
Aedes larval bionomics and implications ... - Parasites & Vectors

Physical and biological controls on larval dispersal and connectivity in a highly energetic
shelf sea
Peter E. Robins,* Simon P. Neill, Luis Gimenez, Stuart R. Jenkins, and Shelagh K. Malham
Centre for Applied Marine Sciences, School of Ocean Sciences, Bangor University, Menai Bridge, Anglesey, United Kingdom
Abstract
Connectivity within marine species plays a fundamental role in population dynamics, genetic diversity, spreadof disease, and resilience to human exploitation. However, for shellfish species that are sessile as adults, the larval-dispersal stage remains largely unresolved. Appreciation of larval connectivity is therefore crucial to populationgenetics and marine management. We describe a coupled three-dimensional hydrodynamic and Lagrangianparticle tracking model used to simulate larval transport and show how temporal and spatial hydrodynamicchanges, together with larval behavior, are likely to affect dispersal. A case study of Irish Sea (United Kingdom)shellfish populations incorporates a wide range of hydrodynamic environments that are prevalent in other marinesystems around the world. Our simulations tested two main processes that control larval dispersal:hydrodynamics and vertical migration. Simulated larval cohorts were released from estuaries and soft sedimentlocations in regions that were oceanographically distinct. Larvae originating from exposed areas could migrateoffshore (low retention and high connectivity) and disperse farther than larvae that remained in flood-dominantestuaries, which promote retention. Simulated self-recruitment and connectivity with neighboring populations(, 50 km apart) were generally high, although well-developed mesoscale residual currents were important,controlling dispersal pathways offshore. Vertical migration strategies, synchronized either with the tide (tidalstream transport) or with the Earth’s rotation (diel transport), enabled more larvae to remain close to the coast,and simulations indicated higher retention than for passive larvae. However, the probability of connectivity withother populations and potential survivorship was greater for tidal strategies than for passive (although passivetransport populated more distinct areas albeit in smaller proportions, as more larvae remained stranded offshore),or diel, where larvae remained close to their release location.
Many natural metapopulations consist of a set of localpopulations composed of reproductive stages that areconnected by migrants (Hanski 1998; Grimm et al. 2003).The long-term persistence of these metapopulations de-pends on the capacity of organisms to be either locallyretained or migrate and reproduce in a distant population.These two processes define two crucial concepts inpopulation biology: self-recruitment, or the capacity ofjuvenile stages to recruit to the parental population, andconnectivity, defined as the dispersal of individuals amonglocal populations (Cowen et al. 2006; Sheaves 2009). Inmany marine invertebrates, the extent to which localpopulations rely on an input of individuals that are locallysourced (i.e., exhibit self-recruitment) or that are connectedwith other populations (i.e., display high connectivity)depends on the transport of larvae (Roughgarden et al.1985; Caley et al. 1996; Swearer et al. 1999) and specificallyon the direction and magnitude of larval transport. Thepattern of larval transport, whether promoting self-recruitment (retention) or high connectivity among localpopulations, is fundamentally important in species living inpatchy habitats such as reefs, intertidal zones, or estuaries.Variability in oceanographic conditions may result in manylarvae being dispersed away from local populations. Forthat reason, predicting the scale of connectivity and themagnitude of self-recruitment and hence the magnitude andsource of new individuals to a region is pivotal to ourunderstanding of population dynamics, genetic structure
and biogeography, and management of resources (Cowenand Sponaugle 2009).
For species living in inshore habitats and estuaries,recruitment success relies on recolonization of the parentalhabitat (self-recruitment) or colonization of a suitablehabitat in another area (connectivity between populations).Recolonization of these habitats takes place in two steps:arrival in a bay or estuary mouth and transport within thehabitat. Transport within these habitats can depend on thecombination of larval behavior in response to predictablecycles in tidal-driven currents (Forward and Tankersley2001; Queiroga et al. 2006; Tilburg et al. 2010). However,larval retention and transport on the shelf depend also oncirculation patterns (Almeida et al. 2006; Yannicelli et al.2006) that fluctuate with variations in the climate (Siegel etal. 2008; Schultz et al. 2011). It is this shelf-scale larvaltransport that is the focus of our study. Larval exchangeacross a shelf can be restricted in summer if an oceano-graphic front develops between two populations (Gilg andHilbish 2003) since the front acts as a conduit for larvaltransport. Apart from temporal variations, interactionsbetween hydrology and topography modify coastal currents,producing eddies (Mann and Lazier 2006) that result inconcentrations of larvae at particular sites (Mace andMorgan 2006) and spatial patterns of recruitment. Inaddition, the spatial configuration of the habitat matrixdetermines the distance among populations and results inspatially varying patterns of connectivity (Borthagaray et al.2009; Shima 2010) due to limitations in the planktonic larvalperiod. As a consequence, connectivity and self-recruitment* Corresponding author: [email protected]
Limnol. Oceanogr., 58(2), 2013, 505–524
E 2013, by the Association for the Sciences of Limnology and Oceanography, Inc.doi:10.4319/lo.2013.58.2.0505
505

depend on the combination of larval behavior with theheterogeneous nature of the marine habitat (Cowen et al.2006; Fox et al. 2009; Shima et al. 2010). Understanding howthe hydrodynamic structure combines with individualbiological traits of larvae is therefore of considerable interestto marine planners, managers, and policymakers (Holt et al.2010).
This study evaluates how spatial variations in oceano-graphic conditions and the interaction of larval behaviorstrategies contribute toward self-recruitment and connec-tivity of populations of organisms confined to estuaries andsoft sediment habitats. We have devised a model-basedbiophysical study to investigate factors that contribute tothese two processes in marine species abundant in the IrishSea (Fig. 1). Our rationale for this study stems from aninsufficient knowledge of the drivers for populationdynamics in a high-energy and hydrodynamically complex
environment such as the Irish Sea. We develop biophysicalmodels to simulate oceanographic circulation and differentlarval behavior strategies known to influence dispersal: dielmigration (Dos Santos et al. 2008), tidal stream migration(Knights et al. 2006; Criales et al. 2007), and passivebehavior. We simulate larval transport and predict patternsof population connectivity and self-recruitment. Our focushabitat, shallow estuarine and sheltered soft sedimentareas, supports the adult benthic stages of many mollusks(e.g., common cockle Cerastoderma edule and musselMytilus edulis), polychaetes, and echinoderms. Althoughfew empirical larval studies exist (Knights et al. 2006), it isthought that these target species for the Irish Sea exhibitvertical migration that are similar to strategies modeledhere. The habitat is typically patchy, isolated by longextensions of exposed sandy or rocky shores. Thus,successful recruitment relies on local retention of larvae
Fig. 1. The hydrodynamic model domain, showing water depths (m) relative to mean sea level (depths in the North Channel exceed200 m). The Irish Sea is approximately bounded by St. George’s Channel and the North Channel. Open boundaries were located fartheroffshore, improving model accuracy of baroclinic circulations within the Irish Sea. Land is colored green. The horizontal grid resolutionis , 1.85 km, and there are 20 terrain-following vertical layers. Locations of 40 larvae spawning sites are also plotted, representingprincipal estuaries and embayments where shellfish populations are found. The names of each site are printed to the right of the map.Other points of interest are as follows: A 5 Pembrokeshire, B 5 Llyn Peninsula, C 5 Anglesey, D 5 the Isle of Man, and E 5 StranraerPeninsula. Data available from location S1 were used to validate simulated sea surface and bottom temperatures.
506 Robins et al.
or export and transport to suitable coastal habitatelsewhere. We outline the following specific questions thatwill be addressed in this study: (1) What is the effect ofrelease location (i.e., local bathymetry and topography) onlarval retention, connectivity, and dispersal? (2) How doestidal energy influence larval retention, connectivity, anddispersal? (3) How does the seasonal evolution of residualcurrents influence larval retention, connectivity, anddispersal? (4) How does larval behavior strategy influenceretention, connectivity, and dispersal?
Methods
This work is based on simulating larval transport in theIrish Sea and obtaining predictions on the proportion oflarvae contributing to self-recruitment and connectivity aswell as the proportion not reaching any appropriate site forsettlement. These predictions are made for three larvalmigration strategies: no migration (passive), diel migration,and tidal migration.
The model domain is shown in Fig. 1. The regionincludes both Irish and UK exclusive economic zones andextends northward from the Celtic Sea to beyond the NorthChannel. Water depths are typically less than 100 m,although depths are greater in the central channel,exceeding 250 m in the North Channel. The Irish Sea is atypical semienclosed mesoscale (10–1000 km) basin, andhence the findings generally relate to analogous marineecosystems. The model focuses on larval transport in theshelf seas; transport within estuaries or bays may dependon local conditions and must be addressed separately(Tilburg et al. 2010; Robins et al. 2012). While the model iscapable of simulating tidal currents in coastal areas,secondary currents generated by local topography may bepoorly simulated because of model resolution. Therefore,larvae were released toward the outer estuary or embay-ment, where they were exposed to shelf sea currents, andtransported to either their natal habitat within the estuary(self-recruitment) or a similar habitat elsewhere (connec-tivity). This assumption implies that larvae are capable ofleaving their natal habitat. The natal habitats are 40geographically distinct locations on the coasts of Wales,England, Scotland, Northern Ireland, Ireland, and the Isle
of Man (see Fig. 1; Table 1). These locations were chosenfrom a number of sources that monitor shellfish popula-tions (e.g., http://data.nbn.org.uk; habitat maps; Robinsonet al. 2010) based on maximum species density.
Modeling is based on two components: a hydrodynamicmodel and a Lagrangian particle (larvae) tracking model,which are explained in the following two sections. The thirdsection describes how model outputs were used to calculateconnectivity and self-recruitment.
Hydrodynamic model
A parallel version of the three-dimensional, primitiveequation, sigma-coordinate, free-surface Princeton OceanModel (POM) was used to simulate the hydrodynamics.The model (in its nonparallel form) is described in detail byBlumberg and Mellor (1987) and hence only brieflyoutlined here. POM incorporates Mellor and Yamada level2.5 turbulence closure (Blumberg and Mellor 1987). It isassumed that the weight of the fluid identically balances thepressure (hydrostatic assumption: valid here since bathy-metric gradients, and hence vertical accelerations remainsmall; Mellor et al. 1994) and that density differences areneglected unless the differences are multiplied by gravity(Boussinesq approximation). The standard prognosticvariables (e.g., velocity, temperature, salinity, and turbu-lence) are solved using finite-difference discretization on astaggered, orthogonal Arakawa-C grid in the horizontalplane and terrain-following sigma layers in the verticalplane.
Accurate simulation of long-term circulation in thecoastal zone is one of the greatest challenges facing oceanmodelers. Confidence in model predictions is achieved onlythrough extensive validation of a range of variables. A yearin the recent past (1990) was chosen as a typical year bothfor validation and to provide input for the ParticleTracking Model (PTM), based on an analysis of bed shearstress, significant wave heights, and temperature recordsfrom a decadal simulation (1989–1998; Neill et al. 2010).The model horizontal cell size is 1/30u (longitude) by 1/60u(latitude), giving a resolution of approximately 1.85 km. Inthe vertical plane, 20 equally segmented sigma layers giveminimum and average resolutions at mean sea level of 9.6
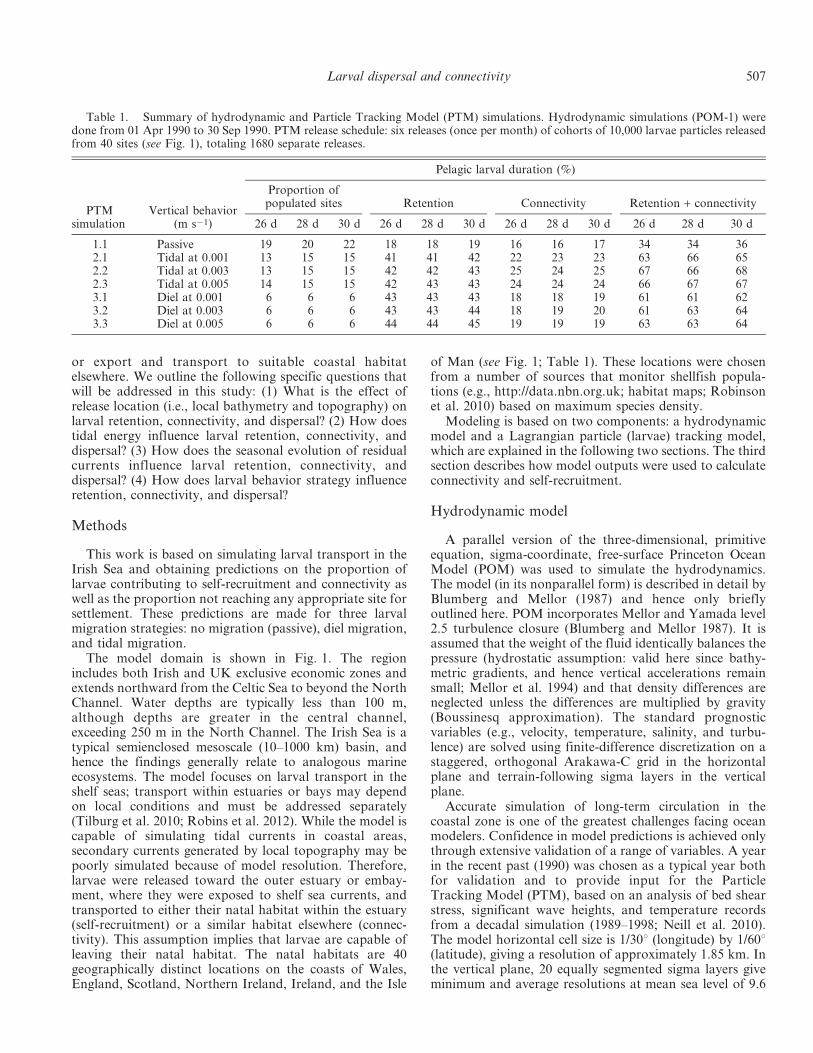
Table 1. Summary of hydrodynamic and Particle Tracking Model (PTM) simulations. Hydrodynamic simulations (POM-1) weredone from 01 Apr 1990 to 30 Sep 1990. PTM release schedule: six releases (once per month) of cohorts of 10,000 larvae particles releasedfrom 40 sites (see Fig. 1), totaling 1680 separate releases.
PTMsimulation
Vertical behavior(m s21)
Pelagic larval duration (%)
Proportion ofpopulated sites Retention Connectivity Retention + connectivity
26 d 28 d 30 d 26 d 28 d 30 d 26 d 28 d 30 d 26 d 28 d 30 d
1.1 Passive 19 20 22 18 18 19 16 16 17 34 34 362.1 Tidal at 0.001 13 15 15 41 41 42 22 23 23 63 66 652.2 Tidal at 0.003 13 15 15 42 42 43 25 24 25 67 66 682.3 Tidal at 0.005 14 15 15 42 43 43 24 24 24 66 67 673.1 Diel at 0.001 6 6 6 43 43 43 18 18 19 61 61 623.2 Diel at 0.003 6 6 6 43 43 44 18 19 20 61 63 643.3 Diel at 0.005 6 6 6 44 44 45 19 19 19 63 63 64
Larval dispersal and connectivity 507
and 4.3 m, respectively. A 6 month, three-dimensional,barotropic + baroclinic simulation was made for the period01 April–30 September 1990 (POM-1; see Table 2). Thesimulation was run for 1 month prior to the 6 monthharmonic analysis so that baroclinic currents were fullydeveloped. The model was forced at the open boundarieswith six tidal constituents including the dominant semidi-urnal M2 (lunar) and S2 (solar) constituents. The source ofsynoptic meteorological fields was the European Centre forMedium-Range Weather Forecasts-Interim reanalysis(Simmons et al. 2006), available every 3 h at a (global)grid resolution of 1.5u. Simulated barotropic circulationwas validated by comparing amplitudes and phases of themodeled M2 and S2 elevations with records of knownvalues from 18 coastal stations within the domain, givingroot-mean square errors of approximately 14 and 6 cm (M2
and S2, respectively) in elevation and 10u and 12u in phase.These values compare well with other models of this region(Xing and Davies 2001; Horsburgh and Hill 2003; Hartnettet al. 2007).
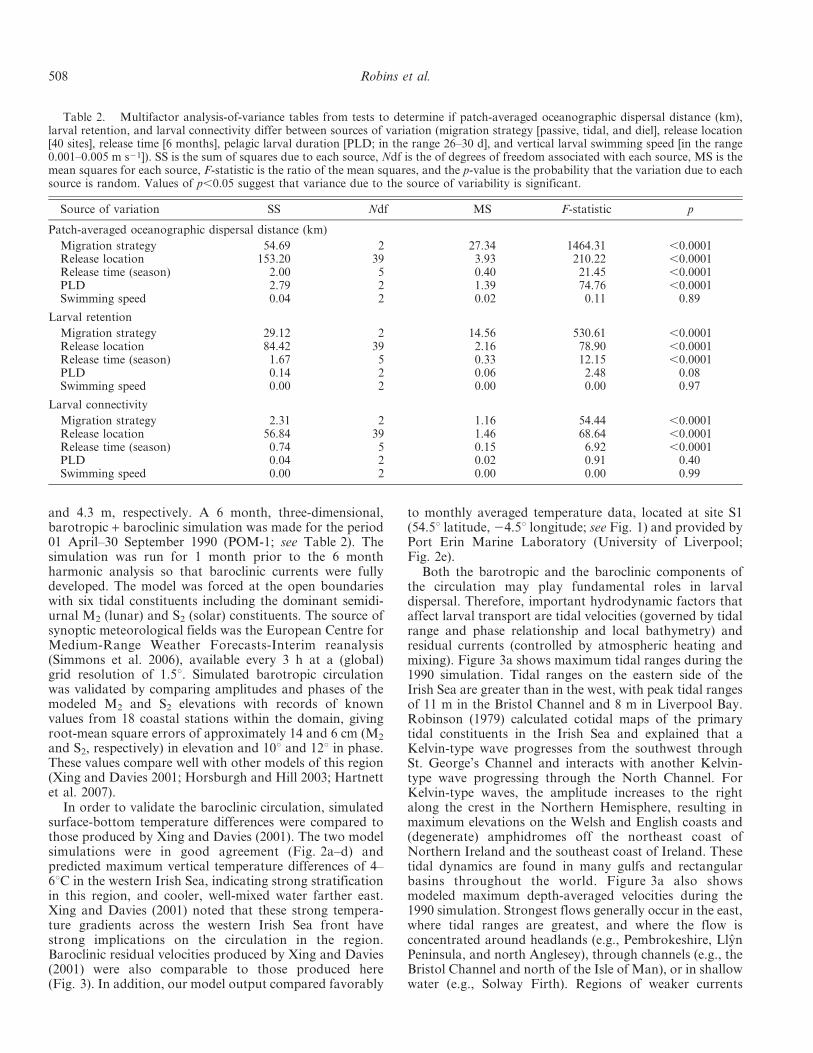
In order to validate the baroclinic circulation, simulatedsurface-bottom temperature differences were compared tothose produced by Xing and Davies (2001). The two modelsimulations were in good agreement (Fig. 2a–d) andpredicted maximum vertical temperature differences of 4–6uC in the western Irish Sea, indicating strong stratificationin this region, and cooler, well-mixed water farther east.Xing and Davies (2001) noted that these strong tempera-ture gradients across the western Irish Sea front havestrong implications on the circulation in the region.Baroclinic residual velocities produced by Xing and Davies(2001) were also comparable to those produced here(Fig. 3). In addition, our model output compared favorably
to monthly averaged temperature data, located at site S1(54.5u latitude, 24.5u longitude; see Fig. 1) and provided byPort Erin Marine Laboratory (University of Liverpool;Fig. 2e).
Both the barotropic and the baroclinic components ofthe circulation may play fundamental roles in larvaldispersal. Therefore, important hydrodynamic factors thataffect larval transport are tidal velocities (governed by tidalrange and phase relationship and local bathymetry) andresidual currents (controlled by atmospheric heating andmixing). Figure 3a shows maximum tidal ranges during the1990 simulation. Tidal ranges on the eastern side of theIrish Sea are greater than in the west, with peak tidal rangesof 11 m in the Bristol Channel and 8 m in Liverpool Bay.Robinson (1979) calculated cotidal maps of the primarytidal constituents in the Irish Sea and explained that aKelvin-type wave progresses from the southwest throughSt. George’s Channel and interacts with another Kelvin-type wave progressing through the North Channel. ForKelvin-type waves, the amplitude increases to the rightalong the crest in the Northern Hemisphere, resulting inmaximum elevations on the Welsh and English coasts and(degenerate) amphidromes off the northeast coast ofNorthern Ireland and the southeast coast of Ireland. Thesetidal dynamics are found in many gulfs and rectangularbasins throughout the world. Figure 3a also showsmodeled maximum depth-averaged velocities during the1990 simulation. Strongest flows generally occur in the east,where tidal ranges are greatest, and where the flow isconcentrated around headlands (e.g., Pembrokeshire, LlynPeninsula, and north Anglesey), through channels (e.g., theBristol Channel and north of the Isle of Man), or in shallowwater (e.g., Solway Firth). Regions of weaker currents
Table 2. Multifactor analysis-of-variance tables from tests to determine if patch-averaged oceanographic dispersal distance (km),larval retention, and larval connectivity differ between sources of variation (migration strategy [passive, tidal, and diel], release location[40 sites], release time [6 months], pelagic larval duration [PLD; in the range 26–30 d], and vertical larval swimming speed [in the range0.001–0.005 m s21]). SS is the sum of squares due to each source, Ndf is the of degrees of freedom associated with each source, MS is themean squares for each source, F-statistic is the ratio of the mean squares, and the p-value is the probability that the variation due to eachsource is random. Values of p,0.05 suggest that variance due to the source of variability is significant.
Source of variation SS Ndf MS F-statistic p
Patch-averaged oceanographic dispersal distance (km)
Migration strategy 54.69 2 27.34 1464.31 ,0.0001Release location 153.20 39 3.93 210.22 ,0.0001Release time (season) 2.00 5 0.40 21.45 ,0.0001PLD 2.79 2 1.39 74.76 ,0.0001Swimming speed 0.04 2 0.02 0.11 0.89
Larval retention
Migration strategy 29.12 2 14.56 530.61 ,0.0001Release location 84.42 39 2.16 78.90 ,0.0001Release time (season) 1.67 5 0.33 12.15 ,0.0001PLD 0.14 2 0.06 2.48 0.08Swimming speed 0.00 2 0.00 0.00 0.97
Larval connectivity
Migration strategy 2.31 2 1.16 54.44 ,0.0001Release location 56.84 39 1.46 68.64 ,0.0001Release time (season) 0.74 5 0.15 6.92 ,0.0001PLD 0.04 2 0.02 0.91 0.40Swimming speed 0.00 2 0.00 0.00 0.99
508 Robins et al.
occur in Cardigan Bay, along the North Wales coast, andin the western Irish Sea.
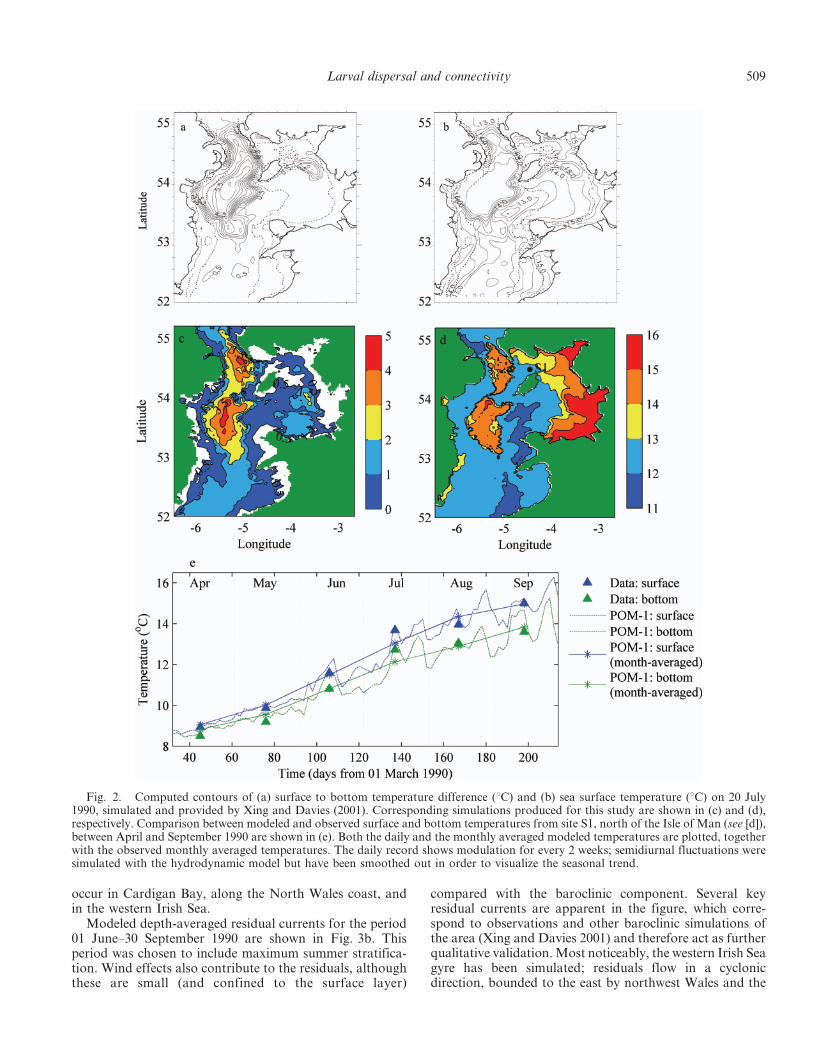
Modeled depth-averaged residual currents for the period01 June–30 September 1990 are shown in Fig. 3b. Thisperiod was chosen to include maximum summer stratifica-tion. Wind effects also contribute to the residuals, althoughthese are small (and confined to the surface layer)
compared with the baroclinic component. Several keyresidual currents are apparent in the figure, which corre-spond to observations and other baroclinic simulations ofthe area (Xing and Davies 2001) and therefore act as furtherqualitative validation. Most noticeably, the western Irish Seagyre has been simulated; residuals flow in a cyclonicdirection, bounded to the east by northwest Wales and the
Fig. 2. Computed contours of (a) surface to bottom temperature difference (uC) and (b) sea surface temperature (uC) on 20 July1990, simulated and provided by Xing and Davies (2001). Corresponding simulations produced for this study are shown in (c) and (d),respectively. Comparison between modeled and observed surface and bottom temperatures from site S1, north of the Isle of Man (see [d]),between April and September 1990 are shown in (e). Both the daily and the monthly averaged modeled temperatures are plotted, togetherwith the observed monthly averaged temperatures. The daily record shows modulation for every 2 weeks; semidiurnal fluctuations weresimulated with the hydrodynamic model but have been smoothed out in order to visualize the seasonal trend.
Larval dispersal and connectivity 509

Isle of Man, and to the west by Ireland. Horsburgh and Hill(2003) explained that the gyre forms during spring andsummer when stratification and weak tidal currents (,0.3 m s21) in the west are bounded by a tidal mixing front(Simpson and Hunter 1974) and strong currents (. 2 m s21)to the east. Residual flow in the western Irish Sea gyre isgenerally between 0.05 and 0.1 m s21, which means thatpassive particles can travel up to 4.5 km because of theresidual alone, over the duration of one tidal cycle. Othersignificant residual currents were simulated flowing south-ward along the eastern coast of Ireland and then westwardinto the Celtic Sea. Although relatively weak (, 0.05 m s21),westward residual currents from South Wales towardIreland, across St. George’s Channel, are likely to beimportant for larval transport. Again, this net flow can beattributed to the observed position of the tidal mixing front(Celtic front) in the southern Irish Sea (Simpson and Hunter1974).
Particle tracking model—Lagrangian PTMs are used tosimulate the individual movement of particles in space andtime, based on advection, subgrid-scale turbulent mixing,and individual particle behavior. In this application, theparticles were used to represent planktonic larvae. Thedispersal probability distributions of the larvae, originatingfrom distinct sites, can be mapped and the connectivitybetween geographically distinct populations estimated.Three-dimensional velocity predictions are output fromthe hydrodynamic model for use, off-line, in PTMs. Thistechnique is sufficiently accurate since the transport oflarvae does not affect the hydrodynamics. The advantageof this method is that both the hydrodynamic and theparticle tracking models are more computationally efficientwhen separated and also that the hydrodynamics from asingle simulation can be used to simulate a large number ofindividual or cluster-release scenarios (as is the case for thepresent study), whereas a particle tracking scheme that isintegrated with the hydrodynamic model would simulate
only one scenario for large computational effort. Thevelocities were trilinearly interpolated from the hydrody-namic model to the position of each larva, and the intervalof hydrodynamic model (15 min) was linearly interpolatedto 5 min for the PTM. Each larva was then iterativelyadvected in space and time.
The larvae are mixed locally through subgrid-scaleturbulence, based on random displacement models (ran-dom walks), where the eastward change in position, Dx (m),over a PTM time step Dt (i.e., 600 s) is given by (Proctor etal. 1994)
Dx~R
rcos (2pR)(2KxDt)1=2 ð1aÞ
where R is a random number in the range [0,1] and r is the
standard deviation of R cos(2pR) with a value of 1=ffiffiffi6p
. Theexpression (R/r)cos(2pR) thus has the necessary propertiesof a mean of zero and a standard deviation of unity (Rossand Sharples 2004). A similar expression for northwarddiffusion is given by
Dy~R
rsin (2pR)(2KyDt)1=2 ð1bÞ
Spatially and temporally varying horizontal diffusivities,Kx and Ky (m2 s21), were output from the hydrodynamicmodel and linearly interpolated in the same way as thevelocities. A random displacement model (Visser 1997;North et al. 2006) was used to simulate subgrid-scalevertical turbulence:
Dz~K0
zDtzR
r(2KzDt)1=2 ð2Þ
where Kz (m2 s21) is the vertical diffusivity, calculated bythe hydrodynamic model and trilinearly interpolated to the
vertical particle position, z, in the PTM algorithm and K0
z 5
hKz/hz is evaluated at depth z.
Fig. 3. (a) Cotidal contours of maximum tidal range (m), superimposed on colored contours of maximum depth-averaged scalarvelocity (m s21). (b) Depth-averaged residual currents over the period 01 June–30 September 1990. Residuals less than 0.02 m s21 havebeen removed for clearer visualization of the stronger currents. The western Irish Sea gyre, where density-driven currents flowcounterclockwise between north Wales, the Isle of Man, and Ireland, can clearly be depicted. Other significant residuals flow west acrossthe Celtic Sea front (St. George’s Channel) and clockwise around southeast Ireland toward the Celtic Sea.
510 Robins et al.

We investigate habitual larval behaviors as controls ondispersal. Three vertical migratory strategies for the larvaehave been implemented into the PTMs: (1) passivetransport, where there is no vertical migration; (2) tidalstream transport, where larvae migrate upward at 3 31023 m s21 during the flood phase of the tide anddownward during the ebb; and (3) diel transport, wherelarvae swim upward, also at 3 3 1023 m s21, during thenight and downward during the day. This generic range ofvertical behaviors has the added advantage that the resultscan be applied to a variety of species. Vertical swimmingspeeds of crustacean larvae reported in the literature varyfrom a few millimeters per second to several centimeters persecond (Michalec et al. 2010; Seuront 2010; Souissi et al.2010). Our chosen larval swimming speed of 3 3 1023 m s21
represents ciliated larvae (Chia et al. 1984) that are found inmollusks (e.g., clams, oysters, scallops, and mussels),polychaetes, and echinoderms. Some crustacean larvae(zoeal stages) swim in the order of several millimeters persecond to centimeters per second, and larger larvae (lobsterand crab megalopae) swim at 5–10 cm per second (Shanks1995). We are not representing these animals. In order toassess the variability of dispersal, our chosen swimmingspeed was modified in the range 0–0.005 m s21. We foundthat, above a threshold speed of 0.001 m s21, dispersalpatterns remained broadly the same (see Discussion).Therefore, our results represent ciliated larvae in the range0.001–0.005 m s21, confirmed by a sensitivity test performedin that range and on all tidal and diel simulations (seeTable 1).
Estimates of early life stage mortality are available for areduced number of species (Peterson and Wrobleski 1984;McGurk 1986; Houde 2002). Mortality rates will affectoverall connectivity but may not affect patterns ofconnectivity unless mortality varies in space (Paris et al.2007). Inclusion of mortality may require species-specificinformation and will prevent an accurate assessment of thephysical and biological controls on larval dispersal in ourgeneralized study. In addition, a mortality function reducesthe number of particles in the analysis. Therefore, we havechosen to assume that larvae remain alive throughout thesimulations. Another important process in the PTM is howlarvae behave when they encounter a coastline. Differentmodeling methods, such as settlement (the effect ifinterpolation between coastal and land cells occurred) orreflection, can affect the overall dispersal. Here, we havechosen to reflect larvae back into the water column, to theirposition at the previous iteration step, if a land boundary(or vertical boundary) is encountered so that maximumdispersal can be investigated (a similar method was used byNorth et al. 2008). Furthermore, keeping more particles incirculation increases the statistical power of the subsequentanalysis.
In order to ensure that sufficient numbers of larvae werereleased in each cohort so that the trajectories are notpolluted by statistical outliers, a simple test was performed.A large number of passive larvae (106) were released from alocation within the domain, and the concentrations oflarvae occurring in selected grid cells at the end of thesimulation were calculated. The PTM run was repeated,
reducing the number of released larvae, until the propor-tion of larvae in each selected cell started to fluctuatesignificantly (i.e., varied by more than 10% of the initial[106] simulation). The experiment was repeated for differentrelease locations, and it was found that a cohort of 10,000larvae is sufficient to satisfactorily simulate larval dispersal.
Cohorts of 10,000 larvae were released instantaneouslyfrom each of the 40 sites and tracked for a pelagic larvalduration (PLD) of 28 d, which is typical for a range ofciliated larvae investigated in the literature (range: 17–49 d;Siegel et al. 2003; Hartnett et al. 2007; Tian et al. 2009). Asensitivity test was also performed on our chosen PLD of28 d by modifying the PLD by 6 10% for all simulations.Larvae were released randomly distributed throughout thewater column. The initial footprint (within which particleswere randomly positioned) of each larvae cluster was chosento coincide with the outer (offshore) region of each estuary,bay, or coast so that the particles have a high chance of beinginfluenced by the offshore currents as well as potentiallybeing advected into the estuary because of onshore coastalcurrents. The above procedure was repeated at six differenttimes throughout spring and summer, and invertebrates arereported to spawn (Hartnett et al. 2007; Ayata et al. 2010)with start dates randomly chosen within the first 2 weeks ofeach month (April–September) so that the variance indispersal caused by tidal phase and season can be captured.The PTM simulations were then repeated for the threevertical migratory behaviors (passive, tidal stream transport,and diel transport), which amounted to a total of 720 PTMreleases, as summarized in Table 1. A selection of thesimulations were repeated at different dates (within eachmonth) in order to assess the variance in dispersal patternsdue to tidal phase and (randomly assigned) initial releasepositions. Although these results are not shown, in each casethe dispersal patterns were similar to the results presentedhereafter, which are therefore representative of all thesimulations. Sensitivity tests on swimming speeds andPLD, described above, were then performed on allsimulations (see Table 1).
Connectivity and self-recruitment—Quantitative parame-terizations of population connectivity have been producedusing source distribution matrices pij. (Ayata et al. 2010),which give the proportion of larvae that successfully settlein population i (i.e., sink population), after the PLD, thatcame from population j (i.e., source population). A valueclose to unity indicates high connectivity, whereas a valueclose to zero indicates low connectivity. Retention at thesource site is determined by pii. The overall success of larvaewithin a population j (i.e., larvae that manage to settle atany site) is given by
Tj{Xi~n
i~1
pij
where n is the number of sink populations. Larval wastageis denoted by 1 2 Tj. On completion of the PTMsimulations, specific criteria were required to determinewhich larvae were successful. We used a similar approachto that taken by Cowen et al. (2006) for their connectivity
Larval dispersal and connectivity 511

criteria of larvae in the Caribbean Sea; any larvae that werewithin 10 km of one of the 40 release sites at the end of thesimulated larval period were deemed to settle at that site.The distance of 10 km was chosen because it is comparableto the tidal excursion of a passive particle (Xing and Davies2001). Larvae that did not meet these criteria (i.e., were notwithin 10 km of one of the 40 sites) were consideredunsuccessful. Evidently, using this approach, measures ofconnectivity are based only on a subset of environmentsthat larvae could possibly live. However, it is thecomparative outputs (e.g., between release locations orlarval behaviors) that are the critical results.
We address the questions raised in the Introductionthrough statistical tests. We use multifactor analysis ofvariance to determine responses of patch-averaged dispers-al distance, retention, and connectivity to the followingvariables: larval behavior strategy, release location (i.e., theeffect of local bathymetry), release time (i.e., seasonaleffects), PLD (in the range 6 10% of 28 d), and larvalswimming speed (in the range 0.001–0.003 m s21). Theinfluence of tidal energy and residual currents on retentionand connectivity were evaluated through quantile regres-sion. This technique estimates trends that are more robustagainst outliers than ordinary least squares regression incases where there is a weak relationship between the meansof such variables. We used oceanographic dispersaldistances (i.e., the total distance traveled by each larvae)and release time as proxies for variations in tidal energyand residual currents, respectively.
Results
Our modeling exercise predicted important effects oflarval behavior, release location, and release time ondispersal distances, connectivity, and self-recruitment. Thiswas confirmed using sensitivity analyses (Table 2). PLDaffected dispersal distance significantly, as one mightexpect, but not connectivity or self-recruitment (Table 2).However, the sensitivity analyses did not show anysignificant effect of swimming speeds in the range 0.001–0.005 m s21 (Table 2). It must be noted that changes inPLD and swimming speed outside our chosen ranges arelikely to affect dispersal, retention, and connectivity.Therefore, the results described below are robust to smallvariations in these PLD and swimming speeds.
General effects of larval behavior—Passive behaviorresulted in lower larval success (connectivity + retention)than active behavior (Fig. 4). In total, 34% of all passivelarvae were either retained or arrived at a suitablesettlement site; this contrasts with 66% and 63% foundfor tidal and diel larvae, respectively (Table 1). Therefore,passive dispersal resulted in high wastage: in 16 out of 40populations, more than 80% of larvae did not reach anysite, and in three populations, wastage was less than 20%(Fig. 5c). By contrast, fewer than seven tidal or dielpopulations predicted high wastage (Fig. 5c). However,the proportion of settlement sites that were populated wasgreater for passive larvae (20%) than for tidal (15%) or diel(6%) strategies (Table 1); that is, passive larvae have the
ability to connect with more settlement sites than either tidalor diel migrators (because the passive cohorts spread outmore).
The reason why less passive larvae reach each site yetmore sites become populated is related to the dispersalpattern. Larvae drifting passively with the currents disperseaway from their natal habitats and occupy extensiveportions of the Irish Sea; more larvae remain offshore indeeper water. Averaged dispersal distances of passivelarvae after 28 d PLD were 622 km (Figs. 6a, 7b), whichwas higher than that of tidal larvae (322 km; Figs. 6b, 7b).Larvae performing tidal migration would be either retainednear the natal shore or transported offshore toward othercoastal locations, leading to a bimodal distribution ofdistances (Fig. 6b). For instance, model simulations pre-dicted that a group of larvae from Milford Havenperforming tidal migration were retained in South Waleswhile a second group crossed the Irish Sea and reached theIrish coast (Fig. 8a). Contrary to passive and tidal larvae,diel larvae remained close to their natal site (i.e., within aradial distance of 50 km; Fig. 6c). Most of these larvaeencountered onshore coastal currents and also spent moretime in weaker bottom currents, promoting retention andconnectivity with nearby populations.
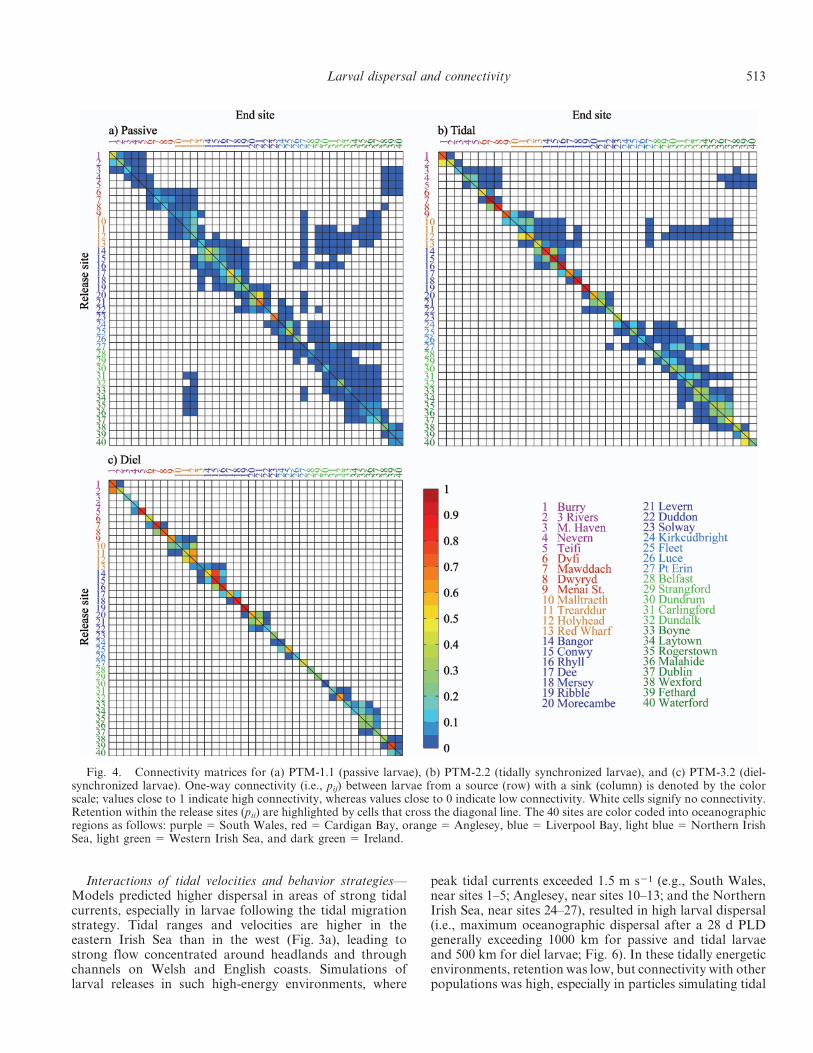
Overall, the above dispersal patterns resulted in retention(Fig. 4; diagonal cells of the connectivity matrices) beinggenerally higher for larvae following diel or tidal migration(42% and 43%, respectively) than for passive larvae (18%;Table 1). However, tidal larvae still favored connectivityacross the shelf (Fig. 4b) without the major losses producedby passive larvae.
Interactions of local bathymetry and behavior strategies—Some of the significant spatial variation in our variables(see Table 2) reflect differences in the bathymetry of theestuary or embayment and the coastline surrounding therelease zone. Relatively long estuaries (e.g., Burry Inlet: site1, Mersey Estuary: site 18, Solway Firth: site 23, andDundalk Estuary: site 32) tended to promote flood-dominant net flow (Robins and Davies 2010), and hencelarvae released here showed lower dispersal distances(Fig. 6) and higher retention (Fig. 4). However, larvaereleased from more exposed coastlines, open bays, andharbors (e.g., Trearddur Bay: site 11, Holyhead: site 12,and Port Erin: site 27) led to greater dispersal (Fig. 6) andreduced retention (Fig. 4). These effects were also influ-enced by strategy. For example, retention levels of 45% forpassive larvae and more than 80% for tidal and diel larvaewere simulated from releases in the Burry Inlet in SouthWales (Fig. 4), which is over 15 km in length. In contrast,larvae released from Trearddur Bay were quickly exposedto strong tidal currents off the north coast of Anglesey;many larvae traveled far (Fig. 6), and retention wasparticularly low (less than 40%), especially for passivelarvae (less than 10%; Fig. 4).
Peninsulas act as a barrier to transport (Drake et al.2011). Predictions here, for instance, show that larvaereleased from the Dyfi (site 6) and Mawddach (site 7)Estuaries remained predominantly in Cardigan Bay,trapped by the Llyn Peninsula to the north (Figs. 4, 6, 8b).
512 Robins et al.
Interactions of tidal velocities and behavior strategies—Models predicted higher dispersal in areas of strong tidalcurrents, especially in larvae following the tidal migrationstrategy. Tidal ranges and velocities are higher in theeastern Irish Sea than in the west (Fig. 3a), leading tostrong flow concentrated around headlands and throughchannels on Welsh and English coasts. Simulations oflarval releases in such high-energy environments, where
peak tidal currents exceeded 1.5 m s21 (e.g., South Wales,near sites 1–5; Anglesey, near sites 10–13; and the NorthernIrish Sea, near sites 24–27), resulted in high larval dispersal(i.e., maximum oceanographic dispersal after a 28 d PLDgenerally exceeding 1000 km for passive and tidal larvaeand 500 km for diel larvae; Fig. 6). In these tidally energeticenvironments, retention was low, but connectivity with otherpopulations was high, especially in particles simulating tidal
Fig. 4. Connectivity matrices for (a) PTM-1.1 (passive larvae), (b) PTM-2.2 (tidally synchronized larvae), and (c) PTM-3.2 (diel-synchronized larvae). One-way connectivity (i.e., pij) between larvae from a source (row) with a sink (column) is denoted by the colorscale; values close to 1 indicate high connectivity, whereas values close to 0 indicate low connectivity. White cells signify no connectivity.Retention within the release sites (pii) are highlighted by cells that cross the diagonal line. The 40 sites are color coded into oceanographicregions as follows: purple 5 South Wales, red 5 Cardigan Bay, orange 5 Anglesey, blue 5 Liverpool Bay, light blue 5 Northern IrishSea, light green 5 Western Irish Sea, and dark green 5 Ireland.
Larval dispersal and connectivity 513

migration or passive dispersal (Fig. 4). For instance,Fig. 8a shows that high dispersal is predicted for MilfordHaven (site 3), Holyhead (site 12), and Dublin (site 37),which are close to strong tidal velocities and also residualcurrents.
By contrast, in Cardigan Bay peak tidal velocities wereless than 0.5 m s21 (Fig. 3a). Simulations of larval releasesfrom this bay (sites 6–9) generally did not result indispersion beyond the bay (e.g., Dyfi Estuary; Figs. 6,8b). A similar pattern is seen in other low-energyenvironments, such as Liverpool Bay (sites 14–23) andthe Western Irish Sea (sites 28–32), where tidal velocitieswere less than 1 m s21 (Fig. 6). As a result, the predictedlarval dispersal was generally low in these areas, especiallyin larvae following diel or tidal migration (e.g., Figs. 6, 8b,which show relatively low dispersal from Dyfi [site 6],Morecambe [Site 20], and Dundalk [site 32]).
The patterns described above are supported by correla-tions between retention (or connectivity) and dispersaldistance, used as a proxy for tidal energy (Fig. 9a,b). Forpassive and tidal strategies, increased dispersal distancesled to reduced retention and also reduced connectivity,suggesting that most connectivity occurs with neighboringsites. Diel migration, however, shows weaker trends inretention and connectivity due to dispersal distance (see95th- quantile regression lines in Fig. 9a,b) because thestrategy dictates that more larvae remain close to the natalsite. This important result implies generally that retentionand connectivity are reduced for larvae that becomeexposed to high-energy environments.
Patterns of connectivity show an effect of migrationstrategy that also depends on the circulation surroundingthe release site. For instance, strong tidal velocities (greaterthan 1.5 m s21; Fig. 3a) in the central Irish Sea causetransport of passive and tidal larvae from Anglesey to theIrish coast but much less transport of diel larvae (Fig. 4).However, the predicted proportion of larvae reaching anew site is low (Fig. 5b): in 17 out of 40 passive populationsand in 20 tidal and 15 diel populations, less than 10% oflarvae reached a new site.
Interactions of residual currents and behavior strategies—Dispersal was influenced by residual flow pathways.Mesoscale density-driven or baroclinic flows develop alongoceanographic fronts where stratified regions meet well-mixed waters and, during summer months, contributesignificantly to the circulation of the Irish Sea (Fig. 3b).Controls on dispersal distance due to residual circulationwere enhanced for tidal and diel strategies, compared withpassive. For example, Liverpool Bay populations (sites 14–23) show considerably more dispersal of passive than activelarvae (Fig. 6) because onshore residual currents in theregion (Fig. 3b) promote retention of active larvae.
High dispersal was predicted, as surface heating led toincreased stratification during summer months, generatingstronger residual baroclinic currents. For example, signif-icant proportions of passive larvae, released in MilfordHaven (site 3) during July and August, reached the Irishcoast, whereas for spring releases, larvae remained closeto Wales (Fig. 10a). Monthly averaged oceanographic
Fig. 5. Graphs that show levels of (a) larval retention, (b) larval connectivity, and (c) larvae that did not reach an end site after 28 dfor passive, tidally synchronized, and diel-synchronized larvae patches from the 40 release sites. For example, (a) shows that (seasonallyaveraged) larval retention was lower than 10% in 18 estuaries or bays for passive larvae and in fewer than five estuaries or bays for larvaewith tidally or diel-synchronized migration.
514 Robins et al.

dispersal distances of passive larvae (Fig. 7a) increasedfrom 500 km (April) to approximately 650 km (June–September). In contrast, tidally synchronized larvae in-creased moderately over the season, and diel larvae showedlittle change (Fig. 7a). Seasonal variability appeared not toinfluence dispersal of diel larvae because, as strongerbaroclinic currents develop during summer months, diellarvae spend longer periods in weaker near-bed currentsbecause of increased daylight hours.
Residual currents caused strong directionalities inconnectivity patterns, but averaged retention decreasedonly slightly over the season (Fig. 9c), and averagedconnectivity showed a weak increasing trend (Fig. 9d).The fate of larvae that are entrained by these residualcurrents is ultimately controlled by them until they reachland, where coastal currents become important for larvaltransport. Vertical migration strategies advanced or re-tarded the advection of larvae but did not notably altertheir trajectories. For example, connectivity of larvae fromSouth Wales (in particular, sites 3–5) with populationsalong the south coast of Ireland (sites 36–40) was low
(, 5%) but evident for passive and tidal strategies (Fig. 4).This pattern is due to the westerly residual currents of theCeltic Sea front (Fig. 3b; see also Milford Haven releases inFig. 10a), whereas there was no reciprocal connectivity ofIrish larval populations with Welsh and English sites.
Another example shows moderate connectivity (13–20%,depending on strategy; Fig. 4) of larvae released fromNorth Wales (sites 14–18) with neighboring estuaries to thewest due to the westward residual current along the NorthWales coast (Fig. 3b). Averaged retention at North Walessites was 33%, 80%, and 73% for passive, tidal, and dielstrategies, respectively (Fig. 4). Considering the WesternIrish Sea gyre and also the southerly residual currents nearthe east coast of Ireland (Fig. 3b), simulated larvae releasedfrom Northern Ireland and the Republic of Ireland (sites28–40) migrated south or southwest and remained in thewestern Irish Sea (e.g., Dublin releases of passive larvae inFig. 8). Therefore, baroclinic currents associated with theWestern Irish Sea gyre increased larval dispersal in tidallycalm environments. However, for some scenarios, connec-tivity of Irish populations with neighboring populations to
Fig. 6. Twenty-eight-day PLD oceanographic dispersal distances (i.e., trajectory distances from source to sink) of larvae from eachrelease site, with (a) passive transport (PTM-1.1), (b) tidal stream transport (PTM-2.2), and (c) diel transport (PTM-3.2). The horizontallength of each graph corresponds to the maximum dispersal distance (in kilometers) from all larvae released at that site, that is, sixsimulations of 10,000 totaling 60,000 larvae. Triangles denote mean values, and the dotted lines correspond to the 25th and 75thpercentiles. Vertical axes represent the proportion of all released larvae from that site and range from 0 (origin) to 30%. The followingoceanographic regions have been highlighted: South Wales, Cardigan Bay, Anglesey, Liverpool Bay, Northern Irish Sea, Western IrishSea, and Ireland.
Larval dispersal and connectivity 515
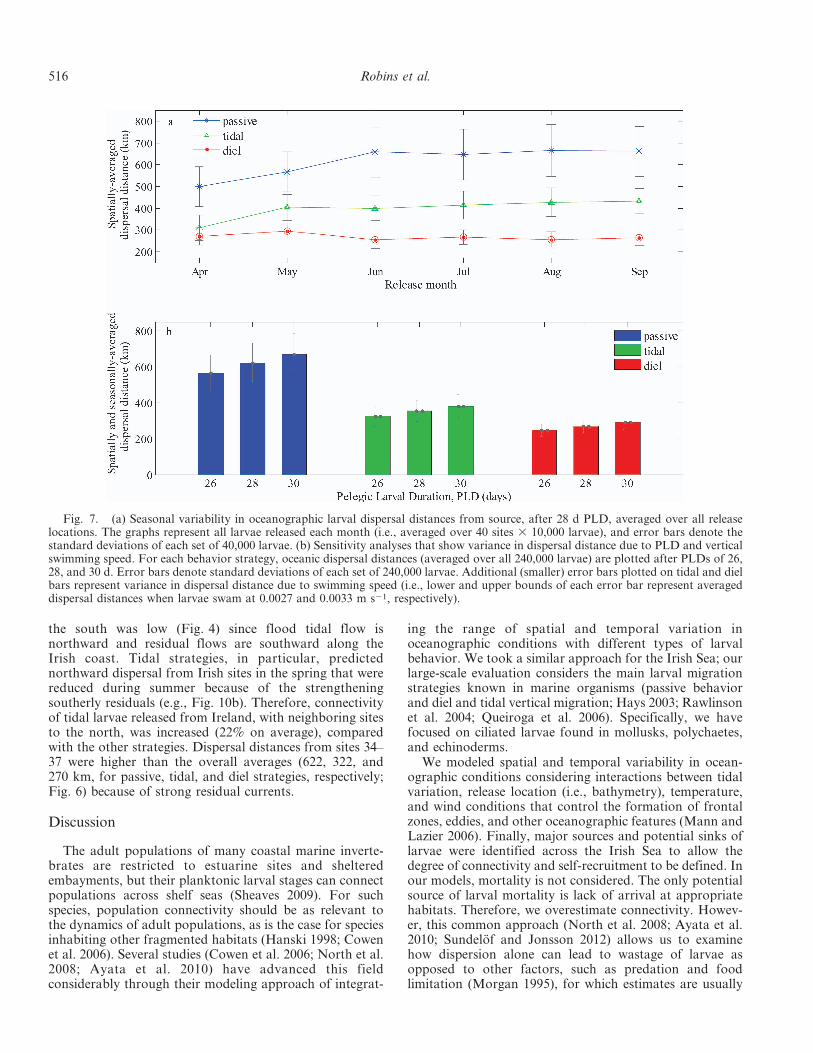
the south was low (Fig. 4) since flood tidal flow isnorthward and residual flows are southward along theIrish coast. Tidal strategies, in particular, predictednorthward dispersal from Irish sites in the spring that werereduced during summer because of the strengtheningsoutherly residuals (e.g., Fig. 10b). Therefore, connectivityof tidal larvae released from Ireland, with neighboring sitesto the north, was increased (22% on average), comparedwith the other strategies. Dispersal distances from sites 34–37 were higher than the overall averages (622, 322, and270 km, for passive, tidal, and diel strategies, respectively;Fig. 6) because of strong residual currents.
Discussion
The adult populations of many coastal marine inverte-brates are restricted to estuarine sites and shelteredembayments, but their planktonic larval stages can connectpopulations across shelf seas (Sheaves 2009). For suchspecies, population connectivity should be as relevant tothe dynamics of adult populations, as is the case for speciesinhabiting other fragmented habitats (Hanski 1998; Cowenet al. 2006). Several studies (Cowen et al. 2006; North et al.2008; Ayata et al. 2010) have advanced this fieldconsiderably through their modeling approach of integrat-
ing the range of spatial and temporal variation inoceanographic conditions with different types of larvalbehavior. We took a similar approach for the Irish Sea; ourlarge-scale evaluation considers the main larval migrationstrategies known in marine organisms (passive behaviorand diel and tidal vertical migration; Hays 2003; Rawlinsonet al. 2004; Queiroga et al. 2006). Specifically, we havefocused on ciliated larvae found in mollusks, polychaetes,and echinoderms.
We modeled spatial and temporal variability in ocean-ographic conditions considering interactions between tidalvariation, release location (i.e., bathymetry), temperature,and wind conditions that control the formation of frontalzones, eddies, and other oceanographic features (Mann andLazier 2006). Finally, major sources and potential sinks oflarvae were identified across the Irish Sea to allow thedegree of connectivity and self-recruitment to be defined. Inour models, mortality is not considered. The only potentialsource of larval mortality is lack of arrival at appropriatehabitats. Therefore, we overestimate connectivity. Howev-er, this common approach (North et al. 2008; Ayata et al.2010; Sundelof and Jonsson 2012) allows us to examinehow dispersion alone can lead to wastage of larvae asopposed to other factors, such as predation and foodlimitation (Morgan 1995), for which estimates are usually
Fig. 7. (a) Seasonal variability in oceanographic larval dispersal distances from source, after 28 d PLD, averaged over all releaselocations. The graphs represent all larvae released each month (i.e., averaged over 40 sites 3 10,000 larvae), and error bars denote thestandard deviations of each set of 40,000 larvae. (b) Sensitivity analyses that show variance in dispersal distance due to PLD and verticalswimming speed. For each behavior strategy, oceanic dispersal distances (averaged over all 240,000 larvae) are plotted after PLDs of 26,28, and 30 d. Error bars denote standard deviations of each set of 240,000 larvae. Additional (smaller) error bars plotted on tidal and dielbars represent variance in dispersal distance due to swimming speed (i.e., lower and upper bounds of each error bar represent averageddispersal distances when larvae swam at 0.0027 and 0.0033 m s21, respectively).
516 Robins et al.

Fig. 8. Dispersal probability distributions (dispersal kernels) of larvae-particles after 28 d PLD from releases at selected high-velocity environments (Milford Haven [site 3]), Holyhead [site 12], and Dublin [site 37]) and low-velocity environments (Dyfi Estuary [site6], Morecambe Bay [site 20], and Dundalk Estuary [site 32]). For each selected release location, dispersal kernels for (a) passive (PTM-1.1.1), (b) tidally synchronized (PTM-2.2), and (c) diel-synchronized (PTM-3.2) larvae are shown. Each dispersal kernel represents theseasonally averaged trajectories, that is, probability distributions of larvae released throughout April–September (six releases of cohortsof 10,000 larvae totaling 60,000 larvae). In reality, dispersal distances were greater during summer, as stratification and hence residualcurrents were stronger (see Fig. 5). The color scale indicates the proportion of total larvae located in each model cell (the scale has beencapped at 0.02% for clearer visualization in less populated cells, although proportions in each cell did not exceed 0.05%).
Larval dispersal and connectivity 517

variable, species dependent, but also unreliable (Morgan1995). Mortality should not change connectivity patternsunless it varies in space (Paris et al. 2007), but meaningfulintroduction of mortality patterns requires knowledge of,for example, predator and prey fields. This will be moreappropriate for species specific models. Some of the keyparameters of our models, such as pelagic durations andswimming speed, affect larval transport (Ayata et al. 2010;Sundelof and Jonsson 2012), but our sensitivity analysesshow that results are robust to small variations in theseparameters. Indeed, simulations with vertical migrationand swimming speeds in the range 2.5 3 102420.005 m s21
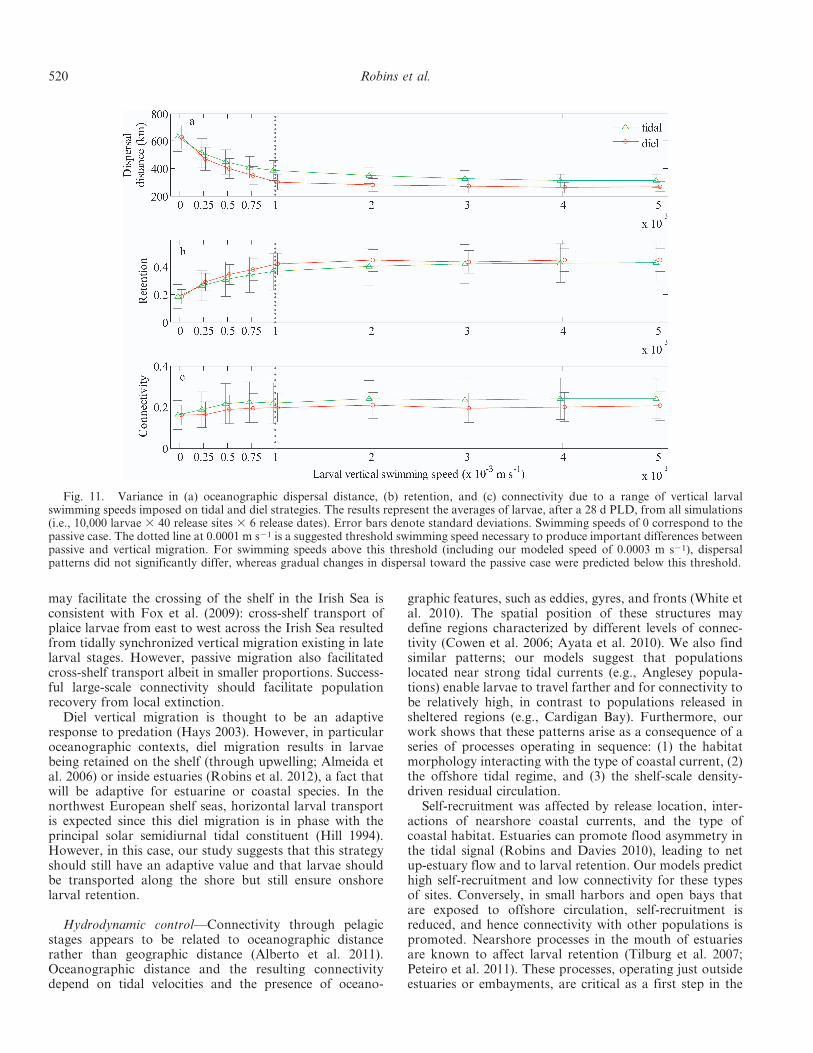
showed that above a threshold speed of 0.001 m s21,dispersal patterns remained similar (Fig. 11). Below thisthreshold, dispersal patterns gradually changed to thepassive case. Ontogenetic changes through the larval stageare also likely to change dispersal patterns, though againthis will be more appropriate for species-specific models.
Our modeling results suggest that connectivity and self-recruitment of these invertebrates are affected by the larvalstrategy but are also controlled by the hydrodynamiccontext, including, importantly, seasonal variability. Stud-ies that make use of only depth-integrated velocities (e.g.
Proctor et al. 2004) may not accurately resolve hydrody-namic dispersal.
Vertical migration strategy—Several studies suggest thatwide-scale dispersal of larvae and, hence, generally highlevels of connectivity may be limited (Jones et al. 1999;Swearer et al. 1999). Our study shows that the potential fordispersal depends strongly on the larval strategy. Passivelarvae showed high dispersal but resulted in high wastageof larvae, tidally synchronized larvae contributed to large-scale connectivity across the shelf sea and to self-recruitment through a minimization of the larval wastage,and daily synchronized larvae contributed to self-recruit-ment and had a low larval wastage. Therefore, a significantconclusion is that tidally or daily synchronized behaviorswill be an advantageous behavioral traits in species living inestuaries and bays.
Passive larvae followed less distinct pathways becauselarvae spread out vertically and, hence, experienceddifferent three-dimensional currents. The connectivitymatrix suggests a localized stepping-stone process withlarvae weakly connecting adjacent sites. This result isstrikingly similar to that of Cowen et al. (2006), who stated
Fig. 9. Controls on retention and connectivity in the light of different behavior strategies: patch-averaged oceanographic dispersaldistance (a proxy for tidal energy; a and b, respectively) and release time (a proxy for the strength of baroclinic currents; c and d,respectively). Each scatter graph contains (a) retention and (b) connectivity values for simulations after 28 d PLD and (where applicable)swimming speeds of 0.003 m s21. Quantile regression (solid) lines plotted in (a) and (b) denote the 95th quantile for each behaviorstrategy, and corresponding dashed lines represent the 95% confidence intervals. R2 values are also presented. Lines in (c) and (d)represent mean retention and mean connectivity, respectively, each month and for each strategy. Error bars denote standard deviations.
518 Robins et al.

that ‘‘passive dispersal is insufficient for populationreplenishment.’’ The concordance in findings between theCaribbean and the Irish Sea suggests that there is potentialfor generalities in this complex area to emerge.
In tidal and diel strategies, larvae should have highertransport speeds than for passive larvae because migrationcauses larvae to remain close together vertically; hence,offshore baroclinic currents will affect the majority of the
cohort similarly. In the case of tidally migrating larvae, thisis consistent with the concept of selective tidal streamtransport (Forward and Tankersley 2001). For a semien-closed body of water such as the Irish Sea, modelpredictions are that either these currents should retainlarvae near the coast or, if transported offshore, larvaewould become entrained in residual pathways and henceencounter the distant coast. The fact that tidal migration
Fig. 10. Dispersal probability distributions (dispersal kernels) after 28 d PLD for (a) passive larvae released from Milford Haven(site 3) and (b) tidal larvae released from Wexford Bay (site 38; release sites are marked with yellow triangles). Each dispersal kernelshows a new simulation of 10,000 larvae released each month (April–September). The color scale indicates the proportion of totalparticles located in each model cell. Successive panels in (a) show that strengthening westward baroclinic currents throughout the seasonled to increased dispersal of the larvae patch and conduit larval transport toward Ireland within strong residual currents. Successivepanels in (b) show that northward flood currents advect tidally synchronized larvae northward. However, strengthening southerlydensity-driven flow, from July on, acts to retard northward dispersal.
Larval dispersal and connectivity 519
may facilitate the crossing of the shelf in the Irish Sea isconsistent with Fox et al. (2009): cross-shelf transport ofplaice larvae from east to west across the Irish Sea resultedfrom tidally synchronized vertical migration existing in latelarval stages. However, passive migration also facilitatedcross-shelf transport albeit in smaller proportions. Success-ful large-scale connectivity should facilitate populationrecovery from local extinction.
Diel vertical migration is thought to be an adaptiveresponse to predation (Hays 2003). However, in particularoceanographic contexts, diel migration results in larvaebeing retained on the shelf (through upwelling; Almeida etal. 2006) or inside estuaries (Robins et al. 2012), a fact thatwill be adaptive for estuarine or coastal species. In thenorthwest European shelf seas, horizontal larval transportis expected since this diel migration is in phase with theprincipal solar semidiurnal tidal constituent (Hill 1994).However, in this case, our study suggests that this strategyshould still have an adaptive value and that larvae shouldbe transported along the shore but still ensure onshorelarval retention.
Hydrodynamic control—Connectivity through pelagicstages appears to be related to oceanographic distancerather than geographic distance (Alberto et al. 2011).Oceanographic distance and the resulting connectivitydepend on tidal velocities and the presence of oceano-
graphic features, such as eddies, gyres, and fronts (White etal. 2010). The spatial position of these structures maydefine regions characterized by different levels of connec-tivity (Cowen et al. 2006; Ayata et al. 2010). We also findsimilar patterns; our models suggest that populationslocated near strong tidal currents (e.g., Anglesey popula-tions) enable larvae to travel farther and for connectivity tobe relatively high, in contrast to populations released insheltered regions (e.g., Cardigan Bay). Furthermore, ourwork shows that these patterns arise as a consequence of aseries of processes operating in sequence: (1) the habitatmorphology interacting with the type of coastal current, (2)the offshore tidal regime, and (3) the shelf-scale density-driven residual circulation.
Self-recruitment was affected by release location, inter-actions of nearshore coastal currents, and the type ofcoastal habitat. Estuaries can promote flood asymmetry inthe tidal signal (Robins and Davies 2010), leading to netup-estuary flow and to larval retention. Our models predicthigh self-recruitment and low connectivity for these typesof sites. Conversely, in small harbors and open bays thatare exposed to offshore circulation, self-recruitment isreduced, and hence connectivity with other populations ispromoted. Nearshore processes in the mouth of estuariesare known to affect larval retention (Tilburg et al. 2007;Peteiro et al. 2011). These processes, operating just outsideestuaries or embayments, are critical as a first step in the
Fig. 11. Variance in (a) oceanographic dispersal distance, (b) retention, and (c) connectivity due to a range of vertical larvalswimming speeds imposed on tidal and diel strategies. The results represent the averages of larvae, after a 28 d PLD, from all simulations(i.e., 10,000 larvae 3 40 release sites 3 6 release dates). Error bars denote standard deviations. Swimming speeds of 0 correspond to thepassive case. The dotted line at 0.0001 m s21 is a suggested threshold swimming speed necessary to produce important differences betweenpassive and vertical migration. For swimming speeds above this threshold (including our modeled speed of 0.0003 m s21), dispersalpatterns did not significantly differ, whereas gradual changes in dispersal toward the passive case were predicted below this threshold.
520 Robins et al.

process of connectivity, as they affect the proportion ofindividuals to be retained or dispersed away from the natalhabitat.
Once larvae are away from the natal habitats, patterns ofretention or dispersal appear to be controlled by barotropicand baroclinic flows. Therefore, these components to thecirculation must be accurately represented in hydrodynam-ic models for the correct application of PTM simulations.The tidal ranges and the associated barotropic velocities aregreater in the eastern Irish Sea than in the west and greaterstill around headlands and through channels. Such hightidal-energy environments, where peak velocities weregreater than 1.5 m s21, led to high larval dispersal,especially in tidally synchronized larvae. While low tidal-energy environments (e.g., Western Irish Sea) produced lesslarval dispersal. This has important implications for theecology and evolution of marine populations: the fitness oflife history strategies based on tidally synchronized larvaewill be very sensitive to local variations in tidal currents ascompared with strategies-based diel migration. In addition,the strong spatial pattern in dispersal predicted for thisstrategy should result in a metapopulation characterized byisolated regions (e.g., Liverpool Bay) and regions showinghigh connectivity. Thus, the capacity to recover from localextinction will vary regionally in response to tidal currents.Weak baroclinic currents maximized retention, especiallyfor tidally synchronized or diel larval strategies. Bycontrast, offshore seasonal baroclinic flows, which generatepersistent residual circulations, also affect larval dispersal.Indeed, it is these currents, rather than oscillatory tidalflows, that control larval dispersal patterns on a time scaleof several weeks. For instance, residual currents areresponsible for considerable transport of plaice eggs andlarvae in the North Sea (Cushing 1990). According toEmsley et al. (2005), larval transport in the Irish Seaappears to depend on whether organisms are trapped in theWestern Irish Sea gyre, a key oceanographic structure ofthe Irish Sea (Horsburgh and Hill 2003). The Irish Sea gyrehas previously been studied in relation to the retention ofNorway lobster larvae (Hill et al. 1996). Our results wereconsistent with the above literature in the fact that larvaldispersal trajectories were longer during the summer thanduring spring, when the gyre is more developed andresidual currents are stronger. Actual traveled distanceswill also be affected by thermal responses of larvaldevelopmental rates (Duarte 2007). The appropriateevaluation of the balance of seasonal changes in residualcirculation and developmental rates will, however, requirespecies-specific studies because these rates vary consider-ably among species (O’Conner et al. 2007; Gimenez 2011).More interesting is the fact that females from a localpopulation releasing larvae at different points in the seasonmay connect with different local populations: this capacitywill vary among species in response to variations in thelength of the spawning season and according to the larvalstrategy.
Another important result is that a residual pathway inthe Irish Sea forms along the Celtic Sea front, and modelspredict that larvae from Welsh populations reach the Irishcoast. Genetic analysis of cockle C. edule samples from
both coasts by Coscia et al. (2012) revealed geneticsimilarities between Irish and Southwest Wales samples,in accordance with our simulations. The population ofBurry Inlet was genetically isolated, again, as our modelspredict.
Thus, the interaction of larval behavior and shelfprocesses is the key to understanding cross-shelf transportand connectivity. While tidal energy and baroclinic flowsaffect the magnitude of dispersal, migration behaviorappears to affect how successfully larvae keep to the shoreand cross the shelf.
A specific application—Our results can be related tospecific cases. For example, Knights et al. (2006) studiedthe mussel M. edulis larvae in the southern Irish Sea (nearsite 38). They found that larvae spawned in late April tolate June and followed tidal migration patterns. Knights etal. (2006) predicted that this strategy would lead to aconsiderable northward movement of larvae since floodflow is northward in this region. Indeed, our simulationspredicted exactly this pattern, with maximum northwardradial dispersal of 180 km from May releases (Fig. 10b).Although Knights et al. (2006) did not consider thebaroclinic component to the circulation, our simulationsshow that northward dispersal of tidal larvae was reducedfrom July to September because of strengthening southerlybaroclinic flow along the western coast of Ireland (Fig. 3b).However, since M. edulis larvae are thought to spawn inspring, the baroclinic component to larval transport maynot be significant.
Our modeling exercise points to the importance ofinteractions between oceanographic processes and larvalbehavior to predict patterns of population connectivity,self-recruitment, and wastage of larvae of coastal marineorganisms. In the case of the Irish Sea, vertical migrationstrategies contribute to both connectivity and self-recruit-ment and limit the extent of larval wastage because theyensure self-recruitment or rapid cross-shelf transport (tidalbehavior). Self-recruitment and connectivity are affected bya series of nearshore processes (e.g., flood-tide asymmetriesproduced by coastal morphology and coastal barriers) andoffshore processes (tidal energy and baroclinic circulation),but the effect of these processes is stronger on tidallysynchronized larvae than with any other behavior. Larvalbehavior should therefore affect the capacity of localpopulations to recover from extinction by recolonizationfrom other populations, but at the same time this capacityshould vary among regions of the metapopulation inresponse to spatial variations in bathymetry, tidal energy,and residual currents. In addition, temporal changes inbaroclinic circulation should produce both intra- andinterspecific variations in the patterns of connectivity inresponse to variations in the timing and extension of thespawning season. These issues deserve further investigation.
Also as a perspective, it is our aim to investigate theeffect of climate change on larval dispersal within the IrishSea region. Predictions are that global atmosphericconditions will substantially influence the northwestEuropean continental shelf environment with far-reachingconsequences for their ecosystems (Holt et al. 2010). The
Larval dispersal and connectivity 521

Irish Sea might warm substantially (21st-century trend:, 0.3uC decade21). As a consequence, frontal densitygradients may increase, strengthening the associated bar-oclinic currents. Sea levels in the Irish Sea are predicted torise 1 m in the next 100 yr, which may reduce tidal ranges andvelocities in shallow areas (Robins and Davies 2010). Otherprocesses may not vary much: interannual variability in thetiming and strength of stratification in the Irish Sea isrelatively small when compared to seasonal change (Youngand Holt 2007). Negligible change in wind stress is predicted(Lowe et al. 2009; Holt et al. 2010). It is our opinion, thatenergy within the system will increase, which will have apositive effect on larval dispersal distances, on the whole, butit is not known how climate change will specifically affectlarval retention, connectivity, and behavior.
AcknowledgmentsThe project was partly funded by the European Union
Regional Development Fund (ERDF) under the Ireland WalesProgramme 2007–2013—Interreg 4A, Project 042. Alan M.Davies and Jiuxing Xing at the National Oceanographic Centre,Liverpool, provided modeled data (Xing and Davies 2001) forvalidation purposes. We would like to thank the reviewers of themanuscript for helpful and constructive comments.
References
ALBERTO, F., P. T. RAIMONDI, D. C. REED, J. R. WATSON, D. A.SIEGEL, S. MITARAI, N. COELHO, AND E. A. SERRAO. 2011.Isolation by oceanographic distance explains genetic structurefor Macrocystis pyrifera in the Santa Barbara Channel. Mol.Ecol. 20: 2543–2554, doi:10.1111/j.1365-294X.2011.05117.x
ALMEIDA, M. J., J. DUBERT, A. PELIZ, AND H. QUEIROGA. 2006.Influence of vertical migration patterns on retention of crablarvae in a seasonal upwelling system. Mar. Ecol. Prog. Ser.307: 1–19, doi:10.3354/meps307001
AYATA, S-D., P. LAZURE, AND E. THIEBAUT. 2010. How does theconnectivity between populations mediate limits of marineinvertebrates? A case study of larval dispersal between theBay of Biscay and the English Channel (North-East Atlantic).Prog. Oceanogr. 87: 18–36, doi:10.1016/j.pocean.2010.09.022
BLUMBERG, A. F., AND G. L. MELLOR. 1987. A description of athree-dimensional coastal ocean circulation model, p. 1–16. InN. S. Heaps [ed.], Three-dimensional coastal ocean models.American Geophysics Union.
BORTHAGARAY, A. I., A. BRAZEIRO, AND L. GIMENEZ. 2009.Connectivity and patch area in a coastal marine landscape: Disen-tangling their influence on local species richness and composi-tion. Aust. Ecol. 34: 641–652, doi:10.1111/j.1442-9993.2009.01969.x
CALEY, M. J., M. H. CARR, M. A. HIXON, T. P. HUGHES, G. P.JONES, AND B. A. MENGE. 1996. Recruitment and the localdynamics of open marine populations. Annu. Rev. Ecol.System. 27: 477–500, doi:10.1146/annurev.ecolsys.27.1.477
CHIA, F. S., J. BUCKLAND-NICKS, AND C. M. YOUNG. 1984.Locomotion of marine invertebrate larvae: A review. Can. J.Zool. 62: 1205–1222, doi:10.1139/z84-176
COSCIA, I., P. E. ROBINS, J. S. PORTER, S. K. MALHAM, AND J. E.IRONSIDE. 2012. Modelled larval dispersal and measured geneflow: Seascape genetics of the common cockle Cerastodermaedule in the southern Irish Sea. Conserv. Genet., doi:10.1007/s10592-012-0404-4
COWEN, R. K., C. B. PARIS, AND A. SRINIVASAN. 2006. Scaling ofconnectivity in marine populations. Science 311: 522–527,doi:10.1126/science.1122039
———, AND S. SPONAUGLE. 2009. Larval dispersal and marinepopulation connectivity. Annu. Rev. Mar. Sci. 1: 443–466,doi:10.1146/annurev.marine.010908.163757
CRIALES, M. M., J. A. BROWDER, C. K. MOOERS, M. B. ROBBLEE,H. CARDENAS, AND T. L. JACKSON. 2007. Cross-shelf transportof pink shrimp larvae: Interactions of tidal currents, larvalvertical migrations and internal tides. Mar. Ecol. Prog. Ser.345: 167–184, doi:10.3354/meps06916
CUSHING, D. H. 1990. Hydrographic containment of a spawninggroup of plaice in the Southern Bight of the North Sea. Mar.Ecol. Prog. Ser. 58: 287–297, doi:10.3354/meps058287
DOS SANTOS, A., A. SANTOS, D. CONWAY, C. BARTILOTTI, P.LOURENCO, AND H. QUEIROGA. 2008. Diel vertical migration ofdecapod larvae in the Portuguese coastal upwelling ecosys-tem: Implications for offshore transport. Mar. Ecol. Prog.Ser. 359: 171–183, doi:10.3354/meps07341
DRAKE, P. T., C. A. EDWARDS, AND J. A. BARTH. 2011. Dispersionand connectivity estimates along the U.S. west coast from arealistic numerical model. J. Mar. Res. 69: 1–37, doi:10.1357/002224011798147615
DUARTE, C. 2007. Marine ecology warms up to theory. TrendsEcol. Evol. 22: 331–333, doi:10.1016/j.tree.2007.04.001
EMSLEY, S. M., G. A. TARLING, AND M. T. BURROWS. 2005. Theeffect of vertical migration strategy on retention anddispersion in the Irish Sea during spring-summer. Fish.Oceanogr. 14: 161–174, doi:10.1111/j.1365-2419.2005.00327.x
FORWARD, R., JR, AND R. A. TANKERSLEY. 2001. Selective tidal-stream transport of marine animals. Ocean. Mar. Biol. Annu.Rev. 39: 305–353.
FOX, C. J., P. MCCLOGHRIE, AND R. M. NASH. 2009. Potentialtransport of plaice eggs and larvae between two apparentlyself-contained populations in the Irish Sea. Estuar. Coast.Shelf Sci. 81: 381–389, doi:10.1016/j.ecss.2008.10.024
GILG, M. R., AND T. J. HILBISH. 2003. The geography of marinelarval dispersal: Coupling genetics with fine-scale physicaloceanography. Ecology 84: 2989–2998, doi:10.1890/02-0498
GIMENEZ, L. 2011. Exploring mechanisms linking temperature increaseand larval phenology: The importance of variance effects. J. Exp.Mar. Biol. Ecol. 400: 227–235, doi:10.1016/j.jembe.2011.02.036
GRIMM, V., K. REISE, AND M. STRASSER. 2003. Marine metapop-ulations: a useful concept? Helgol. Mar. Res. 56: 222–228.
HANSKI, I. 1998. Metapopulation dynamics. Nature 396: 41–49,doi:10.1038/23876
HARTNETT, M., A. BERRY, O. TULLY, AND T. DABROWSKI. 2007.Investigations into the transport and pathways for scalloplarvae—the use of numerical models for managing fish stocks.J. Environ. Monit. 9: 403–410, doi:10.1039/b617035h
HAYS, G. C. 2003. A review of the adaptive significanceand ecosystem consequences of zooplankton diel verticalmigration. Hydrobiologia 503: 163–170, doi:10.1023/B:HYDR.0000008476.23617.b0
HILL, A. E. 1994. Horizontal zooplankton dispersal by dielvertical migration in S2 tidal currents on the northwestEuropean continental shelf. Cont. Shelf Res. 14: 491–506,doi:10.1016/0278-4343(94)90100-7
———, J. BROWN, AND L. FERNAND. 1996. The western Irish Seagyre: A retention system for Norway lobster (Nephropsnorvegicus)? Oceanologica Acta 19: 357–368.
HOLT, J. T., S. WAKELIN, J. LOWE, AND J. TINKER. 2010. Thepotential impacts of climate change on the hydrography of thenorthwest European continental shelf. Prog. Oceanogr. 86:361–379, doi:10.1016/j.pocean.2010.05.003
HORSBURGH, K. J., AND A. E. HILL. 2003. A three-dimensionalmodel of density-driven circulation in the Irish Sea. J. Phys.Oceanogr. 33: 343–365, doi:10.1175/1520-0485(2003)033,0343:ATDMOD.2.0.CO;2
522 Robins et al.

HOUDE, E. D. 2002. Mortality, p. 64–87. In L. A. Fuiman andR. G. Werner [eds.], Fishery science: The unique contribu-tions of early life stages. Blackwell Science.
JONES, G. P., M. J. MILICICH, M. J. EMSLIE, AND C. LUNOW. 1999.Self-recruitment in a coral reef fish population. Nature 402:802–804, doi:10.1038/45538
KNIGHTS, A. M., T. P. CROWE, AND G. BURNELL. 2006.Mechanisms of larval transport: Vertical distribution ofbivalve larvae varies with tidal conditions. Mar. Ecol. Prog.Ser. 326: 167–174, doi:10.3354/meps326167
LOWE, J., T. HOWARD, A. PARDAENS, J. TINKER, J. HOLT, S.WAKELIN, G. MILNE, J. LEAKE, J. WOLF, K. HORSBURGH, T.REEDER, G. JENKINS, J. RIDLEY, S. DYE, AND S. BRADLEY.2009. UK Climate Projections science report: Marine andcoastal projections. Met Office Hadley Centre, UK.
MACE, A. J., AND S. G. MORGAN. 2006. Larval accumulation in thelee of a small headland: Implications for the design of marinereserves. Mar. Ecol. Prog. Ser. 318: 19–29, doi:10.3354/meps318019
MANN, K. H., AND J. R. N. LAZIER. 2006. Dynamics of marineecosystems: Biological-physical interactions in the oceans.Wiley-Blackwell.
MCGURK, M. D. 1986. Natural mortality of marine pelagic fisheggs and larvae: Role of spatial patchiness. Mar. Ecol. Prog.Ser. 34: 227–242, doi:10.3354/meps034227
MELLOR, G. L., T. EZER, AND L. Y. OEY. 1994. The pressuregradient conundrum of sigma coordinate ocean models. J.Atmos. Ocean. Technol. 11: 1126–1134, doi:10.1175/1520-0426(1994)011,1126:TPGCOS.2.0.CO;2
MICHALEC, F.-G., S. SOUISSI, G. DUR, M.-S. MAHJOUB, F. G.SCHMITT, AND J.-S. HWANG. 2010. Differences in behavioralresponses of Eurytemora affinis (Copepoda, Calanoida)reproductive stages to salinity variations. J. Plankton Res.32: 805–813, doi:10.1093/plankt/fbq006
MORGAN, S. G. 1995. Life and death in the plankton: Larvalmortality and adaptation, p. 279–321. In L. McEdward [ed.],Ecology of marine invertebrate larvae. CRC Press.
NEILL, S. P., J. D. SCOURSE, AND K. UEHARA. 2010. Evolution ofbed shear stress distribution over the northwest Europeanshelf seas during the last 12,000 years. Ocean Dyn. 60:1139–1156, doi:10.1007/s10236-010-0313-3
NORTH, E. W., R. R. HOOD, S.-Y. CHAO, AND L. P. SANFORD. 2006.Using a random displacement model to simulate turbulentparticle motion in a baroclinic frontal zone: A newimplementation scheme and model performance tests. J.Mar. Sys. 60: 365–380, doi:10.1016/j.jmarsys.2005.08.003
———, Z. SCHLAG, R. R. HOOD, L. ZHONG, T. GROSS, AND V. S.KENNEDY. 2008. Vertical swimming behaviour influences thedispersal of simulated oyster larvae in a coupled particle-tracking and hydrodynamic model of Chesapeake Bay. Mar.Ecol. Prog. Ser. 359: 99–115, doi:10.3354/meps07317
O’CONNOR, M., J. BRUNO, S. GAINES, B. HALPERIN, S. LESTER, B.KINLAN, AND J. WEISS. 2007. Temperature control of larvaldispersal and the implications for marine ecology, evolution,and conservation. Proc. Natl. Acad. Sci. USA 104:1266–1271, doi:10.1073/pnas.0603422104
PARIS, C. B., L. M. CHERUBIN, AND R. K. COWEN. 2007. Surfing,spinning, or diving from reef to reef: Effects on populationconnectivity. Mar. Ecol. Prog. Ser. 347: 285–300, doi:10.3354/meps06985
PETEIRO, L. G., U. LABARTA, M. J. FERNANDEZ-REIRIZ, X. A.ALVAREZ-SALGADO, R. FILGUEIRA, AND S. PIEDRACOBA. 2011.Influence of intermittent-upwelling on Mytilus galloprovincia-lis settlement patterns in the Rıa de Ares-Betanzos. Mar. Ecol.Prog Ser. 443: 111–127, doi:10.3354/meps09433
PETERSON, I., AND J. S. WROBLESKI. 1984. Mortality rate of fishesin the pelagic ecosystem. Can. J. Fish. Aquat. Sci. 41:1117–1120, doi:10.1139/f84-131
PROCTOR, R., R. A. FLATHER, AND A. J. ELLIOTT. 1994. Modellingtides and surface drift in the Arabian Gulf—application to theGulf oil spill. Cont. Shelf Res. 14: 531–545, doi:10.1016/0278-4343(94)90102-3
QUEIROGA, H., M.-J. ALMEIDA, T. ALPUIM, A. V. FLORES, S.FRANCISCO, I. GONZALEZ-GORDILLO, A. I. MIRANDA, I. SILVA,AND J. PAULA. 2006. Tide and wind control of megalopal supplyto estuarine crab populations on the Portuguese west coast.Mar. Ecol. Prog. Ser. 307: 21–36, doi:10.3354/meps307021
RAWLINSON, K., J. DAVENPORT, AND D. BARNES. 2004. Verticalmigration strategies with respect to advection and stratifica-tion in a semi-enclosed lough: A comparison of mero- andholozooplankton. Mar. Biol. 144: 935–946, doi:10.1007/s00227-003-1261-y
ROBINS, P. E., AND A. G. DAVIES. 2010. Morphological controls insandy estuaries: The influence of tidal flats and bathymetry onsediment transport. Ocean Dyn. 60: 503–517, doi:10.1007/s10236-010-0268-4
———, S. P. NEILL, AND L. GIMENEZ. 2012. A numerical study ofmarine larval dispersal in the presence of an axial convergentfront. Estuar. Coast. Shelf Sci. 100: 172–185, doi:10.1016/j.ecss.2012.02.001
ROBINSON, I. S. 1979. The tidal dynamics of the Irish and CelticSeas. Geophys. J. R. Astron. Soc. 56: 159–197, doi:10.1111/j.1365-246X.1979.tb04774.x
ROBINSON, K. A., K. RAMSAY, C. LINDERBAUM, N. FROST,J. MOORE, A. P. WRIGHT, AND D. PETREY. 2010. Predictingthe distribution of seabed biotopes in the southern IrishSea. Cont. Shelf Res. 31: 120–131, doi:10.1016/j.csr.2010.01.010
ROSS, O. N., AND J. SHARPLES. 2004. Recipe for 1-D Lagrangianparticle tracking models in space-varying diffusivity. Limnol.Oceanogr. 2: 289–302, doi:10.4319/lom.2004.2.289
ROUGHGARDEN, J., Y. IWASA, AND C. BAXTER. 1985. Demographictheory for an open marine population with space limitedrecruitment. Ecology 66: 54–67, doi:10.2307/1941306
SCHULTZ, S. T., J. H. R. GODDARD, T. M. GOSLINER, D. E. MASON,W. E. PENCE, G. R. MCDONALD, V. B. PEARSE, AND J. S.PEARSE. 2011. Climate-index response profiling indicateslarval transport is driving population fluctuations in nudi-branch gastropods from the northeast Pacific Ocean. Limnol.Oceanogr. 56: 749–763, doi:10.4319/lo.2011.56.2.0749
SEURONT, L. 2010. How does salinity influence the swimmingspeed of the estuarine calanoid copepod Eurytemora affinis? J.Plankton Res. 32: 1223–1225, doi:10.1093/plankt/fbq040
SHANKS, A. 1995. Mechanisms of cross-shelf dispersal of larvalinvertebrates and fish, p. 323–367. In L. McEdward [ed.],Ecology of marine invertebrate larvae. CRC Press.
SHEAVES, M. 2009. Consequences of ecological connectivity: Thecoastal ecosystem mosaic. Mar. Ecol. Prog. Ser. 391: 107–115,doi:10.3354/meps08121
SHIMA, J. S., E. G. NOONBURG, AND N. E. PHILLIPS. 2010. Lifehistory and matrix heterogeneity interact to shape metapop-ulation connectivity in spatially structured environments.Ecology 91: 1215–1224, doi:10.1890/08-2058.1
SIEGEL, D. A., B. P. KINLAN, B. GAYLORD, AND S. D. GAINES. 2003.Lagrangian descriptions of marine larval dispersion. Mar.Ecol. Prog. Ser. 260: 83–96, doi:10.3354/meps260083
———, S. MITARAI, C. J. COSTELLO, S. D. GAINES, B. E. KENDALL,R. R. WARNER, AND K. B. WINTERS. 2008. The stochasticnature of larval connectivity among nearshore marinepopulations. Proc. Natl. Acad. Sci. USA 105: 8974–8979,doi:10.1073/pnas.0802544105
Larval dispersal and connectivity 523

SIMMONS, A., S. UPPALA, D. DEE, AND S. KOBAYASHI. 2006. ERAinterim: New ECMWF reanalysis products from 1989onwards. ECMWF Newsl. 110: 25–35.
SIMPSON, J. H., AND J. R. HUNTER. 1974. Fronts in the Irish Sea.Nature 250: 404–406, doi:10.1038/250404a0
SOUISSI, S., F.-G. MICHALEC, G. DUR, M.-S. MAHJOUB, F. G.SCHMITT, AND J.-S. WANG. 2010. How does salinity influencethe swimming speed of the estuarine calanoid copepodEurytemora affinis? Reply. J. Plankton Res. 32: 1227–1229,doi:10.1093/plankt/fbq059
SUNDELOF, A., AND P. R. JONSSON. 2012. Larval dispersal and verticalmigration behaviour—a simulation study for short dispersal times.Mar. Ecol. 33: 183–193, doi:10.1111/j.1439-0485.2011.00485.x
SWEARER, S. E., J. E. CASELLE, D. W. LEA, AND R. R. WARNER.1999. Larval retention and recruitment in an island populationof a coral-reef fish. Nature 402: 799–802, doi:10.1038/45533
TIAN, R., C. CHEN, K. STOKEBURY, B. ROTHSCHILD, Q. XU, S. HU,G. COWLES, B. P. HARRIS, AND M. C. MARINO. 2009.Sensitivity analysis of sea scallop (Placopecten magellanicus)larvae trajectories to hydrodynamic model configuration onGeorges Bank and adjacent coastal regions. Fish. Oceanogr.18: 1365–2419, doi:10.1111/j.1365-2419.2009.00506.x
TILBURG, C., A. I. DITTEL, AND C. E. EPIFANIO. 2007. Retention ofcrab larvae in a coastal null zone. Estuar. Coast. Shelf Sci. 67:570–578, doi:10.1016/j.ecss.2006.11.030
———, J. E. SEAY, T. D. BISHOP, H. L. I. MILLER, AND C. MEILE.2010. Distribution and retention of Petrolisthes armatus in acoastal plain estuary: The role of vertical movement in larvaltransport. Estuar. Coast. Shelf Sci. 88: 260–266, doi:10.1016/j.ecss.2010.04.004
VISSER, A. W. 1997. Using random walk models to simulatethe vertical distribution of particles in a turbulent watercolumn. Mar. Ecol. Prog. Ser. 158: 275–281, doi:10.3354/meps158275
WHITE, C., K. A. SELKOE, J. WATSON, D. A. SIEGEL, D. C.ZACHERL, AND R. J. TOONEN. 2010. Ocean currents helpexplain population genetic structure. Proc. R. Soc. B Biol. Sci.277: 1685–1694, doi:10.1098/rspb.2009.2214
XING, J., AND A. M. DAVIES. 2001. A three-dimensional baroclinicmodel of the Irish Sea: Information of the thermal fronts andassociated circulation. J. Phys. Oceanogr. 31: 94–114,doi:10.1175/1520-0485(2001)031,0094:ATDBMO.2.0.CO;2
YANNICELLI, B., L. R. CASTRO, A. VALLE-LEVINSON, L. ATKINSON,AND D. FIGUEROA. 2006. Vertical distribution of decapodlarvae in the entrance of an equatorward facing bay of centralChile: Implications for transport. J. Plankton Res. 28: 19–37,doi:10.1093/plankt/fbi098
YOUNG, E. F., AND J. T. HOLT. 2007. Prediction and analysis oflong-term variability of temperature and salinity in the IrishSea. J. Geophys. Res. 112: C01008, doi:10.1029/2005JC003386
Associate editor: James J. Leichter
Received: 14 May 2012Accepted: 18 October 2012Amended: 23 October 2012
524 Robins et al.
Copyright © 2022 FDOKUMEN




















