PHYLOGENETIC POSITION OF ZYGOGONIUM ERICETORUM (ZYGNEMATOPHYCEAE, CHAROPHYTA) FROM A HIGH ALPINE...
14
PHYLOGENETIC POSITION OF ZYGOGONIUM ERICETORUM (ZYGNEMATOPHYCEAE, CHAROPHYTA) FROM A HIGH ALPINE HABITAT AND ULTRASTRUCTURAL CHARACTERIZATION OF UNUSUAL APLANOSPORES 1 Rosalina Stancheva Department of Biological Sciences, California State University San Marcos, San Marcos, California 92096, USA John D. Hall Department of Plant Science and Landscape Architecture, University of Maryland, College Park, Maryland 20742, USA Department of Botany, Academy of Natural Sciences, Philadelphia, Pennsylvania 19103, USA Klaus Herburger Institute of Botany, University of Innsbruck, Sternwartestraße 15, Innsbruck A-6020, Austria Louise A. Lewis Department of Ecology and Evolutionary Biology, University of Connecticut, Storrs, Connecticut 06269, USA Richard M. McCourt Department of Botany, Academy of Natural Sciences, Philadelphia, Pennsylvania 19103, USA Department of Biodiversity, Earth, and Environmental Sciences, Drexel University, Philadelphia, Pennsylvania 19104, USA Robert G. Sheath Department of Biological Sciences, California State University San Marcos, San Marcos, California 92096, USA and Andreas Holzinger 2 Institute of Botany, University of Innsbruck, Sternwartestraße 15, Innsbruck A-6020, Austria Zygogonium ericetorum, the type species of the genus, was studied from a natural population collected in Mt. Sch€ onwieskopf, Tyrol, Austria. Generic concepts of Zygogonium and Zygnema were tested with atpB, psbC, and rbcL gene sequence analysis, which showed a sister relationship between Z. ericetorum and Mesotaenium, in an early branching clade sister to a grouping of Zygnema and several other filamentous and unicellular zygnematalean taxa. A variety of light, confocal, transmission electron microscopy, and cytochemical techniques provided new data on the variable chloroplast shape of Z. ericetorum, and its aplanospore structure and development, which has been previously considered taxonomically important but has been ambiguously interpreted. Zygogonium can be distinguished from other zygnematophytes (particularly Zygnema), based on the combination of two characters: (i) irregular, compressed plate-like chloroplasts and (ii) residual cytoplasmic content left in sporangia outside of the fully developed aplanospores or zygospores. The presence of a sporangial wall that separates the spores from the parent cell should be excluded from the definition of Zygogonium, because it is also observed in Zygnema. Similarly, the ecological characterization of Zygogonium as acidophilic is not unique to the genus. The names of 18 species currently belonging to Zygogonium are here changed to Zygnema, because of incompatibility with this new proposed Zygogonium concept. In the species transferred to Zygnema, chloroplasts are typically stellate in three-dimensions, and the entire content of fertile cells is transformed into the spore, so there is no cytoplasmic residue. Key index words: alpine habitat; generic and subgeneric concept; green algae; morphology; phylogeny; repro- duction; Zygnema; Zygnematophyceae; Zygogonium Abbreviations : atpB, ATP-synthetase subunit B; BBM, Bold’s Basal Medium; BI, Bayesian inference; CLSM, confocal laser scanning microscopy; ML, maximum likelihood; MP, maximum parsimony; psbC, photosys- tem II CP43 gene; rbcL, RUBISCO large subunit; TEM, transmission electron microscopy The enigmatic green algal genus Zygogonium K€ utzing and its type species, Zygogonium ericetorum, have confounded phycologists for decades with their 1 Received 5 May 2014. Accepted 21 July 2014. 2 Author for correspondence: e-mail [email protected]. Editorial Responsibility: L. Graham (Associate Editor) J. Phycol. 50, 790–803 (2014) © 2014 The Authors Journal of Phycology published by Wiley Periodicals, Inc. on behalf of Phycological Society of America. This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited. DOI: 10.1111/jpy.12229 790
Transcript of PHYLOGENETIC POSITION OF ZYGOGONIUM ERICETORUM (ZYGNEMATOPHYCEAE, CHAROPHYTA) FROM A HIGH ALPINE...

PHYLOGENETIC POSITION OF ZYGOGONIUM ERICETORUM (ZYGNEMATOPHYCEAE,CHAROPHYTA) FROM A HIGH ALPINE HABITAT AND ULTRASTRUCTURAL
CHARACTERIZATION OF UNUSUAL APLANOSPORES1
Rosalina Stancheva
Department of Biological Sciences, California State University San Marcos, San Marcos, California 92096, USA
John D. Hall
Department of Plant Science and Landscape Architecture, University of Maryland, College Park, Maryland 20742, USA
Department of Botany, Academy of Natural Sciences, Philadelphia, Pennsylvania 19103, USA
Klaus Herburger
Institute of Botany, University of Innsbruck, Sternwartestraße 15, Innsbruck A-6020, Austria
Louise A. Lewis
Department of Ecology and Evolutionary Biology, University of Connecticut, Storrs, Connecticut 06269, USA
Richard M. McCourt
Department of Botany, Academy of Natural Sciences, Philadelphia, Pennsylvania 19103, USA
Department of Biodiversity, Earth, and Environmental Sciences, Drexel University, Philadelphia, Pennsylvania 19104, USA
Robert G. Sheath
Department of Biological Sciences, California State University San Marcos, San Marcos, California 92096, USA
and Andreas Holzinger2
Institute of Botany, University of Innsbruck, Sternwartestraße 15, Innsbruck A-6020, Austria
Zygogonium ericetorum, the type species of the genus,was studied from a natural population collected in Mt.Sch€onwieskopf, Tyrol, Austria. Generic concepts ofZygogonium and Zygnema were tested with atpB, psbC,and rbcL gene sequence analysis, which showed a sisterrelationship between Z. ericetorum and Mesotaenium, inan early branching clade sister to a grouping of Zygnemaand several other filamentous and unicellularzygnematalean taxa. A variety of light, confocal,transmission electron microscopy, and cytochemicaltechniques provided new data on the variablechloroplast shape of Z. ericetorum, and its aplanosporestructure and development, which has been previouslyconsidered taxonomically important but has beenambiguously interpreted. Zygogonium can bedistinguished from other zygnematophytes (particularlyZygnema), based on the combination of two characters:(i) irregular, compressed plate-like chloroplasts and (ii)residual cytoplasmic content left in sporangia outsideof the fully developed aplanospores or zygospores. Thepresence of a sporangial wall that separates the sporesfrom the parent cell should be excluded from the
definition of Zygogonium, because it is also observed inZygnema. Similarly, the ecological characterization ofZygogonium as acidophilic is not unique to the genus.The names of 18 species currently belonging toZygogonium are here changed to Zygnema, because ofincompatibility with this new proposed Zygogoniumconcept. In the species transferred to Zygnema,chloroplasts are typically stellate in three-dimensions,and the entire content of fertile cells is transformedinto the spore, so there is no cytoplasmic residue.
Key index words: alpine habitat; generic and subgenericconcept; green algae; morphology; phylogeny; repro-duction; Zygnema; Zygnematophyceae; Zygogonium
Abbreviations: atpB, ATP-synthetase subunit B; BBM,Bold’s Basal Medium; BI, Bayesian inference; CLSM,confocal laser scanning microscopy; ML, maximumlikelihood; MP, maximum parsimony; psbC, photosys-tem II CP43 gene; rbcL, RUBISCO large subunit;TEM, transmission electron microscopy
The enigmatic green algal genus ZygogoniumK€utzing and its type species, Zygogonium ericetorum,have confounded phycologists for decades with their
1Received 5 May 2014. Accepted 21 July 2014.2Author for correspondence: e-mail [email protected] Responsibility: L. Graham (Associate Editor)
J. Phycol. 50, 790–803 (2014)© 2014 The Authors Journal of Phycology published by Wiley Periodicals, Inc. on behalf of Phycological Society of America.This is an open access article under the terms of the Creative Commons Attribution License,which permits use, distribution and reproduction in any medium, provided the original work is properly cited.DOI: 10.1111/jpy.12229
790

peculiar vegetative and reproductive morphology.Czurda (1932) considered Zygogonium and ZygnemaC. Agardh synonymous because of vegetative similari-ties. Transeau (1933) amended Zygogonium based onthree characteristics: (i) vegetative cells with a pairof rounded, plate-like or irregular pillow-shapedchloroplasts, each with a central pyrenoid; (ii) for-mation of a sporangium wall which divides the gam-etangium before the formation of the spore wall,and (iii) presence of cytoplasmic residue in themature gametangia. Despite the clear delineation ofchloroplast shape by Transeau (1933), this impor-tant taxonomic feature has been subsequentlydescribed for some Zygogonium species in a question-able fashion. For instance, in Zygogonium tunetanumGauthier-Li�evre the chloroplasts have been reportedas irregularly spherical or stellate (Gauthier-Li�evre1965, Wei et al. 1989, Wei and Yung 2000). Someauthors considered both genera Zygnema and Zygogo-nium to have stellate chloroplasts (Kadlubowska1984) and thus to be closely related phylogenetically(Gontcharov et al. 2003). A large number of speciesof Zygogonium with variable vegetative characteristicsand ecological preferences have been reported (e.g.,Randhawa 1959, Kadlubowska 1984).
Stancheva et al. (2012) showed that some Zygnemaspecies also possess a combination of reproductiveand vegetative features noted as characteristic of Zy-gogonium by many authors (e.g., Transeau 1951,Gauthier-Li�evre 1965, Kadlubowska 1984, Rundina1998), such as zygospores formed in the conjuga-tion tubes and separated from the gametangia by acellulosic sporangial wall, predominant asexualreproduction by aplanospores and akinetes, pres-ence of short branches, single-celled or filamentousrhizoidal outgrowths, thickened vegetative cell walls,purple-colored cell content, and transverse walls.These observations confound the existing genericclassification systems of Zygnemataceae, which arebased on the shape of chloroplasts and structure offertile cells (for historical review see Yamagishi1963). Molecular phylogenetic investigations deter-mined that Zygogonium was sister to a clade of Zyg-nema spp. based on the only available strain of Z.tunetanum, in contrast to Zygnemopsis, which was dis-tantly related to Zygnema (Hall et al. 2008, Stanche-va et al. 2012). Stancheva et al. (2012) concludedthat many species of Zygogonium are probably besttreated as Zygnema and described two new specieswith many vegetative and reproductive characteris-tics of Zygogonium, except for the presence of cyto-plasmic residue as Zygnema. However, thephylogenetic position of genus Zygogonium sensustricto and its existence as a taxonomic entity dis-tinct from Zygnema, remained unclear until the typespecies (Z. ericetorum) could be confidently identi-fied and thoroughly studied.
Because of the known difficulties in distinguish-ing between Zygnema and Zygogonium, we will limitfurther discussion of systematics and ecology of Zy-
gogonium to those few well-documented records thatare entirely consistent with that of the type species.Z. ericetorum is a cosmopolitan species and based onthe few confirmed sites, occasionally forms large sin-gle-species communities in acid pools, sphagnumpeat bogs and on wet acid soils (Transeau 1933). Itsability to withstand desiccation in terrestrial habitatshas been known for a long time (Fritsch 1916, Frit-sch and Haines 1923), and this is an ecological abil-ity has been observed in other representatives ofZygnematophyceae (for summary see Holzinger andKarsten 2013, Karsten and Holzinger 2014). How-ever, many records of Z. ericetorum are based on veg-etative filaments and the mostly acidic habitat inwhich the organism was distributed (e.g., Lynn andBrock 1969, Hoppert et al. 2004, Kleeberg et al.2006, Newsome and van Breemen 2012), and nei-ther of these characters is diagnostic (Stanchevaet al. 2012).Holzinger et al. (2010) and Aigner et al. (2013)
recorded Z. ericetorum in high-alpine ephemeralstreamlets with slightly acidic water in the Alps andprovided electron microscopy visualization ofvegetative cells along with ecophysiological data.Zygogonium ericetorum was demonstrated to bedesiccation-tolerant in the vegetative state in fieldsamples, showing intact cell organelles in fully desic-cated samples (Holzinger et al. 2010). Aigner et al.(2013) reported substantially elevated levels of solu-ble compounds, such as phenolics and hydrolysabletannins, in purple-colored filaments of Z. ericetorumisolated mainly from the partially desiccated top lay-ers of the mat, compared to the green filamentsfrom lower layers.Despite the interesting ecophysiological and ultra-
structural features described, no phylogenetic char-acterization of Z. ericetorum has been performed todate, and such an analysis is critical to determinethe placement of filaments attributed to this genus(Stancheva et al. 2012). Therefore, we conductedthis investigation of Z. ericetorum, using fresh livingmaterial from the Alps to (i) evaluate its phyloge-netic placement within Zygnematophyceae and (ii)determine vegetative and reproductive charactersrelevant to generic concepts of Zygogonium and Zyg-nema.
MATERIALS AND METHODS
Algal material. This study was conducted on a natural pop-ulation of Z. ericetorum collected from Mt. Sch€onwieskopf(46°500 998 N, 11°000 903 E), at 2,350 m a.s.l. near Oberg-urgl, Tyrol, Austria as previously reported (Holzinger et al.2010, Aigner et al. 2013). Climate and microclimate data forthe collection site are already available (Karsten et al. 2010).Zygogonium ericetorum samples were collected by hand fromfour different areas in a spring pool on July 18, 2013 andkept at moderate temperature until processed in the labora-tory in Innsbruck for confocal laser scanning microscopy(CLSM) and fixation for transmission electron microscopy(TEM; see below). Zygogonium ericetorum filaments were incu-
PHYLOGENETIC POSITION OF Z. ERICETORUM 791

bated in water from the habitat and placed in plastic petridishes with Bold’s Basal Medium (BBM; Nichols and Bold1965) with 1.5% agarose added (hard BBM).
Two samples were transported to the laboratory at Califor-nia State University San Marcos, California, USA within 5 dof their collection, with light microscopic observations madedirectly on fresh field material and from material grown inculture as described below. Two macroscopically distinguish-able morphs of filaments were collected from mats: purple-tinted filaments from the surface layers, which were exposedto high sun light and desiccation, and green filaments fromthe lower layers, which received less radiation and containedmore moisture. Each filament morph was cultured in bothliquid BBM and 2% agar BBM, latter sprayed every 3 d withdistilled water, both on 12:12 light:dark cycle at 12°C (Ander-sen 2005). The filaments were checked every 3 d up to 35 dand different stages of filament and aplanospore develop-ment under various media conditions were documented bymicrophotographs from living material.
Phylogenetic analysis. Filaments of Z. ericetorum were isolatedusing forceps for DNA extraction. Two extractions were madefrom the field sample described above (strains JH1396 andJH1397). The DNA was extracted using the Nucleon Phyto-pure DNA extraction kit (GE Healthcare, Pittsburgh, PA,USA), as previously described (Hall et al. 2008). Portions ofthe chloroplast genes atpB, psbC, and rbcL were amplifiedusing published primers and protocols (Hall et al. 2008, Stan-cheva et al. 2013, Per�ez et al. 2014). Attempts to amplifyother commonly used loci from the samples were unsuccess-ful (not shown). DNA sequencing and phylogenetic analyseswere performed as previously reported (Stancheva et al.2013). Briefly, maximum parsimony (MP) analyses were per-formed in PAUP* (Swofford 2003) with 10 random additionalsequences. Bootstrap support was estimated using 500 boot-strap replicates. Maximum likelihood (ML) analyses were per-formed using RAxML (Stamatakis et al. 2005, 2008) via theCipres Science Gateway (Miller et al. 2010). Analyses usingBayesian inference (BI) (Ronquist and Huelsenbeck 2003)were also performed via Cipres (Miller et al. 2010) using twochains run for 10 million generations with the first quarter ofthe trees discarded as burnin. Loci were analyzed individuallyand in combined analyses with all three loci. In addition tothese analyses, partitioned data sets were also analyzed usingMrBayes v. 3.2.2 (Ronquist and Huelsenbeck 2003); eachlocus partitioned by codon position and a combined (3 locus)data set partitioned by locus. Finally, a supplementary analysisof rbcL with three additional sequences distantly relatedto Z. ericetorum (two Mesotaenium strains as well as Fotteapyrenoidosa Broady) was performed following the methodsdescribed above and presented in Figure S1 in the Support-ing Information.
Light and confocal microscopy. An Olympus BX41 lightmicroscope and Olympus IX50 inverted fluorescence micro-scope with an attached Olympus MicroFire S99809 digitalcamera (Olympus Imaging America Inc., Center Valley, PA,USA) were used for specimen observation and microphotog-raphy. The size data given in the descriptions are based on aminimum of 20 specimens measured and analyzed using Rin-con image analysis software (Imaging Planet, Goleta, CA,USA). The chemical composition of the cell wall was exam-ined using cytochemical techniques for the localization ofpectic substances (Ruthenium Red, Jensen 1962) and cellu-lose (Calcofluor White [CFW], Krishnamurthy 1999), usingchemicals from Fisher Scientific, Pittsburg, PA, USA. CFWselectively binds to cellulose and chitin. The dye fluoresceswhen exposed to UV light (340–380 nm) and offers a verysensitive method for direct microscopic examination of cellwall content and structure. For chlorophyll autofluorescence,
filaments were excited with FITC filter (excitation: 467-498nm, emission: 513–556 nm, Olympus Scientific SolutionsAmericas in Waltham, MA, USA).
CLSM was performed with a Zeiss Pascal 5 on a Zeiss Axi-overt 200 M microscope (Carl Zeiss, Jena, Germany),equipped with a 6391.4 NA objective lens. An argon laserwas used at 488 nm in single track mode (21% laser power,main dichroic beam splitter a HFT 488, 93 lm pinhole), forexcitation of the chloroplast autofluorescence and the emit-ted signal was collected with a 560-nm long pass filter andfalse colored red. In parallel, a bright field image was col-lected at channel D with a transmission photomultiplier tube.The two images were either merged or only the chloroplastautofluorescence is shown here.
Transmission electron microscopy. TEM was performed withfield-collected samples according to the methods describedby Holzinger et al. (2010). Briefly, field samples were fixedfor 1.5 h in 10 mM sodium cacodylate buffer, pH 6.8, con-taining 1.25% glutaraldehyde. After several rinsing steps, sam-ples were postfixed over night in 1% OsO4 at 4°C,dehydrated in increasing ethanol concentrations and trans-ferred to modified Spurr’s resin (Ellis 2006) via propyleneoxide (2:1 propyleneoxide:Suprr’s resin, 1:2 propylenoxide:Spurr’s resin), for 2.5 h each, continuing by an overnightinfiltration step in pure Spurr’s resin. Samples were then heatpolymerized at 80°C for 8 h, sectioned on a Leica ultracut,counterstained with uranyl acetate and Reynold’s lead citrate.Sections were examined by a Zeiss LIBRA 120 TEM at 80 kV.A Proscan 2 k SSCCD camera (Proscan Electronic Systems,Lagerlechfeld, Germany) was used for image generation.Images were processed with Adobe Photoshop 7.0 software(Adobe Systems Inc., San Jos�e, CA, USA).
RESULTS
Zygogonium ericetorum phylogeny. Using PCR, wewere only able to obtain one sequence of atpB fromextraction JH1396 (Table S1 in the SupportingInformation). We obtained identical psbC sequencesfrom two different extractions. We also obtainedidentical sequences of rbcL from two differentextractions, although one was only a partialsequence. These two sequences were highly similarto an rbcL sequence obtained by coauthor Lewisfrom a previous, unrelated extraction (data notshown). Data sets for these three chloroplast locicontaining a large number of conjugating greenalgae (Zygnematophyceae) and Coleochaete (as anoutgroup) were assembled. The atpB data setincluded 62 taxa and 1,207 nt characters of which544 characters were variable and 504 characterswere parsimony informative. The psbC data setincluded 38 taxa and 1,202 nt of which 518 werevariable and 468 were parsimony informative. TherbcL data set included 62 taxa and 1,335 nt of which567 were variable and 512 were parsimony informa-tive. The combined, 3-locus data set contained 3,744characters of which 1,629 characters were variableand 1,482 characters were parsimony informative.Each locus was aligned and analyzed indepen-
dently. In all analyses, Z. ericetorum was sister to Meso-taenium sp. JH0031 (or a clade of Mesotaenium sp.JH0031, Mesotaenium sp. AG2009-1 and F. pyrenoidosa
792 ROSALINA STANCHEVA ET AL.

in the supplementary analysis of rbcL (Fig. S1) withvariable moderate to high levels of statistical supportdepending on the method. No strains of conjugat-ing green algae were closely related to Z. ericetorumcompared to genus-level relationships in otherclades (e.g., within Desmidiales).
In the supplementary analysis of rbcL, the clade ofZ. ericetorum, Mesotaenium sp. JH0031, Mesotaeniumsp. AG2009-1, and F. pyrenoidosa was embedded in aclade containing Mesotaenium cf. chlamydosporum(Fig. S1). This clade received high statistical sup-port, but its placement in the overall phylogeny ofconjugating green algae was not strongly supported.Because of missing data in these Mesotaenium andFottea strains and because no strain was more closelyrelated to Z. ericetorum than Mesotaenium sp. JH0031,further results and discussion will be limited to thecore data sets.
Placement for the Z. ericetorum and Mesotaenium sp.JH0031 clade varied considerably among analyseswith respect to the branching order of other earlydiverging zygnematophytes. Additionally, statisticalsupport for the placement of the early divergingclades was very low in most analyses of most loci (datanot shown). Because there was no statistically strongdisagreement among loci in bootstrap analyses of sin-gle locus data sets, these loci were combined into a3-locus data set that was analyzed as one large parti-tion as well as with three partitions corresponding toeach gene. In the single-partition 3-locus data set, Z.ericetorum was sister to Mesotaenium sp. JH0031 withhigh statistical support (98% bootstrap support inParsimony analysis, 100% bootstrap support in RAx-ML Maximum Likelihood analysis and 1.0 posteriorprobability in BI; Fig. 1). Inclusion of these taxa in alarger clade containing all other Zygnematalesexcept Spirogyra and Netrium digitus (Br�ebisson ex Ral-fs) Itzigsohn and Rothe was strongly supported inlikelihood analyses (100, 1.0), but poorly supported(74) in parsimony analyses (Fig. 1).
Sister relationships between the Z. ericetorum + Mes-otaenium sp. JH0031 clade and other Zygnematalesvaried among analyses. In the MP analysis, the Z. eri-cetorum + Mesotaenium sp. JH0031 clade was observedto be sister to a clade containing Zygnemopsis, Zygnem-a spp. (including the dubious Zygogonium tunetanum),Mesotaenium kramstai Lemmermann, and most Cylin-drocystis spp. In the BI analyses of the combined dataset, the Z. ericetorum + Mesotaenium sp. JH0031 cladewas found in a polytomy with Cylindrocystis brebissonii(Ralfs) DeBary UTEX1259 and a clade containingmost Zygnematales (except Spirogyra and N. digitus)(not shown). ML analysis of the 3-locus data set inRAxML found the Z. ericetorum + Mesotaenium sp.JH0031 clade sister to all other Zygnematales (exclud-ing Spirogyra and N. digitus; Fig. 1). Support for theinclusion of Z. ericetorum was high (100), but theplacement of the Z. ericetorum + Mesotaenium sp.JH0031 clade with respect to other lineages in theZygnematales remained poorly supported (Fig. 1).
The topology of the 3-locus data set partitionedby locus found using BI was the same as the topol-ogy from the RAxML analysis of the combined dataset (data not shown, cf. Fig. 1), although statisticalsupport and branch length differed. Similarly, therewas strong support for the sister relationshipbetween Z. ericetorum and Mesotaenium sp. JH0031(1.0), and strong support for their inclusion in themain clade of Zygnematales (1.0), but weak supportfor relationships among deeper clades of the Zyg-nematales (data not shown). In all analyses of alldata sets, Z. ericetorum was sister with Mesotaenium sp.JH0031 and outside of the main clade of Zygnemaspp. (including Z. tunetanum).Light and confocal microscopy of the vegetative and
reproductive morphology of Z. ericetorum. Filament mor-phology: Filaments were unbranched or occasionallywith short branches, and unicellular or multicellularrhizoidal outgrowths. Vegetative cells were cylindri-cal, 15–31 lm wide, 10–123 lm long. The cell wallwas two-layered with the inner layer cellulosic, andouter layer pectic. Filaments of the green morph,grown in liquid BBM, had short cells, large, deepgreen chloroplasts, thin cell walls and colorless cellcontent (Fig. 2A). When grown on agar, green fila-ments formed thicker, H-shaped pieces in the pecticlayer of the cell wall and often dissociated intoshort, few-celled segments with rounded cell ends;cells were occasionally slightly inflated. Filaments ofthe purple morph, grown on agar, had more elon-gate cells with pinkish colored cell content(Fig. 2E), proportionally smaller light green chlo-roplasts, thicker multilayered cell walls with pecticmaterial deposited between the cellulosic layers, andfrequent H-shaped wall structures. Filaments withpurple cell content, directly observed from the fieldmaterial, formed numerous aplanospores (Figs. 2G;4, A–C; and 5, described below). By contrast, akine-tes were rarely observed. Akinetes were cylindrical-ovoid to oblong, 19–25 lm wide, 28-52 lm long,completely filling the cell, and were covered bythick multilayered colorless wall (Fig. 4D). Zygo-spore formation has not been observed.Chloroplasts: Each cell contained two chloroplasts
with highly variable morphology (Figs. 2; 3; and S2in the Supporting Information). In the shorter,actively dividing cells of green filaments the twochloroplasts almost completely covered the cell cir-cumference (Fig. 2A). Each chloroplast appeared asan irregularly rounded plate with two to five, typi-cally four, irregular peripheral protrusions in oneplane and a massive, swollen, pillow-shaped centralregion holding a large, and distinct pyrenoid(Fig. 2, A, B, E, and F). Both chloroplasts were bentagainst the walls and positioned opposite to eachother with their massive central regions facing thecell walls and protrusions folded toward the middleof the cell. In the longer cells with purple cell con-tents, the chloroplasts occupied only the centralportion of the cell and were obscured by the pres-
PHYLOGENETIC POSITION OF Z. ERICETORUM 793

99/100/1.0
100/100/1.0
98 /100/1.0
8 2 / 9 9/1.0
8 0 / 8 1/0.9967
96/100/1 .0
99 /100/1.06 2 / 9 1/0.9999
9 1 / 8 8/0.9925
- / - / 0 . 9 9 8 3
- / 6 0/0.6548 0 / 8 4/0.7984
99/100/1.0
- / - /1.0
100/100/1.0
- / 5 1/1.0
100/100/1.0
- / 6 0/0.9991
- / - /-
6 1 / 9 6/0.9999
- / 5 3/0.9859
98/100/1.0
99 /100/1.0- / 5 1/-
100/100/1.0
74 / 100/1.0
100/100/1.0
9 9 / 9 8/0.9779
5 9 / 7 3/- - / - / 1 . 0
100/100/1.0
100/100/1.0
100/100/1.0
100/100/1.0
100/100/1.0
98 /100/0.9999
100/100/1.0
8 2 / 8 6/-
6 6 / 5 3/0.631
9 4 / 9 9/0.9921
100/100/1.0
100 /100/1.0
- / - / 1 . 0
100/100/1.0
- / 5 1/ 0.9904
7 4 / 7 9/-
7 2 / 8 2/1.0
92 /100/0.9998
- / - / 0 . 9 9 9 8
100/100/1.0
100/100/1.0
100/100/1.0
- / 7 8/0.9996
97/100/1.0
100/100/1.0
100/100/1.0
100/100/1.0
100/100/1.0
100/100/1.0
0.09 substitutions/site
Actinotaenium curcurbitinum ACOI901
Closterium ehrenbergii var. malinvernianum JH0013
Haplotaenium minutum SVCK302
Teilingia granulata SAG25.88
Penium margaritaceum ACOI330
Hyalotheca dissiliens SAG384-2
Penium cylindrus ACOI780
Bambusina borreri JH0199
Cosmarium botrytis UTEX301
Desmidium aptogonum SVCK108
Euastrum crassum var. michiganense JH0018
Phymatodocis nordstedtiana SAG47.89
Staurastrum punctulatum SAG679-1
Heimansia pusilla SVCK428Spondylosium pulchellum SVCK365
Staurastrum arctiscon JH0014
Micrasterias rotata UTEX1941
Desmidium grevillei SVCK113
Xanthidium antilopaeum var. polymazum JH0054Staurodesmus convergens UTEX2508
Cosmocladium saxonicum ACOI95
Cylindrocystis brebissonii UTEX1259
Closterium libellula JH0021
Zygnemopsis minuta ACOI60
Cylindrocystis sp. JH0038
Spirogyra borgeana RSS002
Zygogonium ericetorum JH1396
Mesotaenium kramstai UTEX1024
Gonatozygon monotaenium SVCK158
Zygema cylindricum SAG698-2
Mesotaenium caldariorum UTEX41
Mesotaenium sp. JH0031
Zygnema sp. JH0044
Spirogyra lutetiana RSS003
Zygnema sp. JH0007
Zygogonium tunetanum UTCC136
Gonatozygon kinahani ACOI350
Cylindrocystis sp. UTEX1925
Closterium acerosum UTEX1075
Netrium digitus UTEX561
Mougeotia sp. UTEX758
Coleochaete nitellarum UTEX1261
Spirogyra majuscula RSS006Spirogyra varians UTEX479
Coleochaete scutata SAG3.9
Coleochaete pulvinata CFD57b6
Spirogyra sp. JH0936
Sirogonium melanosporum ARL700
Spirogyra pratensis UTEX1746
Coleochaete divergens CFD300d1
Spirogyra gracilis UTEX1743
Spirogyra grevilleana UTEX477
Spirogyra tenuissima ACOI1925
Spirogyra juergensii UTEX1742
Spirogyra longata RSS031
Spirogyra croasdalea JH0425
Spirogyra liana UTEX1745
Coleochaete sieminskiana CFD10d1
Spirogyra condensata UTEX1744
Spirogyra maxima UTEX2495
Spirogyra pratensis UTEX928
Spirogyra notabilis UTEX1682
FIG. 1. Phylogeny of Zygnematophyceae based on a multigene RAxML analysis of the chloroplast genes atpB, psbC and rbcL analyzed asa single partition. Zygogonium ericetorum is highlighted in gray. Numbers above the branches represent bootstrap values from maximum par-simony and RAxML analyses and posterior probabilities from Bayesian inference, respectively. Bootstrap values less than 50 and posteriorprobabilities less than 0.5 are indicated with a dash.
794 ROSALINA STANCHEVA ET AL.

ence of lipid droplets and starch grains. The chlo-roplasts were more rounded (Figs. 2, C and D and3, A and B) or taeniform (Fig. 2, E and F) with veryshort or typically lacking peripheral protrusions.Both chloroplasts were slightly diagonally positionedat variable angles to each other, which showed theirwide front face and narrow profile (Figs. 2D and 3,B and D). The chloroplasts were in close contact,touching each other at just one point, as shown byconfocal microscopy (Fig. 3). This linkage keepsboth chloroplasts together, but on an angle, withtheir opposite ends free. The arrangement was alsoobserved in the aplanospores (Fig. 2, G and H).Aplanospores: In purple (surface layer) filaments,
numerous vegetative cells formed aplanospores.Their morphology and developmental stages areillustrated in Figures 4 and 5. Aplanospores were
globose, ovoid or cylindrical-ovoid, 15–23 lm wide,and 13–28 lm long. They occupied only a portionof the vegetative cell lumen near the transverse wall.Aplanospore development started with compactionof the chloroplasts, nucleus, and other cell organ-elles, all of which migrated toward the transversewall in the direction opposite to a large vacuole withpurple content (Fig. 4A). Then the aplanosporeformed its own smooth, colorless cellulosic wall thatseparated the chloroplasts and nucleus (Figs. 4,A and B and 5A) from a purple transparent cyto-plasmic residue consisting of a large vacuole, anddense, cap-like structure near the aplanospore(Figs. 4, A and B and 5A). Small spherical bodies,and occasionally small chloroplast fragments werevisible outside the aplanospore (Figs. 4A and 5A).CFW staining showed a ring-like transverse rupturein the cellulosic inner layer of the vegetative cellwall above the middle area of the aplanospore(Fig. 5B).Aplanospores did not develop sutures as typically
observed in Zygnema, and their chloroplastsremained intact during the resting period, a mini-mum of 30 d. Longitudinal and transverse cell wallsof filaments thickened with the aging of aplanosp-ores, and cytoplasmic residue became more con-densed and purple-brownish (Figs. 4B and 5, A–C).We observed aplanospore germination into the 30thday of inoculation of the field material in liquidBBM (Figs. 4C and 5, C and D). Germination beganwith the enlargement of the aplanospores and com-paction of purple-brownish cytoplasmic residue, fol-lowed by the transverse cleavage of the aplanosporeinto two nearly equal halves (Figs. 4, B and C and5, C and D). CFW staining showed that the aplanos-pore and its daughter cell were enclosed in thicktransverse walls of original vegetative cell, and inthin longitudinal walls formed de novo in the area
FIG. 2. Light microscopic (A, C, E, G) and fluorescence (B, D, F, H) images of chloroplasts in Zygogonium ericetorum: (A, B) short vege-tative cells with large deep green chloroplasts each with three to five protrusions, (C, D) short vegetative cells with light purple cell con-tent and small rounded plate-like chloroplasts, (E, F) long vegetative cells with light purple cell content and longitudinally elongatedplate- or ribbon-like chloroplasts, (G, H) cells with aplanospores each with two chloroplasts. Arrows show pyrenoids. Scale bars: 10 lm.
FIG. 3. Confocal laser scanning microscopic images of chlo-roplasts in Zygogonium ericetorum: (A, B) cells with small roundedplate-like chloroplasts, (C, D) cells with irregular plate-like chlo-roplasts and short protrusions. Note the diagonal position ofchloroplasts in each cell which displaced their wide front faceand narrow profile. Arrows show the contact area between twochloroplast. Scale bars: 10 lm.
PHYLOGENETIC POSITION OF Z. ERICETORUM 795

of the ring-like rupture around the aplanospore(Fig. 5D). In this way, each aplanospore formed ashort filament of 3–4 new green cells within the spo-rangium, separated by highly reduced, dense, dark-brown cytoplasmic residues (Fig. 4C). These shortfew-celled fragments eventually escaped, growinginto new filaments. Rarely did the aplanospore, afterits enlargement and cell division, break down thesporangium wall to form a laterally growing fila-ment.TEM of vegetative cells and aplanospores. Vegetative
cells and aplanospores were investigated by TEM(Fig. 6). Vegetative cells contained large, electrontranslucent vacuoles, that covered the entire surfaceof the cell and only a small central cytoplasmic por-tion was evident (Fig. 6A). The cytoplasmic portion
facing the cell walls was extremely thin (~50 nm),but clearly visible (Fig. 6A). The chloroplasts con-tained a central, pillow-shaped pyrenoid with clearlyvisible starch grains (Figs. 6A and S3, A–B in theSupporting Information). In cross-sections, the flatparts of the chloroplast appeared as thin lamina upto 300–400 nm thick (Figs. 6A and S3A), containingabout 15 parallel thylakoids, each with a diameter of15–16 nm (Fig. S3C). In the vicinity of the pyre-noid, plastoglobules with a diameter of ~140 to220 nm were situated (Fig. S3, A, B, and D).Aplanospores occupied nearly one-third of the
original cell (Fig. 6B) and were covered by asmooth, ~250 nm-thick cell wall (Fig. 6, B and C).Precipitates were visible on the outer surface of theaplanospore wall and in the inner corners of theoriginal cell wall (Fig. 6, B and C). The aplanosp-ores contained the same organelles as the vegetativecells. The vacuoles of the aplanospores appearedelectron translucent (Fig. 6, B and C). The chlorop-lasts had pyrenoids and plastoglobules (Figs. 6C andS3E). In the cellular space outside the aplanosporeswall, a membrane-enclosed vacuole was visible(Fig. 6B). The granular structure of this vacuolehad varying electron densities, while the region
FIG. 4. Light microscopic images of reproductive morphologyin Zygogonium ericetorum: (A) filament in early stages of aplanos-pore development showing: (i) cells with compacted chloroplasts,nucleus and other cell organelles located near transverse wall inopposite direction to a large vacuole with purple content (blackarrows), (ii) completely developed aplanospore (white arrows)separated from a purple-colored vacuole by a colorless wall, (iii)remnant of chloroplast outside aplanospore (black arrowhead),(B) filament in late stages of aplanospore development followingthe resting period, note enlarged aplanospore with cell wall cleav-age (white arrows), and compacted purple-brownish cytoplasmicresidue (black arrow), (C) filament with two neighboring apl-anospores (white arrows) each germinated into two new vegeta-tive cells (black arrowheads), note very compacted brownishresidue (black arrows), (D) akinetes with thick multilayered color-less walls, white arrowhead show H-shaped structures in the pecticlayer of cell wall. Scale bars: 10 lm.
FIG. 5. Light (A, C) and fluorescence (B, D) microscopicimages of aplanospores in Zygogonium ericetorum: (A) filament withcompletely developed aplanospores, note dark-purple vacuolatedcytoplasmic residue with small dense granules (arrows), (B) CFWstaining for cellulose, note the ring-like transverse rupture in thecellulose inner layer of vegetative cell wall above the equatorialregion of the aplanospore (arrows), (C) Filament with two germi-nating aplanospores (white arrows) in different stages of cell divi-sion and resulting daughter cell (arrowhead), note dense browncytoplasmic residue (black arrows), (D) CFW staining for cellu-lose, showing transverse cell wall formation in germinating apl-anospores (arrows) and a daughter cell (arrowhead). FilamentsCFW and 10% KOH treated. Scale bars: 10 lm.
796 ROSALINA STANCHEVA ET AL.
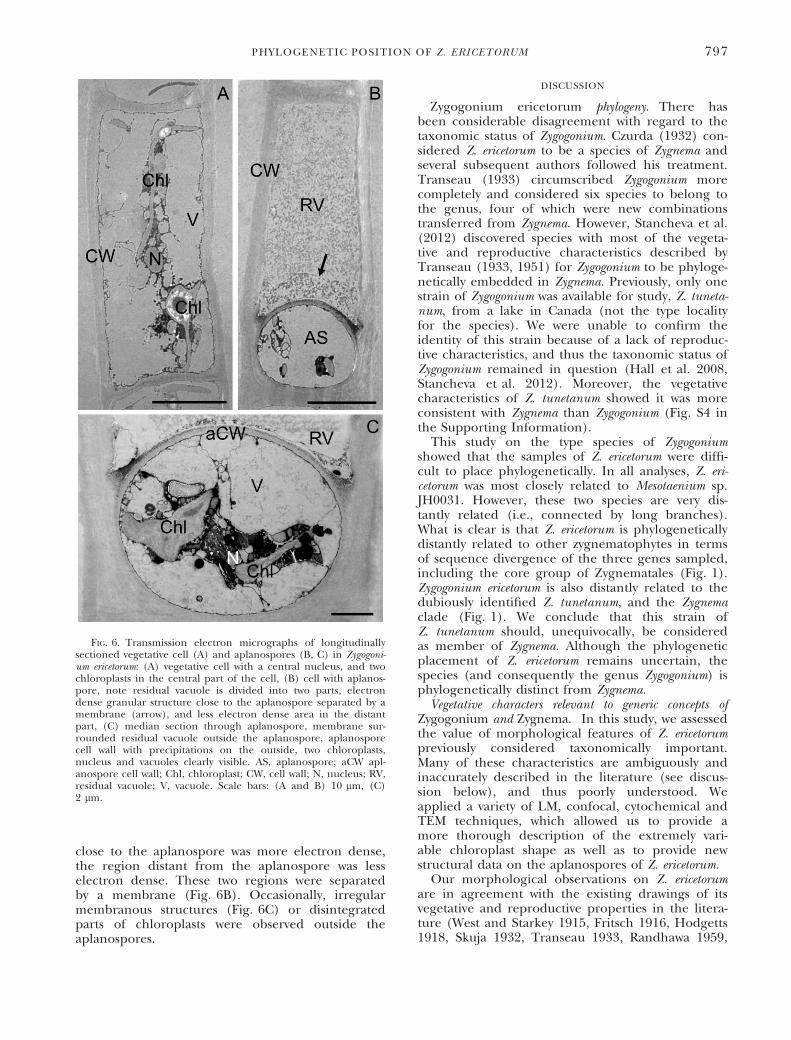
close to the aplanospore was more electron dense,the region distant from the aplanospore was lesselectron dense. These two regions were separatedby a membrane (Fig. 6B). Occasionally, irregularmembranous structures (Fig. 6C) or disintegratedparts of chloroplasts were observed outside theaplanospores.
DISCUSSION
Zygogonium ericetorum phylogeny. There hasbeen considerable disagreement with regard to thetaxonomic status of Zygogonium. Czurda (1932) con-sidered Z. ericetorum to be a species of Zygnema andseveral subsequent authors followed his treatment.Transeau (1933) circumscribed Zygogonium morecompletely and considered six species to belong tothe genus, four of which were new combinationstransferred from Zygnema. However, Stancheva et al.(2012) discovered species with most of the vegeta-tive and reproductive characteristics described byTranseau (1933, 1951) for Zygogonium to be phyloge-netically embedded in Zygnema. Previously, only onestrain of Zygogonium was available for study, Z. tuneta-num, from a lake in Canada (not the type localityfor the species). We were unable to confirm theidentity of this strain because of a lack of reproduc-tive characteristics, and thus the taxonomic status ofZygogonium remained in question (Hall et al. 2008,Stancheva et al. 2012). Moreover, the vegetativecharacteristics of Z. tunetanum showed it was moreconsistent with Zygnema than Zygogonium (Fig. S4 inthe Supporting Information).This study on the type species of Zygogonium
showed that the samples of Z. ericetorum were diffi-cult to place phylogenetically. In all analyses, Z. eri-cetorum was most closely related to Mesotaenium sp.JH0031. However, these two species are very dis-tantly related (i.e., connected by long branches).What is clear is that Z. ericetorum is phylogeneticallydistantly related to other zygnematophytes in termsof sequence divergence of the three genes sampled,including the core group of Zygnematales (Fig. 1).Zygogonium ericetorum is also distantly related to thedubiously identified Z. tunetanum, and the Zygnemaclade (Fig. 1). We conclude that this strain ofZ. tunetanum should, unequivocally, be consideredas member of Zygnema. Although the phylogeneticplacement of Z. ericetorum remains uncertain, thespecies (and consequently the genus Zygogonium) isphylogenetically distinct from Zygnema.Vegetative characters relevant to generic concepts of
Zygogonium and Zygnema. In this study, we assessedthe value of morphological features of Z. ericetorumpreviously considered taxonomically important.Many of these characteristics are ambiguously andinaccurately described in the literature (see discus-sion below), and thus poorly understood. Weapplied a variety of LM, confocal, cytochemical andTEM techniques, which allowed us to provide amore thorough description of the extremely vari-able chloroplast shape as well as to provide newstructural data on the aplanospores of Z. ericetorum.Our morphological observations on Z. ericetorum
are in agreement with the existing drawings of itsvegetative and reproductive properties in the litera-ture (West and Starkey 1915, Fritsch 1916, Hodgetts1918, Skuja 1932, Transeau 1933, Randhawa 1959,
FIG. 6. Transmission electron micrographs of longitudinallysectioned vegetative cell (A) and aplanospores (B, C) in Zygogoni-um ericetorum: (A) vegetative cell with a central nucleus, and twochloroplasts in the central part of the cell, (B) cell with aplanos-pore, note residual vacuole is divided into two parts, electrondense granular structure close to the aplanospore separated by amembrane (arrow), and less electron dense area in the distantpart, (C) median section through aplanospore, membrane sur-rounded residual vacuole outside the aplanospore, aplanosporecell wall with precipitations on the outside, two chloroplasts,nucleus and vacuoles clearly visible. AS, aplanospore; aCW apl-anospore cell wall; Chl, chloroplast; CW, cell wall; N, nucleus; RV,residual vacuole; V, vacuole. Scale bars: (A and B) 10 lm, (C)2 lm.
PHYLOGENETIC POSITION OF Z. ERICETORUM 797

Tiffany and Britton 1971), and with the LM andTEM visualization of sterile filaments (Hoppertet al. 2004, Lewis and Entwisle 2007, Holzingeret al. 2010, Aigner et al. 2013). This species wasoriginally described as Conferva ericetorum by Bory deSaint-Vincent (1797) from the soil of a moor in St.Magne, south-western France, and then illustratedby him as Leda ericetorum (Bory de Saint-Vincent1822a, fig. 8, B–E). Specimens of Z. ericetorum fromAustria studied here corresponded well with originaldescriptions and illustrations by Bory de Saint-Vin-cent (1797, 1822a), which are somewhat vague (seecomments below). Confusion in Z. ericetorumnomenclature is partially due to K€utzing (1843),who proposed a new genus Zygogonium, referencinga specimen in his exsiccatae (K€utzing Alg. Dec. No.51). In the exsiccatae, K€utzing lists the author of C.ericetorum as Roth with no reference to Bory deSaint-Vincent (K€utzing’s Exsiccate viewed at theAcademy of Natural Sciences, Philadelphia). Roth(1800) provided a Latin description of C. erictorum,but when Bory de Saint-Vincent (1822b) transferredC. ericetorum to Leda, he stated that “our C. ericetorumadopted by Roth is probably one of the species.”Because C. ericetorum Roth is a later homonym ofC. ericetorum Bory de Saint-Vincent, Guiry (2013)concluded that C. ericetorum Roth was illegitimateand that K€utzing should be listed as the author ofZ. ericetorum and we follow his suggestion. Regardlessof the nomenclatural status of Z. ericetorum, this islikely the same species as C. ericetorum Bory and C.ericetorum Roth (illegitimate) (Guiry 2013).
Several authors commented on the occurrence inZ. ericetorum of thick, multilayered cell walls with H-shaped structures in response to desiccation (Westand Starkey 1915, Fritsch 1916, Transeau 1933, Hop-pert et al. 2004, Lewis and Entwisle 2007, Holzingeret al. 2010, Aigner et al. 2013). However, similar mul-tilayered cell walls have been observed in some spe-cies of Zygnema (Stancheva et al. 2012) and Mougeotia(Gauthier-Li�evre 1965) distributed in arid conditions.Rundina (1998) attributed the formation of H-shaped structures in the pectic layer of the cell wall insome Zygogonium, Zygnema, Mougeotia, and Spirogyraspecies to their terrestrial habitat and desiccationevents with subsequent rewetting of the filaments,which cause rupture of cell walls. Therefore, the cellwall specifics in Z. ericetorum are of little taxonomicvalue, but most likely indicative for the environmen-tal conditions in the habitat, where desiccation eventsare common (Holzinger et al. 2010).
The chloroplast structure was one of the out-standing cellular characteristics of Z. ericetorum andconsidered to have a high taxonomic value for dis-tinguishing Zygogonium from the other members ofZygnematophyceae. Bory de Saint-Vincent (1822a)on figure 8, B–E provided drawings of three fila-ments with several vegetative cells, each containingchloroplasts with variable shape, ranging from twoprominent rounded plates to elongated narrow
bands, very closely placed and visible as a singlestructure. Some early authors (West and Starkey1915, Fritsch 1916) expressed doubts about thenumber of chloroplasts per cell, reporting a singlechloroplast with one or two pyrenoids in shortercells of Z. ericetorum. However, using a regular LM,both chloroplasts could be hardly visible as two dis-tinct structures connected by an “exceedingly deli-cate bridge,” as observed only in the long cells byFritsch (1916). Transeau (1933) characterized thechloroplasts of Zygogonium as two rounded plate-likeor irregular pillow-shaped, and many phycologistsaccepted this view (Randhawa 1959, Gauthier-Li�evre1965, Tiffany and Britton 1971, Bourrelly 1990, Run-dina 1998, Johnson 2002, Lewis and Entwisle 2007).In our observations, the chloroplasts in Z. ericeto-
rum were determined to be two per cell of an irreg-ular, compressed shape, which was clearly illustratedby fluorescence LM, CLSM and TEM images ofchloroplasts placed at an angle in a cell which dis-placed their wide front face and narrow profile(Figs. 2D; 3, B and D; and 6A and S2). This under-standing corresponds to Bory de Saint-Vincent’s(1822a) illustration of chloroplasts as either largerounded structures, or very narrow bands, clearlyindicating their different front and side view as aplate. Similar paired, disc-shaped, variously orien-tated plastids are characteristic of another genus,Pleurodiscus Lagerheim (Smith 1950, Transeau 1951,Randhawa 1959, Bourrelly 1990), considered closelyrelated to Zygogonium (Skuja 1932, Yamagishi 1963).This finding is in contrast to the radial symmetry
of the chloroplasts in Zygnema, nevertheless someauthors considered Zygnema and Zygogonium havingsimilar stellate chloroplasts (Kadlubowska 1984,Gontcharov et al. 2003). Because of the variablemorphology of the chloroplasts in Z. ericetorum,which might also be the case for other members ofthe genus, many filaments in different life stagesneed to be analyzed to evaluate the correct chloro-plast shape in Zygogonium and to distinguish thegenus from Zygnema using regular LM. Currently,the genus Zygogonium contains over 30 species (Gui-ry and Guiry 2014), many of which have chloroplastscharacteristic of Zygnema (see introduction forexample), and therefore the latter genus needscareful taxonomic revision.Reproductive characters relevant to generic concepts of
Zygogonium and Zygnema. Taxonomically, repro-ductive characteristics are very important for defin-ing groups of filamentous Zygnematophyceae.Unfortunately, we were not able to observe sexualreproduction in Z. erictorum, but according to Tran-seau (1951) sexual and asexual reproductive struc-tural characters (i.e., gametangial and sporangialfeatures, and mesospore color and ornamentation)are consistent within the same species. In the stud-ied material, aplanospore formation was a commonmode of reproduction. However, we did not observea sporangial wall dividing the aplanospore from the
798 ROSALINA STANCHEVA ET AL.

rest of the cell, which is considered by Transeau(1933) as distinguishing characteristic of Zygogonium.On the other hand, the aplanospores and zygosp-ores in Zygnema aplanosporum Stancheva, J. D. Hallet Sheath, illustrated by Stancheva et al. (2012),were separated from the sporangium by a sporangialwall, which is a stable character observed in all pop-ulations collected from a large geographic area (R.Stancheva, personal observations). Therefore, thissporangial wall is not unique to Zygogonium, as asimilar structure has been observed in some speciesof Zygnema (Stancheva et al. 2012) and Mougeotia(Yamagishi 1963). In fact, our observations suggestthat it may not occur in Z. ericetorum.
However, the peculiar development of the apl-anospore may be characteristic of Z. ericetorum. Weobserved that aplanospores germinated after a 30 dresting period into new vegetative cells by direct celldivision. Each aplanospore produced a few new cellswithin the old filament along with almost completeresorption of the purple residue. Our view is incomplete accordance with Fritsch (1916) whodescribed in great detail the formation of daughter-cells by cell-division of the aplanospore where a sep-tum divided the aplanospore into two approximatelyequal halves. Fritsch (1916) illustrated groups offour cells, produced from a single aplanospore(Fig. 2, G and H), similar to our Figure 4C. Westand Starkey (1915) provided an illustration of simi-lar process of aplanospores germination (Fig. 4F),but the authors stated that aplanospores may escapefrom the filament by breaking through the mother-cell wall. The germination of aplanospores hadbeen recorded approximately a month after theirformation (West and Starkey 1915), or immediatelywith a return of moisture (Fritsch 1916). Fritsch(1916) concluded “the response to drought is thusof very simple kind.” Fritsch (1916) used the term“akinete” for the structure we interpret to be an apl-anospore. The aplanospores in Z. ericetorum are verysimple, compared to typical aplanospores in Zygne-matophyceae (for example in Z. aplanosporum, LMand SEM illustrated by Stancheva et al. 2012). Apl-anospores of Z. ericetorum differ from those of mostfilamentous zygnematophytes in that the wall is notmultilayered, but it is colorless, unornamented andlacks an obvious suture. Besides the structural char-acteristics of the aplanospore, the developmentalprocess by which the spores are formed is unique tothis organism. Tiffany and Britton (1971) illustratedthe aplanospore germination process in Z. ericeto-rum, where the new germling filament developedthrough the opening of the aplanospore wall, simi-lar to aplanospore germination in Zygnema, but wecannot confirm that observation.
The presence of cytoplasmic residue left in thegametangia after zygospore (and aplanospore) for-mation is the third diagnostic characteristic ofZygogonium proposed by Transeau (1933). Our datashowed that only a part of protoplast was used in
aplanospore formation in Z. ericetorum, and thus apurple cytoplasmic residue was left in the mothercell (sporangium), outside the aplanospore. Theseobservations of aplanospore structure and associatedpurple residue agreed well with detailed descrip-tions and illustrations by West and Starkey (1915),Fritsch (1916), Transeau (1933), as well as withmore recent authors (Transeau 1951, Randhawa1959, Tiffany and Britton 1971, Rundina 1998,Lewis and Entwisle 2007, Johnson 2002). Our TEMimages showed new details of the aplanospore cellwall, a cap-like structure near the aplanospore andthe ultrastructure of the cytoplasmic residue. Toour knowledge, aplanospores and the cytoplasmicresidue have been studied in Z. ericetorum for thefirst time by TEM. While the aplanospores containall organelles also found in the vegetative cell, thecytoplasmic residue consists of a large vacuole withgranular content of varying electron density. Thegranular material is tightly packed against the newlyformed aplanospore wall. Moreover, the area outsidethe aplanospores included small granules and rarelychloroplast fragments, but no intact organelles.Yamagishi (1963) considered the presence of cyto-
plasmic residue in gametangia (and sporangia) andsporangial wall surrounding the spore as importanttaxonomic features in defining genera of Zygnemat-ophyceae. According to his concept, conjugation inMougeotia C. Agardh and Zygogonium are similar,because the zygospore is surrounded by a sporangialwall separating it from the gametangial cells, andcytoplasmic residue is left in gametangia. However,the structure and composition of cytoplasmic resi-due is also a character that differs among these gen-era. For instance in Mougeotia, MougeotiellaYamagishi and Neozygnema Yamagishi described agranular appearance or irregularly shrunken mem-branous structure (Yamagishi 1963, Rundina 1998),while in Zygnemopsis (Skuja) Transeau and DebaryaWittr. (syn. Transeauina Guiry) the lumen of thegametangia outside the spore is filled completelywith a bluish-white, refractive pectic or cellulosiclayer as the protoplast contracts to the middle ofthe cell (Transeau 1951). Therefore, we confirm thetaxonomic value of the presence and characteristicsof cytoplasmic residue in Zygogonium, which is astrong diagnostic feature by which to distinguishZygogonium from Zygnema.Possible function of purple pigmentation and aplanos-
pore formation. The Zygnematophyceae are knownto synthesize phenolic compounds (e.g., Han et al.2007), which might be adaptive in harsh environ-ments. For example galloyl glucose derivatives weredescribed in Spirogyra varians (Hassall) K€utzing(Nishizawa et al. 1985, Cannell et al. 1988). Morerecently, unusual phenolic compounds have beendescribed in different arctic and Antarctic Zygnemaspp. (Pichrtov�a et al. 2013). Purple to brownish pig-mentation of vacuoles has been reported repeatedlyin Zygnematophyceae living in extreme habitats like
PHYLOGENETIC POSITION OF Z. ERICETORUM 799

bare ice (e.g., in Mesotaenium berggrenii (Wittrock)Lagerheim or Ancylonema nordenski€oldii BerggernRemias et al. 2009, 2012b) and attributed to gallo-ylglucopyranose (Remias et al. 2012a).
Evidence suggests that this pigmented residuemight be adaptive in environments of high light. InZ. ericetorum, the purple pigmentation may have UVshading and antioxidative capacities due to highamounts of soluble phenolic compounds identified(Holzinger et al. 2010, Aigner et al. 2013). Alston(1958) proposed that the purple pigment is an iron-tannin chemical compound. Recently, Newsomeand van Breemen (2012) suggested that it is ahighly branched polymer of glucose, containingester linked polyphenolic moieties such as gallicacid, complexed by ferric iron.
At the present stage, we can only speculate aboutthe importance of aplanospores and cytoplasmic res-idue in vegetative propagation of Z. erictorum.Besides their function as reproductive and restingstructures providing protection during unfavorableenvironmental conditions (Fritsch 1916), they mightserve in biochemical cycles in the cell as well. Thecytoplasmic residue (i.e., the material that isexcluded from the aplanospore) is visibly dark pur-ple or brownish. We hypothesize that this materialis excreted from the newly formed aplanospores forprotective purposes. The above described chemicalcomposition of the purple pigmentation may bebeneficial to the algal cells through antioxidativeproperties; however such pigmented residues canalso be toxic when occurring in higher concentra-tions, in which case they are stored in vacuoles(e.g., Takahama 2004). The newly formed aplanosp-ores of Zygogonium do not have a purple appearanceand the new filaments formed from these aplanosp-ores appear mostly green due to the pigments ofthe larger chloroplast. Under the influence of highUV irradiation occurring under the natural alpineconditions (e.g., Blumthaler 2012), the purple pig-mentation is produced again.Conclusions. Our results require a careful recon-
sideration of the taxonomy and ecology of Zygogoni-um. Our collection of Z. ericetorum was determinedto be distantly related to other zygnematophytes(including Zygnema spp.), although confident phylo-genetic placement was not possible with our data.Structurally, our specimen of Z. ericetorum was largelyconsistent with its original description and theobservations made by subsequent authors (e.g., Frit-sch 1916, Transeau 1933) for that species. However,we did not find a sporangial wall separating the apl-anospore from the sporangium (as previous authorshad suggested) in Zygogonium. In contrast, a sporan-gial wall separated zygospores formed in conjuga-tion tubes, and aplanospores in two Zygnema species(Stancheva et al. 2012). Therefore, we conclude thatthe presence of a sporangial wall should beexcluded from the definition of Zygogonium. In ourobservations, Zygogonium can be distinguished from
other zygnematophytes (particularly Zygnema) onlywhen both vegetative and reproductive material ispresent, based on the combination of two charac-ters: (i) irregular, compressed plate-shaped chlorop-lasts and (ii) residual cytoplasmic content left insporangia outside the aplanospore and zygospore.Z. ericetorum seems to be extraordinarily welladapted to the terrestrial environment as indicatedby its unusual physiology.Taxonomic changes. Based on our observations of
the type species, Z. ericetorum, only a few species areconsistent with its peculiar developmental character-istics. Other taxa currently classified as Zygogonium,having radial, nearly stellate chloroplasts and lack-ing cytoplasmic residue after spore formationshould be transferred to other genera, most likelyZygnema. Many of these species have zygospores withmultilayered colored and ornamented mesospores,and are surrounded by a sporangial wall, similar toother Zygnema species (Stancheva et al. 2012).Below, we summarize the taxonomic changes neces-sary to reflect our current understanding of thegenus Zygogonium. We followed Guiry and Guiry(2014) for the list of Zygogonium species currentlyaccepted taxonomically. The lack of cytoplasmic res-idue in gametangia was confirmed in all 18 speciestransferred to Zygnema, based on the originaldescriptions (see below). When the chloroplastshape was described it was stellate, except forthe irregular sub-globular to globular chloroplastsin Zygogonium sinense C.-C. Jao (Jao 1988) andZ. sphagnophilum Gauthier-Li�evre (Gauthier-Li�evre1965).Zygnema aquaticum (Gauthier-Li�evre) Stancheva, J.
D. Hall, McCourt et Sheath comb. nov.Basionym: Zygogonium aquaticum Gauthier-Li�evre
1965, Beihefte zur Nova Hedwigia 20: p. 87, Pl. 25,figure A a–c.Zygnema cyanosphaeroidicum (O. Bock and W.
Bock)Synonym: Zygogonium cyanosphaeroidicum (O. Bock
and W. Bock) Kadłubowska 1972Zygnema exuvielliforme (C.-C. Jao) Krieger in Kolk-
witz and Krieger 1944Basionym: Zygogonium exuvielliforme C.-C. Jao in
Taylor 1935Zygnema guineense (Gauthier-Li�evre) Stancheva, J.
D. Hall, McCourt et Sheath comb. nov.Basionym: Zygogonium guineense Gauthier-Li�evre
1965, Beihefte zur Nova Hedwigia 20: p. 89, Pl. 26,figure A.Zygnema heydrichii Schmidle 1897Synonym: Zygogonium heydrichii (Schmidle) Tran-
seau 1933Zygnema heydrichii var. indicum Randhawa 1938Synonym: Zygogonium indicum (Randhawa) Tran-
seau 1951Zygnema jaoi Krieger in Kolkwitz and Krieger 1944Synonyms: Zygogonium orientale Y. X. Wei 1979, and
Zygogonium orientale var. reniforme H. J. Hu 1979
800 ROSALINA STANCHEVA ET AL.

(according to Jao 1988), Zygogonium reniforme (H. J.Hu) Kadłubowska 1983. Based on Zygogonium sinenseC.-C. Jao 1935
Zygnema laetevirens Klebs 1886Synonym: Zygogonium laetevirens (Klebs) Migula
1907Zygnema maghrebianum (Gauthier-Li�evre) Stanche-
va, J. D. Hall, McCourt et Sheath comb. nov.Basionym: Zygogonium maghrebianum Gauthier-
Li�evre 1965. Beihefte zur Nova Hedwigia 20: p. 89,Pl. 27, figure A.
Zygnema marocanum (Gauthier-Li�evre) Stancheva,J. D. Hall, McCourt et Sheath comb. nov.
Basionym: Zygogonium marocanum Gauthier-Li�evre1965, Beihefte zur Nova Hedwigia 20: p. 90, Pl. 25,figure D.
Zygnema mayyanadense (Kothari) Stancheva, J. D.Hall, McCourt et Sheath comb. nov.
Basionym: Zygogonium mayyanadense Kothari 1971,Phykos 10, p. 106, figs. 1–5.
Zygnema norvegicum (Kadłubowska et Langangen)Stancheva, J. D. Hall, McCourt et Sheath comb. nov.
Basionym: Zygogonium norvegicum Kadłubowska etLangangen 1998, Nova Hedwigia 66: p. 504, figures1 and 2.
Zygnema plakountiosporum (C.-C. Jao) Krieger inKolkwitz and Krieger 1944
Basionym: Zygogonium plakountiosporum C.-C. Jao inTaylor 1935
Zygnema seuratii Gauthier-Li�evre 1941Synonym: Zygogonium seuratii (Gauthier-Li�evre)
Gauthier-Li�evre 1965Zygnema sphagnophilum (Gauthier-Li�evre) Stanche-
va, J. D. Hall, McCourt et Sheath comb. nov.Basionym: Zygogonium sphagnophilum Gauthier-
Li�evre 1965, Beihefte zur Nova Hedwigia 20: p. 91,Pl. 26, figure C a–f.
Zygnema stephensiae Transeau in Transeau et al.1934
Synonym: Zygogonium stephensiae (Transeau) Tran-seau 1951
Zygnema sudanense (Gauthier-Li�evre) Stancheva,J. D. Hall, McCourt et Sheath comb. nov.
Basionym: Zygogonium sudanense Gauthier-Li�evre1965, Beihefte zur Nova Hedwigia 20: p. 92, Pl. 27,figure D.
Zygnema tunetanum (Gauthier-Li�evre) Stancheva,J. D. Hall, McCourt et Sheath comb. nov.
Basionym: Zygogonium tunetanum Gauthier-Li�evre1965, Beihefte zur Nova Hedwigia 20: p. 92, Pl. 27,figure E.
We thank Stefan Grozev for culturing and maintaining thealgal material, Christina Fuller for the help with staining tech-niques and French translation, and Xizhe Wang for theChinese translation. We gratefully acknowledge Dr. Steve Bar-low at the San Diego State University, Electron MicroscopeFacility, for his assistance in confocal microscope imaging.Moreover, we would like to acknowledge the technical assis-tance of Beatrix Jungwirth, Institute of Botany, Innsbruck inTEM sectioning. Traveling of A. H. was supported by a grant
from the University of Innsbruck, International RelationsOffice. The study was supported by funding from the Califor-nia State Water Resources Control Board Consolidated Grantsand SWAMP Programs to R. G. S. and R. S., by NSF awardsDEB-1036466 to L. L. and DEB-1036478 to R. M. M. K. H.acknowledges a PhD scholarship (Doktoratsstipendium Neu,3. Trache) from the University of Innsbruck, Austria. Thismaterial is based in part work performed while R. M.McCourt worked at the National Science Foundation. Anyopinion, findings, and conclusions or recommendationsexpressed in this material are those of the authors and donot necessarily reflect the views of the National Science Foun-dation. This study was supported by the Austrian ScienceFund (FWF) grant P24242-B16 to A. H.
Aigner, S., Remias, D., Karsten, U. & Holzinger, A. 2013. Unusualphenolic compounds contribute to ecophysiological perfor-mance in the purple-colored green alga Zygogonium ericetorum(Zygnematophyceae, Streptophyta) from a high-alpine habi-tat. J. Phycol. 49:648–60.
Alston, R. E. 1958. An investigation of the purple vacuolar pig-ment of Zygogonium ericetorum and the status of ‘algal antho-cyanins’ and ‘phycoporphyrins’. Am. J. Bot. 45:688–92.
Andersen, R. A. 2005. Algal Culturing Techniques. Elsevier Aca-demic Press, Burlington, Vermont, 596 pp.
Blumthaler, M. 2012. Solar radiation of the high Alps. In L€utz, C.[Ed.] Plants in Alpine Regions. Springer, Wien, New York, pp.11–20.
Bock, O. & Bock, W. 1956. Zygnemales aus Unterfranken. Arch.Hydrobiol. 52:406–48.
Bory de Saint-Vincent, J.B.M. 1797. M�emoire sur les genres Confervaet Byssus, du chevalier O. Linn�e. Louis Carazza, Bordeaux,France, 58 pp.
Bory de Saint-Vincent, J. B. M. 1822a. Dictionaire Classique d’His-toire Naturelle, Vol. VII. Imprimeurs de la Soci�et�e d‘histoireNaturelle, Paris, France, 512 pp.
Bory de Saint-Vincent, J. B. M. 1822b. Dictionaire Classique d’His-toire Naturelle, Vol. I. Imprimeurs de la Soci�et�e d‘histoire Nat-urelle, Paris, France, 604 pp.
Bourrelly, P. 1990. Les Algues d’eau Douce. Initiation �a la Syst�ema-tique. Tome I: Les Algues Vertes. Soci�et�e Nouvelle des�Editions Boub�ee, Paris, France, 572 pp.
Cannell, R. J. P., Farmer, P. W. & John, M. 1988. Purification andcharacterization of pentagalloylglucose, an a-glucosidaseinhibitor/antibiotic from the freshwater green alga Spirogyravarians. Biochem. J. 255:937–41.
Czurda, V. 1932. Zygnemales. Verlag von Gustav Fischer, Jena, Ger-many, 226 pp.
Ellis, E. A. 2006. Solutions to the problem of substitution of ERL4221 for vinyl cyclohexene dioxide in Spurr low viscosityembedding formulations. Microscopy Today 14:32–3.
Fritsch, F. E. 1916. The morphology and ecology of an extremeterrestrial form of Zygnema (Zygogonium) ericetorum (Kuetz.).Hansg. Ann. Bot. 30:135–49.
Fritsch, F. E. & Haines, F. M. 1923. The moisture relations of terres-trial algae. II. The changes during exposure to drought andtreatment with hypertonic solutions. Ann. Bot. 37:683–728.
Gauthier-Li�evre, L. 1941. Algues de eaux continentals africaines:Algues du Sahara septentrional et central. Bull. Soc. Hist. Nat.Afr. Nord. 32:79–152 (in French).
Gauthier-Li�evre, L. 1965. Zygnematacees Afrikaines. Nova Hedwi-gia Beih. 20:1–210 (in French).
Gontcharov, A. A., Marin, B. & Melkonian, M. 2003. Molecularphylogeny of conjugating green algae (Zygnemophyceae,Streptophyta) inferred from SSU rDNA sequence compari-sons. J. Mol. Evol. 56:89–104.
Guiry, M. D. 2013. Taxonomy and nomenclature of the Conjugat-ophyceae (=Zygnematophyceae). Algae 28:1–29.
Guiry, M. D. & Guiry, G. M. 2014. AlgaeBase. World-wide elec-tronic publication. National University of Ireland, Galway,Ireland. http://www.algaebase.org (accessed on 20 April2014).
PHYLOGENETIC POSITION OF Z. ERICETORUM 801

Hall, J. D., Karol, K. G., McCourt, R. M. & Delwiche, C. F. 2008.Phylogeny of the conjugating green algae based on chloro-plast and mitochondrial nucleotide sequence data. J. Phycol.44:467–77.
Han, J. W., Yoon, M., Lee, K. P. & Kim, G. H. 2007. Isolation oftotal RNA from a freshwater green alga Zygnema cruciatum,containing high levels of pigments. Algae 22:125–9.
Hodgetts, W. J. 1918. The conjugation of Zygogonium ericetorumK€utz. New Phytol. 17:238–51.
Holzinger, A. & Karsten, U. 2013. Desiccation stress and tolerancein green algae: consequences for ultrastructure, physiologi-cal, and molecular mechanisms. Front. Plant Sci. 4:327.
Holzinger, A., Tschaikner, A. & Remias, D. 2010. Cytoarchitectureof the desiccation-tolerant green alga Zygogonium ericetorum.Protoplasma 243:15–24.
Hoppert, M., Reimer, R., Kemmling, A., Schr€oder, A., G€unzl, B.& Heinken, T. 2004. Structure and reactivity of a biologicalsoil crust from axeric sandy soil in central Europe. Geomicrobi-ol J. 21:183–91.
Jao, C.C. 1935. Studies on the freshwater Algae of China. I. Zyg-nemataceae from Szechwan. Sinensia 6:551–620.
Jao, C.C., 1988. Flora Algarum Sinicarum Aquae Dulcis. Tomus 1,Zygnemataceae. Science Press, Beijing, China, 228 pp. (inChinese).
Jensen, W. A. 1962. Botanical Histochemistry. W. H. Freeman andCo., San Francisco, California, 408 pp.
Johnson, L. R. 2002. Family Zygnemataceae. In John, D. M., Whit-ton, B. A. & Brook, A. J. [Eds.] The Freshwater Algal Flora ofthe British Isles. An Identification Guide to Freshwater and Terres-trial Algae. Cambridge University Press, Cambridge, UK, pp.480–510.
Kadlubowska, J. Z. 1984. Conjugatophyceae I. Chlorophyta VIII.Zygnemales. In Ettl, H., Gerloff, J., Heynig, H. & Mollenhau-er, D. [Eds.] S€usswasserflora von Mitteleuropa. Band 16. G.Fischer, Stuttgart, Germany, pp. 1–532 (in German).
Kadłubowska, J. Z. 1983. On changes in the nomenclature ofsome taxa of Zygnemaceae. Acta Soc. Bot. Pol. 52:315–20.
Kadłubowska, J. Z. & Langangen, A. 1998. Zygogonium norvegicumsp. nov. (Zygnematales, Chlorophyta) from Norway. NovaHedwigia 66:503–5.
Karsten, U. & Holzinger, A. 2014. Green algae in alpine biologicalsoil crust communities: acclimation strategies against ultravio-let radiation and dehydration. Biodiv. Cons. 23:1845–58.
Karsten, U., L€utz, C. & Holzinger, A. 2010. Ecophysiological per-formance of the aeroterrestrial green alga Klebsormidium cre-nulatum (Charophyceae, Streptophyta) isolated from analpine soil crust with an emphasis on desiccation stress.J. Phycol. 46:1187–97.
Klebs, G. A. 1886. €Uber die Organisation der Gallerte bei einigenAlgen und Flagellaten. Untersuchungen Botanische InstitutT€ubingen 2:333–417.
Kleeberg, A., Schubert, H., Koschorreek, M. & Nixdorf, B. 2006.Abundance and primary production of filamentous greenalgae Zygogonium ericetorum in an extremely acid (pH 2.9)mining lake and its impact on alkalinity generation. Freshw.Biol. 51:925–37.
Kolkwitz, R. & Krieger, H. 1944. Zygnemales. In R. Kolwitz [Ed.]Dr. L. Rabenhorst’s Kryptogamen-Flora von Deutschland und derSchweiz, XIII. Band 2. Akademische Verlagsgesellschaft, Leip-zig, Germany, pp. 1–499.
Kothari, N. M. 1971. Two new members of Zygnemataceae. Phykos10:106–11.
Krishnamurthy, K. V. 1999. Methods in Cell Wall Cytochemistry. CRCPress, Boca Raton, Florida, 318 pp.
K€utzing, F. T. 1843. Phycologia generalis oder Anatomie, Physiologieund Systemkunde der Tange. . . Mit 80 farbig gedruckten Tafeln,gezeichnet und gravirt vom Verfasser. pp. [part 1]: [i]-ii, [1]-142, [part 2:] 143-458, 1, err.], pls 1-80. F.A. Brockhaus,Leipzig, Germany, 668 pp.
Lewis, S. H. & Entwisle, T. J. 2007. Zygnemaceae. In Entwisle, T. J.,Skinner, S., Lewis, S. H. & Foard, H. J. [Eds.] Algae of Austra-lia. Batrachospermales. Thoreales, Oedogoniales and Zygnemaceae.
ABRS, Canberra, Australia, and CSIRO Publishing, Mel-bourne, Australia, pp. 112–56.
Lynn, R. & Brock, T. D. 1969. Notes on the ecology of a speciesof Zygogonium (K€utz) in Yellowstone National Park. J. Phycol.5:181–5.
Migula, W. 1907. Kryptogamen-Flora von Deutschland, Deutsch- €Osterr-eich und der Schweiz. Band II. Algen. 1. Teil. Cyanophyceae, Di-atomaceae, Chlorophyceae. H. Bergm€uhler, Berlin, Germany,914 pp.
Miller, M. A., Pfieffer, W. & Schwartz, T. 2010. Creating the CI-PRES Science Gateway for inference of large phylogenetictrees. Proceedings of the Gateway Computing EnvironmentsWorkshop (GCE), New Orleans, Louisiana.
Newsome, A. G. & van Breemen, R. B. 2012. Characterization ofthe purple vacuolar pigment of Zygogonium ericetorum alga.Planta Med. 78:PJ20. doi:10.1055/s-0032-1321180
Nichols, H. W. & Bold, H. C. 1965. Trichosarcina polymorpha gen.et sp. nov. J. Phycol. 1:34–8.
Nishizawa, M., Yamagishi, T., Nonaka, G., Nishioka, I. & Ragan,M. A. 1985. Gallotannins of the freshwater green alga Spiro-gyra sp. Phytochemistry 24:2411–3.
Per�ez, W., Hall, J. D., McCourt, R. M. & Karol, K. G. (2014) Phy-logeny of North American Tolypella (Charophyceae, Char-ophyta) based on plastid DNA sequences with a descriptionof Tolypella ramosissima sp. nov. J. Phycol. DOI: 10.1111/jpy.12219.
Pichrtov�a, M., Remias, D., Lewis, L. A. & Holzinger, A. 2013.Changes in phenolic compounds and cellular ultrastructureof arctic and Antarctic strains of Zygnema (Zygnematales,Streptophyta) after exposure to experimentally enhancedUV to PAR ratio. Microbial Ecol. 65:68–83.
Randhawa, M. S. 1938. Observations on some Zygnemales fromnorthern India. Part I. Proc. Indian Acad. Sci. B. 8:109–50.
Randhawa, M. S. 1959. Zygnemaceae. Indian Council of Agricul-tural Research, New Delhi, India, 478 pp.
Remias, D., Holzinger, A., Aigner, S. & L€utz, C. 2012b. Ecophysi-ology and ultrastructure of Ancylonema nordenski€oldii (Zyg-nematales, Streptophyta), causing brown ice on glaciers inSvalbard (high arctic). Polar Biol. 35:899–908.
Remias, D., Holzinger, A. & L€utz, C. 2009. Ultrastructure andphysiological characterization of the ice alga Mesotaeniumberggrenii (Zygnemaphyceae, Chlorophyta) from glaciers inthe European Alps. Phycologia 48:302–12.
Remias, D., Schwaiger, S., Aigner, S., Leya, T., Stuppner, H. &L€utz, C. 2012a. Characterization of an UV- and VIS-absorb-ing, purpurogallin-derived secondary pigment new to algaeand highly abundant in Mesotaenium berggrenii (Zygnemato-phyceae, Chlorophyta), an extremophyte living on glaciers.FEMS Microbiol. Ecol. 79:638–48.
Ronquist, F. & Huelsenbeck, J. P. 2003. MrBayes 3: Bayesian phy-logenetic inference under mixed models. Bioinformatics19:1572–4.
Roth, A. W. 1800. Tentamen florae Germanicae continens enumeratio-nem plantarum in Germania sponte nascentium. Tomus III. Parsprior. Bibliopolio Gleditschiano, Lipsiae, Germany, 578 pp.
Rundina, L. A. 1998. The Zygnematales of Russia (Chlorophyta: Zygne-matophyceae, Zygnematales). Nauka, St. Peterburg, Russia, 346pp. (in Russian).
Schmidle, M. W. 1897. Zur Entwicklung einiger Zygnema und Calo-thrix, Flora oder. Allgemeine Botanische Zeitung 84:167–88.
Skuja, H. 1932. Le genre Pleurodiscus doit-il etre maintenu? Rev.Algol. 6:137–46.
Smith, G. M. 1950. The Fresh-Water Algae of the United States, 2ndedn. McGraw-Hill Book Co., New York, 719 pp.
Stamatakis, A., Hoover, P. & Rougemont, J. 2008. A fast boot-strapping algorithm for the RAxML web-servers. Syst. Biol.57:758–71.
Stamatakis, A, Ott, M. & Ludwig, T. 2005. RAxML-OMP: an effi-cient program for phylogenetic inference on SMPs. Proceed-ings of the 8th International Conference on ParallelComputing Technologies (PaCT2005), Lecture Notes inComputer Science, Springer-Verlag.
802 ROSALINA STANCHEVA ET AL.

Stancheva, R., Hall, J. D. & Sheath, R. G. 2012. Systematics of thegenus Zygnema (Zygnematophyceae, Charophyta) from Cali-fornian watersheds. J. Phycol. 48:409–22.
Stancheva, R., Hall, J. D., McCourt, R. M. & Sheath, R. G. 2013.Identity and phylogenetic placement of Spirogyra species(Zygnemaophyceae, Charophyta) from California streamsand elsewhere. J. Phycol. 49:588–607.
Swofford, D. L. 2003. PAUP*: Phylogenetic Analysis Using Parsimony(*and Other Methods). Version 4. Sinauer Associates, Sunder-land, Massachusetts.
Takahama, U. 2004. Oxidation of vacuolar and apoplastic pheno-lic substrates by peroxidase: physiological significance of theoxidation reactions. Phytochem. Rev. 3:207–19.
Taylor, W. R. 1935. Alpine algae from the Santa Marta Moun-tains, Colombia. Am. J. Bot. 22:763–81.
Tiffany, L. H. & Britton, M. E. 1971. The Algae of Illinois. HafnerPublishing Company, New York, 407 pp.
Transeau, E. N. 1933. The genus Zygogonium. Ohio J. Sci. 33:156–62.
Transeau, E. N. 1951. The Zygnemataceae. The Ohio State Univer-sity Press, Columbus, Ohio, 327 pp.
Transeau, E. N., Tiffany, L. H., Taft, C. E. & Li, L. C. 1934. Newspecies of Zygnemataceae. T. Am. Microsc. Soc. 53:208–30.
Wei, Y.X. & Yung, Y.K. 2000. Reproduction of five species of Zyg-nemataceae (Chlorophyta) in Ontario lakes, Canada. Algol.Stud. 98:91–108.
Wei, Y.X., Yung, Y.K., Jackson, M. B. & Sawa, T. 1989. Some Zyg-nemataceae (Chlorophyta) of Ontario, Canada, includingdescription of two new species. Can. J. Bot. 67:3233–47.
West, G. S. & Starkey, G. B. 1915. A contribution to the cytologyand life-history of Zygnema ericetorum (K€utz.) Hansg., withsome remarks on the genus Zygogonium. New Phytol. 14:194–205.
Yamagishi, T. 1963. Classification of the Zygnemataceae. ScienceRep Tokyo Kyoiku Daigaku 11:191–210.
Supporting Information
Additional Supporting Information may befound in the online version of this article at thepublisher’s web site:
Figure S1. Phylogeny of Zygnematophyceaebased on a RAxML analysis of rbcL showing therelationship between Zygogonium ericetorum and abroader sampling of Mesotaenium strains. Supportvalues follow Figure 1.
Figure S2. Confocal laser scanning microscopicimages of chloroplasts in Zygogonium ericetorum,showing their variable morphology: (A–D) sterilevegetative filaments, (E–J) filaments with apl-anospores. Scale bars: 10 lm.
Figure S3. Transmission electron micrographsof Zygogonium ericetorum chloroplast ultrastructurein vegetative cells (A–D) and aplanospore (E).(A) transverse section through chloroplast, pillow-shaped central part with starch grains clearly visi-ble, flat parts emerging to both sides, numerousplastoglobues (arrow) adjacent to the starchgrains, (B) Pyrenoid surrounded by starch grains,plastoglobules (arrow) in the close to starchgrains, (C) fine structure of thylakoids in theedge of a chloroplast wing, (D) surface sectionthrough central part of chloroplast showingstarch grains and numerous plastoglobues, (E)irregular arrangement of thylakoids and plastoglo-bules in aplanospore. PG, plastoglobules; Py, pyre-noid; S, starch; V, vacuole. Scale bars: (A) 2 lm;(B, D–E) 1 lm; (C) 200 nm.
Figure S4. Light and fluorescence microscopicimages of chloroplasts in Zygogonium tunetanumUTCC136. Scale bar: 10 lm.
Table S1. List of strains used in this study aswell as the GenBank number for the sequencedloci. Newly determined sequences are presentedin bold typeface.
PHYLOGENETIC POSITION OF Z. ERICETORUM 803