
Alginate NiFe2O4 Nanoparticles Cryogel for Electrochemical ...
Upload
independentCategory
view
1download
0
SPINE Volume 31, Number 9, pp 959–966©2006, Lippincott Williams & Wilkins, Inc.
Platelet-Rich Plasma (PRP) Stimulates the ExtracellularMatrix Metabolism of Porcine Nucleus Pulposus andAnulus Fibrosus Cells Cultured in Alginate Beads
Koji Akeda, MD,*� Howard S. An, MD,* Rajeswari Pichika, PhD,*Mohamed Attawia, MD,§ Eugene J.-M. A. Thonar, PhD,*†‡Mary Ellen Lenz, MS,† Atsumasa Uchida, MD, PhD,� and Koichi Masuda, MD*†
Study Design. In vitro assessment of the effects ofplatelet-rich plasma on the extracellular matrix metabo-lism of porcine intervertebral disc cells.
Objectives. To determine whether platelet-rich plasmais effective in stimulating cell proliferation and extracellu-lar matrix metabolism by porcine disc cells cultured inalginate beads.
Summary of Background Data. Platelet-rich plasma isused to accelerate wound healing and tissue regenera-tion. Activated platelets release multiple growth factorsthat regulate cell proliferation, differentiation, and mor-phogenesis. Individual growth factors present in platelet-rich plasma have been demonstrated to affect the metab-olism of intervertebral disc cells.
Methods. Platelet-poor and platelet-rich plasma was iso-lated from fresh porcine blood using a commercially avail-able platelet concentration system. After preculture for 7days and serum starvation for 24 hours, the beads contain-ing nucleus pulposus and anulus fibrosus cells were thencultured for another 72 hours in serum-free medium, 10%fetal bovine serum, 10% platelet-poor plasma, or 10% plate-let-rich plasma. The synthesis of proteoglycans and colla-gen, the accumulation of proteoglycans, and the DNA con-tent were biochemically assessed.
Results. Platelet-rich plasma had a mild stimulatory ef-fect on cell proliferation of intervertebral disc cells. Platelet-rich plasma treatment significantly upregulated proteogly-can and collagen synthesis and proteoglycan accumulationwhen compared with platelet-poor plasma.
Conclusions. Platelet-rich plasma was effective in stim-ulating cell proliferation and extracellular matrix metabo-lism. The response to platelet-rich plasma was greaterin the case of anulus fibrosus cells than of nucleus
pulposus cells. The local administration of platelet-richplasma might stimulate intervertebral disc repair. In addi-tion, given the risks of using animal serum for tissue engi-neering, autologous blood may gain favor as a source ofgrowth factors and serum supplements needed for stimu-lating cells to engineer intervertebral disc tissues.
Key words: platelet-rich plasma, growth factor, interver-tebral disc, extracellular matrix, proteoglycan, collagen.Spine 2006;31:959–966
The relatively few cells that reside in the anulus fibrosus(AF) and nucleus pulposus (NP) maintain intervertebraldisc (IVD) matrix metabolism homeostasis. Althoughthe regulatory mechanisms are not well understood, nu-trition to the disc, cytokines, and growth factors arethought to play a role.1,2 Recent studies have shown thatgrowth factors can upregulate the production of matrixmolecules and also stimulate the accumulation of matrixconstituents during culture.3,4 The stimulation of cellmetabolism by growth factors positively affects the per-turbation of matrix maintenance even when the bio-chemical and biomechanical properties of disc tissues arecompromised.5 A recent study showed that the injectionof a growth factor into normal IVDs6 and the discs in arabbit disc degeneration model induced an increase indisc height,7 suggesting that growth factors may have atherapeutic use in the treatment of IVD degeneration. Inthose cases of IVD degeneration that exhibit advancedpathologic changes, apoptosis of IVD cells8 and struc-tural defects, such as an anulus tear, can be observed.
Several growth factors have been shown to positivelymodulate the metabolism of IVD cells. Transforminggrowth factor-� (TGF-�), insulin-like growth factor-1(IGF-1), platelet-derived growth factor (PDGF), and epi-dermal growth factor (EGF) stimulate cell proliferationand proteoglycan (PG) synthesis in vitro.9–12 PDGF alsoreduces the rate of cell apoptosis.11 More recently, Ma-suda et al3 and Yoon et al4 showed that bone morpho-genetic protein-7 (BMP-7, also known as osteogenic pro-tein-1, OP-1) and BMP-2 both enhance PG metabolismin IVD cells. Furthermore, as cited above, the injection ofBMP-7 was effective in increasing the disc height of de-generated rabbit IVDs.7 Therefore, the application ofgrowth factors via direct injection of recombinant proteinsinto the NP or the AF may be an effective therapeutic ap-proach in the treatment of IVD degeneration. For repair oflarger defects, alternative therapeutic approaches, such
From the Departments of *Orthopedic Surgery, †Biochemistry, and‡Internal Medicine, Rush Medical College at Rush University MedicalCenter, Chicago, IL; §DePuy Biologics, DePuy Spine Incorporation,Raynham, MA; and �Department of Orthopedic Surgery, Mie Univer-sity School of Medicine, Mie, Japan.Acknowledgment date: July 7, 2004. First revision date: April 29,2005. Acceptance date: June 1, 2005.Supported by a research grant from DePuy Spine Inc. and in part byNIH grants 2-P50-AR39239 and 1-P01-AR48152.The device(s)/drug(s) that is/are the subject of this manuscript is/are notFDA-approved or approved by corresponding national agency for thisindication.Corporate/Industry, Federal, and Institutional funds were received insupport of this work. Although one or more of the author(s) has/havereceived or will receive benefits for personal or professional use from acommercial party related directly or indirectly to the subject of thismanuscript, benefits will be directed solely to a research fund, founda-tion, educational institution, or other nonprofit organization which theauthor(s) has/have been associated.Address correspondence and reprint requests to Koichi Masuda, MD,1735 W. Harrison St., Cohn 720, Chicago, IL 60612. E-mail:[email protected]
959

as the insertion of functional cells or tissue-engineeredIVD constructs cultured with growth factor supplemen-tation, may prove helpful in treating lumbar disc diseasebiologically rather than by conventional surgery.
Platelet-rich plasma (PRP) is a fraction of plasma thatcan be produced by centrifugal separation of wholeblood in the operating room. Because PRP contains mul-tiple growth factors concentrated at high levels,14–17
PRP from patients is used as an autologous source ofgrowth factors for soft tissue and bone repair in severalclinical settings.18–29
A platelet contains the vast majority of biologicallyactive molecules required for blood coagulation, such asadhesive proteins, coagulation factors and protease in-hibitors, within cytoplasmic �-granules.30 In addition tothe factors that coagulate blood, growth factors such asTGF-�, PDGF, EGF, vascular endothelial growth factor,and IGF-1 are released from �-granules30,31 when plate-lets are activated. These growth factors are known toincrease collagen content, accelerate epithelial regenera-tion, promote angiogenesis, and ultimately improvewound healing in skin and periodontal tissue,21,32–34 aswell as to stimulate IVD metabolism as described above.
Because growth factors found in PRP have been dif-ferently demonstrated to enhance the cell viability or thematrix metabolism of IVD cells,9–12 the authors hypoth-esized that PRP, as a highly concentrated natural cocktailof growth factors, is able to stimulate proliferation andextracellular matrix metabolism of IVD cells. The pur-pose of this study was to examine the effect of PRP on cellproliferation and matrix metabolism of porcine IVD cellscultured in vitro using the alginate bead culture system.
Materials and Methods
Animals: Blood and IVD Collection. Eight mini-pigs (Sin-clair Research Center, Inc., Columbia, MO) were used to iso-late PRP, platelet-poor plasma (PPP) and IVD cells with theapproval of the authors’ institutional Animal Care Committee.Animals were first anesthetized by subcutaneous injections ofTelazol (4.4 mg/kg, Fort Dodge Animal Health, Fort Dodge,IA) and xylazine (2.2 mg/kg, Xyla-Ject, Phoenix Pharmaceuti-cals, Inc., St. Joseph, MO). Blood (55 mL) was drawn from thepopliteal vein with a 14G needle into a 60-mL syringe treatedwith an anticoagulant, citrate dextrose solution (BoehringerLaboratories Inc., Norristown, PA). After collecting peripheralblood, the animals were killed by intravenously injecting super-saturated pentobarbital (Euthanasia B solution, Henry ScheinInc., Washington Port, NY) and the lumbar spinal columns(L1–L6) were dissected en bloc.
Preparation of PRP and PPP. PRP and PPP were isolatedfrom fresh porcine blood (about 55 mL) using the SYMPHONY2 Platelet Concentration System (DePuy Spine, Raynham, MA)according to the manufacturer’s protocol. To facilitate clot for-mation and also eliminate the influence of the biologic effects ofthrombin, both PRP and PPP were clotted by adding a 10%thrombin solution (vol/vol, 1,000 U/mL in 100 mmol/L CaCl2)to yield a final thrombin concentration of 100 U/mL. SolublePRP and PPP releasates from the clotted preparations were
isolated by centrifugation (1,500g for 5 minutes) and clearedby ultra-filtration (0.22 �m). The final soluble releasate prep-arations (equivalent to serum) of PRP and PPP were frozen at�80 C until used.
Isolation of IVD Cells. Porcine IVDs cells were separatelyisolated from both NP and AF tissues by sequential enzymaticdigestion, essentially as previously described.3 Because of thelimitation of cell numbers from a single animal, tissues fromtwo pigs were pooled. The outermost layer of the AF (about 0.5mm) was sharply dissected and discarded to prevent the con-tamination of cells from ligaments surrounding the IVD. TheNP and AF were bluntly separated, and cells were isolated fromNP and AF tissues by sequential enzyme digestion with 0.4%pronase (EMD Bioscience, La Jolla, CA) for 1 hour, and0.025% collagenase P (Roche Applied Science, Indianapolis,IN) and 0.001% deoxy-ribonuclease 2 (DNase 2, Sigma Chem-ical, St. Louis, MO) for 16 hours in a 5% CO2:95% air incu-bator at 37 °C, as previously described.35
Cell Culture in Alginate Beads. After several washes inDMEM/F12 media, the cells isolated from each IVD tissuewere resuspended in 1.2% low-viscosity sterile pharmaceuticalgrade alginate (Keltone LV, a gift from ISP Alginate Inc., SanDiego, CA) solution at 4 million cells/mL.35 The cultures, con-taining 9 beads per well in a 24-well plate, were maintained in400 �L of complete medium [Dulbecco’s modified Eagle me-dium and Ham’s F-12 medium (DMEM/F12: Mediatech,Herndon, VA) containing 10% fetal bovine serum (FBS: Hy-clone, Logan, UT), 25 �g/mL ascorbic acid (Sigma Chemical),360 �g/mL L-glutamine (Mediatech), and 50 �g/mL gentami-cin (Invitrogen, Carlsbad, CA)] in a 5% CO2:95% air incuba-tor. The media were changed daily.
Effects of PRP Releasate, PPP Releasate, and Fetal Bo-vine Serum (FBS). After 7 days of culture in complete me-dium, the cells were precultured in serum-free medium (SFM),which consisted of DMEM/F12 containing 25 �g/mL ascorbicacid, 360 �g/mL L-glutamine and 50 �g/mL gentamicin, for 24hours. The cells were then cultured for another 72 hours underfour different conditions: SFM, FBS (10% FBS in SFM), 10%PPP (10% PPP releasate in SFM), or PRP (10% PRP releasate inthe SFM). The cultures, containing 9 beads per well in a 24-wellplate, were incubated in 800 �L of each medium for 72 hourswithout changing the medium during the treatment period. Apreliminary experiment demonstrated that the rate of incorpo-ration of 35S-sulfate into PG by cells cultured under these con-ditions was constant throughout the 72-hour treatment period(data not shown).
Measurement of DNA Content. To evaluate cellular prolif-eration after the 72-hour treatment period, the DNA content inalginate beads was determined using the bisbenzimidazole flu-orescent dye (Hoechst 33258: Polysciences, Warrington, PA)method.36 Standard curves were generated at the time of eachmeasurement using known concentration of calf thymus DNA.These DNA values were used to normalize the value obtainedfrom the following analyses.
Measurement of Proteoglycan Synthesis. The incorpora-tion of radiolabeled 35S into sulfated proteoglycans was mea-sured as an indicator of proteoglycan synthesis. During the last
960 Spine • Volume 31 • Number 9 • 2006
four hours of the 72-hour treatment period, the cells were cul-tured in the presence of 35S-sulfate (final concentration: 20�Ci/mL; PerkinElmer Life and Analytical Sciences, Boston,MA). After the 72-hour treatments, the beads and culture me-dia were collected and the cell-associated and the further re-moved matrix compartments (CM and FRM, respectively)were separated by dissolving the beads followed by mild cen-trifugation as previously described.37 The CM fractions wereextracted for 48 hours at 4 °C with 4 mol/L guanidine-HCl inthe presence of proteinase inhibitors.38 The amount of radio-labeled 35S-PGs in the CM and FRM fractions and in the me-dium was quantified by a rapid filtration assay following pre-cipitation of the glycosaminoglycans with Alcian blue.39
Characterization of 35S-Labeled Proteoglycan. The size ofnewly synthesized PGs was assessed using CL-2B sieve chro-matography under dissociative conditions, as previously de-scribed.37 The partition coefficient of PGs in each column frac-tion was calculated as follows; (Ve � Vo/Vt � Vo) where Ve
represents the elution volume for each column fraction. Pro-portions of large (Kd � 0.35–0.45) and small (Kd � 0.7–0.8)PGs were calculated by using a Peak Fit v4.12 software (Sea-Solve Software. Inc., Framingham, MA) that performs nonlin-ear least squares curve-fitting using the Marquardt-Levenbergalgorithm to find the true absolute minimum value of the sumof the squared deviations.35
Measurement of Total PG Content. After dissolving thealginate beads, the matrix compartments (CM and FRM) wereseparated and further digested with papain (Sigma Chemical)at 60 C for 16 hours.35 The total sulfated PG content wasmeasured in each compartment using a modified dimethylm-ethylene blue dye-binding method40 in the presence of 0.03%alginate.41 The sum of the total sulfated PG contents of the CMand FRM was used to represent the total content of PG.
Measurement of Collagen Synthesis. The incorporation ofradiolabeled proline into pepsin-resistant protein was mea-sured as an indicator of collagen synthesis. During the last 16hours of the 72-hour treatment period, the cultures were radio-labeled with L-[2,3,4,5-3H]-proline (final concentration: 50�Ci/mL; Amersham Biosciences Corp., Piscataway, NJ). Afterisolation, the CM, FRM, and medium fractions were digestedwith 0.5 mol/L acetic acid including pepsin (Sigma Chemical;100 �g/mL) at 4 °C for 16 hours. An aliquot (25 �L) of eachsample and 125 �L of 30% trichloroacetic acid (final concen-tration 25%; Sigma) were pipetted into a well of a 96-wellMultiScreen filtration plate assembly with gentle agitation forone hour at 4 C.42 The formed precipitate, which containedpepsin-resistant 3H-labeled protein, was collected on the filtermembrane and unincorporated 3H-proline was removed bywashing each well three times with 200 �L of 10% trichloro-acetic acid followed by vacuum filtration through the mem-brane. The membrane in each well was punched out into ascintillation vial with 500 �L of dissolving buffer (2% SDS in0.1 N NaOH solution) followed by gentle shaking overnight at37 °C and the radioactivity of samples counted after adding 2.5mL of Hydrofluor scintillation fluid (National Diagnostics, At-lanta, GA). The types of newly synthesized collagens present inthe CM, FRM, and medium fractions were assessed by SDS-PAGE fluorography, as previously reported.3
Statistical Analyses. All values are reported as the mean �standard deviation of the results of analyses of three separatecultures of 9 beads. Experiments were repeated at least threetimes and representative data are presented. Analysis of vari-ance was used to assess the effects of culture conditions on theresults. When significant differences were detected, hypothesistesting (unpaired T test) was performed to compare values forthe two culture conditions.
Results
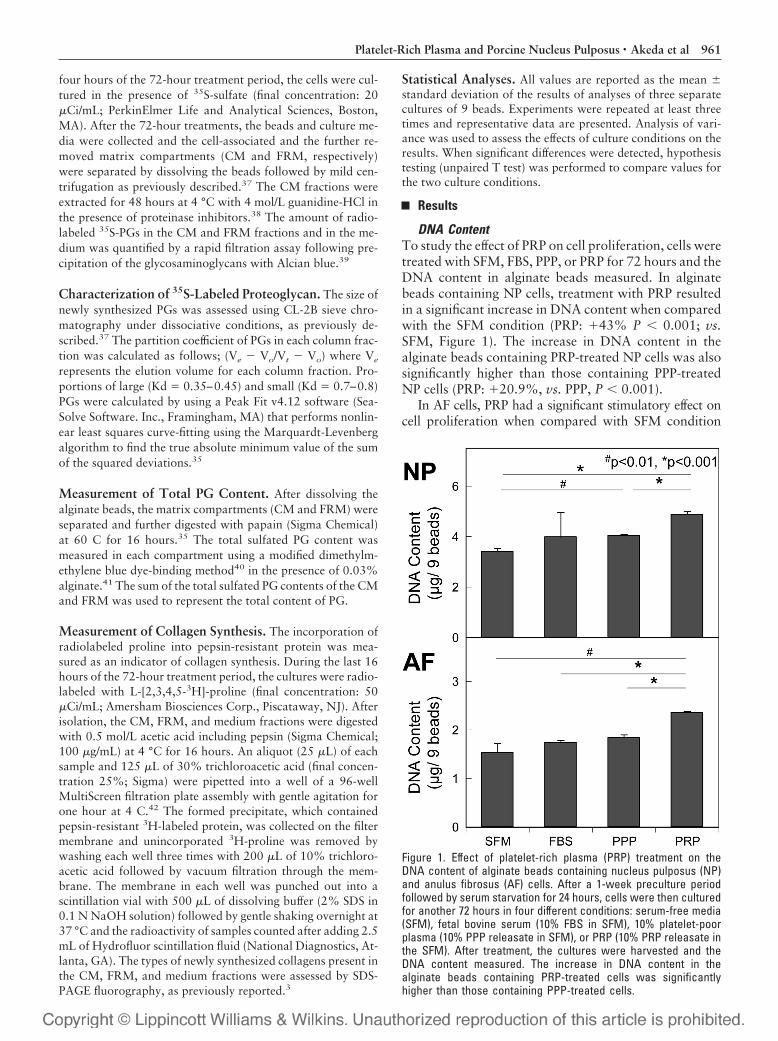
DNA ContentTo study the effect of PRP on cell proliferation, cells weretreated with SFM, FBS, PPP, or PRP for 72 hours and theDNA content in alginate beads measured. In alginatebeads containing NP cells, treatment with PRP resultedin a significant increase in DNA content when comparedwith the SFM condition (PRP: �43% P � 0.001; vs.SFM, Figure 1). The increase in DNA content in thealginate beads containing PRP-treated NP cells was alsosignificantly higher than those containing PPP-treatedNP cells (PRP: �20.9%, vs. PPP, P � 0.001).
In AF cells, PRP had a significant stimulatory effect oncell proliferation when compared with SFM condition
Figure 1. Effect of platelet-rich plasma (PRP) treatment on theDNA content of alginate beads containing nucleus pulposus (NP)and anulus fibrosus (AF) cells. After a 1-week preculture periodfollowed by serum starvation for 24 hours, cells were then culturedfor another 72 hours in four different conditions: serum-free media(SFM), fetal bovine serum (10% FBS in SFM), 10% platelet-poorplasma (10% PPP releasate in SFM), or PRP (10% PRP releasate inthe SFM). After treatment, the cultures were harvested and theDNA content measured. The increase in DNA content in thealginate beads containing PRP-treated cells was significantlyhigher than those containing PPP-treated cells.
961Platelet-Rich Plasma and Porcine Nucleus Pulposus • Akeda et al
(PRP: �53.7%, P � 0.01; vs. SFM). The DNA content ofalginate beads containing PRP-treated AF cells was alsosignificantly higher than those of the FBS- and PPP-treatedAF cells (PRP: �35.7% vs. FBS; �28.5%, vs. PPP, bothP � 0.001).
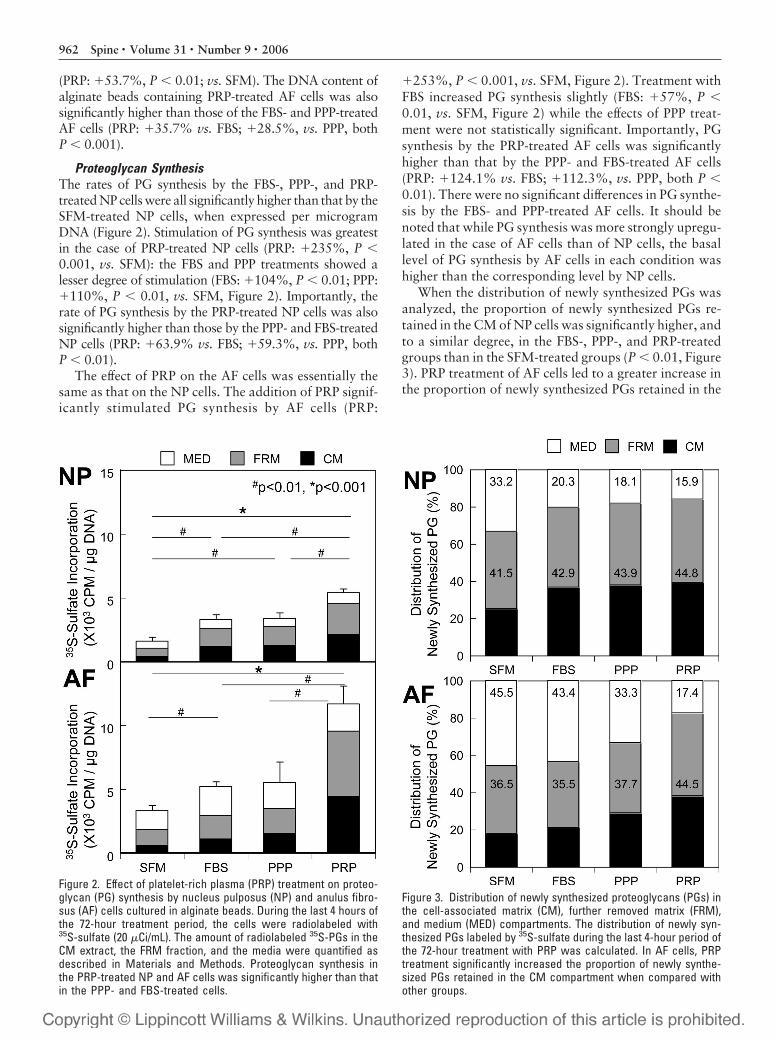
Proteoglycan SynthesisThe rates of PG synthesis by the FBS-, PPP-, and PRP-treated NP cells were all significantly higher than that by theSFM-treated NP cells, when expressed per microgramDNA (Figure 2). Stimulation of PG synthesis was greatestin the case of PRP-treated NP cells (PRP: �235%, P �0.001, vs. SFM): the FBS and PPP treatments showed alesser degree of stimulation (FBS: �104%, P � 0.01; PPP:�110%, P � 0.01, vs. SFM, Figure 2). Importantly, therate of PG synthesis by the PRP-treated NP cells was alsosignificantly higher than those by the PPP- and FBS-treatedNP cells (PRP: �63.9% vs. FBS; �59.3%, vs. PPP, bothP � 0.01).
The effect of PRP on the AF cells was essentially thesame as that on the NP cells. The addition of PRP signif-icantly stimulated PG synthesis by AF cells (PRP:
�253%, P � 0.001, vs. SFM, Figure 2). Treatment withFBS increased PG synthesis slightly (FBS: �57%, P �0.01, vs. SFM, Figure 2) while the effects of PPP treat-ment were not statistically significant. Importantly, PGsynthesis by the PRP-treated AF cells was significantlyhigher than that by the PPP- and FBS-treated AF cells(PRP: �124.1% vs. FBS; �112.3%, vs. PPP, both P �0.01). There were no significant differences in PG synthe-sis by the FBS- and PPP-treated AF cells. It should benoted that while PG synthesis was more strongly upregu-lated in the case of AF cells than of NP cells, the basallevel of PG synthesis by AF cells in each condition washigher than the corresponding level by NP cells.
When the distribution of newly synthesized PGs wasanalyzed, the proportion of newly synthesized PGs re-tained in the CM of NP cells was significantly higher, andto a similar degree, in the FBS-, PPP-, and PRP-treatedgroups than in the SFM-treated groups (P � 0.01, Figure3). PRP treatment of AF cells led to a greater increase inthe proportion of newly synthesized PGs retained in the
Figure 2. Effect of platelet-rich plasma (PRP) treatment on proteo-glycan (PG) synthesis by nucleus pulposus (NP) and anulus fibro-sus (AF) cells cultured in alginate beads. During the last 4 hours ofthe 72-hour treatment period, the cells were radiolabeled with35S-sulfate (20 �Ci/mL). The amount of radiolabeled 35S-PGs in theCM extract, the FRM fraction, and the media were quantified asdescribed in Materials and Methods. Proteoglycan synthesis inthe PRP-treated NP and AF cells was significantly higher than thatin the PPP- and FBS-treated cells.
Figure 3. Distribution of newly synthesized proteoglycans (PGs) inthe cell-associated matrix (CM), further removed matrix (FRM),and medium (MED) compartments. The distribution of newly syn-thesized PGs labeled by 35S-sulfate during the last 4-hour period ofthe 72-hour treatment with PRP was calculated. In AF cells, PRPtreatment significantly increased the proportion of newly synthe-sized PGs retained in the CM compartment when compared withother groups.
962 Spine • Volume 31 • Number 9 • 2006
CM than other forms of treatment (PRP: vs. SFM, P �0.001; vs. FBS, P � 0.01, vs. PPP, P � 0.05, Figure 3).
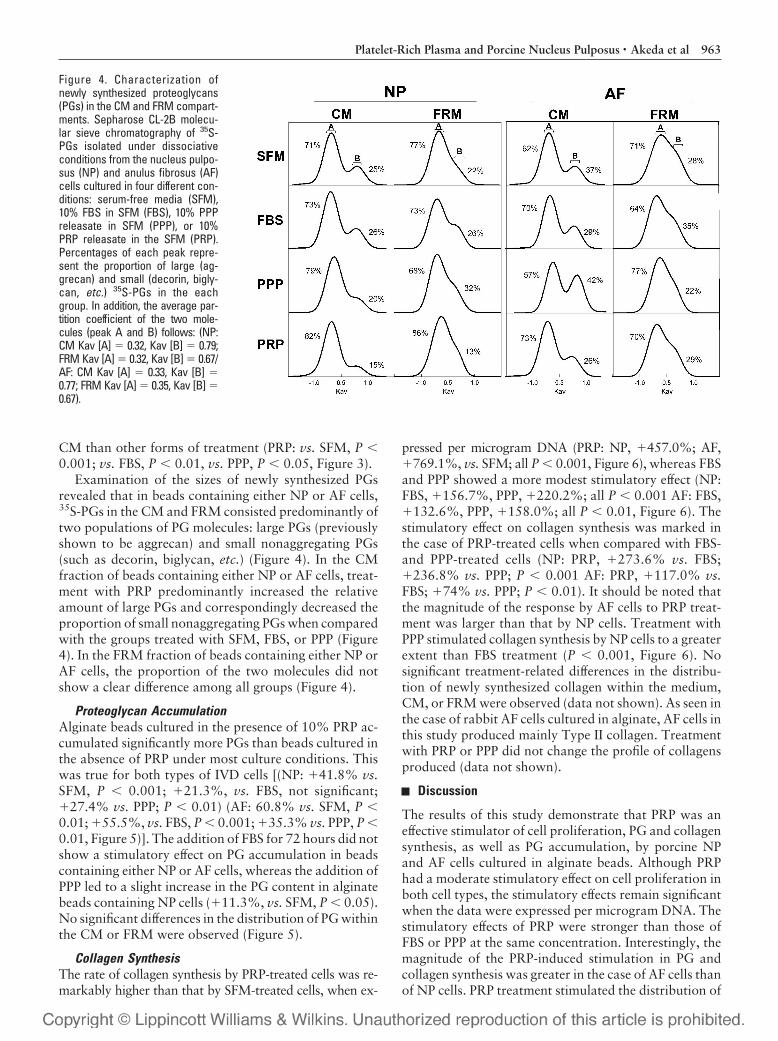
Examination of the sizes of newly synthesized PGsrevealed that in beads containing either NP or AF cells,35S-PGs in the CM and FRM consisted predominantly oftwo populations of PG molecules: large PGs (previouslyshown to be aggrecan) and small nonaggregating PGs(such as decorin, biglycan, etc.) (Figure 4). In the CMfraction of beads containing either NP or AF cells, treat-ment with PRP predominantly increased the relativeamount of large PGs and correspondingly decreased theproportion of small nonaggregating PGs when comparedwith the groups treated with SFM, FBS, or PPP (Figure4). In the FRM fraction of beads containing either NP orAF cells, the proportion of the two molecules did notshow a clear difference among all groups (Figure 4).
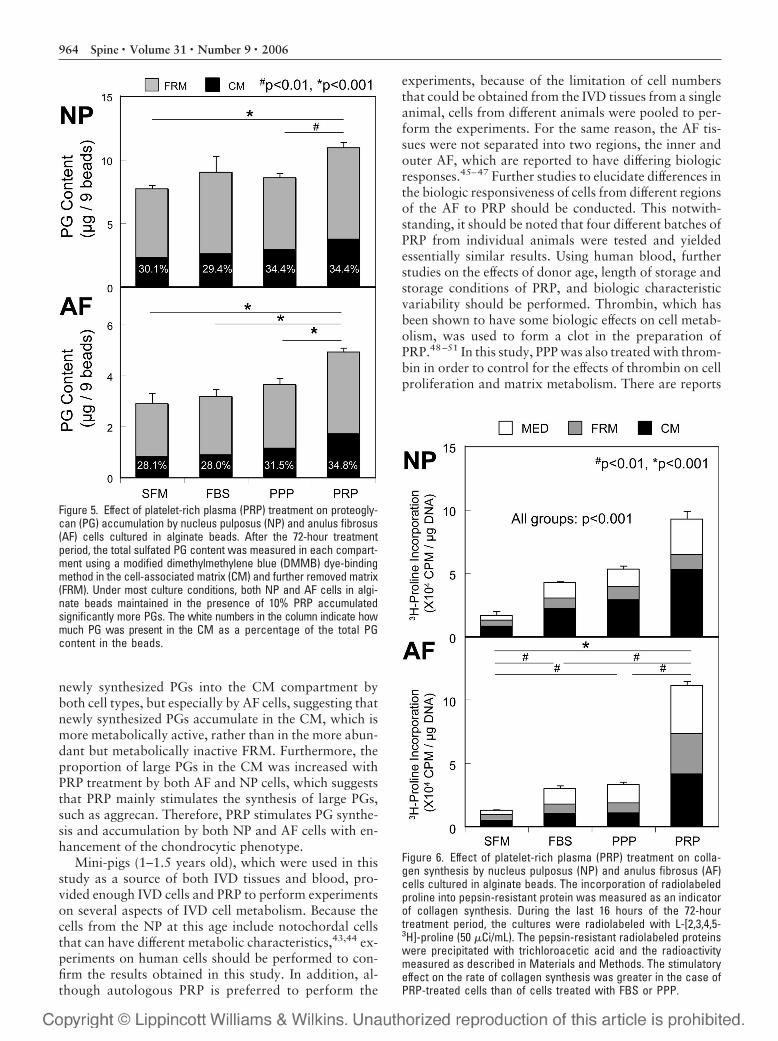
Proteoglycan AccumulationAlginate beads cultured in the presence of 10% PRP ac-cumulated significantly more PGs than beads cultured inthe absence of PRP under most culture conditions. Thiswas true for both types of IVD cells [(NP: �41.8% vs.SFM, P � 0.001; �21.3%, vs. FBS, not significant;�27.4% vs. PPP; P � 0.01) (AF: 60.8% vs. SFM, P �0.01; �55.5%, vs. FBS, P � 0.001; �35.3% vs. PPP, P �0.01, Figure 5)]. The addition of FBS for 72 hours did notshow a stimulatory effect on PG accumulation in beadscontaining either NP or AF cells, whereas the addition ofPPP led to a slight increase in the PG content in alginatebeads containing NP cells (�11.3%, vs. SFM, P � 0.05).No significant differences in the distribution of PG withinthe CM or FRM were observed (Figure 5).
Collagen SynthesisThe rate of collagen synthesis by PRP-treated cells was re-markably higher than that by SFM-treated cells, when ex-
pressed per microgram DNA (PRP: NP, �457.0%; AF,�769.1%, vs. SFM; all P � 0.001, Figure 6), whereas FBSand PPP showed a more modest stimulatory effect (NP:FBS, �156.7%, PPP, �220.2%; all P � 0.001 AF: FBS,�132.6%, PPP, �158.0%; all P � 0.01, Figure 6). Thestimulatory effect on collagen synthesis was marked inthe case of PRP-treated cells when compared with FBS-and PPP-treated cells (NP: PRP, �273.6% vs. FBS;�236.8% vs. PPP; P � 0.001 AF: PRP, �117.0% vs.FBS; �74% vs. PPP; P � 0.01). It should be noted thatthe magnitude of the response by AF cells to PRP treat-ment was larger than that by NP cells. Treatment withPPP stimulated collagen synthesis by NP cells to a greaterextent than FBS treatment (P � 0.001, Figure 6). Nosignificant treatment-related differences in the distribu-tion of newly synthesized collagen within the medium,CM, or FRM were observed (data not shown). As seen inthe case of rabbit AF cells cultured in alginate, AF cells inthis study produced mainly Type II collagen. Treatmentwith PRP or PPP did not change the profile of collagensproduced (data not shown).
Discussion
The results of this study demonstrate that PRP was aneffective stimulator of cell proliferation, PG and collagensynthesis, as well as PG accumulation, by porcine NPand AF cells cultured in alginate beads. Although PRPhad a moderate stimulatory effect on cell proliferation inboth cell types, the stimulatory effects remain significantwhen the data were expressed per microgram DNA. Thestimulatory effects of PRP were stronger than those ofFBS or PPP at the same concentration. Interestingly, themagnitude of the PRP-induced stimulation in PG andcollagen synthesis was greater in the case of AF cells thanof NP cells. PRP treatment stimulated the distribution of
Figure 4. Characterization ofnewly synthesized proteoglycans(PGs) in the CM and FRM compart-ments. Sepharose CL-2B molecu-lar sieve chromatography of 35S-PGs isolated under dissociativeconditions from the nucleus pulpo-sus (NP) and anulus fibrosus (AF)cells cultured in four different con-ditions: serum-free media (SFM),10% FBS in SFM (FBS), 10% PPPreleasate in SFM (PPP), or 10%PRP releasate in the SFM (PRP).Percentages of each peak repre-sent the proportion of large (ag-grecan) and small (decorin, bigly-can, etc.) 35S-PGs in the eachgroup. In addition, the average par-tition coefficient of the two mole-cules (peak A and B) follows: (NP:CM Kav [A] � 0.32, Kav [B] � 0.79;FRM Kav [A] � 0.32, Kav [B] � 0.67/AF: CM Kav [A] � 0.33, Kav [B] �0.77; FRM Kav [A] � 0.35, Kav [B] �0.67).
963Platelet-Rich Plasma and Porcine Nucleus Pulposus • Akeda et al
newly synthesized PGs into the CM compartment byboth cell types, but especially by AF cells, suggesting thatnewly synthesized PGs accumulate in the CM, which ismore metabolically active, rather than in the more abun-dant but metabolically inactive FRM. Furthermore, theproportion of large PGs in the CM was increased withPRP treatment by both AF and NP cells, which suggeststhat PRP mainly stimulates the synthesis of large PGs,such as aggrecan. Therefore, PRP stimulates PG synthe-sis and accumulation by both NP and AF cells with en-hancement of the chondrocytic phenotype.
Mini-pigs (1–1.5 years old), which were used in thisstudy as a source of both IVD tissues and blood, pro-vided enough IVD cells and PRP to perform experimentson several aspects of IVD cell metabolism. Because thecells from the NP at this age include notochordal cellsthat can have different metabolic characteristics,43,44 ex-periments on human cells should be performed to con-firm the results obtained in this study. In addition, al-though autologous PRP is preferred to perform the
experiments, because of the limitation of cell numbersthat could be obtained from the IVD tissues from a singleanimal, cells from different animals were pooled to per-form the experiments. For the same reason, the AF tis-sues were not separated into two regions, the inner andouter AF, which are reported to have differing biologicresponses.45–47 Further studies to elucidate differences inthe biologic responsiveness of cells from different regionsof the AF to PRP should be conducted. This notwith-standing, it should be noted that four different batches ofPRP from individual animals were tested and yieldedessentially similar results. Using human blood, furtherstudies on the effects of donor age, length of storage andstorage conditions of PRP, and biologic characteristicvariability should be performed. Thrombin, which hasbeen shown to have some biologic effects on cell metab-olism, was used to form a clot in the preparation ofPRP.48–51 In this study, PPP was also treated with throm-bin in order to control for the effects of thrombin on cellproliferation and matrix metabolism. There are reports
Figure 5. Effect of platelet-rich plasma (PRP) treatment on proteogly-can (PG) accumulation by nucleus pulposus (NP) and anulus fibrosus(AF) cells cultured in alginate beads. After the 72-hour treatmentperiod, the total sulfated PG content was measured in each compart-ment using a modified dimethylmethylene blue (DMMB) dye-bindingmethod in the cell-associated matrix (CM) and further removed matrix(FRM). Under most culture conditions, both NP and AF cells in algi-nate beads maintained in the presence of 10% PRP accumulatedsignificantly more PGs. The white numbers in the column indicate howmuch PG was present in the CM as a percentage of the total PGcontent in the beads.
Figure 6. Effect of platelet-rich plasma (PRP) treatment on colla-gen synthesis by nucleus pulposus (NP) and anulus fibrosus (AF)cells cultured in alginate beads. The incorporation of radiolabeledproline into pepsin-resistant protein was measured as an indicatorof collagen synthesis. During the last 16 hours of the 72-hourtreatment period, the cultures were radiolabeled with L-[2,3,4,5-3H]-proline (50 �Ci/mL). The pepsin-resistant radiolabeled proteinswere precipitated with trichloroacetic acid and the radioactivitymeasured as described in Materials and Methods. The stimulatoryeffect on the rate of collagen synthesis was greater in the case ofPRP-treated cells than of cells treated with FBS or PPP.
964 Spine • Volume 31 • Number 9 • 2006

that the biologic behavior of IVD cells is affected at alower oxygen tension52,53 and a low pH54 condition thatdiffer from the standard in vitro culture conditions thatwe used in these experiments. The effect of PRP, espe-cially on human IVD cells, at different oxygen tensionsand pH, should be further investigated.
The effect of PRP on PG and collagen synthesis was morepronounced in the case of AF cells than of NP cells. Thisdiffers from results obtained with OP-1, which showed amore pronounced stimulatory effect on NP cells.3 It shouldalso be noted that the effects of PRP were more pronouncedon collagen synthesis, especially in the case of AF cells.Because the AF matrix is rich in collagen, this might suggestthat PRP is a good candidate to induce AF regeneration. Amore detailed analysis of growth factors contained in PRPmight shed light on the mechanisms responsible for thevarying effects of PRP in different tissues and the metabo-lism of different extracellular matrix molecules. In the fu-ture, the therapeutic use of PRP and/or a combination ofgrowth factors that target the repair of a specific tissue ormolecule might be possible.
Because tissue engineering is gaining acceptance as atherapeutic approach for the biologic repair of the IVD,another usage of PRP might be for in vitro tissue engi-neering purposes.1,55 Currently, most tissue engineeringapproaches involve the use of animal serum, such as FBS,during culture. From the regulatory standpoint, the useof autologous serum and/or PRP may gain favor as asource of growth factors and serum supplements.
The use of PRP as a local injection might be one prac-tical approach to stimulate cells for tissue regeneration,as was shown to be effective in the case of a recombinantgrowth factor protein.7
One limitation of the use of PRP as a local injection is theconcentration of growth factors in the preparation. Theconcentration of TGF-�, IGF-1, and PDGF in PRP is lim-ited to approximately a range of 80 to 500 ng/mL.14–17
The amounts of growth factors in the injectable volume(1–2 mL) might be too low to induce the types of biologiceffects that were observed when OP-1 was used at 2 �g/IVDin the rabbit.6 Although multiple growth factors might ex-hibit synergistic effects, the in vivo effects of injectableamounts of PRP should be tested in an experimental animalmodel.
The use of PRP has several advantages over the use ofrecombinant growth factors or products of animal ori-gin. As a point of care approach, the autologous prepa-ration of PRP avoids the complex regulatory pathway. Inaddition, safety issues, such as disease transmission andimmunologic reaction, are of much less concern. As aconsequence, the cost of using PRP will be considerablycheaper than that associated with the use of recombinantproteins.
Conclusion
PRP may prove useful as a culture supplement for tissueengineering or for the biologic repair of the interverte-bral disc. The results of this study provide preliminary
evidence for the possible clinical use of PRP to facilitatebiologic disc repair. Further studies to confirm the effi-cacy and reproducibility of PRP in stimulating humanIVD cell metabolism and its applicability to in vivo ani-mal experiments should be performed to advance thissimple and safe new approach to repair IVD tissue bio-logically.
Key Points
● Platelet-rich plasma is effective in stimulating cellproliferation and extracellular matrix metabolismof porcine intervertebral disc cells cultured in algi-nate beads.● The magnitude of the stimulation by platelet-richplasma was significantly greater than that by eitherfetal bovine serum or platelet-poor plasma.● The response to platelet-rich plasma was greaterin the case of anulus fibrosus cells than of nucleuspulposus cells.● The results raise the possibility of using platelet-rich plasma as a culture supplement for tissue en-gineering or as a local injection to stimulate inter-vertebral disc repair.
References
1. An HS, Thonar EJ, Masuda K. Biological repair of intervertebral disc. Spine2003;28(suppl):86–92.
2. Masuda K, Oegema TR Jr, An HS. Growth factors and treatment of inter-vertebral disc degeneration. Spine 2004;29:2757–69.
3. Masuda K, Takegami K, An H, et al. Recombinant osteogenic protein-1upregulates extracellular matrix metabolism by rabbit annulus fibrosus andnucleus pulposus cells cultured in alginate beads. J Orthop Res 2003;21:922–30.
4. Yoon TS, Su Kim K, Li J, et al. The effect of bone morphogenetic protein-2 onrat intervertebral disc cells in vitro. Spine 2003;28:1773–80.
5. Takegami K, Thonar EJ, An HS, et al. Osteogenic protein-1 enhances matrixreplenishment by intervertebral disc cells previously exposed to interleu-kin-1. Spine 2002;27:1318–25.
6. An HS, Takegami K, Kamada H, et al. Intradiscal administration of osteo-genic protein-1 increases intervertebral disc height and proteoglycan contentin the nucleus pulposus in normal adolescent rabbits. Spine 2005;30:25–31.
7. Masuda K, Imai Y, Okuma M, et al. Osteogenic protein-1 (OP-1) injectioninto a degenerated disc induces the restoration of disc height and structuralchanges in the rabbit annular puncture model. Spine In press.
8. Gruber HE, Hanley EN Jr. Analysis of aging and degeneration of the humanintervertebral disc: comparison of surgical specimens with normal controls.Spine 1998;23:751–7.
9. Thompson JP, Oegema TR Jr, Bradford DS. Stimulation of mature canineintervertebral disc by growth factors. Spine 1991;16:253–60.
10. Gruber HE, Fisher EC Jr, Desai B, et al. Human intervertebral disc cells fromthe annulus: three-dimensional culture in agarose or alginate and responsive-ness to TGF-beta1. Exp Cell Res 1997;235:13–21.
11. Gruber HE, Norton HJ, Hanley EN Jr. Anti-apoptotic effects of IGF-1 andPDGF on human intervertebral disc cells in vitro. Spine 2000;25:2153–7.
12. Osada R, Ohshima H, Ishihara H, et al. Autocrine/paracrine mechanism ofinsulin-like growth factor-1 secretion, and the effect of insulin-like growthfactor-1 on proteoglycan synthesis in bovine intervertebral discs. J OrthopRes 1996;14:690–9.
13. Deleted in proof.14. Weibrich G, Kleis WK, Hafner G, et al. Growth factor levels in platelet-rich
plasma and correlations with donor age, sex, and platelet count. J Crani-omaxillofac Surg 2002;30:97–102.
15. Okuda K, Kawase T, Momose M, et al. Platelet-rich plasma contains highlevels of platelet-derived growth factor and transforming growth factor-betaand modulates the proliferation of periodontally related cells in vitro.J Periodontol 2003;74:849–57.
965Platelet-Rich Plasma and Porcine Nucleus Pulposus • Akeda et al

16. Dugrillon A, Eichler H, Kern S, et al. Autologous concentrated platelet-richplasma (cPRP) for local application in bone regeneration. Int J Oral Maxil-lofac Surg 2002;31:615–9.
17. Landesberg R, Roy M, Glickman RS. Quantification of growth factor levelsusing a simplified method of platelet-rich plasma gel preparation. J OralMaxillofac Surg 2000;58:297–300; discussion 301.
18. Anitua E. Plasma rich in growth factors: preliminary results of use in thepreparation of future sites for implants. Int J Oral Maxillofac Implants1999;14:529–35.
19. Anitua E. The use of plasma-rich growth factors (PRGF) in oral surgery.Pract Proc Aesthet Dent 2001;13:487–93; quiz 493.
20. Marx RE, Carlson ER, Eichstaedt RM, et al. Platelet-rich plasma: growthfactor enhancement for bone grafts. Oral Surg Oral Med Oral Pathol OralRadiol Endod 1998;85:638–46.
21. Carter CA, Jolly DG, Worden CE Sr, et al. Platelet-rich plasma gel promotesdifferentiation and regeneration during equine wound healing. Exp MolPathol 2003;74:244–55.
22. Crovetti G, Martinelli G, Issi M, et al. Platelet gel for healing cutaneouschronic wounds. Transfus Apheresis Sci 2004;30:145–51.
23. Choi BH, Im CJ, Huh JY, et al. Effect of platelet-rich plasma on bone regen-eration in autogenous bone graft. Int J Oral Maxillofac Surg 2004;33:56–9.
24. Fennis JP, Stoelinga PJ, Jansen JA. Mandibular reconstruction: a histologicaland histomorphometric study on the use of autogenous scaffolds, particulatecortico-cancellous bone grafts and platelet rich plasma in goats. Int J OralMaxillofac Surg 2004;33:48–55.
25. Carlson NE, Roach RB Jr. Platelet-rich plasma: clinical applications in den-tistry. J Am Dent Assoc 2002;133:1383–6.
26. Li H, Zou X, Xue Q, et al. Anterior lumbar interbody fusion with carbonfiber cage loaded with bioceramics and platelet-rich plasma: an experimentalstudy on pigs. Eur Spine J 2004;13:354–8.
27. Lozada JL, Caplanis N, Proussaefs P, et al. Platelet-rich plasma applicationin sinus graft surgery: I. Background and processing techniques. J Oral Im-plantol 2001;27:38–42.
28. Anitua E, Andia I, Ardanza B, et al. Autologous platelets as a source ofproteins for healing and tissue regeneration. Thromb Haemost 2004;91:4–15.
29. Hiramatsu T, Okamura T, Imai Y, et al. Effects of autologous platelet con-centrate reinfusion after open heart surgery in patients with congenital heartdisease. Ann Thorac Surg 2002;73:1282–5.
30. Harrison P, Cramer EM. Platelet alpha-granules. Blood Rev 1993;7:52–62.31. Assoian RK, Komoriya A, Meyers CA, et al. Transforming growth factor-
beta in human platelets: identification of a major storage site, purification,and characterization. J Biol Chem 1983;258:7155–60.
32. Declair V. The importance of growth factors in wound healing. OstomyWound Manage 1999;45:64–8.
33. Ross R. Platelet-derived growth factor. Annu Rev Med 1987;38:71–9.34. Kawase T, Okuda K, Wolff LF, et al. Platelet-rich plasma-derived fibrin clot
formation stimulates collagen synthesis in periodontal ligament and osteo-blastic cells in vitro. J Periodontol 2003;74:858–64.
35. Chiba K, Andersson GB, Masuda K, et al. Metabolism of the extracellularmatrix formed by intervertebral disc cells cultured in alginate. Spine 1997;22:2885–93.
36. Kim YJ, Sah RL, Doong JY, et al. Fluorometric assay of DNA in cartilageexplants using Hoechst 33258. Anal Biochem 1988;174:168–76.
37. Mok SS, Masuda K, Hauselmann HJ, et al. Aggrecan synthesized by mature
bovine chondrocytes suspended in alginate: identification of two distinctmetabolic matrix pools. J Biol Chem 1994;269:33021–7.
38. Hauselmann HJ, Fernandes RJ, Mok SS, et al. Phenotypic stability of bovinearticular chondrocytes after long-term culture in alginate beads. J Cell Sci1994;107(Pt 1):17–27.
39. Masuda K, Shirota H, Thonar EJ. Quantification of 35S-labeled proteogly-cans complexed to alcian blue by rapid filtration in multiwell plates. AnalBiochem 1994;217:167–75.
40. Chandrasekhar S, Esterman MA, Hoffman HA. Microdetermination of pro-teoglycans and glycosaminoglycans in the presence of guanidine hydrochlo-ride. Anal Biochem 1987;161:103–8.
41. D’Souza AL, Masuda K, Otten LM, et al. Differential effects of interleukin-1on hyaluronan and proteoglycan metabolism in two compartments of thematrix formed by articular chondrocytes maintained in alginate. Arch Bio-chem Biophys 2000;374:59–65.
42. Koyano Y, Hammerle H, Mollenhauer J. Analysis of 3H-proline-labeledprotein by rapid filtration in multiwell plates for the study of collagen me-tabolism. Biotechniques 1997;22:706–14.
43. Oegema TR Jr. The role of disc cell heterogeneity in determining disc bio-chemistry: a speculation. Biochem Soc Trans 2002;30:839–44.
44. Aguiar DJ, Johnson SL, Oegema TR. Notochordal cells interact with nucleuspulposus cells: regulation of proteoglycan synthesis. Exp Cell Res 1999;246:129–37.
45. Horner HA, Roberts S, Bielby RC, et al. Cells from different regions of theintervertebral disc: effect of culture system on matrix expression and cellphenotype. Spine 2002;27:1018–28.
46. Maeda S, Kokubun S. Changes with age in proteoglycan synthesis in cellscultured in vitro from the inner and outer rabbit annulus fibrosus: responsesto interleukin-1 and interleukin-1 receptor antagonist protein. Spine 2000;25:166–9.
47. Zhang Y, An HS, Song S, et al. Growth factor osteogenic protein-1: differingeffects on cells from three distinct zones in the bovine intervertebral disc.Am J Phys Med Rehabil 2004;83:515–21.
48. Fujii N, Kaji T, Akai T, et al. Thrombin reduces large heparan sulfate pro-teoglycan molecules in cultured vascular endothelial cell layers through in-hibition of core protein synthesis. Thromb Res 1997;88:299–307.
49. Chan B, Merchan JR, Kale S, et al. Antiangiogenic property of human throm-bin. Microvasc Res 2003;66:1–14.
50. Cizmeci-Smith G, Carey DJ. Thrombin stimulates syndecan-1 promotor ac-tivity and expression of a form of syndecan-1 that binds antithrombin III invascular smooth muscle cells. Arterioscler Thromb Vasc Biol 1997;17:2609–16.
51. Haralabopoulos GC, Grant DS, Kleinman HK, et al. Thrombin promotesendothelial cell alignment in Matrigel in vitro and angiogenesis in vivo. Am JPhysiol 1997;273:C239–45.
52. Holm S, Maroudas A, Urban JP, et al. Nutrition of the intervertebral disc:solute transport and metabolism. Connect Tissue Res 1981;8:101–19.
53. Ishihara H, Urban JP. Effects of low oxygen concentrations and metabolicinhibitors on proteoglycan and protein synthesis rates in the intervertebraldisc. J Orthop Res 1999;17:829–35.
54. Bibby SR, Urban JP. Effect of nutrient deprivation on the viability of inter-vertebral disc cells. Eur Spine J 2004.
55. Sun Y, Hurtig M, Pilliar RM, et al. Characterization of nucleus pulposus-liketissue formed in vitro. J Orthop Res 2001;19:1078–84.
966 Spine • Volume 31 • Number 9 • 2006
Copyright © 2022 FDOKUMEN




















