
Phylogenetic analysis of Alysiella and related genera of Neisseriaceae: Proposal of Alysiella crassa...
10
Introduction The genus Alysiella Langeron 1923 consists of only the type species, A. filiformis isolated from sheep saliva, and its phylogenetic position remains uncertain. The phenotypic characteristics of Alysiella are very similar to those of the genus Simonsiella Simons 1922, which has also gliding motility and filamentous cells. These strictly aerobic, gram-negative bacteria are found in the oral cavities of animals and human beings and are considered to be nonpathogenetic. They are easily recognized by their typical multicellular micro- morphology. The latest edition of Bergey’s Manual of Systematic Bacteriology (Garrity and Holt, 2001) has presented Alysiella as belonging to the family Neisseri- aceae of b -Proteobacteria, but this classification has not been confirmed by the 16S rDNA sequence data. Historically, the systematics of the family Neisseri- aceae by K. Bøvre in 1984 has been problematic and has required frequent readjustment. The genera Acinetobacter, Moraxella and Psychrobacter, and some species of Kingella have been removed from this family using newly possible techniques. Currently this family includes Nesisseria, Eikenella, Kingella, Simon- siella and Alysiella emended by Rossau et al. (1989) on the basis of an rRNA cistron similarities study, which showed that the genus Simonsiella was geno- Phylogenetic analysis of Alysiella and related genera of Neisseriaceae: Proposal of Alysiella crassa comb. nov., Conchiformibium steedae gen. nov., comb. nov., Conchiformibium kuhniae sp. nov. and Bergeriella denitrificans gen. nov., comb. nov. Cheng-Hui Xie* and Akira Yokota Institute of Molecular and Cellular Biosciences, The University of Tokyo, Bunkyo-ku, Tokyo 113–0032, Japan (Received July 22, 2003; Accepted October 12, 2004) A phylogenetic analysis based on 16S rRNA gene sequences reveals that Alysiella filiformis be- longs to the family Neisseriaceae. The genus Simonsiella is phylogenetically separated by the genera Kingella and Neisseria. The species Simonsiella crassa and A. filiformis show a close phylogenetic relationship, with the 16S rDNA sequence similarity and the DNA-DNA hybridiza- tion representing 98.7% and 35%, respectively. Therefore, S. crassa should be transferred from the genus Simonsiella to the genus Alysiella as Alysiella crassa comb. nov. Simonsiella steedae and Simonsiella sp. of cat origin show strong genetic affinities and are distantly related with the type species of Simonsiella, S. mulleri. Thus, a new genus, Conchiformibium is proposed; Conchiformibium steedae comb. nov. and Conchiformibium kuhniae sp. nov. are accommodated in this new genus. On the basis of the phylogenetic, phenotypic and chemotaxonomic distinc- tion from the genus Neisseria, N. denitrificans should be reclassified, for which a new genus and new combination Bergeriella denitrificans are proposed. Key Words——Alysiella crassa comb. nov.; Bergeriella denitrificans gen. nov., comb. nov.; Conchi- formibium kuhniae sp. nov.; Conchiformibium steedae gen. nov., comb. nov.; Neisseriaceae; Simonsiella J. Gen. Appl. Microbiol., 51, 1–10 (2005) Full Paper * Address reprint requests to: Dr. Cheng-Hui Xie, Institute of Molecular and Cellular Biosciences, The University of Tokyo, 1–1–1 Yayoi, Bunkyo-ku, Tokyo 113–0032, Japan. E-mail: [email protected]
Transcript of Phylogenetic analysis of Alysiella and related genera of Neisseriaceae: Proposal of Alysiella crassa...

Introduction
The genus Alysiella Langeron 1923 consists of onlythe type species, A. filiformis isolated from sheepsaliva, and its phylogenetic position remains uncertain.The phenotypic characteristics of Alysiella are verysimilar to those of the genus Simonsiella Simons 1922,which has also gliding motility and filamentous cells.These strictly aerobic, gram-negative bacteria arefound in the oral cavities of animals and human beingsand are considered to be nonpathogenetic. They are
easily recognized by their typical multicellular micro-morphology. The latest edition of Bergey’s Manual ofSystematic Bacteriology (Garrity and Holt, 2001) haspresented Alysiella as belonging to the family Neisseri-aceae of b-Proteobacteria, but this classification hasnot been confirmed by the 16S rDNA sequence data.Historically, the systematics of the family Neisseri-aceae by K. Bøvre in 1984 has been problematic andhas required frequent readjustment. The generaAcinetobacter, Moraxella and Psychrobacter, andsome species of Kingella have been removed from thisfamily using newly possible techniques. Currently thisfamily includes Nesisseria, Eikenella, Kingella, Simon-siella and Alysiella emended by Rossau et al. (1989)on the basis of an rRNA cistron similarities study,which showed that the genus Simonsiella was geno-
Phylogenetic analysis of Alysiella and related genera of Neisseriaceae:Proposal of Alysiella crassa comb. nov., Conchiformibium steedae
gen. nov., comb. nov., Conchiformibium kuhniae sp. nov. and Bergeriella denitrificans gen. nov., comb. nov.
Cheng-Hui Xie* and Akira Yokota
Institute of Molecular and Cellular Biosciences, The University of Tokyo, Bunkyo-ku, Tokyo 113–0032, Japan
(Received July 22, 2003; Accepted October 12, 2004)
A phylogenetic analysis based on 16S rRNA gene sequences reveals that Alysiella filiformis be-longs to the family Neisseriaceae. The genus Simonsiella is phylogenetically separated by thegenera Kingella and Neisseria. The species Simonsiella crassa and A. filiformis show a closephylogenetic relationship, with the 16S rDNA sequence similarity and the DNA-DNA hybridiza-tion representing 98.7% and 35%, respectively. Therefore, S. crassa should be transferred fromthe genus Simonsiella to the genus Alysiella as Alysiella crassa comb. nov. Simonsiella steedaeand Simonsiella sp. of cat origin show strong genetic affinities and are distantly related with thetype species of Simonsiella, S. mulleri. Thus, a new genus, Conchiformibium is proposed;Conchiformibium steedae comb. nov. and Conchiformibium kuhniae sp. nov. are accommodatedin this new genus. On the basis of the phylogenetic, phenotypic and chemotaxonomic distinc-tion from the genus Neisseria, N. denitrificans should be reclassified, for which a new genus andnew combination Bergeriella denitrificans are proposed.
Key Words——Alysiella crassa comb. nov.; Bergeriella denitrificans gen. nov., comb. nov.; Conchi-formibium kuhniae sp. nov.; Conchiformibium steedae gen. nov., comb. nov.; Neisseriaceae; Simonsiella
J. Gen. Appl. Microbiol., 51, 1–10 (2005)
Full Paper
* Address reprint requests to: Dr. Cheng-Hui Xie, Institute ofMolecular and Cellular Biosciences, The University of Tokyo,1–1–1 Yayoi, Bunkyo-ku, Tokyo 113–0032, Japan.
E-mail: [email protected]

typically heterogeneous and that its species were dis-tributed all through the Neisseriaceae cluster. Rossauet al. (1989) also pointed out that all the species ofNeisseria were clustered to a group except the threespecies: i.e., N. canis, N. animalis and N. dentrificans.Therefore, the allocation of these three species in thegenus Neisseria is not certain. These results wereproved by the present studies of 16S rDNA analysis. Inthis paper we present substantial evidence, mainlyfrom the 16S rDNA sequence, fatty acid composition,quinone system and biochemical tests to elucidate thetaxonomic position of the genus Alysiella and are alsoconcerned with the phylogenetic analyses of the re-lated genera of the family Neisseriaceae Prevot 1933,119AL.
Materials and Methods
Bacterial strains and culture media. Strains A. fili-formis IAM 14968T (�ATCC 15532T) and IAM 14895T
(�NCTC 10282T) were used in this study, while NCTC10282T originated from ATCC 15532T. Simonsiellacrassa IAM 14969T (�ATCC 15533T), S. steedae IAM14972T (�ATCC 27409T), Simonsiella sp. IAM 15036(�ATCC 29436), IAM 15037 (�ATCC 29437), IAM15038 (�ATCC 29465) and N. denitrificans IAM14975T (�ATCC 14686T) were also used in this study.Bacterial strains were incubated in ATCC medium 653(Gherna et al., 1992) at 37°C except for N. denitrifi-cans IAM 14975T, which was incubated in ATCCmedium 4 (Gherna et al., 1992).
Electron microscopy. Cells grown on solid mediumwere fixed in a solution of 1% glutaraldehyde in 0.01 M
phosphate buffer (pH 7.2) for 2 h in room temperature.Cells were spread on a slide and air-dried. Water wasremoved gradually by suspending the samples in anacetone series (30%, 50%, 70%, 90%, 95% and100%) and 100% 2-methyl-2-propanol solution. Sam-ples were freeze-dried, spatter-coated with platinumunder a vacuum, and observed with a model HitachiS4500 scanning electron microscope.
Biochemical analysis. API 20NE and 50CHL(BioMérieux, S. A., Marcy-I’Etoile, France) were usedto determine the physiological and biochemical char-acteristics. The API strips were incubated for 2 days at37°C.
Chemotaxonomic investigations. Cellular fatty acidmethyl esters were prepared, separated and identifiedby using the Microbial Identification system as de-
scribed by Xie and Yokota (2003). The respiratoryquinone system was extracted and determined byHPLC (Shimadzu, Kyoto, Japan), and the genomicDNA extraction, PCR-mediated amplification of 16SrDNA, and sequencing of the PCR products were car-ried out as described previously (Xie and Yokota,2003).
Analysis of sequence data and construction of aphylogenetic tree. The DNA sequences of A. fili-formis were compared with the sequences obtainedfrom the DNA database. The sequences were alignedwith the CLUSTAL W software package (Thompson etal., 1994) and evolutionary distances and the Knuc
value (Kimura, 1980) were generated. Alignment gapsand ambiguous bases were not taken into considera-tion and 1,204 bases were compared because ofsome substitutions, insertions and deletions that couldbe attributed to possible sequencing errors in thesepublished sequences (i.e., an error rate of 0.78%)(Harmsen et al., 2001). A phylogenetic tree was con-structed by using the neighbor-joining method (Saitouand Nei, 1987) and the Knuc value. The confidence oftree topology was evaluated by bootstrap resamplinganalysis (Felsenstein, 1985) with 1,000 replicates. Thesimilarity values were calculated using PAUP 4.0b1(Swofford, 1998). For further phylogenetic analysis,the dataset was calculated by maximum likelihoodmethod using PAUP Program, and the bootstrap val-ues were also calculated.
Nucleotide sequence accession numbers. The16S rDNA sequences determined in this study wereregistered in DDBJ as the accession numbers: A. filiformis IAM 14968T (AB087262), IAM 14895T
(AB087263), S. crassa IAM 14969T (AB087264), S.steedae IAM 14972T (AB087261) and N. denitrificansIAM 14975T (AB087265).
Results
Morphological characteristics The scanning electron micrographs of S. crassa IAM
14969T, S. steedae IAM 14972T and A. filiformis IAM14968T are shown in Fig. 1. These microorganismspresent flat multicellular filaments that may reach from2 to 10 mm in width and 50 mm or more in length, andthey possess bilateral symmetry. The cells of S. crassaand A. filiformis are filamentous, paired, smooth andplump, whereas those of S. steedae are concave onone side, and have an appearance similar to those of
2 XIE and YOKOTA Vol. 51
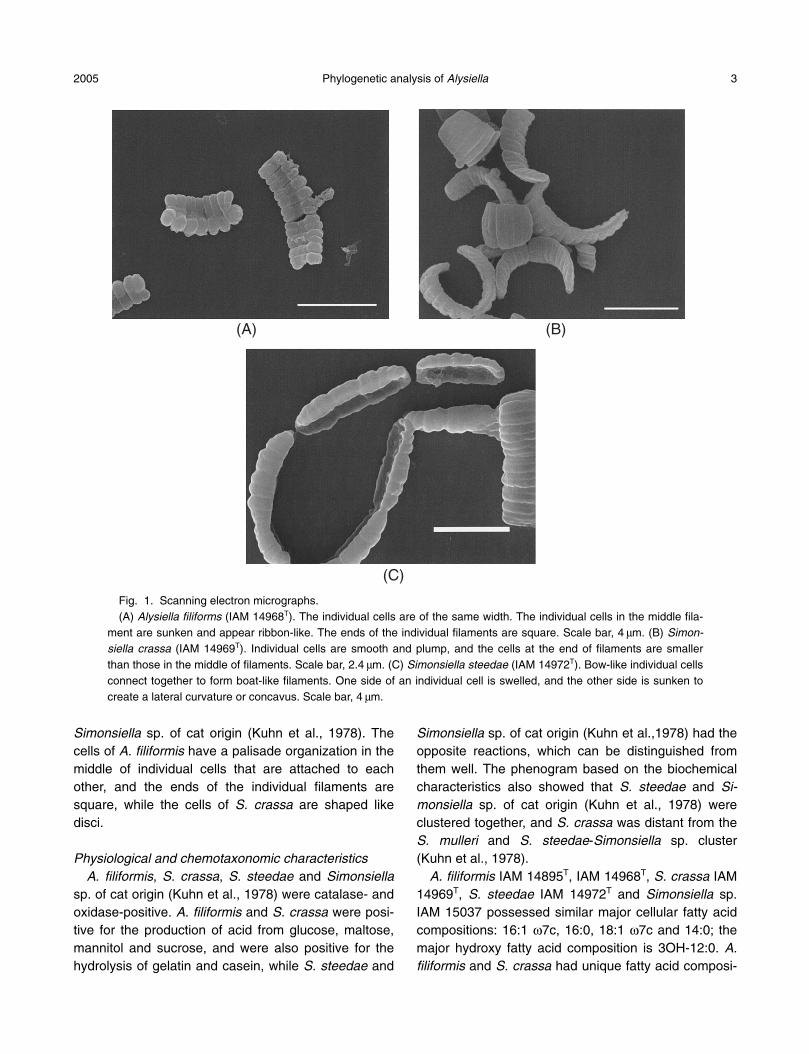
Simonsiella sp. of cat origin (Kuhn et al., 1978). Thecells of A. filiformis have a palisade organization in themiddle of individual cells that are attached to eachother, and the ends of the individual filaments aresquare, while the cells of S. crassa are shaped likedisci.
Physiological and chemotaxonomic characteristicsA. filiformis, S. crassa, S. steedae and Simonsiella
sp. of cat origin (Kuhn et al., 1978) were catalase- andoxidase-positive. A. filiformis and S. crassa were posi-tive for the production of acid from glucose, maltose,mannitol and sucrose, and were also positive for thehydrolysis of gelatin and casein, while S. steedae and
Simonsiella sp. of cat origin (Kuhn et al.,1978) had theopposite reactions, which can be distinguished fromthem well. The phenogram based on the biochemicalcharacteristics also showed that S. steedae and Si-monsiella sp. of cat origin (Kuhn et al., 1978) wereclustered together, and S. crassa was distant from theS. mulleri and S. steedae-Simonsiella sp. cluster(Kuhn et al., 1978).
A. filiformis IAM 14895T, IAM 14968T, S. crassa IAM14969T, S. steedae IAM 14972T and Simonsiella sp.IAM 15037 possessed similar major cellular fatty acidcompositions: 16:1 w7c, 16:0, 18:1 w7c and 14:0; themajor hydroxy fatty acid composition is 3OH-12:0. A.filiformis and S. crassa had unique fatty acid composi-
2005 Phylogenetic analysis of Alysiella 3
(A) (B)
(C)
Fig. 1. Scanning electron micrographs.(A) Alysiella filiforms (IAM 14968T). The individual cells are of the same width. The individual cells in the middle fila-
ment are sunken and appear ribbon-like. The ends of the individual filaments are square. Scale bar, 4 mm. (B) Simon-siella crassa (IAM 14969T). Individual cells are smooth and plump, and the cells at the end of filaments are smallerthan those in the middle of filaments. Scale bar, 2.4 mm. (C) Simonsiella steedae (IAM 14972T). Bow-like individual cellsconnect together to form boat-like filaments. One side of an individual cell is swelled, and the other side is sunken tocreate a lateral curvature or concavus. Scale bar, 4 mm.
tions (ECL 15.487) with more than 1%, while S.steedae and Simonsiella sp. of cat origin (Kuhn et al.,1978) contained none or less than 0.1% (shown inTable 1). All species of Simonsiella and Alysiella hadthe respiratory quinone (ubiquinone-8), by which the b-Proteobacteria is characterized. The G�C content ofthe DNA of A. filiformis IAM 14895T and S. crassa IAM14969T were around 45 mol%, which is 10 mol% lessthan that of S. steedae and Simonsiella sp. of cat ori-gin (Kuhn et al., 1978).
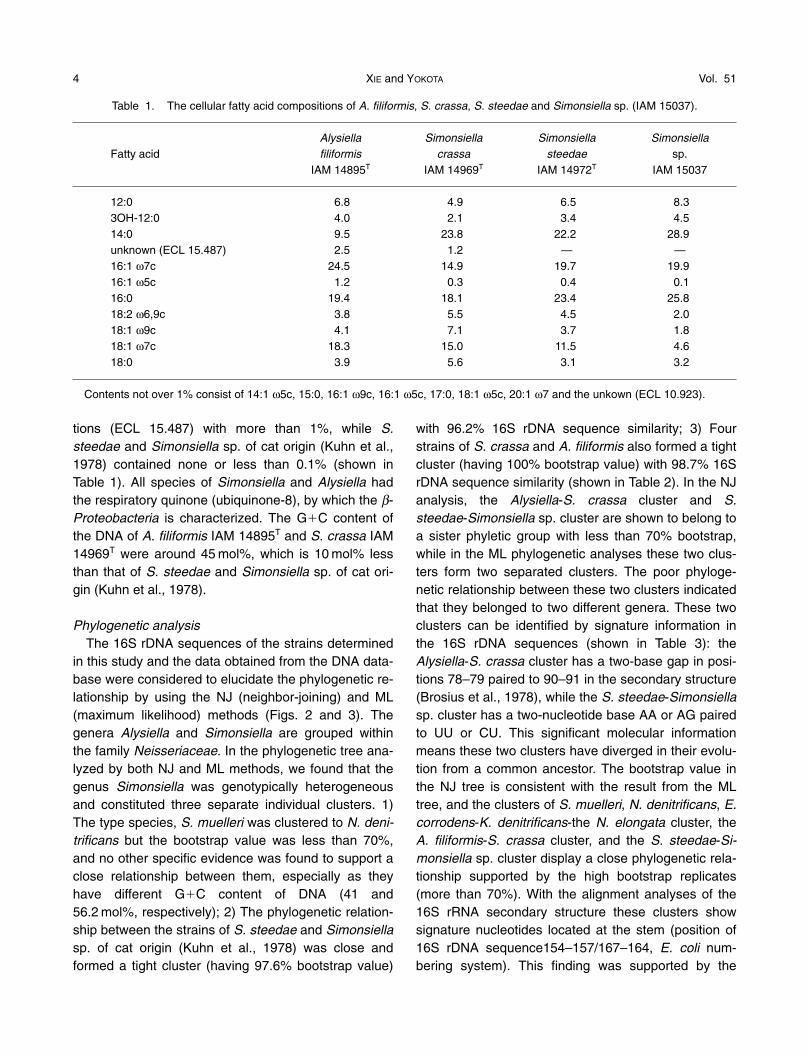
Phylogenetic analysisThe 16S rDNA sequences of the strains determined
in this study and the data obtained from the DNA data-base were considered to elucidate the phylogenetic re-lationship by using the NJ (neighbor-joining) and ML(maximum likelihood) methods (Figs. 2 and 3). Thegenera Alysiella and Simonsiella are grouped withinthe family Neisseriaceae. In the phylogenetic tree ana-lyzed by both NJ and ML methods, we found that thegenus Simonsiella was genotypically heterogeneousand constituted three separate individual clusters. 1)The type species, S. muelleri was clustered to N. deni-trificans but the bootstrap value was less than 70%,and no other specific evidence was found to support aclose relationship between them, especially as theyhave different G�C content of DNA (41 and56.2 mol%, respectively); 2) The phylogenetic relation-ship between the strains of S. steedae and Simonsiellasp. of cat origin (Kuhn et al., 1978) was close andformed a tight cluster (having 97.6% bootstrap value)
with 96.2% 16S rDNA sequence similarity; 3) Fourstrains of S. crassa and A. filiformis also formed a tightcluster (having 100% bootstrap value) with 98.7% 16SrDNA sequence similarity (shown in Table 2). In the NJanalysis, the Alysiella-S. crassa cluster and S.steedae-Simonsiella sp. cluster are shown to belong toa sister phyletic group with less than 70% bootstrap,while in the ML phylogenetic analyses these two clus-ters form two separated clusters. The poor phyloge-netic relationship between these two clusters indicatedthat they belonged to two different genera. These twoclusters can be identified by signature information inthe 16S rDNA sequences (shown in Table 3): theAlysiella-S. crassa cluster has a two-base gap in posi-tions 78–79 paired to 90–91 in the secondary structure(Brosius et al., 1978), while the S. steedae-Simonsiellasp. cluster has a two-nucleotide base AA or AG pairedto UU or CU. This significant molecular informationmeans these two clusters have diverged in their evolu-tion from a common ancestor. The bootstrap value inthe NJ tree is consistent with the result from the MLtree, and the clusters of S. muelleri, N. denitrificans, E.corrodens-K. denitrificans-the N. elongata cluster, theA. filiformis-S. crassa cluster, and the S. steedae-Si-monsiella sp. cluster display a close phylogenetic rela-tionship supported by the high bootstrap replicates(more than 70%). With the alignment analyses of the16S rRNA secondary structure these clusters showsignature nucleotides located at the stem (position of16S rDNA sequence154–157/167–164, E. coli num-bering system). This finding was supported by the
4 XIE and YOKOTA Vol. 51
Table 1. The cellular fatty acid compositions of A. filiformis, S. crassa, S. steedae and Simonsiella sp. (IAM 15037).
Alysiella Simonsiella Simonsiella SimonsiellaFatty acid filiformis crassa steedae sp.
IAM 14895T IAM 14969T IAM 14972T IAM 15037
12:0 6.8 4.9 6.5 8.33OH-12:0 4.0 2.1 3.4 4.514:0 9.5 23.8 22.2 28.9unknown (ECL 15.487) 2.5 1.2 — —16:1 w7c 24.5 14.9 19.7 19.916:1 w5c 1.2 0.3 0.4 0.116:0 19.4 18.1 23.4 25.818:2 w6,9c 3.8 5.5 4.5 2.018:1 w9c 4.1 7.1 3.7 1.818:1 w7c 18.3 15.0 11.5 4.618:0 3.9 5.6 3.1 3.2
Contents not over 1% consist of 14:1 w5c, 15:0, 16:1 w9c, 16:1 w5c, 17:0, 18:1 w5c, 20:1 w7 and the unkown (ECL 10.923).
study of Harmsen et al. 2001, which showed the posi-tions 54 to 510 of 16S rDNA sequence of Neisseri-aceae were a hypervariable rDNA stretch, flanked by astrongly conserved region and can be used for molec-ular species identification.
Discussion
Rossau et al. 1989 determined the DNA-DNA relat-edness among the strains of both genera Alysiella andSimonsiella. As a result, S. crassa IAM 14969T and A.filiformis IAM 14968T possessed 35% relatedness,whereas S. steedae IAM 14972T and Simonsiella sp.
2005 Phylogenetic analysis of Alysiella 5
Fig. 2. Neighbor-joining tree showing phylogenetic relationships among some members of the family of Neisseri-aceae based on 1,204 nucleotide positions of 16S rDNA sequences.
Only bootstrap values over 70% are shown.

ATCC 27381 showed 40% relatedness. Kuhn et al.1977 determined the G+C contents of the DNA of the50 strains of Simonsiella and Alysiella; we found thatthey could be divided into three distinct groups. TheG�C contents of the DNA of S. muelleri strains (ATCC29462, ATCC 29453T, ATCC 29441 and ATCC 29433)were approximately 40 mol%; those of the DNA of A.
filiformis (ATCC 29469T) and S. crassa (ATCC 27504T,ATCC 29446, ATCC 29447 and ATCC 29448) wereapproximately 45 mol%; those of the DNA of S.steedae (ATCC 27409T, ATCC 29457, ATCC 29445,ATCC 29435, ATCC 27398 and ATCC 27410) and Si-monsiella sp. (ATCC 27381T, ATCC 29465, ATCC29437 and ATCC 29436) were around 55 mol%.
6 XIE and YOKOTA Vol. 51
Fig. 3. Maximum-likelihood tree showing phylogenetic relationships among some members of the family of Neis-seriaceae based on 1,204 nucleotide positions of 16S rDNA sequences.
Only bootstrap values over 70% are shown.

Based on the results mentioned above and Rossauet al. 1989, the genus Simonsiella should be dividedinto three different taxonomic groups: i.e., the S. muel-leri cluster, the S. steedae-Simonsiella sp. cluster andthe S. crassa-A. filiformis cluster. A close phylogeneticrelationship, but not more than 97% of the 16S rDNAsequence similarity level, was found between A. fili-formis and S. crassa, which were within the commonindex of 16S rDNA sequence for the species-level dif-ferentiation (Gillis et al., 2001). Therefore, S. crassaand A. filiformis were suggested to be the samegenus, and S. crassa should be transferred to thegenus Alysiella as Alysiella crassa. S. steedae and thestrains of Simonsiella sp. cluster were not included inthe genus Alysiella, and their genetic distance is toofar from the type species of the genus Simonsiella, S.muelleri. Therefore, a new genus Conchiformibium isproposed for them, and S. steedae and Simonsiellasp. of cat origin (Kuhn et al., 1978) should then betransferred to the new genus as Conchiformibiumsteedae comb. nov. and Conchiformibium kuhniae
comb. nov., respectively.The discrepancies in phylogenetic placement of the
genera Neisseria and Kingella display a distinct profilein the analyses of 16S rDNA sequences by NJ and MLmethods. N. elongata, K. denitrificans and E. corro-dens form a cluster with the 16S rDNA sequence simi-larity of more than 97%. The phylogenetic position ofK. denitrificans is distant from that of the other Kingellaspecies with only 95% 16S rDNA sequence similarity.Some biochemical characteristics of K. denitrificansare quite different from those of the genus Kingella butsimilar to those of E. corrodens (Dewhirst et al., 1989).N. elongata displays lower genetic affinities to the clas-sical concept of the Neisseriaceae which was verifiedby the phylogenetic analyses of argF, recA and rho(Smith et al., 1999) and by the biochemical characteri-zation (Bøvre, 1984). The exact taxonomic allocationof N. elongata and K. denitrificans needs to be studiedmore in the future. The phylogenetic position of N.denitrificans is distant from that of the Neisseria clusterwith the type species N. gonorrhoeae and forms an in-
2005 Phylogenetic analysis of Alysiella 7
Table 2. The comparison of 16S rDNA similarity of the strains in the genera Simonsiella, Alysiella, Kingella, Eikenella and Neisseria.
Similarity (%)Bacteria
1 2 3 4 5 6 7 8 9
1 N. elongata ATCC 25295T —2 E. corrodens ATCC 23834T 97.1 —3 K. denitrificans ATCC 33394T 97.2 97.4 —4 S. muelleri ATCC 29453T 95.1 95.9 95.3 —5 N. denitrificans ATCC 14686T 97.0 96.8 97.0 96.6 —6 K. kingae ATCC 23330T 97.2 94.8 95.4 94.8 95.1 —7 S. steedae IAM 14972T 95.3 94.5 94.3 93.2 95.2 95.3 —8 Simonsiella sp. ATCC 27381 94.4 94.5 95.3 93.6 94.9 95.3 96.2 —9 S. crassa IAM 14969T 93.6 93.8 94.9 94.3 94.7 95 94.5 94.4 —
10 A. filiformis IAM 14895T 93.7 94.0 94.6 94.6 94.5 94.8 94.6 94.7 98.7
Table 3. The signature nucleotide positions define for the clusters of Alysiella-S. crassa (I), E. corrodens-K. denitrificans-N. elon-gata (II), N. denitrificans (III), S. muelleri (IV) and S. steedae-Simonsiella sp. (V).
Position of base pair I II III IV V
154-167 U-A C-G U-A U-G U-A155-166 G-U A-U A-U A-U U-A156-165 U-A A-U A-U U-A G-U157-164 C-G U-A U-A C-G C-G
Nucleotide positions are according to the E. coli numbering (Brosius et al., 1978).
8 XIE and YOKOTA Vol. 51
Tabl
e4.
Diff
eren
tial c
hara
cter
istic
s of
the
spec
ies
in th
e ge
nera
Eik
enel
la, A
lysi
ella
, Kin
gella
, Nei
sser
iaan
d C
onch
iform
ibiu
m.
Cha
ract
eris
tics
N. e
long
ataa
K. d
enitr
ifica
nsa
E. c
orro
dens
aN
. den
itrifi
cans
aA
. filif
orm
isb
A. c
rass
aC
. ste
edae
C. k
uhni
aeb
Rod
s�
��
�fil
amen
tfil
amen
tfil
amen
tfil
amen
tS
hort
cha
ins
��
��
��
��
Yello
wis
h pi
gmen
tw
eak
��
��
��
�
Hem
olys
is o
n bl
ood
agar
��
��
��
�
Oxi
dase
��
��
��
��
Cat
alas
e�
��
��
��
�
Aci
d pr
oduc
ed fr
om:
Glu
cose
��
��
��
��
Mal
tose
��
��
��
��
Fru
ctos
e�
��
��
��
�
Suc
rose
��
��
��
��
Man
nose
��
��
��
��
Lact
ose
��
��
��
��
Nitr
ate
redu
ctio
n�
��
��
��
�
Nitr
ite r
educ
tion
��
��
��
��
Gas
from
nitr
ite�
��
��
��
�
Syn
thes
is o
f pol
ysac
char
ide
�N
DN
D�
ND
ND
ND
ND
Trib
utyr
in h
ydro
lysi
s�
ND
ND
�N
DN
DN
DN
DP
rolin
e am
inop
eptid
ase
��
��
ND
ND
ND
ND
Maj
or fa
tty a
cid
16:1
, 16:
016
:0, 1
4:0
18:1
w9c
, 16:
1 w
7c16
:1, 1
6:0
16:1
w7c
, 16:
014
:0, 1
6:0
16:0
, 14:
014
:0, 1
6:1
w9c
Mol
% G
�C
of D
NA
5354
–57
56–5
856
44.3
–45.
443
.9–4
4.9
51–5
453
–55
aD
ata
from
Bø
vre
1984
, bda
ta fr
om K
uhn
et a
l., 1
978.
�, a
ll sp
ecie
s in
the
genu
s ar
e po
sitiv
e; �
/�, s
ome
spec
ies
in th
e ge
nus
are
posi
tive
or n
egat
ive;
�, a
ll sp
ecie
s in
gen
us a
re n
egat
ive;
ND
, no
data
or
not t
este
d.

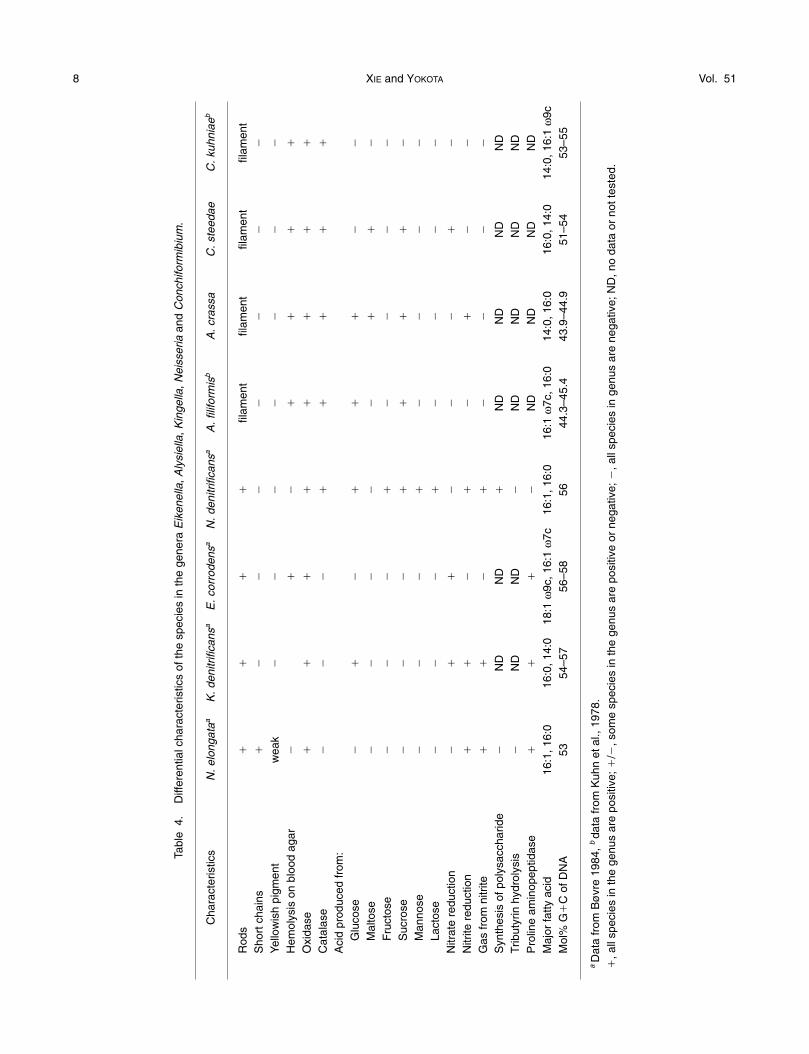
dependent cluster supported by the low bootstrapvalue (less than 70%). The acid production from man-nose, and presence of a large amount of 3OH-14:0 butabsence 3OH-12:0, were significantly different fromother Neisseria species. The G�C content of the DNAis 56.6 mol%, which is 3–7 and 13 mol% higher thanfor other Neisseria species and S. muelleri, respec-tively. N. denitrificans is also characterized by a uniquesignature sequence of 16S rDNA differentiated fromother members of Neisseriaceae. We agree with aview of Rossau et al. (1989) that the taxonomic place-ment of N. denitrificans should be reclassified, forwhich Bergerieella gen. nov. is proposed and referredto as Bergeriella denitrificans sp. nov.
Conclusion
In summary, based on the genotypic and phenotypiccomparisons, S. crassa should be accommodated inthe genus Alysiella. Simonsiella steedae and Simon-siella sp. of cat origin (Kuhn et al., 1978) should be re-classified to a new genus, Conchiformibium. Neisseriadenitrificans was reclassified as Bergeriella denitrifi-cans gen. nov., sp. nov. These results are supportedby the data of rRNA cistron similarities and DNA-DNArelatedness (Rossau et al., 1989), and might be con-firmed by phylogenetic analysis (Dewhirst et al., 1989).
Description of Alysiella crassa comb. nov.Basonym: Simonsiella crassa Schmid in Simons
1922, 509AL.Alysiella crassa (cras’sa. L. fem. adj. crassa thick).Cells are Gram-negative, filamentous, and show
gliding motility. Growth occurs at 27–43°C, at pH 6–8and at 1.5% NaCl. Oxidase and catalase are positive.Acid is produced from glucose, maltose, trehalose, ri-bose, fructose, sucrose and mannitol. Gelatin, caseinand serum are degraded. Reduction of nitrate to nitriteis negative, while reduction of nitrite to nitrogen gas ispositive. The quinone system is ubiquinone-8. Themajor fatty acid compositions are 16:1 w7c, 16:0, 18:1w7c, 14:0. The major hydroxy fatty acid is 3OH-12:0.The G�C content of the DNA is 45–50 mol%. Habitat:Oral cavity of sheep.
The type strain is IAM 14969T (�ATCC 15533T). Ac-cording to Kuhn et al. 1977, the additional strains alsoisolated from the oral cavity of sheep are ATCC 27504,ATCC 29446, ATCC 29447 and ATCC 29448.
Description of Conchiformibium gen. nov.Conchiformibium (Con.chi’for.mi.bium. L. n. conchi-
formis shell-shaped; Gr. n. bium life; Conchiformibiumshell-shaped life).
Cells have lateral curvature of the filaments, andshow gliding motility. Grows at 37°C, but cannot growat 27°C or 43°C. Grow at pH 7.0 but not at pH 6.0. Ox-idase and catalase are positive. Acid is not producedfrom glucose, maltose, trehalose, ribose, fructose, su-crose, mannitol or salicin. Gelatin and casein are notdegraded. Reduction of nitrite to nitrogen gas is nega-tive. Quinone system is ubiquinone-8. The G�C con-tent of the DNA is 50–55 mol%. Habitat: Oral cavity ofanimals (dogs and cats). This genus consists of twospecies, the type species C. steedae and C. kuhniae.
Description of Conchiformibium steedae comb. nov.Basonym: Simonsiella steedae Kuhn and Gregory
1978, 13AL.Conchiformibium steedae (stee’da.e N. L. gen. n.
steedae of Steed; named for P. Steed Glaister, whofirst isolated axenic cultures of Simonsiella anderected the family Simonsiellaceae).
Cells are filamentous, and show gliding motility.Grows at pH 8.0 and at 1.5% NaCl. Reduction of ni-trate to nitrite is positive. Quinone system isubiquinone-8. The major fatty acid compositions are16:0, 14:0, 16:1 w7c, and 18:1 w7c. The major hy-droxy fatty acid is 3OH-12:0. The G�C content of theDNA is 50–55 mol%. Habitat: Oral cavity of dogs.
The type strain of C. steedae is IAM 14972T
(�ATCC 27409T). According to Kuhn et al. 1977, addi-tional strains isolated from oral cavity of dogs areATCC 29457, ATCC 29445 and ATCC 29435.
Description of Conchiformibium kuhniae sp. nov.Conchiformibium kuhniae (kuhni.ae N. L. gen. n.
kuhniae of Kuhn; name for D. A. Kuhn, who first de-scribed these organisms).
Cells are filamentous, and show gliding motility.Grows at pH 8.0 and at 1.5% NaCl. Reduction of ni-trate to nitrite is negative. The major fatty acid compo-sitions are 16:0, 14:0, 16:1 w7c and 18:1 w7c. Themajor hydroxy fatty acid is 3OH-12:0. The G�C con-tent of the DNA is 50–55 mol%. The respiratoryquinone is Q-8. Habitat: Oral cavity of cats.
The type strain of C. kuhniae is IAM 15037T
(�ATCC 29437T). According Kuhn et al. 1977, addi-tional strains isolated from the oral cavity of cat are
2005 Phylogenetic analysis of Alysiella 9

IAM 15038 (�ATCC 29465), IAM 15036 (�ATCC29436) and ATCC 27381.
Description of Bergeriella gen. nov.Bergeriella (Ber.ge.rie.l’la. N. L. fem. n. Berger
named after H. Berger, who first described these or-ganisms, ending-ella; N. L. fem. n. Bergeriella, organ-ism of Berger).
Cells are cocci and aerobic. Catalase is absent. Nopigment is produced. Unlike the other Neisseriaspecies, it can produce acid from mannose, and acidis produced from glucose, fructose and sucrose, butnot from maltose. Nitrate reduction is negative, but ni-trite reduction is positive. Carbonic anhydrase is pro-duced. The major cellular fatty acid composition is16:0, 16:1 w7c and 12:0; the major hydroxy fatty acidcomposition is 3OH-14:0 (Lambert et al., 1971). TheG�C content of the DNA is 55.6 mol%.
The type species of the genus Bergeriella is B. deni-trificans.
Description of Bergeriella denitrificans comb. nov. (ba-sonym “Neisseria denitrificans” Berger 1962, 455AL)
Bergeriella denitrificans (de.ni.tri’fi.cans. L.prep. deaway from; L. nitrum soda; N. L. n. nitrum nitrate; N. L.v. denitrifico to denitrify; N. L. part. adj. denitrificansdenitrifying).
Cells are cocci. Lysine decarboxylase, ornithine de-carboxylase, proline aminopeptidase and alkalinephosphatase were not presented. Resaurin reductionis positive. Its name is based on its ability to reduce ni-trite to nitrogen gas. Habitat: Throats of guinea pigs.
The type strain is ATCC 14686T (�CCUG 2155T
�NCTC 10295T). The G�C content of the type strainis 55.6%.
References
Bøvre, K. (1984) Family VIII. Neisseriaceae Prévot 1933, 119AL.In Bergey’s Manual of Systematic Bacteriology, Vol. 1, ed.by Krieg, N. R. and Holt, J. G., Williams and Wilkins, Balti-more, USA, pp. 288–309.
Brosius, J., Palmer, J. L., Kennedy, J. P., and Noller, H. F.(1978) Complete nucleotide sequence of a 16S ribosomalRNA gene from Escherichia coli. Proc. Natl. Acad. Sci.USA, 75, 4801–4805.
Dewhirst, F., Paster, B., and Bright, P. (1989) Chromobacterium,Eikenella, Kingella, Neisseria, Simonsiella, and Vitreoscillaspecies comprise a major branch of the beta group pro-teobacteria by 16S ribosomal ribonucleic acid sequencecomparison: Transfer of Eikenela and Simonsiella to thefamily Neisseriaceae (emend). Int. J. Syst. Bacteriol., 39,
258–264.Felsenstein, J. (1985) Confidence limits on phylogenies: An ap-
proach using the bootstrap. Evolution, 39, 783–791.Garrity, G. M. and Holt, J. G. (2001) The road map to the Man-
ual. In Bergey’s Manual of Systematic Bacteriology, 2nded., Vol. 1, ed. by Boone, D. R. and Castenholz, R. W.,Springer-Verlag, Heidelberg, New York, Berlin, pp.119–166.
Gherna, R., Pienta P., and Cote, R. (1992). American Type Cul-ture Collection Catalogue of Bacteria and Phages, p. 639,p. 415.
Gillis, M., Vandamme, P., De Vos, O., Swings, J., and Kersters,K. (2001) Polyphasic taxonomy. In Bergey’s Manual of Sys-tematic Bacteriology, 2nd ed., Vol. 1, by Boone, D. R., Cas-tenholz, R. W., and Garrity, G. M., Springer-Verlag, Heidel-berg, New York, Berlin, pp. 43–88.
Harmsen, D., Singer, C., Rothganger, J., Tonjum, T., de Hoog,G. S., Shah, H., Albert, J., and Frosch, M. (2001) Diagnos-tics of Neisseriaceae and Moraxellaceae by ribosomal DNAsequencing: Ribosomal differentiation of medical microor-ganisms. J. Clin. Microbiol., 39, 936–942.
Kimura, M. (1980) A simple method for estimating evolutionaryrates of base substitutions through comparative studies ofnucleotide sequences. J. Mol. Evol., 16, 111–120.
Kuhn, D., Gregory, D., Buchanan, G., and Nydy, M. (1978) Iso-lation, characterization, and numerical taxonomy of Simon-siella strains from the oral cavities of cats, dogs, sheep,and humans. Arch. Microbiol., 118, 235–241.
Kuhn, D., Gregory, D., and Nydy, M. (1977) Deoxyribonucleicacid base composition of Simonsiellaceae. Arch. Microbiol.,113, 205–207.
Lambert, M. A., Hollis, D. G., Moss, C. W., Weaver, R. E., andThomas, M. L. (1971) Cellular fatty acids of non-pathogenicNeisseria. Can. J. Microbiol., 17, 1491–1502.
Rossau, R., Vandenbussche, G., and Thielemans, S. (1989) Ri-bosomal ribonucleic acid cistron similarities and deoxyri-bonucleic acid homologies of Neisseria, Kingella, Eikenella,Simonsiella, Alysiella, and centers for disease controlgroups EF-4 and M-5 in the emend family Neisseriaceae.Int. J. Syst. Bacteriol., 39, 185–198.
Saitou, N. and Nei, M. (1987) The neighbor-joining method: Anew method for reconstructing phylogenetic tree. Mol. Biol.Evol., 4, 406–425.
Smith, N. H., Holmes, E. C., Donovan, G. M., Carpenter, G. A.,and Spratt, B. G. (1999) Networks and groups within thegenus Neisseria: Analysis of argF, recA, rho, and 16SrRNA sequences from human Neisseria species. Mol. Biol.Evol., 16, 773–783.
Swofford, D. L. (1998) PAUP*: Phylogenetic analysis using par-simony (* and other methods), version 4.0. Sinauer Associ-ates, Sunderland, MA.
Thompson, J. D., Higgins, D. G., and Gibson, T. J. (1994)CLUSTAL W: improving the sensitivity of progressive multi-ple sequence alignment through sequence weighting, posi-tion specific gap penalties and weight matrix choice. Nu-cleic Acids Res., 22, 4673–4680.
Xie, C. and Yokota, A. (2003) Phylogenetic analysis of Lampro-pedia hyalina based on the 16S rRNA gene sequence. J.Gen. Appl. Microbiol., 49, 345–349.
10 XIE and YOKOTA Vol. 51




















