
Periphyton and phytoplankton associated with the tropical carnivorous plant Utricularia foliosa
Upload
independentCategory
view
1download
0
Hydrobiologia 325 : 1 8 3 -192, 1996 .
183© 1996 Kluwer Academic Publishers. Printed in Belgium.
Algal seston and periphyton distribution along a stream linking a chain oflakes on the Canadian Shield
Antonella CattaneoDepartement de sciences biologiques, Universite de Montreal, C . P. 6126, succ. Centre Ville Montreal,Quebec, H3C 3J7, Canada
Received 15 June 1995 ; in revised form 19 September 1995 ; accepted 29 November 1995
Key words : periphyton, seston, stream, impoundment, Quebec, lake outlet, chlorophyll
Abstract
The effect of natural impoundments on the longitudinal distribution of summer sestonic and periphytic algal biomasswas assessed in a stream linking a chain of lakes in the Laurentian mountains of Quebec . Algal seston biomasswas higher at lake outlets but declined rapidly downstream . Periphyton on gravel generally had larger biomasses atlake outlets, but on boulders, the dominant substratum, periphyton was unaffected by distance from the upstreamlake. However, the only lake in the set larger than 100 ha, Lac Achigan, had a significant positive effect on thebiomass of all the algal assemblages . Periphyton on boulders was significantly correlated with the distance fromthe headwater. The longitudinal trend in periphyton biomass was unimodal, with the peak around 4 km from theheadwater. When the effect of lakes was statistically removed, algal seston distribution also followed a unimodallongitudinal trend parallel to that of periphyton . Simple geomorphic variables like distance downstream from thenearest lake and distance from headwater account for a significant portion of the variation of algal seston (52%)and periphyton (64%) in this stream .
Introduction
The River Continuum Concept (Vannote et al ., 1980)describes general changes in stream community distri-bution along the stream's length . Because natural andman-made impoundments are common on most rivers,a corollary, the Serial Discontinuity Concept, has beenproposed to accommodate shifts in stream variablesrelated to the presence of impoundments (Ward &Stanford, 1983) . The application of these conceptsshould be especially pertinent to streams on the Cana-dian Shield where lakes often lie in chains linked bylengths of stream. These natural impoundments mayaffect the physical, chemical, and biological character-istics downstream, altering the longitudinal continuumof stream communities .
Although there is general agreement that sestonincreases within an impoundment, the downstream fateof suspended algae is less clear. They could be trapped
* Contribution No . 400 of the Groupe d'Ecologie des Eauxdouces de 1'Universite de Montreal .
by the aquatic vegetation (Chandler, 1937), ingestedby invertebrate filter feeders that are usually abundantat lake outlets (Mackay & Waters, 1986 : Morin &Peters, 1988; Robinson & Minshall, 1990 ; Richardson& Mackay, 1991), or damaged by abrasion producedby the strong current .
The effect of the lakes on stream periphyton devel-opment is debatable. Benthic algal proliferations havebeen observed downstream from deep release dams(Dufford et al ., 1987; Marcus, 1980), where nutri-ents concentrations are higher and currents and tem-peratures are more stable than in unregulated sections(Lowe, 1979 ; Richardson & Mackay, 1991) . On theother hand, outlets of lakes or impoundments with sur-face release could be nutrient deficient, resulting inlower benthic biomass than in sections farther fromlakes (Newbold, 1987) .
Existing descriptions of longitudinal trends in algalseston and periphyton usually apply only to a large spa-tial scale . In general, benthic algae reach their maxi-mum development in mid reaches whereas phytoplank-

184
ton becomes prevalent only in eutrophic large orderrivers (Cushing et al ., 1983 ; Minshall et al., 1983) .Nevertheless, it is possible that changes in stream com-munities would be detectable on the scale of a fewkilometres .
This study measures the effect of natural impound-ments on the longitudinal patterns of algal biomass .An apparently pristine stream connecting many lakesin the Laurentian mountains of Quebec provides theopportunity to repeat the measurement of the effectof lakes at increasing distances from the headwater .In particular, I determine the effect of distance down-stream from a lake and distance from the headwateron the distribution of stream seston and periphyton . Ifthese simple geomorphic variables are related to peri-phyton and seston biomass, they could be useful inmodelling the distribution of stream primary produc-ers.
Methods
Rivi6re de L'Achigan flows through the Lower Lau-rentian mountains (Quebec) and is a tributary of theSt. Lawrence River. The headwater (Lac Pilon) is aground-water fed lake that receives only intermittentsurface inflows . The stream section considered in thisstudy (45 ° 59'N, 74 ° 01'W) is about 15 km long,extending from the Lac Pilon to Lac de l'Achigan andlinking a chain of small lakes (Figure 1) . In summer,these lakes release epilimnetic water to the stream out-lets. Only two tributaries have some relevance ; oneenters shortly upstream from Lac des Chutes and a larg-er one joins about 1 km upstream from Lac Achigan .Sampling stations were chosen upstream and down-stream of five of these lakes, for a total of 14 sites (Fig-ure 1 and Table 1). The outlets of Lac Cromwell andLac Corriveau were best suited for intensive sampling ;in each of these sections, I established four stationsat increasing distance from the lake (5, 65, 100-200,> 1000 m) . In this study, I refer to the distance of eachstation from the headwater Lac Pilon as `distance fromheadwater' ; `distance downstream from lake' refersinstead to the distance from the nearest upstream lake .
All stations were in unshaded riffles . The streamsubstratum was dominated by boulders (15-<100 cmdiameter) with some gravel (2-4 cm diameter) inter-posed among them. The bedrock was Precambriananorthosite (McGerrigle, 1976). Because sand andcobbles were rare, and absent at some stations, Isampled only boulders and gravel. The sampling was
Table 1 . Stream order, distance from headwater, and distance down-stream from a lake for the fourteen stations . The lakes upstream fromthe stations are listed with their surface area in parentheses.
Table 2 . Physico-chemical characteristics of the sectionof Riviere Achigan considered in this study. The meanand the range of values observed at the fourteen stationsduring the three summer samplings (n = 42) are listed .
repeated three times, in May, June, and July of 1991 .At each sampling, one sample of water was collect-ed for subsequent seston and chemical analyses . Eightreplicate samples of gravel and boulders were collect-ed haphazardly within 5 m of the point where the watersample was collected . Gravel was gently lifted by handfrom the water and stored in plastic bags . Fragmentsof boulder surface were chipped off with hammer andchisel and placed in plastic bags . At times, some moss-es were intermingled with periphyton . Because of theirclose physical connection and their similar function as
Station Streamorder
Distancefromsource(m)
Distancedownstreamlake(m)
Lake andsurface area
(ha)
1 1 60 60 Pilon (31)2 2 1460 10 Long (18)3 2 4860 400 Quatre (7)4 2 5460 5 Cromwell (9)5 2 5515 60 Cromwell (9)6 2 5580 125 Cromwell (9)7 2 6460 1000 Cromwell (9)8 2 6820 5 Corriveau (6)9 2 6880 65 Corriveau (6)
10 2 6980 165 Corriveau (6)11 2 9020 2200 Corriveau (6)12 2 9620 200 Des Chutes (3)13 3 11620 1780 Des Chutes (3)14 3 15620 5 Achigan (529)
Parameter Mean Range
Current (cm s -1 ) 13 1-26Depth (cm) 18 7-35Color (mg Pt 1 -1 ) 24 .6 2 .4-53 .1pH 6.69 5 .95-8 .02NH4-N (µg 1- ') 42 .9 20.6-102 .5N01 -N (µg 1-' ) 59 .3 5 .4-203N02-N (µg 1-1 ) 3 .6 0 .8-8 .6Total N (µg 1 -1 ) 262 128-518Total dissolved P (µg 1 -1 ) 3 .1 0 .2-10 .4Total P (µg 1 -1 ) 7 .3 1 .3-57 .2
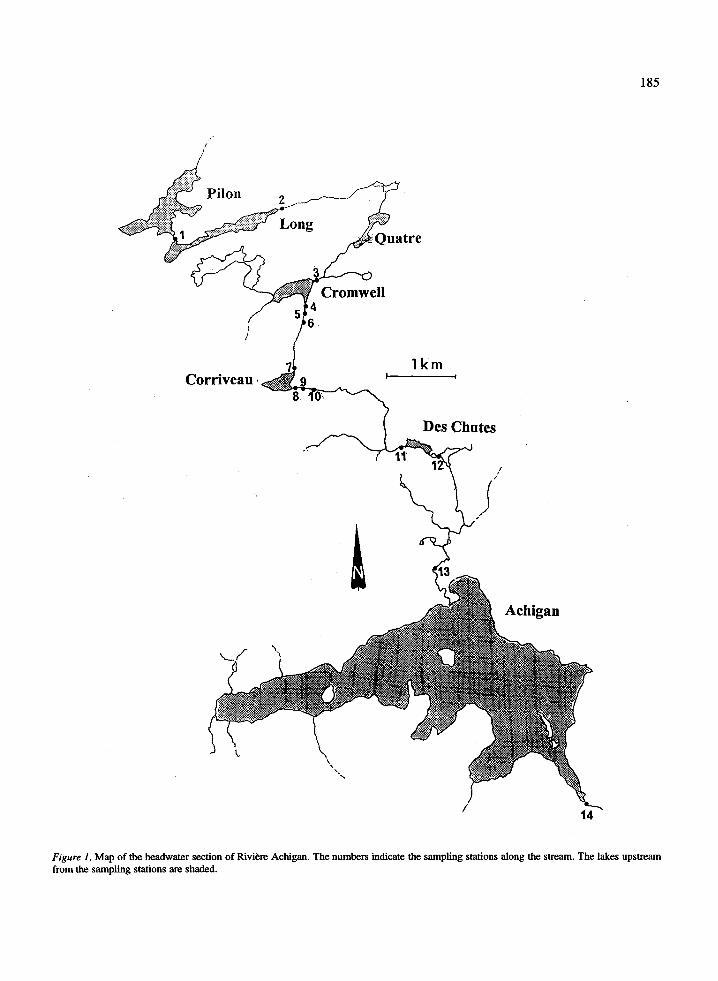
1 85
Figure 1 . Map of the headwater section of RiviBre Achigan . The numbers indicate the sampling stations along the stream . The lakes upstreamfrom the sampling stations are shaded .

186
Table 3. Multiple regression analyses of the biomass (chlorophyll a) of seston and of periphyton on gravel and on boulders . The variables testedwere: distance downstream from lake (DL ; m), distance from headwater (DH ; m), lake surface (S ; ha), color (Col ; mg Pt 1 -1 ), and ammoniaconcentration (NH4-N ; pg 1 -1 ) . The regressions were calculated using the data from all stations (all) and then repeated omitting Lac Achigan'soutlet (station 14 ; - Ach .) . Partial coefficients of determination (R2) and probability levels (* p<0.05, ** p<0.01, and *** p<0 .001) are listedunder each variable . For all the regressions, p<0 .0001 . @ In this equation, 1/s was used instead of S .
primary producers, I did not attempt to separate themfrom the rest of the sample. The samples were keptcold, and then frozen 3-4 h later upon arriving at the`Station Biologique des Laurentides' . Current veloci-ty, measured with a Pygmy current meter held at about0.6 of total depth, and water depth were recorded closeto the location of each periphyton replicate .
Periphyton and algal seston biomasses were esti-mated as chlorophyll a concentration . Gravel and boul-der fragments were extracted in 96% ethanol for 24 hand chlorophyll a levels were determined spectropho-tometrically (Bergman & Peters, 1980) . The colonizedsurface of the boulder fragments was determined bycarefully covering the colonized area with aluminiumfoil of known weight per unit area and by convertingweight of foil to surface area. For gravel, the totalsurface was estimated from three diameters measuredalong the three main axes (Graham et al ., 1988). Toestimate the colonized surface, I considered that, onaverage, only 2/3 of the calculated total gravel surfacewas available for algal growth, for 1/3 was in con-tact with the stream bed (Biggs & Close, 1989) . Ses-ton chlorophyll was estimated by filtering 1 1 of waterthrough Gelman glass fiber filters (1 .2 µm porosity) .These filters were kept frozen until analysis, when thechlorophyll was extracted in 96% ethanol and mea-sured spectrophotometrically .
Various physico-chemical variables were analyzed(Table 2). Color was determined by measuring theabsorbance at 440 nm of water samples filtered throughGelman glass fibre filters ; absorbances were trans-
formed to standard Hazen color units following Bowl-ing et al. (1986). Unfiltered samples were used todetermine total phosphorus and total nitrogen concen-trations ; total dissolved phosphorus, nitrate, nitrite,and ammonia were measured after filtration throughprerinsed 0.45 tm Millipore filters . All these analyseswere done with a Technicon autoanalyzer. Total anddissolved phosphorus were analyzed with the ascorbicacid modification of the molybdenum blue technique(Strickland & Parson, 1972) preceded by digestionunder pressure with potassium persulfate (Menzel &Corwin, 1965) . Total nitrogen was determined after awet-oxidation with peroxydisulfate digestion (Raim-bault & Slawyk, 1991) . The analyses of the otherchemicals followed standard methods (APHA, 1989) .
The data were analyzed by simple and multipleregression . Curvilinear regressions with quadratic vari-ables (square of the distance from headwater) were alsocalculated to fit the longitudinal trend in algal biomass-es (Sokal & Rohlf, 1981) . Prior to analyses, the datawere transformed to achieve the most normal distribu-tion . Square root transformation was the most suitablefor seston and gravel periphyton biomasses, while peri-phyton biomass on boulders, nutrients, and distancefrom lake were normalized by a log transformation .The remaining variables did not need transformation .These analyses and a non-parametric test (Wilcoxonsigned rank test) were performed using a statisticalpackage (Statistix, version 4 .0, Analytical Software,St. Paul, Minnesota) .
Intercept Log DL DH DH' Col NH4-N S n R2 F
1) Sqrt seston (all) 0 .824 -2 .44E-01 1 .99E-04 -1 .40E-08 9.87E-03 1 .85E-03 42 0.58 12.5*** (0 .22)*** (0 .02)*** (0.24)** (0.06)* (0.04)*
2) Sqrt seston (- Ach .) 0 .884 -2 .42E-01 1 .94E-04 -1 .38E-08 9 .15E-03 39 0.58 14.1*** (0 .19)*** (0 .09)*** (0 .27)** (0.03)
3) Sqrt gravel (all) 1 .094 -1 .13E-01 9.40E-05 -5 .80E-09 - -2.572@ 42 0.39 7 .6*** (0 .22)* (0 .01)* (0 .04)* (0.12)***
4) Sqrt gravel (- Ach.) 3 .795 -5 .61E-01 39 0.27 14 .9*** 0.27)***
5) Log boulders (all) 1 .075 1 .68E-04 -1 .87E-08 - 5.53E-03 4.51E-03 42 0.68 23 .2*** (0 .07)*** (0 .40)*** (0.04)* (0 .17)***
6) Log boulders (- Ach.) 1 .212 1 .47E-04 -1 .77E-08 - 5.30E-03 - 39 0.69 29 .4(0 .26)*** (0 .40)*** (0.03)*
Results
Distance from lake
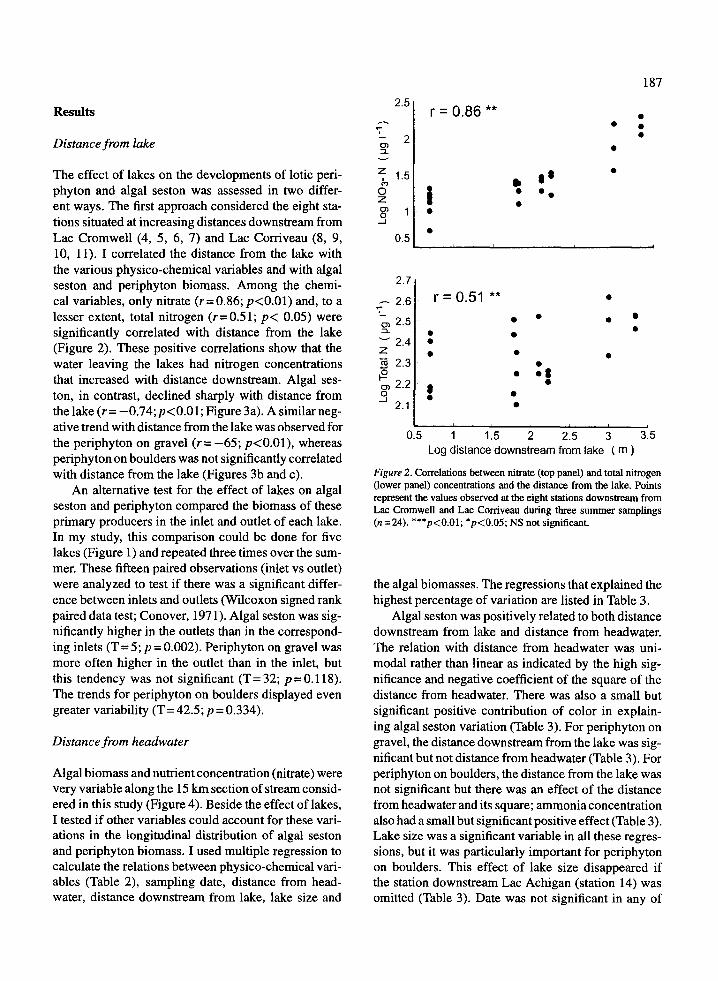
The effect of lakes on the developments of lotic peri-phyton and algal seston was assessed in two differ-ent ways . The first approach considered the eight sta-tions situated at increasing distances downstream fromLac Cromwell (4, 5, 6, 7) and Lac Corriveau (8, 9,10, 11) . I correlated the distance from the lake withthe various physico-chemical variables and with algalseston and periphyton biomass. Among the chemi-cal variables, only nitrate (r= 0 .86; p<0.01) and, to alesser extent, total nitrogen (r=0 .51 ; p< 0.05) weresignificantly correlated with distance from the lake(Figure 2). These positive correlations show that thewater leaving the lakes had nitrogen concentrationsthat increased with distance downstream . Algal ses-ton, in contrast, declined sharply with distance fromthe lake (r= -0.74;p<0.01 ; Figure 3a). A similar neg-ative trend with distance from the lake was observed forthe periphyton on gravel (r= -65 ; p<0.01), whereasperiphyton on boulders was not significantly correlatedwith distance from the lake (Figures 3b and c) .
An alternative test for the effect of lakes on algalseston and periphyton compared the biomass of theseprimary producers in the inlet and outlet of each lake .In my study, this comparison could be done for fivelakes (Figure 1) and repeated three times over the sum-mer. These fifteen paired observations (inlet vs outlet)were analyzed to test if there was a significant differ-ence between inlets and outlets (Wilcoxon signed rankpaired data test; Conover, 1971) . Algal seston was sig-nificantly higher in the outlets than in the correspond-ing inlets (T = 5 ; p = 0.002) . Periphyton on gravel wasmore often higher in the outlet than in the inlet, butthis tendency was not significant (T=32 ; p=0.118) .The trends for periphyton on boulders displayed evengreater variability (T = 42 .5 ; p = 0 .334) .
Distance from headwater
Algal biomass and nutrient concentration (nitrate) werevery variable along the 15 km section of stream consid-ered in this study (Figure 4) . Beside the effect of lakes,I tested if other variables could account for these vari-ations in the longitudinal distribution of algal sestonand periphyton biomass . I used multiple regression tocalculate the relations between physico-chemical vari-ables (Table 2), sampling date, distance from head-water, distance downstream from lake, lake size and
2 .5
M0z0) 1J
0 .5
2
2 .7- 2.6
a, 2 .52 .4z
m 2.3
a, 2 .20-' 2 .1
0.5
1
1 .5
2
2.5
3
3.5Log distance downstream from lake (m )
Figure 2 . Correlations between nitrate (top panel) and total nitrogen(lower panel) concentrations and the distance from the lake . Pointsrepresent the values observed at the eight stations downstream fromLac Cromwell and Lac Corriveau during three summer samplings(n=24). ***p<0 .01 ; *p<0 .05 ; NS not significant .
the algal biomasses . The regressions that explained thehighest percentage of variation are listed in Table 3 .
Algal seston was positively related to both distancedownstream from lake and distance from headwater.The relation with distance from headwater was uni-modal rather than linear as indicated by the high sig-nificance and negative coefficient of the square of thedistance from headwater. There was also a small butsignificant positive contribution of color in explain-ing algal seston variation (Table 3) . For periphyton ongravel, the distance downstream from the lake was sig-nificant but not distance from headwater (Table 3) . Forperiphyton on boulders, the distance from the lake wasnot significant but there was an effect of the distancefrom headwater and its square ; ammonia concentrationalso had a small but significant positive effect (Table 3) .Lake size was a significant variable in all these regres-sions, but it was particularly important for periphytonon boulders . This effect of lake size disappeared ifthe station downstream Lac Achigan (station 14) wasomitted (Table 3) . Date was not significant in any of
r = 0 .86 **
r = 0.51 **•
• •
•
11
1 87

1 88
0J
0.5
1
1 .5
2
2.5
3
3.5Log distance downstream from lake (m )
Figure 3 . Correlations between the biomass (as chlorophyll) of ses-ton (top panel), of periphyton on gravel (middle panel), or on boul-ders (lower panel) and the distance from the lake. Points represent thevalues observed at the eight stations downstream from Lac Cromwelland Lac Corriveau during three summer samplings (n = 24) .
the regressions indicating a stability of this system overthe summer .
To illustrate the longitudinal patterns in algalbiomass after accounting for the effect of lakes, Isolved equations 2 and 6 (Table 3) keeping the dis-tance from upstream lake, color (in equation 2), andammonia (in equation 6) constant at their average val-ues (431 m, 24.6 mg Pt 1 -t , 42.9 ltg 1 -t respectively) .
Figure 4 . The longitudinal trends of nitrates, seston, periphyton ongravel, and periphyton on boulders . The points represent the averageover the season at each station and the bars the standard error (n = 3) .The stations immediately downstream from a lake are representedby open circles ; all the other stations are represented by solid circles .
4-4.5 Gravel r=-0.65**E 4
E 3.5• •
~, 3L If 00 2.50L
2U •
•1 .501 I
.
I
1
.
I . I
N 2.2 - Boulders • r = - 0 .17 NS'E 2rnE 1 .8 t

60 I 1 .4
E~.- .0t
300C,201
0V_ In 10. 0
a ,0 0
2
4
6
8
10
12
140 .2
Distance from headwater (km )
Figure 5. Model of the longitudinal distribution of seston and peri-phyton biomass along the stream before entering Lac Achigan afterstatistically accounting for the effect of lakes . Curves were obtainedby fitting the equations in Table 3 keeping constant the distance fromthe lake .
The development of both algal seston and periphytonon boulders followed a unimodal trend with distancefrom headwater. Periphyton peaked after 4 km, where-as the maximum of algal seston was reached 2-3 kmfurther downstream (Figure 5) . Because periphyton ongravel was not correlated to distance from headwater,it is not represented in Figure 5. Among the physico-chemical variables measured, pH and color also exhib-ited a unimodal longitudinal trend (Figure 6) .
Discussion
The Serial Discontinuity Concept (Ward & Stanford,1983) predicts that impoundments disrupt the naturalcontinuum of abiotic and biotic stream characteristics .This disruption of longitudinal pattern was observedin this lake chain for the concentrations of nitrates andsestonic chlorophyll a (Figure 2 and 3) . Nitrates weredepleted in the water leaving the lakes, but their con-centration recovered about 1 km downstream probablydue to groundwater inputs. Hynes (1960) suggestedthat lakes act as a nutrient trap . This suggestion isconfirmed in this system at least for nitrogen . Theeffect was not significant for either soluble reactiveor total phosphorus, but the results could have beenconfused by some analytical variability encounteredin the analyses of such low concentration samples .Nutrient reduction downstream from impoundmentswith surface release has been observed in a SouthAfrican stream (Palmer & O' Keeffe, 1990), whereasdeep release dams usually discharge nutrient enrichedwater (Marcus, 1980) . The outlets in this study that
1 .2
z1 vr
0 .8 0°0
0 .6 0C
0 .4 aCO
8.5
8
7.5
0. 7
6.5
6
5 .5
0.25
0 .2
n 0 .15rnE
0 .1000 0.05
00
5
10
15
20Distance from headwater (km )
189
Figure 6 . pH (top panel) and color (lower panel) values observed atthe different stations along the stream . These variables were weak-ly but significantly correlated with distance from the headwater(DH) and its square (DH2) : pH=6.647-4.78E-05 DH +6.11E-09 DH2 , R2 =0.37, P=0.00001, n=42 ; Color= 16.05+3.57E-03DH - 2.58E-07 DH2 , R 2 =0.27, P=0 .002, n=42.
receive epilimnetic water are more comparable to sur-face release impoundments .
Sestonic chlorophyll a immediately below the lakeswas higher than in the inlets, but was reduced by 50%less than 100 m below the lake . Similar rates of declinehave been observed in streams elsewhere . Reif (1939)observed a 50% reduction of the biomass 180 m belowa lake outlet in Minnesota, and Ruttner (1956) mea-sured even faster decline rates (72% in the first 50 mand 90% after 1100 m) downstream from an Austri-an mountain lake . An exponential decrease in sestonicchlorophyll a was observed in a Montana lake outletbut only when discharge was less than 1 m3 s-1 (Vade-bancoeur, 1994). Macioleck & Tunzi (1968) report-ed a 60% decline after 400 m . I did not measure thebiomass of filter feeding invertebrates at my samplingsites. However, they were abundant in the outlet ofLac Quatre, a lake in the same chain (Morin & Harper,1986) and could account at least partly for the sestondecrease . Part of the sestonic algae may have settled tothe bottom. The slightly elevated biomass of periphy-

190
ton on gravel observed at lake outlets may reflect thisrecruitment .
In a model of phosphorus spiralling in streams,Newbold (1987) predicted that impoundments, byslowing the river flow, would discharge water rich inplankton but poor in nutrients and consequently reducethe downstream benthic biomass . In this study, nutrientdepletion was not translated into a depression of peri-phyton biomass . According to Bothwell (1989), thegrowth rate of thin periphyton assemblages is saturat-ed at nutrient concentrations much lower than thoserecorded in this system ; so gravel periphyton wasnot likely to be affected by nutrient depletion . Thisdepletion could be more severe for the thicker mats onboulders, but the nutrient-biomass relation in periphy-ton is often obscured by the effect of other variables(Cattaneo, 1987) . For example, longitudinal changesin nutrient concentration along a Tennessee woodlandstream affected periphyton alkaline phosphatase activ-ity and phosphorus content, but not its biomass andproductivity which were apparently controlled by her-bivory (Mulholland & Rosemond, 1992) . Colonizationfrom algal seston, as previously stated, could also bal-ance the negative effect of nutrient depletion . Whateverthe mechanism, in this system, the presence of lakesdid not affect periphyton biomass on boulders whichwere the dominant substrata in this stream . Because allour measurements were taken during the summer, theresults cannot be extrapolated to other seasons withoutfurther tests .
In all models of algal biomass, there was a signif-icant positive effect of lake size (Table 3) that disap-peared when the site downstream from Lac Achiganwas omitted . This lake has a surface area about 50times larger than the other lakes in this set (Table 1) .It was not possible to establish whether the size effectwas general or simply a reflection of some local pecu-liarities . The importance of lake size should be furtherexplored by sampling a series of outlets covering alarger range of lake size . Lake size could be relat-ed to retention time and consequently to the extent ofchanges in water chemistry . There is some indicationthat larger reservoirs affect downstream water chem-istry and transport more than smaller impoundments(Palmer & O' Keeffe, 1990) .
It is somewhat surprising that even at the smallscale of this study, where variations in stream widthand depth were minimal (Table 2), distance from head-water could explain such a significant portion (60%)of periphyton variation on boulders . Distance is nodoubt a surrogate variable for other important factors
regulating periphyton biomass . Among the variablesmeasured, color and pH showed longitudinal trends(Figure 6), but they were less effective, either aloneor together, than distance in predicting periphyton .Lithology appears homogenous in this basin (McGer-rigle, 1976), but bog development is more patchy.The presence of bogs could affect color, pH, andunmeasured variables like humic substances, DOC,and cation levels which are particularly important formoss distribution (Glime & Vitt, 1987) . The inputsfrom the tributary appear negligible in the upper sec-tion of the stream, but could have affected the chem-istry of the lower stations .
In this system, the periphyton growing on graveldiffered greatly from periphyton on boulders . Biomasson gravel was quite variable and unrelated to distancefrom headwater. Biomass on boulders was at least 5times larger than on gravel (Figure 4) . Mosses (Fonti-nalis sp .) and macroalgae (Draparnaldia sp ., Batra-chospermum sp ., andStiaonema sp .) were often promi-nent on boulders, but usually absent on gravel . Theselarge forms grow slowly and therefore need a more sta-ble substratum for their development . The gravel wasinterspersed among the boulders in crevices where cur-rent and consequently water renewal was slower, andwhere boulders may have provided shade . These dif-ferences in taxonomy and stream-bed position betweenthe assemblages could partly explain their differentresponses to the geomorphic variables like distancedownstream from lake and distance from headwater.
Naiman (1983) showed the importance of geomor-phic variables in predicting the development of differ-ent primary producers from headwaters to large rivers .My study indicates that simple geomorphic variablessuch as distance from headwater and downstream fromlake are also useful on a much smaller scale (15 km)where they account for more than 50% of the variationin algal seston and periphyton biomass. Because theyare simple and inexpensive to measure, these variablesshould be included in stream models . Distance fromlake is important in modelling algal seston and conse-quently the distribution of filter feeding invertebrates(Morin & Peters, 1988 ; Richardson & Mackay, 1991) .For periphyton, neither the models showing reductionin benthic algae (Newbold, 1987) or the observationsof enhancement downstream of deep release reservoirsapply to the outlets of these small lakes. A positiveeffect on periphyton biomass of large lakes was sug-gested by the Lac Achigan outlet, but this needs con-firmation. The significant relationship between algalbiomass and distance from headwater suggests that

exploration of likely correlates with distance may befruitful . Lithology, watershed slope, vegetation, andland use could be powerful variables for stream com-munities as they seem to be for lake plankton (Ras-mussen et al ., 1989; Wetzel, 1992; D'Arcy & Carig-nan, 1996) .
Acknowledgments
This work was funded by an operating grant from theNatural Sciences and Engineering Research Council ofCanada and a team grant from the FCAR funds of theQuebec Ministry of Education . Caroline Brazeau andMarie Christine Amireault assisted in the field and inthe laboratory. Peter Harper kindly shared his knowl-edge in selecting the sampling sites . Robert Peters com-mented on an earlier draft .
References
APHA, 1989. Standard methods for the examination of water andwastewater. 17th edition . American Public Health Association,Washington, D .C.
Bergman, M. & R . H . Peters, 1980. A simple reflectance methodfor the measurements of particulate pigment in lake water and itsapplication to phosphorus-chlorophyll-seston relationships . Can .J . Fish . aquat. Sci . 37 : 111-114 .
Biggs, B . J. F. & M . E . Close, 1989 . Periphyton biomass dynamicsin gravel bed rivers : the relative effects of flow and nutrients .Freshwat . Biol . 22 : 209-231 .
Bothwell, M. L., 1989 . Phosphorus-limited growth dynamics oflotic periphytic diatom communities : areal biomass and cellulargrowth rate responses . Can . J. Fish. aquat. Sci . 46 : 1293-1301 .
Bowling, L. C ., M . C . Steane & P. A. Tyler, 1986 . The spectral dis-tribution and attenuation of underwater irradiance in Tasmanianinland waters . Freshwat. Biol. 16 : 313-335 .
Cattaneo, A ., 1987. Periphyton in lakes of different trophy . Can. J .Fish. aquat . Sci. 44 : 296-303 .
Chandler, D. C., 1937 . Fate of typical lake plankton in streams . Ecol .Monogr. 16: 445-479 .
Conover, W. J ., 1971 . Practical nonparametric statistics. John Wileyand Sons, New York . 462 pp.
Cushing, C. E., K . W. Cummins, G. W. Minshall & R. L. Vannote,1983 . Periphyton, chlorophyll a, and diatoms of the Middle Forkof the Salmon River, Idaho . Holart . Ecol. 6 : 221-227 .
D'Arcy, P. & R . Carignan, 1996 . Influence of watershed topographyon water quality in the Shield lakes of southeastern Quebec . Can .J . Fish . aquat . Sci. (in press) .
Dufford, R . G ., H. J . Zimmermann, L . D. Cline & J . V. Ward, 1987 .Responses of epilithic algae to regulation of Rocky Mountainstreams . In J. F. Craig & J . B . Kemper (eds), Regulated streams.Advances in ecology. Plenum Press, New York: 383-390 .
Glime, J. M . & D . H . Vitt, 1987. A comparison of bryophyte speciesdiversity and niche structure of montane streams and streambanks . Can. J . Bot. 65 : 1824-1837 .
191
Graham, A . A ., D. J . McCaughan & F. S . McKee, 1988 . Measure-ment of surface area of stones . Hydrobiologia 157 : 85-87 .
Hynes, H . B . N ., 1960 . The biology of polluted waters . LiverpoolUniversity Press, Liverpool . 555 pp .
Lowe, R. L ., 1979 . Phytobenthic ecology and regulated streams .In J . V. Ward & J . A. Stanford (eds), The ecology of regulatedstreams . Plenum Press, New York: 25-34.
Maciolek, J . A. & M. G. Tunzi, 1968 . Microseston dynamics in asimple Sierra Nevada lake-stream system. Ecology 49 : 60-75 .
Mackay, R. J. & T. F. Waters, 1986 . Effects of small impoundmentson hydropsychid caddisfly production in Valley Creek, Minneso-ta. Ecology 67 : 1680-1686.
Marcus, M. D ., 1980 . Periphytic community response to chronicnutrient enrichment by a reservoir discharge. Ecology 61 : 387-399 .
McGerrigle, J . L ., 1976 . Region de Saint-Hippolyte/Saint-Adele(Quebec) . Minist6re des Richesse Naturelles, Ottawa. Rapportgeologique no. 179 .
Menzel, D. W. & H . Corwin, 1965 . The measurement of total phos-phorus based on the liberation of organically bound fractions bypersulfate oxidation . Limnol . Oceanogr. 10 : 280-282 .
Minshall, G. W., R . C . Petersen, K. W. Cummins, T. L . Bott,J . R . Sedell, C. E . Cushing & R . L . Vannote, 1983 . Interbiomecomparison of stream ecosystem dynamics . Ecol. Monogr. 53 :125 .
Morin, A. & P. P. Harper, 1986 . Phenologie et microdistribution desadultes et des larves de Trichopteres filtreurs dans un ruisseau desBasses Laurentides (Quebec) . Arch . Hydrobiol . 108 : 167-183 .
Morin, A . & R . H. Peters, 1988 . Effect of micro-habitat features,seston quality, and periphyton on the abundance of overwinteringblack fly larvae. Limnol . Oceanogr. 33 : 431-446.
Mulholland, P. J . & A . D . Rosemond, 1992 . Periphyton response tolongitudinal nutrient depletion in a woodland stream : evidenceof upstream-downstream linkage . J . N . am. benthol . Soc. 11 :405-419.
Naiman, R . J ., 1983 . A geomorphic approach for examining the roleof periphyton in large watersheds . In R . G . Wetzel (ed .), Periphy-ton of Freshwater Ecosystems . Developments in Hydrobiology17. Dr W. Junk Publishers, The Hague : 191-198 .
Newbold, J . D ., 1987 . Phosphorus spiralling in rivers and river-reservoir systems : implications of a model . In J. F. Craig &J . B . Kemper (eds), Regulated streams. Advances in ecology .Plenum Press, New York : 303-327 .
Palmer, R . W. & J . H . O'Keeffe, 1990 . Downstream effects ofimpoundments on the water chemistry of the Buffalo River (East-ern Cape), South Africa . Hydrobiologia 202 : 71-83 .
Raimbault, P. & G . Slawyk, 1991 . A semiautomatic, wet-oxidationmethod for the determination of particulate organic nitrogen col-lected on filters . Limnol. Oceanogr. 36: 405-408 .
Rasmussen, J . B ., L . Godbout & M . Schallenberg, 1989 . The humiccontent of lake water and its relationship to watershed and lakemorphometry. Limnol . Oceanogr. 34 : 1336-1343 .
Reif, C . B ., 1939 . The effect of stream conditions on lake plankton .Trans . am. Micros . Soc . 58 : 398-403 .
Richardson, J. S . & R . J. Mackay, 1991 . Lake outlets and the distri-bution of filter feeders : an assessment of hypotheses . Oikos 62 :370-380.
Robinson, C . T . & G . W. Minshall, 1990 . Longitudinal develop-ment of macroinvertebrate communities below oligotrophic lakeoutlets . Great Basin Naturalist 50 : 303-311 .
Rutmer, F., 1956 . Einige Beobachtungen fiber das Verhalten desPlanktons in Seeabfliissen . Osterreich botanical Zeitung 103 : 98-109 .

192
Sokal, R. R . & F. J . Rohlf, 1981 . Biometry. W. H. Freeman andCompany, New York. 859 pp .
Strickland, J . D . H. & T. R . Parson, 1972. A practical handbook ofseawater a nalysis . 2nd. ed . Bull. Fish. Res . B d Can. 167 : 310 pp .
Vadebancoeur, Y., 1994 . Longitudinal dynamics of seston concen-tration and composition in a lake outlet stream. J. N . am. benthol.Soc. 13 : 181-189 .
Vannote, R. L., G. W. Minshall, K . W. Cummins, J . R . Sedell &C. E . Cushing, 1980 . The river continuum concept . Can . J . Fish .
aquat . Sci . 37 : 130-137 .Ward, J . V. & J . A. Stanford, 1983 . The serial discontinuity concept
of lotic ecosystems . In T. D. Fontaine & S. M . Bartell (eds) . Thedynamics of lotic ecosystems . Ann Arbor Science Publishers,Ann Arbor, Michigan: 161-180 .
Wetzel, R. G ., 1992. Gradient-dominated ecosystems : sources andregulatory functions of dissolved organic matter in freshwaterecosystems . Hydrobiologia 229 : 181-198 .
Copyright © 2022 FDOKUMEN




















