducts. This is supportive of earlier
reports suggesting that levuglandins bind far more avidly to LDL compared to HNE or
MDA.31,81 So the latter may be cleared before they have an opportunity to form adducts.
140

-1
1
3
5
7
9
11
13
15
17
0
2
4
6
8
10
12
14
Rel
ativ
e in
tens
ity (O
D)
(HN
E-im
mun
orea
ctiv
e ba
nds)
Controls POAG
p≤0.0003
BR
elat
ive
inte
nsity
(OD
) (is
o[4]
LGE 2
imm
unor
eact
ive
band
s)
p≤2E-11
Controls POAG
A
-1
1
3
5
7
9
11
13
15
17
0
2
4
6
8
10
12
14
Rel
ativ
e in
tens
ity (O
D)
(HN
E-im
mun
orea
ctiv
e ba
nds)
Controls POAG
p≤0.0003
BR
elat
ive
inte
nsity
(OD
) (is
o[4]
LGE 2
imm
unor
eact
ive
band
s)
p≤2E-11p≤2E-11
Controls POAG
A
Figure 4.11: Quantification of iso[4]LGE2 and HNE immunoreactive bands in 1D
Western blots. Intensity for each of the lanes detected (all the bands in the lane included)
in the Western blots was normalized with respect to the total of all the bands detected in
that blot (details of calculations in the experimental section). The minimum intensity for
band detection was fixed for each of the blots. ◊ – Relative intensity of a control sample
on the blot; □ – OD of a POAG tissue on the blot. A. Levels of iso[4]LGE2 modified
proteins (tissue donors – 23 POAGs, 22 controls, shown in Figures 4.3 and 4.1S
(appendix)), B. Levels of HNE modified proteins (tissue donors – 23 POAGs, 22
controls, shown in Figures 4.4 and 4.2S (appendix)). ‘p’ values calculated by students t-
test using Microsoft ® Excel 2003.
Additionally, a moderate increase in the levels of argpyrimidine modification was
evident through Western blot analysis using antibodies against argpyrimidine in
glaucomatous TM compared to the controls (p < 0.0035). Though the analysis did not
take into account whether the subjects are diabetic or not, the difference in these levels
along with the evidence for increased lipid oxidation, may point to an increase in the
141

contribution of oxidative lipid fragmentations to the production of methyl glyoxal. This is
additional evidence to previous reports that suggested a contribution of oxidative stress to
the formation of methyl glyoxal in addition to fragmentation of sugars and glycolysis
products.69 There is also increase in the amount of oxidized tryptophan as measured by
the amount of OHKYN using the anti-OHKYN antibodies.
2
3
4
5
0
5
10
15
20
25
30
35
40
Rel
ativ
e in
tens
ity (O
D)
(Arg
pyrim
idin
e im
mun
orea
ctiv
e ba
nds)
Rel
ativ
e in
tens
ity (O
D)
(OH
KYN
imm
unor
eact
ive
band
s)
p≤0.0015p≤0.001
Controls POAGControls POAG
BA
2
3
4
5
0
5
10
15
20
25
30
35
40
Rel
ativ
e in
tens
ity (O
D)
(Arg
pyrim
idin
e im
mun
orea
ctiv
e ba
nds)
Rel
ativ
e in
tens
ity (O
D)
(OH
KYN
imm
unor
eact
ive
band
s)
p≤0.0015p≤0.0015p≤0.001p≤0.001
Controls POAGControls POAG
BA
Figure 4.12: Quantification of argpyrimidine and OHKYN immunoreactive bands in 1D
Western blots. Relative intensity of optical density (OD) for each of the lanes (all the
bands in each lane included) in the Western blots was normalized for each of the blots (8
lanes each). The minimum intensity for band detection was fixed for each of the blots. ◊ –
Relative intensity of a control sample on the blot; □ – relative intensity of a POAG tissue
on the blot. A. Levels of argpyrimidine modified proteins (tissue donors – 12 POAGs, 12
controls, shown in Figures 4.5 and 4.3S (appendix)). B. Levels of OHKYN modified
proteins (tissue donors – 8 POAGs, 8 controls, shown in Figures 4.6). ‘p’ values
calculated by students t-test using Microsoft ® Excel 2003.
Based on our data from the immunoblots, we performed the immunoprecipitation
(IP) of the modified proteins from the glaucomatous TM. IP was done using antibody
142

(Iso[4]LGE2 and HNE) covalently coupled with sepharose A beads. The
immunoprecipitated proteins were analyzed for their immunoreactivity by immunoblot
analysis (Figure 5, Lane 1’). Following 1D SDS PAGE of the immunoprecipitated
proteins, the gel bands were excised, and the proteins analyzed by mass spectrometry. In
both of the IPs a majority of the proteins identified are the extracellular matrix (ECM)
proteins, suggesting that these proteins are especially vulnerable to oxidative protein
modifications. The proteins identified by immunoprecipitation to be oxidatively modified
were also reported in our earlier proteomic analysis of the TM.82
Keratoepithelin (TGF-βig-h3) one of the proteins identified, is found in many
connective tissues and considered to have a binding function of these tissues.83 Elevated
levels and/or abnormal aggregation of keratoepithelin has been associated with type 2
diabetic nephropathy and corneal dystrophies.84 We also found β-IGH3, a protein
localized in the corneal epithelial tissue that is apparently modified by lipid oxidation
products. However, the possibility that these deposits in the TM originate by detachment
from the corneal epithelium, dispersal into the aqueous humor, and deposition in the TM
cannot be ruled out. Protein disulphide isomerase (PDI),85 an enzyme involved in
formation of disulphide bonds in native proteins that also acts as a chaperone, is
immunoprecipitated by HNE antibodies. Modification of this enzyme could cause protein
misfolding and aggregation. Other prominent proteins that immunoprecipitated are the
extracellular matrix associated proteins (viz, decorin, lumican, prolargin and vitronectin)
and those involved in energy metabolism (aldehyde dehydrogenase and pyruvate kinase).
This observation suggests that increase in oxidative stress can potentially compromise the
morphology of the cells lining the TM of POAG as reported earlier.86
143

In addition to IP, 2D SDS PAGE analysis of the TM extract was also performed
for identification of the oxidatively modified proteins. Owing to better separation of
proteins, this analysis resulted in more protein hits compared to the IP. The mass
spectrometric analysis of immunoreactive spots revealed several ECM matrix proteins,
corroborating the results from the IP. Oxidative stress conditions have been shown to
lessen the adhesion of cultured TM cells to ECM proteins.86 This could induce changes in
the cell cycle 87 or apoptosis.88 Identification of transferrin, a iron regulating protein
shown to be elevated in retinal tissue of experimental glaucoma in monkey,89 shows the
presence of antioxidant systems.
The changes observed in the molecular weight of the proteins identified by 2D
PAGE may reflect aggregation by crosslinking (increased MW) caused by the oxidized
lipids or oxidative protein degradation (decreased MW), while the changes in the pI
reflect charge changes due to modifications and/or transformation. Some of the protein
spots identified on the 2D gel had molecular weight that corresponds to a dimer of the
protein identified. Table 4.4 shows a list of the proteins that were identified by mass
spectrometry of the anti-iso[4]LGE2 pAbs and the anti-HNE pAbs immunoreactive spots
on the 2D PAGE (Figure 4.10).
α-Actinin is a homodimer involved in the crosslinking of actins, which is
important in the formation of actin filaments.90 Actinin appears at ~52 kDa (spots 11, 36,
Figure 4.10) that corresponds to the monomer. The list includes a cysteine protease
(cathepsin k) that cleaves collagens I and II in bone and has been implicated in diseases
such as osteoporosis91 and amyloidosis.92 Several of the ECM proteins were also
144

identified to be crosslinked by these lipid oxidation products. Crosslinking of proteins
can affect the mobility and functions of the proteins.
Table 4.4: List of putatively crosslinked proteins identified by 2D PAGE and mass
spectrometric analyses of glaucomatous TM.
Actual Observed ImmunoreactiveProtein Identified MW
(kDa) pI MW (kDa) pI isoLGE HNE
Peptide match
Accession No.
cathepsin K 37 8.8 98 7.3 √ 1 P43235
Keratocan 40 7.2 93 8.4 √ 2 O62702
Lumican 38 6.5 92, 67 - √ √ 2 P51884
prolargin 44 9.6 100-45 - √ √ 5 P51888
Lumican is a kertansulphate proteoglycan found mostly in the sclera and some
connective tissues involved in modulating collagen fibril formation. The 2D shows the
presence of lumican crosslinked as a high molecular weight species (putative dimer and
trimer) in the 2D PAGE. Crosslinking of this protein could result in malformed collagen,
a key structural protein in the TM.93 Prolargin, a proline-arginine-rich end leucine-rich
repeat protein, anchors basement membranes to the connective tissue.94 Prolargin also
appears as a high molecular weight species (putative dimer) (Table 4.4). Earlier reports of
the protein crosslinking ability of iso[4]LGE2 and HNE,30,95 are in accord with these
results.
As the TM isolated from the corneal rim region or by trabeculectomy is likely to
have some of the surrounding corneal, scleral or iris tissue, the proteins identified cannot
145

be unambiguously attributed to the TM region. To localize the modified proteins,
immunohistochemical analysis was performed in the TM region using the iso[4]LGE2
and HNE antibodies. There is an increased amount of staining in the TM region of the
glaucomatous TM sections for the lipid specific antibodies compared to that of the
preimmune IgG treated controls (Figure 4.7, Figure 4.8). Preimmune staining of the
sections eliminated the presence of any non-specific binding artifacts.
We postulate that oxidative modifications, including covalent crosslinks,
accumulate in TM and contribute to aqueous outflow blockage and onset of POAG. The
trabecular meshwork environment is conducive to oxidative damage especially with
age.16,17 The risk of glaucoma increases with age, with POAG at age 80 being 5 to 10
times more prevalent than at age 40.96,97 The body combats free radicals and other
reactive oxidants through a highly efficient and adaptive antioxidant system (enzymatic
and non-enzymatic). It has been shown that there is a decrease in the levels of GSH 98,
while no difference in the activity of MPO and catalase in plasma, whilst a big difference
in the MDA derived protein modifications72 is present in POAG compared to that of
controls. Decrease in the activity of proteasome function and GSH levels, and increase in
lipid peroxides 99 and a stress response mediated by ELAM-1100 have been observed in
cultured TM cells under stress. TM cells have been shown to be able to synthesize a
specific set of proteins that may act as molecular chaperones to prevent oxidative or heat
shock damage.16 In addition to these reports, our findings suggest an oxidative damage
component in glaucoma pathogenesis, and raise many additional questions about the role
of lipid peroxidation in glaucoma. Is it the increase of lipid oxidation-regulating machine
146

that is affected or is it the cellular degradation machinery that is impaired during the
process of aging?
Our findings might thus represent identification of an oxidative stress response
component in glaucoma. It is important, now, to address this question directly by testing
the efficacy of glaucoma therapies that target antioxidant systems. Potentially
encouraging is the finding that antioxidant therapy can be beneficial in certain forms of
ARMD and cataract.11 However, enthusiasm for this possibility must be tempered by the
report that antioxidant therapy is not effective in age-related cataract, a disease also
linked to oxidative stress and iron abnormalities.101
147

4.4. Experimental Procedures.
4.4.1. General methods. All chemicals used were of high purity analytical grade. The
following commercially available materials were used as received: bovine serum albumin
(BSA, fraction V, 96-99%) and human serum albumin (HSA, fraction V) were from
Sigma (St. Louis, MO); goat anti-rabbit IgG-alkaline phos¬phatase (Boehringer-
Mannheim); p-(N,N-dimethylamino) benzaldehyde (DMAB, Aldrich, WI). Spectrapor
membrane tubing (Mr cutoff 14,000 No. 2) for dialysis was obtained from Fisher
Scientific Co. Phosphate buffered saline (PBS) was prepared from a pH 7.4 stock
solution containing 0.2 M NaH2PO4/Na2HPO4, 3.0 M NaCl, and 0.02% NaN3 (w/w).
This solution was diluted to 10 mM or as needed. ON-KLH antibodies102 and
iso[4]LGE2-KLH antibodies6 were prepared as described previously. Iso[4]LGE2 was
prepared by Jim Laird and ON-BSA was prepared by Liang Lu in our laboratory. The
iso[4]LGE2 –HSA, iso[4]LGE2-BSA103 and ON/PPC-HSA104 were prepared as described
previously. The monoclonal anti-argpyrimidine antibody was obtained as gift from Dr.
Kochi Uchida (Nagoya University, Japan) and MG-BSA56 was obtained from Dr. Ram H.
Nagaraj (Case Western Reserve University, USA).
4.4.2. Tissue procurement. Human eyes from 52 normal donors and 52 POAG donors,
all between the ages of 43 and 95 years of age, used in this study were obtained through
the Cleveland Eye Bank and the Cleveland Clinic Foundation (Clinical practices of Drs.
Edward Rockwood and Scott Smith). Eyes were enucleated within 10 h of death and
stored at -80 ºC until TM tissue was isolated by dissection. Control eyes were from
donors with no visual field defects, no evidence of glaucoma and without any disease of
148

the central nervous system. Fixed human TM tissues used for immunohistochemistry
were obtained from the National Disease Research Interchange (NDRI, Philadelphia,
PA). Glaucomatous eyes and tissues were from clinically documented POAG donors.
Glaucomatous TM tissues (~1-2 mm3) were obtained by trabeculectomy of POAG
patients in the Cole Eye Institute, of the Cleveland Clinic Foundation with institutional
review board approval. Human tissues obtained by trabeculectomy were predominantly
TM but possible contamination with small amounts of surrounding tissue (e.g., sclera)
cannot be excluded.
4.4.3. Protein extraction. TM from cadaver and trabeculectomy samples was extracted
by homogenization in 100 mM Tris-Cl buffer pH 7.8 containing 5 mM dithiothreitol,
1mM SnCl2, 50 mM NaHPO4, 1mM diethylenetriaminepentaacetic acid, 100 mM
butylated hydroxy toluene and 0.5% SDS. Insoluble material was removed by
centrifugation (8000g for 5 min). Soluble protein was quantified by the Bradford assay 105
using amino acid quantified BSA as standard, yielding ~15-20 µg total soluble protein
per trabeculectomy tissue sample (~1-2 mm3). Protein extracts were subjected to SDS-
PAGE on 10% gels (Bio-Rad Laboratories, Hercules, CA) and the gels were used either
for Western analyses or for mass spectrometric proteomic analyses.12,82 The soluble
protein was subjected to SDS-PAGE or kept at -80 ºC until analyzed.
4.4.4. Western analysis. Electrophoresis and Western blotting were carried out
according to published protocols.12,82 Protein extracts (~ 10-15 µg) from TM tissue were
suspended in Laemmli sample buffer,82 fractionated on a 10% SDS PAGE, electro blotted
149

on to polyvinyl difluoride (PVDF) membrane (Millipore, Bedford, MA) and probed using
various antibodies: rabbit polyclonal anti-HNE,41,102 rabbit polyclonal anti-iso[4]LGE2 ,33
mouse monoclonal anti-methylglyoxal (argpyrimidine)56 and anti-hydroxykynurenine
mAb.59 As the amount of protein extracted from TM was very low, equal loading was
verified by Coomassie blue stained gels concomitantly with Bradford assay.105 The
immunoreactive bands were quantified using GS-710 scanner and QuantityOne ®
software from Biorad™ (Hercules, CA).Quantification of the Western blot were
performed in two steps. Firstly, the Western blots were normalized based on the
Coomassie blue stained gels to ensure the incorporation of variations in the protein
amounts. Secondly, normalization was performed between Western blots to exclude any
variations between the blots.
4.4.5. Histochemical analysis. Immunohistochemical analyses to localize modified
proteins in ocular tissue were performed with cadaver eyes enucleated within six hours of
death and fixed immediately with calcium acetate buffered 4% para-formaldehyde.
Paraffin embedded tissue was blocked and sectioned (10 µm). The sections were serially
incubated in 1% BSA in phosphate buffered saline pH 7.5, then with 10-100 ng antibody
(rabbit polyclonal anti-(HNE) 41 or rabbit polyclonal anti-Iso[4]LGE2 33) overnight at 4
°C. The sections were washed with a solution of 1% BSA in PBS (3 x 5 min) and
subsequently incubated with 10 ng secondary antibody conjugated with Alexa 594 or
Alexa 488 (Molecular Probes Inc, Eugene, OR) for one hour at room temperature.
Sections were washed with 1% BSA in PBS as mentioned above and sealed with
Vectashield® and analyzed with a Nikon EFD-3 fluorescence microscope attached to a
150

CCD camera. For localization of TM, slides were stained with hematoxylin by serially
incubating in staining solution for 3 min, tap water for 5 min and finally in distilled water
for 5 min. The slides were then dried in air and visualized by light microscopy. The
microscopic images were assembled in Adobe® Photoshop® CS.
4.4.6. Immunoprecipitation. Immunoprecipitation required protein extracts without any
SDS as this might hinder the antibody antigen interaction. Protein was extracted using
100 mM Tris-Cl buffer pH 7.5, 50 mM NaCl and 0.01% genapol unlike the methods
mentioned above. Antibody-coupled protein A beads were used for all
immunoprecipitations. About 100 µg of swelled protein A sepharose CL-4B beads
(Amersham Pharmacia Biotech, CA) was coupled with 100 µg antibody using 4 x 25 mg
dimethylpimelimidate (DMP), each with 2 h incubation at room temperature in 50 mM
sodium borate buffer pH 8.3. During each of the washes, the beads were centrifuged at
2000 rpm for 2 min and the supernatant carefully drained. Following incubation, the
beads were washed with 200 mM ethanolamine pH 8.0 and incubated in the same
solution for 2 h. Beads (100 µg antibody-coupled beads) were finally washed in
phosphate buffered saline pH 7.4 and incubated with protein extracts (~30 µg) prepared
in 100 mM Tris-Cl buffer pH 7.5, 50 mM NaCl and 0.01% genapol. After incubation, the
beads were recovered by centrifugation at 2500 g for 5 minutes and washed 3 x 500 µL
of 100 mM Tris-Cl buffer pH 7.5, 100 mM NaCl and 0.02% genapol. The beads were
boiled with 5 µL Laemmli buffer82 for 2 minutes and separated on a 10% SDS-PAGE.
The gels were either subjected to Coomassie staining and/or subsequent LC MS/MS of
gel bands or Western analyses.
151

4.4.7. 2D Gel electrophoresis. Protein extract from a dissected TM of a 79 year old
Caucasian male, extracted using solution B (7 M urea, 4% Chaps, 2 M thiourea, and 0.5%
Triton-X), was used for the two dimensional electrophoresis. Protein extract (~80 µg)
was transferred to 7 cm IPGphore coffins (Amersham Pharmacia Biosciences,
Piscataway, NJ) and the volume adjusted to 300 µL with additional detergent solution. A
7 cm precast ProteomIQ™ immobilized non-linear pH gradient strip gel (IPG, pH 3-10
NL) was placed in the coffin and overlaid with 600 µL of mineral oil. First dimension
IEF was performed using the Pharmacia IPGphor and the following programmed voltage
gradient; IPG strips were rehydrated with sample at 30 V/6 h; then 300V/1 min; followed
by a linear increase to 3500 V/8 h, held at 3500 V/1 h; then at 8000 V/21 h to reach
approximately 80 kVh. For the second dimension, the IPG strip was reduced in a buffer
(50 mM TrisCl pH 8.8, 6 M urea, 30% glycerol, 2% SDS) containing 1% dithiothreitol
(DTT) for 12 min and followed by alkylation using 0.25% iodoacetamide with a trace of
bromophenol as described earlier.71 The equilibrated IPG strip was embedded in 0.7%
w/v agarose on the top of a 10% acrylamide gel containing a 4% stacking gel. Second
dimension SDS-PAGE (2h at 80 V constant voltage) was performed with Bio-rad
electrophoresis equipment. The gel was stained using GelCode Blue® (Pierce) and
scanned with a GS-710 Imaging Densitometer™ (BioRad). The coordinates of spots on
the 2D image was digitalized using Origin® 7.5 software assuming a linear scale between
the least measurable units shown (both pI and molecular weight).
152

4.4.8. LC MS/MS analysis and protein identification. Following the SDS PAGE, the
Gelcode Blue® stained gel bands were excised in 1mm3 slices and destained according to
standard protocol for in-gel digestion.12,82 The tryptic digested proteins were analyzed by
LC-MS/MS using a quadrupole-time of flight (QTof2) mass spectrometer equipped with
a CapLC system (Waters, Millford, MA), ProteinLynx Global Server acquisition and
processing software. Peptide digests were trapped on a precolumn (0.3 x 1mm, 5µC18,
LC Packing) with 0.1% formic acid in 2% acetonitrile as loading solvent then eluted onto
a capillary column (PicoFrit 0.050 x 50mm, 5µ tip ID; New Objective Inc., Woburn,
MA). Chromatography was performed with a flow of 250 nL/min with aqueous
acetonitrile/formic acid solvents and 100% of the eluant was directed towards the mass
spectrometry. The mass spectrometer was operated in standard MS/MS switching mode
with the three most intense ions in each survey scan subjected to MS/MS analysis.
Protein identifications from MS/MS data utilized ProteinLynxTM Global Server (Waters
Corporation) and Mascot (Matrix Science) search engines and the Swiss-Protein and
NCBI protein sequence databases.12,82
153

4.5 References
(1) Stadtman, E. R. Science 1992, 257, 1220-4. (2) Marnett, L. J.; Hurd, H. K.; Hollstein, M. C.; Levin, D. E.; Esterbauer, H.; Ames,
B. N. Mutat Res 1985, 148, 25-34. (3) Fraga, C. G.; Shigenaga, M. K.; Park, J. W.; Degan, P.; Ames, B. N. Proc Natl
Acad Sci U S A 1990, 87, 4533-7. (4) Murphy, T. C.; Poppe, C.; Porter, J. E.; Montine, T. J.; Picklo, M. J., Sr. J
Neurochem 2004, 89, 1462-70. (5) Podrez, E. A.; Poliakov, E.; Shen, Z.; Zhang, R.; Deng, Y.; Sun, M.; Finton, P. J.;
Shan, L.; Gugiu, B.; Fox, P. L.; Hoff, H. F.; Salomon, R. G.; Hazen, S. L. J Biol Chem 2002, 277, 38503-16.
(6) Salomon, R. G.; Subbanagounder, G.; O'Neil, J.; Kaur, K.; Smith, M. A.; Hoff, H. F.; Perry, G.; Monnier, V. M. Chem Res Toxicol 1997, 10, 536-45.
(7) Salomon, R. G.; Kaur, K.; Podrez, E.; Hoff, H. F.; Krushinsky, A. V.; Sayre, L. M. Chem Res Toxicol 2000, 13, 557-64.
(8) Salomon, R. G. Antioxid Redox Signal 2005, 7, 185-201. (9) Gu, X.; Meer, S. G.; Miyagi, M.; Rayborn, M. E.; Hollyfield, J. G.; Crabb, J. W.;
Salomon, R. G. J Biol Chem 2003, 278, 42027-35. (10) Cringle, S. J.; Yu, D. Y.; Yu, P. K.; Su, E. N. Invest Ophthalmol Vis Sci 2002, 43,
1922-7. (11) Beatty, S.; Koh, H.; Phil, M.; Henson, D.; Boulton, M. Surv Ophthalmol 2000, 45,
115-34. (12) Crabb, J. W.; Miyagi, M.; Gu, X.; Shadrach, K.; West, K. A.; Sakaguchi, H.;
Kamei, M.; Hasan, A.; Yan, L.; Rayborn, M. E.; Salomon, R. G.; Hollyfield, J. G. Proc Natl Acad Sci U S A 2002, 99, 14682-7.
(13) Stillwell, W.; Wassall, S. R. Chem Phys Lipids 2003, 126, 1-27. (14) Quigley, H. A. Br J Ophthalmol 1996, 80, 389-93. (15) Oyster, C. W. The Human Eye Structure and Function; Sinauer Associates, Inc.:
Sunderland, Massachusetts., 1999. (16) Tamm, E. R.; Russell, P.; Johnson, D. H.; Piatigorsky, J. Invest Ophthalmol Vis
Sci 1996, 37, 2402-13. (17) De La Paz, M. A.; Epstein, D. L. Invest Ophthalmol Vis Sci 1996, 37, 1849-53. (18) Green, K. Ophthalmic Res 1995, 27 Suppl 1, 143-9. (19) Babizhayev, M. A.; Bunin, A. Acta Ophthalmol (Copenh) 1989, 67, 371-7. (20) Yan, D. B.; Trope, G. E.; Ethier, C. R.; Menon, I. A.; Wakeham, A. Invest
Ophthalmol Vis Sci 1991, 32, 2515-20. (21) Russell, P.; Johnson, D. H. J Glaucoma 1996, 5, 317-24. (22) Russell, P.; Garland, D.; Epstein, D. L. Exp Eye Res 1989, 48, 251-60. (23) Gherghel, D.; Griffiths, H. R.; Hilton, E. J.; Cunliffe, I. A.; Hosking, S. L. Invest.
Ophthalmol. Vis. Sci. 2005, 46, 877-883. (24) Bunin, A. Vestn Oftalmol 2000, 116, 24-7. (25) Veach, J. Br J Nutr 2004, 91, 809-29. (26) Padgaonkar, V. A.; Giblin, F. J.; Leverenz, V.; Reddy, V. N. J. Glaucoma 1994,
3, 123-131.
154

(27) Salomon, R. G.; Miller, D. B. Adv Prostaglandin Thromboxane Leukot Res 1985, 15, 323-6.
(28) Morrow, J. D.; Harris, T. M.; Roberts, L. J., 2nd Anal Biochem 1990, 184, 1-10. (29) Salomon, R. G. Chem Phys Lipids 2005, 134, 1-20. (30) Iyer, R. S.; Ghosh, S.; Salomon, R. G. Prostaglandins 1989, 37, 471-80. (31) Hoppe, G.; Subbanagounder, G.; O'Neil, J.; Salomon, R. G.; Hoff, H. F. Biochim
Biophys Acta 1997, 1344, 1-5. (32) Davies, S. S.; Amarnath, V.; Montine, K. S.; Bernoud-Hubac, N.; Boutaud, O.;
Montine, T. J.; Roberts, L. J., 2nd Faseb J 2002, 16, 715-7. (33) Salomon, R. G.; Batyreva, E.; Kaur, K.; Sprecher, D. L.; Schreiber, M. J.; Crabb,
J. W.; Penn, M. S.; DiCorletoe, A. M.; Hazen, S. L.; Podrez, E. A. Biochim Biophys Acta 2000, 1485, 225-35.
(34) Salomon, R. G.; Sha, W.; Brame, C.; Kaur, K.; Subbanagounder, G.; O'Neil, J.; Hoff, H. F.; Roberts, L. J., 2nd J Biol Chem 1999, 274, 20271-80.
(35) Murphy, M. E.; Kehrer, J. P. Chem Biol Interact 1989, 69, 101-73. (36) Tak, P. P.; Zvaifler, N. J.; Green, D. R.; Firestein, G. S. Immunol Today 2000, 21,
78-82. (37) Shacter, E. Drug Metab Rev 2000, 32, 307-26. (38) Esterbauer, H.; Schaur, R. J.; Zollner, H. Free Radic Biol Med 1991, 11, 81-128. (39) Zarkovic, N. Mol Aspects Med 2003, 24, 281-91. (40) Zarkovic, K. Mol Aspects Med 2003, 24, 293-303. (41) Uchida, K.; Toyokuni, S.; Nishikawa, K.; Kawakishi, S.; Oda, H.; Hiai, H.;
Stadtman, E. R. Biochemistry 1994, 33, 12487-94. (42) Amarnath, V.; Valentine, W. M.; Montine, T. J.; Patterson, W. H.; Amarnath, K.;
Bassett, C. N.; Graham, D. G. Chem Res Toxicol 1998, 11, 317-328. (43) Liu, Y.; Sun, G.; David, A.; Sayre, L. M. Chem Res Toxicol 2004, 17, 110-8. (44) Nadkarni, D. V.; Sayre, L. M. Chem Res Toxicol 1995, 8, 284-91. (45) Hoff, H.; O'Neil, J.; Chisolm, G., 3d; Cole, T.; Quehenberger, O.; Esterbauer, H.;
Jurgens, G. Arterioscler Thromb Vasc Biol 1989, 9, 538-549. (46) Sayre, L. M.; Zelasko, D. A.; Harris, P. L.; Perry, G.; Salomon, R. G.; Smith, M.
A. J Neurochem 1997, 68, 2092-7. (47) Verdejo, C.; Marco, P.; Renau-Piqueras, J.; Pinazo-Duran, M. D. Eye 1999, 13 (
Pt 2), 183-8. (48) Sell, D. R.; Lane, M. A.; Johnson, W. A.; Masoro, E. J.; Mock, O. B.; Reiser, K.
M.; Fogarty, J. F.; Cutler, R. G.; Ingram, D. K.; Roth, G. S.; Monnier, V. M. Proc Natl Acad Sci U S A 1996, 93, 485-90.
(49) Araki, N.; Ueno, N.; Chakrabarti, B.; Morino, Y.; Horiuchi, S. J Biol Chem 1992, 267, 10211-4.
(50) Gerhardinger, C.; Taneda, S.; Marion, M. S.; Monnier, V. M. J Biol Chem 1994, 269, 27297-302.
(51) Du Yan, S.; Zhu, H.; Fu, J.; Yan, S. F.; Roher, A.; Tourtellotte, W. W.; Rajavashisth, T.; Chen, X.; Godman, G. C.; Stern, D.; Schmidt, A. M. Proc Natl Acad Sci U S A 1997, 94, 5296-301.
(52) Vitek, M. P.; Bhattacharya, K.; Glendening, J. M.; Stopa, E.; Vlassara, H.; Bucala, R.; Manogue, K.; Cerami, A. Proc Natl Acad Sci U S A 1994, 91, 4766-70.
155

(53) Yan, S. D.; Chen, X.; Schmidt, A. M.; Brett, J.; Godman, G.; Zou, Y. S.; Scott, C. W.; Caputo, C.; Frappier, T.; Smith, M. A.; et al. Proc Natl Acad Sci U S A 1994, 91, 7787-91.
(54) Miyata, T.; Inagi, R.; Iida, Y.; Sato, M.; Yamada, N.; Oda, O.; Maeda, K.; Seo, H. J Clin Invest 1994, 93, 521-8.
(55) Neeper, M.; Schmidt, A. M.; Brett, J.; Yan, S. D.; Wang, F.; Pan, Y. C.; Elliston, K.; Stern, D.; Shaw, A. J Biol Chem 1992, 267, 14998-5004.
(56) Oya, T.; Hattori, N.; Mizuno, Y.; Miyata, S.; Maeda, S.; Osawa, T.; Uchida, K. J Biol Chem 1999, 274, 18492-502.
(57) Baynes, J. W. Exp Gerontol 2001, 36, 1527-37. (58) Van Heijst, J. W. J.; Niessen, H. W. M.; Hoekman, K.; Schalkwijk, C. G. Ann NY
Acad Sci 2005, 1043, 725-733. (59) Staniszewska, M. M.; Nagaraj, R. H. J. Biol. Chem. 2005, 280, 22154-22164. (60) Goldstein, L. E.; Leopold, M. C.; Huang, X.; Atwood, C. S.; Saunders, A. J.;
Hartshorn, M.; Lim, J. T.; Faget, K. Y.; Muffat, J. A.; Scarpa, R. C.; Chylack, L. T., Jr.; Bowden, E. F.; Tanzi, R. E.; Bush, A. I. Biochemistry 2000, 39, 7266-75.
(61) Reynolds, G.; Pearson, S. Lancet 1989, 2, 979-80. (62) Ogawa, T.; Matson, W.; Beal, M.; Myers, R.; Bird, E.; Milbury, P.; Saso, S.
Neurology 1992, 42, 1702-6. (63) Sardar, A.; Bell, J.; Reynolds, G. J Neurochem 1995, 64, 932-5. (64) Vazquez, S.; Aquilina, J. A.; Jamie, J. F.; Sheil, M. M.; Truscott, R. J. W. J. Biol.
Chem. 2002, 277, 4867-4873. (65) Hood, B. D.; Garner, B.; Truscott, R. J. J Biol Chem 1999, 274, 32547-50. (66) Garner, B.; Vazquez, S.; Griffith, R.; Lindner, R. A.; Carver, J. A.; Truscott, R. J.
J Biol Chem 1999, 274, 20847-54. (67) Vazquez, S.; Garner, B.; Sheil, M.; Truscott, R. Free Radic Res 2000, 32, 11-23. (68) Garner, B.; Shaw, D. C.; Lindner, R. A.; Carver, J. A.; Truscott, R. J. Biochim
Biophys Acta 2000, 1476, 265-78. (69) Shamsi, F. A.; Lin, K.; Sady, C.; Nagaraj, R. H. Invest Ophthalmol Vis Sci 1998,
39, 2355-64. (70) Doolittle, M. H.; Ben-Zeev, O.; Briquet-Laugier, V. J. Lipid Res. 1998, 39, 934-
942. (71) Miyagi, M.; Sakaguchi, H.; Darrow, R. M.; Yan, L.; West, K. A.; Aulak, K. S.;
Stuehr, D. J.; Hollyfield, J. G.; Organisciak, D. T.; Crabb, J. W. Mol Cell Proteomics 2002, 1, 293-303.
(72) Yildirim, O.; Ates, N. A.; Ercan, B.; Muslu, N.; Unlu, A.; Tamer, L.; Atik, U.; Kanik, A. Eye 2004, 1-4.
(73) Choi, J.; Rees, H. D.; Weintraub, S. T.; Levey, A. I.; Chin, L. S.; Li, L. J Biol Chem 2005, 280, 11648-55.
(74) Moreno, M. C.; Campanelli, J.; Sande, P.; Sanez, D. A.; Keller Sarmiento, M. I.; Rosenstein, R. E. Free Radic Biol Med 2004, 37, 803-12.
(75) Zarbin, M. A. Arch Ophthalmol 2004, 122, 598-614. (76) Ferreira, S. M.; Lerner, S. F.; Brunzini, R.; Evelson, P. A.; Llesuy, S. F. Am J
Ophthalmol 2004, 137, 62-9. (77) Izzotti, A.; Sacca, S. C.; Cartiglia, C.; De Flora, S. Am J Med 2003, 114, 638-46. (78) Chen, J. Z.; Kadlubar, F. F. Am J Med 2003, 114, 697-8.
156

(79) Kaur, K.; Salomon, R. G.; O'Neil, J.; Hoff, H. F. Chem Res Toxicol 1997, 10, 1387-96.
(80) Crabb, J. W.; O'Neil, J.; Miyagi, M.; West, K.; Hoff, H. F. Protein Sci 2002, 11, 831-40.
(81) Brame, C. J.; Salomon, R. G.; Morrow, J. D.; Roberts, L. J., 2nd J Biol Chem 1999, 274, 13139-46.
(82) Bhattacharya, S. K.; Rockwood, E. J.; Smith, S. D.; Bonilha, V. L.; Crabb, J. S.; Kuchtey, R. W.; Robertson, N. G.; Peachey, N. S.; Morton, C. C.; Crabb, J. W. J Biol Chem 2005, 280, 6080-4.
(83) Streeten, B. W.; Qi, Y.; Klintworth, G. K.; Eagle, R. C., Jr.; Strauss, J. A.; Bennett, K. Arch Ophthalmol 1999, 117, 67-75.
(84) Ha, S. W.; Kim, H. J.; Bae, J. S.; Jeong, G. H.; Chung, S. C.; Kim, J. G.; Park, S. H.; Kim, Y. L.; Kam, S.; Kim, I. S.; Kim, B. W. Diabetes Res Clin Pract 2004, 65, 167-73.
(85) Wang, C. C.; Tsou, C. L. Faseb J 1993, 7, 1515-7. (86) Zhou, L.; Li, Y.; Yue, B. Y. J Cell Physiol 1999, 180, 182-9. (87) Giancotti, F. G. Curr Opin Cell Biol 1997, 9, 691-700. (88) Frisch, S. M.; Ruoslahti, E. Curr Opin Cell Biol 1997, 9, 701-6. (89) Farkas, R. H.; Chowers, I.; Hackam, A. S.; Kageyama, M.; Nickells, R. W.;
Otteson, D. C.; Duh, E. J.; Wang, C.; Valenta, D. F.; Gunatilaka, T. L.; Pease, M. E.; Quigley, H. A.; Zack, D. J. Invest Ophthalmol Vis Sci 2004, 45, 1410-7.
(90) Matsudaira, P. Trends Biochem Sci 1991, 16, 87-92. (91) Motyckova, G.; Weilbaecher, K.; Horstmann, M.; Rieman, D.; Fisher, D.; Fisher,
D. Proc Nat Acad Sci U S A 2001, 98, 5798-803. (92) Rocken, C.; Stix, B.; Bromme, D.; Ansorge, S.; Roessner, A.; Buhling, F. Am J
Pathol 2001, 158, 1029-1038. (93) Dunlevy, J. R.; Rada, J. A. S. Invest. Ophthalmol. Vis. Sci. 2004, 45, 3849-3856. (94) http://ca.expasy.org/uniprot/P51888, 2005. (95) Jirousek, M. R.; Murthi, K. K.; Salomon, R. G. Prostaglandins 1990, 40, 187-
203. (96) Wilson, M. R.; Mansour, M.; Atud, A. E.; Casey, R.; Fobi, G.; Moukouri, E.;
Alemayehu, W.; Martone, J. F.; Ross-Degnan, D. Arch Ophthalmol 1996, 114, 464-8.
(97) Gordon, M. O.; Beiser, J. A.; Brandt, J. D.; Heuer, D. K.; Higginbotham, E. J.; Johnson, C. A.; Keltner, J. L.; Miller, J. P.; Parrish, R. K., 2nd; Wilson, M. R.; Kass, M. A. Arch Ophthalmol 2002, 120, 714-20; discussion 829-30.
(98) Gherghel, D.; Griffiths, H. R.; Hilton, E. J.; Cunliffe, I. A.; Hosking, S. L. Invest Ophthalmol Vis Sci 2005, 46, 877-83.
(99) Caballero, M.; Liton, P. B.; Epstein, D. L.; Gonzalez, P. Biochem Biophys Res Commun 2003, 308, 346-52.
(100) Wang, N.; Chintala, S. K.; Fini, M. E.; Schuman, J. S. Nat Med 2001, 7, 304-9. (101) Jr, C. L.; Brown, N.; Bron, A.; Hurst, M.; Kopcke, W.; Thien, U.; Schalch, W.
Ophthal Epidemiol 2002, 9, 49-80. (102) Sayre, L. M.; Sha, W.; Xu, G.; Kaur, K.; Nadkarni, D.; Subbanagounder, G.;
Salomon, R. G. Chem Res Toxicol 1996, 9, 1194-201.
157

(103) Subbanagounder, G.; Salomon, R. G.; Murthi, K. K.; Brame, C.; Roberts, L. J. J Org Chem 1997, 62, 7658-7666.
(104) Xu, G. Ph.D, Case Western Reserve University, 1998. (105) Bradford, M. M. Anal Biochem 1976, 72, 248-54.
158

Chapter 5
Iso[4]LGE2 Modified Proteins in Trabecular Meshwork of Glaucomatous DBA/2J
Mice
159

5.1. Background
Glaucoma, affects 70 million people worldwide,1 and in one of the leading causes
of irreversible blindness. Glaucoma refers to a group of eye diseases that progressively
steal sight, mostly without any initial symptoms. Primary open angle glaucoma (POAG)
is a form of glaucoma in which there usually is an increase in the intraocular pressure
(IOP), except normal pressure glaucoma, leading to progressive optic nerve damage and
vision loss. Secondary glaucomas may result from an injury, inflammation or tumor2 and
may also occur in advanced cases of cataract, diabetes or as a result of the use of
steroids.3
Increased IOP is a major risk factor for glaucoma. The insidious nature of
pressure-induced changes in the glaucomatous eye makes it difficult to study in humans.
Also, as the disease condition occurs in elderly patients, it is additionally confounded by
other medical complications.4 Several animal models of glaucomas have been developed
that have broadened our understanding of the disease. The diversity from species to
species with regard to differences in the anatomical features, especially, the outflow
angle, has substantially broadened the view on our understanding of anterior segment
diseases, e.g., glaucoma and cataract.4 A detailed discussion of these animal models for
glaucoma is presented in Chapter 1.
5.1.1 Oxidative stress and inflammation pathways. The anterior segment is constantly
exposed to oxidative stress due to the formation of reactive oxygen species (ROS) by
light catalyzed reactions. The increase in ROS can trigger formation of lipid and
160

carbohydrate-derived reactive fragments, leading to oxidative protein modifications,5 and
accumulation of these modified proteins may be pathological.
Lipid Peroxides
NF-кB
IL-18
ROS TNF-α
COX-2LO
Oxidative Stress
Prostaglandins, levuglandins and other lipid peroxidation products
Inflammation
Free radical pathway
Enzyme pathway
Lipid Peroxides
NF-кB
IL-18
ROS TNF-α
COX-2LO
Oxidative Stress
Prostaglandins, levuglandins and other lipid peroxidation products
Inflammation
Free radical pathway
Enzyme pathway
Scheme 5.1. Diagrammatic representation of activation and expression of proteins
involved in inflammation pathway by oxidative stress. Details of the scheme are
discussed in the text. (ROS – reactive oxygen species, COX-2 – cyclooxygenase-2, LO –
lipooxygenase, NF-кB – nuclear factor- kappa B, IL-18 – interleukin 18, TNF – tumor
necrosis factor).
Formation of ROS participate in a positive feedback loop that activates and/or
promotes expression of proteins that are involved inflammatory processes, and which
inturn may be involved in generation of reactive oxygen species (Scheme 5.1). NF-κB, an
161

oxidative stress-responsive transcription factor, is known to be activated, by several
factors including oxidative stress (H2O26,7 or by lipid peroxidation8), to express
proinflammatory cytokines.6 For example, the proinflammatory cytokine TNF-α, may be
induced by oxidative stress directly or through NF-κB.7,9 TNF-α can, in turn, induce
several cascades of activation or inhibition of pathways that are involved in
inflammation, immunity, and cell death.10-12 Interleukin-18 (IL-18), a regulator of innate
and acquired immunity, is involved in a variety of pathways including the release of
cytokines, activation of NF-κB and upregulation of cyclooxygenase-2.13 There are reports
of NF-κB regulating the expression of IL-18.14
5.1.2. Different forms of secondary glaucoma. There are several forms of secondary
glaucoma viz, pigmentary dispersion, traumatic, pseudoexfoliation and neovascular
glaucoma. Pigmentary glaucoma is caused by release of pigments present in the inner
portion of the iris, which get deposited near the trabecular meshwork (TM) region
causing increased resistance to aqueous outflow. Traumatic glaucoma, as the name
suggests, is caused by an injury that has traumatic impact on the eye, causing an increase
in the IOP and causing optic nerve damage. In the case of the pseudoexfoliation
syndrome, thin flaky dandruff-like material, peeling off from the lens, clogs the outflow
facility and causes the IOP to increase. Neovascular glaucoma occurs due to the
formation of blood vessels on the iris and over the drainage canals of the anterior
chamber of the eye (www.glaucoma.org/ learn). There are a number of factors that are
attributed to be bases for the optic nerve degeneration in glaucoma viz, genetic
predisposition,15-19 involvement of receptors and transporters for glutamate,20,21 nitric
162

oxide19 or γ-synuclein22 and of heat shock proteins23 and retinal autoantibodies.24,25
However, detailed molecular mechanisms involved in ganglion cell death are still poorly
understood.26
Pigmentary glaucoma, caused by pigment dispersion syndrome, is one of the
common forms of secondary glaucoma affecting about 0.22 million Americans.13 This is
an open angle form of secondary glaucoma that presents a heavily pigmented TM
towards the end stages. Although use of drugs that lower the IOP of the eye help in
slowing the progression of the disease, but the molecular and biochemical mechanisms
underlying the disease have yet to be elucidated.27
5.1.3. DBA/2J, mouse model for glaucoma. The DBA/2J mouse model for glaucoma
presents several hallmarks of the human pigmentary glaucoma including iris stromal
atrophy (ISA), pigment dispersion (IPD), retinal ganglion loss and optic nerve
cupping.28,29 Iridial pigment dispersal in DBA/2J mice is first clinically evident at around
6-9 months. By 9–10 months iris depigmentation is pronounced and occurs with
increased IOP, resulting in anterior chamber enlargement and glaucoma.28 The
depigmenting iris disease of DBA/2J mice is genetically separable into two distinct
phenotypes, IPD and ISA, which appear to be caused by mutations in the Gpnmb and
Tyrp1 genes respectively. The two missense mutations (Cys110Tyr, Arg326His) in the
tyrosinase related protein-1(TYRP1) gene cause disrupted melanosomes and clumping of
pigments, and a mutation on the stop codon of Gpnmb (Arg150X) causes iris pigmentary
dispersion.30,31 These mutations are suggested to be involved in the leakage of toxic
intermediates of pigment production from the melanosomes causing ISA and IPD.
163

However, the hypopigmented DBA/2J mouse strain does not exhibit iris pigment
dispersion, iris stromal atrophy or glaucoma despite homozygosity for the Gpnmb and
Tyrp1 mutations. Hence, all the factors contributing to elevated IOP in DBA/2J mice
remain to be identified. Nevertheless, DBA/2J mice provide a valuable resource for
defining factors that may contribute to human glaucoma.
Recently, Zhou et. al.13 showed that there is a considerable increase in the
expression of IL-18 mRNA in the iris/ciliary body, and a consequent increase in IL-18
levels in the aqueous humor, of DBA/2J mice with age. This study further showed that
there are increases in the amounts of activated NF-κB and caspase-3. Additionally, they
noted elevation in the expression of matrix metallo-proteinase-1 (MMP-1) and apoptosis
related genes. MMP’s are known to be involved in the degradation of extracellular matrix
proteins in response to cytokines and inflammatory mediators.32,33 Thus, this study
suggests an involvement of inflammation, degradation and apoptosis in the iris/ciliary
body of the DBA/2J mouse model of glaucoma.
Since, oxidative stress plays a role in the regulation of factors involved in the
inflammation process (Scheme 5.1), the study of their levels in the TM of DBA/2J mice
may be important for understanding the changes occurring in the outflow pathway. In this
chapter, the levels of oxidative stress in the TM of DBA/2J mice will be assessed based
on the levels of isolevuglandin-modified proteins.
5.1.4. Iso[4]Levuglandin E2 – formation and pathology. Previously, Dr. Salomon’s lab
discovered that spontaneous rearrangements of prostaglandin endoperoxides generate
levulinaldehyde derivatives, with prostaglandin side chains appended at the carbons α
164

and β to the aldehyde group.34,35 Levuglandins (Scheme 4.1, page 163),36,37 seco
prostanoic acids, are co-generated with prostaglandins (PGs)38,39 by rearrangements of
the endoperoxide PGH2, which occur readily under the free radical-induced and/or
cyclooxygenase -promoted biosynthetic pathway from arachidonic acid (AA).36,40
C5H11
(CH2)3COOH
C5H11
(CH2)3COOHOO
OH
C5H11
(CH2)3COOH
OHCOH
O
Levuglandin E2
Cyclooxygenase
Phospholipase A2
NProtein
NProtein
X
O
Pyrrole Adduct
Lactam (X = H) Hydroxylactam (X = OH)
C5H11
(CH2)3COOPC
OO
(CH2)3COOPC
OH
C5H11
OHC
(CH2)3COOR
OH
C5H11
O
AA-PC
Iso[4] levuglandin E2
Free radical induced
R= H or PC
Other PG's
Scheme 5.2: Generation of levuglandins from arachidonic acid by enzyme mediated and
free radical mechanisms (Detailed scheme presented in Scheme 4.1, page 163).
Iso[4]LGE2 is formed by a free-radical pathway unlike its structural isomer LGE2,
which can be formed through either a free radical or enzyme mediated pathway. These
165

reactive aldehydes bind avidly with proteins forming a protein-bound pyrrole, e.g.,
iso[4]LGE2-pyrrole, as well as protein-protein41,42 and DNA-protein crosslinks.43 We
have also reported mass spectral characterization of several lysine-based modifications
that are generated by covalent adduction of levuglandins with proteins.41 Levels of
levuglandin-protein adducts are markedly elevated in the blood of atherosclerosis and end
stage renal disease patients versus healthy controls.44,45
These highly reactive oxidized lipids, the LGs and isoLGs, form protein
aggregates by cross linking proteins at a very high rate compared to other lipid
peroxidation products viz, HNE and MDA.46 This suggests that they might play a
prominent role in causing oxidative injury. These adducts as well as free LGs have been
shown to hinder the activity of the 20S proteasome, a component of cytosolic protein
degradation machinery, in a dose dependent manner.47 The formation of stable adducts
by LGs and their localization in the TM reflects the localized oxidative stress at TM.
166

5.2. Results
5.2.1. Clinical examination of DBA/2J mice. Examinations of the DBA/2J mice were
performed, as described earlier using a slit lamp, visually as well as by sectioning the eye
post-mortem.28 The sectioning of the eyes was performed transversely for assessing the
TM. The optic nerve was assessed by analyzing both sagital and transverse sections.
Sagital sections pass through the longitudinal axis of the eye from anterior to the
posterior regions, while the transverse sections are perpendicular to the sagital sections
(Figure 5.1). Haematoxylin stained sections were used as reference to locate the TM
region.
Figure 5.1: Diagrammatic illustration of sagittal and transverse sections of the eye. The
sagital section passes through the longitudinal axis of the eye from the anterior to the
posterior region. The transverse section passes perpendicularly axis to the sagittal axis.
Pigment dispersion. DBA/2J mice develop a form of glaucoma with similarities to human
pigmentary dispersion syndrome. A visual inspection of the ~ 6-month-old DBA/2J
mouse eye clearly showed the signs of degenerating iris with pigment dispersed across
167

the pupil (Figure 5.2A). By 8-9 months, iris depigmentation is pronouncedly increased
(Figure 5.2C) and elevated IOP has developed, resulting in anterior chamber enlargement
and glaucoma.
A B CA B C
Figure 5.2. Anterior segment assessment of DBA/2J mice. The anterior segment pictures
were taken with a Leica MZ stereomicroscope (Leica Microsystems Inc., Bannockburn,
IL) equipped with a SpotCam RT KE digital camera (Sterling Heights, MI). A. 6-month-
old DBA/2J mouse, B. 8-month-old and C. 14-month-old. Arrows show the iris
degeneration (A), heavy prigmentation (B) and complete iris degeneration and
depigmentation (C).
Optic nerve damage. The optic nerve sections of 8-month DBA/2J and C57BL6 mice
were performed using both sagital as well as transverse sections. The sections were
stained with haematoxylin stain. The longitudinal section shows the optic cupping
feature, reflecting the optic nerve excavations (arrow in Figure 5.3A), which is a feature
of the optic nerve degeneration compared with the normal optic nerve in Figure 5.3B.
The cross sections across the optic nerve show a distinct difference in the myelination of
the nerve fibres, a characteristic of optic neuropathy. Figure 5.3C shows disorganized
nerve fibres (arrows) compared to the normal control in Figure 5.3D.
168

DBA/2J C57BL6
A B
C D
DBA/2J C57BL6
A B
C D
DBA/2J C57BL6
A B
C D
Figure 5.3. Histochemical assessment of mouse optic nerve. The mouse eye sections (10
µm) passing though optic nerve were stained with hematoxylin. A. Sagittal section of 8
months old DBA/2J optic nerve (x 20) and C. a transverse section of DBA/2J optic nerve
(x 100) B. Sagittal section of age matched C57BL6 control optic nerve (x 20) and D. a
transverse section of age matched C57BL6 control optic nerve (x 100). Arrows denote
the optic nerve excavations (A) and demylenation of the nerve fibres (C).
5.2.2. Increase in levels of iso[4]LGE2 modified proteins in DBA/2J with age. At age
8-10 weeks, elevated IOP is typically exhibited by DBA/2J mice in contrast to C57BL6
controls. The TM of DBA/2J mice was isolated surgically and the proteins were extracted
immediately.
169

A
B
2.5
mon
ths
8 m
onth
sD
BA/
2J
C57
BL6
2.5
mon
ths
8 m
onth
s
BS
A-Is
o[4]
LGE
2
11898
56
37
29
20
11898
56
37
29
20
0
10
20
30
40
50
2.5
mnt
h
8 m
nth
2.5
mnt
h
8 m
nth
Rel
ativ
e in
tens
ity (%
)
C57BL6
DBA/2J
CA
B
2.5
mon
ths
8 m
onth
sD
BA/
2J
C57
BL6
2.5
mon
ths
8 m
onth
s
BS
A-Is
o[4]
LGE
2
11898
56
37
29
20
11898
56
37
29
20
11898
56
37
29
20
11898
56
37
29
20
0
10
20
30
40
50
2.5
mnt
h
8 m
nth
2.5
mnt
h
8 m
nth
Rel
ativ
e in
tens
ity (%
)
C57BL6
DBA/2J
0
10
20
30
40
50
2.5
mnt
h
8 m
nth
2.5
mnt
h
8 m
nth
Rel
ativ
e in
tens
ity (%
)
C57BL6
DBA/2J
C
Figure 5.4. Western analysis of DBA/2J TM proteins with iso[4]LGE2 antibodies. TM
proteins (~5 µg/lane) were subjected to 1D SDS/PAGE, electroblotted onto a
poly(vinylidene difluoride) membrane, and probed with anti-iso[4]LGE2 antibodies.
Bovine serum albumin modified with iso[4]LGE2 was used as a positive control. A.
Immunoblot of DBA/2J and C57BL6 TM proteins. B. Coomassie stained immunoblot.C.
Densitometric quantification of the Western blot (n=1), see experimental section p. 182
for details.
170

As described previously for our studies of oxidative stress in human TM (chapter
4), we used iso[4]LGE2-protein adduct as a marker for oxidative stress. We probed the
TM extract of DBA/2J mice for iso[4]LGE2-derived protein modifications by Western
analyses using anti-iso[4]LGE2 polyclonal antibodies (pAb). The immunoblots showed
the presence of background levels of some modified proteins at 3 weeks to 5 months old
DBA/2J mice and in control mice of comparable ages. Nevertheless, there is a significant
difference in the number and intensity of immunoreactive bands at 8 months in TM of
DBA/2J compared to controls (Figure 5.4). Subsequent time course analyses in DBA/2J
revealed the presence of iso[4]LGE2 modification prominently at 8 weeks of age (see
Appendix). There is intense staining for iso[4]LGE2 modification in the high molecular
weight region of the SDS-PAGE.
5.2.3. Immunohistochemical localization of iso[4]LGE2 modified proteins. To
visualize the localization of iso[4]LGE2 modified proteins in the limbus region of the
DBA/2J mice, a transverse section of the anterior segment was analyzed. The anti-
iso[4]LGE2 antibody stained section showed the presence of immunoreactivity in the TM
in 6 out of 8 slides analyzed (4 DBA/2J and 4 C57BL6 mice were used), while
comparable sections of age-matched C57BL6 mice showed much less staining.
Additionally, serial sections of the slides mentioned above were also stained with pre-
immune antibodies, that served as controls. The green channel was used to detect the
interfering autofluorescence of the sections. Nuclear staining was performed using 4',6-
diamidino-2-phenylindole (DAPI).
171

Iso[4]LGE2 Pre-Immune
DBA/2J C57BL6
Iso[4]LGE2
A
C D
E
F
B
Iso[4]LGE2 Pre-Immune
DBA/2J C57BL6
Iso[4]LGE2
A
C D
E
F
B
Figure 5.5. Histochemical localization of isoLGE2 modified proteins in TM.
Representative immunohistochemical analyses with a rabbit polyclonal anti-isoLGE2 are
shown. Rhodamine conjugated secondary antibody with red fluorescence (Alexa 594)
was used for detection of iso[4]LGE2-modified proteins. A. DBA/2J mice TM, 8 months
(green channel) B. DBA/2J mice TM, 8 months(red channel) C. C57BL6 mice TM,
8months (green channel). D. C57BL6 mice TM, 8 months (red channel). Red channel –
immunofluorescence of iso[4]LGE2 modified proteins; green channel – autofluorescence.
5.2.4. Immunoprecipitation of iso[4]LGE2-modified proteins. To identify the proteins
that were modified by iso[4]LGE2, protein extracts from the TM were immunoprecipitat-
ed using rabbit anti-iso[4]LGE2-pAb linked to protein A sepharose beads. The beads
were prepared using protocols described in Chapter 4. Each of the IP experiments utilized
172

protein extracts from TM of both eyes from one DBA/2J mouse. The Coomassie blue
stained gels showed the protein bands that were immunoprecipitated as well as the long
chain of the IgGs. As mentioned in Chapter 4, The bands detected at ~55 kDa that is
common to both the lane 2 and 4 (shown with dotted lines in Figure 5.6) corresponds to
the long chain of IgG originating from the cleavage of IgG chains from the antibody-
coupled beads as reported previously.48
11898
56
37
29
20
IgG
Lane 1 2 3 4 5
11
2
34
56
11898
56
37
29
20
IgG
Lane 1 2 3 4 5
11
2
34
56
Figure 5.6: Anti-iso[4]LGE2 immunoprecipitation of TM proteins from an 8 month old
DBA/2J mouse. Immunoprecipitations (IPs) utilized antibody coupled beads (10 µg) and
TM protein extracts (10 µg). They were analyzed by SDS-PAGE and mass spectrometry.
A Coomassie blue stained gel is shown: lane 1, molecular weight markers; lane 2,
antibody coupled beads without TM; lane 3, TM protein extract (5 µg); lane 4, anti-
iso[4]LGE2 IP products; lane 5, supernatant containing unbound proteins that did not
immunoprecipiate. Bands corresponding to proteins excised and identified in Table 5.1
are numbered 1-6.
173

5.2.5. Identification of modified proteins using LC-MS/MS. Identification of the
immunoprecipitation products was performed as described earlier.
Table 5.1: Proteins immunoprecipitated from TM extract of DBA/2J mice using
iso[4]LGE2-pAb. Gel slices from top to the bottom of lane 4 of the gel in Figure 5.6 were
digested using trypsin and the proteins identified using LC-MS/MS and MassLynx™
software with the Swissprot database.
Proteins Lysine
Content(%)
BandNo.
Peptides Matched
Calculated1 MW(kDa)
Observed MW(kDa) AccessionNo.
Actin 5.0 3 1 42 120 P68134
Calcium/calmodulin-dependent
phosphodiesterase 6 5 1 64 67 Q61481
Collagen alpha 1(I) 3.8 1 1 137 ≥2204 P11087
F-box/LRR-repeat protein 20 4.4 1 1 48 ≥2204 Q9CZV8
GFAP2 4.7 1 1 50 ≥2204 P03995
Hemoglobin 7.8 1 4 15 ≥2204 P01942
Histone H1 27 1,5 1 21 67, ≥2204 P15864
Histone H2 10 1,5 1 14 67, ≥2204 P22752
Keratin, cytoskeletal 3.7 6 5 58 65 P02535
Liprin-beta 2 6.8 4 1 98 98 O35711 PI4K3 5.2 2 1 90 170 Q8BKC8
Serum albumin 8.4 5 1 68 68 P07724
Alpha-tubulin 1 4.2 2,5 2 50 170 P68369
Enamelin precursor 4.0 3 1 128 120 O97939
1 For monomeric protein 2GFAP – Glial acidic fibrillary protein; 3PI4K - Phosphatidylinositol 4-kinase; 4
High molecular weight bands at the top of the gel
174

The proteins identified included extracellular matrix proteins as well as some proteins
from blood. There were fewer proteins detected from IP of DBA/2J TM extracts than
from IP of human TM proteins probably due to the lesser amount of IP proteins in mouse
TM than from the human TM. However, a lack in overlap between the proteins identified
from DBA/2J and human was not completely understood. The observed molecular weight
is an estimate based on SDS PAGE and the molecular weight maker positions using the
Digitizer® function of the program Origin® version 7.5.
175

5.3. Discussion
The importance of oxidative stress as a contributing factor in the progression of
glaucoma has been previously hypothesized. This study was directed towards
understanding the pathological consequences of oxidative stress in glaucoma.49 Animal
models to study the basic aspects of the disease have benefits, both in terms of time and
effort. DBA/2J has been well established as mouse model of glaucoma with a well-
defined period of increase in the intraocular pressure followed by irreversible damage to
the optic nerve. Herein we sought to determine whether there is an increase in the levels
of oxidative damage in the DBA/2J mouse TM concurrently with damage of the optic
nerve.
Western analyses of the TM extracts from DBA/2J mice at different ages
compared to age matched control C57BL6 mice using iso[4]LGE2 antibodies shows a
distinct increase in the amount of immunoreactive bands in older DBA/2J mice. Also, of
note, a number of immunoreactive bands appear in the higher molecular weight region of
the SDS PAGE. This indicates the presence of cross-linked proteins. Since the amount of
protein in the higher molecular weight region of the Coomassie stained gel appeared to be
very low (Figure 5.4), we concluded that the identification of proteins from this 1D
PAGE would not be easy. So, to concentrate and characterize the iso[4]LGE2-modified
protein, we performed immunoprecipitation with isoLGE2 antibody coupled protein A
sepharose beads. Iso[4]LGE2 immunoprecipitation of the DBA/2J mouse TM extract
enriched some proteins compared to that of the total protein extract (Figure 5.6). A lack
of iso[4]LGE2-modified proteins was demonstrated in C57BL6 and the putative
iso[4]LGE2-modified proteins were not immunoprecipitated with preimmune rabbit
176

serum (see Appendix 5.1S). The immunoprecipitated proteins from DBA/2J mice (n=5)
examined were identified by using LC-MS/MS and are listed in the Table 5.1.
Because they appear as oligomers on a denaturing PAGE gel (Figure 5.4 and
Table 5.1), many of the proteins from the IP may be crosslinked. Some of the
immunoprecipitated proteins that were identified are actin, collagen I, liprin, keratin
(cytoskeletal) and tubulin, which are involved in the structural organization of the TM.
The structural protein actin (band 3, Figure 5.4) appears at ~120 kDa, while it’s
molecular weight is 42 kDa. This implies that it may be crosslinked with itself or other
proteins. Collagen I, is also a structural protein present in TM. There are reports that the
levels of collagen I increase with hydrostatic pressure in cell cultures,50 a model of
increased intraocular pressure (IOP). It is tempting to speculate that the increased IOP
associated with glaucomas promote lipid oxidation and consequent protein
oligomerization and accumulation. Both Liprin-β2, which functions as a cytoskeletal
anchoring protein, and GFAP, a class-III intermediate filament in astrocytes,51 appear to
be modified. GFAP appears to be extensively oligomerized.
Modification of nuclear proteins may have profound effects on the molecular
processes that are involved in cell division and cell differentiation. Tubulin is a major
constituent of microtubules that is involved in mitosis. Dr. Salomon’s lab has previously
shown that this protein can be crosslinked, by exogenous LGE2, and LGE2 also inhibits
microtubule assembly in vitro and cell division in vivo.52 In the present mouse model of
glaucoma, we found tubulin to be crosslinked, possibly as a trimer. Histones are nuclear
proteins that are associated with DNA and play an important role in structural
organization of nucleosomes and regulation of transcription. Histones are highly enriched
177

with lysine residues (10-27%), which are targets for LGs. In the IP, histones (H1 and H2)
appear in the very high molecular region (band 1 and 2, Figure 5.6); possibly, due to high
lysine content that makes them prone to extensive crosslinking. Previously, LGs were
shown to crosslink DNA in vivo with protein that is presumably histones.43,53
Another protein that appears to be modified extensively (perhaps oligomerized) is
the F-box protein Fbx2, which binds specifically to proteins attached to N-linked high-
mannose oligosaccharides and subsequently contributes to ubiquitination of N-
glycosylated proteins.54 This implies that this protein is involved in the substrate
recognition of ubiquitin-proteosomal degradation machinery and it’s modification can
possibly lead to cellular instability.55
Phosphoinsitol 4-kinase (PI4K) plays a vital role in cell growth and is expressed
in higher amounts during cell stress.56 The IP also shows that PI4K (band 2, Figure 5.6) is
significantly modified and is perhaps crosslinked as a dimer.
Preliminary results using Western blot analysis using anti-proteasomal antibody
that recognizes the 23 kDa representing proteasome 20S also suggested that the
proteasome is also modified in the TM of 8-month-old DBA/2J mice (Figure 5.7). It
appears as a rather narrow band with an apparent molecular weight of 120 kDa, which
suggests a pentamer. Earlier reports of chronic inhibition of proteasome in TM cultures in
response to oxidative stress has been noted.57
178

2.5
mon
ths
8 m
onth
s
11898
56
37
29
20
2.5
mon
ths
8 m
onth
s
Iso[4
]LGE 2
Prot
easo
me
Western Blots
2.5
mon
ths
8 m
onth
s
11898
56
37
29
20
11898
56
37
29
20
2.5
mon
ths
8 m
onth
s
Iso[4
]LGE 2
Prot
easo
me
Western Blots
Figure 5.7. Proteasome crosslinked in 8-month-old DBA/2J mice. A. Western blot using
iso[4]LGE2 polyclonal antibody (portion of Figure 5.4). B. The immunoblot in Figure
5.4A was probed with anti-rabbit proteasome polyclonal antibody that detects the 23 kDa
representing the proteasome 20S (arrow shows the position of high molecular weight
band immunoreactive for the proteasome antibody).
These studies suggest that: (1) there is an increase in the oxidative stress levels of
the glaucomatous DBA/2J mice and (2) the structural proteins are modified extensively.
The oxidative modifications that we detected are those produced through a free radical
pathway. Further studies are needed to establish whether free radical and enzyme
catalyzed lipid oxidation are important in glaucoma pathogenesis.
179

5.4. Experimental procedures
5.4.1. Tissue procurement. DBA/2J and C57BL/6 control mice were obtained (Jackson
ImmunoResearch Laboratories Inc., West Grove, PA) and bred in the Cole Eye Institute
animal facility. Trabecular meshwork tissues were excised from these mice at different
stages of their lifetime in accordance with the animal procedures approved by CCF
IACUC and stored at -80º C. Whole eyes from these mice were removed and fixed in 4%
formaldehyde solution and embedded using standard protocols for immunohistochemical
analysis.
5.4.2. Protein extraction. The excised TM tissue was incubated on ice in 50 µL of 100
mM Tris-Cl (pH 7.8) containing 8 mM SnCl2, 50 mM NaHPO4, 5 mM DTT
(dithiothreitol), 1mM DTPA (diethlenetriaminepentacetic acid), 100 mM BHT (butylated
hydroxytoluene) and 0.5% SDS (sodium dodecylsulphate) for 30 minutes. The tissue was
homogenized using a hand held homogenizer. The solution was subsequently centrifuged
at 8K rpm for 2 min. The concentration of the soluble protein fraction was determined
using the Bradford method 58 and amino acid quantified BSA (National Bureau of
Standards) as standard. The solution were stored at -80 ºC until analyzed.
5.4.3. Western analysis. Electrophoretic separation of protein extracts (~5 µg) from TM
tissue, in Laemmli sample buffer59, was performed on a 10% SDS PAGE with a 4%
stacking gel using the Bio-Rad Mini-Protein II system following standard protocols.60
The gel was electroblotted onto a polyvinyl difluoride (PVDF) membrane (Millipore,
Bedford, MA), and probed with anti-iso[4]LGE2 polyclonal antibodies (pAb)
180

antibodies.45 To evaluate whether equal amounts of protein were used for Western
analyses, PVDF membranes were stained with Coomassie blue post-transfer. Western
analyses were performed with antibodies to modifications of proteins by iso[4]LGE245
and anti-rabbit proteasome polyclonal antibody. The anti-proteasome polyclonal antibody
that detects the 23 kDa proteasome subunit was obtained from Abcam Ltd (Cambridge,
MA). Iso[4]LGE2 modified BSA was used as positive control. The immunoreactive bands
were quantified using GS-710 Densitometer scanner and QuantityOne ® software from
Biorad™ (Hercules, CA).
5.4.4. Immunohistochemistry. Eyes fixed in 4% paraformaldehyde were cryoprotected
by incubating them sequentially in 10%, 20% and 30% sucrose for 1 h each until the eyes
completely sank. The eyes were then incubated in 30% sucrose in O.C.T. embedding
medium for 15 min. The eyes were quickly embedded into O.C.T. medium on liquid
nitrogen. After the O.C.T. solidified, the eyes were stored in a -20 ºC freezer only to be
removed when ready to section onto slides. The sections (7-10 µm) were then placed onto
microscopic slides. The sections were incubated in phosphate buffered saline (PBS)
solution followed by a solution of 100 mM glycine in PBS (pH 7.4) for 1 h each. The
tissues were then covered with the antibody to be probed at 1:100 dilution in PBS for 1 h.
The tissue was then washed thrice with PBS solution for 10 min each, and then incubated
in 10 ng secondary antibody conjugated with Alexa 594 or Alexa 488 (Molecular Probes
Inc, Eugene, OR) for 1 h. The sections were sealed with Vectashield® Mounting Medium
(H-1000) and analyzed with a Nikon EFD-3 fluorescence microscope attached to a CCD
181

camera. Hematoxylin stained slides were used to locate the TM in the sections. The
microscopic images were assembled in Adobe® Photoshop CS.
5.4.5. Immunoprecipitation. Antibody-coupled protein A beads , prepared using
suitable modifications of standard protocols,61 were used for all immunoprecipitations.
About 100 µg of swelled protein A sepharose CL-4B beads (Amersham Pharmacia
Biotech, CA) was coupled with 100 µg of antibody using 4 x 25 mg
dimethylpimelimidate, each time with a 2 h incubation at room temperature for each
aliquot, in 50 mM sodium borate buffer pH 8.3. Following incubation, the beads were
washed, and then incubated for 2 h with 200 mM ethanolamine pH 8.0. Beads (20 µg
antibody-coupled beads) were finally washed in phosphate buffered saline pH 7.4 and
incubated with protein extracts (~20 µg) prepared in 100 mM Tris-Cl buffer pH 7.5, 50
mM NaCl and 0.01% genapol. After incubation, the beads were recovered by
centrifugation at 2500 x g for 5 minutes and washed 3 x 500 µl of 100 mM Tris-Cl buffer
pH 7.5, 100 mM NaCl and 0.02% genapol. The beads were boiled with 30 µL Laemmli
buffer59 for 2 min, and then separated on a 10% SDS-PAGE. The gels were subjected to
staining with Coomassie stain and subsequent LC MS/MS of in-gel tryptic digested
proteins bands or Western analyses.
5.4.6. LC MS/MS analysis. Following electrophoresis, the gel bands were excised in
1mm slices and destained and processed using standard protocol for in-gel digestion 60 in
a 96-well plate. The in-gel tryptic digested proteins were subjected to LC-MS/MS
analysis using a quadrupole-time of flight (QToF2) mass spectrometer equipped with a
182

CapLC system (Micromass, Milford, MA), ProteinLynx™ Global Server acquisition and
processing software. Peptide digests were trapped on a precolumn (0.3 x 1mm, 5µC18,
LC Packing) with 0.1% formic acid in 2% acetonitrile as loading solvent, and then eluted
onto a capillary column (PicoFrit 0.050 x 50mm, 5µ tip ID, New Objective Inc).
Chromatography was performed with a flow of 250 nl/min with aqueous
acetonitrile/formic acid solvents and 100% of the eluant was directed towards the mass
spectrometry. The mass spectrometer was operated in standard MS/MS switching mode
with the three most intense ions in each survey scan subjected to MS/MS analysis.
183

5.5. References
(1) Quigley, H. A. Br J Ophthalmol 1996, 80, 389-93. (2) http://www.glaucoma.org/learn/; Vol. 2005. (3) Oyster, C. W. The Human Eye Structure and Function; Sinauer Associates, Inc.:
Sunderland, Massachusetts., 1999. (4) Chew, S. In The Glaucomas: Basic Sciences, Anatomy and Pathology; 2 ed.;
Krupin, T., Ed.; Mosby: New York, 1996; Vol. 1. (5) Green, K. Ophthalmic Res 1995, 27 Suppl 1, 143-9. (6) Li, N.; Karin, M. Faseb J 1999, 13, 1137-43. (7) Schreck, R.; Rieber, P.; Baeuerle, P. A. Embo J 1991, 10, 2247-58. (8) Bowie, A. G.; Moynagh, P. N.; O'Neill, L. A. J Biol Chem 1997, 272, 25941-50. (9) Verhasselt, V.; Goldman, M.; Willems, F. Eur J Immun 1998, 28, 3886-3890. (10) Liao, F.; Andalibi, A.; deBeer, F. C.; Fogelman, A. M.; Lusis, A. J. J Clin Invest
1993, 91, 2572-9. (11) Liao, F.; Andalibi, A.; Qiao, J. H.; Allayee, H.; Fogelman, A. M.; Lusis, A. J. J
Clin Invest 1994, 94, 877-84. (12) Finkel, T.; Holbrook, N. J. Nature 2000, 408, 239-47. (13) Zhou, X.; Li, F.; Kong, L.; Tomita, H.; Li, C.; Cao, W. J Biol Chem 2005, 280,
31240-8. (14) Chandrasekar, B.; Colston, J. T.; de la Rosa, S. D.; Rao, P. P.; Freeman, G. L.
Biochem Biophys Res Commun 2003, 303, 1152-8. (15) Mandelkorn, R. M.; Hoffman, M. E.; Olander, K. W.; Zimmerman, T. J.; Harsha,
D. Ophthalmic Paediatr Genet 1985, 6, 325-31. (16) Stone, E. M.; Fingert, J. H.; Alward, W. L. M.; Nguyen, T. D.; Polansky, J. R.;
Sunden, S. L. F.; Nishimura, D.; Clark, A. F.; Nystuen, A.; Nichols, B. E.; Mackey, D. A.; Ritch, R.; Kalenak, J. W.; Craven, E. R.; Sheffield, V. C. Science 1997, 275, 668-670.
(17) Fingert, J.; Heon, E.; Liebmann, J.; Yamamoto, T.; Craig, J.; Rait, J.; Kawase, K.; Hoh, S.; Buys, Y.; Dickinson, J.; Hockey, R.; Williams-Lyn, D.; Trope, G.; Kitazawa, Y.; Ritch, R.; Mackey, D.; Alward, W.; Sheffield, V.; Stone, E. Hum Mol Genet 1999, 8, 899-905.
(18) Stoilov, I. R.; Costa, V. P.; Vasconcellos, J. P.; Melo, M. B.; Betinjane, A. J.; Carani, J. C.; Oltrogge, E. V.; Sarfarazi, M. Invest Ophthalmol Vis Sci 2002, 43, 1820-7.
(19) Vincent, A. L.; Billingsley, G.; Buys, Y.; Levin, A. V.; Priston, M.; Trope, G.; Williams-Lyn, D.; Heon, E. Am J Hum Genet 2002, 70, 448-60.
(20) Liu, B.; Neufeld, A. H. Arch Ophthalmol 2001, 119, 240-5. (21) Lam, T. T.; Abler, A. S.; Kwong, J. M.; Tso, M. O. Invest Ophthalmol Vis Sci
1999, 40, 2391-7. (22) Surgucheva, I.; McMahan, B.; Ahmed, F.; Tomarev, S.; Wax, M. B.; Surguchov,
A. J Neurosci Res 2002, 68, 97-106. (23) Tezel, G.; Hernandez, R.; Wax, M. B. Arch Ophthalmol 2000, 118, 511-8. (24) Ikeda, Y.; Ohguro, H.; Maruyama, I. Jpn J Ophthalmol 2000, 44, 648-52. (25) Schori, H.; Kipnis, J.; Yoles, E.; WoldeMussie, E.; Ruiz, G.; Wheeler, L. A.;
Schwartz, M. Proc Natl Acad Sci U S A 2001, 98, 3398-403.
184

(26) Dyka, F. M.; May, C. A.; Enz, R. J Neurochem 2004, 90, 190-202. (27) Jacobi, P. C.; Dietlein, T. S.; Krieglstein, G. n. K. Ophthalmology 2000, 107, 417-
421. (28) John, S. W.; Smith, R. S.; Savinova, O. V.; Hawes, N. L.; Chang, B.; Turnbull,
D.; Davisson, M.; Roderick, T. H.; Heckenlively, J. R. Invest Ophthalmol Vis Sci 1998, 39, 951-62.
(29) Schuettauf, F.; Quinto, K.; Naskar, R.; Zurakowski, D. Vision Res 2002, 42, 2333-7.
(30) Anderson, M. G.; Smith, R. S.; Hawes, N. L.; Zabaleta, A.; Chang, B.; Wiggs, J. L.; John, S. W. Nat Genet 2002, 30, 81-5.
(31) Chang, B.; Smith, R. S.; Hawes, N. L.; Anderson, M. G.; Zabaleta, A.; Savinova, O.; Roderick, T. H.; Heckenlively, J. R.; Davisson, M. T.; John, S. W. Nat Genet 1999, 21, 405-9.
(32) Ishida, Y.; Migita, K.; Izumi, Y.; Nakao, K.; Ida, H.; Kawakami, A.; Abiru, S.; Ishibashi, H.; Eguchi, K.; Ishii, N. FEBS Lett 2004, 569, 156-60.
(33) Zhang, B.; Wu, K. F.; Cao, Z. Y.; Rao, Q.; Ma, X. T.; Zheng, G. G.; Li, G. Leuk Res 2004, 28, 91-5.
(34) Salomon, R. Acc Chem Res 1985, 18, 294 (35) Salomon, R. G.; Miller, D. B.; Zagorski, M. G.; Coughlin, D. J. J Am Chem Soc
1984, 106, 6049. (36) Salomon, R. G.; Miller, D. B. Adv Prostaglandin Thromboxane Leukot Res 1985,
15, 323-6. (37) Morrow, J. D.; Harris, T. M.; Roberts, L. J., 2nd Anal Biochem 1990, 184, 1-10. (38) Hamberg, M.; Svensson, J.; Samuelsson, B. Proc Natl Acad Sci USA 1974, 71,
3824-3828. (39) Nugteren, D.; Hazelhof, E. Biochim Biophys Acta 1973, 326, 448-61. (40) Nugteren, D.; Christ-Hazelhof, E. Adv Prostaglandin Thromboxane Res 1980, 6,
129-37. (41) Brame, C. J.; Salomon, R. G.; Morrow, J. D.; Roberts, L. J., 2nd J Biol Chem
1999, 274, 13139-46. (42) Foreman, D.; Zuk, L.; Miller, D. B.; Salomon, R. G. Prostaglandins Other Lipid
Mediators 1987, 34, 91-8. (43) Murthi, K. K.; Friedman, L. R.; Oleinick, N. L.; Salomon, R. G. Biochemistry
1993, 32, 4090-7. (44) Salomon, R. G.; Subbanagounder, G.; O'Neil, J.; Kaur, K.; Smith, M. A.; Hoff, H.
F.; Perry, G.; Monnier, V. M. Chem Res Toxicol 1997, 10, 536-45. (45) Salomon, R. G.; Batyreva, E.; Kaur, K.; Sprecher, D. L.; Schreiber, M. J.; Crabb,
J. W.; Penn, M. S.; DiCorletoe, A. M.; Hazen, S. L.; Podrez, E. A. Biochim Biophys Acta 2000, 1485, 225-35.
(46) Iyer, R. S.; Ghosh, S.; Salomon, R. G. Prostaglandins 1989, 37, 471-80. (47) Davies, S. S.; Amarnath, V.; Montine, K. S.; Bernoud-Hubac, N.; Boutaud, O.;
Montine, T. J.; Roberts, L. J., 2nd Faseb J 2002, 16, 715-7. (48) Doolittle, M. H.; Ben-Zeev, O.; Briquet-Laugier, V. J Lipid Res 1998, 39, 934-
942. (49) Chen, J. Z.; Kadlubar, F. F. Am J Med 2003, 114, 697-8. (50) Yang, J.; Neufeld, A.; Zorn, M.; Hernandez, M. Exp Eye Res 1993, 56, 567-74.
185

(51) Nielsen, A. L.; Jorgensen, A. L. J Biol Chem. 2004, 279, 41537-41545. (52) Murthi, K. K.; Salomon, R. G.; Sternlicht, H. Prostaglandins 1990, 39, 611-22. (53) Boutaud, O.; Andreasson, K.; Zagol-Ikapitte, I.; Oates, J. Brain pathol (Zurich,
Switzerland) 2005, 15, 139-42. (54) Yoshida, Y.; Chiba, T.; Tokunaga, F.; Kawasaki, H.; Iwai, K.; Suzuki, T.; Ito, Y.;
Matsuoka, K.; Yoshida, M.; Tanaka, K.; Tai, T. Nature (London) 2002, 418, 438-42.
(55) Ciechanover, A.; Brundin, P. Neuron 2003, 40,427-46. (56) Audhya, A.; Emr, S. Dev Cell 2002, 2, 593-605. (57) Caballero, M.; Liton, P. B.; Epstein, D. L.; Gonzalez, P. Biochem Biophys Res
Commun 2003, 308, 346-52. (58) Bradford, M. M. Anal Biochem 1976, 72, 248-54. (59) Laemmli, U. K. Nature 1970, 227, 680-5. (60) Miyagi, M.; Sakaguchi, H.; Darrow, R. M.; Yan, L.; West, K. A.; Aulak, K. S.;
Stuehr, D. J.; Hollyfield, J. G.; Organisciak, D. T.; Crabb, J. W. Mol Cell Proteomics 2002, 1, 293-303.
(61) Harlow, E.; Lane, D. Antibodies: a laboratory manual; Cold Spring Harbor Laboratory, 1988.
186

Chapter 6
Part A: Pilot Studies Towards Identification of Levuglandin Modified Proteins in
Macrophages
Part B: Initial Studies Towards Developing a Model System to Differentiate Enzyme
Mediated and Free- Radical Mediated Formation of Levuglandins
187

6.1. Background
6.1.1. Macrophages, inflammation and atherosclerosis. Monocytes present in the
blood are progenitors of macrophages. Insults to the endothelial layer of the blood
vessels, such as hypercholesterolemia or cytokines, can stimulate the production of
leukocyte chemo-attractant molecules and expression of adhesion molecules that bind to
the monocytes and make them adhere to the endothelial layer through integrins (Figure
6.1).1 A family of low molecular weight chemotactic cytokine proteins, called
chemokines, e.g., monocyte chemoattractant protein-1 (MCP-1), recruit the monocytes to
the site of inflammation.2 Once the monocytes are inside the arterial intima, they are
differentiated into macrophages through the influence of the cytokine, macrophage-
colony stimulating factor (MCS-F).3 Macrophages are involved in both specific
immunity, e.g., engulf and degrade foreign material, as well as non-specific immunity,
e.g., process and present antigens to immune cells.
A crucial role of macrophages in atherosclerosis was established through a mouse
model that is deficient for both apoE and MCS-F, which showed dramatic protection
from atherosclerosis.4 Macrophages can be activated by cytokines, including interferon
(INF) γ and tumor necrosis factor (TNF) α. Furthermore, activated macrophages can
promote the production of cytokines.5 It has been shown that apoE deficient mice lacking
the INFγ receptor, which down regulates expression of the ABCA-1 receptor and
consequently inhibits cholesterol efflux from the macrophages. This is proatherogenic,
because it accelerates the formation of foam cells.6 Altered cholesterol homeostasis is
critical factor in foam cell formation as the macrophage internalizes large amounts of
cholesterol derived from lipoproteins. Uptake of oxidized or modified lipoproteins occurs
188

through scavenger receptors expressed on macrophages. Components of oxLDLs are
degraded and processed by macrophages to present the antigens present in them and
additionally, these oxLDL can act as activators of the macrophages itself. For example,
the oxLDL may contain strongly proinflammatory platelet activating factor (PAF)-like
molecules which can activate the macrophages as well as endothelial cells.7 A role for
oxLDL in the atherosclerosis is discussed in the following sections of this chapter.
igure 6.1: Schematic description of one of the factors contributing to foam cell
LDL
monocytes
macrophage oxLDL
Foam cell
Lum
enVe
ssel
wal
l
Endothelial layerradicals
peroxides
LDL
monocytes
macrophage oxLDL
Foam cell
Lum
enVe
ssel
wal
l
Endothelial layerradicals
peroxides
F
formation through endocytosis of oxLDL by macrophages.
189

6.1.2. Macrophages and COX-2. Cyclooxygenase enzymes are involved in the
rmal
X-1,
n
itial studies of atherogenic processes using non specific inhibitors of COX
enzyme be
s,
.1.3. Lipopolysaccharides (LPS) and cyclooxygenase (COX) expression. LPS is a
s
ation
formation, from fatty acids (vide supra), of eicosanoids, that are involved in the no
physiological functions. They are known to modulate physiological processes in
atherosclerosis and thrombosis. Two-isoforms of the enzymes are known viz, CO
which is constitutively expressed in most human tissues,8 and COX-2, whose expressio
is readily induced by inflammatory stimuli such as LPS and cytokines in a variety of
cells.9,10
In
s showed its anti-atherogenic effects in animal models.11 COX-2 is known to
upregulated in activated macrophages through proinflammatory agents like IL-1, TNFα,
LPS and TGFβ. COX-2 mediated production of PGs by activated macrophages in arterial
wall has been linked to atherosclerosis through a number of mechanisms including
activation of chemotaxis, propagation of the inflammatory cascade through cytokine
and induction of vascular permeability.11 More direct evidence for the role of COX-2 in
atherosclerosis was provided by a study using an LDL receptor negative (LDLR-/-) mouse
that is deficient in the COX-2 gene had significantly fewer atherosclerotic lesions
compared an LDLR-/- mouse with the COX-2 gene.12
6
structural element of Gram-negative bacterial cell wall consisting of lipids and polymer
of carbohydrates. LPS is an endotoxin, which acts as a ligand for receptors that can
trigger proinflammatory cytokines in many cell types.13 Among the other systemic
effects that are caused by LPS are the generation of secondary mediators of inflamm
190

viz, prostaglandins (PGs) and platelet activating factors (PAFs).14 In the case of
macrophages, LPS stimulation causes the secretion of PGs (mainly PGE2) and
proinflammatory cytokines, which, in turn, act in an autocrine or paracrine man
regulate the host response.
ner to
in the synthesis of PGs by LPS stimulation is the enzyme
cycloox
to
.1.4. Distinguishing levuglandins and isolevuglandins. Cyclooxygenation of
des
2,
rm
α, β-
.
15
The rate-limiting step
ygenase (COX).16 COX-2 expression in response to LPS in human and mouse
macrophages has been reported to be induced via NF-κB, indicating a transcriptional
mechanism for this potent stimulus.17 LPS-treated macrophages have also been shown
release the proinflammatory cytokines IL-1ß and TNF-α.15
6
polyunsaturated fatty acids (PUFAs) is known to generate prostanoid endoperoxi
through enzymatic or non-enzymatic processes. The prostaglandin endoperoxide, PGH
is converted enzymatically into a wide range of products that are receptor ligands in
normal physiological processes. PGH2 is also known to spontaneously rearrange to fo
PG isomers, PGD2 and PGE2. Additionally, Dr. Salomon discovered that PGH2
rearranges nonenzymatically to give γ-ketoaldehyde functional compounds with
substituted prostanoid chains. These γ-ketoaldehydes were named levuglandins (LGs)
Free radical mediated cyclooxygenation of the PUFAs can also generate PGs and the
corresponding LGs. The isomers of LGs generated by free radical processes include a
variety of structural isomers that are not produced through the enzyme-mediated proces
which only generates
s,
stereoisomers of PGs (Scheme 1.7, page 9). Since, wide ranges of
PUFAs are present in the LDL particle; oxidation of LDL can generate an array of PGs as
191

well as isoLGs. Differences in the amounts of LGs that are generated via enzyme-
mediated or free radical mediated processes can potentially be an effective tool to
compare their contributions towards the total oxidative stress and pathobiology.
The LGs and isoLGs can modify proteins with greater avidity, forming “stable”
adducts
ls
y mass
estigate the pharmacological and clinical significance of various LG
derived
m the
ove, LPS can trigger the expression of COX-2 and subsequently
generat
, than most other lipid oxidation products e.g., HNE and MDA.18 This feature
makes them an attractive “bio-marker” tool to assess the oxidative stress in the tissues.
Our lab has generated antibodies against several of the LG adducts to measure their leve
by immunochemical techniques. Using these antibodies, levels of adducts formed by LGs
or isoLGs, have been shown to be elevated in atherosclerosis19 and glaucoma.20
Elevated levels of LG adducts in Alzheimer’s disease have been recently shown b
spectrometric methods.21 Adduction by LGs have shown to cause loss of protein function,
e.g., loss of tublin function,22 and formation of protein crosslinks23 and DNA-protein
crosslinks.24
To inv
adducts requires an in vivo model system. The model system could also be
utilized in distinguishing the levels of enzyme mediated lipid oxidation products fro
free radical mediated processes under a variety of conditions. This can be achieved
immunochemically by using antibodies that are specific to LG adducts derived from
either of the processes.
As mentioned ab
e LGs or PGs. Part A of this chapter describes the use of this method in cultured
mouse peritoneal macrophages (MPM) to identify the proteins that are susceptible to
oxidative modifications. In part B of this chapter, cornea is used as an in vivo model
192

system to study the LPS-induced formation of LGs and isoLGs through corneal
inflammation or keratitis.
193

6.2. Results.
PART A
6.2.1. LPS stimulates formation of LGE2 and iso[4]LGE2 modified proteins in mouse
peritoneal macrophage cell cultures. Macrophages were cultured in the presence or
absence of LPS (10 µg/mL) for 24 h and probed for LGE and iso[4]LGE modified
proteins by Western blot analysis. The proteins from the LPS treated cells showed
increased immunoreactivity with anti-LGE2-KLH polyclonal antibodies (pAb) compared
that of the cells that were not treated (Figure 6.2).
20
2837
50
88114250
LMW
1 2 1 2LMW
LMW low molecular weight marker1-control2-LPS
Coomassie Western blotA B C
32
68
0
100
Control LPS
20
2837
50
88114250
LMW
1 2 1 2LMW
LMW low molecular weight marker1-control2-LPS
Coomassie Western blotA B C
32
68
0
100
Control LPS
Figure 6.2: LGE2-modified proteins in LPS stimulated macrophages. A. Coomassie
stained SDS PAGE proteins from macrophages. B. Western blot analysis for LGE2
immunoreactivity in the electroblotted PVDF membrane. C. Densitometric quantification
of the total immunoreactive bands in each of the lanes (as relative intensity). 1- control
macrophages, 2 - LPS (10 µg/mL) treated macrophages (n=2).
194

When the cell culture was treated with indomethacin, a COX inhibitor, before and
after LPS treatment, staining by the anti-LGE2 pAb was absent (see Appendix). The
electroblots were also probed with anti-iso[4]LGE2 pAb and a small elevation in the
levels of immunoreactivity was observed in LPS treated compared to untreated cells
(Figure 6.3, B & C).
LMW
2029
36
50
89
114250
LMW
Coomassie Western blotA B
1 2 1 2
LMW low molecular weight marker1-control2-LPS
C
4456
0
100
Control LPS
LMW
2029
36
50
89
114250
LMW
Coomassie Western blotA B
1 2 1 2
LMW low molecular weight marker1-control2-LPS
C
4456
0
100
Control LPS
Figure 6.3: Iso[4]LGE2-modified proteins in LPS stimulated macrophages. A.
Coomassie stained SDS PAGE of proteins from macrophages. B. Iso[4]LGE2
immunoreactivity of the electro-blotted PVDF membrane. C. Densitometric
quantification of the total immunoreactive bands in each of the lanes (as relative
intensity).
This suggests that the LPS may induce COX-2 expression, which results in the
generation of LGE2. However, some contribution of the free radical-induced oxidation is
also indicated by the small elevation in iso[4]LGE2 immunoreactivity that ensues upon
195
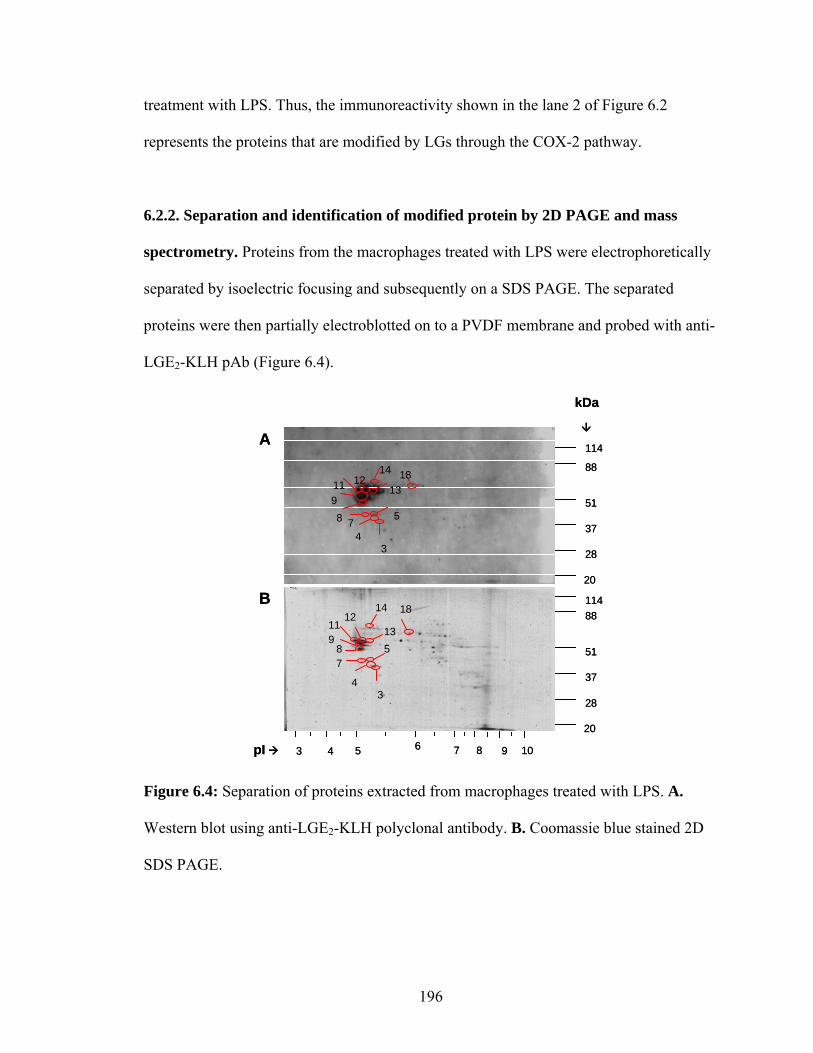
treatment with LPS. Thus, the immunoreactivity shown in the lane 2 of Figure 6.2
represents the proteins that are modified by LGs through the COX-2 pathway.
6.2.2. Separation and identification of modified protein by 2D PAGE and mass
spectrometry. Proteins from the macrophages treated with LPS were electrophoretically
separated by isoelectric focusing and subsequently on a SDS PAGE. The separated
proteins were then partially electroblotted on to a PVDF membrane and probed with anti-
LGE2-KLH pAb (Figure 6.4).
pI
A
B
20
28
37
51
88114
20
28
37
51
88
114
34
578
911
1213
14 18
34
578
911 12
13
14 18
kDa
3 10987654pI
A
B
20
28
37
51
88114
20
28
37
51
88
114
34
578
911
1213
14 18
34
578
911 12
13
14 18
kDa
3 10987654
Figure 6.4: Separation of proteins extracted from macrophages treated with LPS. A.
Western blot using anti-LGE2-KLH polyclonal antibody. B. Coomassie blue stained 2D
SDS PAGE.
196

The Western blots were aligned with the Coomassie stained gel and the protein
spots corresponding to immunoreactive spots were excised. The gel pieces were analyzed
for proteins using the protocol reported earlier.25 The proteins identified by MS/MS
using NCBI database are listed in Table 6.2.
Table 6.1: List of LGE2 – modified proteins electrophoretically separated by 2D gel
electrophoresis and identified by mass spectrometry.
Molecular Mass (kDa/pI) Spot No
Proteins (immunoreactive to
LGE2)
Peptides Matched Measured Calculated
Accession No.
3 β-actin 3 1 42.5/5.3 41.7/5.4 P53486
3 IgG 1 42.5/5.4 11.5/5.8 P01834
3 Tubulin β-1 1 42.5/5.5 49.8/4.8 P09203
4 Rab11 1 43.0/5.2 69.5/9.4 Q8R361
4 Serpin 12 1 43.0/5.3 45.7/6.1 Q9JK88
4 Eucaryotic initiation factor 1 43.0/5.4 44.5/5.3 P87206
5,7,18 Rab11 1 49.5/5.1 92/8.4, 175/7,
55/6.5 Q8R362
8-13 Vimentin 5 55.0/5.1 52-97/7.3-8.7 P48670
14 Keratin, cytoskeletal 4 77.0/5.2 65.0/9.1 P04104
The cytoskeletal proteins viz, actin (actin 3, spot 3), microtubules (tubulin β-1,
spot 3), and two major intermediate filaments vimentin (spots 8-13) and cytokeratin I
(spot 14) were identified. Vimentin and Rab11 have altered masses that correspond to
dimers or trimers (Table 6.2). A detailed discussion is presented in section 6.4.
197

6.2.3. Immunohistochemical analysis of macrophages. Difference in the amount of
LGE2 modified proteins in naive macrophages compared to the cells treated with LPS or
with LPS and indomethacin were found using immunohistochemical analysis. The
macrophages were labeled with 4’,6-diamidino-2-phenylindole (DAPI), a nucleus
specific stain with blue fluorescence and the LGE2-modified proteins were labeled using
a green fluorescent tagged secondary antibody, Alexa 488.
B
A C
DB
A C
D
Figure 6.5: Immunohistochemical analysis of macrophages for LGE2 modified proteins.
macrophages grown to confluency were plated on to a glass slide and treated with or
without LPS. A. Cells treated with LPS and stained with pre-immune serum. B. Controls
cells stained with anti-LGE2-KLH pAb. C. Cells treated with LPS stained with anti-
LGE2-KLH pAb D. Cells treated with LPS along with indomethacin for 24 h and stained
with anti-LGE2-KLH pAb (green fluorescence). Macrophage nuclei are stained with
DAPI (blue fluorescence).
198

As shown in Figure 6.5, there is an increase in the staining (green fluorescence)
of cells treated with LPS (panel C, Figure 6.5) compared to the untreated control cells
(panel B, Figure 6.5). Also, macrophages that were treated with indomethacin before and
after LPS treatment appeared to show less fluorescence (panel D, Figure 6.5) compared
to LPS treated slides, but more than the control cells. This suggests that only some, if
any, LGE2 immunoreactivity is produced through COX pathway, and some is produced
through free radical-induced cyclooxygenation.
199
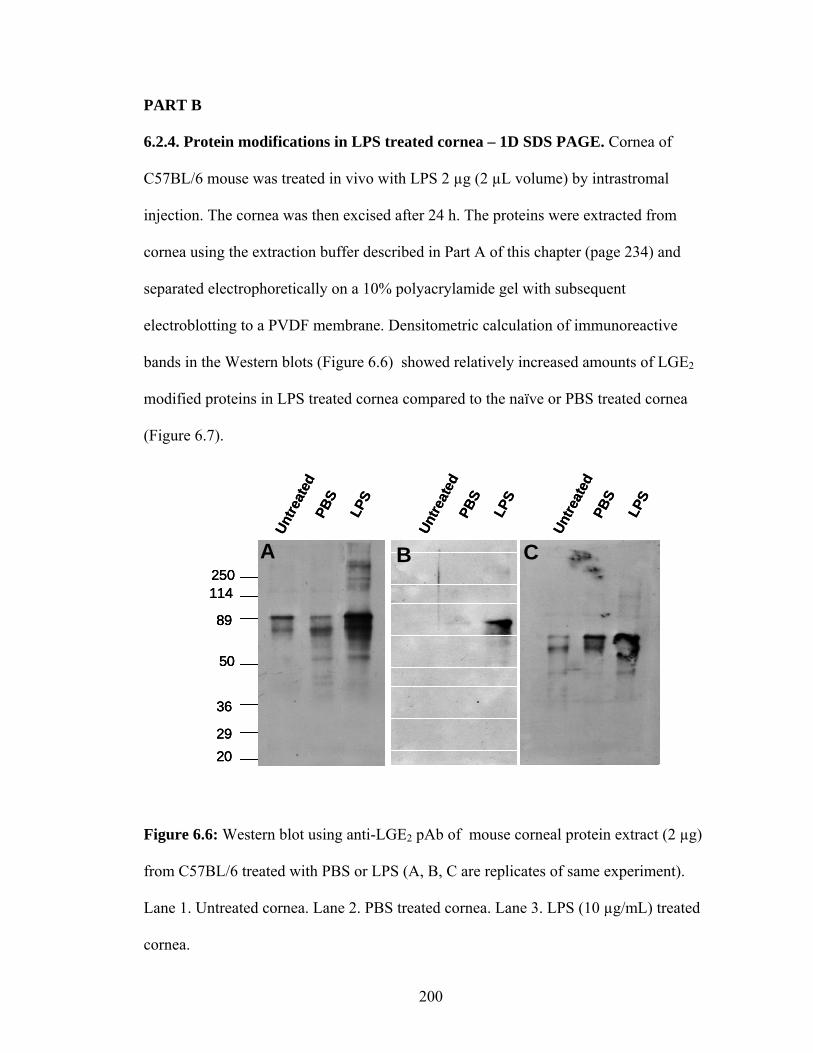
PART B
6.2.4. Protein modifications in LPS treated cornea – 1D SDS PAGE. Cornea of
C57BL/6 mouse was treated in vivo with LPS 2 µg (2 µL volume) by intrastromal
injection. The cornea was then excised after 24 h. The proteins were extracted from
cornea using the extraction buffer described in Part A of this chapter (page 234) and
separated electrophoretically on a 10% polyacrylamide gel with subsequent
electroblotting to a PVDF membrane. Densitometric calculation of immunoreactive
bands in the Western blots (Figure 6.6) showed relatively increased amounts of LGE2
modified proteins in LPS treated cornea compared to the naïve or PBS treated cornea
(Figure 6.7).
2029
36
50
89
114250
Untre
ated
PBS
LPS
Untre
ated
PBS
LPS
Untre
ated
PBS
LPS
A B C
2029
36
50
89
114250
2029
36
50
89
114250
Untre
ated
PBS
LPS
Untre
ated
PBS
LPS
Untre
ated
PBS
LPS
A B C
Figure 6.6: Western blot using anti-LGE2 pAb of mouse corneal protein extract (2 µg)
from C57BL/6 treated with PBS or LPS (A, B, C are replicates of same experiment).
Lane 1. Untreated cornea. Lane 2. PBS treated cornea. Lane 3. LPS (10 µg/mL) treated
cornea.
200

0
20
40
60
80
Untreated PBS LPS
Rel
ativ
e in
tens
ity (%
)
Figure 6.7: Relative amount of LGE2 immunoreactivity. Densitometric quantification of
immunoreactive bands of the Western blots probed with LGE2-pAb. untreated – corneal
proteins of untreated C57BL/6 mouse, PBS – corneal proteins of mice treated with PBS,
LPS - corneal proteins of mice treated with LPS (10 µg/mL). Error bars indicate standard
deviation for three experiments.
6.2.5. Immunohistochemical analysis. To localize the LGE2 and iso[4]LGE2 modified
proteins in the cornea and to determine the effect of LPS on the generation of LGs,
sections of cornea were stained by treatment with the corresponding antibodies. The
stained section shows increased staining in corneal stroma in the LPS treated cornea
compared to that of untreated or PBS treated cornea. The PBS treated cornea shows some
immunoreactivity that could be contributed by intra-stromal injection-induced
inflammation. The amount of green fluorescence in the stromal region of LPS treated
corneal sections stained with LGE2 –pAb (panel L, Figure 6.8), was visibly greater than
201

that of the corresponding stromal region of the iso[4]LGE2 stained sections (panel K,
Figure 6.8).
LGE2-pAbPreimmune
Unt
reat
ed24
h PB
S6h
LP
S24
h LP
S
Iso[4]LGE2-pAb
A
D
G
J K
H
E
B C
F
I
L
Epithelial layer
Stroma
LGE2-pAbPreimmune
Unt
reat
ed24
h PB
S6h
LP
S24
h LP
S
Iso[4]LGE2-pAb
A
D
G
J K
H
E
B C
F
I
L
Epithelial layer
Stroma
Figure 6.8: Cornea of C57BL/6 mouse was treated with LPS or PBS in vivo and
preserved after 6 h and 24 h. Sections (5 µm) of cornea was stained with LGE2 and
iso[4]LGE2 pAb. Untreated corneal sections were used as additional controls.
A,B,C – Untreated cornea; D,E,F – PBS treated (24 h); G,H,I – LPS treated (6 h); J,K,L
– LPS treated (24 h). Panels treated with preimmune, LGE2 and iso[4]LGE2 pAbs are
indicated in the figure.
202

6.3. Discussion.
PART A
Oxidative damage of LDL particles and their subsequent internalization by macrophages
through scavenger receptors leads to foam cell formation, an early step in the
pathobiology of atherosclerosis. Dr. Salomon’s lab previously reported that LDL
modified with LGE2 can bind to the macrophage scavenger receptor26 and additionally,
has identified a set of oxidized phospholipids that can act as ligands for the scavenger
receptor CD36.27 Inefficient processing of oxidatively modified LDL leads to
accumulation of oxLDL in macrophages, and formation of “foam cells”. To understand
the pathological alterations that interfere with metabolism of endocytosed oxLDL by
macrophages, identification of the modified proteins that are present in macrophages can
be helpful. Part A of this chapter is a pilot project in identifying the proteins that are
modified by levuglandins in macrophages as a consequence of activation by
lipopolysaccharides (LPS). This study was conducted in collaboration with Dr. Eugene
Podrez (Cleveland Clinic Foundation).
LPS induced increase in the COX-2 expression along with increase in the
expression of inflammatory cytokines has been reported for various cell types including
macrophages. Mouse peritoneal macrophage cells were cultured and exposed to LPS.
Increases in the amount of COX-2 can subsequently catalyze the formation of LGs. A
definite difference in the amount of LGE2-modified proteins in the LPS stimulated
macrophages compared to the untreated cells was detected by Western blot analysis of
the cellular proteins. Proteins from macrophages pre-treated with a COX inhibitor,
203

indomethacin, showed no immunoreactivity for LGE2 pAb upon exposure to LPS. This
establishes that the protein modification was promoted by COX.
Identification of the LGE2-modified proteins was performed by 2D PAGE
separation followed by identification of the immunoreactive proteins by mass
spectrometric analysis. The proteins identified included several cytoskeletal proteins
crucial to cell stability, cell migration, cell division, intracellular transport, vesicular
trafficking, and organelle morphogenesis. All three types of protein filaments were
affected (Table 6.2): actin, microtubules, and two major intermediate filaments vimentin
and cytokeratin I. Of these, vimentin is critical for post-lysosomal trafficking of lipo-
protein cholesterol in SR-BI-expressing cells.28,29 and tubulin is also associated with lipid
efflux.30 We suggest that, if the function of these proteins is compromised by covalent
modification, deficient delivery of cholesterol for efflux could result. We also detected
modification of Rab11 interacting protein (Rip 11) and eukaryotic initiation factor 4A
(elF4A). Rip11 appears to regulate the trafficking of Rab11,31 which in turn is an
essential regulator of the dynamics of recycling endosomes that is critical in cellular
cholesterol transport.32 ElF4A is a subunit of the initiation factor complex elF4F that
mediates binding of mRNA to the ribosome.33 Consequently, modification of these two
proteins suggests that LGs can affect, respectively, cholesterol trafficking and/or the gene
expression profile in macrophages.
204

PART B
Cornea is a clear transparent tissue that is ideal for in vivo visualization and
quantitation of biological experiments through fluorescently-tagged probes. This feature
of cornea is exploited in studying models of angiogenesis,34 tumor growth,35 tissue
preservation,36 and inflammation.37 Here, part B of this chapter deals with developing
cornea as an in vivo model system for generation of LGs and isoLGs upon activation by
LPS (in collaboration with Dr. Eric Carlson and Dr. Victor Perez at Cleveland Clinic
Foundation).
Intrastromal injection of LPS in the cornea is known to induce COX-2
expression.38 We extracted the proteins from cornea 6h and 24 h after LPS injection and
analyzed for LGE2 immunoreactivity using Western blot analysis. There were increased
amounts of LGE2 modified proteins (62%) compared to the untreated (15%) or PBS
(22%) treated cornea (Figure 6.7). Also, immunohistochemical data suggests that there
was an increased amount of modified proteins in LPS treated cornea compared to the
controls (Figure 6.8).
Further experiments in continuation of this pilot project will be focused on identification
of methods to inhibit COX-2 and study the influence of light on generation of various
LGs and isoLGs in cornea.
205

6.4. Experimental Procedures.
6.4.1. General methods. The following commercially available materials were used as
received: bovine serum albumin (BSA, fraction V, 96-99%) and human serum albumin
(HSA, fraction V) were from Sigma (St. Louis, MO); goat anti-rabbit IgG-alkaline
phosphatase (Boehringer-Mannheim); anti-rabbit IgG coupled with Alexa 488 (a green
fluorescent tag) (Molecular probes, OR); Phosphate buffered saline (PBS) was prepared
from a pH 7.4 stock solution containing 0.2 M NaH2PO4/Na2HPO4, 3.0 M NaCl, and
0.02% NaN3 (w/w). This solution was diluted to 10 mM or as needed. Anti-LGE2-KLH
antibodies19 and anti-iso[4]LGE2-KLH39 antibodies were prepared as described
previously. Iso[4]LGE2 was prepared by Mr. Jim Laird in our lab. The iso[4]LGE2 –
HSA and iso[4]LGE2-BSA40 were prepared as described previously.
6.4.2. Protein extraction. Thioglycollate-elicited mouse peritoneal macrophage (MPM)
cells from wild-type (C57BL/6) mice were isolated and cultured by Dr. Podrez.41 The
cells were scraped, pelleted and stored at -20 ºC until used. The pelleted cells were
extracted by homogenization in 100 mM Tris-Cl buffer pH 7.8 containing 5 mM
dithiothreitol, 1mM SnCl2, 50 mM NaHPO4, 1mM diethylenetriaminepentaacetic acid,
100 mM butylated hydroxytoluene and 0.5% SDS. Insoluble material was removed by
centrifugation (8000g for 5 min). Soluble protein was quantified by the Bradford assay42
using amino acid quantified BSA as standard, yielding ~1 µg/µL of soluble protein.
Protein extracts were subjected to SDS-PAGE on 10% gels (Bio-Rad Laboratories,
Hercules, CA) and the gels were used either for Western analyses or for mass
spectrometric proteomic analyses.43,44
206

6.4.3. Western analysis. Electrophoresis and Western blotting methods were carried out
according to published protocols.43 Protein extract (10 µg) was suspended in Laemmli
sample buffer 45 and fractionated on a 10% SDS PAGE and partially electro blotted onto
polyvinyl difluoride (PVDF) membrane (Millipore, Bedford, MA). The PVDF
membranes were blocked with 5% NFDM milk and probed using various antibodies: 10
µg rabbit polyclonal anti-LGE2 and 4.5 µg of rabbit polyclonal anti-iso[4]LGE246 as
described in Chapter 4 and 10 -100 ng anti-rabbit secondary antibody.
6.2.4. Histochemical analysis. Cultured macrophages were fixed on slides and processed
for immunohistochemical analysis. The cells were blocked with 1% BSA solution in PBS
for 1 h and washed with PBS (10 mM) thrice. Following which the cells were incubated
with 1:100 dilution antibody solutions (0.8 µg/µL rabbit polyclonal anti-Iso[4]LGE2 or
1.5 µg/µL anti-LGE2) for 1 h. The sections were then washed with a solution of 1% BSA
in PBS (3 x 5 min) and subsequently incubated with 10 ng anti-rabbit secondary antibody
conjugated with Alexa 488 (Molecular Probes Inc, Eugene, OR) for one hour at room
temperature. Sections were washed with 1% BSA in PBS as mentioned above, sealed
with Vectashield®, and analyzed with a Nikon EFD-3 fluorescence microscope attached
to a CCD camera. The microscopic images were assembled in Adobe® Photoshop® CS.
(Experiment performed by Dr. Eugene Podrez)
Immunohistochemical analyses to localize modified proteins in corneal tissue
were performed with eyes enucleated from C57BL/6 mice. The corneal processing was
done by enucleating the eye at the specific time period and snap freezing it in OCT®.
5 µm corneal sections were then cut, fixed with 4% paraformaldehyde. The sections were
207

serially incubated in 1% BSA in phosphate buffered saline pH 7.5, then with 10-100 ng
primary antibody (rabbit polyclonal anti-Iso[4]LGE2 or anti-LGE2) overnight at 4°C. The
sections were then washed with a solution of 1% BSA in PBS (3 x 5 min) and
subsequently incubated with 10 ng anti-rabbit secondary antibody conjugated with Alexa
488 (Molecular Probes Inc, Eugene, OR) for one hour at room temperature. Sections
were washed with 1% BSA in PBS as mentioned above, and sealed with Vectashield®,
and analyzed with a Nikon EFD-3 fluorescence microscope attached to a CCD camera.
The microscopic images were assembled in Adobe® Photoshop® CS. (Experiment
performed by Dr. Eric Carlson)
6.4.5. 2D gel electrophoresis. Protein extract from LPS treated macrophages (80 µg)
were used for the two dimensional electrophoresis as discussed in Chapter 4 (page 151).
6.4.6. LC MS/MS analysis and protein identification. Following the SDS PAGE, the
Gelcode Blue® stained gel bands were excised in 1 mm3 slices and destained according
to standard protocol for in-gel digestion.43 The tryptic digested proteins were analyzed by
LC-MS/MS using a quadrupole-time of flight (QTof2) mass spectrometer equipped with
a CapLC system (Waters, Millford, MA), ProteinLynx Global Server acquisition and
processing software. Peptide digests were trapped on a precolumn (0.3 x 1 mm, 5 µC18,
LC Packing) with 0.1% formic acid in 2% acetonitrile as loading solvent then eluted onto
a capillary column (PicoFrit 0.050 x 50 mm, 5 µ tip ID; New Objective Inc., Woburn,
MA). Chromatography was performed at 250 nL/min with aqueous acetonitrile/formic
acid solvents and 100% of the eluant was directed towards the mass spectrometry. The
208

MS/MS data for representative peptides were collected for the mass range of 50-2000.
Protein identifications from MS/MS data utilized ProteinLynxTM Global Server (Waters
Corporation) and Mascot (Matrix Science) search engines and the Swiss-Protein and
NCBI protein sequence databases.43
209

6.5. Reference.
(1) Ross, R. N Engl J Med 1999, 340, 115-26. (2) Peters, W.; Charo, I. F. Curr Opin Lipidol 2001, 12, 175-80. (3) Hansson, G. K. Arterioscler Thromb Vasc Biol 2001, 21, 1876-90. (4) Smith, J. D.; Trogan, E.; Ginsberg, M.; Grigaux, C.; Tian, J.; Miyata, M. Proc
Natl Acad Sci U S A 1995, 92, 8264-8. (5) Lee, T. S.; Yen, H. C.; Pan, C. C.; Chau, L. Y. Arterioscler Thromb Vasc Biol
1999, 19, 734-42. (6) Panousis, C. G.; Zuckerman, S. H. Arterioscler Thromb Vasc Biol 2000, 20, 1565-
71. (7) Frostegard, J.; Huang, Y. H.; Ronnelid, J.; Schafer-Elinder, L. Arterioscler
Thromb Vasc Biol 1997, 17, 963-8. (8) O'Neill, G. P.; Ford-Hutchinson, A. W. FEBS Lett 1993, 330, 156-60. (9) Habib, A.; Creminon, C.; Frobert, Y.; Grassi, J.; Pradelles, P.; Maclouf, J. J Biol
Chem 1993, 268, 23448-23454. (10) O'Banion, M. K.; Winn, V. D.; Young, D. A. Proc Natl Acad Sci U S A 1992, 89,
4888-92. (11) Linton, M.; Fazio, S. Int J Obesity 2003, 27 Suppl 3, S35-40. (12) Burleigh, M. E.; Babaev, V. R.; Oates, J. A.; Harris, R. C.; Gautam, S.; Riendeau,
D.; Marnett, L. J.; Morrow, J. D.; Fazio, S.; Linton, M. F. Circulation 2002, 105, 1816-1823.
(13) http://en.wikipedia.org/wiki/Lipopolysaccharide, 2005; Vol. 2005. (14) Dal Nogare, A. R. Am J Med Sci 1991, 302, 50-65. (15) Barrios-Rodiles, M.; Tiraloche, G.; Chadee, K. J Immunol 1999, 163, 963-969. (16) Cao, C.; Matsumura, K.; Yamagata, K.; Watanabe, Y. Am J Physiol Regul Integr
Comp Physiol 1997, 272, R1712-1725. (17) D'Acquisto, F.; Iuvone, T.; Rombola, L.; Sautebin, L.; Di Rosa, M.; Carnuccio, R.
FEBS Lett 1997, 418, 175-8. (18) Davies, S. S.; Amarnath, V.; Montine, K. S.; Bernoud-Hubac, N.; Boutaud, O.;
Montine, T. J.; Roberts, L. J., 2nd Faseb J 2002, 16, 715-7. (19) Salomon, R. G.; Subbanagounder, G.; O'Neil, J.; Kaur, K.; Smith, M. A.; Hoff, H.
F.; Perry, G.; Monnier, V. M. Chem Res Toxicol 1997, 10, 536-45. (20) Annangudi, S. P.; Rockwood, E. J.; Smith, S. D.; Salomon, R. G.; Bhattacharya,
S. K.; Crabb, J. W. Invest. Ophthalmol. Vis. Sci. 2005, 46, E-Abstract 3694. (21) Zagol-Ikapitte, I.; Masterson, T. S.; Amarnath, V.; Montine, T. J.; Andreasson, K.
I.; Boutaud, O.; Oates, J. A. J Neurochem 2005, 94, 1140-5. (22) Murthi, K. K.; Salomon, R. G.; Sternlicht, H. Prostaglandins 1990, 39, 611-22. (23) Jirousek, M. R.; Murthi, K. K.; Salomon, R. G. Prostaglandins 1990, 40, 187-
203. (24) Murthi, K. K.; Friedman, L. R.; Oleinick, N. L.; Salomon, R. G. Biochemistry
1993, 32, 4090-7. (25) Miyagi, M.; Sakaguchi, H.; Darrow, R. M.; Yan, L.; West, K. A.; Aulak, K. S.;
Stuehr, D. J.; Hollyfield, J. G.; Organisciak, D. T.; Crabb, J. W. Mol Cell Proteomics 2002, 1, 293-303.
210

(26) Hoppe, G.; Subbanagounder, G.; O'Neil, J.; Salomon, R. G.; Hoff, H. F. Biochim Biophys Acta 1997, 1344, 1-5.
(27) Podrez, E. A.; Poliakov, E.; Shen, Z.; Zhang, R.; Deng, Y.; Sun, M.; Finton, P. J.; Shan, L.; Gugiu, B.; Fox, P. L.; Hoff, H. F.; Salomon, R. G.; Hazen, S. L. J Biol Chem 2002, 277, 38503-16.
(28) Sarria, A. J.; Panini, S. R.; Evans, R. M. In J Biol Chem, 1992; Vol. 267. (29) Holwell, T. A.; Schweitzer, S. C.; Reyland, M. E.; Evans, R. M. J Lipid Res 1999,
40, 1440-52. (30) Rogler, G.; Aschenbrenner, E.; Gross, V.; Stange, E. F.; Scholmerich, J. Digestion
2000, 61, 47-58. (31) Junutula, J. R.; Schonteich, E.; Wilson, G. M.; Peden, A. A.; Scheller, R. H.;
Prekeris, R. J Biol Chem 2004, 279, 33430-7. (32) Holtta-Vuori, M.; Tanhuanpaa, K.; Mobius, W.; Somerharju, P.; Ikonen, E. Mol
Biol Cell 2002, 13, 3107-22. (33) Svitkin, Y. V.; Pause, A.; Haghighat, A.; Pyronnet, S.; Witherell, G.; Belsham, G.
J.; Sonenberg, N. RNA 2001, 7, 382-94. (34) Kenyon, B.; Voest, E.; Chen, C.; Flynn, E.; Folkman, J.; D'Amato, R. Invest
Ophthalmol Vis Sci 1996, 37, 1625-1632. (35) Gimbrone, M. A., Jr.; Cotran, R. S.; Leapman, S. B.; Folkman, J. J Natl Cancer
Inst 1974, 52, 413-27. (36) Chen, C. H.; Rama, P.; Chen, S. C.; Sansoy, F. N. Transplantation 1997, 63, 656-
63. (37) Pfister, R.; Haddox, J.; Sommers, C. Invest Ophthalmol Vis Sci 1998, 39, 1744-
1750. (38) Garcia-Cabanes, C.; Palmero, M.; Bellot, J. L.; Castillo, M.; Orts, A. J Ocul
Pharmacol Ther 2001, 17, 67-74. (39) Salomon, R. G.; Sha, W.; Brame, C.; Kaur, K.; Subbanagounder, G.; O'Neil, J.;
Hoff, H. F.; Roberts, L. J., 2nd J Biol Chem 1999, 274, 20271-80. (40) Subbanagounder, G.; Salomon, R. G.; Murthi, K. K.; Brame, C.; Roberts, L. J. J
Org Chem 1997, 62, 7658-7666. (41) Podrez, E. A.; Schmitt, D.; Hoff, H. F.; Hazen, S. L. J Clin Invest 1999, 103,
1547-60. (42) Bradford, M. M. Anal Biochem 1976, 72, 248-54. (43) Crabb, J. W.; Miyagi, M.; Gu, X.; Shadrach, K.; West, K. A.; Sakaguchi, H.;
Kamei, M.; Hasan, A.; Yan, L.; Rayborn, M. E.; Salomon, R. G.; Hollyfield, J. G. Proc Natl Acad Sci U S A 2002, 99, 14682-7.
(44) Bhattacharya, S. K.; Rockwood, E. J.; Smith, S. D.; Bonilha, V. L.; Crabb, J. S.; Kuchtey, R. W.; Robertson, N. G.; Peachey, N. S.; Morton, C. C.; Crabb, J. W. J Biol Chem 2005, 280, 6080-4.
(45) Laemmli, U. K. Nature 1970, 227, 680-5. (46) Salomon, R. G.; Batyreva, E.; Kaur, K.; Sprecher, D. L.; Schreiber, M. J.; Crabb,
J. W.; Penn, M. S.; DiCorletoe, A. M.; Hazen, S. L.; Podrez, E. A. Biochim Biophys Acta 2000, 1485, 225-35.
211

Appendix
213

Chapter 2
O OTs
2.14
a)
b)
Figure 2.1S: a) 13C NMR and b) 1H NMR of 2.14
214

O
OTs
OHO
2.36
Figure 2.2S: 1H NMR of 2.38 (crude)
OC5H11
OHO
2.37
Figure 2.3S: a) 13C NMR and b) 1H NMR of 2.37
215

HO
O
O
OTs
2.17 a)
b)
Figure 2.4S: a) 13C NMR and b) 1H NMR of 2.17
216

O
OTBDMS
OHO
2.38 a)
b)
Figure 2.5S: a) 13C NMR and b) 1H NMR of 2.38
217
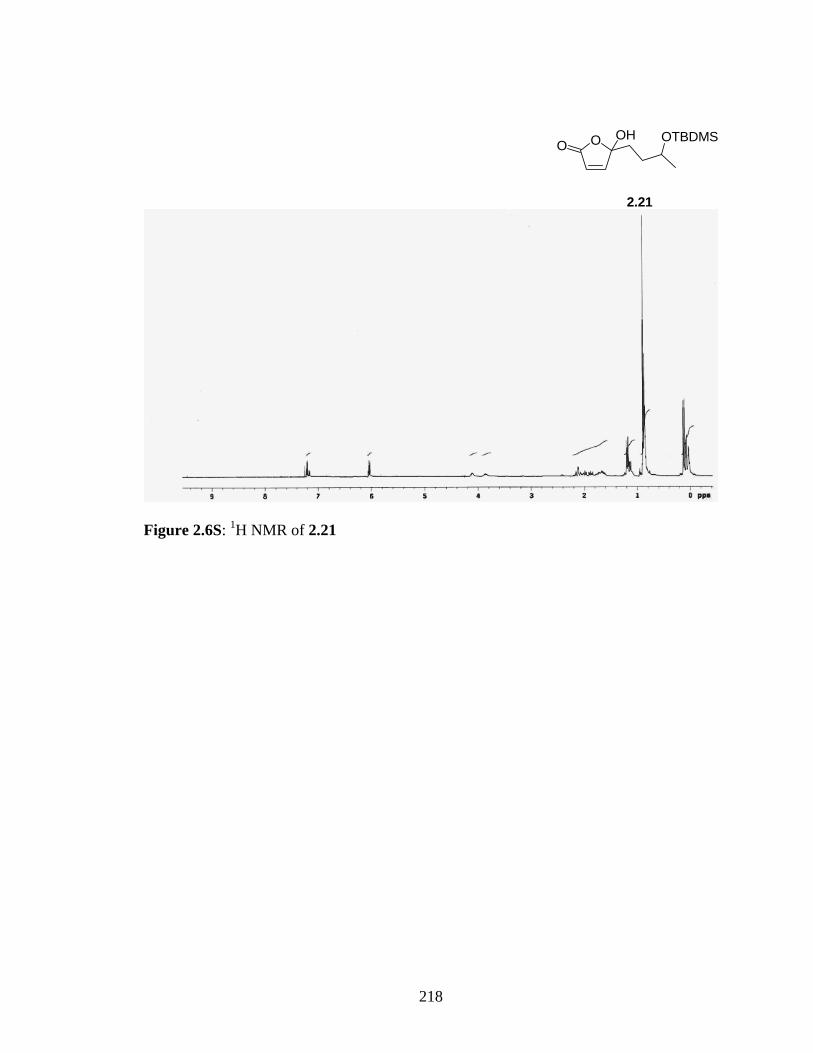
O OHO
OTBDMS
2.21
Figure 2.6S: 1H NMR of 2.21
218

a)
HO
O
O
OTBDMS
2.22
b)
Figure 2.7S: a) 13C NMR and b) 1H NMR of 2.22
219

O
OTHP
OHO
2.39
Figure 2.8S: 1H NMR of 2.39 (crude)
220

O
O
O
OTHP
2.40 a)
b)
Figure 2.9S: a) 13C (APT) NMR and b) 1H NMR of sec-butyl ester of 2.40
221

2.27
Figure 2.10S: 1H NMR of 2.27 (crude)
222

2.26
Figure 2.11S : 1H NMR of 2.26
223

Chapter 3
HD2C
D2C O
SO
O
3.3
Figure 3.1S: 1H NMR of 3.3
224

HD2C
D2C I
3.4
Figure 3.2S: 2H NMR of 3.4
225

a)
OOHD2C
CD2 OH
3.6
b)
Figure 3.3S: a) 1H NMR b) 13C NMR of 3.6
226
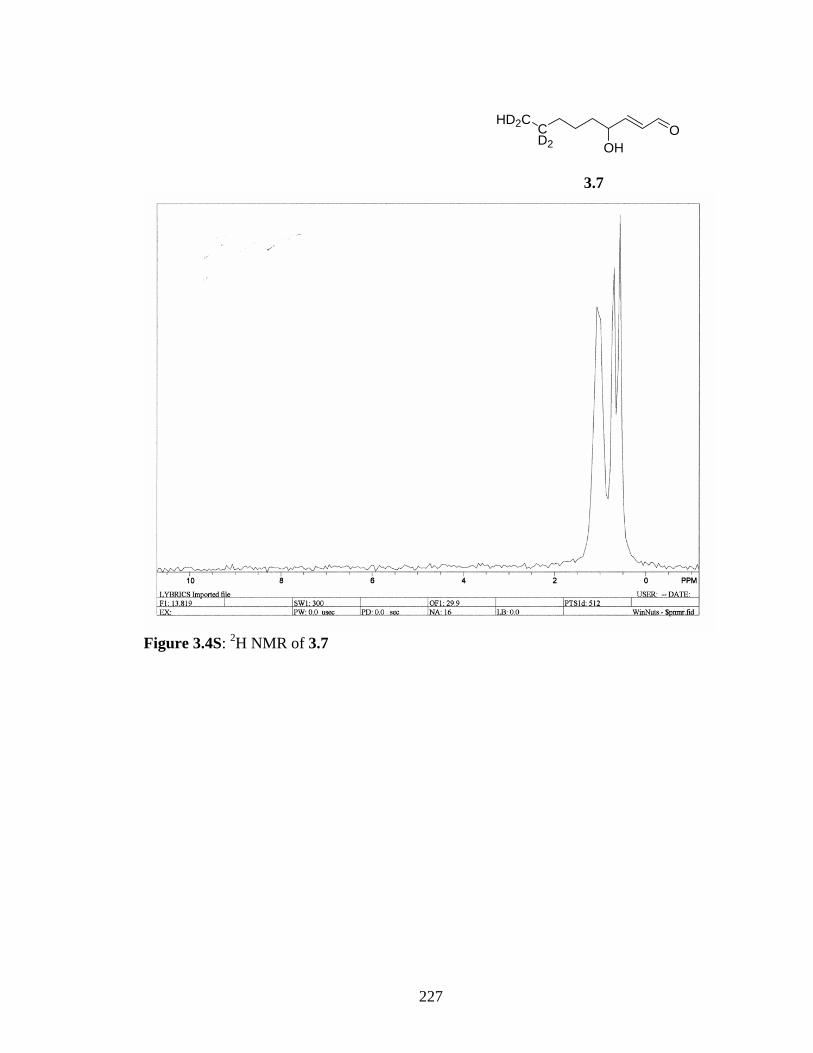
OHD2C
CD2 OH
3.7
Figure 3.4S: 2H NMR of 3.7
227

OHD2C
CD2 OH
3.7a)
b)
Figure 3.5S: a) 1H NMR and b) 13C NMR of 3.7
228

TOF – time of flight analyzerMSMS – tandem mass 260.00 – mass of parent ion analyzedES+ - Electrospray positive mode
Intensity ions formed
TOF – time of flight analyzerMSMS – tandem mass 260.00 – mass of parent ion analyzedES+ - Electrospray positive mode
Intensity ions formed
Figure 3.6S: How to read a mass spectra?
H2NOH
O
N
NH
O
198 m/z
CH2CO
H3NOH
O
N
NH
156 m/z
HCOOH
HN
N
NH2
110 m/z
OHO
N
NH
139 m/z
HN
NH
82 m/z
HCN
H2N
N
NH
O
154 m/z
NH
H2N
44 m/z
44 m/z
NH3
229

Scheme 3.1S: Fragmentation of N-acetyl histidine
HNOH
O
N
N
O
O
H2NOH
O
N
N
O
C15H25N3O4Exact Mass: 311.1845
C17H27N3O5Exact Mass: 353.1951
N
N
N
O
O
C16H25N3O3Exact Mass: 307.1896
-carboxyl
H3N
N
N
O
HCOOH
C12H19N3O•+
Exact Mass: 221.1523
HO
HOHO
HO
-acetyl
N
NH
OHO
C14H23N2O2+
Exact Mass: 251.1754
C14H24N3O2+
Exact Mass: 266.1863
NH3
1,2-hydride shift
HN
N
N
OH
HN
N
NH
OHO
-acetyl
-acetyl -carboxyl
Scheme 3.2S: Fragmentation of (N-acetyl-His)-HNE Michael adduct
HN
O
H2N
ONH
OO
O
HO
HNO
NH2
O
NHO
O
OHN
O
HN
ONH
OO
HO
HO
O
-H20
Retro-Michael
H2NO
NH
HN
O
O
HO
O
McL -acetyl
-H20
m/z 416
m/z 139
m/z 372 m/z 372
m/z 398
230

Scheme 3.3S: Suggested fragmentation of (N-acetyl-Gly-Lys-OMe)-HNE Michael (1:1)
adduct.
HNO
N
O
NH
O
OOH
H2NO
N
NH
O
OOH
-acetyl
N
NH3
O
O
m/z 281
NOH
O
m/z 250
Nm/z 204
NOH
222 m/z
m/z 354
H2O
Amide cleavage
CO2
-NH3, -CH3
H2O
m/z 398
Scheme 3.5S: Suggested fragmentation of (N-acetyl-Gly-Lys-OMe)-HNE Schiff base
adduct
231

NHO
N
OHN
O
O
NH3
O
N
HN
O
O
N
N
NH3O
O
m/z 380
m/z 338
m/z 204 m/z 281
Scheme 3.6S: Fragmentation of (N-acetyl-Gly-Lys-OMe)-HNE pyrrole adduct
His-HNE 10ug0.00000000
2.00 4.00 6.00 8.00 10.00 12.00 14.00 16.00 18.00Time0
100
%
0
100
%
0
100
%
0
100
%
0
100
%
0
100
%
HIS_HNE-07 SIR of 13 Channels ES+ 696
8.17e411.16
10.291.68
17.4415.8414.5712.10 13.90 18.38
HIS_HNE-07 SIR of 13 Channels ES+ 692
8.37e417.44
15.7013.7011.901.28
19.58
HIS_HNE-07 SIR of 13 Channels ES+ 688
7.66e418.5815.84
14.6313.7011.83
1.61
17.57
HIS_HNE-07 SIR of 13 Channels ES+ 684
3.70e519.51
15.8414.70 18.44
HIS_HNE-07 SIR of 13 Channels ES+ 678
1.70e515.70
14.63
13.6317.51 19.64
HIS_HNE-07 SIR of 13 Channels ES+ 674
4.60e515.70
14.6319.78
232

Figure 3.7S: ESI-TOF-SIR of N-acetyl-His/HNE reaction mixture. Channels shown here
m/z 696, m/z 692, m/z 688, m/z 684, m/z 678 and m/z 674.
Figure 3.8S: ESI-TOF-SIR of (N-acetyl-Gly-Lys-OMe)/HNE reaction mixture. Channels
shown here are from m/z 384, 402, 420, 580, 640, 644, 652, 656, 736 and 740.
233

HN
OHO
S
O
NH
HO O
S
O
HN
OHO
S
O
S HN
OHO
S
O
NH2
HO O
S
m/z 281m/z 208
HN
OHO
S
O
NH
O
S
O
m/z 307
m/z 324-H2O
-acetylα−cleavage
Scheme 3.7S: Fragmentation of (N-acetyl-Cys)-(N-acetyl-Cys) disulphide adduct
HN
HO
O
SO
OHO
NHHO
O
SH
O
O
NH3HO
O
S
O
NH2
O
S
O
m/z 302
m/z 260m/z 242
m/z 319
-acetyl
-H2O
-H2O
Scheme 3.8S: Fragmentation of (N-acetyl-Cys)-HNE Michael adduct
234

Cys-HNE10ug0.00000000
2.00 4.00 6.00 8.00 10.00 12.00 14.00 16.00 18.00Time0
100
%
0
100
%
0
100
%
0
100
%
0
100
%
0
100
%
CystHNE-10 SIR of 13 Channels ES+ 360
4.12e611.03
10.43 15.5711.5013.63
CystHNE-10 SIR of 13 Channels ES+ 356
4.35e610.16 10.89
11.5013.70
CystHNE-10 SIR of 13 Channels ES+ 352
4.42e511.09
10.89
15.7715.1711.50 14.03
CystHNE-10 SIR of 13 Channels ES+ 326
3.23e511.76
10.161.34 10.49
17.3114.17
13.63 15.77
CystHNE-10 SIR of 13 Channels ES+ 320
2.38e59.76
8.822.280.2117.3114.0311.5610.96
13.03 16.3715.7718.17
CystHNE-10 SIR of 13 Channels ES+ TIC
1.05e711.09
9.8311.50
15.7715.2413.70 17.31
Figure 3.9S: ESI-TOF-SIR of (N-acetyl-Cys)/HNE reaction mixture. Channels shown
here are from channels monitored at m/z 320, m/z 326, m/z 352, m/z 356 and m/z 360.
235

O OH
HNOH
O
N
N
O
OH
O
OHOH
H3NOH
O
N
N
O
O
m/z 468
OHOH
H2N
N
N
O
O
m/z 422
OHOH
H2N
N
N
O
O
O
m/z 466
OHOH
OHO
N
N
O
O
m/z 451
OHOH
N
N
O
O
m/z 407
-acetyl
-carboxyl
-NH3
m/z 510
-acetyl -carboxyl
-acetyl
Scheme 3.9S: Suggested fragmentation of (N-acetyl-His)-HNE 1:2 adduct
OHOH
HNOH
O
N
N
O
O
HO
O
O
OHOH
HNOH
O
N
N
O
O
m/z 394
OHOH
HNOH
O
N
N
O
O
m/z 466
m/z 665
O OHOH
HO
O
O
m/z 467
Scheme 3.10S: Suggested fragmentation of (N-acetyl-His)-HNE 1:2 adduct
236

Table 3.1 Summary of tandem MS/MS fragments incorporating the C5 alkyl chain of
HNE derived from the precursor ions 354, 358, 510, 514, and 510 thus showing the 4 Da
difference.
Fragments with mass difference of 4 Da
Precursor ion (m/z)
Mass Range (Da)
354
358
510
514
518
550-500 - - 510 514 518
500-450 - - 464, 466, 468 468, 470, 472 472, 474, 476
450-425 - - 422 426 430
425-400 - - 407 411 415
375-345 354 358 354
348
354, 358
348, 352
358
352
345-300 308, 312
336
312, 316
340
308, 312
336
312, 316
336, 340
312, 316
340
300-275 277
290, 294
281
294, 298
277
290,294
277, 281
290, 294, 298
281
294, 298
275-245 248, 251
259, 266
252,255
263, 270
248, 251
259, 266
248,251,252,255
259,263,266,270
252,255
263, 270
245-200
-
221, 233
-
-
225, 237
-
209
221, 233
239
209,213
221,225, 233,237
239,243
213
225, 237
243
200-100 139 143 139 139,143 143
237

Chapter 4
43FW
77M
W
85FW
52M
W
49M
W
85FW
62FW
67FW
62FW
77M
W
46FW
77M
W
85FW
81FW
82FW
63M
W
86FW
57FW
61FW
59FW
74M
W
Normal TM Glaucomatous TM
11488
50
35
28
20
25043
FW
77M
W
85FW
52M
W
49M
W
85FW
62FW
67FW
62FW
77M
W
46FW
77M
W
85FW
81FW
82FW
63M
W
86FW
57FW
61FW
59FW
74M
W
Normal TM Glaucomatous TM
11488
50
35
28
20
250
11488
50
35
28
20
250
Figure 4.1S: Western blot analyses of normal and POAG trabecular meshwork using
anti-iso[4]LGE2 antibodies as described in Section 4.4.4.
43FW
77M
W
85FW
52M
W
49M
W
85FW
62FW
67FW
62FW
77M
W
46FW
77M
W
85FW
81FW
82FW
63M
W
86FW
57FW
61FW
59FW
74M
W
Normal TM Glaucomatous TM
11488
50
35
28
20
250
43FW
77M
W
85FW
52M
W
49M
W
85FW
62FW
67FW
62FW
77M
W
46FW
77M
W
85FW
81FW
82FW
63M
W
86FW
57FW
61FW
59FW
74M
W
Normal TM Glaucomatous TM
11488
50
35
28
20
250
11488
50
35
28
20
250
Figure 4.2S: Western blot analyses of normal and POAG trabecular meshwork using
anti-HNE antibodies as described in Section 4.4.4.
238

88M
W
74 M
W
87 M
W
84 M
W
80 F
W
72 M
W
87 M
W
87 M
W
LMW
c
11488
50
35
28
20
Normal TM Glaucomatous TM
11488
50
35
28
20
88M
W
74 M
W
87 M
W
84 M
W
80 F
W
72 M
W
87 M
W
87 M
W
LMW
c
11488
50
35
28
20
11488
50
35
28
20
Normal TM Glaucomatous TM
11488
50
35
28
20
11488
50
35
28
20
Figure 4.3S: Western blot analyses of normal and POAG trabecular meshwork using
anti-argpyrimidine antibodies as described in Section 4.4.4.
LMW
Prot
ein e
xtrac
t
IP u
sing
iso[4
]LGE
2 pA
bIP
usin
g Pr
eimm
une
se
114
88
50
35
28
20
250
LMW
Prot
ein e
xtrac
t
IP u
sing
iso[4
]LGE
2 pA
bIP
usin
g Pr
eimm
une
se
LMW
Prot
ein e
xtrac
t
IP u
sing
iso[4
]LGE
2 pA
bIP
usin
g Pr
eimm
une
se
114
88
50
35
28
20
250
114
88
50
35
28
20
250
Figure 4.4S: Immunoprecipitation of POAG trabecular meshwork using anti-iso[4]LGE2
pAb and preimmune serum antibodies as described in Section 4.4.6
239

Chapter 5
A
B
7 da
ys
15 d
ays
3 w
eeks
1 m
onth
s
2.5
mon
ths
8 m
onth
s
9 m
onth
s
BS
A-Is
o[4]
LGE
2
LMWA
B
7 da
ys
15 d
ays
3 w
eeks
1 m
onth
s
2.5
mon
ths
8 m
onth
s
9 m
onth
s
BS
A-Is
o[4]
LGE
2
LMW
Figure 5.1S: Western blot of TM proteins from DBA/2J mouse of different age groups..
5 µg of protein was electrophoretically separated on a SDS PAGE and electroblotted on
a PVDF membrane and probed with iso[4]LGE2 pAb. Time course of DBA/2J mice. A.
Western blot. B. Coomassie stained SDS PAGE.
240
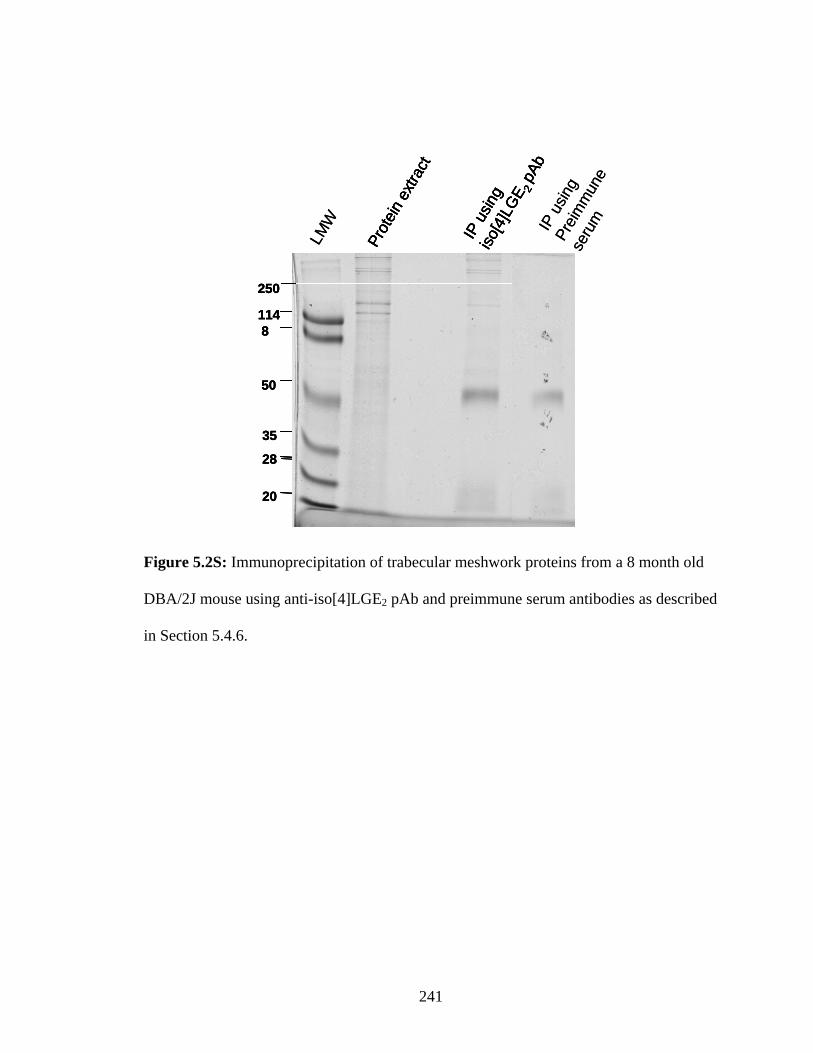
Prot
ein e
xtrac
t
IP u
sing
iso[4
]LGE
2 pA
b
1148
50
35
28
20
250
LMW
Prot
ein e
xtrac
t
IP u
sing
iso[4
]LGE
2 pA
bIP
usin
g Pr
eimm
une
seru
m
Prot
ein e
xtrac
t
IP u
sing
iso[4
]LGE
2 pA
b
1148
50
35
28
20
250
1148
50
35
28
20
250
LMW
Prot
ein e
xtrac
t
IP u
sing
iso[4
]LGE
2 pA
bIP
usin
g Pr
eimm
une
seru
m
Figure 5.2S: Immunoprecipitation of trabecular meshwork proteins from a 8 month old
DBA/2J mouse using anti-iso[4]LGE2 pAb and preimmune serum antibodies as described
in Section 5.4.6.
241

Chapter 6
1 2LMW
LMW low molecular weight marker1-control2-LPS3-treated with indomethacin
Western blot
31 2LMW
LMW low molecular weight marker1-control2-LPS3-treated with indomethacin
Western blot
3
Figure 6.1S: LGE2-modified proteins in LPS stimulated MPM cells. Western blot
analysis for LGE2 immunoreactivity in the electroblotted PVDF membrane.
242

Thesis Conclusion
This thesis primarily focuses on the levels of lipid derived protein modifications
in glaucoma. Previously, other studies, used spectrophotometric methods to quantify the
lipid oxidation products in glaucoma, or investigated the biological consequences of
oxidative stress in cell culture models of glaucoma. The present study, using antibodies
against protein modifications (iso[4] levuglandin E2 and 4-hydroxynonenal) derived from
lipid oxidation, provides the first direct evidence for elevated levels of lipid-derived
oxidative protein modifications in the trabecular meshwork of primary open angle
glaucoma compared to the controls. The oxidatively modified proteins were identified
using immunoprecipitation, 2D gel electrophoresis and mass spectrometry. Proteins
identified to be putatively modified, included several structural proteins, which may be
important in maintaining the structure as well as the function of the trabecular meshwork.
Future work may be focused on the role of these modified proteins in the increase of
intraocular pressure that may augment our understanding of glaucoma pathogenesis.
Additionally, an increase in the levels of proteins modified by iso[4] levuglandin
E2 was observed in the trabecular meshwork of glaucomatous DBA/2J mice, a model for
glaucoma. This model can be valuable for studying the role of the modified proteins in
glaucoma pathogenesis. Using models for these analyses is desirable, inter alia, because
tissue samples from human glaucoma patients are difficult to obtain.
Multiple additions of the lipid oxidation product 4-hydroxynonenal (HNE) to side
chain residues of lysine and histidine were detected and characterized using deuterated
HNE and mass spectrometry. Future work may focus on the detection of such multiple
243

HNE adducts in vivo and determine their role in detoxification and/or pathology of
oxidative insult.
244

Bibliography
245

Adger BM, Barrett C, Brennan J, McKervey MA & Murray RW. (1991). Oxidation of furans with dimethyldioxirane. . J Chem Soc, Chem Comm 21, 1553-1554.
Aldini G, Granata P & Carini M. (2002). Detoxification of cytotoxic alpha,beta-unsaturated aldehydes by carnosine: characterization of conjugated adducts by electrospray ionization tandem mass spectrometry and detection by liquid chromatography/mass spectrometry in rat skeletal muscle. J Mass Spectrom 37, 1219-1228.
Allingham MT, Howard-Jones A, Murphy PJ, Thomas DA & Caulkett PWR. (2003). Synthesis and applications of C2-symmetric guanidine bases. Tetrahedron Lett 44, 8677-8680.
Alvarado J, Murphy C, Polansky J & Juster R. (1981). Age-related changes in trabecular meshwork cellularity. Invest Ophthalmol Vis Sci 21, 714-727.
Amarnath V, Valentine WM, Amarnath K, Eng MA & Graham DG. (1994). The mechanism of nucleophilic substitution of alkylpyrroles in the presence of oxygen. Chem Res Toxicol 7, 56-61.
Amarnath V, Valentine WM, Montine TJ, Patterson WH, Amarnath K, Bassett CN & Graham DG. (1998). Reactions of 4-Hydroxy-2(E)-nonenal and Related Aldehydes with Proteins Studied by Carbon-13 Nuclear Magnetic Resonance Spectroscopy. Chem Res Toxicol 11, 317-328.
Amarnath V, Valentine WM, Montine TJ, Patterson WH, Amarnath K, Bassett CN & Graham DG. (1998). Reactions of 4-Hydroxy-2(E)-nonenal and Related Aldehydes with Proteins Studied by Carbon-13 Nuclear Magnetic Resonance Spectroscopy. Chem Res Toxicol 11, 317-328.
Anderson MG, Smith RS, Hawes NL, Zabaleta A, Chang B, Wiggs JL & John SW. (2002). Mutations in genes encoding melanosomal proteins cause pigmentary glaucoma in DBA/2J mice. Nat Genet 30, 81-85.
Annangudi SP, Sun M & Salomon RG. (2005). An Efficient Synthesis of 4-Oxoalkenoic Acids from 2-Alkylfurans. Synlett 2005, 1468-1470.
Annangudi SP, Rockwood EJ, Smith SD, Salomon RG, Bhattacharya SK & Crabb JW. (2005). Oxidative Protein Modifications in Glaucomatous Trabecular Meshwork. Invest Ophthalmol Vis Sci 46, E-Abstract 3694.
Appleton D, Duguid AB, Lee S-K, Ha Y-J, Ha H-J & Leeper FJ. (1998). Synthesis of analogs of 5-aminolevulinic acid and inhibition of 5-aminolevulinic acid dehydratase. J Chem Soc, Perkin Trans 1: Org Bio-Org Chem, 89-101.
246

Araki N, Ueno N, Chakrabarti B, Morino Y & Horiuchi S. (1992). Immunochemical evidence for the presence of advanced glycation end products in human lens proteins and its positive correlation with aging. J Biol Chem 267, 10211-10214.
Arlt S, Kontush A, Muller-Thomsen T & Beisiegel U. (2001). [Lipid peroxidation as a common pathomechanism in coronary heart disease and Alzheimer disease]. Z Gerontol Geriatr 34, 461-465.
Asaoka M, Yanagida N, Sugimura N & Takei H. (1980). The reaction of 2-(trialkylsiloxy)furans with lead(IV) acetate. The synthesis of dl-pyrenophorin. . Bull Chem Soc Jpn 53, 1061-1064.
Audhya A & Emr S. (2002). Stt4 PI 4-kinase localizes to the plasma membrane and functions in the Pkc1-mediated MAP kinase cascade. Dev Cell 2, 593-605.
Babizhayev MA & Brodskaya MW. (1989). Fibronectin detection in drainage outflow system of human eyes in ageing and progression of open-angle glaucoma. Mech Ageing Dev 47, 145-157.
Babizhayev MA & Bunin A. (1989). Lipid peroxidation in open-angle glaucoma. Acta Ophthalmol (Copenh) 67, 371-377.
Babizhayev MA & Costa EB. (1994). Lipid peroxide and reactive oxygen species generating systems of the crystalline lens. Biochim Biophys Acta 1225, 326-337.
Balamraju YN, Sun M & Salomon RG. (2004). Gamma-hydroxyalkenals are oxidatively cleaved through Michael addition of acylperoxy radicals and fragmentation of intermediate beta-hydroxyperesters. J Am Chem Soc 126, 11522-11528.
Ballini R & Bosica G. (1998). Synthesis of (E)-4-oxonon-2-enoic acid, a natural antibiotic produced by streptomyces olivaceus. J Nat Prod 61, 673-674.
Ballini R, Bosica G, Fiorini D, Gil MV & Petrini M. (2001). Stereoselective Synthesis of (E)-4-Alkylidenecyclopent-2-en-1-ones by a Tandem Ring Closure-Michael Addition-Elimination. Org Lett 3, 1265-1267.
Barco A, Benetti S, De Risi C, Pollini G & Zanirato V. (1995a). A [3+2] Nitrile Oxide Cycloaddition Approach to (-)-Pyrenophorin, and Rosefuran. Tetrahedron 51, 7721.
Barrios-Rodiles M, Tiraloche G & Chadee K. (1999). Lipopolysaccharide Modulates Cyclooxygenase-2 Transcriptionally and Posttranscriptionally in Human Macrophages Independently from Endogenous IL-1{beta} and TNF-{alpha}. J Immunol 163, 963-969.
Baynes JW. (2001). The role of AGEs in aging: causation or correlation. Exp Gerontol 36, 1527-1537.
Beatty S, Koh H, Phil M, Henson D & Boulton M. (2000). The role of oxidative stress in the pathogenesis of age-related macular degeneration. Surv Ophthalmol 45, 115-134.
247

Bergamini C, Gambetti S, Dondi A & Cervellati C. (2004). Oxygen, reactive oxygen species and tissue damage. Curr Pharma Design 10, 1611-1626.
Bhattacharya SK, Rockwood EJ, Smith SD, Bonilha VL, Crabb JS, Kuchtey RW, Robertson NG, Peachey NS, Morton CC & Crabb JW. (2005). Proteomics reveal Cochlin deposits associated with glaucomatous trabecular meshwork. J Biol Chem 280, 6080-6084.
Birch AJ & Walker KAM. (1966). J Chem Soc (C), 1894.
Bonini C, Chiummiento L, Evidente A & Funicello M. (1995). First enantioselective synthesis of (-)-seiridin the major phytotoxic metabolite of Seiridium species pathogenic for cypress. Tetrahedron Lett 36, 7285-7286.
Boon PJM, Marinho HS, Oosting R & Mulder GJ. (1999). Glutathione Conjugation of 4-Hydroxy-trans-2,3-nonenal in the Rat in Vivo, the Isolated Perfused Liver and Erythrocytes. Toxicol Appl Pharmacol 159, 214-223.
Bouchain G, Leit S, Frechette S, Khalil E, Lavoie R, Moradei O, Woo S, Fournel M, Yan P, Kalita A, Trachy-Bourget M, Beaulieu C, Li Z, Robert M, MacLeod A, Besterman J & Delorme D. (2003). Development of potential antitumor agents. Synthesis and biological evaluation of a new set of sulfonamide derivatives as histone deacetylase inhibitors. J Med Chem 46, 820-830.
Boukouvalas J & Lachance N. (1998). A mild, efficient, and general method for the synthesis of trialkylsilyl (Z)-4-oxo-2-alkenoates and .gamma.-hydroxybutenolides. Synlett, 31-32.
Boutaud O, Andreasson K, Zagol-Ikapitte I & Oates J. (2005). Cyclooxygenase-dependent lipid-modification of brain proteins. Brain pathol (Zurich, Switzerland) 15, 139-142.
Boutaud O, Li J, Zagol I, Shipp EA, Davies SS, Roberts LJ, 2nd & Oates JA. (2003). Levuglandinyl adducts of proteins are formed via a prostaglandin H2 synthase-dependent pathway after platelet activation. J Biol Chem 278, 16926-16928.
Bowie AG, Moynagh PN & O'Neill LA. (1997). Lipid peroxidation is involved in the activation of NF-kappaB by tumor necrosis factor but not interleukin-1 in the human endothelial cell line ECV304. Lack of involvement of H2O2 in NF-kappaB activation by either cytokine in both primary and transformed endothelial cells. J Biol Chem 272, 25941-25950.
Bracher F & Schulte B. (2003). Total synthesis of both enantiomers of the macrocyclic lactone citreofuran. Nat Prod Res 17, 293-299.
Bradford MM. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72, 248-254.
248

Brame CJ, Salomon RG, Morrow JD & Roberts LJ, 2nd. (1999). Identification of extremely reactive gamma-ketoaldehydes (isolevuglandins) as products of the isoprostane pathway and characterization of their lysyl protein adducts. J Biol Chem 274, 13139-13146.
Brown MS & Goldstein JL. (1983). Lipoprotein metabolism in the macrophage: implications for cholesterol deposition in atherosclerosis. Annu Rev Biochem 52, 223-261.
Bucolo C, Campana G, Di Toro R, Cacciaguerra S & Spampinato S. (1999). Sigma1 recognition sites in rabbit iris-ciliary body: topical sigma1-site agonists lower intraocular pressure. J Pharmacol Exp Ther 289, 1362-1369.
Bunin A, Filina AA & Erichev VP. (1992). [A glutathione deficiency in open-angle glaucoma and the approaches to its correction]. Vestn Oftalmol 108, 13-15.
Bunin A. (2000). [Pathogenetic factors of destructive process in trabecular tissues in primary open-angle glaucoma]. Vestn Oftalmol 116, 24-27.
Burleigh ME, Babaev VR, Oates JA, Harris RC, Gautam S, Riendeau D, Marnett LJ, Morrow JD, Fazio S & Linton MF. (2002). Cyclooxygenase-2 Promotes Early Atherosclerotic Lesion Formation in LDL Receptor-Deficient Mice. Circulation 105, 1816-1823.
Caballero M, Liton PB, Epstein DL & Gonzalez P. (2003). Proteasome inhibition by chronic oxidative stress in human trabecular meshwork cells. Biochem Biophys Res Commun 308, 346-352.
Cao C, Matsumura K, Yamagata K & Watanabe Y. (1997). Involvement of cyclooxygenase-2 in LPS-induced fever and regulation of its mRNA by LPS in the rat brain. Am J Physiol Regul Integr Comp Physiol 272, R1712-1725.
Carles L, Narkunan K, Penlou S, Rousset L, Bouchu D & Ciufolini MA. (2002). 2-Pyridones from Cyanoacetamides and Enecarbonyl Compounds: Application to the Synthesis of Nothapodytine B. J Org Chem 67, 4304-4308.
Castellani RJ, Perry G, Harris PL, Cohen ML, Sayre LM, Salomon RG & Smith MA. (1998). Advanced lipid peroxidation end-products in Alexander's disease. Brain Res 787, 15-18.
Chakraborti T, Ghosh SK, Michael JR, Batabyal SK & Chakraborti S. (1998). Targets of oxidative stress in cardiovascular system. Mol Cell Biochem 187, 1-10.
Chandrasekar B, Colston JT, de la Rosa SD, Rao PP & Freeman GL. (2003). TNF-alpha and H2O2 induce IL-18 and IL-18R beta expression in cardiomyocytes via NF-kappa B activation. Biochem Biophys Res Commun 303, 1152-1158.
249

Chang B, Smith RS, Hawes NL, Anderson MG, Zabaleta A, Savinova O, Roderick TH, Heckenlively JR, Davisson MT & John SW. (1999). Interacting loci cause severe iris atrophy and glaucoma in DBA/2J mice. Nat Genet 21, 405-409.
Chen CH, Rama P, Chen SC & Sansoy FN. (1997). Efficacy of organ preservation media enriched with nonlactate-generating substrate for maintaining tissue viability: a transplantation study. Transplantation 63, 656-663.
Chen JZ & Kadlubar FF. (2003). A new clue to glaucoma pathogenesis. Am J Med 114, 697-698.
Chew S. (1996). Animal models of glaucoma. In The Glaucomas: Basic Sciences, Anatomy and Pathology, 2 edn, ed. Ritch R, Shields M & Krupin T. Mosby, New York.
Choi J, Rees HD, Weintraub ST, Levey AI, Chin LS & Li L. (2005). Oxidative modifications and aggregation of Cu,Zn-superoxide dismutase associated with Alzheimer and Parkinson diseases. J Biol Chem 280, 11648-11655.
Ciechanover A & Brundin P. (2003). The ubiquitin proteasome system in neurodegenerative diseases: sometimes the chicken, sometimes the egg. Neuron 40, 427-446.
Cottier L, Descotes G, Eymard L & Rapp K. (1995). Syntheses of g-oxo acids or g-oxo esters by photooxygenation of furanic compounds and reduction under ultrasound: application to the synthesis of 5-aminolevulinic acid hydrochloride. Synthesis, 303-306.
Crabb JW, Miyagi M, Gu X, Shadrach K, West KA, Sakaguchi H, Kamei M, Hasan A, Yan L, Rayborn ME, Salomon RG & Hollyfield JG. (2002a). Drusen proteome analysis: an approach to the etiology of age-related macular degeneration. Proc Natl Acad Sci U S A 99, 14682-14687.
Crabb JW, O'Neil J, Miyagi M, West K & Hoff HF. (2002b). Hydroxynonenal inactivates cathepsin B by forming Michael adducts with active site residues. Protein Sci 11, 831-840.
Cringle SJ, Yu DY, Yu PK & Su EN. (2002). Intraretinal oxygen consumption in the rat in vivo. Invest Ophthalmol Vis Sci 43, 1922-1927.
D'Acquisto F, Iuvone T, Rombola L, Sautebin L, Di Rosa M & Carnuccio R. (1997). Involvement of NF-kappaB in the regulation of cyclooxygenase-2 protein expression in LPS-stimulated J774 macrophages. FEBS Lett 418, 175-178.
Dal Nogare AR. (1991). Septic shock. Am J Med Sci 302, 50-65.
D'Auria M, Piancatelli G & Scettri A. (1980). Synthesis of 4-ylidenebutenolides and 4-oxo-2-enoic acid methyl esters from 5-methoxy-2-furyl carbinols. Tetrahedron 36, 3071-3074.
250

Davies SS, Amarnath V, Montine KS, Bernoud-Hubac N, Boutaud O, Montine TJ & Roberts LJ, 2nd. (2002). Effects of reactive gamma-ketoaldehydes formed by the isoprostane pathway (isoketals) and cyclooxygenase pathway (levuglandins) on proteasome function. Faseb J 16, 715-717.
De La Paz MA & Epstein DL. (1996). Effect of age on superoxide dismutase activity of human trabecular meshwork. Invest Ophthalmol Vis Sci 37, 1849-1853.
De Mattia G, Bravi MC, Laurenti O, Cassone-Faldetta M, Proietti A, De Luca O, Armiento A & Ferri C. (1998). Reduction of oxidative stress by oral N-acetyl-L-cysteine treatment decreases plasma soluble vascular cell adhesion molecule-1 concentrations in non-obese, non-dyslipidaemic, normotensive, patients with non-insulin-dependent diabetes. Diabetologia 41, 1392-1396.
Deng Y & Salomon RG. (1998). Synthesis of [9-3H]-trans-4-Hydroxy-2-nonenal. J Org Chem 63, 3504-3507.
Deng Y. (2000). Oxidized Lipids: Part I: Radiochemical Studies On HNE-Protein Adduction Part II: Total Synthesis Of Oxidized Phospholipids And Cholesterol Esters. In Department of Chemistry. Case Western Reserve University, Cleveland.
DiFranco E, Subbanagounder G, Kim S, Murthi K, Taneda S, Monnier VM & Salomon RG. (1995). Formation and stability of pyrrole adducts in the reaction of levuglandin E2 with proteins. Chem Res Toxicol 8, 61-67.
Dittami JP, Xu F, Qi H, Martin MW, Bordner J, Decosta DL, Kiplinger J, Reiche P & Ware R. (1995). Preparation of N-alkyl pyrrolidinones via photocyclization of g-keto-a,b-unsaturated amides. Tetrahedron Lett 36, 4197-4200.
Doolittle MH, Ben-Zeev O & Briquet-Laugier V. (1998). Enhanced detection of lipoprotein lipase by combining immunoprecipitation with Western blot analysis. J Lipid Res 39, 934-942.
Doorn JA & Petersen DR. (2003). Covalent adduction of nucleophilic amino acids by 4-hydroxynonenal and 4-oxononenal. Chem Biol Interact 143-144, 93-100.
Doreleijers JF. (1999). Validation of Biomolecular NMR Structures. Universiteit Utrecht.
Du Yan S, Zhu H, Fu J, Yan SF, Roher A, Tourtellotte WW, Rajavashisth T, Chen X, Godman GC, Stern D & Schmidt AM. (1997). Amyloid-beta peptide-receptor for advanced glycation endproduct interaction elicits neuronal expression of macrophage-colony stimulating factor: a proinflammatory pathway in Alzheimer disease. Proc Natl Acad Sci U S A 94, 5296-5301.
Dunlevy JR & Rada JAS. (2004). Interaction of Lumican with Aggrecan in the Aging Human Sclera. Invest Ophthalmol Vis Sci 45, 3849-3856.
251

Dyka FM, May CA & Enz R. (2004). Metabotropic glutamate receptors are differentially regulated under elevated intraocular pressure. J Neurochem 90, 190-202.
Edwards GL, Muldoon CA & Sinclair DJ. (1996). Cyclic Enol Ether Synthesis via Arenesulfonyl Iodide Additions to Alkynols. Tetrahedron 52, 7779-7788.
Enoiu M, Herber R, Wennig R, Marson C, Bodaud H, Leroy P, Mitrea N, Siest G & Wellman M. (2002). gamma-Glutamyltranspeptidase-dependent metabolism of 4-hydroxynonenal-glutathione conjugate. Arch Biochem Biophys 397, 18-27.
Epstein DL. (1997). Chandler and Grant's glaucoma. Williams & Wilkins, co.,, Baltimore, Md.
Esterbauer H, Schaur RJ & Zollner H. (1991). Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic Biol Med 11, 81-128.
Farkas RH, Chowers I, Hackam AS, Kageyama M, Nickells RW, Otteson DC, Duh EJ, Wang C, Valenta DF, Gunatilaka TL, Pease ME, Quigley HA & Zack DJ. (2004). Increased expression of iron-regulating genes in monkey and human glaucoma. Invest Ophthalmol Vis Sci 45, 1410-1417.
Farooqui AA & Horrocks LA. (1998). Lipid Peroxides in the Free Radical Pathophysiology of Brain Diseases. Cell Mol Neurobiology 18, 599-608.
Felder S & Rowan DD. (1999). Synthesis of deuterated C-6 and C-9 flavour volatiles. J Labelled Compounds Radiopharm 42, 83 - 92.
Ferreira SM, Lerner SF, Brunzini R, Evelson PA & Llesuy SF. (2004). Oxidative stress markers in aqueous humor of glaucoma patients. Am J Ophthalmol 137, 62-69.
Fingert J, Heon E, Liebmann J, Yamamoto T, Craig J, Rait J, Kawase K, Hoh S, Buys Y, Dickinson J, Hockey R, Williams-Lyn D, Trope G, Kitazawa Y, Ritch R, Mackey D, Alward W, Sheffield V & Stone E. (1999). Analysis of myocilin mutations in 1703 glaucoma patients from five different populations. Hum Mol Genet 8, 899-905.
Finkel T & Holbrook NJ. (2000). Oxidants, oxidative stress and the biology of ageing. Nature 408, 239-247.
Finlay JM, M. Anthony; Gunaratne, H. Q. Nimal. (1998). Oxidations Catalysed by Rhenium(V) Oxo Species 1. Conversion of Furans to Enediones using Methyltrioxorhenium and Urea Hydrogen Peroxide. Tetrahedron lett 39, 5651.
Foreman D, Zuk L, Miller DB & Salomon RG. (1987). Effects of E2 levuglandins on the contractile activity of the rat uterus. Prostaglandins Other Lipid Mediators 34, 91-98.
Fraga CG, Shigenaga MK, Park JW, Degan P & Ames BN. (1990). Oxidative damage to DNA during aging: 8-hydroxy-2'-deoxyguanosine in rat organ DNA and urine. Proc Natl Acad Sci U S A 87, 4533-4537.
252

Franzen R, Tanabe K & Morita M. (1998). Ring-chain tautomerism of chlorinated hydroxyfuranones and reaction with nucleosides. Chemosphere 38, 973-980.
Frisch SM & Ruoslahti E. (1997). Integrins and anoikis. Curr Opin Cell Biol 9, 701-706.
Frostegard J, Huang YH, Ronnelid J & Schafer-Elinder L. (1997). Platelet-activating factor and oxidized LDL induce immune activation by a common mechanism. Arterioscler Thromb Vasc Biol 17, 963-968.
Gaasterland D & Kupfer C. (1974). Experimental glaucoma in the rhesus monkey. Invest Ophthalmol 13, 455-457.
Garcia-Cabanes C, Palmero M, Bellot JL, Castillo M & Orts A. (2001). Inhibition of COX in ocular tissues: an in vitro model to identify selective COX-2 inhibitors. J Ocul Pharmacol Ther 17, 67-74.
Gardner HW & Deighton N. (2001). Effect of 4-hydroxy-2(E)-nonenal on soybean lipoxygenase-1. Lipids 36, 623-628.
Garner B, Shaw DC, Lindner RA, Carver JA & Truscott RJ. (2000). Non-oxidative modification of lens crystallins by kynurenine: a novel post-translational protein modification with possible relevance to ageing and cataract. Biochim Biophys Acta 1476, 265-278.
Garner B, Vazquez S, Griffith R, Lindner RA, Carver JA & Truscott RJ. (1999). Identification of glutathionyl-3-hydroxykynurenine glucoside as a novel fluorophore associated with aging of the human lens. J Biol Chem 274, 20847-20854.
Gerhardinger C, Taneda S, Marion MS & Monnier VM. (1994). Isolation, purification, and characterization of an Amadori product binding protein from a Pseudomonas sp. soil strain. J Biol Chem 269, 27297-27302.
Gherghel D, Griffiths HR, Hilton EJ, Cunliffe IA & Hosking SL. (2005). Systemic reduction in glutathione levels occurs in patients with primary open-angle glaucoma. Invest Ophthalmol Vis Sci 46, 877-883.
Giancotti FG. (1997). Integrin signaling: specificity and control of cell survival and cell cycle progression. Curr Opin Cell Biol 9, 691-700.
Gimbrone MA, Jr., Cotran RS, Leapman SB & Folkman J. (1974). Tumor growth and neovascularization: an experimental model using the rabbit cornea. J Natl Cancer Inst 52, 413-427.
Gioacchini AM, Calonghi N, Boga C, Cappadone C, Masotti L, Roda A & Traldi P. (1999). Determination of 4-hydroxy-2-nonenal at cellular levels by means of electrospray mass spectrometry. Rapid Commun Mass Spectrom 13, 1573-1579.
253

Girotti AW. (1998). Lipid hydroperoxide generation, turnover, and effector action in biological systems. J Lipid Res 39, 1529-1542.
Goldstein LE, Leopold MC, Huang X, Atwood CS, Saunders AJ, Hartshorn M, Lim JT, Faget KY, Muffat JA, Scarpa RC, Chylack LT, Jr., Bowden EF, Tanzi RE & Bush AI. (2000). 3-Hydroxykynurenine and 3-hydroxyanthranilic acid generate hydrogen peroxide and promote alpha-crystallin cross-linking by metal ion reduction. Biochemistry 39, 7266-7275.
Gollnick K & Griesbeck A. (1985). Singlet oxygen photooxygenation of furans. Isolation and reactions of (4 + 2)-cycloaddition products (unsaturated sec.-ozonides). Tetrahedron 41, 2057-2068.
Gonzalez-Avila G, Ginebra M, Hayakawa T, Vadillo-Ortega F, Teran L & Selman M. (1995). Collagen metabolism in human aqueous humor from primary open-angle glaucoma. Decreased degradation and increased biosynthesis play a role in its pathogenesis. Arch Ophthalmol 113, 1319-1323.
Gordon MO, Beiser JA, Brandt JD, Heuer DK, Higginbotham EJ, Johnson CA, Keltner JL, Miller JP, Parrish RK, 2nd, Wilson MR & Kass MA. (2002). The Ocular Hypertension Treatment Study: baseline factors that predict the onset of primary open-angle glaucoma. Arch Ophthalmol 120, 714-720; discussion 829-730.
Gotz ME, Kunig G, Riederer P & Youdim MB. (1994). Oxidative stress: free radical production in neural degeneration. Pharmacol Ther 63, 37-122.
Grandjean EM, Berthet P, Ruffmann R & Leuenberger P. (2000). Efficacy of oral long-term N-acetylcysteine in chronic bronchopulmonary disease: a meta-analysis of published double-blind, placebo-controlled clinical trials. Clin Ther 22, 209-221.
Green K. (1995). Free radicals and aging of anterior segment tissues of the eye: a hypothesis. Ophthalmic Res 27 Suppl 1, 143-149.
Gross JH. (2004). Mass Spectrometry- A Textbook. Springer-Verlag, Heidelberg.
Gross RL, Ji J, Chang P, Pennesi ME, Yang Z, Zhang J & Wu SM. (2003). A mouse model of elevated intraocular pressure: retina and optic nerve findings. Trans Am Ophthalmol Soc 101, 163-169; discussion 169-171.
Gu X, Meer SG, Miyagi M, Rayborn ME, Hollyfield JG, Crabb JW & Salomon RG. (2003). Carboxyethylpyrrole protein adducts and autoantibodies, biomarkers for age-related macular degeneration. J Biol Chem 278, 42027-42035.
Gunn BP & Brooks DW. (1985). Total synthesis of (+-)-12-hydroxy-5,8,14-cis-10-trans-eicosatetraenoic acid (12-HETE). J Org Chem 50, 4417-4418.
254

Gunn BP. (1985). An efficient and general methodology for the synthesis of the HETES: synthesis of (+-)-5-hydroxy-6-trans-8,11,14-cis-eicosatetraenoic acid (5-HETE). Tetrahedron Lett 26, 2869-2872.
Ha SW, Kim HJ, Bae JS, Jeong GH, Chung SC, Kim JG, Park SH, Kim YL, Kam S, Kim IS & Kim BW. (2004). Elevation of urinary betaig-h3, transforming growth factor-beta-induced protein in patients with type 2 diabetes and nephropathy. Diabetes Res Clin Pract 65, 167-173.
Habib A, Creminon C, Frobert Y, Grassi J, Pradelles P & Maclouf J. (1993). Demonstration of an inducible cyclooxygenase in human endothelial cells using antibodies raised against the carboxyl-terminal region of the cyclooxygenase-2. J Biol Chem 268, 23448-23454.
Halland N, Aburel PS & Jorgensen KA. (2003). Highly enantioselective organocatalytic conjugate addition of malonates to acyclic a,b-unsaturated enones. Angew Chem Int Edn 42, 661-665.
Halland N, Hazell RG & Jorgensen KA. (2002). Organocatalytic Asymmetric Conjugate Addition of Nitroalkanes to a,b-Unsaturated Enones Using Novel Imidazolidine Catalysts. J Org Chem 67, 8331-8338.
Hamberg M, Svensson J & Samuelsson B. (1974). Prostaglandin Endoperoxides. A New Concept concerning the Mode of Action and Release of Prostaglandins. Proc Natl Acad Sci U S A 71, 3824-3828.
Hansson GK. (2001). Immune mechanisms in atherosclerosis. Arterioscler Thromb Vasc Biol 21, 1876-1890.
Harlow E & Lane D. (1988). Antibodies: a laboratory manual. Cold Spring Harbor Laboratory.
Harrison KA & Murphy RC. (1995). Isoleukotrienes are biologically active free radical products of lipid peroxidation. J Biol Chem 270, 17273-17278.
Hase TA & Nylund EL. (1979). Synthesis of the macrolide antibiotic A26771B methyl ester. Tetrahedron Lett 2633-2636.
Hazen SL & Chisolm GM. (2002). Oxidized phosphatidylcholines: pattern recognition ligands for multiple pathways of the innate immune response. Proc Natl Acad Sci U S A 99, 12515-12517.
Hirsch JA & Szur AJ. (1972). Reactions of acyclic enediones with menthofuran. Tetrahedron 28, 2961-2967.
Hoff H, O'Neil J, Chisolm G, 3d, Cole T, Quehenberger O, Esterbauer H & Jurgens G. (1989). Modification of low density lipoprotein with 4-hydroxynonenal induces uptake by macrophages. Arterioscler Thromb Vasc Biol 9, 538-549.
255

Holtta-Vuori M, Tanhuanpaa K, Mobius W, Somerharju P & Ikonen E. (2002). Modulation of cellular cholesterol transport and homeostasis by Rab11. Mol Biol Cell 13, 3107-3122.
Holwell TA, Schweitzer SC, Reyland ME & Evans RM. (1999). Vimentin-dependent utilization of LDL-cholesterol in human adrenal tumor cells is not associated with the level of expression of apoE, sterol carrier protein-2, or caveolin. J Lipid Res 40, 1440-1452.
Hood BD, Garner B & Truscott RJ. (1999). Human lens coloration and aging. Evidence for crystallin modification by the major ultraviolet filter, 3-hydroxy-kynurenine O-beta-D-glucoside. J Biol Chem 274, 32547-32550.
Hoppe G, Subbanagounder G, O'Neil J, Salomon RG & Hoff HF. (1997). Macrophage recognition of LDL modified by levuglandin E2, an oxidation product of arachidonic acid. Biochim Biophys Acta 1344, 1-5.
Hrbac J & Kohen R. (2000). Biological redox activity: Its importance, methods for its quantification and implication for health and disease. Drug Development Research 50, 516-527.
http://ca.expasy.org/uniprot/P51888. (2005).
http://en.wikipedia.org/wiki/Lipopolysaccharide. (2005).
http://www.ayurveda-ayurvedic.com/e-zine/health-ezine-404.html. (Volume 1, Issue 4. April 2004). ANCIENT VEDIC WISDOM FOR A HEALTHY HEART.
http://www.glaucoma.org/learn/. Glaucoma.
Ikeda Y, Ohguro H & Maruyama I. (2000). Two cases of primary open angle glaucoma with serum autoantibody against retinal ganglion cells. Jpn J Ophthalmol 44, 648-652.
Ishida Y, Migita K, Izumi Y, Nakao K, Ida H, Kawakami A, Abiru S, Ishibashi H, Eguchi K & Ishii N. (2004). The role of IL-18 in the modulation of matrix metalloproteinases and migration of human natural killer (NK) cells. FEBS Lett 569, 156-160.
Isom AL, Barnes S, Wilson L, Kirk M, Coward L & Darley-Usmar V. (2004). Modification of Cytochrome c by 4-hydroxy- 2-nonenal: evidence for histidine, lysine, and arginine-aldehyde adducts. J Am Soc Mass Spectrom 15, 1136-1147.
Itakura K, Osawa T & Uchida K. (1998). Structure of a Fluorescent Compound Formed from 4-Hydroxy-2-nonenal and N(alpha)-Hippuryllysine: A Model for Fluorophores Derived from Protein Modifications by Lipid Peroxidation. J Org Chem 63, 185-187.
256

Itakura K, Oya-Ito T, Osawa T, Yamada S, Toyokuni S, Shibata N, Kobayashi M & Uchida K. (2000). Detection of lipofuscin-like fluorophore in oxidized human low-density lipoprotein. FEBS Lett 473, 249-253.
Iyer RS, Ghosh S & Salomon RG. (1989). Levuglandin E2 crosslinks proteins. Prostaglandins 37, 471-480.
Izzotti A, Sacca SC, Cartiglia C & De Flora S. (2003). Oxidative deoxyribonucleic acid damage in the eyes of glaucoma patients. Am J Med 114, 638-646.
Jacobi PC, Dietlein TS & Krieglstein GnK. (2000). Effect of trabecular aspiration on intraocular pressure in pigment dispersion syndrome and pigmentary glaucomaThe authors have no proprietary interest in the development or marketing of this or any competing piece of equipment. Ophthalmology 107, 417-421.
Jirousek MR, Murthi KK & Salomon RG. (1990). Electrophilic levuglandin E2-protein adducts bind glycine: a model for protein crosslinking. Prostaglandins 40, 187-203.
John SW, Smith RS, Savinova OV, Hawes NL, Chang B, Turnbull D, Davisson M, Roderick TH & Heckenlively JR. (1998). Essential iris atrophy, pigment dispersion, and glaucoma in DBA/2J mice. Invest Ophthalmol Vis Sci 39, 951-962.
Jr CL, Brown N, Bron A, Hurst M, Kopcke W, Thien U & Schalch W. (2002). The Roche European American Cataract Trial (REACT): a randomized clinical trial to investigate the efficacy of an oral antioxidant micronutrient mixture to slow progression of age-related cataract. Ophthalmic Epidemiology 9, 49-80.
Junutula JR, Schonteich E, Wilson GM, Peden AA, Scheller RH & Prekeris R. (2004). Molecular characterization of Rab11 interactions with members of the family of Rab11-interacting proteins. J Biol Chem 279, 33430-33437.
Jurgens G, Lang J & Esterbauer H. (1986). Modification of human low-density lipoprotein by the lipid peroxidation product 4-hydroxynonenal. Biochim Biophys Acta 875, 103-114.
Kahn MG, Giblin FJ & Epstein DL. (1983). Glutathione in calf trabecular meshwork and its relation to aqueous humor outflow facility. Invest Ophthalmol Vis Sci 24, 1283-1287.
Kalita D, Khan AT, Barua NC & Bez G. (1999). Total synthesis of (R)-(+)-Patulolide A and (R)-(-)-Patulolide B: the macrolides isolated from Penicillium urticae mutant. Tetrahedron 55, 5177-5184.
Kates M. (1986). Techniques of lipidology : isolation, analysis, and identification of lipids. Elsevier, Amsterdam,The Netherlands.
Kaur K, Salomon RG, O'Neil J & Hoff HF. (1997). (Carboxyalkyl)pyrroles in human plasma and oxidized low-density lipoproteins. Chem Res Toxicol 10, 1387-1396.
257

Kawashima M, Sato T & Fujisawa T. (1988). Regio- and stereoselective ring opening of w-alkenyllactones using organocopper reagents. Bull Chem Soc Jpn 61, 3255-3264.
Kenyon B, Voest E, Chen C, Flynn E, Folkman J & D'Amato R. (1996). A model of angiogenesis in the mouse cornea. Invest Ophthalmol Vis Sci 37, 1625-1632.
Kingston D. (1974). Intramolecular hydrogen transfer in mass spectra. II. The McLafferty Rearrangement and related reactions. Chem Rev 74, 215.
Ko ML, Hu DN, Ritch R & Sharma SC. (2000). The combined effect of brain-derived neurotrophic factor and a free radical scavenger in experimental glaucoma. Invest Ophthalmol Vis Sci 41, 2967-2971.
Kobayashi Y, Kishihara K & Watatani K. (1996). Two-step Conversion of 2-Substituted Furans into γ-Oxo-α,β-unsaturated Carboxylic Acids. Formal Synthesis of (+)-Patulolide A and (−)-Pyrenophorin. Tetrahedron Lett 37, 4385-4388.
Kobayashi Y, Kumar GB, Kurachi T, Acharya HP, Yamazaki T & Kitazume T. (2001). Furan ring oxidation strategy for the synthesis of macrosphelides A and B. J Org Chem 66, 2011-2018.
Kobayashi Y, Nakano M, Kumar GB & Kishihara K. (1998). Efficient Conditions for Conversion of 2-Substituted Furans into 4-Oxygenated 2-Enoic Acids and Its Application to Synthesis of (+)-Aspicilin, (+)-Patulolide A, and (-)-Pyrenophorin. J Org Chem 63, 7505-7515.
Kondo Y, Murakami S, Oda H & Nagate T. (2000). Taurine reduces atherosclerotic lesion development in apolipoprotein E-deficient mice. Adv Exp Med Biol 483, 193-202.
Kopitz J, Holz FG, Kaemmerer E & Schutt F. (2004). Lipids and lipid peroxidation products in the pathogenesis of age-related macular degeneration. Biochimie 86, 825-831.
Kurysheva NI, Vinetskaia MI, Erichev VP, Demchuk ML & Kuryshev SI. (1996). [Contribution of free-radical reactions of chamber humor to the development of primary open-angle glaucoma]. Vestn Oftalmol 112, 3-5.
Laemmli UK. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680-685.
Lam TT, Abler AS, Kwong JM & Tso MO. (1999). N-methyl-D-aspartate (NMDA)--induced apoptosis in rat retina. Invest Ophthalmol Vis Sci 40, 2391-2397.
Lee SH & Blair IA. (2000). Characterization of 4-oxo-2-nonenal as a novel product of lipid peroxidation. Chem Res Toxicol 13, 698-702.
258

Lee TS, Yen HC, Pan CC & Chau LY. (1999). The role of interleukin 12 in the development of atherosclerosis in ApoE-deficient mice. Arterioscler Thromb Vasc Biol 19, 734-742.
Li N & Karin M. (1999). Is NF-kappaB the sensor of oxidative stress? Faseb J 13, 1137-1143.
Liao F, Andalibi A, deBeer FC, Fogelman AM & Lusis AJ. (1993). Genetic control of inflammatory gene induction and NF-kappa B-like transcription factor activation in response to an atherogenic diet in mice. J Clin Invest 91, 2572-2579.
Liao F, Andalibi A, Qiao JH, Allayee H, Fogelman AM & Lusis AJ. (1994). Genetic evidence for a common pathway mediating oxidative stress, inflammatory gene induction, and aortic fatty streak formation in mice. J Clin Invest 94, 877-884.
Linton M & Fazio S. (2003). Macrophages, inflammation, and atherosclerosis. International J Obesity 27 Suppl 3, S35
Liu B & Neufeld AH. (2001). Nitric oxide synthase-2 in human optic nerve head astrocytes induced by elevated pressure in vitro. Arch Ophthalmol 119, 240-245.
Liu Y, Jinno H, Kurihara M, Miyata N & Toyo'oka T. (1999). Determination of 4-hydroxy-2-nonenal in primary rat hepatocyte cultures by liquid chromatography with laser induced fluorescence detection. Biomed Chromatogr : BMC 13, 75-80.
Liu Y, Sun G, David A & Sayre LM. (2004). Model studies on the metal-catalyzed protein oxidation: structure of a possible His-Lys cross-link. Chem Res Toxicol 17, 110-118.
Lukevics EY, Ignatovich LM, Gol'dberg YS & Shymanskaya MV. (1986). Alkylation of furan and thiophene with tert-butanol in the presence of the strong acid cation exchanger Amberlyst 15. Khimiya Geterotsiklicheskikh Soedinenii, 853-854.
Macdonald S, Dowle M, Harrison L, Clarke G, Inglis G, Johnson M, Shah P, Smith R, Amour A, Fleetwood G, Humphreys D, Molloy C, Dixon M, Godward R, Wonacott A, Singh O, Hodgson S & Hardy G. (2002). Discovery of further pyrrolidine trans-lactams as inhibitors of human neutrophil elastase (HNE) with potential as development candidates and the crystal structure of HNE complexed with an inhibitor (GW475151). J Med Chem 45, 3878-3890.
Mandelkorn RM, Hoffman ME, Olander KW, Zimmerman TJ & Harsha D. (1985). Inheritance and the pigmentary dispersion syndrome. Ophthalmic Paediatr Genet 6, 325-331.
Marnett LJ, Hurd HK, Hollstein MC, Levin DE, Esterbauer H & Ames BN. (1985). Naturally occurring carbonyl compounds are mutagens in Salmonella tester strain TA104. Mutat Res 148, 25-34.
259

Matsudaira P. (1991). Modular organization of actin crosslinking proteins. Trends Biochem Sci 16, 87-92.
Matsushita Y-i, Sugamoto K & Matsui T. (1998). Syntheses of fatty acid derivatives derived from lipid peroxidation by the application of cobalt porphyrin-catalyzed reduction-oxygenation. Tennen Yuki Kagobutsu Toronkai Koen Yoshishu 40th, 613-618.
McCall MR & Frei B. (1999). Can antioxidant vitamins materially reduce oxidative damage in humans? Free Radic Biol Med 26, 1034-1053.
Mikheytseva IN, Kashintseva LT, Krizhanovsky GN, Kopp OP & Lipovetskaya EM. (2004). The influence of the calcium channel blocker verapamil on experimental glaucoma. Int Ophthalmol 25, 75-79.
Miyagi M, Sakaguchi H, Darrow RM, Yan L, West KA, Aulak KS, Stuehr DJ, Hollyfield JG, Organisciak DT & Crabb JW. (2002). Evidence that light modulates protein nitration in rat retina. Mol Cell Proteomics 1, 293-303.
Miyata T, Inagi R, Iida Y, Sato M, Yamada N, Oda O, Maeda K & Seo H. (1994). Involvement of beta 2-microglobulin modified with advanced glycation end products in the pathogenesis of hemodialysis-associated amyloidosis. Induction of human monocyte chemotaxis and macrophage secretion of tumor necrosis factor-alpha and interleukin-1. J Clin Invest 93, 521-528.
Mlakar A & Spiteller G. (1996). Previously unknown aldehydic lipid peroxidation compounds of arachidonic acid. Chem Phy Lipids 79, 47-53.
Moreno MC, Campanelli J, Sande P, Sanez DA, Keller Sarmiento MI & Rosenstein RE. (2004). Retinal oxidative stress induced by high intraocular pressure. Free Radic Biol Med 37, 803-812.
Morrison JC, Moore CG, Deppmeier LM, Gold BG, Meshul CK & Johnson EC. (1997). A rat model of chronic pressure-induced optic nerve damage. Exp Eye Res 64, 85-96.
Morrow JD, Harris TM & Roberts LJ, 2nd. (1990). Noncyclooxygenase oxidative formation of a series of novel prostaglandins: analytical ramifications for measurement of eicosanoids. Anal Biochem 184, 1-10.
Motyckova G, Weilbaecher K, Horstmann M, Rieman D, Fisher D & Fisher D. (2001). Linking osteopetrosis and pycnodysostosis: regulation of cathepsin K expression by the microphthalmia transcription factor family. Proc Natl Acad Sci U S A 98, 5798-5803.
Murphy ME & Kehrer JP. (1989). Oxidative stress and muscular dystrophy. Chem Biol Interact 69, 101-173.
Murphy TC, Poppe C, Porter JE, Montine TJ & Picklo MJ, Sr. (2004). 4-Hydroxy-trans-2-nonenoic acid is a gamma-hydroxybutyrate receptor ligand in the cerebral cortex and hippocampus. J Neurochem 89, 1462-1470.
260

Murthi KK, Friedman LR, Oleinick NL & Salomon RG. (1993). Formation of DNA-protein cross-links in mammalian cells by levuglandin E2. Biochemistry 32, 4090-4097.
Murthi KK, Salomon RG & Sternlicht H. (1990). Levuglandin E2 inhibits mitosis and microtubule assembly. Prostaglandins 39, 611-622.
Nadkarni DV & Sayre LM. (1995). Structural definition of early lysine and histidine adduction chemistry of 4-hydroxynonenal. Chem Res Toxicol 8, 284-291.
Nakamura I, Saito S & Yamamoto Y. (2000). Hydrofurylation of Alkylidenecyclopropanes Catalyzed by Palladium. J Am Chem Soc 122, 2661-2662.
Neeper M, Schmidt AM, Brett J, Yan SD, Wang F, Pan YC, Elliston K, Stern D & Shaw A. (1992). Cloning and expression of a cell surface receptor for advanced glycosylation end products of proteins. J Biol Chem 267, 14998-15004.
Newcombe J, Li H & Cuzner ML. (1994). Low density lipoprotein uptake by macrophages in multiple sclerosis plaques: implications for pathogenesis. Neuropathol Appl Neurobiol 20, 152-162.
Ngooi T. (1989). A chemoenzymatic route to (-)-pyrenophorin. J Org Chem 54, 911.
Nielsen AL & Jorgensen AL. (2004). Self-assembly of the Cytoskeletal Glial Fibrillary Acidic Protein Is Inhibited by an Isoform-specific C Terminus. J Biol Chem 279, 41537-41545.
Noordermeer MA, Feussner I, Kolbe A, Veldink GA & Vliegenthart JF. (2000). Oxygenation of (3Z)-alkenals to 4-hydroxy-(2E)-alkenals in plant extracts: a nonenzymatic process. Biochem Biophys Res Commun 277, 112-116.
Noyori R, Sato T & Kobayashi H. (1983). C-Nucleoside synthesis. 18. The Baeyer-Villiger oxidation of 8-oxabicyclo[3.2.1]octan-3-ones. Substituent effects on the regioselectivity. Bull Chem Soc Jpn 56, 2661-2679.
Nozoe S, Hirai K, Tsuda K, Ishibashi K, Shirasaka M & Grove JF. (1965). Structure of pyrenophorin. Tetrahedron Lett, 4675-4677.
Nugteren D & Christ-Hazelhof E. (1980). Chemical and enzymic conversions of the prostaglandin endoperoxide PGH2. Adv Prostaglandin Thromboxane Res 6, 129-137.
Nugteren D & Hazelhof E. (1973). Isolation and properties of intermediates in prostaglandin biosynthesis. Biochim Biophy Acta 326, 448-461.
O'Banion MK, Winn VD & Young DA. (1992). cDNA cloning and functional activity of a glucocorticoid-regulated inflammatory cyclooxygenase. Proc Natl Acad Sci U S A 89, 4888-4892.
261

Obrecht D & Weiss B. (1989). A new method for the preparation of (E)-3-acylprop-2-enoic acids. Helv Chim Acta 72, 117-122.
Ochiai M, Arimoto M & Fujita E. (1981). Umpolung of reactivity of allylsilane, allylgermane, and allylstannane via their reaction with thallium(III) salt. A new allylation reaction for aromatic compounds. Tetrahedron Lett 22, 4491-4494.
Ogawa T, Matson W, Beal M, Myers R, Bird E, Milbury P & Saso S. (1992). Kynurenine pathway abnormalities in Parkinson's disease. Neurology 42, 1702-1706.
Oishi S & Nelson SD. ( 1992). Evidence for the formation of heterocyclic arene oxides and a .gamma.-keto enal by reaction of menthofuran with dimethyldioxirane. J Org Chem 57, 2744-2747.
O'Neil JA, Lindell SD, Simpson TJ & Willis CL. (1994). A simple enantioselective synthesis of g-valerolactone. Tetrahedron: Asymm 5, 117-118.
O'Neill GP & Ford-Hutchinson AW. (1993). Expression of mRNA for cyclooxygenase-1 and cyclooxygenase-2 in human tissues. FEBS Lett 330, 156-160.
O'Neill JA, Lindell SD, Simpson TJ & Willis, Christine L. (1994). A simple enantioselective synthesis of [gamma]-valerolactone. Tetrahedron: Asymm 5, 117-118.
Oya T, Hattori N, Mizuno Y, Miyata S, Maeda S, Osawa T & Uchida K. (1999). Methylglyoxal modification of protein. Chemical and immunochemical characterization of methylglyoxal-arginine adducts. J Biol Chem 274, 18492-18502.
Oyster CW. (1999). The Human Eye Structure and Function. Sinauer Associates, Inc., Sunderland, Massachusetts.
Padgaonkar VA, Giblin FJ, Leverenz V & Reddy VN. (1994). Studies of H2O2-induced effects on cultured bovine trabecular meshwork cells. J Glaucoma 3, 123-131.
Panousis CG & Zuckerman SH. (2000). Interferon-gamma induces downregulation of Tangier disease gene (ATP-binding-cassette transporter 1) in macrophage-derived foam cells. Arterioscler Thromb Vasc Biol 20, 1565-1571.
Pappalardo P, Ehlinger E & Magnus P. (1982). Silicon in synthesis. 1-Trimethylsilyl-1-methoxy allene: a reagent for the direct conversion of aliphatic aldehydes into 2-trimethylsilyl furans. Tetrahedron Lett 23, 309-312.
Parthasarathy S, Quinn MT, Schwenke DC, Carew TE & Steinberg D. (1989). Oxidative modification of beta-very low density lipoprotein. Potential role in monocyte recruitment and foam cell formation. Arteriosclerosis 9, 398-404.
Peters W & Charo IF. (2001). Involvement of chemokine receptor 2 and its ligand, monocyte chemoattractant protein-1, in the development of atherosclerosis: lessons from knockout mice. Curr Opin Lipidol 12, 175-180.
262

Pfefferle C, Kempter C, Metzger JW & Fiedler HP. (1996). (E)-4-oxonon-2-enoic acid, an antibiotically active fatty acid produced by Streptomyces olivaceus Tu 4018. J Antibiot (Tokyo) 49, 826-828.
Pfister R, Haddox J & Sommers C. (1998). Injection of chemoattractants into normal cornea: a model of inflammation after alkali injury. Invest Ophthalmol Vis Sci 39, 1744-1750.
Podrez EA, Poliakov E, Shen Z, Zhang R, Deng Y, Sun M, Finton PJ, Shan L, Febbraio M, Hajjar DP, Silverstein RL, Hoff HF, Salomon RG & Hazen SL. (2002a). A novel family of atherogenic oxidized phospholipids promotes macrophage foam cell formation via the scavenger receptor CD36 and is enriched in atherosclerotic lesions. J Biol Chem 277, 38517-38523.
Podrez EA, Schmitt D, Hoff HF & Hazen SL. (1999). Myeloperoxidase-generated reactive nitrogen species convert LDL into an atherogenic form in vitro. J Clin Invest 103, 1547-1560.
Poli G & Schaur RJ. (2000). 4-Hydroxynonenal in the pathomechanisms of oxidative stress. IUBMB Life 50, 315-321.
Pryor WA & Porter NA. (1990). Suggested mechanisms for the production of 4-hydroxy-2-nonenal from the autoxidation of polyunsaturated fatty acids. Free Radic Biol Med 8, 541-543.
Quigley HA. (1996). Number of people with glaucoma worldwide. Br J Ophthalmol 80, 389-393.
Quigley HA. (1998). The search for glaucoma genes--implications for pathogenesis and disease detection. N Engl J Med 338, 1063-1064.
Rao AVR, Reddy KB & Dhar TGM. (1986). Synthesis of (+-)-ethyl (E)-7-acetoxy-4-oxo-2-octenoate: the key synthon of pyrenophorin. Ind J Chem, Section B: Org Chem Med Chem 25B, 1014-1016.
Rathahao E, Peiro G, Martins N, Alary J, Gueraud F & Debrauwer L. (2005). Liquid chromatography-multistage tandem mass spectrometry for the quantification of dihydroxynonene mercapturic acid (DHN-MA), a urinary end-metabolite of 4-hydroxynonenal. Anal Bioanal Chem 381, 1532-1539.
Reynolds G & Pearson S. (1989). Increased brain 3-hydroxykynurenine in Huntington's disease. The Lancet 2, 979-980.
Ritch R, Shields M & Krupin T. (1996). The Glaucomas, vol. 1. Mosby, St. Louis, MO.
Rocken C, Stix B, Bromme D, Ansorge S, Roessner A & Buhling F. (2001). A Putative Role for Cathepsin K in Degradation of AA and AL Amyloidosis. Am J Pathol 158, 1029-1038.
263

Rodriguez-Martinez E, Rugerio-Vargas C, Rodriguez AI, Borgonio-Perez G & Rivas-Arancibia S. (2004). Antioxidant effects of taurine, vitamin C, and vitamin E on oxidative damage in hippocampus caused by the administration of 3-nitropropionic acid in rats. Int J Neurosci 114, 1133-1145.
Rogler G, Aschenbrenner E, Gross V, Stange EF & Scholmerich J. (2000). Intracellular transport of high-density lipoprotein 3 in intestinal epithelial cells (Caco-2) is tubulin associated. Digestion 61, 47-58.
Ronsheim MD & Zercher CK. (2003). Ring Expansions of .beta.-Keto Lactones with Zinc Carbenoids: Syntheses of (+)-Patulolide A and (.+-.)-Patulolide B. J Org Chem 68, 1878-1885.
Ross R. (1999). Atherosclerosis--an inflammatory disease. N Engl J Med 340, 115-126.
Russell P & Johnson DH. (1996). Enzymes protective of oxidative damage present in all decades of life in the trabecular meshwork, as detected by two-dimensional gel electrophoresis protein maps. J Glaucoma 5, 317-324.
Russell P, Garland D & Epstein DL. (1989). Analysis of the proteins of calf and cow trabecular meshwork: development of a model system to study aging effects and glaucoma. Exp Eye Res 48, 251-260.
Saldabol NO, Popelis J & Slavinska V. (2002). Thiocyanation, halogenation, dehalogenation, transhalogenation, and nitration of 2-substituted 4-(2-furyl)thiazoles. Chem of Heterocyclic Compounds (New York, NY, United States)(Translation of Khimiya Geterotsiklicheskikh Soedinenii) 38, 873-881.
Salomon R. (1985). Prostaglandin endoperoxide reaction mechanisms and the discovery of levuglandins. Acc Chem Res 18, 294
Salomon RG & Miller DB. (1985). Levuglandins: isolation, characterization, and total synthesis of new secoprostanoid products from prostaglandin endoperoxides. Adv Prostaglandin Thromboxane Leukot Res 15, 323-326.
Salomon RG, Batyreva E, Kaur K, Sprecher DL, Schreiber MJ, Crabb JW, Penn MS, DiCorletoe AM, Hazen SL & Podrez EA. (2000a). Isolevuglandin-protein adducts in humans: products of free radical-induced lipid oxidation through the isoprostane pathway. Biochim Biophys Acta 1485, 225-235.
Salomon RG, Kaur K, Podrez E, Hoff HF, Krushinsky AV & Sayre LM. (2000). HNE-derived 2-pentylpyrroles are generated during oxidation of LDL, are more prevalent in blood plasma from patients with renal disease or atherosclerosis, and are present in atherosclerotic plaques. Chem Res Toxicol 13, 557-564.
Salomon RG, Miller DB, Zagorski MG & Coughlin DJ. (1984). Solvent induced fragmentation of prostaglandin endoperoxides. New aldehyde products from PGH 2 and a
264

novel intramolecular 1, 2-hydride shift during endoperoxide fragmentation in aqueous solution. J Am Chem Soc 106, 6049.
Salomon RG, Sha W, Brame C, Kaur K, Subbanagounder G, O'Neil J, Hoff HF & Roberts LJ, 2nd. (1999). Protein adducts of iso[4]levuglandin E2, a product of the isoprostane pathway, in oxidized low density lipoprotein. J Biol Chem 274, 20271-20280.
Salomon RG, Subbanagounder G, O'Neil J, Kaur K, Smith MA, Hoff HF, Perry G & Monnier VM. (1997). Levuglandin E2-protein adducts in human plasma and vasculature. Chem Res Toxicol 10, 536-545.
Salomon RG. (2005a). Distinguishing levuglandins produced through the cyclooxygenase and isoprostane pathways. Chem Phys Lipids 134, 1-20.
Salomon RG. (2005b). Levuglandins and isolevuglandins: stealthy toxins of oxidative injury. Antioxid Redox Signal 7, 185-201.
Sardar A, Bell J & Reynolds G. (1995). Increased concentrations of the neurotoxin 3-hydroxykynurenine in the frontal cortex of HIV-1-positive patients. J NeuroChem 64, 932-935.
Sarria AJ, Panini SR & Evans RM. (1992). A functional role for vimentin intermediate filaments in the metabolism of lipoprotein-derived cholesterol in human SW-13 cells. In J Biol Chem, pp. 19455-19463.
Sayre LM, Arora PK, Iyer RS & Salomon RG. (1993). Pyrrole formation from 4-hydroxynonenal and primary amines. Chem Res Toxicol 6, 19-22.
Sayre LM, Sha W, Xu G, Kaur K, Nadkarni D, Subbanagounder G & Salomon RG. (1996). Immunochemical evidence supporting 2-pentylpyrrole formation on proteins exposed to 4-hydroxy-2-nonenal. Chem Res Toxicol 9, 1194-1201.
Sayre LM, Zelasko DA, Harris PL, Perry G, Salomon RG & Smith MA. (1997). 4-Hydroxynonenal-derived advanced lipid peroxidation end products are increased in Alzheimer's disease. J Neurochem 68, 2092-2097.
Schneider C, Tallman KA, Porter NA & Brash AR. (2001a). Two distinct pathways of formation of 4-hydroxynonenal. Mechanisms of nonenzymatic transformation of the 9- and 13-hydroperoxides of linoleic acid to 4-hydroxyalkenals. J Biol Chem 276, 20831-20838.
Schori H, Kipnis J, Yoles E, WoldeMussie E, Ruiz G, Wheeler LA & Schwartz M. (2001). Vaccination for protection of retinal ganglion cells against death from glutamate cytotoxicity and ocular hypertension: implications for glaucoma. Proc Natl Acad Sci U S A 98, 3398-3403.
265

Schreck R, Rieber P & Baeuerle PA. (1991). Reactive oxygen intermediates as apparently widely used messengers in the activation of the NF-kappa B transcription factor and HIV-1. Embo J 10, 2247-2258.
Schuettauf F, Quinto K, Naskar R & Zurakowski D. (2002). Effects of anti-glaucoma medications on ganglion cell survival: the DBA/2J mouse model. Vision Res 42, 2333-2337.
SDBS Web : http://www.aist.go.jp/RIODB/SDBS/ (National Institute of Advanced Industrial Science and Technology.
Sekiguchi J, Kuroda H, Yamada Y & Okada H. (1985). Structure of patulolide A, a new macrolide from Penicillium urticae mutants. Tetrahedron Lett 26, 2341-2342.
Sell DR, Lane MA, Johnson WA, Masoro EJ, Mock OB, Reiser KM, Fogarty JF, Cutler RG, Ingram DK, Roth GS & Monnier VM. (1996). Longevity and the genetic determination of collagen glycoxidation kinetics in mammalian senescence. Proc Natl Acad Sci U S A 93, 485-490.
Selley ML, Bartlett MR, McGuiness JA, Hapel AJ & Ardlie NG. (1989). Determination of the lipid peroxidation product trans-4-hydroxy-2-nonenal in biological samples by high-performance liquid chromatography and combined capillary column gas chromatography-negative-ion chemical ionisation mass spectrometry. J Chromatogr 488, 329-340.
Serhan CN & Ward PA, ed. (1999). Molecular and Cellular Basis of Inflammation. Humana Press, Totowa, New Jersey.
Shacter E. (2000). Quantification and significance of protein oxidation in biological samples. Drug Metab Rev 32, 307-326.
Shamsi FA, Lin K, Sady C & Nagaraj RH. (1998). Methylglyoxal-derived modifications in lens aging and cataract formation. Invest Ophthalmol Vis Sci 39, 2355-2364.
Shet J, Desai V & Tilve S. (2004). Domino primary alcohol oxidation-Wittig reaction: Total synthesis of ABT-418 and (E)-4-Oxonon-2-enoic acid. Synthesis, 1859-1863.
Smith JD, Trogan E, Ginsberg M, Grigaux C, Tian J & Miyata M. (1995). Decreased atherosclerosis in mice deficient in both macrophage colony-stimulating factor (op) and apolipoprotein E. Proc Natl Acad Sci U S A 92, 8264-8268.
Sowka J. (2004). Pigment dispersion syndrome and pigmentary glaucoma. Optometry 75, 115-122.
Srivastata SK, Awasthi S, Wang L, Bhatnagar A, Awasthi YC & Ansari NH. (1996). Attenuation of 4-hydroxynonenal-induced cataractogenesis in rat lens by butylated hydroxytoluene. Curr Eye Res 15, 749-754.
266

Stadtman ER. (1992). Protein oxidation and aging. Science 257, 1220-1224.
Staniszewska MM & Nagaraj RH. (2005). 3-Hydroxykynurenine-mediated Modification of Human Lens Proteins: STRUCTURE DETERMINATION OF A MAJOR MODIFICATION USING A MONOCLONAL ANTIBODY. J Biol Chem 280, 22154-22164.
Steen H & Mann M. (2001). Similarity between condensed phase and gas phase chemistry: fragmentation of peptides containing oxidized cysteine residues and its implications for proteomics. J Am Soc Mass Spectrom 12, 228-232.
Stillwell W & Wassall SR. (2003). Docosahexaenoic acid: membrane properties of a unique fatty acid. Chem Phys Lipids 126, 1-27.
Stoilov IR, Costa VP, Vasconcellos JP, Melo MB, Betinjane AJ, Carani JC, Oltrogge EV & Sarfarazi M. (2002). Molecular genetics of primary congenital glaucoma in Brazil. Invest Ophthalmol Vis Sci 43, 1820-1827.
Stone EM, Fingert JH, Alward WLM, Nguyen TD, Polansky JR, Sunden SLF, Nishimura D, Clark AF, Nystuen A, Nichols BE, Mackey DA, Ritch R, Kalenak JW, Craven ER & Sheffield VC. (1997). Identification of a Gene That Causes Primary Open Angle Glaucoma. Science 275, 668-670.
Streeten BW, Qi Y, Klintworth GK, Eagle RC, Jr., Strauss JA & Bennett K. (1999). Immunolocalization of beta ig-h3 protein in 5q31-linked corneal dystrophies and normal corneas. Arch Ophthalmol 117, 67-75.
Subbanagounder G, Salomon RG, Murthi KK, Brame C & Roberts LJ. (1997). Total Synthesis of Iso[4]-levuglandin E2. J Org Chem 62, 7658-7666.
Sugamoto K, Matsushita Y-i & Matsui T. (1998). Direct hydroperoxygenation of conjugated olefins catalyzed by cobalt(II) porphyrin. J Chem Soc, Perkin Trans 1: Org Bio-Org Chem, 3989-3998.
Sun AY & Chen YM. (1998). Oxidative stress and neurodegenerative disorders. J Biomed Sci 5, 401-414.
Sun M, Deng Y, Batyreva E, Sha W & Salomon RG. (2002). Novel bioactive phospholipids: practical total syntheses of products from the oxidation of arachidonic and linoleic esters of 2-lysophosphatidylcholine(1). J Org Chem 67, 3575-3584.
Sun M. (2004). Synthetic and Mechanistic Studies of Lipid Peroxidation in vitro and in vivo. In Department of Chemistry. Case Western Reserve University, Cleveland, Ohio.
Surgucheva I, McMahan B, Ahmed F, Tomarev S, Wax MB & Surguchov A. (2002). Synucleins in glaucoma: implication of gamma-synuclein in glaucomatous alterations in the optic nerve. J Neurosci Res 68, 97-106.
267

Svitkin YV, Pause A, Haghighat A, Pyronnet S, Witherell G, Belsham GJ & Sonenberg N. (2001). The requirement for eukaryotic initiation factor 4A (elF4A) in translation is in direct proportion to the degree of mRNA 5' secondary structure. RNA 7, 382-394.
Szweda LI, Uchida K, Tsai L & Stadtman ER. (1993). Inactivation of glucose-6-phosphate dehydrogenase by 4-hydroxy-2-nonenal. Selective modification of an active-site lysine. J Biol Chem 268, 3342-3347.
Tak PP, Zvaifler NJ, Green DR & Firestein GS. (2000). Rheumatoid arthritis and p53: how oxidative stress might alter the course of inflammatory diseases. Immunology Today 21, 78-82.
Takeya H. (1995). Photochemical oxidation process for preparing water-soluble org oxides such as 5-aminolevulinic acid. In Eur Pat Appl, pp. 8 pp. (Cosmo Research Institute, Japan; Cosmo Oil Co., Ltd.). Ep.
Tamm ER, Russell P, Johnson DH & Piatigorsky J. (1996). Human and monkey trabecular meshwork accumulate alpha B-crystallin in response to heat shock and oxidative stress. Invest Ophthalmol Vis Sci 37, 2402-2413.
Tapiero H, Ba GN, Couvreur P & Tew KD. (2002). Polyunsaturated fatty acids (PUFA) and eicosanoids in human health and pathologies. Biomed Pharmacother 56, 215-222.
Tezel G, Hernandez R & Wax MB. (2000). Immunostaining of heat shock proteins in the retina and optic nerve head of normal and glaucomatous eyes. Arch Ophthalmol 118, 511-518.
Tezel G, Yang X & Cai J. (2005). Proteomic identification of oxidatively modified retinal proteins in a chronic pressure-induced rat model of glaucoma. Invest Ophthalmol Vis Sci 46, 3177-3187.
Tripathi RC, Li J, Chan WF & Tripathi BJ. (1994). Aqueous humor in glaucomatous eyes contains an increased level of TGF-beta 2. Exp Eye Res 59, 723-727.
Uchida K, Toyokuni S, Nishikawa K, Kawakishi S, Oda H, Hiai H & Stadtman ER. (1994). Michael addition-type 4-hydroxy-2-nonenal adducts in modified low-density lipoproteins: markers for atherosclerosis. Biochemistry 33, 12487-12494.
Uchida K. (2003). 4-Hydroxy-2-nonenal: a product and mediator of oxidative stress. Prog Lipid Res 42, 318-343.
Uchida K. (2003). Histidine and lysine as targets of oxidative modification. Amino Acids 25, 249-257.
Van Heijst JWJ, Niessen HWM, Hoekman K & Schalkwijk CG. (2005). Advanced Glycation End Products in Human Cancer Tissues: Detection of N{epsilon}-(Carboxymethyl)lysine and Argpyrimidine. Ann NY Acad Sci 1043, 725-733.
268

van Kuijk FJ, Siakotos AN, Fong LG, Stephens RJ & Thomas DW. (1995). Quantitative measurement of 4-hydroxyalkenals in oxidized low-density lipoprotein by gas chromatography-mass spectrometry. Anal Biochem 224, 420-424.
Vazquez S, Aquilina JA, Jamie JF, Sheil MM & Truscott RJW. (2002). Novel Protein Modification by Kynurenine in Human Lenses. J Biol Chem 277, 4867-4873.
Vazquez S, Garner B, Sheil M & Truscott R. (2000). Characterisation of the major autoxidation products of 3-hydroxykynurenine under physiological conditions. Free Radical Res 32, 11-23.
Veach J. (2004). Functional dichotomy: glutathione and vitamin E in homeostasis relevant to primary open-angle glaucoma. Br J Nutr 91, 809-829.
Verdejo C, Marco P, Renau-Piqueras J & Pinazo-Duran MD. (1999). Lipid peroxidation in proliferative vitreoretinopathies. Eye 13 ( Pt 2), 183-188.
Verhasselt V, Goldman M & Willems F. (1998). Oxidative stress up-regulates IL-8 and TNF-&agr; synthesis by human dendritic cells. Eur J Immunol 28, 3886-3890.
Vincent AL, Billingsley G, Buys Y, Levin AV, Priston M, Trope G, Williams-Lyn D & Heon E. (2002). Digenic inheritance of early-onset glaucoma: CYP1B1, a potential modifier gene. Am J Hum Genet 70, 448-460.
Vitek MP, Bhattacharya K, Glendening JM, Stopa E, Vlassara H, Bucala R, Manogue K & Cerami A. (1994). Advanced glycation end products contribute to amyloidosis in Alzheimer disease. Proc Natl Acad Sci U S A 91, 4766-4770.
Wagberg M, Jansson AH, Westerlund C, Ostlund-Lindqvist AM, Sarnstrand B, Bergstrand H & Pettersson K. (2001). N,N'-diacetyl-L-cystine (DiNAC), the disulphide dimer of N-acetylcysteine, inhibits atherosclerosis in WHHL rabbits: evidence for immunomodulatory agents as a new approach to prevent atherosclerosis. J Pharmacol Exp Ther 299, 76-82.
Walton HM. (1957). Potential antimicrobial agents. I. Alkyl 4-oxo-2-alkenoates. J Org Chem 22, 308-312.
Wang CC & Tsou CL. (1993). Protein disulfide isomerase is both an enzyme and a chaperone. Faseb J 7, 1515-1517.
Wang N, Chintala SK, Fini ME & Schuman JS. (2001). Activation of a tissue-specific stress response in the aqueous outflow pathway of the eye defines the glaucoma disease phenotype. Nat Med 7, 304-309.
Williams P & Eugene L. ( 1981). Enedione-functionalized macrocycles via oxidative ring opening of furans. J Org Chem 46(21), 4143-4147.
269

Wilson MR, Mansour M, Atud AE, Casey R, Fobi G, Moukouri E, Alemayehu W, Martone JF & Ross-Degnan D. (1996). A population-based study of xerophthalmia in the extreme North Province of Cameroon, West Africa. Arch Ophthalmol 114, 464-468.
Winyard PG, Tatzber F, Esterbauer H, Kus ML, Blake DR & Morris CJ. (1993). Presence of foam cells containing oxidised low density lipoprotein in the synovial membrane from patients with rheumatoid arthritis. Ann Rheum Dis 52, 677-680.
Witztum JL & Berliner JA. (1998). Oxidized phospholipids and isoprostanes in atherosclerosis. Curr Opin Lipidol 9, 441-448.
Xu G & Sayre LM. (1998). Structural characterization of a 4-hydroxy-2-alkenal-derived fluorophore that contributes to lipoperoxidation-dependent protein cross-linking in aging and degenerative disease. Chem Res Toxicol 11, 247-251.
Xu G, Liu Y & Sayre LM. (2000). Polyclonal antibodies to a fluorescent 4-hydroxy-2-nonenal (HNE)-derived lysine-lysine cross-link: characterization and application to HNE-treated protein and in vitro oxidized low-density lipoprotein. Chem Res Toxicol 13, 406-413.
Xu G. (1998). In Department of Chemistry. Case Western Reserve University, Cleveland.
Yan DB, Trope GE, Ethier CR, Menon IA & Wakeham A. (1991). Effects of hydrogen peroxide-induced oxidative damage on outflow facility and washout in pig eyes. Invest Ophthalmol Vis Sci 32, 2515-2520.
Yan SD, Chen X, Schmidt AM, Brett J, Godman G, Zou YS, Scott CW, Caputo C, Frappier T, Smith MA & et al. (1994). Glycated tau protein in Alzheimer disease: a mechanism for induction of oxidant stress. Proc Natl Acad Sci U S A 91, 7787-7791.
Yan X, Tezel G, Wax MB & Edward DP. (2000). Matrix metalloproteinases and tumor necrosis factor alpha in glaucomatous optic nerve head. Arch Ophthalmol 118, 666-673.
Yang J, Neufeld A, Zorn M & Hernandez M. (1993). Collagen type I mRNA levels in cultured human lamina cribrosa cells: effects of elevated hydrostatic pressure. Exp Eye Res 56, 567-574.
Yildirim O, Ates NA, Ercan B, Muslu N, Unlu A, Tamer L, Atik U & Kanik A. (2004). Role of oxidative stress enzymes in open-angle glaucoma. Eye, 1-4.
Yoritaka A, Hattori N, Uchida K, Tanaka M, Stadtman ER & Mizuno Y. (1996). Immunohistochemical detection of 4-hydroxynonenal protein adducts in Parkinson disease. Proc Natl Acad Sci U S A 93, 2696-2701.
Yoshida Y, Chiba T, Tokunaga F, Kawasaki H, Iwai K, Suzuki T, Ito Y, Matsuoka K, Yoshida M, Tanaka K & Tai T. (2002). E3 ubiquitin ligase that recognizes sugar chains. Nature (London) 418, 438-442.
270

Yu J-Q & Corey EJ. (2003). A Mild, Catalytic, and Highly Selective Method for the Oxidation of a,b-Enones to 1,4-Enediones. J Am Chem Soc 125, 3232-3233.
Zagol-Ikapitte I, Masterson TS, Amarnath V, Montine TJ, Andreasson KI, Boutaud O & Oates JA. (2005). Prostaglandin H(2)-derived adducts of proteins correlate with Alzheimer's disease severity. J Neurochem 94, 1140-1145.
Zarbin MA. (2004). Current Concepts in the Pathogenesis of Age-Related Macular Degeneration. Arch Ophthalmol 122, 598-614.
Zarkovic K. (2003a). 4-hydroxynonenal and neurodegenerative diseases. Mol Aspects Med 24, 293-303.
Zhang B, Wu KF, Cao ZY, Rao Q, Ma XT, Zheng GG & Li G. (2004). IL-18 increases invasiveness of HL-60 myeloid leukemia cells: up-regulation of matrix metalloproteinases-9 (MMP-9) expression. Leuk Res 28, 91-95.
Zhou L, Li Y & Yue BY. (1999). Oxidative stress affects cytoskeletal structure and cell-matrix interactions in cells from an ocular tissue: the trabecular meshwork. J Cell Physiol 180, 182-189.
Zhou X, Li F, Kong L, Tomita H, Li C & Cao W. (2005). Involvement of inflammation, degradation and apoptosis in a mouse model of glaucoma. J Biol Chem, M502641200.
271




















