
The centric diatom Puncticulata balatonis (Pantocsek) Wojtal et Budzy½ska, comb. nov., in the...
16
509 Nova Hedwigia Vol. 93 issue 3–4, 509–524 Stuttgart, November 2011 Article © 2011 J. Cramer in Gebr. Borntraeger Verlagsbuchhandlung, Stuttgart, Germany www.borntraeger-cramer.de DOI: 10.1127/0029-5035/2011/0093-0509 0029-5035/2011/0093-0509 $ 4.00 The centric diatom Puncticulata balatonis (Pantocsek) Wojtal et Budzy½ska, comb. nov., in the plankton of eutrophic-hypertrophic Rusa»ka Lake (Western Poland) Agnieszka Budzyn ´ ska 1 and Agata Z. Wojtal 2 1 Department of Water Protection, Adam Mickiewicz University, Umultowska 89, 61-614 Poznañ, Poland. e-mail: [email protected] 2 Department of Phycology, W. Szafer Institute of Botany, Polish Academy of Sciences, Lubicz 46, 31-512 Kraków, Poland With 24 figures and 3 tables Budzyńska, A. & A.Z. Wojtal (2011): The centric diatom Puncticulata balatonis (Pantocsek) Wojtal et Budzyńska, comb. nov., in the plankton of eutrophic-hypertrophic Rusałka Lake (Western Poland). – Nova Hedwigia 93: 509–524. Abstract: Cyclotella balatonis Pantocsek is probably a common and widely distributed component of the phytoplankton in mesotrophic to eutrophic European lakes and rivers. However, the distribution and ecology of the species are still unclear, since the species remained hidden in the broad C. radiosa (Grunow) Lemmermann - concept throughout most of the 20 th century. Cyclotella balatonis was found to form quite a large population in the plankton of a small and shallow, urban, eutrophic to hypereutrophic lake (Rusałka Lake, Western Poland). The species occurred in the lake throughout the study period, from February till December 2007, with its highest number in the winter - spring period. The morphology and fine valve structure of the species are described in the paper, documented with light microscope (LM) and scanning electron microscope (SEM) micrographs, and discussed along with the species ecology. Additionally, Cyclotella balatonis is formally transferred into the genus Puncticulata [P. balatonis (Pant.) Wojtal et Budzyńska, comb nov.], and compared with the morphologically similar taxa – P. radiosa (Lemmermann) Håkansson, P. comta (Ehrenberg) Håkansson and P. prae- termissa (Lund) Håkansson. Key words: Bacillariophyta, Puncticulata, ecology, morphology, Poland . Introduction Centric diatoms are an important component of freshwater ecosystems throughout the world. They often reach high numbers in eutrophic lakes and rivers (Genkal & Kiss 1993). As their occurrence is usually related to the trophic status of inhabited water bodies, they are particularly useful bio-indicators in monitoring works and paleolimnological studies (Wunsam et al. 1995, Siver 1999, Wojtal & Kwandrans 2006, Rimet et al. 2009, Houk et al. 2010).
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of The centric diatom Puncticulata balatonis (Pantocsek) Wojtal et Budzy½ska, comb. nov., in the...

509
Nova Hedwigia Vol 93 issue 3ndash4 509ndash524Stuttgart November 2011
Article
copy 2011 J Cramer in Gebr Borntraeger Verlagsbuchhandlung Stuttgart Germany wwwborntraeger-cramerdeDOI 1011270029-503520110093-0509 0029-503520110093-0509 $ 400
The centric diatom Puncticulata balatonis (Pantocsek)Wojtal et Budzyfrac12ska comb nov in the plankton ofeutrophic-hypertrophic Rusaraquoka Lake (Western Poland)
Agnieszka Budzynska1 and Agata Z Wojtal2
1 Department of Water Protection Adam Mickiewicz University Umultowska 89 61-614Poznantilde Poland e-mail abudzamuedupl
2 Department of Phycology W Szafer Institute of Botany Polish Academy of SciencesLubicz 46 31-512 Krakoacutew Poland
With 24 figures and 3 tables
Budzyńska A amp AZ Wojtal (2011) The centric diatom Puncticulata balatonis (Pantocsek) Wojtal etBudzyńska comb nov in the plankton of eutrophic-hypertrophic Rusałka Lake (Western Poland) ndashNova Hedwigia 93 509ndash524
Abstract Cyclotella balatonis Pantocsek is probably a common and widely distributed component ofthe phytoplankton in mesotrophic to eutrophic European lakes and rivers However the distributionand ecology of the species are still unclear since the species remained hidden in the broad C radiosa(Grunow) Lemmermann - concept throughout most of the 20th century Cyclotella balatonis was foundto form quite a large population in the plankton of a small and shallow urban eutrophic to hypereutrophiclake (Rusałka Lake Western Poland) The species occurred in the lake throughout the study periodfrom February till December 2007 with its highest number in the winter - spring period The morphologyand fine valve structure of the species are described in the paper documented with light microscope(LM) and scanning electron microscope (SEM) micrographs and discussed along with the speciesecology Additionally Cyclotella balatonis is formally transferred into the genus Puncticulata[P balatonis (Pant) Wojtal et Budzyńska comb nov] and compared with the morphologicallysimilar taxa ndash P radiosa (Lemmermann) Haringkansson P comta (Ehrenberg) Haringkansson and P prae-termissa (Lund) Haringkansson
Key words Bacillariophyta Puncticulata ecology morphology Poland
Introduction
Centric diatoms are an important component of freshwater ecosystems throughoutthe world They often reach high numbers in eutrophic lakes and rivers (Genkal ampKiss 1993) As their occurrence is usually related to the trophic status of inhabitedwater bodies they are particularly useful bio-indicators in monitoring works andpaleolimnological studies (Wunsam et al 1995 Siver 1999 Wojtal amp Kwandrans2006 Rimet et al 2009 Houk et al 2010)
510
Despite some of the members of Cyclotella genus are known for up to 170 years theecology of many species still remains uncertain Moreover there are also manyuncertainties within the genus concerning taxonomy and nomenclature (Hausmannamp Lotter 2001 Haringkansson 2002 Houk et al 2010) Recently some species of thenot-uniform genus were moved to several newly-established genera (short review inHouk et al 2010) In 2002 the genus Puncticulata was established comprisingspecies characterised by concentrically undulated central part of valve face andnumerous scattered or radially arranged areolae and fultoportulae (or only areolaeor only fultoportulae) with P comta as the type (Haringkansson 2002) The genus hasbeen generally accepted (eg Tanaka 2007 Hayashi 2007 Kobayasi et al 2006Cremer amp Koolmees 2010) though some uncertainties are still discussed (Houk etal 2010)
One of the species sharing the above-mentioned characteristics of the genusPuncticulata (though not transferred to the genus till now) is Cyclotella balatonisand its variety C balatonis var binotata described by Pantocsek (1901 p 104 figs319 331 332) from lake Balaton near Sioacutefok in Hungary Originally the specieswas illustrated by three drawings of valves with scattered (Pantocsek 1901 PlateXV figs 319 332) and radially arranged (Pantocsek 1901 Plate XV fig 331)puncta in the middle part of valve face After Pantocsekrsquos description C balatonisremained practically forgotten and was reported only a few times (eg Handmann1913 Hindaacutek amp Hindaacutekovaacute 2008) It was generally considered a synonym ofCyclotella radiosa (eg Krammer amp Lange-Bertalot 1991 fig 62 11 12) followingthe broad species concept that was in use at that time Thus specimens of a broadmorphological range present in a variety of water bodies were identified as C radi-osa Cyclotella balatonis might have also been misidentified as other species of thecomplex such as P praetermissa (syn Cyclotella praetermissa) or Puncticulatacomta (syn Cyclotella comta) Recently as a result of precise study by Houk et al(2010) using SEM Cyclotella balatonis has been resurrected and the morphologicalranges of similar species were discussed However due to the proceeding long periodof taxonomical confusion the species is almost absent in the literature and needsmuch attention particularly in terms of distribution and autecology
In this paper a population of C balatonis in a small and shallow urban eutrophic tohypereutrophic lake (Rusałka Lake Western Poland) is presented with its quantitativecharacteristics along an annual cycle Morphological features of the diatom aredescribed in detail and documented with LM photographs of live specimens andcleaned frustules as well as with the SEM micrographs Cyclotella balatonis wasalso compared with morphologically similar species namely Puncticulata comtaP praetermissa and P radiosa Due to the great morphological similarity of Cyclotellabalatonis to these and other Puncticulata-species the transfer of Cyclotella balatonisto the genus Puncticulata has been proposed
Study site
Rusałka Lake is situated in the city of Poznań Western Poland (N52deg25E16deg52) It was formed in1943 by damming the Bogdanka River and subsequent flooding of its valley The lake covers the area
511
of 367 ha Although its maximum depth is 90 meters the average does not exceed 2 meters As aresult of the morphometry the lake is mainly polymictic with thermal stratification developing only ina small area around its deepest point in summer (Gołdyn et al 2010)
The lake is strongly eutrophic or hypereutrophic according to different criteria (Table 1) Cyanobacterialblooms occur in summer and autumn (Gołdyn et al 2010) Due to its location and surroundingattractive areas (mainly forests) the lake is intensely used for recreation
Materials and methods
Water samples for the phytoplankton analysis were taken at every meter depth at the deepest part of thelake in 2007 once a month from February till October and once in December Quantitative sampleswere preserved with Lugolrsquos solution and analysed with the use of inverted microscope PZO aftersedimentation in cylindrical chambers according to the Utermoumlhl method (Wetzel amp Likens 1991)
All diatoms from the family Stephanodiscaceae were counted as one taxon The relative frequency ofPuncticulata balatonis and other species belonging to this family was then estimated by counting 100frustules in hydrogene peroxide-treated samples using an Olympus BX-60 microscope Measurementsand the micrographs of live diatoms were done with the use of the same LM and AnalySIS programme
A qualitative analysis of the diatom material was performed after removing carbonates (addition of10 HCl to the samples followed by boiling taking place after 24 h for 15 minutes) and organic matter(boiling in 30 H
2O
2 with small amounts of KClO
3) After air-drying the material was mounted in
Naphraxreg Observations of the diatoms were performed with a Nikon Eclipse 80i light microscopeequipped with Differential Interference Contrast The micrographs were taken with a Nikon DS-Fi 1camera SEM analyses were performed on the material coated with gold using a Hitachi S-4700scanning electron microscope SEM micrographs were taken in the Laboratory of Electron and ConfocalMicroscopy at the Faculty of Biology Adam Mickiewicz University in Poznań and in the Laboratoryof Field Emission Scanning Electron Microscopy and Microanalysis at the Institute of GeologicalSciences of the Jagiellonian University in Krakoacutew Density of valve face structures (eg costae) in10 microm in a line tangential to the radius was calculated based on the formula in Genkal (1977)Additionally data on the morphology of Cyclotella balatonis were obtained from the type material(Balaton Lake Siofok samples 156 157) hosted in the Natural History Museum in Budapest (Hungary)
Table 1 Estimation of the trophic state of Rusałka Lake according to Carlson index (1977) and OECD(1982) criteria (acc to Gołdyn et al 2010)
Carlson indexTSI Values in Boundary values Trophic state
Rusałka lake for eutrophy
TSI (TP) 7437 gt70 HypertrophyTSI (ChL) 7171 53ndash70 HypertrophyTSI (SD) 6737 53ndash70 Eutrophy
OECDCharacteristic Values in Boundary values Trophic state
Rusałka lake for eutrophy
Mean TP (microgl) 96 gt100 EutrophyMean chlorophyll-a (microgl) 347 gt25 HypertrophyMax chlorophyll-a (microgl) 802 gt75 HypertrophyMean Secchi depth (m) 08 lt15 HypertrophyMin Secchi depth (m) 06 lt07 Hypertrophy
512
Water temperature pH and conductivity were measured in situ using YSI 610-DM-meter Watertransparency was expressed as the visibility of the Secchi disc For the statistical graphs Statistica 80software was used
Results
Morphology of Puncticulata balatonis in Rusałka Lake
Puncticulata balatonis (Pantocsek) Wojtal et Budzyńska comb nov Figs 1ndash12 amp 15ndash22
BASIONYM Cyclotella balatonis Pantocsek Die Kieselalgen oder Bacillarien des Balaton p 104 PlateXV figs 319 332 1901
TYPE LOCALITY Balaton Lake near Sioacutefok Hungary
Cells solitary or in short 2ndash7-cell chains (Fig 1) containing multiple discoidchloroplasts The valve diameter ranges between 87 and 210 microm Stria density is15ndash185 in 10 microm with 35ndash47 costae (thickened ribs) in 10 microm Striae are ofunequal length with inserted shorter (Figs 15ndash17) or divided striae (Fig 19) formingan irregularly circular central area (Figs 6 9 12 17) The central area is concentricallyslightly elevated (Figs 15 17) with numerous scattered openings of valve facefultoportulae (externally visible as minor puncta Fig 16) and areolae (externallyvisible as bigger puncta Figs 15ndash17) External openings of the rimopor-tulae arepresent at the end of the shortened striae (Figs 15 16) The internal opening of thesessile rimoportulae has a radially or slightly obliquely positioned slit (Figs 18 20)situated close to internal alveolar openings (towards the valve face margin) Marginalfultoportula openings with two satellite pores are situated (internally) on thethickened costae (shadow lines) separated by 3ndash5 thinner costae Valve facefultoportula openings are internally covered with domed cribra and surrounded bythree satellite pores (Figs 19 20) The morphology of the valve face internal alveolaropenings and the location of internal rimoportula openings correspond well withthe morphology of the specimens from the type material of Cyclotella balatonis(Figs 13 14 amp 21 22 Table 3)
Table 2 Physio-chemical parameters of water of Rusałka Lake in 2007
Temperature pH Coductivity Transparencyndash(degC) (microScm) Secchi depth (m)
20 II 31ndash41 72ndash78 836ndash841 1526 III 49ndash71 79ndash81 866ndash868 1820 IV 134ndash103 79ndash80 no data 1018 V 110ndash156 77ndash82 820ndash840 0819 VI 112ndash232 73ndash84 760ndash878 0918 VII 126ndash240 75ndash83 697ndash754 08520 VIII 145ndash220 75ndash91 627ndash739 0612 IX 144ndash148 83ndash85 646ndash652 0822 X 85ndash89 80ndash83 668ndash680 10503 XII 35ndash38 81ndash82 740ndash744 135
513
Ecological data
The water of Rusałka Lake was alkaline throughout the study period showing highconductivity Water transparency was relatively low ranging from 06 to 18 m ofSecchi depth (Table 2)
Puncticulata balatonis was present in the lake in all the seasons of 2007 It wasfound to form chains only in February March and April The diatom valve sizedistribution pattern varied in subsequent months with the largest average valve sizein February (Fig 23) Its abundance in the lake was also higher in the winter-springperiod (from February till May fig 24) with the maximum value of 35 times 103 cellsml-1 and a biomass of 298 mg l-1 (19 of the total phytoplankton cell number and525 of the total biomass at 6 m depth in March) In other spring samples
Figs 1ndash14 Puncticulata balatonis (Pantocsek) Wojtal et Budzyńska Light microscopy Figs 1ndash12 Specimens from the Rusałka Lake Figs 13ndash14 P balatonis type material sample no 152Fig 1 Short chain of alive cells in girdle view Figs 5ndash6 The same valve at different focus levels OnFigs 6 12ndash13 the characteristic gaps (notae) are marked with an arrow Scale bar represents 10 microm
514
Figs 15ndash22 Puncticulata balatonis (Pantocsek) Wojtal et Budzyńska Scanning electron microscopyFigs 15ndash20 specimens from the Rusałka Lake Figs 21ndash22 P balatonis type material samples no152 amp 156 Fig 15 external view of the rimoportula opening at the end of shortened striae ndash notae
515
P balatonis contributed between 63 and 187 to the total phytoplankton numbersand between 21 and 47 to the total biomass From June until October the numberof the diatom did not exceed 300 frustules per 1 ml (Fig 24) and no valve wasfound in September At the beginning of December P balatonis number increasedslightly again
At the time of the bloom of P balatonis chrysoflagellates (Erkenia subaequiciliataSkuja Chrysococcus triporus Matvienko) pennate diatoms (Fragilaria sp) andorcoenobia-forming Chlorococcales (Didymocystis inconspicua Koršikov Tetastrumsp div Scenedesmus sp div Coelastrum microporum Naumlgeli Crucigenia tetrapediaKirchner) were also abundant Other diatoms such as Aulacoseira ambigua (Grunow)Simonsen Cyclotella meneghiniana Kuumltzing C ocellata Pantocsek Cyclostephanosdelicatus (Genkal) Casper C dubius (Fricke) Round Stephanodiscus parvus Stoermeramp Haringkansson Asterionella formosa Hassal and Navicula sp were also observed
Discussion
Morphology of Puncticulata balatonis (Pant) Wojtal et Budzyńska
Puncticulata balatonis shares important morphological characters assigned to thegenus justifying its formal transfer to this genus a slightly concentrically undulatemiddle part of the valve face with scattered or radially arranged numerous areolaewith internally domed cribra and numerous fultoportulae Additionally valves ofP balatonis have internal alveolar openings with marginal fultoportulae located onthicker costae rimoportulae located externally at the top of shortened striae andinternally in the marginal part of the valve face
The morphology of P balatonis valves from Rusałka Lake corresponds well withthe morphology of the specimens from the Pantocsekrsquos type material (Figs 13 1421 22 Table 3) though the diameter range and the number of striae in 10 microm werefound to be different Pantocsek (1901) stated in his diagnosis (p 104) that thevalves of Cyclotella balatonis are 16ndash20 microm in diameter but we have found bothsmaller and larger specimens (Table 3) The wider diameter range in our observationcould be related to a more complete morphological range of the species since thediatom population in Rusałka Lake was abundant However the time of samplingmay have also been of importance as the medium valve size of the species in our
(arrowed) at concentrically elevated valve face Fig 16 external view of the openings of central (right)and marginal (left) fultoportulae (arrowed) Fig 17 external view of the short and long striae of themarginal area Fig18 internal view of the rimoportula opening (arrowed) located close to the valvemargin and internal alveolar opening and central openings of numerous areolae and fultoportulae(arrowed) Fig 19 internal view of the divided thinner costa (marked) and internal opening of centralfultoportula (arrowed) Fig 20 internal view of the marginal fultoportulae located on the thicker costae(arrowed) Fig 21 external view of the valve face with marked notae Fig 22 internal view of thealveolar openings with thicker costa bearing marginal fultoportulae and internal rimoportula openinglocated near the margin (arrowed) Scale bar represents 5 microm for Figs 15 18 amp 21 4 microm for Fig 163 microm for Figs 19 amp 20 and 2 microm for Figs 17 amp 22
516
Tab
le 3
Com
pari
son
of P
unct
icul
ata
bala
toni
s an
d m
orph
olog
ical
ly s
imila
r sp
ecie
s C
F ndash
cent
ral
fulto
port
ulae
MF
ndash m
argi
nal
fulto
port
ulae
o
urob
serv
atio
n ty
pe m
ater
ial S
iofoacute
k s
ampl
e no
152
our
obse
rvat
ion
base
d on
the
fig
62
11
12
in K
ram
mer
amp L
ange
-Ber
talo
t 199
1 w
hich
sho
ws
spec
imen
s fr
om th
e ty
pe m
ater
ial o
f Cyc
lote
lla
bala
toni
s Pa
ntoc
sek
(as
C r
adio
sa)
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
Ref
eren
ces
Pant
ocse
kH
ouk
et a
lL
und
1951
H
aringkan
sson
H
ouk
et a
lH
aringkan
sson
H
ouk
et a
lH
aringkan
sson
H
ouk
et a
l19
01 p
104
20
10 p
39
p 9
820
02 p
116
20
10 p
41
2002
p 1
142
010
p 3
720
02 p
113
20
10 p
36
Plat
e X
V
Plat
es 2
69
Figs
1 2
Figs
422
ndashPl
ates
280
Fi
gs 4
15ndash
Plat
es 2
61
Figs
410
ndash P
late
s 25
7 1
ndashFi
gs 3
19
1ndash11
270
42
61ndash
11 2
81
421
1ndash11
262
41
423
258
1ndash1
133
21ndash
6 2
71
1ndash6
1ndash6
1ndash12
263
25
9 1
ndash12
1ndash5
264
1ndash6
260
1ndash8
Col
onie
sno
dat
a2ndash
82ndash
7 ce
lls2ndash
8 ce
lls
shor
tC
ells
solit
ary
no d
ata
mos
tlyce
lls s
olita
ryno
dat
alo
osel
ylo
osel
ylo
osel
yor
in s
hort
solit
ary
rare
ly in
unite
dun
ited
unite
d ch
ains
loos
ely
unite
dsh
ort c
hain
sch
ains
2(8
)ce
llsD
iam
eter
16ndash2
016
ndash22
87ndash
210
8ndash25
8ndash25
8ndash30
7ndash35
7ndash25
8ndash50
5ndash32
(microm
)St
riae
20 p
er 1
0 microm
ca
16
per
15ndash1
85
per
13ndash1
9 pe
r13
ndash15
per
15ndash1
8 pe
r 10
no d
ata
15ndash1
8 pe
r 10
no d
ata
14ndash1
8 pe
r10
microm
of
10 micro
m10
microm
10 micro
mmicrom
of n
earl
ymicrom
of n
early
10 micro
mne
arly
equ
aleq
ual l
engt
heq
ual l
engt
hle
ngth
occ
upy
occu
py 1
2ndash1
4ta
king
abo
ut1
3ndash1
2 of
of th
e va
lvae
12
of th
e va
lve
valv
e fa
cedi
amet
er fa
ce d
iam
eter
diam
eter
517
Cen
tral
are
a
flat
tofla
t to
slig
htly
flat t
o sl
ight
lyce
ntra
l are
aco
ncen
trica
llyfla
t or s
light
lysl
ight
ly c
on-
dist
inct
ly c
on-f
lat t
o co
n-fl
at to
con
-sl
ight
ly c
on-
conc
entri
cally
conc
entri
cally
with
mor
e or
undu
late
co
ncen
trica
llyce
ntri
cally
cent
rica
llyce
ntri
cally
cent
rica
llyce
ntric
ally
un-
undu
late
un
dula
te
less
regu
larl
yw
ith sc
atte
red
undu
late
un
dula
te
undu
late
and
undu
late
un
dula
te
dula
te w
ithw
ith C
F an
dw
ith n
ume-
arra
nged
or ra
dial
lyce
ntra
lra
dial
lyra
dial
lysm
ooth
smoo
th C
Fnu
mer
ous C
Far
eola
ero
us C
F an
dra
dial
arra
nged
CF
areo
lae
and
wrin
kled
C
Fw
rinkl
ed C
Fan
d ar
eola
ean
d ar
eola
ear
rang
ed in
areo
lae
ope-
punc
taan
d ar
eola
eC
F sc
atte
red
and
areo
lae
and
areo
lae
scat
tere
d or
open
ings
mor
e or
less
ning
s sca
ttere
dor
arr
ange
dar
rang
ed in
alw
ays
arra
nged
insc
atte
red
orra
dial
row
sor
occ
asio
nal-
in ra
dial
radi
al p
atte
rnar
rang
ed in
radi
al ro
ws
arra
nged
inly
arr
ange
d in
row
sra
dial
row
sra
dial
pat
tern
radi
al p
atte
rn(F
igs
8 amp
9)
Rim
o-2
ndash3 in
the
(1)2
ndash5 in
the
1ndash2(
3) in
the
no d
ata
at le
ast o
ne
1ndash5
in th
eat
leas
t one
at
leas
t one
in
at le
ast o
ne
at le
ast o
ne
port
ulae
mar
gina
lm
argi
nal
mar
gina
l
Inte
rnal
lyon
the
valv
aem
argi
nal a
rea
on th
e va
lve
the
mar
gina
lon
the v
alva
eon
the
valv
aear
ea n
ear t
hear
ea n
ear t
hear
ea n
ear t
hea
sess
ile la
bi-
face
at t
hecl
ose
to li
mit
face
at t
hear
ea c
lose
tofa
ce a
t the
face
at t
heva
lve
mar
gin
valv
e mar
gin
valv
e mar
gin
um in
obl
ique
end
of s
hor-
of th
e ce
ntra
len
d of
sho
r-lim
it of
the
end
of s
hor-
end
of s
light
lyE
xter
nal o
pe-
Ext
erna
lE
xter
nal
posi
tion
lo-
tene
d st
ria
area
Ext
erna
lte
ned
stri
ace
ntra
l are
ate
ned
stri
ash
orte
ned
stri
ani
ngs
at th
eop
enin
gs a
top
enin
gs a
tca
ted
betw
een
Inte
rnal
ly a
open
ings
on
Inte
rnal
ly a
Ext
erna
l ope
-In
tern
ally
aIn
tern
ally
a la
rge
end
of s
hor-
the
end
ofth
e en
d of
alve
olar
sess
ile la
bi-
a hy
alin
e ar
eare
lativ
ely
ning
s on
ala
rge
sess
ilese
ssile
labi
umte
ned
stri
ash
orte
ned
shor
tene
dop
enin
gsum
in ra
dial
at th
e en
d of
larg
e se
ssile
hyal
ine
area
labi
um in
in o
bliq
ue o
rIn
tern
ally
ast
ria
Inte
r-st
ria
Inte
r-an
d ce
ntra
lpo
sitio
n lo
-a
slig
htly
labi
um in
slit
at th
e en
dob
lique
po-
circ
umfe
rent
ial
smal
l ses
sile
nally
a se
ssile
nally
a sm
all
area
cate
d m
id-
shor
tene
dpo
sitio
n lo
-of
a s
light
lysi
tion
loca
-po
sitio
n lo
cate
dla
bium
clo
sela
bium
ver
yse
ssile
labi
-w
ay b
etw
een
stria
Int
erna
l-ca
ted
betw
een
shor
tene
dte
d be
twee
nin
the
mar
gina
lto
inte
rnal
clos
e to
inte
r-um
clo
se to
alve
olar
ope
-ly
a s
essi
leal
veol
ar o
pe-
stri
a In
ter-
alve
olar
ope
-ar
ea n
ear t
o th
eal
veol
ar o
pe-
nal a
lveo
lar
inte
rnal
alv
e-ni
ngs
and
labi
um in
ning
s an
dna
lly a
ses
-ni
ngs
and
cent
ral a
rea
ning
s w
ithop
enin
gs
olar
ope
ning
sce
ntra
l are
aob
lique
to c
ir-ce
ntra
l are
asi
le la
bium
cent
ral a
rea
radi
ally
or
with
radi
ally
with
radi
ally
cum
fere
ntia
lw
ith a
slit
ori-
obliq
uely
po-
or o
bliq
uely
or o
bliq
uely
slit
posi
tion
ente
d ob
lique
-si
tione
d sl
itpo
sitio
ned
posi
tione
d sl
itly
to n
earl
ysl
itci
rcum
fere
n-tia
llyIn
tern
alsm
all
cost
aesm
all
cost
aesm
all
cost
ae
radi
ally
radi
ally
elon
-ra
dial
ly e
lon-
smal
l co
stae
smal
l co
stae
smal
l co
stae
smal
l co
stae
alve
olar
bear
ing
MF
bear
ing
MF
bear
ing
MF
elon
gate
dga
ted
cos
tae
gate
d c
osta
ebe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fop
enin
gscl
early
larg
ercl
early
larg
ercl
early
larg
erco
stae
bea
-be
arin
g M
Fbe
arin
g M
Fcl
earl
y la
rger
clea
rly la
rger
clea
rly la
rger
clea
rly
larg
erth
an th
in ri
bsth
an th
in ri
bsth
an th
in ri
bsri
ng M
Fcl
early
larg
ersl
ight
ly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
than
thin
ribs
clea
rly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
518
Lak
e B
alat
onM
esot
roph
ic
Eut
roph
ic
Plan
kton
of a
no d
ata
Plan
kton
or
no d
ata
Plan
kton
of
no d
ata
foss
il di
atom
near
Sioacute
fok
eutr
ophi
chy
pere
utro
-nu
mbe
r of
litto
ral o
ligo
olig
om
eso-
lake
s re
ser
ph
ic re
serv
oir
lake
s po
orm
esot
roph
ictr
ophi
csl
owly
run
-in
org
anic
lake
sm
ainl
y su
b-ni
ng w
ater
sm
atte
r and
alpi
ne la
kes
One
of t
heso
me
quar
rym
ost c
omm
onpo
ols
inE
urop
ean
Eng
lish
Lak
esp
ecie
s of
the
Dis
tric
tC
-rad
iosa
com
plex
Tab
le 3
con
tinue
d
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
519
samples declined in subsequent winter-spring months which is a typical consequenceof diatom cell division The number of striae in 10 microm was found to be lower (11 to17 striae in 10 microm) in some specimens from Rusałka Lake than has been originallystated by Pantocsek (1901) It might be related to the striae counting method becauseeven at original Pantocseks drawings (Plate XV figs 319 332) number of striae in10 microm is lower when the Genkalrsquos method (1977) is applied It also remains inaccordance with our observations of the type material (Table 3) and the observationsin Houk et al (2010)
Puncticulata balatonis is morphologically similar to several species of the complexmaking it difficult for a non-specialist to distinguish them The species resemblesP radiosa but differs from it in valve face ultrastructure such as the location of theinternal rimoportula openings The valves of P radiosa have a more coarse andradially wrinkled central area in the valve face whereas P balatonis has a smootherand finely silicified valve face Additionally the radial pattern of the valve facefultoportulae and areolae is very distinct in P radiosa The external rimoportulaopenings of P balatonis are situated at the end of shortened stria well discernible inLM as gaps (notae) whereas according to Houk et al (2010) the external openingsof P radiosa are situated at the end of only slightly shortened striae The differencesare clearly visible from the internal valvae view because P balatonis has valve facerimoportula openings located close to the internal alveolar openings whereas the
Fig 23 The variation in Puncticulata balatonis valve size in Rusałka Lake in subsequent months in2007
520
rimoportula openings in P radiosa are located more close to the central area ofthe valves Such a close location of the internal rimoportula openings to the valvemargin is typical only for P balatonis within the discussed group (Table 3)
Another diatom showing a similar morphology to Puncticulata balatonis is P prae-termissa (Lund) Haringkansson Most of the morphological features are similar forboth species (Table 3) but the later possesses radially elongated relatively largeinternal alveolar openings Moreover the costae bearing marginal fultoportulaeare only slightly thicker than the rest and the internal rimoportula openings arecloser to the central part of the valve face (Table 3) The last feature as well as themarginal stria length enable also the morphological distinction between P balatonisand P comta
Ecology and distribution
The distinction between the species of the Puncticulata radiosa-complex includingPuncticulata balatonis is not easy for a plankton ecologist or other researcher whois not a centric diatom specialist The correct identification requires detailedmorphological studies with SEM andor specialized knowledge That is why apartfrom the long-lasting taxonomical confusion the real distribution of the speciesfrom the complex is uncertain According to Houk et al (2010) Puncticulata balatonis(as Cyclotella balatonis) is one of the most common members of the P radiosa (as
Fig 24 The number of Puncticulata balatonis frustules in the water column of Rusałka Lake in 2007
521
Cyclotella radiosa) complex in Europe found in numerous mesotrophic and eutrophiclakes and slowly running waters in Hungary Germany and Czech Republic Howeverreports of the species are not numerous Till now P balatonis has also never beenreported from Poland
The limited number of reports on the occurrence of P balatonis may be related to itsmisidentificationmisinterpretation as Cyclotella comta (Ehrenb) Kuumltz (Kiss amp Pająk1994 fig 13) found in the plankton of the eutrophic Vistula River as Cyclotellapraetermissa quadrijuncta (Wunsam et al 1995) or as Puncticulata radiosa (Wojtalamp Kwandrans 2006) found in the eutrophic Rudawa River We are convinced that apart of other records of P comta the most commonly reported species of the complexin Poland (Siemińska amp Wołowski 2003) in fact might concern P balatonis InRusałka Lake the species may have also been mistaken for P comta (asC comta) reported from the lake by Kotlińska (1976) It is also likely that P bala-tonis in Poland may have been misidentified as P praetermissa although the latterspecies was only reported several times including findings in the benthos of CzarneLake and the Kamionka River NE Poland (Rakowska 1996) in the plankton ofseveral lakes of the Drawa National Park NW Poland (Gołdyn amp Szeląg-Wasielewska2004) in a sand-pit lake in Krakoacutew and a spring fed pond in Krztynia in S Poland(Wojtal unpublished data)
Up to now there has also been much confusion around the ecology of the speciesfrom the complex For instance P praetermissa described by Lund (1951) fromlakes with a low organic matter content was later found both in the plankton and thebenthos in a wide variety of natural and man-made lakes including alpine andsubalpine lakes (Wunsam et al 1995 Lange-Bertalot amp Metzeltin 1996 Guumlttingeramp Straub 1998 Tolotti et al 2004) as well as in more eutrophic habitats (egSchmidt et al 2002 Hindaacutek amp Hindaacutekovaacute 2003 Tanaka 2007) Houk et al (2010)cleared up the ambiguities ascribing P praetermissa and P radiosa to oligo-mesotrophic habitats According to the authors P comta (as Cyclotella comta) is tillnow known only from fossil materials P balatonis (as Cyclotella balatonis) inturn has been assigned to meso-eutrophic lakes and rivers consistently with thetrophic character of its type locality ndash lake Balaton Our results correspond with theautecology of P balatonis (as Cyclotella balatonis) proposed by Houk et al (2010)widening even slightly the preferences of the species for the trophic conditionsdetermined by Houk et al (2010) as it was also able to develop a large populationin an eutrophic to hypereutrophic water body
The highest numbers of P balatonis in Rusałka Lake were observed in the winter-spring period The diatom bloom in the spring plankton is a well-known phenomenon(Sommer et al 1986) ndash sometimes also occurring during the winter period (Kiss ampGenkal 1993) ndash and is usually associated with a changing silica andor phosphorusavailability during that period (Lampert amp Sommer 2001) Sinking was also provedto be an essential factor hampering diatom success during the thermal stratificationperiod (Jewson et al 1981 Ferris amp Lehman 2007) explaining probably the seasonalityof the P balatonis growth in Rusałka Lake as its high development finished withthe stratification onset Relatively high numbers of frustules in deeper water layersmight also indicate a significant role of the sedimentation process
522
Conclusions
The morphology of Puncticulata balatonis in Rusałka Lake corresponds well toother species around P comta with the exception of the location of marginalfultoportula openings and the length of the shortened striae (notae) Analysis of themorphological characters enabled the transfer of Cyclotella balatonis into the genusPuncticulata A large population of the species in the eutrophic to hypereutrophicRusałka Lake remains congruent with its tolerance ranges with respect to the trophicstate proposed by Houk et al (2010) The variation in the cell number and averagedimension of frustules indicated the species preference to develop large populationsin spring
Acknowledgements
We thank Prof Ryszard Gołdyn (AMickiewicz University in Poznań) for his inspiration researchsupervision and useful comments to the manuscript We are also grateful to Dr Bart Van de Vijver andtwo anonymous reviewers for their valuable suggestions and to Dr Krisztina Buczko for enabling theanalysis of the type material of Cyclotella balatonis from the Pantocsekrsquos collection at HungarianNatural History Museum Sampling on Rusałka Lake was part of a project financially supported by theDistrict and Municipial Funds of Enviromental Protection and Water Management in Poznań (Poland)
References
CARLSON RE (1977) A trophic state index for lakes ndash Limnol Oceanogr 22 (2) 361ndash369
CREMER H amp H KOOLMEES (2010) Common diatoms (Centrales and Fragilariaceae Bacillario-phyta) of modern and fossil freshwater environments in the Netherlands ndash Nova Hedwigia 90(3ndash4)343ndash381
FERRIS JA amp JT LEHMAN (2007) Interannual variation in diatom bloom dynamics Roles ofhydrology nutrient limitation sinking and whole lake manipulation ndash Water Res 41 2551ndash2562
GENKAL SI (1977) On the method of calculation of some taxonomicaly significant structuralelements of the valve in the diatoms of the family Thalassiosiraceae Lebour emend Hasle (Bacillariophyta)ndash Bot Zhurn 62 848ndash851 (in Russian)
GENKAL SI amp KT KISS (1993) Morphological variability of the diatom Cyclotella atomusHustedt var atomus and C atomus var gracilis var nov ndash Hydrobiologia 269270 39ndash47
GOŁDYN R S PODSIADŁOWSKI K KOWALCZEWSKA-MADURA R DONDAJEWSKAE SZELĄG-WASIELEWSKA A BUDZYŃSKA P DOMEK amp W ROMANOWICZ (2010)Functioning of the Rusałka Lake ecosystem in Poznań (west Poland) ndash Ocean Hydrobiol Stud39(3) 65ndash80
GOŁDYN R amp E SZELĄG-WASIELEWSKA (2004) Changes in the phytoseston of a river-lakesystem in Drawieński National Park ndash Ocean Hydrobiol Stud 33(2) 17ndash28
GUumlTTINGER W amp F STRAUB (1998) Diatoms of Lake Cadagno ndash In PEDUZZI RR BACHOFEN amp M TONOLLA (eds) Lake Cadagno a meromictic Alpine lake ndash Doc Ist ItalIdrobiol 63 57ndash64
HANDMANN R (1913) Die Diatomeenflora des Almfeegebiets ndash In MANZ GJ (ed) Mitteilungendes Mikrologischen Vereins Linz ndash Mikrolog Ver Regensburg (in German)
HAUSMANN S amp AF LOTTER (2001) Morphological variation within the diatom taxon Cyclotellacomensis and its importance for quantitative temperature reconstructions ndash Freshw Biol 46 1323ndash1333
523
HAYASHI T (2007) Puncticulata versiformis sp nov and Cyclotella kathmanduensis sp nov(Bacillariophyta) new fossil species from middle Pleistocene lacustrine sediments Kathmandu NepalHilalaya ndash J Phycol 43(2) 304ndash318
HAringKANSSON H (2002) A compilation and evaluation of species in the genera StephanodiscusCyclostephanos and Cylotella with a new genus in the family Stephanodiscaceae ndash Diatom Res 17(1)1ndash139
HINDAacuteK F amp A HINDAacuteKOVAacute (2003) Diversity of cyanobacteria and algae of urban gravel pitlakes in Bratislava Slovakia a survey ndash Hydrobiologia 506ndash509 155ndash162
HINDAacuteK F amp A HINDAacuteKOVAacute (2008) Diversity of cyanobacteria and algae of the NationalNature Reserve Čiacutečovskeacute mŕtve rameno oxbow (Southern Slovakia) ndash Bulletin SBS 30 (1) 11ndash19 (inSlovak with an English abstract)
HOUK V R KLEE amp H TANAKA (2010) Atlas of freshwater centric diatoms with a brief key anddescriptions Part III Stephanodiscaceae A Cyclotella Tertiarius Discostella ndash Fottea 10 (Supplement)1ndash498
JEWSON DH BH RIPPEY amp WJK GILMORE (1981) Loss rates from sedimentation parasitismand grazing during the growth nutrient limitation and dormancy of a diatom crop ndash Limnol Oceanogr26 1045ndash1056
KISS KT amp SI GENKAL (1993) Winter blooms of centric diatoms in the River Danube and in itsside arms near Budapest ndash Hydrobiologia 269270 317ndash325
KISS KT amp G PAJĄK (1994) Seasonal change of diatom s in the plankton of the Vistula riverabove and below the Goczalkowice reservoir Poland ndash In KOCIOLEK JP (ed) Proceedings ofthe 11th International Diatom Symposium San Francisco Aug 12ndash17 1990 Memoirs of the CaliforniaAcademy of Sciences 17 517ndash524 San Francisco California Academy of Sciences
KOBAYASI H M IDEI S MAYAMA T NAGUMO amp K OSADA (2006) H KobayasirsquosAtlas of Japanese diatoms based on electron microscopy 1 1ndash531 Uchida Rokakuho Publishing CoLtd Tokyo
KOTLIŃSKA E (1976) Fitoplankton Jeziora Rusałka In Fitoplankton sztucznych jezior położonychna terenie Poznania 1ndash54 Poznańskie Tow Przyjacioacuteł Nauk Prace Komisji Biologicznej (in Polish)
KRAMMER K amp H LANGE-BERTALOT (1991) Bacillariophyceae 3 Teil Centrales Fragilaria-ceae Eunotiaceae ndash In ETTL H J GERLOFF H HEYNIGH amp D MOLLENHAUER (eds)Suumlsswasserflora von Mitteleuropa 2(3) 1ndash576 Gustav Fischer Verlag Stuttgart Jena
LAMPERT W amp U SOMMER (2001) Ekologia woacuted śroacutedlądowych ndash PWN Warszawa
LANGE-BERTALOT H amp D METZELTIN (1996) Indicators of oligotrophy 800 representativeof three ecologically distinct lake types Carbonate buffered-Oligotrophic-Weakly buffered soft waterndash Iconogr Diatomol 2 1ndash390
LUND JWG (1951) Contribution to our knowledge of British algae ndash Hydrobiologia 3(1) 93ndash100
ORGANIZATION FOR ECONOMIC COOPERATION AND DEVELOPMENT (1982)Eutrophication of Waters ndash Monitoring Assessment and Control Final Report OECD Paris
PANTOCSEK J (1901) Die Kieselalgen oder Bacillarien des Balaton 1ndash112 XVI Tafeln ndash Druckder K und K Hofbuchdruckerei des Victor Hornyaacutenszky Budapest
RAKOWSKA B (1996) Preliminary elaboration of the diatom communities occuring in the benthosof the Kamionka River and Lake Czarne at the Pojezierze Suwalskie (1993ndash1994) ndash Fragm FloristGeobot Polon 3 221ndash238
RIMET F JC DRUART amp O ANNEVILLE (2009) Exploring the dynamics of plankton diatomcommunities in Lake Geneva using emergent self-organizing maps (1974ndash2007) ndash Ecol Inform4 99ndash110
524
SCHMIDT R R PSENNER J MUumlLLER P INDINGER amp C KAMENIK (2002) Impact of thelate glacial climate variations on stratification and trophic state of the meromictic lake Laumlngsee (Austria)validation of a conceptual model by multiproxy studies ndash J Limnol 61(1) 49ndash60
SIEMIŃSKA J amp K WOŁOWSKI (2003) Catalogue of Polish prokaryotic and eukaryotic algaendash W Szafer Institute of Botany Polish Academy of Sciences Krakoacutew
SIVER PA (1999) Development of paleolimnological inference models for pH total nitrogen andspecific conductivity based on planktonic diatoms ndash J Paleolimnol 21 45ndash59
SOMMER U ZM GLIWICZ W LAMPERT amp A DUNCAN (1986) The PEG-model of seasonalsuccession of planktonic events in fresh waters ndash Arch Hydrobiol 106 433ndash471
TANAKA H (2007) Taxonomic studies of the genera Cyclotella (Kuumltzing) Breacutebisson DiscostellaHouk et Klee and Puncticulata Haringkansson in the family Stepanodiscaceae Glezer et Makarova(Bacillariophyta) in Japan ndash Biblioth Diatomol 53 1ndash205
TOLOTTI M D CALLIARI amp F CORRADINI (2004) Interannual variability of the phytoplanktonof Tovel lake (Trentino Italy) ndash Studi Trent di Scienze Naturali Acta Biol 81 Suppl 2 327ndash340 (inItalian)
WETZEL RG amp GE LIKENS (1991) Limnological analyses 1ndash391 ndash Springer-Verlag NewYork Berlin Heidelberg
WOJTAL AZ amp J KWANDRANS (2006) Diatoms of the Wyżyna Krakowsko-Częstochowskaupland (S Poland) ndash Coscinodiscophyceae (Thalassiosirophycidae) ndash Polish Bot J 51(2) 177ndash207
WUNSAM S R SCHMIDT amp R KLEE (1995) Cyclotella-taxa (Bacillariophyceae) in lakes of theAlpine region and their relationship to environmental variables ndash Aquat Sci 57(4) 360ndash381
Received 25 March 2010 accepted in revised form 31 March 2011

510
Despite some of the members of Cyclotella genus are known for up to 170 years theecology of many species still remains uncertain Moreover there are also manyuncertainties within the genus concerning taxonomy and nomenclature (Hausmannamp Lotter 2001 Haringkansson 2002 Houk et al 2010) Recently some species of thenot-uniform genus were moved to several newly-established genera (short review inHouk et al 2010) In 2002 the genus Puncticulata was established comprisingspecies characterised by concentrically undulated central part of valve face andnumerous scattered or radially arranged areolae and fultoportulae (or only areolaeor only fultoportulae) with P comta as the type (Haringkansson 2002) The genus hasbeen generally accepted (eg Tanaka 2007 Hayashi 2007 Kobayasi et al 2006Cremer amp Koolmees 2010) though some uncertainties are still discussed (Houk etal 2010)
One of the species sharing the above-mentioned characteristics of the genusPuncticulata (though not transferred to the genus till now) is Cyclotella balatonisand its variety C balatonis var binotata described by Pantocsek (1901 p 104 figs319 331 332) from lake Balaton near Sioacutefok in Hungary Originally the specieswas illustrated by three drawings of valves with scattered (Pantocsek 1901 PlateXV figs 319 332) and radially arranged (Pantocsek 1901 Plate XV fig 331)puncta in the middle part of valve face After Pantocsekrsquos description C balatonisremained practically forgotten and was reported only a few times (eg Handmann1913 Hindaacutek amp Hindaacutekovaacute 2008) It was generally considered a synonym ofCyclotella radiosa (eg Krammer amp Lange-Bertalot 1991 fig 62 11 12) followingthe broad species concept that was in use at that time Thus specimens of a broadmorphological range present in a variety of water bodies were identified as C radi-osa Cyclotella balatonis might have also been misidentified as other species of thecomplex such as P praetermissa (syn Cyclotella praetermissa) or Puncticulatacomta (syn Cyclotella comta) Recently as a result of precise study by Houk et al(2010) using SEM Cyclotella balatonis has been resurrected and the morphologicalranges of similar species were discussed However due to the proceeding long periodof taxonomical confusion the species is almost absent in the literature and needsmuch attention particularly in terms of distribution and autecology
In this paper a population of C balatonis in a small and shallow urban eutrophic tohypereutrophic lake (Rusałka Lake Western Poland) is presented with its quantitativecharacteristics along an annual cycle Morphological features of the diatom aredescribed in detail and documented with LM photographs of live specimens andcleaned frustules as well as with the SEM micrographs Cyclotella balatonis wasalso compared with morphologically similar species namely Puncticulata comtaP praetermissa and P radiosa Due to the great morphological similarity of Cyclotellabalatonis to these and other Puncticulata-species the transfer of Cyclotella balatonisto the genus Puncticulata has been proposed
Study site
Rusałka Lake is situated in the city of Poznań Western Poland (N52deg25E16deg52) It was formed in1943 by damming the Bogdanka River and subsequent flooding of its valley The lake covers the area
511
of 367 ha Although its maximum depth is 90 meters the average does not exceed 2 meters As aresult of the morphometry the lake is mainly polymictic with thermal stratification developing only ina small area around its deepest point in summer (Gołdyn et al 2010)
The lake is strongly eutrophic or hypereutrophic according to different criteria (Table 1) Cyanobacterialblooms occur in summer and autumn (Gołdyn et al 2010) Due to its location and surroundingattractive areas (mainly forests) the lake is intensely used for recreation
Materials and methods
Water samples for the phytoplankton analysis were taken at every meter depth at the deepest part of thelake in 2007 once a month from February till October and once in December Quantitative sampleswere preserved with Lugolrsquos solution and analysed with the use of inverted microscope PZO aftersedimentation in cylindrical chambers according to the Utermoumlhl method (Wetzel amp Likens 1991)
All diatoms from the family Stephanodiscaceae were counted as one taxon The relative frequency ofPuncticulata balatonis and other species belonging to this family was then estimated by counting 100frustules in hydrogene peroxide-treated samples using an Olympus BX-60 microscope Measurementsand the micrographs of live diatoms were done with the use of the same LM and AnalySIS programme
A qualitative analysis of the diatom material was performed after removing carbonates (addition of10 HCl to the samples followed by boiling taking place after 24 h for 15 minutes) and organic matter(boiling in 30 H
2O
2 with small amounts of KClO
3) After air-drying the material was mounted in
Naphraxreg Observations of the diatoms were performed with a Nikon Eclipse 80i light microscopeequipped with Differential Interference Contrast The micrographs were taken with a Nikon DS-Fi 1camera SEM analyses were performed on the material coated with gold using a Hitachi S-4700scanning electron microscope SEM micrographs were taken in the Laboratory of Electron and ConfocalMicroscopy at the Faculty of Biology Adam Mickiewicz University in Poznań and in the Laboratoryof Field Emission Scanning Electron Microscopy and Microanalysis at the Institute of GeologicalSciences of the Jagiellonian University in Krakoacutew Density of valve face structures (eg costae) in10 microm in a line tangential to the radius was calculated based on the formula in Genkal (1977)Additionally data on the morphology of Cyclotella balatonis were obtained from the type material(Balaton Lake Siofok samples 156 157) hosted in the Natural History Museum in Budapest (Hungary)
Table 1 Estimation of the trophic state of Rusałka Lake according to Carlson index (1977) and OECD(1982) criteria (acc to Gołdyn et al 2010)
Carlson indexTSI Values in Boundary values Trophic state
Rusałka lake for eutrophy
TSI (TP) 7437 gt70 HypertrophyTSI (ChL) 7171 53ndash70 HypertrophyTSI (SD) 6737 53ndash70 Eutrophy
OECDCharacteristic Values in Boundary values Trophic state
Rusałka lake for eutrophy
Mean TP (microgl) 96 gt100 EutrophyMean chlorophyll-a (microgl) 347 gt25 HypertrophyMax chlorophyll-a (microgl) 802 gt75 HypertrophyMean Secchi depth (m) 08 lt15 HypertrophyMin Secchi depth (m) 06 lt07 Hypertrophy
512
Water temperature pH and conductivity were measured in situ using YSI 610-DM-meter Watertransparency was expressed as the visibility of the Secchi disc For the statistical graphs Statistica 80software was used
Results
Morphology of Puncticulata balatonis in Rusałka Lake
Puncticulata balatonis (Pantocsek) Wojtal et Budzyńska comb nov Figs 1ndash12 amp 15ndash22
BASIONYM Cyclotella balatonis Pantocsek Die Kieselalgen oder Bacillarien des Balaton p 104 PlateXV figs 319 332 1901
TYPE LOCALITY Balaton Lake near Sioacutefok Hungary
Cells solitary or in short 2ndash7-cell chains (Fig 1) containing multiple discoidchloroplasts The valve diameter ranges between 87 and 210 microm Stria density is15ndash185 in 10 microm with 35ndash47 costae (thickened ribs) in 10 microm Striae are ofunequal length with inserted shorter (Figs 15ndash17) or divided striae (Fig 19) formingan irregularly circular central area (Figs 6 9 12 17) The central area is concentricallyslightly elevated (Figs 15 17) with numerous scattered openings of valve facefultoportulae (externally visible as minor puncta Fig 16) and areolae (externallyvisible as bigger puncta Figs 15ndash17) External openings of the rimopor-tulae arepresent at the end of the shortened striae (Figs 15 16) The internal opening of thesessile rimoportulae has a radially or slightly obliquely positioned slit (Figs 18 20)situated close to internal alveolar openings (towards the valve face margin) Marginalfultoportula openings with two satellite pores are situated (internally) on thethickened costae (shadow lines) separated by 3ndash5 thinner costae Valve facefultoportula openings are internally covered with domed cribra and surrounded bythree satellite pores (Figs 19 20) The morphology of the valve face internal alveolaropenings and the location of internal rimoportula openings correspond well withthe morphology of the specimens from the type material of Cyclotella balatonis(Figs 13 14 amp 21 22 Table 3)
Table 2 Physio-chemical parameters of water of Rusałka Lake in 2007
Temperature pH Coductivity Transparencyndash(degC) (microScm) Secchi depth (m)
20 II 31ndash41 72ndash78 836ndash841 1526 III 49ndash71 79ndash81 866ndash868 1820 IV 134ndash103 79ndash80 no data 1018 V 110ndash156 77ndash82 820ndash840 0819 VI 112ndash232 73ndash84 760ndash878 0918 VII 126ndash240 75ndash83 697ndash754 08520 VIII 145ndash220 75ndash91 627ndash739 0612 IX 144ndash148 83ndash85 646ndash652 0822 X 85ndash89 80ndash83 668ndash680 10503 XII 35ndash38 81ndash82 740ndash744 135
513
Ecological data
The water of Rusałka Lake was alkaline throughout the study period showing highconductivity Water transparency was relatively low ranging from 06 to 18 m ofSecchi depth (Table 2)
Puncticulata balatonis was present in the lake in all the seasons of 2007 It wasfound to form chains only in February March and April The diatom valve sizedistribution pattern varied in subsequent months with the largest average valve sizein February (Fig 23) Its abundance in the lake was also higher in the winter-springperiod (from February till May fig 24) with the maximum value of 35 times 103 cellsml-1 and a biomass of 298 mg l-1 (19 of the total phytoplankton cell number and525 of the total biomass at 6 m depth in March) In other spring samples
Figs 1ndash14 Puncticulata balatonis (Pantocsek) Wojtal et Budzyńska Light microscopy Figs 1ndash12 Specimens from the Rusałka Lake Figs 13ndash14 P balatonis type material sample no 152Fig 1 Short chain of alive cells in girdle view Figs 5ndash6 The same valve at different focus levels OnFigs 6 12ndash13 the characteristic gaps (notae) are marked with an arrow Scale bar represents 10 microm
514
Figs 15ndash22 Puncticulata balatonis (Pantocsek) Wojtal et Budzyńska Scanning electron microscopyFigs 15ndash20 specimens from the Rusałka Lake Figs 21ndash22 P balatonis type material samples no152 amp 156 Fig 15 external view of the rimoportula opening at the end of shortened striae ndash notae
515
P balatonis contributed between 63 and 187 to the total phytoplankton numbersand between 21 and 47 to the total biomass From June until October the numberof the diatom did not exceed 300 frustules per 1 ml (Fig 24) and no valve wasfound in September At the beginning of December P balatonis number increasedslightly again
At the time of the bloom of P balatonis chrysoflagellates (Erkenia subaequiciliataSkuja Chrysococcus triporus Matvienko) pennate diatoms (Fragilaria sp) andorcoenobia-forming Chlorococcales (Didymocystis inconspicua Koršikov Tetastrumsp div Scenedesmus sp div Coelastrum microporum Naumlgeli Crucigenia tetrapediaKirchner) were also abundant Other diatoms such as Aulacoseira ambigua (Grunow)Simonsen Cyclotella meneghiniana Kuumltzing C ocellata Pantocsek Cyclostephanosdelicatus (Genkal) Casper C dubius (Fricke) Round Stephanodiscus parvus Stoermeramp Haringkansson Asterionella formosa Hassal and Navicula sp were also observed
Discussion
Morphology of Puncticulata balatonis (Pant) Wojtal et Budzyńska
Puncticulata balatonis shares important morphological characters assigned to thegenus justifying its formal transfer to this genus a slightly concentrically undulatemiddle part of the valve face with scattered or radially arranged numerous areolaewith internally domed cribra and numerous fultoportulae Additionally valves ofP balatonis have internal alveolar openings with marginal fultoportulae located onthicker costae rimoportulae located externally at the top of shortened striae andinternally in the marginal part of the valve face
The morphology of P balatonis valves from Rusałka Lake corresponds well withthe morphology of the specimens from the Pantocsekrsquos type material (Figs 13 1421 22 Table 3) though the diameter range and the number of striae in 10 microm werefound to be different Pantocsek (1901) stated in his diagnosis (p 104) that thevalves of Cyclotella balatonis are 16ndash20 microm in diameter but we have found bothsmaller and larger specimens (Table 3) The wider diameter range in our observationcould be related to a more complete morphological range of the species since thediatom population in Rusałka Lake was abundant However the time of samplingmay have also been of importance as the medium valve size of the species in our
(arrowed) at concentrically elevated valve face Fig 16 external view of the openings of central (right)and marginal (left) fultoportulae (arrowed) Fig 17 external view of the short and long striae of themarginal area Fig18 internal view of the rimoportula opening (arrowed) located close to the valvemargin and internal alveolar opening and central openings of numerous areolae and fultoportulae(arrowed) Fig 19 internal view of the divided thinner costa (marked) and internal opening of centralfultoportula (arrowed) Fig 20 internal view of the marginal fultoportulae located on the thicker costae(arrowed) Fig 21 external view of the valve face with marked notae Fig 22 internal view of thealveolar openings with thicker costa bearing marginal fultoportulae and internal rimoportula openinglocated near the margin (arrowed) Scale bar represents 5 microm for Figs 15 18 amp 21 4 microm for Fig 163 microm for Figs 19 amp 20 and 2 microm for Figs 17 amp 22
516
Tab
le 3
Com
pari
son
of P
unct
icul
ata
bala
toni
s an
d m
orph
olog
ical
ly s
imila
r sp
ecie
s C
F ndash
cent
ral
fulto
port
ulae
MF
ndash m
argi
nal
fulto
port
ulae
o
urob
serv
atio
n ty
pe m
ater
ial S
iofoacute
k s
ampl
e no
152
our
obse
rvat
ion
base
d on
the
fig
62
11
12
in K
ram
mer
amp L
ange
-Ber
talo
t 199
1 w
hich
sho
ws
spec
imen
s fr
om th
e ty
pe m
ater
ial o
f Cyc
lote
lla
bala
toni
s Pa
ntoc
sek
(as
C r
adio
sa)
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
Ref
eren
ces
Pant
ocse
kH
ouk
et a
lL
und
1951
H
aringkan
sson
H
ouk
et a
lH
aringkan
sson
H
ouk
et a
lH
aringkan
sson
H
ouk
et a
l19
01 p
104
20
10 p
39
p 9
820
02 p
116
20
10 p
41
2002
p 1
142
010
p 3
720
02 p
113
20
10 p
36
Plat
e X
V
Plat
es 2
69
Figs
1 2
Figs
422
ndashPl
ates
280
Fi
gs 4
15ndash
Plat
es 2
61
Figs
410
ndash P
late
s 25
7 1
ndashFi
gs 3
19
1ndash11
270
42
61ndash
11 2
81
421
1ndash11
262
41
423
258
1ndash1
133
21ndash
6 2
71
1ndash6
1ndash6
1ndash12
263
25
9 1
ndash12
1ndash5
264
1ndash6
260
1ndash8
Col
onie
sno
dat
a2ndash
82ndash
7 ce
lls2ndash
8 ce
lls
shor
tC
ells
solit
ary
no d
ata
mos
tlyce
lls s
olita
ryno
dat
alo
osel
ylo
osel
ylo
osel
yor
in s
hort
solit
ary
rare
ly in
unite
dun
ited
unite
d ch
ains
loos
ely
unite
dsh
ort c
hain
sch
ains
2(8
)ce
llsD
iam
eter
16ndash2
016
ndash22
87ndash
210
8ndash25
8ndash25
8ndash30
7ndash35
7ndash25
8ndash50
5ndash32
(microm
)St
riae
20 p
er 1
0 microm
ca
16
per
15ndash1
85
per
13ndash1
9 pe
r13
ndash15
per
15ndash1
8 pe
r 10
no d
ata
15ndash1
8 pe
r 10
no d
ata
14ndash1
8 pe
r10
microm
of
10 micro
m10
microm
10 micro
mmicrom
of n
earl
ymicrom
of n
early
10 micro
mne
arly
equ
aleq
ual l
engt
heq
ual l
engt
hle
ngth
occ
upy
occu
py 1
2ndash1
4ta
king
abo
ut1
3ndash1
2 of
of th
e va
lvae
12
of th
e va
lve
valv
e fa
cedi
amet
er fa
ce d
iam
eter
diam
eter
517
Cen
tral
are
a
flat
tofla
t to
slig
htly
flat t
o sl
ight
lyce
ntra
l are
aco
ncen
trica
llyfla
t or s
light
lysl
ight
ly c
on-
dist
inct
ly c
on-f
lat t
o co
n-fl
at to
con
-sl
ight
ly c
on-
conc
entri
cally
conc
entri
cally
with
mor
e or
undu
late
co
ncen
trica
llyce
ntri
cally
cent
rica
llyce
ntri
cally
cent
rica
llyce
ntric
ally
un-
undu
late
un
dula
te
less
regu
larl
yw
ith sc
atte
red
undu
late
un
dula
te
undu
late
and
undu
late
un
dula
te
dula
te w
ithw
ith C
F an
dw
ith n
ume-
arra
nged
or ra
dial
lyce
ntra
lra
dial
lyra
dial
lysm
ooth
smoo
th C
Fnu
mer
ous C
Far
eola
ero
us C
F an
dra
dial
arra
nged
CF
areo
lae
and
wrin
kled
C
Fw
rinkl
ed C
Fan
d ar
eola
ean
d ar
eola
ear
rang
ed in
areo
lae
ope-
punc
taan
d ar
eola
eC
F sc
atte
red
and
areo
lae
and
areo
lae
scat
tere
d or
open
ings
mor
e or
less
ning
s sca
ttere
dor
arr
ange
dar
rang
ed in
alw
ays
arra
nged
insc
atte
red
orra
dial
row
sor
occ
asio
nal-
in ra
dial
radi
al p
atte
rnar
rang
ed in
radi
al ro
ws
arra
nged
inly
arr
ange
d in
row
sra
dial
row
sra
dial
pat
tern
radi
al p
atte
rn(F
igs
8 amp
9)
Rim
o-2
ndash3 in
the
(1)2
ndash5 in
the
1ndash2(
3) in
the
no d
ata
at le
ast o
ne
1ndash5
in th
eat
leas
t one
at
leas
t one
in
at le
ast o
ne
at le
ast o
ne
port
ulae
mar
gina
lm
argi
nal
mar
gina
l
Inte
rnal
lyon
the
valv
aem
argi
nal a
rea
on th
e va
lve
the
mar
gina
lon
the v
alva
eon
the
valv
aear
ea n
ear t
hear
ea n
ear t
hear
ea n
ear t
hea
sess
ile la
bi-
face
at t
hecl
ose
to li
mit
face
at t
hear
ea c
lose
tofa
ce a
t the
face
at t
heva
lve
mar
gin
valv
e mar
gin
valv
e mar
gin
um in
obl
ique
end
of s
hor-
of th
e ce
ntra
len
d of
sho
r-lim
it of
the
end
of s
hor-
end
of s
light
lyE
xter
nal o
pe-
Ext
erna
lE
xter
nal
posi
tion
lo-
tene
d st
ria
area
Ext
erna
lte
ned
stri
ace
ntra
l are
ate
ned
stri
ash
orte
ned
stri
ani
ngs
at th
eop
enin
gs a
top
enin
gs a
tca
ted
betw
een
Inte
rnal
ly a
open
ings
on
Inte
rnal
ly a
Ext
erna
l ope
-In
tern
ally
aIn
tern
ally
a la
rge
end
of s
hor-
the
end
ofth
e en
d of
alve
olar
sess
ile la
bi-
a hy
alin
e ar
eare
lativ
ely
ning
s on
ala
rge
sess
ilese
ssile
labi
umte
ned
stri
ash
orte
ned
shor
tene
dop
enin
gsum
in ra
dial
at th
e en
d of
larg
e se
ssile
hyal
ine
area
labi
um in
in o
bliq
ue o
rIn
tern
ally
ast
ria
Inte
r-st
ria
Inte
r-an
d ce
ntra
lpo
sitio
n lo
-a
slig
htly
labi
um in
slit
at th
e en
dob
lique
po-
circ
umfe
rent
ial
smal
l ses
sile
nally
a se
ssile
nally
a sm
all
area
cate
d m
id-
shor
tene
dpo
sitio
n lo
-of
a s
light
lysi
tion
loca
-po
sitio
n lo
cate
dla
bium
clo
sela
bium
ver
yse
ssile
labi
-w
ay b
etw
een
stria
Int
erna
l-ca
ted
betw
een
shor
tene
dte
d be
twee
nin
the
mar
gina
lto
inte
rnal
clos
e to
inte
r-um
clo
se to
alve
olar
ope
-ly
a s
essi
leal
veol
ar o
pe-
stri
a In
ter-
alve
olar
ope
-ar
ea n
ear t
o th
eal
veol
ar o
pe-
nal a
lveo
lar
inte
rnal
alv
e-ni
ngs
and
labi
um in
ning
s an
dna
lly a
ses
-ni
ngs
and
cent
ral a
rea
ning
s w
ithop
enin
gs
olar
ope
ning
sce
ntra
l are
aob
lique
to c
ir-ce
ntra
l are
asi
le la
bium
cent
ral a
rea
radi
ally
or
with
radi
ally
with
radi
ally
cum
fere
ntia
lw
ith a
slit
ori-
obliq
uely
po-
or o
bliq
uely
or o
bliq
uely
slit
posi
tion
ente
d ob
lique
-si
tione
d sl
itpo
sitio
ned
posi
tione
d sl
itly
to n
earl
ysl
itci
rcum
fere
n-tia
llyIn
tern
alsm
all
cost
aesm
all
cost
aesm
all
cost
ae
radi
ally
radi
ally
elon
-ra
dial
ly e
lon-
smal
l co
stae
smal
l co
stae
smal
l co
stae
smal
l co
stae
alve
olar
bear
ing
MF
bear
ing
MF
bear
ing
MF
elon
gate
dga
ted
cos
tae
gate
d c
osta
ebe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fop
enin
gscl
early
larg
ercl
early
larg
ercl
early
larg
erco
stae
bea
-be
arin
g M
Fbe
arin
g M
Fcl
earl
y la
rger
clea
rly la
rger
clea
rly la
rger
clea
rly
larg
erth
an th
in ri
bsth
an th
in ri
bsth
an th
in ri
bsri
ng M
Fcl
early
larg
ersl
ight
ly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
than
thin
ribs
clea
rly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
518
Lak
e B
alat
onM
esot
roph
ic
Eut
roph
ic
Plan
kton
of a
no d
ata
Plan
kton
or
no d
ata
Plan
kton
of
no d
ata
foss
il di
atom
near
Sioacute
fok
eutr
ophi
chy
pere
utro
-nu
mbe
r of
litto
ral o
ligo
olig
om
eso-
lake
s re
ser
ph
ic re
serv
oir
lake
s po
orm
esot
roph
ictr
ophi
csl
owly
run
-in
org
anic
lake
sm
ainl
y su
b-ni
ng w
ater
sm
atte
r and
alpi
ne la
kes
One
of t
heso
me
quar
rym
ost c
omm
onpo
ols
inE
urop
ean
Eng
lish
Lak
esp
ecie
s of
the
Dis
tric
tC
-rad
iosa
com
plex
Tab
le 3
con
tinue
d
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
519
samples declined in subsequent winter-spring months which is a typical consequenceof diatom cell division The number of striae in 10 microm was found to be lower (11 to17 striae in 10 microm) in some specimens from Rusałka Lake than has been originallystated by Pantocsek (1901) It might be related to the striae counting method becauseeven at original Pantocseks drawings (Plate XV figs 319 332) number of striae in10 microm is lower when the Genkalrsquos method (1977) is applied It also remains inaccordance with our observations of the type material (Table 3) and the observationsin Houk et al (2010)
Puncticulata balatonis is morphologically similar to several species of the complexmaking it difficult for a non-specialist to distinguish them The species resemblesP radiosa but differs from it in valve face ultrastructure such as the location of theinternal rimoportula openings The valves of P radiosa have a more coarse andradially wrinkled central area in the valve face whereas P balatonis has a smootherand finely silicified valve face Additionally the radial pattern of the valve facefultoportulae and areolae is very distinct in P radiosa The external rimoportulaopenings of P balatonis are situated at the end of shortened stria well discernible inLM as gaps (notae) whereas according to Houk et al (2010) the external openingsof P radiosa are situated at the end of only slightly shortened striae The differencesare clearly visible from the internal valvae view because P balatonis has valve facerimoportula openings located close to the internal alveolar openings whereas the
Fig 23 The variation in Puncticulata balatonis valve size in Rusałka Lake in subsequent months in2007
520
rimoportula openings in P radiosa are located more close to the central area ofthe valves Such a close location of the internal rimoportula openings to the valvemargin is typical only for P balatonis within the discussed group (Table 3)
Another diatom showing a similar morphology to Puncticulata balatonis is P prae-termissa (Lund) Haringkansson Most of the morphological features are similar forboth species (Table 3) but the later possesses radially elongated relatively largeinternal alveolar openings Moreover the costae bearing marginal fultoportulaeare only slightly thicker than the rest and the internal rimoportula openings arecloser to the central part of the valve face (Table 3) The last feature as well as themarginal stria length enable also the morphological distinction between P balatonisand P comta
Ecology and distribution
The distinction between the species of the Puncticulata radiosa-complex includingPuncticulata balatonis is not easy for a plankton ecologist or other researcher whois not a centric diatom specialist The correct identification requires detailedmorphological studies with SEM andor specialized knowledge That is why apartfrom the long-lasting taxonomical confusion the real distribution of the speciesfrom the complex is uncertain According to Houk et al (2010) Puncticulata balatonis(as Cyclotella balatonis) is one of the most common members of the P radiosa (as
Fig 24 The number of Puncticulata balatonis frustules in the water column of Rusałka Lake in 2007
521
Cyclotella radiosa) complex in Europe found in numerous mesotrophic and eutrophiclakes and slowly running waters in Hungary Germany and Czech Republic Howeverreports of the species are not numerous Till now P balatonis has also never beenreported from Poland
The limited number of reports on the occurrence of P balatonis may be related to itsmisidentificationmisinterpretation as Cyclotella comta (Ehrenb) Kuumltz (Kiss amp Pająk1994 fig 13) found in the plankton of the eutrophic Vistula River as Cyclotellapraetermissa quadrijuncta (Wunsam et al 1995) or as Puncticulata radiosa (Wojtalamp Kwandrans 2006) found in the eutrophic Rudawa River We are convinced that apart of other records of P comta the most commonly reported species of the complexin Poland (Siemińska amp Wołowski 2003) in fact might concern P balatonis InRusałka Lake the species may have also been mistaken for P comta (asC comta) reported from the lake by Kotlińska (1976) It is also likely that P bala-tonis in Poland may have been misidentified as P praetermissa although the latterspecies was only reported several times including findings in the benthos of CzarneLake and the Kamionka River NE Poland (Rakowska 1996) in the plankton ofseveral lakes of the Drawa National Park NW Poland (Gołdyn amp Szeląg-Wasielewska2004) in a sand-pit lake in Krakoacutew and a spring fed pond in Krztynia in S Poland(Wojtal unpublished data)
Up to now there has also been much confusion around the ecology of the speciesfrom the complex For instance P praetermissa described by Lund (1951) fromlakes with a low organic matter content was later found both in the plankton and thebenthos in a wide variety of natural and man-made lakes including alpine andsubalpine lakes (Wunsam et al 1995 Lange-Bertalot amp Metzeltin 1996 Guumlttingeramp Straub 1998 Tolotti et al 2004) as well as in more eutrophic habitats (egSchmidt et al 2002 Hindaacutek amp Hindaacutekovaacute 2003 Tanaka 2007) Houk et al (2010)cleared up the ambiguities ascribing P praetermissa and P radiosa to oligo-mesotrophic habitats According to the authors P comta (as Cyclotella comta) is tillnow known only from fossil materials P balatonis (as Cyclotella balatonis) inturn has been assigned to meso-eutrophic lakes and rivers consistently with thetrophic character of its type locality ndash lake Balaton Our results correspond with theautecology of P balatonis (as Cyclotella balatonis) proposed by Houk et al (2010)widening even slightly the preferences of the species for the trophic conditionsdetermined by Houk et al (2010) as it was also able to develop a large populationin an eutrophic to hypereutrophic water body
The highest numbers of P balatonis in Rusałka Lake were observed in the winter-spring period The diatom bloom in the spring plankton is a well-known phenomenon(Sommer et al 1986) ndash sometimes also occurring during the winter period (Kiss ampGenkal 1993) ndash and is usually associated with a changing silica andor phosphorusavailability during that period (Lampert amp Sommer 2001) Sinking was also provedto be an essential factor hampering diatom success during the thermal stratificationperiod (Jewson et al 1981 Ferris amp Lehman 2007) explaining probably the seasonalityof the P balatonis growth in Rusałka Lake as its high development finished withthe stratification onset Relatively high numbers of frustules in deeper water layersmight also indicate a significant role of the sedimentation process
522
Conclusions
The morphology of Puncticulata balatonis in Rusałka Lake corresponds well toother species around P comta with the exception of the location of marginalfultoportula openings and the length of the shortened striae (notae) Analysis of themorphological characters enabled the transfer of Cyclotella balatonis into the genusPuncticulata A large population of the species in the eutrophic to hypereutrophicRusałka Lake remains congruent with its tolerance ranges with respect to the trophicstate proposed by Houk et al (2010) The variation in the cell number and averagedimension of frustules indicated the species preference to develop large populationsin spring
Acknowledgements
We thank Prof Ryszard Gołdyn (AMickiewicz University in Poznań) for his inspiration researchsupervision and useful comments to the manuscript We are also grateful to Dr Bart Van de Vijver andtwo anonymous reviewers for their valuable suggestions and to Dr Krisztina Buczko for enabling theanalysis of the type material of Cyclotella balatonis from the Pantocsekrsquos collection at HungarianNatural History Museum Sampling on Rusałka Lake was part of a project financially supported by theDistrict and Municipial Funds of Enviromental Protection and Water Management in Poznań (Poland)
References
CARLSON RE (1977) A trophic state index for lakes ndash Limnol Oceanogr 22 (2) 361ndash369
CREMER H amp H KOOLMEES (2010) Common diatoms (Centrales and Fragilariaceae Bacillario-phyta) of modern and fossil freshwater environments in the Netherlands ndash Nova Hedwigia 90(3ndash4)343ndash381
FERRIS JA amp JT LEHMAN (2007) Interannual variation in diatom bloom dynamics Roles ofhydrology nutrient limitation sinking and whole lake manipulation ndash Water Res 41 2551ndash2562
GENKAL SI (1977) On the method of calculation of some taxonomicaly significant structuralelements of the valve in the diatoms of the family Thalassiosiraceae Lebour emend Hasle (Bacillariophyta)ndash Bot Zhurn 62 848ndash851 (in Russian)
GENKAL SI amp KT KISS (1993) Morphological variability of the diatom Cyclotella atomusHustedt var atomus and C atomus var gracilis var nov ndash Hydrobiologia 269270 39ndash47
GOŁDYN R S PODSIADŁOWSKI K KOWALCZEWSKA-MADURA R DONDAJEWSKAE SZELĄG-WASIELEWSKA A BUDZYŃSKA P DOMEK amp W ROMANOWICZ (2010)Functioning of the Rusałka Lake ecosystem in Poznań (west Poland) ndash Ocean Hydrobiol Stud39(3) 65ndash80
GOŁDYN R amp E SZELĄG-WASIELEWSKA (2004) Changes in the phytoseston of a river-lakesystem in Drawieński National Park ndash Ocean Hydrobiol Stud 33(2) 17ndash28
GUumlTTINGER W amp F STRAUB (1998) Diatoms of Lake Cadagno ndash In PEDUZZI RR BACHOFEN amp M TONOLLA (eds) Lake Cadagno a meromictic Alpine lake ndash Doc Ist ItalIdrobiol 63 57ndash64
HANDMANN R (1913) Die Diatomeenflora des Almfeegebiets ndash In MANZ GJ (ed) Mitteilungendes Mikrologischen Vereins Linz ndash Mikrolog Ver Regensburg (in German)
HAUSMANN S amp AF LOTTER (2001) Morphological variation within the diatom taxon Cyclotellacomensis and its importance for quantitative temperature reconstructions ndash Freshw Biol 46 1323ndash1333
523
HAYASHI T (2007) Puncticulata versiformis sp nov and Cyclotella kathmanduensis sp nov(Bacillariophyta) new fossil species from middle Pleistocene lacustrine sediments Kathmandu NepalHilalaya ndash J Phycol 43(2) 304ndash318
HAringKANSSON H (2002) A compilation and evaluation of species in the genera StephanodiscusCyclostephanos and Cylotella with a new genus in the family Stephanodiscaceae ndash Diatom Res 17(1)1ndash139
HINDAacuteK F amp A HINDAacuteKOVAacute (2003) Diversity of cyanobacteria and algae of urban gravel pitlakes in Bratislava Slovakia a survey ndash Hydrobiologia 506ndash509 155ndash162
HINDAacuteK F amp A HINDAacuteKOVAacute (2008) Diversity of cyanobacteria and algae of the NationalNature Reserve Čiacutečovskeacute mŕtve rameno oxbow (Southern Slovakia) ndash Bulletin SBS 30 (1) 11ndash19 (inSlovak with an English abstract)
HOUK V R KLEE amp H TANAKA (2010) Atlas of freshwater centric diatoms with a brief key anddescriptions Part III Stephanodiscaceae A Cyclotella Tertiarius Discostella ndash Fottea 10 (Supplement)1ndash498
JEWSON DH BH RIPPEY amp WJK GILMORE (1981) Loss rates from sedimentation parasitismand grazing during the growth nutrient limitation and dormancy of a diatom crop ndash Limnol Oceanogr26 1045ndash1056
KISS KT amp SI GENKAL (1993) Winter blooms of centric diatoms in the River Danube and in itsside arms near Budapest ndash Hydrobiologia 269270 317ndash325
KISS KT amp G PAJĄK (1994) Seasonal change of diatom s in the plankton of the Vistula riverabove and below the Goczalkowice reservoir Poland ndash In KOCIOLEK JP (ed) Proceedings ofthe 11th International Diatom Symposium San Francisco Aug 12ndash17 1990 Memoirs of the CaliforniaAcademy of Sciences 17 517ndash524 San Francisco California Academy of Sciences
KOBAYASI H M IDEI S MAYAMA T NAGUMO amp K OSADA (2006) H KobayasirsquosAtlas of Japanese diatoms based on electron microscopy 1 1ndash531 Uchida Rokakuho Publishing CoLtd Tokyo
KOTLIŃSKA E (1976) Fitoplankton Jeziora Rusałka In Fitoplankton sztucznych jezior położonychna terenie Poznania 1ndash54 Poznańskie Tow Przyjacioacuteł Nauk Prace Komisji Biologicznej (in Polish)
KRAMMER K amp H LANGE-BERTALOT (1991) Bacillariophyceae 3 Teil Centrales Fragilaria-ceae Eunotiaceae ndash In ETTL H J GERLOFF H HEYNIGH amp D MOLLENHAUER (eds)Suumlsswasserflora von Mitteleuropa 2(3) 1ndash576 Gustav Fischer Verlag Stuttgart Jena
LAMPERT W amp U SOMMER (2001) Ekologia woacuted śroacutedlądowych ndash PWN Warszawa
LANGE-BERTALOT H amp D METZELTIN (1996) Indicators of oligotrophy 800 representativeof three ecologically distinct lake types Carbonate buffered-Oligotrophic-Weakly buffered soft waterndash Iconogr Diatomol 2 1ndash390
LUND JWG (1951) Contribution to our knowledge of British algae ndash Hydrobiologia 3(1) 93ndash100
ORGANIZATION FOR ECONOMIC COOPERATION AND DEVELOPMENT (1982)Eutrophication of Waters ndash Monitoring Assessment and Control Final Report OECD Paris
PANTOCSEK J (1901) Die Kieselalgen oder Bacillarien des Balaton 1ndash112 XVI Tafeln ndash Druckder K und K Hofbuchdruckerei des Victor Hornyaacutenszky Budapest
RAKOWSKA B (1996) Preliminary elaboration of the diatom communities occuring in the benthosof the Kamionka River and Lake Czarne at the Pojezierze Suwalskie (1993ndash1994) ndash Fragm FloristGeobot Polon 3 221ndash238
RIMET F JC DRUART amp O ANNEVILLE (2009) Exploring the dynamics of plankton diatomcommunities in Lake Geneva using emergent self-organizing maps (1974ndash2007) ndash Ecol Inform4 99ndash110
524
SCHMIDT R R PSENNER J MUumlLLER P INDINGER amp C KAMENIK (2002) Impact of thelate glacial climate variations on stratification and trophic state of the meromictic lake Laumlngsee (Austria)validation of a conceptual model by multiproxy studies ndash J Limnol 61(1) 49ndash60
SIEMIŃSKA J amp K WOŁOWSKI (2003) Catalogue of Polish prokaryotic and eukaryotic algaendash W Szafer Institute of Botany Polish Academy of Sciences Krakoacutew
SIVER PA (1999) Development of paleolimnological inference models for pH total nitrogen andspecific conductivity based on planktonic diatoms ndash J Paleolimnol 21 45ndash59
SOMMER U ZM GLIWICZ W LAMPERT amp A DUNCAN (1986) The PEG-model of seasonalsuccession of planktonic events in fresh waters ndash Arch Hydrobiol 106 433ndash471
TANAKA H (2007) Taxonomic studies of the genera Cyclotella (Kuumltzing) Breacutebisson DiscostellaHouk et Klee and Puncticulata Haringkansson in the family Stepanodiscaceae Glezer et Makarova(Bacillariophyta) in Japan ndash Biblioth Diatomol 53 1ndash205
TOLOTTI M D CALLIARI amp F CORRADINI (2004) Interannual variability of the phytoplanktonof Tovel lake (Trentino Italy) ndash Studi Trent di Scienze Naturali Acta Biol 81 Suppl 2 327ndash340 (inItalian)
WETZEL RG amp GE LIKENS (1991) Limnological analyses 1ndash391 ndash Springer-Verlag NewYork Berlin Heidelberg
WOJTAL AZ amp J KWANDRANS (2006) Diatoms of the Wyżyna Krakowsko-Częstochowskaupland (S Poland) ndash Coscinodiscophyceae (Thalassiosirophycidae) ndash Polish Bot J 51(2) 177ndash207
WUNSAM S R SCHMIDT amp R KLEE (1995) Cyclotella-taxa (Bacillariophyceae) in lakes of theAlpine region and their relationship to environmental variables ndash Aquat Sci 57(4) 360ndash381
Received 25 March 2010 accepted in revised form 31 March 2011

511
of 367 ha Although its maximum depth is 90 meters the average does not exceed 2 meters As aresult of the morphometry the lake is mainly polymictic with thermal stratification developing only ina small area around its deepest point in summer (Gołdyn et al 2010)
The lake is strongly eutrophic or hypereutrophic according to different criteria (Table 1) Cyanobacterialblooms occur in summer and autumn (Gołdyn et al 2010) Due to its location and surroundingattractive areas (mainly forests) the lake is intensely used for recreation
Materials and methods
Water samples for the phytoplankton analysis were taken at every meter depth at the deepest part of thelake in 2007 once a month from February till October and once in December Quantitative sampleswere preserved with Lugolrsquos solution and analysed with the use of inverted microscope PZO aftersedimentation in cylindrical chambers according to the Utermoumlhl method (Wetzel amp Likens 1991)
All diatoms from the family Stephanodiscaceae were counted as one taxon The relative frequency ofPuncticulata balatonis and other species belonging to this family was then estimated by counting 100frustules in hydrogene peroxide-treated samples using an Olympus BX-60 microscope Measurementsand the micrographs of live diatoms were done with the use of the same LM and AnalySIS programme
A qualitative analysis of the diatom material was performed after removing carbonates (addition of10 HCl to the samples followed by boiling taking place after 24 h for 15 minutes) and organic matter(boiling in 30 H
2O
2 with small amounts of KClO
3) After air-drying the material was mounted in
Naphraxreg Observations of the diatoms were performed with a Nikon Eclipse 80i light microscopeequipped with Differential Interference Contrast The micrographs were taken with a Nikon DS-Fi 1camera SEM analyses were performed on the material coated with gold using a Hitachi S-4700scanning electron microscope SEM micrographs were taken in the Laboratory of Electron and ConfocalMicroscopy at the Faculty of Biology Adam Mickiewicz University in Poznań and in the Laboratoryof Field Emission Scanning Electron Microscopy and Microanalysis at the Institute of GeologicalSciences of the Jagiellonian University in Krakoacutew Density of valve face structures (eg costae) in10 microm in a line tangential to the radius was calculated based on the formula in Genkal (1977)Additionally data on the morphology of Cyclotella balatonis were obtained from the type material(Balaton Lake Siofok samples 156 157) hosted in the Natural History Museum in Budapest (Hungary)
Table 1 Estimation of the trophic state of Rusałka Lake according to Carlson index (1977) and OECD(1982) criteria (acc to Gołdyn et al 2010)
Carlson indexTSI Values in Boundary values Trophic state
Rusałka lake for eutrophy
TSI (TP) 7437 gt70 HypertrophyTSI (ChL) 7171 53ndash70 HypertrophyTSI (SD) 6737 53ndash70 Eutrophy
OECDCharacteristic Values in Boundary values Trophic state
Rusałka lake for eutrophy
Mean TP (microgl) 96 gt100 EutrophyMean chlorophyll-a (microgl) 347 gt25 HypertrophyMax chlorophyll-a (microgl) 802 gt75 HypertrophyMean Secchi depth (m) 08 lt15 HypertrophyMin Secchi depth (m) 06 lt07 Hypertrophy
512
Water temperature pH and conductivity were measured in situ using YSI 610-DM-meter Watertransparency was expressed as the visibility of the Secchi disc For the statistical graphs Statistica 80software was used
Results
Morphology of Puncticulata balatonis in Rusałka Lake
Puncticulata balatonis (Pantocsek) Wojtal et Budzyńska comb nov Figs 1ndash12 amp 15ndash22
BASIONYM Cyclotella balatonis Pantocsek Die Kieselalgen oder Bacillarien des Balaton p 104 PlateXV figs 319 332 1901
TYPE LOCALITY Balaton Lake near Sioacutefok Hungary
Cells solitary or in short 2ndash7-cell chains (Fig 1) containing multiple discoidchloroplasts The valve diameter ranges between 87 and 210 microm Stria density is15ndash185 in 10 microm with 35ndash47 costae (thickened ribs) in 10 microm Striae are ofunequal length with inserted shorter (Figs 15ndash17) or divided striae (Fig 19) formingan irregularly circular central area (Figs 6 9 12 17) The central area is concentricallyslightly elevated (Figs 15 17) with numerous scattered openings of valve facefultoportulae (externally visible as minor puncta Fig 16) and areolae (externallyvisible as bigger puncta Figs 15ndash17) External openings of the rimopor-tulae arepresent at the end of the shortened striae (Figs 15 16) The internal opening of thesessile rimoportulae has a radially or slightly obliquely positioned slit (Figs 18 20)situated close to internal alveolar openings (towards the valve face margin) Marginalfultoportula openings with two satellite pores are situated (internally) on thethickened costae (shadow lines) separated by 3ndash5 thinner costae Valve facefultoportula openings are internally covered with domed cribra and surrounded bythree satellite pores (Figs 19 20) The morphology of the valve face internal alveolaropenings and the location of internal rimoportula openings correspond well withthe morphology of the specimens from the type material of Cyclotella balatonis(Figs 13 14 amp 21 22 Table 3)
Table 2 Physio-chemical parameters of water of Rusałka Lake in 2007
Temperature pH Coductivity Transparencyndash(degC) (microScm) Secchi depth (m)
20 II 31ndash41 72ndash78 836ndash841 1526 III 49ndash71 79ndash81 866ndash868 1820 IV 134ndash103 79ndash80 no data 1018 V 110ndash156 77ndash82 820ndash840 0819 VI 112ndash232 73ndash84 760ndash878 0918 VII 126ndash240 75ndash83 697ndash754 08520 VIII 145ndash220 75ndash91 627ndash739 0612 IX 144ndash148 83ndash85 646ndash652 0822 X 85ndash89 80ndash83 668ndash680 10503 XII 35ndash38 81ndash82 740ndash744 135
513
Ecological data
The water of Rusałka Lake was alkaline throughout the study period showing highconductivity Water transparency was relatively low ranging from 06 to 18 m ofSecchi depth (Table 2)
Puncticulata balatonis was present in the lake in all the seasons of 2007 It wasfound to form chains only in February March and April The diatom valve sizedistribution pattern varied in subsequent months with the largest average valve sizein February (Fig 23) Its abundance in the lake was also higher in the winter-springperiod (from February till May fig 24) with the maximum value of 35 times 103 cellsml-1 and a biomass of 298 mg l-1 (19 of the total phytoplankton cell number and525 of the total biomass at 6 m depth in March) In other spring samples
Figs 1ndash14 Puncticulata balatonis (Pantocsek) Wojtal et Budzyńska Light microscopy Figs 1ndash12 Specimens from the Rusałka Lake Figs 13ndash14 P balatonis type material sample no 152Fig 1 Short chain of alive cells in girdle view Figs 5ndash6 The same valve at different focus levels OnFigs 6 12ndash13 the characteristic gaps (notae) are marked with an arrow Scale bar represents 10 microm
514
Figs 15ndash22 Puncticulata balatonis (Pantocsek) Wojtal et Budzyńska Scanning electron microscopyFigs 15ndash20 specimens from the Rusałka Lake Figs 21ndash22 P balatonis type material samples no152 amp 156 Fig 15 external view of the rimoportula opening at the end of shortened striae ndash notae
515
P balatonis contributed between 63 and 187 to the total phytoplankton numbersand between 21 and 47 to the total biomass From June until October the numberof the diatom did not exceed 300 frustules per 1 ml (Fig 24) and no valve wasfound in September At the beginning of December P balatonis number increasedslightly again
At the time of the bloom of P balatonis chrysoflagellates (Erkenia subaequiciliataSkuja Chrysococcus triporus Matvienko) pennate diatoms (Fragilaria sp) andorcoenobia-forming Chlorococcales (Didymocystis inconspicua Koršikov Tetastrumsp div Scenedesmus sp div Coelastrum microporum Naumlgeli Crucigenia tetrapediaKirchner) were also abundant Other diatoms such as Aulacoseira ambigua (Grunow)Simonsen Cyclotella meneghiniana Kuumltzing C ocellata Pantocsek Cyclostephanosdelicatus (Genkal) Casper C dubius (Fricke) Round Stephanodiscus parvus Stoermeramp Haringkansson Asterionella formosa Hassal and Navicula sp were also observed
Discussion
Morphology of Puncticulata balatonis (Pant) Wojtal et Budzyńska
Puncticulata balatonis shares important morphological characters assigned to thegenus justifying its formal transfer to this genus a slightly concentrically undulatemiddle part of the valve face with scattered or radially arranged numerous areolaewith internally domed cribra and numerous fultoportulae Additionally valves ofP balatonis have internal alveolar openings with marginal fultoportulae located onthicker costae rimoportulae located externally at the top of shortened striae andinternally in the marginal part of the valve face
The morphology of P balatonis valves from Rusałka Lake corresponds well withthe morphology of the specimens from the Pantocsekrsquos type material (Figs 13 1421 22 Table 3) though the diameter range and the number of striae in 10 microm werefound to be different Pantocsek (1901) stated in his diagnosis (p 104) that thevalves of Cyclotella balatonis are 16ndash20 microm in diameter but we have found bothsmaller and larger specimens (Table 3) The wider diameter range in our observationcould be related to a more complete morphological range of the species since thediatom population in Rusałka Lake was abundant However the time of samplingmay have also been of importance as the medium valve size of the species in our
(arrowed) at concentrically elevated valve face Fig 16 external view of the openings of central (right)and marginal (left) fultoportulae (arrowed) Fig 17 external view of the short and long striae of themarginal area Fig18 internal view of the rimoportula opening (arrowed) located close to the valvemargin and internal alveolar opening and central openings of numerous areolae and fultoportulae(arrowed) Fig 19 internal view of the divided thinner costa (marked) and internal opening of centralfultoportula (arrowed) Fig 20 internal view of the marginal fultoportulae located on the thicker costae(arrowed) Fig 21 external view of the valve face with marked notae Fig 22 internal view of thealveolar openings with thicker costa bearing marginal fultoportulae and internal rimoportula openinglocated near the margin (arrowed) Scale bar represents 5 microm for Figs 15 18 amp 21 4 microm for Fig 163 microm for Figs 19 amp 20 and 2 microm for Figs 17 amp 22
516
Tab
le 3
Com
pari
son
of P
unct
icul
ata
bala
toni
s an
d m
orph
olog
ical
ly s
imila
r sp
ecie
s C
F ndash
cent
ral
fulto
port
ulae
MF
ndash m
argi
nal
fulto
port
ulae
o
urob
serv
atio
n ty
pe m
ater
ial S
iofoacute
k s
ampl
e no
152
our
obse
rvat
ion
base
d on
the
fig
62
11
12
in K
ram
mer
amp L
ange
-Ber
talo
t 199
1 w
hich
sho
ws
spec
imen
s fr
om th
e ty
pe m
ater
ial o
f Cyc
lote
lla
bala
toni
s Pa
ntoc
sek
(as
C r
adio
sa)
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
Ref
eren
ces
Pant
ocse
kH
ouk
et a
lL
und
1951
H
aringkan
sson
H
ouk
et a
lH
aringkan
sson
H
ouk
et a
lH
aringkan
sson
H
ouk
et a
l19
01 p
104
20
10 p
39
p 9
820
02 p
116
20
10 p
41
2002
p 1
142
010
p 3
720
02 p
113
20
10 p
36
Plat
e X
V
Plat
es 2
69
Figs
1 2
Figs
422
ndashPl
ates
280
Fi
gs 4
15ndash
Plat
es 2
61
Figs
410
ndash P
late
s 25
7 1
ndashFi
gs 3
19
1ndash11
270
42
61ndash
11 2
81
421
1ndash11
262
41
423
258
1ndash1
133
21ndash
6 2
71
1ndash6
1ndash6
1ndash12
263
25
9 1
ndash12
1ndash5
264
1ndash6
260
1ndash8
Col
onie
sno
dat
a2ndash
82ndash
7 ce
lls2ndash
8 ce
lls
shor
tC
ells
solit
ary
no d
ata
mos
tlyce
lls s
olita
ryno
dat
alo
osel
ylo
osel
ylo
osel
yor
in s
hort
solit
ary
rare
ly in
unite
dun
ited
unite
d ch
ains
loos
ely
unite
dsh
ort c
hain
sch
ains
2(8
)ce
llsD
iam
eter
16ndash2
016
ndash22
87ndash
210
8ndash25
8ndash25
8ndash30
7ndash35
7ndash25
8ndash50
5ndash32
(microm
)St
riae
20 p
er 1
0 microm
ca
16
per
15ndash1
85
per
13ndash1
9 pe
r13
ndash15
per
15ndash1
8 pe
r 10
no d
ata
15ndash1
8 pe
r 10
no d
ata
14ndash1
8 pe
r10
microm
of
10 micro
m10
microm
10 micro
mmicrom
of n
earl
ymicrom
of n
early
10 micro
mne
arly
equ
aleq
ual l
engt
heq
ual l
engt
hle
ngth
occ
upy
occu
py 1
2ndash1
4ta
king
abo
ut1
3ndash1
2 of
of th
e va
lvae
12
of th
e va
lve
valv
e fa
cedi
amet
er fa
ce d
iam
eter
diam
eter
517
Cen
tral
are
a
flat
tofla
t to
slig
htly
flat t
o sl
ight
lyce
ntra
l are
aco
ncen
trica
llyfla
t or s
light
lysl
ight
ly c
on-
dist
inct
ly c
on-f
lat t
o co
n-fl
at to
con
-sl
ight
ly c
on-
conc
entri
cally
conc
entri
cally
with
mor
e or
undu
late
co
ncen
trica
llyce
ntri
cally
cent
rica
llyce
ntri
cally
cent
rica
llyce
ntric
ally
un-
undu
late
un
dula
te
less
regu
larl
yw
ith sc
atte
red
undu
late
un
dula
te
undu
late
and
undu
late
un
dula
te
dula
te w
ithw
ith C
F an
dw
ith n
ume-
arra
nged
or ra
dial
lyce
ntra
lra
dial
lyra
dial
lysm
ooth
smoo
th C
Fnu
mer
ous C
Far
eola
ero
us C
F an
dra
dial
arra
nged
CF
areo
lae
and
wrin
kled
C
Fw
rinkl
ed C
Fan
d ar
eola
ean
d ar
eola
ear
rang
ed in
areo
lae
ope-
punc
taan
d ar
eola
eC
F sc
atte
red
and
areo
lae
and
areo
lae
scat
tere
d or
open
ings
mor
e or
less
ning
s sca
ttere
dor
arr
ange
dar
rang
ed in
alw
ays
arra
nged
insc
atte
red
orra
dial
row
sor
occ
asio
nal-
in ra
dial
radi
al p
atte
rnar
rang
ed in
radi
al ro
ws
arra
nged
inly
arr
ange
d in
row
sra
dial
row
sra
dial
pat
tern
radi
al p
atte
rn(F
igs
8 amp
9)
Rim
o-2
ndash3 in
the
(1)2
ndash5 in
the
1ndash2(
3) in
the
no d
ata
at le
ast o
ne
1ndash5
in th
eat
leas
t one
at
leas
t one
in
at le
ast o
ne
at le
ast o
ne
port
ulae
mar
gina
lm
argi
nal
mar
gina
l
Inte
rnal
lyon
the
valv
aem
argi
nal a
rea
on th
e va
lve
the
mar
gina
lon
the v
alva
eon
the
valv
aear
ea n
ear t
hear
ea n
ear t
hear
ea n
ear t
hea
sess
ile la
bi-
face
at t
hecl
ose
to li
mit
face
at t
hear
ea c
lose
tofa
ce a
t the
face
at t
heva
lve
mar
gin
valv
e mar
gin
valv
e mar
gin
um in
obl
ique
end
of s
hor-
of th
e ce
ntra
len
d of
sho
r-lim
it of
the
end
of s
hor-
end
of s
light
lyE
xter
nal o
pe-
Ext
erna
lE
xter
nal
posi
tion
lo-
tene
d st
ria
area
Ext
erna
lte
ned
stri
ace
ntra
l are
ate
ned
stri
ash
orte
ned
stri
ani
ngs
at th
eop
enin
gs a
top
enin
gs a
tca
ted
betw
een
Inte
rnal
ly a
open
ings
on
Inte
rnal
ly a
Ext
erna
l ope
-In
tern
ally
aIn
tern
ally
a la
rge
end
of s
hor-
the
end
ofth
e en
d of
alve
olar
sess
ile la
bi-
a hy
alin
e ar
eare
lativ
ely
ning
s on
ala
rge
sess
ilese
ssile
labi
umte
ned
stri
ash
orte
ned
shor
tene
dop
enin
gsum
in ra
dial
at th
e en
d of
larg
e se
ssile
hyal
ine
area
labi
um in
in o
bliq
ue o
rIn
tern
ally
ast
ria
Inte
r-st
ria
Inte
r-an
d ce
ntra
lpo
sitio
n lo
-a
slig
htly
labi
um in
slit
at th
e en
dob
lique
po-
circ
umfe
rent
ial
smal
l ses
sile
nally
a se
ssile
nally
a sm
all
area
cate
d m
id-
shor
tene
dpo
sitio
n lo
-of
a s
light
lysi
tion
loca
-po
sitio
n lo
cate
dla
bium
clo
sela
bium
ver
yse
ssile
labi
-w
ay b
etw
een
stria
Int
erna
l-ca
ted
betw
een
shor
tene
dte
d be
twee
nin
the
mar
gina
lto
inte
rnal
clos
e to
inte
r-um
clo
se to
alve
olar
ope
-ly
a s
essi
leal
veol
ar o
pe-
stri
a In
ter-
alve
olar
ope
-ar
ea n
ear t
o th
eal
veol
ar o
pe-
nal a
lveo
lar
inte
rnal
alv
e-ni
ngs
and
labi
um in
ning
s an
dna
lly a
ses
-ni
ngs
and
cent
ral a
rea
ning
s w
ithop
enin
gs
olar
ope
ning
sce
ntra
l are
aob
lique
to c
ir-ce
ntra
l are
asi
le la
bium
cent
ral a
rea
radi
ally
or
with
radi
ally
with
radi
ally
cum
fere
ntia
lw
ith a
slit
ori-
obliq
uely
po-
or o
bliq
uely
or o
bliq
uely
slit
posi
tion
ente
d ob
lique
-si
tione
d sl
itpo
sitio
ned
posi
tione
d sl
itly
to n
earl
ysl
itci
rcum
fere
n-tia
llyIn
tern
alsm
all
cost
aesm
all
cost
aesm
all
cost
ae
radi
ally
radi
ally
elon
-ra
dial
ly e
lon-
smal
l co
stae
smal
l co
stae
smal
l co
stae
smal
l co
stae
alve
olar
bear
ing
MF
bear
ing
MF
bear
ing
MF
elon
gate
dga
ted
cos
tae
gate
d c
osta
ebe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fop
enin
gscl
early
larg
ercl
early
larg
ercl
early
larg
erco
stae
bea
-be
arin
g M
Fbe
arin
g M
Fcl
earl
y la
rger
clea
rly la
rger
clea
rly la
rger
clea
rly
larg
erth
an th
in ri
bsth
an th
in ri
bsth
an th
in ri
bsri
ng M
Fcl
early
larg
ersl
ight
ly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
than
thin
ribs
clea
rly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
518
Lak
e B
alat
onM
esot
roph
ic
Eut
roph
ic
Plan
kton
of a
no d
ata
Plan
kton
or
no d
ata
Plan
kton
of
no d
ata
foss
il di
atom
near
Sioacute
fok
eutr
ophi
chy
pere
utro
-nu
mbe
r of
litto
ral o
ligo
olig
om
eso-
lake
s re
ser
ph
ic re
serv
oir
lake
s po
orm
esot
roph
ictr
ophi
csl
owly
run
-in
org
anic
lake
sm
ainl
y su
b-ni
ng w
ater
sm
atte
r and
alpi
ne la
kes
One
of t
heso
me
quar
rym
ost c
omm
onpo
ols
inE
urop
ean
Eng
lish
Lak
esp
ecie
s of
the
Dis
tric
tC
-rad
iosa
com
plex
Tab
le 3
con
tinue
d
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
519
samples declined in subsequent winter-spring months which is a typical consequenceof diatom cell division The number of striae in 10 microm was found to be lower (11 to17 striae in 10 microm) in some specimens from Rusałka Lake than has been originallystated by Pantocsek (1901) It might be related to the striae counting method becauseeven at original Pantocseks drawings (Plate XV figs 319 332) number of striae in10 microm is lower when the Genkalrsquos method (1977) is applied It also remains inaccordance with our observations of the type material (Table 3) and the observationsin Houk et al (2010)
Puncticulata balatonis is morphologically similar to several species of the complexmaking it difficult for a non-specialist to distinguish them The species resemblesP radiosa but differs from it in valve face ultrastructure such as the location of theinternal rimoportula openings The valves of P radiosa have a more coarse andradially wrinkled central area in the valve face whereas P balatonis has a smootherand finely silicified valve face Additionally the radial pattern of the valve facefultoportulae and areolae is very distinct in P radiosa The external rimoportulaopenings of P balatonis are situated at the end of shortened stria well discernible inLM as gaps (notae) whereas according to Houk et al (2010) the external openingsof P radiosa are situated at the end of only slightly shortened striae The differencesare clearly visible from the internal valvae view because P balatonis has valve facerimoportula openings located close to the internal alveolar openings whereas the
Fig 23 The variation in Puncticulata balatonis valve size in Rusałka Lake in subsequent months in2007
520
rimoportula openings in P radiosa are located more close to the central area ofthe valves Such a close location of the internal rimoportula openings to the valvemargin is typical only for P balatonis within the discussed group (Table 3)
Another diatom showing a similar morphology to Puncticulata balatonis is P prae-termissa (Lund) Haringkansson Most of the morphological features are similar forboth species (Table 3) but the later possesses radially elongated relatively largeinternal alveolar openings Moreover the costae bearing marginal fultoportulaeare only slightly thicker than the rest and the internal rimoportula openings arecloser to the central part of the valve face (Table 3) The last feature as well as themarginal stria length enable also the morphological distinction between P balatonisand P comta
Ecology and distribution
The distinction between the species of the Puncticulata radiosa-complex includingPuncticulata balatonis is not easy for a plankton ecologist or other researcher whois not a centric diatom specialist The correct identification requires detailedmorphological studies with SEM andor specialized knowledge That is why apartfrom the long-lasting taxonomical confusion the real distribution of the speciesfrom the complex is uncertain According to Houk et al (2010) Puncticulata balatonis(as Cyclotella balatonis) is one of the most common members of the P radiosa (as
Fig 24 The number of Puncticulata balatonis frustules in the water column of Rusałka Lake in 2007
521
Cyclotella radiosa) complex in Europe found in numerous mesotrophic and eutrophiclakes and slowly running waters in Hungary Germany and Czech Republic Howeverreports of the species are not numerous Till now P balatonis has also never beenreported from Poland
The limited number of reports on the occurrence of P balatonis may be related to itsmisidentificationmisinterpretation as Cyclotella comta (Ehrenb) Kuumltz (Kiss amp Pająk1994 fig 13) found in the plankton of the eutrophic Vistula River as Cyclotellapraetermissa quadrijuncta (Wunsam et al 1995) or as Puncticulata radiosa (Wojtalamp Kwandrans 2006) found in the eutrophic Rudawa River We are convinced that apart of other records of P comta the most commonly reported species of the complexin Poland (Siemińska amp Wołowski 2003) in fact might concern P balatonis InRusałka Lake the species may have also been mistaken for P comta (asC comta) reported from the lake by Kotlińska (1976) It is also likely that P bala-tonis in Poland may have been misidentified as P praetermissa although the latterspecies was only reported several times including findings in the benthos of CzarneLake and the Kamionka River NE Poland (Rakowska 1996) in the plankton ofseveral lakes of the Drawa National Park NW Poland (Gołdyn amp Szeląg-Wasielewska2004) in a sand-pit lake in Krakoacutew and a spring fed pond in Krztynia in S Poland(Wojtal unpublished data)
Up to now there has also been much confusion around the ecology of the speciesfrom the complex For instance P praetermissa described by Lund (1951) fromlakes with a low organic matter content was later found both in the plankton and thebenthos in a wide variety of natural and man-made lakes including alpine andsubalpine lakes (Wunsam et al 1995 Lange-Bertalot amp Metzeltin 1996 Guumlttingeramp Straub 1998 Tolotti et al 2004) as well as in more eutrophic habitats (egSchmidt et al 2002 Hindaacutek amp Hindaacutekovaacute 2003 Tanaka 2007) Houk et al (2010)cleared up the ambiguities ascribing P praetermissa and P radiosa to oligo-mesotrophic habitats According to the authors P comta (as Cyclotella comta) is tillnow known only from fossil materials P balatonis (as Cyclotella balatonis) inturn has been assigned to meso-eutrophic lakes and rivers consistently with thetrophic character of its type locality ndash lake Balaton Our results correspond with theautecology of P balatonis (as Cyclotella balatonis) proposed by Houk et al (2010)widening even slightly the preferences of the species for the trophic conditionsdetermined by Houk et al (2010) as it was also able to develop a large populationin an eutrophic to hypereutrophic water body
The highest numbers of P balatonis in Rusałka Lake were observed in the winter-spring period The diatom bloom in the spring plankton is a well-known phenomenon(Sommer et al 1986) ndash sometimes also occurring during the winter period (Kiss ampGenkal 1993) ndash and is usually associated with a changing silica andor phosphorusavailability during that period (Lampert amp Sommer 2001) Sinking was also provedto be an essential factor hampering diatom success during the thermal stratificationperiod (Jewson et al 1981 Ferris amp Lehman 2007) explaining probably the seasonalityof the P balatonis growth in Rusałka Lake as its high development finished withthe stratification onset Relatively high numbers of frustules in deeper water layersmight also indicate a significant role of the sedimentation process
522
Conclusions
The morphology of Puncticulata balatonis in Rusałka Lake corresponds well toother species around P comta with the exception of the location of marginalfultoportula openings and the length of the shortened striae (notae) Analysis of themorphological characters enabled the transfer of Cyclotella balatonis into the genusPuncticulata A large population of the species in the eutrophic to hypereutrophicRusałka Lake remains congruent with its tolerance ranges with respect to the trophicstate proposed by Houk et al (2010) The variation in the cell number and averagedimension of frustules indicated the species preference to develop large populationsin spring
Acknowledgements
We thank Prof Ryszard Gołdyn (AMickiewicz University in Poznań) for his inspiration researchsupervision and useful comments to the manuscript We are also grateful to Dr Bart Van de Vijver andtwo anonymous reviewers for their valuable suggestions and to Dr Krisztina Buczko for enabling theanalysis of the type material of Cyclotella balatonis from the Pantocsekrsquos collection at HungarianNatural History Museum Sampling on Rusałka Lake was part of a project financially supported by theDistrict and Municipial Funds of Enviromental Protection and Water Management in Poznań (Poland)
References
CARLSON RE (1977) A trophic state index for lakes ndash Limnol Oceanogr 22 (2) 361ndash369
CREMER H amp H KOOLMEES (2010) Common diatoms (Centrales and Fragilariaceae Bacillario-phyta) of modern and fossil freshwater environments in the Netherlands ndash Nova Hedwigia 90(3ndash4)343ndash381
FERRIS JA amp JT LEHMAN (2007) Interannual variation in diatom bloom dynamics Roles ofhydrology nutrient limitation sinking and whole lake manipulation ndash Water Res 41 2551ndash2562
GENKAL SI (1977) On the method of calculation of some taxonomicaly significant structuralelements of the valve in the diatoms of the family Thalassiosiraceae Lebour emend Hasle (Bacillariophyta)ndash Bot Zhurn 62 848ndash851 (in Russian)
GENKAL SI amp KT KISS (1993) Morphological variability of the diatom Cyclotella atomusHustedt var atomus and C atomus var gracilis var nov ndash Hydrobiologia 269270 39ndash47
GOŁDYN R S PODSIADŁOWSKI K KOWALCZEWSKA-MADURA R DONDAJEWSKAE SZELĄG-WASIELEWSKA A BUDZYŃSKA P DOMEK amp W ROMANOWICZ (2010)Functioning of the Rusałka Lake ecosystem in Poznań (west Poland) ndash Ocean Hydrobiol Stud39(3) 65ndash80
GOŁDYN R amp E SZELĄG-WASIELEWSKA (2004) Changes in the phytoseston of a river-lakesystem in Drawieński National Park ndash Ocean Hydrobiol Stud 33(2) 17ndash28
GUumlTTINGER W amp F STRAUB (1998) Diatoms of Lake Cadagno ndash In PEDUZZI RR BACHOFEN amp M TONOLLA (eds) Lake Cadagno a meromictic Alpine lake ndash Doc Ist ItalIdrobiol 63 57ndash64
HANDMANN R (1913) Die Diatomeenflora des Almfeegebiets ndash In MANZ GJ (ed) Mitteilungendes Mikrologischen Vereins Linz ndash Mikrolog Ver Regensburg (in German)
HAUSMANN S amp AF LOTTER (2001) Morphological variation within the diatom taxon Cyclotellacomensis and its importance for quantitative temperature reconstructions ndash Freshw Biol 46 1323ndash1333
523
HAYASHI T (2007) Puncticulata versiformis sp nov and Cyclotella kathmanduensis sp nov(Bacillariophyta) new fossil species from middle Pleistocene lacustrine sediments Kathmandu NepalHilalaya ndash J Phycol 43(2) 304ndash318
HAringKANSSON H (2002) A compilation and evaluation of species in the genera StephanodiscusCyclostephanos and Cylotella with a new genus in the family Stephanodiscaceae ndash Diatom Res 17(1)1ndash139
HINDAacuteK F amp A HINDAacuteKOVAacute (2003) Diversity of cyanobacteria and algae of urban gravel pitlakes in Bratislava Slovakia a survey ndash Hydrobiologia 506ndash509 155ndash162
HINDAacuteK F amp A HINDAacuteKOVAacute (2008) Diversity of cyanobacteria and algae of the NationalNature Reserve Čiacutečovskeacute mŕtve rameno oxbow (Southern Slovakia) ndash Bulletin SBS 30 (1) 11ndash19 (inSlovak with an English abstract)
HOUK V R KLEE amp H TANAKA (2010) Atlas of freshwater centric diatoms with a brief key anddescriptions Part III Stephanodiscaceae A Cyclotella Tertiarius Discostella ndash Fottea 10 (Supplement)1ndash498
JEWSON DH BH RIPPEY amp WJK GILMORE (1981) Loss rates from sedimentation parasitismand grazing during the growth nutrient limitation and dormancy of a diatom crop ndash Limnol Oceanogr26 1045ndash1056
KISS KT amp SI GENKAL (1993) Winter blooms of centric diatoms in the River Danube and in itsside arms near Budapest ndash Hydrobiologia 269270 317ndash325
KISS KT amp G PAJĄK (1994) Seasonal change of diatom s in the plankton of the Vistula riverabove and below the Goczalkowice reservoir Poland ndash In KOCIOLEK JP (ed) Proceedings ofthe 11th International Diatom Symposium San Francisco Aug 12ndash17 1990 Memoirs of the CaliforniaAcademy of Sciences 17 517ndash524 San Francisco California Academy of Sciences
KOBAYASI H M IDEI S MAYAMA T NAGUMO amp K OSADA (2006) H KobayasirsquosAtlas of Japanese diatoms based on electron microscopy 1 1ndash531 Uchida Rokakuho Publishing CoLtd Tokyo
KOTLIŃSKA E (1976) Fitoplankton Jeziora Rusałka In Fitoplankton sztucznych jezior położonychna terenie Poznania 1ndash54 Poznańskie Tow Przyjacioacuteł Nauk Prace Komisji Biologicznej (in Polish)
KRAMMER K amp H LANGE-BERTALOT (1991) Bacillariophyceae 3 Teil Centrales Fragilaria-ceae Eunotiaceae ndash In ETTL H J GERLOFF H HEYNIGH amp D MOLLENHAUER (eds)Suumlsswasserflora von Mitteleuropa 2(3) 1ndash576 Gustav Fischer Verlag Stuttgart Jena
LAMPERT W amp U SOMMER (2001) Ekologia woacuted śroacutedlądowych ndash PWN Warszawa
LANGE-BERTALOT H amp D METZELTIN (1996) Indicators of oligotrophy 800 representativeof three ecologically distinct lake types Carbonate buffered-Oligotrophic-Weakly buffered soft waterndash Iconogr Diatomol 2 1ndash390
LUND JWG (1951) Contribution to our knowledge of British algae ndash Hydrobiologia 3(1) 93ndash100
ORGANIZATION FOR ECONOMIC COOPERATION AND DEVELOPMENT (1982)Eutrophication of Waters ndash Monitoring Assessment and Control Final Report OECD Paris
PANTOCSEK J (1901) Die Kieselalgen oder Bacillarien des Balaton 1ndash112 XVI Tafeln ndash Druckder K und K Hofbuchdruckerei des Victor Hornyaacutenszky Budapest
RAKOWSKA B (1996) Preliminary elaboration of the diatom communities occuring in the benthosof the Kamionka River and Lake Czarne at the Pojezierze Suwalskie (1993ndash1994) ndash Fragm FloristGeobot Polon 3 221ndash238
RIMET F JC DRUART amp O ANNEVILLE (2009) Exploring the dynamics of plankton diatomcommunities in Lake Geneva using emergent self-organizing maps (1974ndash2007) ndash Ecol Inform4 99ndash110
524
SCHMIDT R R PSENNER J MUumlLLER P INDINGER amp C KAMENIK (2002) Impact of thelate glacial climate variations on stratification and trophic state of the meromictic lake Laumlngsee (Austria)validation of a conceptual model by multiproxy studies ndash J Limnol 61(1) 49ndash60
SIEMIŃSKA J amp K WOŁOWSKI (2003) Catalogue of Polish prokaryotic and eukaryotic algaendash W Szafer Institute of Botany Polish Academy of Sciences Krakoacutew
SIVER PA (1999) Development of paleolimnological inference models for pH total nitrogen andspecific conductivity based on planktonic diatoms ndash J Paleolimnol 21 45ndash59
SOMMER U ZM GLIWICZ W LAMPERT amp A DUNCAN (1986) The PEG-model of seasonalsuccession of planktonic events in fresh waters ndash Arch Hydrobiol 106 433ndash471
TANAKA H (2007) Taxonomic studies of the genera Cyclotella (Kuumltzing) Breacutebisson DiscostellaHouk et Klee and Puncticulata Haringkansson in the family Stepanodiscaceae Glezer et Makarova(Bacillariophyta) in Japan ndash Biblioth Diatomol 53 1ndash205
TOLOTTI M D CALLIARI amp F CORRADINI (2004) Interannual variability of the phytoplanktonof Tovel lake (Trentino Italy) ndash Studi Trent di Scienze Naturali Acta Biol 81 Suppl 2 327ndash340 (inItalian)
WETZEL RG amp GE LIKENS (1991) Limnological analyses 1ndash391 ndash Springer-Verlag NewYork Berlin Heidelberg
WOJTAL AZ amp J KWANDRANS (2006) Diatoms of the Wyżyna Krakowsko-Częstochowskaupland (S Poland) ndash Coscinodiscophyceae (Thalassiosirophycidae) ndash Polish Bot J 51(2) 177ndash207
WUNSAM S R SCHMIDT amp R KLEE (1995) Cyclotella-taxa (Bacillariophyceae) in lakes of theAlpine region and their relationship to environmental variables ndash Aquat Sci 57(4) 360ndash381
Received 25 March 2010 accepted in revised form 31 March 2011
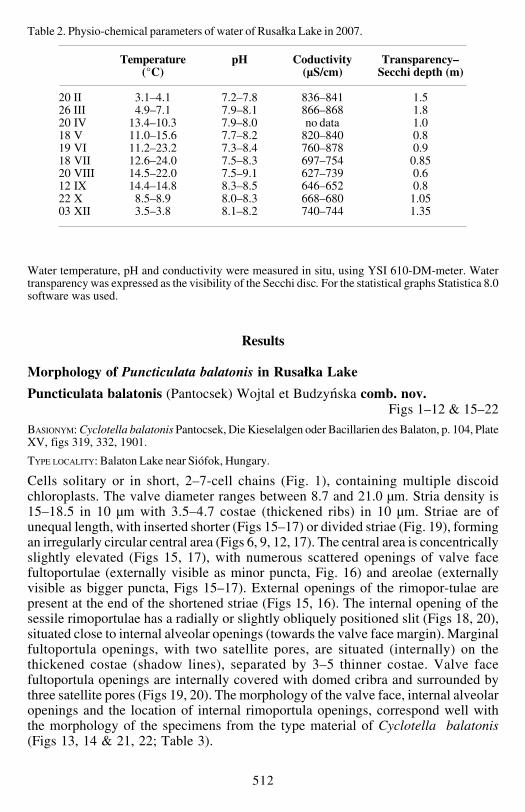
512
Water temperature pH and conductivity were measured in situ using YSI 610-DM-meter Watertransparency was expressed as the visibility of the Secchi disc For the statistical graphs Statistica 80software was used
Results
Morphology of Puncticulata balatonis in Rusałka Lake
Puncticulata balatonis (Pantocsek) Wojtal et Budzyńska comb nov Figs 1ndash12 amp 15ndash22
BASIONYM Cyclotella balatonis Pantocsek Die Kieselalgen oder Bacillarien des Balaton p 104 PlateXV figs 319 332 1901
TYPE LOCALITY Balaton Lake near Sioacutefok Hungary
Cells solitary or in short 2ndash7-cell chains (Fig 1) containing multiple discoidchloroplasts The valve diameter ranges between 87 and 210 microm Stria density is15ndash185 in 10 microm with 35ndash47 costae (thickened ribs) in 10 microm Striae are ofunequal length with inserted shorter (Figs 15ndash17) or divided striae (Fig 19) formingan irregularly circular central area (Figs 6 9 12 17) The central area is concentricallyslightly elevated (Figs 15 17) with numerous scattered openings of valve facefultoportulae (externally visible as minor puncta Fig 16) and areolae (externallyvisible as bigger puncta Figs 15ndash17) External openings of the rimopor-tulae arepresent at the end of the shortened striae (Figs 15 16) The internal opening of thesessile rimoportulae has a radially or slightly obliquely positioned slit (Figs 18 20)situated close to internal alveolar openings (towards the valve face margin) Marginalfultoportula openings with two satellite pores are situated (internally) on thethickened costae (shadow lines) separated by 3ndash5 thinner costae Valve facefultoportula openings are internally covered with domed cribra and surrounded bythree satellite pores (Figs 19 20) The morphology of the valve face internal alveolaropenings and the location of internal rimoportula openings correspond well withthe morphology of the specimens from the type material of Cyclotella balatonis(Figs 13 14 amp 21 22 Table 3)
Table 2 Physio-chemical parameters of water of Rusałka Lake in 2007
Temperature pH Coductivity Transparencyndash(degC) (microScm) Secchi depth (m)
20 II 31ndash41 72ndash78 836ndash841 1526 III 49ndash71 79ndash81 866ndash868 1820 IV 134ndash103 79ndash80 no data 1018 V 110ndash156 77ndash82 820ndash840 0819 VI 112ndash232 73ndash84 760ndash878 0918 VII 126ndash240 75ndash83 697ndash754 08520 VIII 145ndash220 75ndash91 627ndash739 0612 IX 144ndash148 83ndash85 646ndash652 0822 X 85ndash89 80ndash83 668ndash680 10503 XII 35ndash38 81ndash82 740ndash744 135
513
Ecological data
The water of Rusałka Lake was alkaline throughout the study period showing highconductivity Water transparency was relatively low ranging from 06 to 18 m ofSecchi depth (Table 2)
Puncticulata balatonis was present in the lake in all the seasons of 2007 It wasfound to form chains only in February March and April The diatom valve sizedistribution pattern varied in subsequent months with the largest average valve sizein February (Fig 23) Its abundance in the lake was also higher in the winter-springperiod (from February till May fig 24) with the maximum value of 35 times 103 cellsml-1 and a biomass of 298 mg l-1 (19 of the total phytoplankton cell number and525 of the total biomass at 6 m depth in March) In other spring samples
Figs 1ndash14 Puncticulata balatonis (Pantocsek) Wojtal et Budzyńska Light microscopy Figs 1ndash12 Specimens from the Rusałka Lake Figs 13ndash14 P balatonis type material sample no 152Fig 1 Short chain of alive cells in girdle view Figs 5ndash6 The same valve at different focus levels OnFigs 6 12ndash13 the characteristic gaps (notae) are marked with an arrow Scale bar represents 10 microm
514
Figs 15ndash22 Puncticulata balatonis (Pantocsek) Wojtal et Budzyńska Scanning electron microscopyFigs 15ndash20 specimens from the Rusałka Lake Figs 21ndash22 P balatonis type material samples no152 amp 156 Fig 15 external view of the rimoportula opening at the end of shortened striae ndash notae
515
P balatonis contributed between 63 and 187 to the total phytoplankton numbersand between 21 and 47 to the total biomass From June until October the numberof the diatom did not exceed 300 frustules per 1 ml (Fig 24) and no valve wasfound in September At the beginning of December P balatonis number increasedslightly again
At the time of the bloom of P balatonis chrysoflagellates (Erkenia subaequiciliataSkuja Chrysococcus triporus Matvienko) pennate diatoms (Fragilaria sp) andorcoenobia-forming Chlorococcales (Didymocystis inconspicua Koršikov Tetastrumsp div Scenedesmus sp div Coelastrum microporum Naumlgeli Crucigenia tetrapediaKirchner) were also abundant Other diatoms such as Aulacoseira ambigua (Grunow)Simonsen Cyclotella meneghiniana Kuumltzing C ocellata Pantocsek Cyclostephanosdelicatus (Genkal) Casper C dubius (Fricke) Round Stephanodiscus parvus Stoermeramp Haringkansson Asterionella formosa Hassal and Navicula sp were also observed
Discussion
Morphology of Puncticulata balatonis (Pant) Wojtal et Budzyńska
Puncticulata balatonis shares important morphological characters assigned to thegenus justifying its formal transfer to this genus a slightly concentrically undulatemiddle part of the valve face with scattered or radially arranged numerous areolaewith internally domed cribra and numerous fultoportulae Additionally valves ofP balatonis have internal alveolar openings with marginal fultoportulae located onthicker costae rimoportulae located externally at the top of shortened striae andinternally in the marginal part of the valve face
The morphology of P balatonis valves from Rusałka Lake corresponds well withthe morphology of the specimens from the Pantocsekrsquos type material (Figs 13 1421 22 Table 3) though the diameter range and the number of striae in 10 microm werefound to be different Pantocsek (1901) stated in his diagnosis (p 104) that thevalves of Cyclotella balatonis are 16ndash20 microm in diameter but we have found bothsmaller and larger specimens (Table 3) The wider diameter range in our observationcould be related to a more complete morphological range of the species since thediatom population in Rusałka Lake was abundant However the time of samplingmay have also been of importance as the medium valve size of the species in our
(arrowed) at concentrically elevated valve face Fig 16 external view of the openings of central (right)and marginal (left) fultoportulae (arrowed) Fig 17 external view of the short and long striae of themarginal area Fig18 internal view of the rimoportula opening (arrowed) located close to the valvemargin and internal alveolar opening and central openings of numerous areolae and fultoportulae(arrowed) Fig 19 internal view of the divided thinner costa (marked) and internal opening of centralfultoportula (arrowed) Fig 20 internal view of the marginal fultoportulae located on the thicker costae(arrowed) Fig 21 external view of the valve face with marked notae Fig 22 internal view of thealveolar openings with thicker costa bearing marginal fultoportulae and internal rimoportula openinglocated near the margin (arrowed) Scale bar represents 5 microm for Figs 15 18 amp 21 4 microm for Fig 163 microm for Figs 19 amp 20 and 2 microm for Figs 17 amp 22
516
Tab
le 3
Com
pari
son
of P
unct
icul
ata
bala
toni
s an
d m
orph
olog
ical
ly s
imila
r sp
ecie
s C
F ndash
cent
ral
fulto
port
ulae
MF
ndash m
argi
nal
fulto
port
ulae
o
urob
serv
atio
n ty
pe m
ater
ial S
iofoacute
k s
ampl
e no
152
our
obse
rvat
ion
base
d on
the
fig
62
11
12
in K
ram
mer
amp L
ange
-Ber
talo
t 199
1 w
hich
sho
ws
spec
imen
s fr
om th
e ty
pe m
ater
ial o
f Cyc
lote
lla
bala
toni
s Pa
ntoc
sek
(as
C r
adio
sa)
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
Ref
eren
ces
Pant
ocse
kH
ouk
et a
lL
und
1951
H
aringkan
sson
H
ouk
et a
lH
aringkan
sson
H
ouk
et a
lH
aringkan
sson
H
ouk
et a
l19
01 p
104
20
10 p
39
p 9
820
02 p
116
20
10 p
41
2002
p 1
142
010
p 3
720
02 p
113
20
10 p
36
Plat
e X
V
Plat
es 2
69
Figs
1 2
Figs
422
ndashPl
ates
280
Fi
gs 4
15ndash
Plat
es 2
61
Figs
410
ndash P
late
s 25
7 1
ndashFi
gs 3
19
1ndash11
270
42
61ndash
11 2
81
421
1ndash11
262
41
423
258
1ndash1
133
21ndash
6 2
71
1ndash6
1ndash6
1ndash12
263
25
9 1
ndash12
1ndash5
264
1ndash6
260
1ndash8
Col
onie
sno
dat
a2ndash
82ndash
7 ce
lls2ndash
8 ce
lls
shor
tC
ells
solit
ary
no d
ata
mos
tlyce
lls s
olita
ryno
dat
alo
osel
ylo
osel
ylo
osel
yor
in s
hort
solit
ary
rare
ly in
unite
dun
ited
unite
d ch
ains
loos
ely
unite
dsh
ort c
hain
sch
ains
2(8
)ce
llsD
iam
eter
16ndash2
016
ndash22
87ndash
210
8ndash25
8ndash25
8ndash30
7ndash35
7ndash25
8ndash50
5ndash32
(microm
)St
riae
20 p
er 1
0 microm
ca
16
per
15ndash1
85
per
13ndash1
9 pe
r13
ndash15
per
15ndash1
8 pe
r 10
no d
ata
15ndash1
8 pe
r 10
no d
ata
14ndash1
8 pe
r10
microm
of
10 micro
m10
microm
10 micro
mmicrom
of n
earl
ymicrom
of n
early
10 micro
mne
arly
equ
aleq
ual l
engt
heq
ual l
engt
hle
ngth
occ
upy
occu
py 1
2ndash1
4ta
king
abo
ut1
3ndash1
2 of
of th
e va
lvae
12
of th
e va
lve
valv
e fa
cedi
amet
er fa
ce d
iam
eter
diam
eter
517
Cen
tral
are
a
flat
tofla
t to
slig
htly
flat t
o sl
ight
lyce
ntra
l are
aco
ncen
trica
llyfla
t or s
light
lysl
ight
ly c
on-
dist
inct
ly c
on-f
lat t
o co
n-fl
at to
con
-sl
ight
ly c
on-
conc
entri
cally
conc
entri
cally
with
mor
e or
undu
late
co
ncen
trica
llyce
ntri
cally
cent
rica
llyce
ntri
cally
cent
rica
llyce
ntric
ally
un-
undu
late
un
dula
te
less
regu
larl
yw
ith sc
atte
red
undu
late
un
dula
te
undu
late
and
undu
late
un
dula
te
dula
te w
ithw
ith C
F an
dw
ith n
ume-
arra
nged
or ra
dial
lyce
ntra
lra
dial
lyra
dial
lysm
ooth
smoo
th C
Fnu
mer
ous C
Far
eola
ero
us C
F an
dra
dial
arra
nged
CF
areo
lae
and
wrin
kled
C
Fw
rinkl
ed C
Fan
d ar
eola
ean
d ar
eola
ear
rang
ed in
areo
lae
ope-
punc
taan
d ar
eola
eC
F sc
atte
red
and
areo
lae
and
areo
lae
scat
tere
d or
open
ings
mor
e or
less
ning
s sca
ttere
dor
arr
ange
dar
rang
ed in
alw
ays
arra
nged
insc
atte
red
orra
dial
row
sor
occ
asio
nal-
in ra
dial
radi
al p
atte
rnar
rang
ed in
radi
al ro
ws
arra
nged
inly
arr
ange
d in
row
sra
dial
row
sra
dial
pat
tern
radi
al p
atte
rn(F
igs
8 amp
9)
Rim
o-2
ndash3 in
the
(1)2
ndash5 in
the
1ndash2(
3) in
the
no d
ata
at le
ast o
ne
1ndash5
in th
eat
leas
t one
at
leas
t one
in
at le
ast o
ne
at le
ast o
ne
port
ulae
mar
gina
lm
argi
nal
mar
gina
l
Inte
rnal
lyon
the
valv
aem
argi
nal a
rea
on th
e va
lve
the
mar
gina
lon
the v
alva
eon
the
valv
aear
ea n
ear t
hear
ea n
ear t
hear
ea n
ear t
hea
sess
ile la
bi-
face
at t
hecl
ose
to li
mit
face
at t
hear
ea c
lose
tofa
ce a
t the
face
at t
heva
lve
mar
gin
valv
e mar
gin
valv
e mar
gin
um in
obl
ique
end
of s
hor-
of th
e ce
ntra
len
d of
sho
r-lim
it of
the
end
of s
hor-
end
of s
light
lyE
xter
nal o
pe-
Ext
erna
lE
xter
nal
posi
tion
lo-
tene
d st
ria
area
Ext
erna
lte
ned
stri
ace
ntra
l are
ate
ned
stri
ash
orte
ned
stri
ani
ngs
at th
eop
enin
gs a
top
enin
gs a
tca
ted
betw
een
Inte
rnal
ly a
open
ings
on
Inte
rnal
ly a
Ext
erna
l ope
-In
tern
ally
aIn
tern
ally
a la
rge
end
of s
hor-
the
end
ofth
e en
d of
alve
olar
sess
ile la
bi-
a hy
alin
e ar
eare
lativ
ely
ning
s on
ala
rge
sess
ilese
ssile
labi
umte
ned
stri
ash
orte
ned
shor
tene
dop
enin
gsum
in ra
dial
at th
e en
d of
larg
e se
ssile
hyal
ine
area
labi
um in
in o
bliq
ue o
rIn
tern
ally
ast
ria
Inte
r-st
ria
Inte
r-an
d ce
ntra
lpo
sitio
n lo
-a
slig
htly
labi
um in
slit
at th
e en
dob
lique
po-
circ
umfe
rent
ial
smal
l ses
sile
nally
a se
ssile
nally
a sm
all
area
cate
d m
id-
shor
tene
dpo
sitio
n lo
-of
a s
light
lysi
tion
loca
-po
sitio
n lo
cate
dla
bium
clo
sela
bium
ver
yse
ssile
labi
-w
ay b
etw
een
stria
Int
erna
l-ca
ted
betw
een
shor
tene
dte
d be
twee
nin
the
mar
gina
lto
inte
rnal
clos
e to
inte
r-um
clo
se to
alve
olar
ope
-ly
a s
essi
leal
veol
ar o
pe-
stri
a In
ter-
alve
olar
ope
-ar
ea n
ear t
o th
eal
veol
ar o
pe-
nal a
lveo
lar
inte
rnal
alv
e-ni
ngs
and
labi
um in
ning
s an
dna
lly a
ses
-ni
ngs
and
cent
ral a
rea
ning
s w
ithop
enin
gs
olar
ope
ning
sce
ntra
l are
aob
lique
to c
ir-ce
ntra
l are
asi
le la
bium
cent
ral a
rea
radi
ally
or
with
radi
ally
with
radi
ally
cum
fere
ntia
lw
ith a
slit
ori-
obliq
uely
po-
or o
bliq
uely
or o
bliq
uely
slit
posi
tion
ente
d ob
lique
-si
tione
d sl
itpo
sitio
ned
posi
tione
d sl
itly
to n
earl
ysl
itci
rcum
fere
n-tia
llyIn
tern
alsm
all
cost
aesm
all
cost
aesm
all
cost
ae
radi
ally
radi
ally
elon
-ra
dial
ly e
lon-
smal
l co
stae
smal
l co
stae
smal
l co
stae
smal
l co
stae
alve
olar
bear
ing
MF
bear
ing
MF
bear
ing
MF
elon
gate
dga
ted
cos
tae
gate
d c
osta
ebe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fop
enin
gscl
early
larg
ercl
early
larg
ercl
early
larg
erco
stae
bea
-be
arin
g M
Fbe
arin
g M
Fcl
earl
y la
rger
clea
rly la
rger
clea
rly la
rger
clea
rly
larg
erth
an th
in ri
bsth
an th
in ri
bsth
an th
in ri
bsri
ng M
Fcl
early
larg
ersl
ight
ly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
than
thin
ribs
clea
rly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
518
Lak
e B
alat
onM
esot
roph
ic
Eut
roph
ic
Plan
kton
of a
no d
ata
Plan
kton
or
no d
ata
Plan
kton
of
no d
ata
foss
il di
atom
near
Sioacute
fok
eutr
ophi
chy
pere
utro
-nu
mbe
r of
litto
ral o
ligo
olig
om
eso-
lake
s re
ser
ph
ic re
serv
oir
lake
s po
orm
esot
roph
ictr
ophi
csl
owly
run
-in
org
anic
lake
sm
ainl
y su
b-ni
ng w
ater
sm
atte
r and
alpi
ne la
kes
One
of t
heso
me
quar
rym
ost c
omm
onpo
ols
inE
urop
ean
Eng
lish
Lak
esp
ecie
s of
the
Dis
tric
tC
-rad
iosa
com
plex
Tab
le 3
con
tinue
d
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
519
samples declined in subsequent winter-spring months which is a typical consequenceof diatom cell division The number of striae in 10 microm was found to be lower (11 to17 striae in 10 microm) in some specimens from Rusałka Lake than has been originallystated by Pantocsek (1901) It might be related to the striae counting method becauseeven at original Pantocseks drawings (Plate XV figs 319 332) number of striae in10 microm is lower when the Genkalrsquos method (1977) is applied It also remains inaccordance with our observations of the type material (Table 3) and the observationsin Houk et al (2010)
Puncticulata balatonis is morphologically similar to several species of the complexmaking it difficult for a non-specialist to distinguish them The species resemblesP radiosa but differs from it in valve face ultrastructure such as the location of theinternal rimoportula openings The valves of P radiosa have a more coarse andradially wrinkled central area in the valve face whereas P balatonis has a smootherand finely silicified valve face Additionally the radial pattern of the valve facefultoportulae and areolae is very distinct in P radiosa The external rimoportulaopenings of P balatonis are situated at the end of shortened stria well discernible inLM as gaps (notae) whereas according to Houk et al (2010) the external openingsof P radiosa are situated at the end of only slightly shortened striae The differencesare clearly visible from the internal valvae view because P balatonis has valve facerimoportula openings located close to the internal alveolar openings whereas the
Fig 23 The variation in Puncticulata balatonis valve size in Rusałka Lake in subsequent months in2007
520
rimoportula openings in P radiosa are located more close to the central area ofthe valves Such a close location of the internal rimoportula openings to the valvemargin is typical only for P balatonis within the discussed group (Table 3)
Another diatom showing a similar morphology to Puncticulata balatonis is P prae-termissa (Lund) Haringkansson Most of the morphological features are similar forboth species (Table 3) but the later possesses radially elongated relatively largeinternal alveolar openings Moreover the costae bearing marginal fultoportulaeare only slightly thicker than the rest and the internal rimoportula openings arecloser to the central part of the valve face (Table 3) The last feature as well as themarginal stria length enable also the morphological distinction between P balatonisand P comta
Ecology and distribution
The distinction between the species of the Puncticulata radiosa-complex includingPuncticulata balatonis is not easy for a plankton ecologist or other researcher whois not a centric diatom specialist The correct identification requires detailedmorphological studies with SEM andor specialized knowledge That is why apartfrom the long-lasting taxonomical confusion the real distribution of the speciesfrom the complex is uncertain According to Houk et al (2010) Puncticulata balatonis(as Cyclotella balatonis) is one of the most common members of the P radiosa (as
Fig 24 The number of Puncticulata balatonis frustules in the water column of Rusałka Lake in 2007
521
Cyclotella radiosa) complex in Europe found in numerous mesotrophic and eutrophiclakes and slowly running waters in Hungary Germany and Czech Republic Howeverreports of the species are not numerous Till now P balatonis has also never beenreported from Poland
The limited number of reports on the occurrence of P balatonis may be related to itsmisidentificationmisinterpretation as Cyclotella comta (Ehrenb) Kuumltz (Kiss amp Pająk1994 fig 13) found in the plankton of the eutrophic Vistula River as Cyclotellapraetermissa quadrijuncta (Wunsam et al 1995) or as Puncticulata radiosa (Wojtalamp Kwandrans 2006) found in the eutrophic Rudawa River We are convinced that apart of other records of P comta the most commonly reported species of the complexin Poland (Siemińska amp Wołowski 2003) in fact might concern P balatonis InRusałka Lake the species may have also been mistaken for P comta (asC comta) reported from the lake by Kotlińska (1976) It is also likely that P bala-tonis in Poland may have been misidentified as P praetermissa although the latterspecies was only reported several times including findings in the benthos of CzarneLake and the Kamionka River NE Poland (Rakowska 1996) in the plankton ofseveral lakes of the Drawa National Park NW Poland (Gołdyn amp Szeląg-Wasielewska2004) in a sand-pit lake in Krakoacutew and a spring fed pond in Krztynia in S Poland(Wojtal unpublished data)
Up to now there has also been much confusion around the ecology of the speciesfrom the complex For instance P praetermissa described by Lund (1951) fromlakes with a low organic matter content was later found both in the plankton and thebenthos in a wide variety of natural and man-made lakes including alpine andsubalpine lakes (Wunsam et al 1995 Lange-Bertalot amp Metzeltin 1996 Guumlttingeramp Straub 1998 Tolotti et al 2004) as well as in more eutrophic habitats (egSchmidt et al 2002 Hindaacutek amp Hindaacutekovaacute 2003 Tanaka 2007) Houk et al (2010)cleared up the ambiguities ascribing P praetermissa and P radiosa to oligo-mesotrophic habitats According to the authors P comta (as Cyclotella comta) is tillnow known only from fossil materials P balatonis (as Cyclotella balatonis) inturn has been assigned to meso-eutrophic lakes and rivers consistently with thetrophic character of its type locality ndash lake Balaton Our results correspond with theautecology of P balatonis (as Cyclotella balatonis) proposed by Houk et al (2010)widening even slightly the preferences of the species for the trophic conditionsdetermined by Houk et al (2010) as it was also able to develop a large populationin an eutrophic to hypereutrophic water body
The highest numbers of P balatonis in Rusałka Lake were observed in the winter-spring period The diatom bloom in the spring plankton is a well-known phenomenon(Sommer et al 1986) ndash sometimes also occurring during the winter period (Kiss ampGenkal 1993) ndash and is usually associated with a changing silica andor phosphorusavailability during that period (Lampert amp Sommer 2001) Sinking was also provedto be an essential factor hampering diatom success during the thermal stratificationperiod (Jewson et al 1981 Ferris amp Lehman 2007) explaining probably the seasonalityof the P balatonis growth in Rusałka Lake as its high development finished withthe stratification onset Relatively high numbers of frustules in deeper water layersmight also indicate a significant role of the sedimentation process
522
Conclusions
The morphology of Puncticulata balatonis in Rusałka Lake corresponds well toother species around P comta with the exception of the location of marginalfultoportula openings and the length of the shortened striae (notae) Analysis of themorphological characters enabled the transfer of Cyclotella balatonis into the genusPuncticulata A large population of the species in the eutrophic to hypereutrophicRusałka Lake remains congruent with its tolerance ranges with respect to the trophicstate proposed by Houk et al (2010) The variation in the cell number and averagedimension of frustules indicated the species preference to develop large populationsin spring
Acknowledgements
We thank Prof Ryszard Gołdyn (AMickiewicz University in Poznań) for his inspiration researchsupervision and useful comments to the manuscript We are also grateful to Dr Bart Van de Vijver andtwo anonymous reviewers for their valuable suggestions and to Dr Krisztina Buczko for enabling theanalysis of the type material of Cyclotella balatonis from the Pantocsekrsquos collection at HungarianNatural History Museum Sampling on Rusałka Lake was part of a project financially supported by theDistrict and Municipial Funds of Enviromental Protection and Water Management in Poznań (Poland)
References
CARLSON RE (1977) A trophic state index for lakes ndash Limnol Oceanogr 22 (2) 361ndash369
CREMER H amp H KOOLMEES (2010) Common diatoms (Centrales and Fragilariaceae Bacillario-phyta) of modern and fossil freshwater environments in the Netherlands ndash Nova Hedwigia 90(3ndash4)343ndash381
FERRIS JA amp JT LEHMAN (2007) Interannual variation in diatom bloom dynamics Roles ofhydrology nutrient limitation sinking and whole lake manipulation ndash Water Res 41 2551ndash2562
GENKAL SI (1977) On the method of calculation of some taxonomicaly significant structuralelements of the valve in the diatoms of the family Thalassiosiraceae Lebour emend Hasle (Bacillariophyta)ndash Bot Zhurn 62 848ndash851 (in Russian)
GENKAL SI amp KT KISS (1993) Morphological variability of the diatom Cyclotella atomusHustedt var atomus and C atomus var gracilis var nov ndash Hydrobiologia 269270 39ndash47
GOŁDYN R S PODSIADŁOWSKI K KOWALCZEWSKA-MADURA R DONDAJEWSKAE SZELĄG-WASIELEWSKA A BUDZYŃSKA P DOMEK amp W ROMANOWICZ (2010)Functioning of the Rusałka Lake ecosystem in Poznań (west Poland) ndash Ocean Hydrobiol Stud39(3) 65ndash80
GOŁDYN R amp E SZELĄG-WASIELEWSKA (2004) Changes in the phytoseston of a river-lakesystem in Drawieński National Park ndash Ocean Hydrobiol Stud 33(2) 17ndash28
GUumlTTINGER W amp F STRAUB (1998) Diatoms of Lake Cadagno ndash In PEDUZZI RR BACHOFEN amp M TONOLLA (eds) Lake Cadagno a meromictic Alpine lake ndash Doc Ist ItalIdrobiol 63 57ndash64
HANDMANN R (1913) Die Diatomeenflora des Almfeegebiets ndash In MANZ GJ (ed) Mitteilungendes Mikrologischen Vereins Linz ndash Mikrolog Ver Regensburg (in German)
HAUSMANN S amp AF LOTTER (2001) Morphological variation within the diatom taxon Cyclotellacomensis and its importance for quantitative temperature reconstructions ndash Freshw Biol 46 1323ndash1333
523
HAYASHI T (2007) Puncticulata versiformis sp nov and Cyclotella kathmanduensis sp nov(Bacillariophyta) new fossil species from middle Pleistocene lacustrine sediments Kathmandu NepalHilalaya ndash J Phycol 43(2) 304ndash318
HAringKANSSON H (2002) A compilation and evaluation of species in the genera StephanodiscusCyclostephanos and Cylotella with a new genus in the family Stephanodiscaceae ndash Diatom Res 17(1)1ndash139
HINDAacuteK F amp A HINDAacuteKOVAacute (2003) Diversity of cyanobacteria and algae of urban gravel pitlakes in Bratislava Slovakia a survey ndash Hydrobiologia 506ndash509 155ndash162
HINDAacuteK F amp A HINDAacuteKOVAacute (2008) Diversity of cyanobacteria and algae of the NationalNature Reserve Čiacutečovskeacute mŕtve rameno oxbow (Southern Slovakia) ndash Bulletin SBS 30 (1) 11ndash19 (inSlovak with an English abstract)
HOUK V R KLEE amp H TANAKA (2010) Atlas of freshwater centric diatoms with a brief key anddescriptions Part III Stephanodiscaceae A Cyclotella Tertiarius Discostella ndash Fottea 10 (Supplement)1ndash498
JEWSON DH BH RIPPEY amp WJK GILMORE (1981) Loss rates from sedimentation parasitismand grazing during the growth nutrient limitation and dormancy of a diatom crop ndash Limnol Oceanogr26 1045ndash1056
KISS KT amp SI GENKAL (1993) Winter blooms of centric diatoms in the River Danube and in itsside arms near Budapest ndash Hydrobiologia 269270 317ndash325
KISS KT amp G PAJĄK (1994) Seasonal change of diatom s in the plankton of the Vistula riverabove and below the Goczalkowice reservoir Poland ndash In KOCIOLEK JP (ed) Proceedings ofthe 11th International Diatom Symposium San Francisco Aug 12ndash17 1990 Memoirs of the CaliforniaAcademy of Sciences 17 517ndash524 San Francisco California Academy of Sciences
KOBAYASI H M IDEI S MAYAMA T NAGUMO amp K OSADA (2006) H KobayasirsquosAtlas of Japanese diatoms based on electron microscopy 1 1ndash531 Uchida Rokakuho Publishing CoLtd Tokyo
KOTLIŃSKA E (1976) Fitoplankton Jeziora Rusałka In Fitoplankton sztucznych jezior położonychna terenie Poznania 1ndash54 Poznańskie Tow Przyjacioacuteł Nauk Prace Komisji Biologicznej (in Polish)
KRAMMER K amp H LANGE-BERTALOT (1991) Bacillariophyceae 3 Teil Centrales Fragilaria-ceae Eunotiaceae ndash In ETTL H J GERLOFF H HEYNIGH amp D MOLLENHAUER (eds)Suumlsswasserflora von Mitteleuropa 2(3) 1ndash576 Gustav Fischer Verlag Stuttgart Jena
LAMPERT W amp U SOMMER (2001) Ekologia woacuted śroacutedlądowych ndash PWN Warszawa
LANGE-BERTALOT H amp D METZELTIN (1996) Indicators of oligotrophy 800 representativeof three ecologically distinct lake types Carbonate buffered-Oligotrophic-Weakly buffered soft waterndash Iconogr Diatomol 2 1ndash390
LUND JWG (1951) Contribution to our knowledge of British algae ndash Hydrobiologia 3(1) 93ndash100
ORGANIZATION FOR ECONOMIC COOPERATION AND DEVELOPMENT (1982)Eutrophication of Waters ndash Monitoring Assessment and Control Final Report OECD Paris
PANTOCSEK J (1901) Die Kieselalgen oder Bacillarien des Balaton 1ndash112 XVI Tafeln ndash Druckder K und K Hofbuchdruckerei des Victor Hornyaacutenszky Budapest
RAKOWSKA B (1996) Preliminary elaboration of the diatom communities occuring in the benthosof the Kamionka River and Lake Czarne at the Pojezierze Suwalskie (1993ndash1994) ndash Fragm FloristGeobot Polon 3 221ndash238
RIMET F JC DRUART amp O ANNEVILLE (2009) Exploring the dynamics of plankton diatomcommunities in Lake Geneva using emergent self-organizing maps (1974ndash2007) ndash Ecol Inform4 99ndash110
524
SCHMIDT R R PSENNER J MUumlLLER P INDINGER amp C KAMENIK (2002) Impact of thelate glacial climate variations on stratification and trophic state of the meromictic lake Laumlngsee (Austria)validation of a conceptual model by multiproxy studies ndash J Limnol 61(1) 49ndash60
SIEMIŃSKA J amp K WOŁOWSKI (2003) Catalogue of Polish prokaryotic and eukaryotic algaendash W Szafer Institute of Botany Polish Academy of Sciences Krakoacutew
SIVER PA (1999) Development of paleolimnological inference models for pH total nitrogen andspecific conductivity based on planktonic diatoms ndash J Paleolimnol 21 45ndash59
SOMMER U ZM GLIWICZ W LAMPERT amp A DUNCAN (1986) The PEG-model of seasonalsuccession of planktonic events in fresh waters ndash Arch Hydrobiol 106 433ndash471
TANAKA H (2007) Taxonomic studies of the genera Cyclotella (Kuumltzing) Breacutebisson DiscostellaHouk et Klee and Puncticulata Haringkansson in the family Stepanodiscaceae Glezer et Makarova(Bacillariophyta) in Japan ndash Biblioth Diatomol 53 1ndash205
TOLOTTI M D CALLIARI amp F CORRADINI (2004) Interannual variability of the phytoplanktonof Tovel lake (Trentino Italy) ndash Studi Trent di Scienze Naturali Acta Biol 81 Suppl 2 327ndash340 (inItalian)
WETZEL RG amp GE LIKENS (1991) Limnological analyses 1ndash391 ndash Springer-Verlag NewYork Berlin Heidelberg
WOJTAL AZ amp J KWANDRANS (2006) Diatoms of the Wyżyna Krakowsko-Częstochowskaupland (S Poland) ndash Coscinodiscophyceae (Thalassiosirophycidae) ndash Polish Bot J 51(2) 177ndash207
WUNSAM S R SCHMIDT amp R KLEE (1995) Cyclotella-taxa (Bacillariophyceae) in lakes of theAlpine region and their relationship to environmental variables ndash Aquat Sci 57(4) 360ndash381
Received 25 March 2010 accepted in revised form 31 March 2011
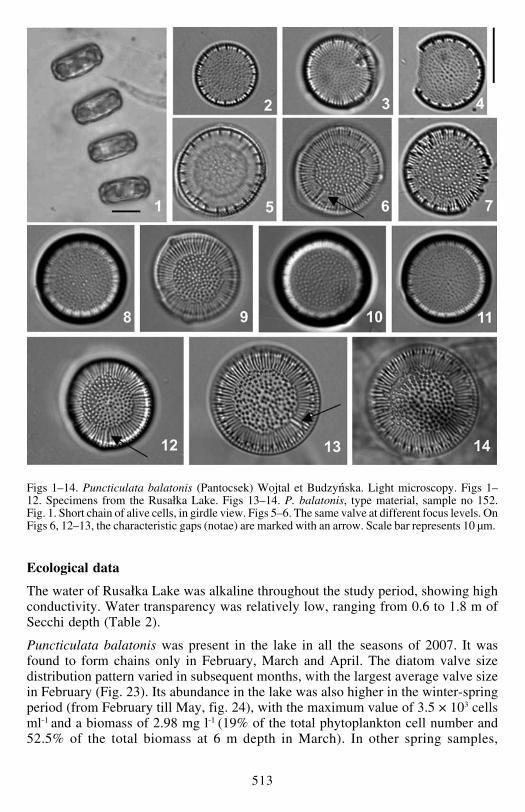
513
Ecological data
The water of Rusałka Lake was alkaline throughout the study period showing highconductivity Water transparency was relatively low ranging from 06 to 18 m ofSecchi depth (Table 2)
Puncticulata balatonis was present in the lake in all the seasons of 2007 It wasfound to form chains only in February March and April The diatom valve sizedistribution pattern varied in subsequent months with the largest average valve sizein February (Fig 23) Its abundance in the lake was also higher in the winter-springperiod (from February till May fig 24) with the maximum value of 35 times 103 cellsml-1 and a biomass of 298 mg l-1 (19 of the total phytoplankton cell number and525 of the total biomass at 6 m depth in March) In other spring samples
Figs 1ndash14 Puncticulata balatonis (Pantocsek) Wojtal et Budzyńska Light microscopy Figs 1ndash12 Specimens from the Rusałka Lake Figs 13ndash14 P balatonis type material sample no 152Fig 1 Short chain of alive cells in girdle view Figs 5ndash6 The same valve at different focus levels OnFigs 6 12ndash13 the characteristic gaps (notae) are marked with an arrow Scale bar represents 10 microm
514
Figs 15ndash22 Puncticulata balatonis (Pantocsek) Wojtal et Budzyńska Scanning electron microscopyFigs 15ndash20 specimens from the Rusałka Lake Figs 21ndash22 P balatonis type material samples no152 amp 156 Fig 15 external view of the rimoportula opening at the end of shortened striae ndash notae
515
P balatonis contributed between 63 and 187 to the total phytoplankton numbersand between 21 and 47 to the total biomass From June until October the numberof the diatom did not exceed 300 frustules per 1 ml (Fig 24) and no valve wasfound in September At the beginning of December P balatonis number increasedslightly again
At the time of the bloom of P balatonis chrysoflagellates (Erkenia subaequiciliataSkuja Chrysococcus triporus Matvienko) pennate diatoms (Fragilaria sp) andorcoenobia-forming Chlorococcales (Didymocystis inconspicua Koršikov Tetastrumsp div Scenedesmus sp div Coelastrum microporum Naumlgeli Crucigenia tetrapediaKirchner) were also abundant Other diatoms such as Aulacoseira ambigua (Grunow)Simonsen Cyclotella meneghiniana Kuumltzing C ocellata Pantocsek Cyclostephanosdelicatus (Genkal) Casper C dubius (Fricke) Round Stephanodiscus parvus Stoermeramp Haringkansson Asterionella formosa Hassal and Navicula sp were also observed
Discussion
Morphology of Puncticulata balatonis (Pant) Wojtal et Budzyńska
Puncticulata balatonis shares important morphological characters assigned to thegenus justifying its formal transfer to this genus a slightly concentrically undulatemiddle part of the valve face with scattered or radially arranged numerous areolaewith internally domed cribra and numerous fultoportulae Additionally valves ofP balatonis have internal alveolar openings with marginal fultoportulae located onthicker costae rimoportulae located externally at the top of shortened striae andinternally in the marginal part of the valve face
The morphology of P balatonis valves from Rusałka Lake corresponds well withthe morphology of the specimens from the Pantocsekrsquos type material (Figs 13 1421 22 Table 3) though the diameter range and the number of striae in 10 microm werefound to be different Pantocsek (1901) stated in his diagnosis (p 104) that thevalves of Cyclotella balatonis are 16ndash20 microm in diameter but we have found bothsmaller and larger specimens (Table 3) The wider diameter range in our observationcould be related to a more complete morphological range of the species since thediatom population in Rusałka Lake was abundant However the time of samplingmay have also been of importance as the medium valve size of the species in our
(arrowed) at concentrically elevated valve face Fig 16 external view of the openings of central (right)and marginal (left) fultoportulae (arrowed) Fig 17 external view of the short and long striae of themarginal area Fig18 internal view of the rimoportula opening (arrowed) located close to the valvemargin and internal alveolar opening and central openings of numerous areolae and fultoportulae(arrowed) Fig 19 internal view of the divided thinner costa (marked) and internal opening of centralfultoportula (arrowed) Fig 20 internal view of the marginal fultoportulae located on the thicker costae(arrowed) Fig 21 external view of the valve face with marked notae Fig 22 internal view of thealveolar openings with thicker costa bearing marginal fultoportulae and internal rimoportula openinglocated near the margin (arrowed) Scale bar represents 5 microm for Figs 15 18 amp 21 4 microm for Fig 163 microm for Figs 19 amp 20 and 2 microm for Figs 17 amp 22
516
Tab
le 3
Com
pari
son
of P
unct
icul
ata
bala
toni
s an
d m
orph
olog
ical
ly s
imila
r sp
ecie
s C
F ndash
cent
ral
fulto
port
ulae
MF
ndash m
argi
nal
fulto
port
ulae
o
urob
serv
atio
n ty
pe m
ater
ial S
iofoacute
k s
ampl
e no
152
our
obse
rvat
ion
base
d on
the
fig
62
11
12
in K
ram
mer
amp L
ange
-Ber
talo
t 199
1 w
hich
sho
ws
spec
imen
s fr
om th
e ty
pe m
ater
ial o
f Cyc
lote
lla
bala
toni
s Pa
ntoc
sek
(as
C r
adio
sa)
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
Ref
eren
ces
Pant
ocse
kH
ouk
et a
lL
und
1951
H
aringkan
sson
H
ouk
et a
lH
aringkan
sson
H
ouk
et a
lH
aringkan
sson
H
ouk
et a
l19
01 p
104
20
10 p
39
p 9
820
02 p
116
20
10 p
41
2002
p 1
142
010
p 3
720
02 p
113
20
10 p
36
Plat
e X
V
Plat
es 2
69
Figs
1 2
Figs
422
ndashPl
ates
280
Fi
gs 4
15ndash
Plat
es 2
61
Figs
410
ndash P
late
s 25
7 1
ndashFi
gs 3
19
1ndash11
270
42
61ndash
11 2
81
421
1ndash11
262
41
423
258
1ndash1
133
21ndash
6 2
71
1ndash6
1ndash6
1ndash12
263
25
9 1
ndash12
1ndash5
264
1ndash6
260
1ndash8
Col
onie
sno
dat
a2ndash
82ndash
7 ce
lls2ndash
8 ce
lls
shor
tC
ells
solit
ary
no d
ata
mos
tlyce
lls s
olita
ryno
dat
alo
osel
ylo
osel
ylo
osel
yor
in s
hort
solit
ary
rare
ly in
unite
dun
ited
unite
d ch
ains
loos
ely
unite
dsh
ort c
hain
sch
ains
2(8
)ce
llsD
iam
eter
16ndash2
016
ndash22
87ndash
210
8ndash25
8ndash25
8ndash30
7ndash35
7ndash25
8ndash50
5ndash32
(microm
)St
riae
20 p
er 1
0 microm
ca
16
per
15ndash1
85
per
13ndash1
9 pe
r13
ndash15
per
15ndash1
8 pe
r 10
no d
ata
15ndash1
8 pe
r 10
no d
ata
14ndash1
8 pe
r10
microm
of
10 micro
m10
microm
10 micro
mmicrom
of n
earl
ymicrom
of n
early
10 micro
mne
arly
equ
aleq
ual l
engt
heq
ual l
engt
hle
ngth
occ
upy
occu
py 1
2ndash1
4ta
king
abo
ut1
3ndash1
2 of
of th
e va
lvae
12
of th
e va
lve
valv
e fa
cedi
amet
er fa
ce d
iam
eter
diam
eter
517
Cen
tral
are
a
flat
tofla
t to
slig
htly
flat t
o sl
ight
lyce
ntra
l are
aco
ncen
trica
llyfla
t or s
light
lysl
ight
ly c
on-
dist
inct
ly c
on-f
lat t
o co
n-fl
at to
con
-sl
ight
ly c
on-
conc
entri
cally
conc
entri
cally
with
mor
e or
undu
late
co
ncen
trica
llyce
ntri
cally
cent
rica
llyce
ntri
cally
cent
rica
llyce
ntric
ally
un-
undu
late
un
dula
te
less
regu
larl
yw
ith sc
atte
red
undu
late
un
dula
te
undu
late
and
undu
late
un
dula
te
dula
te w
ithw
ith C
F an
dw
ith n
ume-
arra
nged
or ra
dial
lyce
ntra
lra
dial
lyra
dial
lysm
ooth
smoo
th C
Fnu
mer
ous C
Far
eola
ero
us C
F an
dra
dial
arra
nged
CF
areo
lae
and
wrin
kled
C
Fw
rinkl
ed C
Fan
d ar
eola
ean
d ar
eola
ear
rang
ed in
areo
lae
ope-
punc
taan
d ar
eola
eC
F sc
atte
red
and
areo
lae
and
areo
lae
scat
tere
d or
open
ings
mor
e or
less
ning
s sca
ttere
dor
arr
ange
dar
rang
ed in
alw
ays
arra
nged
insc
atte
red
orra
dial
row
sor
occ
asio
nal-
in ra
dial
radi
al p
atte
rnar
rang
ed in
radi
al ro
ws
arra
nged
inly
arr
ange
d in
row
sra
dial
row
sra
dial
pat
tern
radi
al p
atte
rn(F
igs
8 amp
9)
Rim
o-2
ndash3 in
the
(1)2
ndash5 in
the
1ndash2(
3) in
the
no d
ata
at le
ast o
ne
1ndash5
in th
eat
leas
t one
at
leas
t one
in
at le
ast o
ne
at le
ast o
ne
port
ulae
mar
gina
lm
argi
nal
mar
gina
l
Inte
rnal
lyon
the
valv
aem
argi
nal a
rea
on th
e va
lve
the
mar
gina
lon
the v
alva
eon
the
valv
aear
ea n
ear t
hear
ea n
ear t
hear
ea n
ear t
hea
sess
ile la
bi-
face
at t
hecl
ose
to li
mit
face
at t
hear
ea c
lose
tofa
ce a
t the
face
at t
heva
lve
mar
gin
valv
e mar
gin
valv
e mar
gin
um in
obl
ique
end
of s
hor-
of th
e ce
ntra
len
d of
sho
r-lim
it of
the
end
of s
hor-
end
of s
light
lyE
xter
nal o
pe-
Ext
erna
lE
xter
nal
posi
tion
lo-
tene
d st
ria
area
Ext
erna
lte
ned
stri
ace
ntra
l are
ate
ned
stri
ash
orte
ned
stri
ani
ngs
at th
eop
enin
gs a
top
enin
gs a
tca
ted
betw
een
Inte
rnal
ly a
open
ings
on
Inte
rnal
ly a
Ext
erna
l ope
-In
tern
ally
aIn
tern
ally
a la
rge
end
of s
hor-
the
end
ofth
e en
d of
alve
olar
sess
ile la
bi-
a hy
alin
e ar
eare
lativ
ely
ning
s on
ala
rge
sess
ilese
ssile
labi
umte
ned
stri
ash
orte
ned
shor
tene
dop
enin
gsum
in ra
dial
at th
e en
d of
larg
e se
ssile
hyal
ine
area
labi
um in
in o
bliq
ue o
rIn
tern
ally
ast
ria
Inte
r-st
ria
Inte
r-an
d ce
ntra
lpo
sitio
n lo
-a
slig
htly
labi
um in
slit
at th
e en
dob
lique
po-
circ
umfe
rent
ial
smal
l ses
sile
nally
a se
ssile
nally
a sm
all
area
cate
d m
id-
shor
tene
dpo
sitio
n lo
-of
a s
light
lysi
tion
loca
-po
sitio
n lo
cate
dla
bium
clo
sela
bium
ver
yse
ssile
labi
-w
ay b
etw
een
stria
Int
erna
l-ca
ted
betw
een
shor
tene
dte
d be
twee
nin
the
mar
gina
lto
inte
rnal
clos
e to
inte
r-um
clo
se to
alve
olar
ope
-ly
a s
essi
leal
veol
ar o
pe-
stri
a In
ter-
alve
olar
ope
-ar
ea n
ear t
o th
eal
veol
ar o
pe-
nal a
lveo
lar
inte
rnal
alv
e-ni
ngs
and
labi
um in
ning
s an
dna
lly a
ses
-ni
ngs
and
cent
ral a
rea
ning
s w
ithop
enin
gs
olar
ope
ning
sce
ntra
l are
aob
lique
to c
ir-ce
ntra
l are
asi
le la
bium
cent
ral a
rea
radi
ally
or
with
radi
ally
with
radi
ally
cum
fere
ntia
lw
ith a
slit
ori-
obliq
uely
po-
or o
bliq
uely
or o
bliq
uely
slit
posi
tion
ente
d ob
lique
-si
tione
d sl
itpo
sitio
ned
posi
tione
d sl
itly
to n
earl
ysl
itci
rcum
fere
n-tia
llyIn
tern
alsm
all
cost
aesm
all
cost
aesm
all
cost
ae
radi
ally
radi
ally
elon
-ra
dial
ly e
lon-
smal
l co
stae
smal
l co
stae
smal
l co
stae
smal
l co
stae
alve
olar
bear
ing
MF
bear
ing
MF
bear
ing
MF
elon
gate
dga
ted
cos
tae
gate
d c
osta
ebe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fop
enin
gscl
early
larg
ercl
early
larg
ercl
early
larg
erco
stae
bea
-be
arin
g M
Fbe
arin
g M
Fcl
earl
y la
rger
clea
rly la
rger
clea
rly la
rger
clea
rly
larg
erth
an th
in ri
bsth
an th
in ri
bsth
an th
in ri
bsri
ng M
Fcl
early
larg
ersl
ight
ly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
than
thin
ribs
clea
rly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
518
Lak
e B
alat
onM
esot
roph
ic
Eut
roph
ic
Plan
kton
of a
no d
ata
Plan
kton
or
no d
ata
Plan
kton
of
no d
ata
foss
il di
atom
near
Sioacute
fok
eutr
ophi
chy
pere
utro
-nu
mbe
r of
litto
ral o
ligo
olig
om
eso-
lake
s re
ser
ph
ic re
serv
oir
lake
s po
orm
esot
roph
ictr
ophi
csl
owly
run
-in
org
anic
lake
sm
ainl
y su
b-ni
ng w
ater
sm
atte
r and
alpi
ne la
kes
One
of t
heso
me
quar
rym
ost c
omm
onpo
ols
inE
urop
ean
Eng
lish
Lak
esp
ecie
s of
the
Dis
tric
tC
-rad
iosa
com
plex
Tab
le 3
con
tinue
d
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
519
samples declined in subsequent winter-spring months which is a typical consequenceof diatom cell division The number of striae in 10 microm was found to be lower (11 to17 striae in 10 microm) in some specimens from Rusałka Lake than has been originallystated by Pantocsek (1901) It might be related to the striae counting method becauseeven at original Pantocseks drawings (Plate XV figs 319 332) number of striae in10 microm is lower when the Genkalrsquos method (1977) is applied It also remains inaccordance with our observations of the type material (Table 3) and the observationsin Houk et al (2010)
Puncticulata balatonis is morphologically similar to several species of the complexmaking it difficult for a non-specialist to distinguish them The species resemblesP radiosa but differs from it in valve face ultrastructure such as the location of theinternal rimoportula openings The valves of P radiosa have a more coarse andradially wrinkled central area in the valve face whereas P balatonis has a smootherand finely silicified valve face Additionally the radial pattern of the valve facefultoportulae and areolae is very distinct in P radiosa The external rimoportulaopenings of P balatonis are situated at the end of shortened stria well discernible inLM as gaps (notae) whereas according to Houk et al (2010) the external openingsof P radiosa are situated at the end of only slightly shortened striae The differencesare clearly visible from the internal valvae view because P balatonis has valve facerimoportula openings located close to the internal alveolar openings whereas the
Fig 23 The variation in Puncticulata balatonis valve size in Rusałka Lake in subsequent months in2007
520
rimoportula openings in P radiosa are located more close to the central area ofthe valves Such a close location of the internal rimoportula openings to the valvemargin is typical only for P balatonis within the discussed group (Table 3)
Another diatom showing a similar morphology to Puncticulata balatonis is P prae-termissa (Lund) Haringkansson Most of the morphological features are similar forboth species (Table 3) but the later possesses radially elongated relatively largeinternal alveolar openings Moreover the costae bearing marginal fultoportulaeare only slightly thicker than the rest and the internal rimoportula openings arecloser to the central part of the valve face (Table 3) The last feature as well as themarginal stria length enable also the morphological distinction between P balatonisand P comta
Ecology and distribution
The distinction between the species of the Puncticulata radiosa-complex includingPuncticulata balatonis is not easy for a plankton ecologist or other researcher whois not a centric diatom specialist The correct identification requires detailedmorphological studies with SEM andor specialized knowledge That is why apartfrom the long-lasting taxonomical confusion the real distribution of the speciesfrom the complex is uncertain According to Houk et al (2010) Puncticulata balatonis(as Cyclotella balatonis) is one of the most common members of the P radiosa (as
Fig 24 The number of Puncticulata balatonis frustules in the water column of Rusałka Lake in 2007
521
Cyclotella radiosa) complex in Europe found in numerous mesotrophic and eutrophiclakes and slowly running waters in Hungary Germany and Czech Republic Howeverreports of the species are not numerous Till now P balatonis has also never beenreported from Poland
The limited number of reports on the occurrence of P balatonis may be related to itsmisidentificationmisinterpretation as Cyclotella comta (Ehrenb) Kuumltz (Kiss amp Pająk1994 fig 13) found in the plankton of the eutrophic Vistula River as Cyclotellapraetermissa quadrijuncta (Wunsam et al 1995) or as Puncticulata radiosa (Wojtalamp Kwandrans 2006) found in the eutrophic Rudawa River We are convinced that apart of other records of P comta the most commonly reported species of the complexin Poland (Siemińska amp Wołowski 2003) in fact might concern P balatonis InRusałka Lake the species may have also been mistaken for P comta (asC comta) reported from the lake by Kotlińska (1976) It is also likely that P bala-tonis in Poland may have been misidentified as P praetermissa although the latterspecies was only reported several times including findings in the benthos of CzarneLake and the Kamionka River NE Poland (Rakowska 1996) in the plankton ofseveral lakes of the Drawa National Park NW Poland (Gołdyn amp Szeląg-Wasielewska2004) in a sand-pit lake in Krakoacutew and a spring fed pond in Krztynia in S Poland(Wojtal unpublished data)
Up to now there has also been much confusion around the ecology of the speciesfrom the complex For instance P praetermissa described by Lund (1951) fromlakes with a low organic matter content was later found both in the plankton and thebenthos in a wide variety of natural and man-made lakes including alpine andsubalpine lakes (Wunsam et al 1995 Lange-Bertalot amp Metzeltin 1996 Guumlttingeramp Straub 1998 Tolotti et al 2004) as well as in more eutrophic habitats (egSchmidt et al 2002 Hindaacutek amp Hindaacutekovaacute 2003 Tanaka 2007) Houk et al (2010)cleared up the ambiguities ascribing P praetermissa and P radiosa to oligo-mesotrophic habitats According to the authors P comta (as Cyclotella comta) is tillnow known only from fossil materials P balatonis (as Cyclotella balatonis) inturn has been assigned to meso-eutrophic lakes and rivers consistently with thetrophic character of its type locality ndash lake Balaton Our results correspond with theautecology of P balatonis (as Cyclotella balatonis) proposed by Houk et al (2010)widening even slightly the preferences of the species for the trophic conditionsdetermined by Houk et al (2010) as it was also able to develop a large populationin an eutrophic to hypereutrophic water body
The highest numbers of P balatonis in Rusałka Lake were observed in the winter-spring period The diatom bloom in the spring plankton is a well-known phenomenon(Sommer et al 1986) ndash sometimes also occurring during the winter period (Kiss ampGenkal 1993) ndash and is usually associated with a changing silica andor phosphorusavailability during that period (Lampert amp Sommer 2001) Sinking was also provedto be an essential factor hampering diatom success during the thermal stratificationperiod (Jewson et al 1981 Ferris amp Lehman 2007) explaining probably the seasonalityof the P balatonis growth in Rusałka Lake as its high development finished withthe stratification onset Relatively high numbers of frustules in deeper water layersmight also indicate a significant role of the sedimentation process
522
Conclusions
The morphology of Puncticulata balatonis in Rusałka Lake corresponds well toother species around P comta with the exception of the location of marginalfultoportula openings and the length of the shortened striae (notae) Analysis of themorphological characters enabled the transfer of Cyclotella balatonis into the genusPuncticulata A large population of the species in the eutrophic to hypereutrophicRusałka Lake remains congruent with its tolerance ranges with respect to the trophicstate proposed by Houk et al (2010) The variation in the cell number and averagedimension of frustules indicated the species preference to develop large populationsin spring
Acknowledgements
We thank Prof Ryszard Gołdyn (AMickiewicz University in Poznań) for his inspiration researchsupervision and useful comments to the manuscript We are also grateful to Dr Bart Van de Vijver andtwo anonymous reviewers for their valuable suggestions and to Dr Krisztina Buczko for enabling theanalysis of the type material of Cyclotella balatonis from the Pantocsekrsquos collection at HungarianNatural History Museum Sampling on Rusałka Lake was part of a project financially supported by theDistrict and Municipial Funds of Enviromental Protection and Water Management in Poznań (Poland)
References
CARLSON RE (1977) A trophic state index for lakes ndash Limnol Oceanogr 22 (2) 361ndash369
CREMER H amp H KOOLMEES (2010) Common diatoms (Centrales and Fragilariaceae Bacillario-phyta) of modern and fossil freshwater environments in the Netherlands ndash Nova Hedwigia 90(3ndash4)343ndash381
FERRIS JA amp JT LEHMAN (2007) Interannual variation in diatom bloom dynamics Roles ofhydrology nutrient limitation sinking and whole lake manipulation ndash Water Res 41 2551ndash2562
GENKAL SI (1977) On the method of calculation of some taxonomicaly significant structuralelements of the valve in the diatoms of the family Thalassiosiraceae Lebour emend Hasle (Bacillariophyta)ndash Bot Zhurn 62 848ndash851 (in Russian)
GENKAL SI amp KT KISS (1993) Morphological variability of the diatom Cyclotella atomusHustedt var atomus and C atomus var gracilis var nov ndash Hydrobiologia 269270 39ndash47
GOŁDYN R S PODSIADŁOWSKI K KOWALCZEWSKA-MADURA R DONDAJEWSKAE SZELĄG-WASIELEWSKA A BUDZYŃSKA P DOMEK amp W ROMANOWICZ (2010)Functioning of the Rusałka Lake ecosystem in Poznań (west Poland) ndash Ocean Hydrobiol Stud39(3) 65ndash80
GOŁDYN R amp E SZELĄG-WASIELEWSKA (2004) Changes in the phytoseston of a river-lakesystem in Drawieński National Park ndash Ocean Hydrobiol Stud 33(2) 17ndash28
GUumlTTINGER W amp F STRAUB (1998) Diatoms of Lake Cadagno ndash In PEDUZZI RR BACHOFEN amp M TONOLLA (eds) Lake Cadagno a meromictic Alpine lake ndash Doc Ist ItalIdrobiol 63 57ndash64
HANDMANN R (1913) Die Diatomeenflora des Almfeegebiets ndash In MANZ GJ (ed) Mitteilungendes Mikrologischen Vereins Linz ndash Mikrolog Ver Regensburg (in German)
HAUSMANN S amp AF LOTTER (2001) Morphological variation within the diatom taxon Cyclotellacomensis and its importance for quantitative temperature reconstructions ndash Freshw Biol 46 1323ndash1333
523
HAYASHI T (2007) Puncticulata versiformis sp nov and Cyclotella kathmanduensis sp nov(Bacillariophyta) new fossil species from middle Pleistocene lacustrine sediments Kathmandu NepalHilalaya ndash J Phycol 43(2) 304ndash318
HAringKANSSON H (2002) A compilation and evaluation of species in the genera StephanodiscusCyclostephanos and Cylotella with a new genus in the family Stephanodiscaceae ndash Diatom Res 17(1)1ndash139
HINDAacuteK F amp A HINDAacuteKOVAacute (2003) Diversity of cyanobacteria and algae of urban gravel pitlakes in Bratislava Slovakia a survey ndash Hydrobiologia 506ndash509 155ndash162
HINDAacuteK F amp A HINDAacuteKOVAacute (2008) Diversity of cyanobacteria and algae of the NationalNature Reserve Čiacutečovskeacute mŕtve rameno oxbow (Southern Slovakia) ndash Bulletin SBS 30 (1) 11ndash19 (inSlovak with an English abstract)
HOUK V R KLEE amp H TANAKA (2010) Atlas of freshwater centric diatoms with a brief key anddescriptions Part III Stephanodiscaceae A Cyclotella Tertiarius Discostella ndash Fottea 10 (Supplement)1ndash498
JEWSON DH BH RIPPEY amp WJK GILMORE (1981) Loss rates from sedimentation parasitismand grazing during the growth nutrient limitation and dormancy of a diatom crop ndash Limnol Oceanogr26 1045ndash1056
KISS KT amp SI GENKAL (1993) Winter blooms of centric diatoms in the River Danube and in itsside arms near Budapest ndash Hydrobiologia 269270 317ndash325
KISS KT amp G PAJĄK (1994) Seasonal change of diatom s in the plankton of the Vistula riverabove and below the Goczalkowice reservoir Poland ndash In KOCIOLEK JP (ed) Proceedings ofthe 11th International Diatom Symposium San Francisco Aug 12ndash17 1990 Memoirs of the CaliforniaAcademy of Sciences 17 517ndash524 San Francisco California Academy of Sciences
KOBAYASI H M IDEI S MAYAMA T NAGUMO amp K OSADA (2006) H KobayasirsquosAtlas of Japanese diatoms based on electron microscopy 1 1ndash531 Uchida Rokakuho Publishing CoLtd Tokyo
KOTLIŃSKA E (1976) Fitoplankton Jeziora Rusałka In Fitoplankton sztucznych jezior położonychna terenie Poznania 1ndash54 Poznańskie Tow Przyjacioacuteł Nauk Prace Komisji Biologicznej (in Polish)
KRAMMER K amp H LANGE-BERTALOT (1991) Bacillariophyceae 3 Teil Centrales Fragilaria-ceae Eunotiaceae ndash In ETTL H J GERLOFF H HEYNIGH amp D MOLLENHAUER (eds)Suumlsswasserflora von Mitteleuropa 2(3) 1ndash576 Gustav Fischer Verlag Stuttgart Jena
LAMPERT W amp U SOMMER (2001) Ekologia woacuted śroacutedlądowych ndash PWN Warszawa
LANGE-BERTALOT H amp D METZELTIN (1996) Indicators of oligotrophy 800 representativeof three ecologically distinct lake types Carbonate buffered-Oligotrophic-Weakly buffered soft waterndash Iconogr Diatomol 2 1ndash390
LUND JWG (1951) Contribution to our knowledge of British algae ndash Hydrobiologia 3(1) 93ndash100
ORGANIZATION FOR ECONOMIC COOPERATION AND DEVELOPMENT (1982)Eutrophication of Waters ndash Monitoring Assessment and Control Final Report OECD Paris
PANTOCSEK J (1901) Die Kieselalgen oder Bacillarien des Balaton 1ndash112 XVI Tafeln ndash Druckder K und K Hofbuchdruckerei des Victor Hornyaacutenszky Budapest
RAKOWSKA B (1996) Preliminary elaboration of the diatom communities occuring in the benthosof the Kamionka River and Lake Czarne at the Pojezierze Suwalskie (1993ndash1994) ndash Fragm FloristGeobot Polon 3 221ndash238
RIMET F JC DRUART amp O ANNEVILLE (2009) Exploring the dynamics of plankton diatomcommunities in Lake Geneva using emergent self-organizing maps (1974ndash2007) ndash Ecol Inform4 99ndash110
524
SCHMIDT R R PSENNER J MUumlLLER P INDINGER amp C KAMENIK (2002) Impact of thelate glacial climate variations on stratification and trophic state of the meromictic lake Laumlngsee (Austria)validation of a conceptual model by multiproxy studies ndash J Limnol 61(1) 49ndash60
SIEMIŃSKA J amp K WOŁOWSKI (2003) Catalogue of Polish prokaryotic and eukaryotic algaendash W Szafer Institute of Botany Polish Academy of Sciences Krakoacutew
SIVER PA (1999) Development of paleolimnological inference models for pH total nitrogen andspecific conductivity based on planktonic diatoms ndash J Paleolimnol 21 45ndash59
SOMMER U ZM GLIWICZ W LAMPERT amp A DUNCAN (1986) The PEG-model of seasonalsuccession of planktonic events in fresh waters ndash Arch Hydrobiol 106 433ndash471
TANAKA H (2007) Taxonomic studies of the genera Cyclotella (Kuumltzing) Breacutebisson DiscostellaHouk et Klee and Puncticulata Haringkansson in the family Stepanodiscaceae Glezer et Makarova(Bacillariophyta) in Japan ndash Biblioth Diatomol 53 1ndash205
TOLOTTI M D CALLIARI amp F CORRADINI (2004) Interannual variability of the phytoplanktonof Tovel lake (Trentino Italy) ndash Studi Trent di Scienze Naturali Acta Biol 81 Suppl 2 327ndash340 (inItalian)
WETZEL RG amp GE LIKENS (1991) Limnological analyses 1ndash391 ndash Springer-Verlag NewYork Berlin Heidelberg
WOJTAL AZ amp J KWANDRANS (2006) Diatoms of the Wyżyna Krakowsko-Częstochowskaupland (S Poland) ndash Coscinodiscophyceae (Thalassiosirophycidae) ndash Polish Bot J 51(2) 177ndash207
WUNSAM S R SCHMIDT amp R KLEE (1995) Cyclotella-taxa (Bacillariophyceae) in lakes of theAlpine region and their relationship to environmental variables ndash Aquat Sci 57(4) 360ndash381
Received 25 March 2010 accepted in revised form 31 March 2011

514
Figs 15ndash22 Puncticulata balatonis (Pantocsek) Wojtal et Budzyńska Scanning electron microscopyFigs 15ndash20 specimens from the Rusałka Lake Figs 21ndash22 P balatonis type material samples no152 amp 156 Fig 15 external view of the rimoportula opening at the end of shortened striae ndash notae
515
P balatonis contributed between 63 and 187 to the total phytoplankton numbersand between 21 and 47 to the total biomass From June until October the numberof the diatom did not exceed 300 frustules per 1 ml (Fig 24) and no valve wasfound in September At the beginning of December P balatonis number increasedslightly again
At the time of the bloom of P balatonis chrysoflagellates (Erkenia subaequiciliataSkuja Chrysococcus triporus Matvienko) pennate diatoms (Fragilaria sp) andorcoenobia-forming Chlorococcales (Didymocystis inconspicua Koršikov Tetastrumsp div Scenedesmus sp div Coelastrum microporum Naumlgeli Crucigenia tetrapediaKirchner) were also abundant Other diatoms such as Aulacoseira ambigua (Grunow)Simonsen Cyclotella meneghiniana Kuumltzing C ocellata Pantocsek Cyclostephanosdelicatus (Genkal) Casper C dubius (Fricke) Round Stephanodiscus parvus Stoermeramp Haringkansson Asterionella formosa Hassal and Navicula sp were also observed
Discussion
Morphology of Puncticulata balatonis (Pant) Wojtal et Budzyńska
Puncticulata balatonis shares important morphological characters assigned to thegenus justifying its formal transfer to this genus a slightly concentrically undulatemiddle part of the valve face with scattered or radially arranged numerous areolaewith internally domed cribra and numerous fultoportulae Additionally valves ofP balatonis have internal alveolar openings with marginal fultoportulae located onthicker costae rimoportulae located externally at the top of shortened striae andinternally in the marginal part of the valve face
The morphology of P balatonis valves from Rusałka Lake corresponds well withthe morphology of the specimens from the Pantocsekrsquos type material (Figs 13 1421 22 Table 3) though the diameter range and the number of striae in 10 microm werefound to be different Pantocsek (1901) stated in his diagnosis (p 104) that thevalves of Cyclotella balatonis are 16ndash20 microm in diameter but we have found bothsmaller and larger specimens (Table 3) The wider diameter range in our observationcould be related to a more complete morphological range of the species since thediatom population in Rusałka Lake was abundant However the time of samplingmay have also been of importance as the medium valve size of the species in our
(arrowed) at concentrically elevated valve face Fig 16 external view of the openings of central (right)and marginal (left) fultoportulae (arrowed) Fig 17 external view of the short and long striae of themarginal area Fig18 internal view of the rimoportula opening (arrowed) located close to the valvemargin and internal alveolar opening and central openings of numerous areolae and fultoportulae(arrowed) Fig 19 internal view of the divided thinner costa (marked) and internal opening of centralfultoportula (arrowed) Fig 20 internal view of the marginal fultoportulae located on the thicker costae(arrowed) Fig 21 external view of the valve face with marked notae Fig 22 internal view of thealveolar openings with thicker costa bearing marginal fultoportulae and internal rimoportula openinglocated near the margin (arrowed) Scale bar represents 5 microm for Figs 15 18 amp 21 4 microm for Fig 163 microm for Figs 19 amp 20 and 2 microm for Figs 17 amp 22
516
Tab
le 3
Com
pari
son
of P
unct
icul
ata
bala
toni
s an
d m
orph
olog
ical
ly s
imila
r sp
ecie
s C
F ndash
cent
ral
fulto
port
ulae
MF
ndash m
argi
nal
fulto
port
ulae
o
urob
serv
atio
n ty
pe m
ater
ial S
iofoacute
k s
ampl
e no
152
our
obse
rvat
ion
base
d on
the
fig
62
11
12
in K
ram
mer
amp L
ange
-Ber
talo
t 199
1 w
hich
sho
ws
spec
imen
s fr
om th
e ty
pe m
ater
ial o
f Cyc
lote
lla
bala
toni
s Pa
ntoc
sek
(as
C r
adio
sa)
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
Ref
eren
ces
Pant
ocse
kH
ouk
et a
lL
und
1951
H
aringkan
sson
H
ouk
et a
lH
aringkan
sson
H
ouk
et a
lH
aringkan
sson
H
ouk
et a
l19
01 p
104
20
10 p
39
p 9
820
02 p
116
20
10 p
41
2002
p 1
142
010
p 3
720
02 p
113
20
10 p
36
Plat
e X
V
Plat
es 2
69
Figs
1 2
Figs
422
ndashPl
ates
280
Fi
gs 4
15ndash
Plat
es 2
61
Figs
410
ndash P
late
s 25
7 1
ndashFi
gs 3
19
1ndash11
270
42
61ndash
11 2
81
421
1ndash11
262
41
423
258
1ndash1
133
21ndash
6 2
71
1ndash6
1ndash6
1ndash12
263
25
9 1
ndash12
1ndash5
264
1ndash6
260
1ndash8
Col
onie
sno
dat
a2ndash
82ndash
7 ce
lls2ndash
8 ce
lls
shor
tC
ells
solit
ary
no d
ata
mos
tlyce
lls s
olita
ryno
dat
alo
osel
ylo
osel
ylo
osel
yor
in s
hort
solit
ary
rare
ly in
unite
dun
ited
unite
d ch
ains
loos
ely
unite
dsh
ort c
hain
sch
ains
2(8
)ce
llsD
iam
eter
16ndash2
016
ndash22
87ndash
210
8ndash25
8ndash25
8ndash30
7ndash35
7ndash25
8ndash50
5ndash32
(microm
)St
riae
20 p
er 1
0 microm
ca
16
per
15ndash1
85
per
13ndash1
9 pe
r13
ndash15
per
15ndash1
8 pe
r 10
no d
ata
15ndash1
8 pe
r 10
no d
ata
14ndash1
8 pe
r10
microm
of
10 micro
m10
microm
10 micro
mmicrom
of n
earl
ymicrom
of n
early
10 micro
mne
arly
equ
aleq
ual l
engt
heq
ual l
engt
hle
ngth
occ
upy
occu
py 1
2ndash1
4ta
king
abo
ut1
3ndash1
2 of
of th
e va
lvae
12
of th
e va
lve
valv
e fa
cedi
amet
er fa
ce d
iam
eter
diam
eter
517
Cen
tral
are
a
flat
tofla
t to
slig
htly
flat t
o sl
ight
lyce
ntra
l are
aco
ncen
trica
llyfla
t or s
light
lysl
ight
ly c
on-
dist
inct
ly c
on-f
lat t
o co
n-fl
at to
con
-sl
ight
ly c
on-
conc
entri
cally
conc
entri
cally
with
mor
e or
undu
late
co
ncen
trica
llyce
ntri
cally
cent
rica
llyce
ntri
cally
cent
rica
llyce
ntric
ally
un-
undu
late
un
dula
te
less
regu
larl
yw
ith sc
atte
red
undu
late
un
dula
te
undu
late
and
undu
late
un
dula
te
dula
te w
ithw
ith C
F an
dw
ith n
ume-
arra
nged
or ra
dial
lyce
ntra
lra
dial
lyra
dial
lysm
ooth
smoo
th C
Fnu
mer
ous C
Far
eola
ero
us C
F an
dra
dial
arra
nged
CF
areo
lae
and
wrin
kled
C
Fw
rinkl
ed C
Fan
d ar
eola
ean
d ar
eola
ear
rang
ed in
areo
lae
ope-
punc
taan
d ar
eola
eC
F sc
atte
red
and
areo
lae
and
areo
lae
scat
tere
d or
open
ings
mor
e or
less
ning
s sca
ttere
dor
arr
ange
dar
rang
ed in
alw
ays
arra
nged
insc
atte
red
orra
dial
row
sor
occ
asio
nal-
in ra
dial
radi
al p
atte
rnar
rang
ed in
radi
al ro
ws
arra
nged
inly
arr
ange
d in
row
sra
dial
row
sra
dial
pat
tern
radi
al p
atte
rn(F
igs
8 amp
9)
Rim
o-2
ndash3 in
the
(1)2
ndash5 in
the
1ndash2(
3) in
the
no d
ata
at le
ast o
ne
1ndash5
in th
eat
leas
t one
at
leas
t one
in
at le
ast o
ne
at le
ast o
ne
port
ulae
mar
gina
lm
argi
nal
mar
gina
l
Inte
rnal
lyon
the
valv
aem
argi
nal a
rea
on th
e va
lve
the
mar
gina
lon
the v
alva
eon
the
valv
aear
ea n
ear t
hear
ea n
ear t
hear
ea n
ear t
hea
sess
ile la
bi-
face
at t
hecl
ose
to li
mit
face
at t
hear
ea c
lose
tofa
ce a
t the
face
at t
heva
lve
mar
gin
valv
e mar
gin
valv
e mar
gin
um in
obl
ique
end
of s
hor-
of th
e ce
ntra
len
d of
sho
r-lim
it of
the
end
of s
hor-
end
of s
light
lyE
xter
nal o
pe-
Ext
erna
lE
xter
nal
posi
tion
lo-
tene
d st
ria
area
Ext
erna
lte
ned
stri
ace
ntra
l are
ate
ned
stri
ash
orte
ned
stri
ani
ngs
at th
eop
enin
gs a
top
enin
gs a
tca
ted
betw
een
Inte
rnal
ly a
open
ings
on
Inte
rnal
ly a
Ext
erna
l ope
-In
tern
ally
aIn
tern
ally
a la
rge
end
of s
hor-
the
end
ofth
e en
d of
alve
olar
sess
ile la
bi-
a hy
alin
e ar
eare
lativ
ely
ning
s on
ala
rge
sess
ilese
ssile
labi
umte
ned
stri
ash
orte
ned
shor
tene
dop
enin
gsum
in ra
dial
at th
e en
d of
larg
e se
ssile
hyal
ine
area
labi
um in
in o
bliq
ue o
rIn
tern
ally
ast
ria
Inte
r-st
ria
Inte
r-an
d ce
ntra
lpo
sitio
n lo
-a
slig
htly
labi
um in
slit
at th
e en
dob
lique
po-
circ
umfe
rent
ial
smal
l ses
sile
nally
a se
ssile
nally
a sm
all
area
cate
d m
id-
shor
tene
dpo
sitio
n lo
-of
a s
light
lysi
tion
loca
-po
sitio
n lo
cate
dla
bium
clo
sela
bium
ver
yse
ssile
labi
-w
ay b
etw
een
stria
Int
erna
l-ca
ted
betw
een
shor
tene
dte
d be
twee
nin
the
mar
gina
lto
inte
rnal
clos
e to
inte
r-um
clo
se to
alve
olar
ope
-ly
a s
essi
leal
veol
ar o
pe-
stri
a In
ter-
alve
olar
ope
-ar
ea n
ear t
o th
eal
veol
ar o
pe-
nal a
lveo
lar
inte
rnal
alv
e-ni
ngs
and
labi
um in
ning
s an
dna
lly a
ses
-ni
ngs
and
cent
ral a
rea
ning
s w
ithop
enin
gs
olar
ope
ning
sce
ntra
l are
aob
lique
to c
ir-ce
ntra
l are
asi
le la
bium
cent
ral a
rea
radi
ally
or
with
radi
ally
with
radi
ally
cum
fere
ntia
lw
ith a
slit
ori-
obliq
uely
po-
or o
bliq
uely
or o
bliq
uely
slit
posi
tion
ente
d ob
lique
-si
tione
d sl
itpo
sitio
ned
posi
tione
d sl
itly
to n
earl
ysl
itci
rcum
fere
n-tia
llyIn
tern
alsm
all
cost
aesm
all
cost
aesm
all
cost
ae
radi
ally
radi
ally
elon
-ra
dial
ly e
lon-
smal
l co
stae
smal
l co
stae
smal
l co
stae
smal
l co
stae
alve
olar
bear
ing
MF
bear
ing
MF
bear
ing
MF
elon
gate
dga
ted
cos
tae
gate
d c
osta
ebe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fop
enin
gscl
early
larg
ercl
early
larg
ercl
early
larg
erco
stae
bea
-be
arin
g M
Fbe
arin
g M
Fcl
earl
y la
rger
clea
rly la
rger
clea
rly la
rger
clea
rly
larg
erth
an th
in ri
bsth
an th
in ri
bsth
an th
in ri
bsri
ng M
Fcl
early
larg
ersl
ight
ly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
than
thin
ribs
clea
rly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
518
Lak
e B
alat
onM
esot
roph
ic
Eut
roph
ic
Plan
kton
of a
no d
ata
Plan
kton
or
no d
ata
Plan
kton
of
no d
ata
foss
il di
atom
near
Sioacute
fok
eutr
ophi
chy
pere
utro
-nu
mbe
r of
litto
ral o
ligo
olig
om
eso-
lake
s re
ser
ph
ic re
serv
oir
lake
s po
orm
esot
roph
ictr
ophi
csl
owly
run
-in
org
anic
lake
sm
ainl
y su
b-ni
ng w
ater
sm
atte
r and
alpi
ne la
kes
One
of t
heso
me
quar
rym
ost c
omm
onpo
ols
inE
urop
ean
Eng
lish
Lak
esp
ecie
s of
the
Dis
tric
tC
-rad
iosa
com
plex
Tab
le 3
con
tinue
d
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
519
samples declined in subsequent winter-spring months which is a typical consequenceof diatom cell division The number of striae in 10 microm was found to be lower (11 to17 striae in 10 microm) in some specimens from Rusałka Lake than has been originallystated by Pantocsek (1901) It might be related to the striae counting method becauseeven at original Pantocseks drawings (Plate XV figs 319 332) number of striae in10 microm is lower when the Genkalrsquos method (1977) is applied It also remains inaccordance with our observations of the type material (Table 3) and the observationsin Houk et al (2010)
Puncticulata balatonis is morphologically similar to several species of the complexmaking it difficult for a non-specialist to distinguish them The species resemblesP radiosa but differs from it in valve face ultrastructure such as the location of theinternal rimoportula openings The valves of P radiosa have a more coarse andradially wrinkled central area in the valve face whereas P balatonis has a smootherand finely silicified valve face Additionally the radial pattern of the valve facefultoportulae and areolae is very distinct in P radiosa The external rimoportulaopenings of P balatonis are situated at the end of shortened stria well discernible inLM as gaps (notae) whereas according to Houk et al (2010) the external openingsof P radiosa are situated at the end of only slightly shortened striae The differencesare clearly visible from the internal valvae view because P balatonis has valve facerimoportula openings located close to the internal alveolar openings whereas the
Fig 23 The variation in Puncticulata balatonis valve size in Rusałka Lake in subsequent months in2007
520
rimoportula openings in P radiosa are located more close to the central area ofthe valves Such a close location of the internal rimoportula openings to the valvemargin is typical only for P balatonis within the discussed group (Table 3)
Another diatom showing a similar morphology to Puncticulata balatonis is P prae-termissa (Lund) Haringkansson Most of the morphological features are similar forboth species (Table 3) but the later possesses radially elongated relatively largeinternal alveolar openings Moreover the costae bearing marginal fultoportulaeare only slightly thicker than the rest and the internal rimoportula openings arecloser to the central part of the valve face (Table 3) The last feature as well as themarginal stria length enable also the morphological distinction between P balatonisand P comta
Ecology and distribution
The distinction between the species of the Puncticulata radiosa-complex includingPuncticulata balatonis is not easy for a plankton ecologist or other researcher whois not a centric diatom specialist The correct identification requires detailedmorphological studies with SEM andor specialized knowledge That is why apartfrom the long-lasting taxonomical confusion the real distribution of the speciesfrom the complex is uncertain According to Houk et al (2010) Puncticulata balatonis(as Cyclotella balatonis) is one of the most common members of the P radiosa (as
Fig 24 The number of Puncticulata balatonis frustules in the water column of Rusałka Lake in 2007
521
Cyclotella radiosa) complex in Europe found in numerous mesotrophic and eutrophiclakes and slowly running waters in Hungary Germany and Czech Republic Howeverreports of the species are not numerous Till now P balatonis has also never beenreported from Poland
The limited number of reports on the occurrence of P balatonis may be related to itsmisidentificationmisinterpretation as Cyclotella comta (Ehrenb) Kuumltz (Kiss amp Pająk1994 fig 13) found in the plankton of the eutrophic Vistula River as Cyclotellapraetermissa quadrijuncta (Wunsam et al 1995) or as Puncticulata radiosa (Wojtalamp Kwandrans 2006) found in the eutrophic Rudawa River We are convinced that apart of other records of P comta the most commonly reported species of the complexin Poland (Siemińska amp Wołowski 2003) in fact might concern P balatonis InRusałka Lake the species may have also been mistaken for P comta (asC comta) reported from the lake by Kotlińska (1976) It is also likely that P bala-tonis in Poland may have been misidentified as P praetermissa although the latterspecies was only reported several times including findings in the benthos of CzarneLake and the Kamionka River NE Poland (Rakowska 1996) in the plankton ofseveral lakes of the Drawa National Park NW Poland (Gołdyn amp Szeląg-Wasielewska2004) in a sand-pit lake in Krakoacutew and a spring fed pond in Krztynia in S Poland(Wojtal unpublished data)
Up to now there has also been much confusion around the ecology of the speciesfrom the complex For instance P praetermissa described by Lund (1951) fromlakes with a low organic matter content was later found both in the plankton and thebenthos in a wide variety of natural and man-made lakes including alpine andsubalpine lakes (Wunsam et al 1995 Lange-Bertalot amp Metzeltin 1996 Guumlttingeramp Straub 1998 Tolotti et al 2004) as well as in more eutrophic habitats (egSchmidt et al 2002 Hindaacutek amp Hindaacutekovaacute 2003 Tanaka 2007) Houk et al (2010)cleared up the ambiguities ascribing P praetermissa and P radiosa to oligo-mesotrophic habitats According to the authors P comta (as Cyclotella comta) is tillnow known only from fossil materials P balatonis (as Cyclotella balatonis) inturn has been assigned to meso-eutrophic lakes and rivers consistently with thetrophic character of its type locality ndash lake Balaton Our results correspond with theautecology of P balatonis (as Cyclotella balatonis) proposed by Houk et al (2010)widening even slightly the preferences of the species for the trophic conditionsdetermined by Houk et al (2010) as it was also able to develop a large populationin an eutrophic to hypereutrophic water body
The highest numbers of P balatonis in Rusałka Lake were observed in the winter-spring period The diatom bloom in the spring plankton is a well-known phenomenon(Sommer et al 1986) ndash sometimes also occurring during the winter period (Kiss ampGenkal 1993) ndash and is usually associated with a changing silica andor phosphorusavailability during that period (Lampert amp Sommer 2001) Sinking was also provedto be an essential factor hampering diatom success during the thermal stratificationperiod (Jewson et al 1981 Ferris amp Lehman 2007) explaining probably the seasonalityof the P balatonis growth in Rusałka Lake as its high development finished withthe stratification onset Relatively high numbers of frustules in deeper water layersmight also indicate a significant role of the sedimentation process
522
Conclusions
The morphology of Puncticulata balatonis in Rusałka Lake corresponds well toother species around P comta with the exception of the location of marginalfultoportula openings and the length of the shortened striae (notae) Analysis of themorphological characters enabled the transfer of Cyclotella balatonis into the genusPuncticulata A large population of the species in the eutrophic to hypereutrophicRusałka Lake remains congruent with its tolerance ranges with respect to the trophicstate proposed by Houk et al (2010) The variation in the cell number and averagedimension of frustules indicated the species preference to develop large populationsin spring
Acknowledgements
We thank Prof Ryszard Gołdyn (AMickiewicz University in Poznań) for his inspiration researchsupervision and useful comments to the manuscript We are also grateful to Dr Bart Van de Vijver andtwo anonymous reviewers for their valuable suggestions and to Dr Krisztina Buczko for enabling theanalysis of the type material of Cyclotella balatonis from the Pantocsekrsquos collection at HungarianNatural History Museum Sampling on Rusałka Lake was part of a project financially supported by theDistrict and Municipial Funds of Enviromental Protection and Water Management in Poznań (Poland)
References
CARLSON RE (1977) A trophic state index for lakes ndash Limnol Oceanogr 22 (2) 361ndash369
CREMER H amp H KOOLMEES (2010) Common diatoms (Centrales and Fragilariaceae Bacillario-phyta) of modern and fossil freshwater environments in the Netherlands ndash Nova Hedwigia 90(3ndash4)343ndash381
FERRIS JA amp JT LEHMAN (2007) Interannual variation in diatom bloom dynamics Roles ofhydrology nutrient limitation sinking and whole lake manipulation ndash Water Res 41 2551ndash2562
GENKAL SI (1977) On the method of calculation of some taxonomicaly significant structuralelements of the valve in the diatoms of the family Thalassiosiraceae Lebour emend Hasle (Bacillariophyta)ndash Bot Zhurn 62 848ndash851 (in Russian)
GENKAL SI amp KT KISS (1993) Morphological variability of the diatom Cyclotella atomusHustedt var atomus and C atomus var gracilis var nov ndash Hydrobiologia 269270 39ndash47
GOŁDYN R S PODSIADŁOWSKI K KOWALCZEWSKA-MADURA R DONDAJEWSKAE SZELĄG-WASIELEWSKA A BUDZYŃSKA P DOMEK amp W ROMANOWICZ (2010)Functioning of the Rusałka Lake ecosystem in Poznań (west Poland) ndash Ocean Hydrobiol Stud39(3) 65ndash80
GOŁDYN R amp E SZELĄG-WASIELEWSKA (2004) Changes in the phytoseston of a river-lakesystem in Drawieński National Park ndash Ocean Hydrobiol Stud 33(2) 17ndash28
GUumlTTINGER W amp F STRAUB (1998) Diatoms of Lake Cadagno ndash In PEDUZZI RR BACHOFEN amp M TONOLLA (eds) Lake Cadagno a meromictic Alpine lake ndash Doc Ist ItalIdrobiol 63 57ndash64
HANDMANN R (1913) Die Diatomeenflora des Almfeegebiets ndash In MANZ GJ (ed) Mitteilungendes Mikrologischen Vereins Linz ndash Mikrolog Ver Regensburg (in German)
HAUSMANN S amp AF LOTTER (2001) Morphological variation within the diatom taxon Cyclotellacomensis and its importance for quantitative temperature reconstructions ndash Freshw Biol 46 1323ndash1333
523
HAYASHI T (2007) Puncticulata versiformis sp nov and Cyclotella kathmanduensis sp nov(Bacillariophyta) new fossil species from middle Pleistocene lacustrine sediments Kathmandu NepalHilalaya ndash J Phycol 43(2) 304ndash318
HAringKANSSON H (2002) A compilation and evaluation of species in the genera StephanodiscusCyclostephanos and Cylotella with a new genus in the family Stephanodiscaceae ndash Diatom Res 17(1)1ndash139
HINDAacuteK F amp A HINDAacuteKOVAacute (2003) Diversity of cyanobacteria and algae of urban gravel pitlakes in Bratislava Slovakia a survey ndash Hydrobiologia 506ndash509 155ndash162
HINDAacuteK F amp A HINDAacuteKOVAacute (2008) Diversity of cyanobacteria and algae of the NationalNature Reserve Čiacutečovskeacute mŕtve rameno oxbow (Southern Slovakia) ndash Bulletin SBS 30 (1) 11ndash19 (inSlovak with an English abstract)
HOUK V R KLEE amp H TANAKA (2010) Atlas of freshwater centric diatoms with a brief key anddescriptions Part III Stephanodiscaceae A Cyclotella Tertiarius Discostella ndash Fottea 10 (Supplement)1ndash498
JEWSON DH BH RIPPEY amp WJK GILMORE (1981) Loss rates from sedimentation parasitismand grazing during the growth nutrient limitation and dormancy of a diatom crop ndash Limnol Oceanogr26 1045ndash1056
KISS KT amp SI GENKAL (1993) Winter blooms of centric diatoms in the River Danube and in itsside arms near Budapest ndash Hydrobiologia 269270 317ndash325
KISS KT amp G PAJĄK (1994) Seasonal change of diatom s in the plankton of the Vistula riverabove and below the Goczalkowice reservoir Poland ndash In KOCIOLEK JP (ed) Proceedings ofthe 11th International Diatom Symposium San Francisco Aug 12ndash17 1990 Memoirs of the CaliforniaAcademy of Sciences 17 517ndash524 San Francisco California Academy of Sciences
KOBAYASI H M IDEI S MAYAMA T NAGUMO amp K OSADA (2006) H KobayasirsquosAtlas of Japanese diatoms based on electron microscopy 1 1ndash531 Uchida Rokakuho Publishing CoLtd Tokyo
KOTLIŃSKA E (1976) Fitoplankton Jeziora Rusałka In Fitoplankton sztucznych jezior położonychna terenie Poznania 1ndash54 Poznańskie Tow Przyjacioacuteł Nauk Prace Komisji Biologicznej (in Polish)
KRAMMER K amp H LANGE-BERTALOT (1991) Bacillariophyceae 3 Teil Centrales Fragilaria-ceae Eunotiaceae ndash In ETTL H J GERLOFF H HEYNIGH amp D MOLLENHAUER (eds)Suumlsswasserflora von Mitteleuropa 2(3) 1ndash576 Gustav Fischer Verlag Stuttgart Jena
LAMPERT W amp U SOMMER (2001) Ekologia woacuted śroacutedlądowych ndash PWN Warszawa
LANGE-BERTALOT H amp D METZELTIN (1996) Indicators of oligotrophy 800 representativeof three ecologically distinct lake types Carbonate buffered-Oligotrophic-Weakly buffered soft waterndash Iconogr Diatomol 2 1ndash390
LUND JWG (1951) Contribution to our knowledge of British algae ndash Hydrobiologia 3(1) 93ndash100
ORGANIZATION FOR ECONOMIC COOPERATION AND DEVELOPMENT (1982)Eutrophication of Waters ndash Monitoring Assessment and Control Final Report OECD Paris
PANTOCSEK J (1901) Die Kieselalgen oder Bacillarien des Balaton 1ndash112 XVI Tafeln ndash Druckder K und K Hofbuchdruckerei des Victor Hornyaacutenszky Budapest
RAKOWSKA B (1996) Preliminary elaboration of the diatom communities occuring in the benthosof the Kamionka River and Lake Czarne at the Pojezierze Suwalskie (1993ndash1994) ndash Fragm FloristGeobot Polon 3 221ndash238
RIMET F JC DRUART amp O ANNEVILLE (2009) Exploring the dynamics of plankton diatomcommunities in Lake Geneva using emergent self-organizing maps (1974ndash2007) ndash Ecol Inform4 99ndash110
524
SCHMIDT R R PSENNER J MUumlLLER P INDINGER amp C KAMENIK (2002) Impact of thelate glacial climate variations on stratification and trophic state of the meromictic lake Laumlngsee (Austria)validation of a conceptual model by multiproxy studies ndash J Limnol 61(1) 49ndash60
SIEMIŃSKA J amp K WOŁOWSKI (2003) Catalogue of Polish prokaryotic and eukaryotic algaendash W Szafer Institute of Botany Polish Academy of Sciences Krakoacutew
SIVER PA (1999) Development of paleolimnological inference models for pH total nitrogen andspecific conductivity based on planktonic diatoms ndash J Paleolimnol 21 45ndash59
SOMMER U ZM GLIWICZ W LAMPERT amp A DUNCAN (1986) The PEG-model of seasonalsuccession of planktonic events in fresh waters ndash Arch Hydrobiol 106 433ndash471
TANAKA H (2007) Taxonomic studies of the genera Cyclotella (Kuumltzing) Breacutebisson DiscostellaHouk et Klee and Puncticulata Haringkansson in the family Stepanodiscaceae Glezer et Makarova(Bacillariophyta) in Japan ndash Biblioth Diatomol 53 1ndash205
TOLOTTI M D CALLIARI amp F CORRADINI (2004) Interannual variability of the phytoplanktonof Tovel lake (Trentino Italy) ndash Studi Trent di Scienze Naturali Acta Biol 81 Suppl 2 327ndash340 (inItalian)
WETZEL RG amp GE LIKENS (1991) Limnological analyses 1ndash391 ndash Springer-Verlag NewYork Berlin Heidelberg
WOJTAL AZ amp J KWANDRANS (2006) Diatoms of the Wyżyna Krakowsko-Częstochowskaupland (S Poland) ndash Coscinodiscophyceae (Thalassiosirophycidae) ndash Polish Bot J 51(2) 177ndash207
WUNSAM S R SCHMIDT amp R KLEE (1995) Cyclotella-taxa (Bacillariophyceae) in lakes of theAlpine region and their relationship to environmental variables ndash Aquat Sci 57(4) 360ndash381
Received 25 March 2010 accepted in revised form 31 March 2011

515
P balatonis contributed between 63 and 187 to the total phytoplankton numbersand between 21 and 47 to the total biomass From June until October the numberof the diatom did not exceed 300 frustules per 1 ml (Fig 24) and no valve wasfound in September At the beginning of December P balatonis number increasedslightly again
At the time of the bloom of P balatonis chrysoflagellates (Erkenia subaequiciliataSkuja Chrysococcus triporus Matvienko) pennate diatoms (Fragilaria sp) andorcoenobia-forming Chlorococcales (Didymocystis inconspicua Koršikov Tetastrumsp div Scenedesmus sp div Coelastrum microporum Naumlgeli Crucigenia tetrapediaKirchner) were also abundant Other diatoms such as Aulacoseira ambigua (Grunow)Simonsen Cyclotella meneghiniana Kuumltzing C ocellata Pantocsek Cyclostephanosdelicatus (Genkal) Casper C dubius (Fricke) Round Stephanodiscus parvus Stoermeramp Haringkansson Asterionella formosa Hassal and Navicula sp were also observed
Discussion
Morphology of Puncticulata balatonis (Pant) Wojtal et Budzyńska
Puncticulata balatonis shares important morphological characters assigned to thegenus justifying its formal transfer to this genus a slightly concentrically undulatemiddle part of the valve face with scattered or radially arranged numerous areolaewith internally domed cribra and numerous fultoportulae Additionally valves ofP balatonis have internal alveolar openings with marginal fultoportulae located onthicker costae rimoportulae located externally at the top of shortened striae andinternally in the marginal part of the valve face
The morphology of P balatonis valves from Rusałka Lake corresponds well withthe morphology of the specimens from the Pantocsekrsquos type material (Figs 13 1421 22 Table 3) though the diameter range and the number of striae in 10 microm werefound to be different Pantocsek (1901) stated in his diagnosis (p 104) that thevalves of Cyclotella balatonis are 16ndash20 microm in diameter but we have found bothsmaller and larger specimens (Table 3) The wider diameter range in our observationcould be related to a more complete morphological range of the species since thediatom population in Rusałka Lake was abundant However the time of samplingmay have also been of importance as the medium valve size of the species in our
(arrowed) at concentrically elevated valve face Fig 16 external view of the openings of central (right)and marginal (left) fultoportulae (arrowed) Fig 17 external view of the short and long striae of themarginal area Fig18 internal view of the rimoportula opening (arrowed) located close to the valvemargin and internal alveolar opening and central openings of numerous areolae and fultoportulae(arrowed) Fig 19 internal view of the divided thinner costa (marked) and internal opening of centralfultoportula (arrowed) Fig 20 internal view of the marginal fultoportulae located on the thicker costae(arrowed) Fig 21 external view of the valve face with marked notae Fig 22 internal view of thealveolar openings with thicker costa bearing marginal fultoportulae and internal rimoportula openinglocated near the margin (arrowed) Scale bar represents 5 microm for Figs 15 18 amp 21 4 microm for Fig 163 microm for Figs 19 amp 20 and 2 microm for Figs 17 amp 22
516
Tab
le 3
Com
pari
son
of P
unct
icul
ata
bala
toni
s an
d m
orph
olog
ical
ly s
imila
r sp
ecie
s C
F ndash
cent
ral
fulto
port
ulae
MF
ndash m
argi
nal
fulto
port
ulae
o
urob
serv
atio
n ty
pe m
ater
ial S
iofoacute
k s
ampl
e no
152
our
obse
rvat
ion
base
d on
the
fig
62
11
12
in K
ram
mer
amp L
ange
-Ber
talo
t 199
1 w
hich
sho
ws
spec
imen
s fr
om th
e ty
pe m
ater
ial o
f Cyc
lote
lla
bala
toni
s Pa
ntoc
sek
(as
C r
adio
sa)
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
Ref
eren
ces
Pant
ocse
kH
ouk
et a
lL
und
1951
H
aringkan
sson
H
ouk
et a
lH
aringkan
sson
H
ouk
et a
lH
aringkan
sson
H
ouk
et a
l19
01 p
104
20
10 p
39
p 9
820
02 p
116
20
10 p
41
2002
p 1
142
010
p 3
720
02 p
113
20
10 p
36
Plat
e X
V
Plat
es 2
69
Figs
1 2
Figs
422
ndashPl
ates
280
Fi
gs 4
15ndash
Plat
es 2
61
Figs
410
ndash P
late
s 25
7 1
ndashFi
gs 3
19
1ndash11
270
42
61ndash
11 2
81
421
1ndash11
262
41
423
258
1ndash1
133
21ndash
6 2
71
1ndash6
1ndash6
1ndash12
263
25
9 1
ndash12
1ndash5
264
1ndash6
260
1ndash8
Col
onie
sno
dat
a2ndash
82ndash
7 ce
lls2ndash
8 ce
lls
shor
tC
ells
solit
ary
no d
ata
mos
tlyce
lls s
olita
ryno
dat
alo
osel
ylo
osel
ylo
osel
yor
in s
hort
solit
ary
rare
ly in
unite
dun
ited
unite
d ch
ains
loos
ely
unite
dsh
ort c
hain
sch
ains
2(8
)ce
llsD
iam
eter
16ndash2
016
ndash22
87ndash
210
8ndash25
8ndash25
8ndash30
7ndash35
7ndash25
8ndash50
5ndash32
(microm
)St
riae
20 p
er 1
0 microm
ca
16
per
15ndash1
85
per
13ndash1
9 pe
r13
ndash15
per
15ndash1
8 pe
r 10
no d
ata
15ndash1
8 pe
r 10
no d
ata
14ndash1
8 pe
r10
microm
of
10 micro
m10
microm
10 micro
mmicrom
of n
earl
ymicrom
of n
early
10 micro
mne
arly
equ
aleq
ual l
engt
heq
ual l
engt
hle
ngth
occ
upy
occu
py 1
2ndash1
4ta
king
abo
ut1
3ndash1
2 of
of th
e va
lvae
12
of th
e va
lve
valv
e fa
cedi
amet
er fa
ce d
iam
eter
diam
eter
517
Cen
tral
are
a
flat
tofla
t to
slig
htly
flat t
o sl
ight
lyce
ntra
l are
aco
ncen
trica
llyfla
t or s
light
lysl
ight
ly c
on-
dist
inct
ly c
on-f
lat t
o co
n-fl
at to
con
-sl
ight
ly c
on-
conc
entri
cally
conc
entri
cally
with
mor
e or
undu
late
co
ncen
trica
llyce
ntri
cally
cent
rica
llyce
ntri
cally
cent
rica
llyce
ntric
ally
un-
undu
late
un
dula
te
less
regu
larl
yw
ith sc
atte
red
undu
late
un
dula
te
undu
late
and
undu
late
un
dula
te
dula
te w
ithw
ith C
F an
dw
ith n
ume-
arra
nged
or ra
dial
lyce
ntra
lra
dial
lyra
dial
lysm
ooth
smoo
th C
Fnu
mer
ous C
Far
eola
ero
us C
F an
dra
dial
arra
nged
CF
areo
lae
and
wrin
kled
C
Fw
rinkl
ed C
Fan
d ar
eola
ean
d ar
eola
ear
rang
ed in
areo
lae
ope-
punc
taan
d ar
eola
eC
F sc
atte
red
and
areo
lae
and
areo
lae
scat
tere
d or
open
ings
mor
e or
less
ning
s sca
ttere
dor
arr
ange
dar
rang
ed in
alw
ays
arra
nged
insc
atte
red
orra
dial
row
sor
occ
asio
nal-
in ra
dial
radi
al p
atte
rnar
rang
ed in
radi
al ro
ws
arra
nged
inly
arr
ange
d in
row
sra
dial
row
sra
dial
pat
tern
radi
al p
atte
rn(F
igs
8 amp
9)
Rim
o-2
ndash3 in
the
(1)2
ndash5 in
the
1ndash2(
3) in
the
no d
ata
at le
ast o
ne
1ndash5
in th
eat
leas
t one
at
leas
t one
in
at le
ast o
ne
at le
ast o
ne
port
ulae
mar
gina
lm
argi
nal
mar
gina
l
Inte
rnal
lyon
the
valv
aem
argi
nal a
rea
on th
e va
lve
the
mar
gina
lon
the v
alva
eon
the
valv
aear
ea n
ear t
hear
ea n
ear t
hear
ea n
ear t
hea
sess
ile la
bi-
face
at t
hecl
ose
to li
mit
face
at t
hear
ea c
lose
tofa
ce a
t the
face
at t
heva
lve
mar
gin
valv
e mar
gin
valv
e mar
gin
um in
obl
ique
end
of s
hor-
of th
e ce
ntra
len
d of
sho
r-lim
it of
the
end
of s
hor-
end
of s
light
lyE
xter
nal o
pe-
Ext
erna
lE
xter
nal
posi
tion
lo-
tene
d st
ria
area
Ext
erna
lte
ned
stri
ace
ntra
l are
ate
ned
stri
ash
orte
ned
stri
ani
ngs
at th
eop
enin
gs a
top
enin
gs a
tca
ted
betw
een
Inte
rnal
ly a
open
ings
on
Inte
rnal
ly a
Ext
erna
l ope
-In
tern
ally
aIn
tern
ally
a la
rge
end
of s
hor-
the
end
ofth
e en
d of
alve
olar
sess
ile la
bi-
a hy
alin
e ar
eare
lativ
ely
ning
s on
ala
rge
sess
ilese
ssile
labi
umte
ned
stri
ash
orte
ned
shor
tene
dop
enin
gsum
in ra
dial
at th
e en
d of
larg
e se
ssile
hyal
ine
area
labi
um in
in o
bliq
ue o
rIn
tern
ally
ast
ria
Inte
r-st
ria
Inte
r-an
d ce
ntra
lpo
sitio
n lo
-a
slig
htly
labi
um in
slit
at th
e en
dob
lique
po-
circ
umfe
rent
ial
smal
l ses
sile
nally
a se
ssile
nally
a sm
all
area
cate
d m
id-
shor
tene
dpo
sitio
n lo
-of
a s
light
lysi
tion
loca
-po
sitio
n lo
cate
dla
bium
clo
sela
bium
ver
yse
ssile
labi
-w
ay b
etw
een
stria
Int
erna
l-ca
ted
betw
een
shor
tene
dte
d be
twee
nin
the
mar
gina
lto
inte
rnal
clos
e to
inte
r-um
clo
se to
alve
olar
ope
-ly
a s
essi
leal
veol
ar o
pe-
stri
a In
ter-
alve
olar
ope
-ar
ea n
ear t
o th
eal
veol
ar o
pe-
nal a
lveo
lar
inte
rnal
alv
e-ni
ngs
and
labi
um in
ning
s an
dna
lly a
ses
-ni
ngs
and
cent
ral a
rea
ning
s w
ithop
enin
gs
olar
ope
ning
sce
ntra
l are
aob
lique
to c
ir-ce
ntra
l are
asi
le la
bium
cent
ral a
rea
radi
ally
or
with
radi
ally
with
radi
ally
cum
fere
ntia
lw
ith a
slit
ori-
obliq
uely
po-
or o
bliq
uely
or o
bliq
uely
slit
posi
tion
ente
d ob
lique
-si
tione
d sl
itpo
sitio
ned
posi
tione
d sl
itly
to n
earl
ysl
itci
rcum
fere
n-tia
llyIn
tern
alsm
all
cost
aesm
all
cost
aesm
all
cost
ae
radi
ally
radi
ally
elon
-ra
dial
ly e
lon-
smal
l co
stae
smal
l co
stae
smal
l co
stae
smal
l co
stae
alve
olar
bear
ing
MF
bear
ing
MF
bear
ing
MF
elon
gate
dga
ted
cos
tae
gate
d c
osta
ebe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fop
enin
gscl
early
larg
ercl
early
larg
ercl
early
larg
erco
stae
bea
-be
arin
g M
Fbe
arin
g M
Fcl
earl
y la
rger
clea
rly la
rger
clea
rly la
rger
clea
rly
larg
erth
an th
in ri
bsth
an th
in ri
bsth
an th
in ri
bsri
ng M
Fcl
early
larg
ersl
ight
ly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
than
thin
ribs
clea
rly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
518
Lak
e B
alat
onM
esot
roph
ic
Eut
roph
ic
Plan
kton
of a
no d
ata
Plan
kton
or
no d
ata
Plan
kton
of
no d
ata
foss
il di
atom
near
Sioacute
fok
eutr
ophi
chy
pere
utro
-nu
mbe
r of
litto
ral o
ligo
olig
om
eso-
lake
s re
ser
ph
ic re
serv
oir
lake
s po
orm
esot
roph
ictr
ophi
csl
owly
run
-in
org
anic
lake
sm
ainl
y su
b-ni
ng w
ater
sm
atte
r and
alpi
ne la
kes
One
of t
heso
me
quar
rym
ost c
omm
onpo
ols
inE
urop
ean
Eng
lish
Lak
esp
ecie
s of
the
Dis
tric
tC
-rad
iosa
com
plex
Tab
le 3
con
tinue
d
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
519
samples declined in subsequent winter-spring months which is a typical consequenceof diatom cell division The number of striae in 10 microm was found to be lower (11 to17 striae in 10 microm) in some specimens from Rusałka Lake than has been originallystated by Pantocsek (1901) It might be related to the striae counting method becauseeven at original Pantocseks drawings (Plate XV figs 319 332) number of striae in10 microm is lower when the Genkalrsquos method (1977) is applied It also remains inaccordance with our observations of the type material (Table 3) and the observationsin Houk et al (2010)
Puncticulata balatonis is morphologically similar to several species of the complexmaking it difficult for a non-specialist to distinguish them The species resemblesP radiosa but differs from it in valve face ultrastructure such as the location of theinternal rimoportula openings The valves of P radiosa have a more coarse andradially wrinkled central area in the valve face whereas P balatonis has a smootherand finely silicified valve face Additionally the radial pattern of the valve facefultoportulae and areolae is very distinct in P radiosa The external rimoportulaopenings of P balatonis are situated at the end of shortened stria well discernible inLM as gaps (notae) whereas according to Houk et al (2010) the external openingsof P radiosa are situated at the end of only slightly shortened striae The differencesare clearly visible from the internal valvae view because P balatonis has valve facerimoportula openings located close to the internal alveolar openings whereas the
Fig 23 The variation in Puncticulata balatonis valve size in Rusałka Lake in subsequent months in2007
520
rimoportula openings in P radiosa are located more close to the central area ofthe valves Such a close location of the internal rimoportula openings to the valvemargin is typical only for P balatonis within the discussed group (Table 3)
Another diatom showing a similar morphology to Puncticulata balatonis is P prae-termissa (Lund) Haringkansson Most of the morphological features are similar forboth species (Table 3) but the later possesses radially elongated relatively largeinternal alveolar openings Moreover the costae bearing marginal fultoportulaeare only slightly thicker than the rest and the internal rimoportula openings arecloser to the central part of the valve face (Table 3) The last feature as well as themarginal stria length enable also the morphological distinction between P balatonisand P comta
Ecology and distribution
The distinction between the species of the Puncticulata radiosa-complex includingPuncticulata balatonis is not easy for a plankton ecologist or other researcher whois not a centric diatom specialist The correct identification requires detailedmorphological studies with SEM andor specialized knowledge That is why apartfrom the long-lasting taxonomical confusion the real distribution of the speciesfrom the complex is uncertain According to Houk et al (2010) Puncticulata balatonis(as Cyclotella balatonis) is one of the most common members of the P radiosa (as
Fig 24 The number of Puncticulata balatonis frustules in the water column of Rusałka Lake in 2007
521
Cyclotella radiosa) complex in Europe found in numerous mesotrophic and eutrophiclakes and slowly running waters in Hungary Germany and Czech Republic Howeverreports of the species are not numerous Till now P balatonis has also never beenreported from Poland
The limited number of reports on the occurrence of P balatonis may be related to itsmisidentificationmisinterpretation as Cyclotella comta (Ehrenb) Kuumltz (Kiss amp Pająk1994 fig 13) found in the plankton of the eutrophic Vistula River as Cyclotellapraetermissa quadrijuncta (Wunsam et al 1995) or as Puncticulata radiosa (Wojtalamp Kwandrans 2006) found in the eutrophic Rudawa River We are convinced that apart of other records of P comta the most commonly reported species of the complexin Poland (Siemińska amp Wołowski 2003) in fact might concern P balatonis InRusałka Lake the species may have also been mistaken for P comta (asC comta) reported from the lake by Kotlińska (1976) It is also likely that P bala-tonis in Poland may have been misidentified as P praetermissa although the latterspecies was only reported several times including findings in the benthos of CzarneLake and the Kamionka River NE Poland (Rakowska 1996) in the plankton ofseveral lakes of the Drawa National Park NW Poland (Gołdyn amp Szeląg-Wasielewska2004) in a sand-pit lake in Krakoacutew and a spring fed pond in Krztynia in S Poland(Wojtal unpublished data)
Up to now there has also been much confusion around the ecology of the speciesfrom the complex For instance P praetermissa described by Lund (1951) fromlakes with a low organic matter content was later found both in the plankton and thebenthos in a wide variety of natural and man-made lakes including alpine andsubalpine lakes (Wunsam et al 1995 Lange-Bertalot amp Metzeltin 1996 Guumlttingeramp Straub 1998 Tolotti et al 2004) as well as in more eutrophic habitats (egSchmidt et al 2002 Hindaacutek amp Hindaacutekovaacute 2003 Tanaka 2007) Houk et al (2010)cleared up the ambiguities ascribing P praetermissa and P radiosa to oligo-mesotrophic habitats According to the authors P comta (as Cyclotella comta) is tillnow known only from fossil materials P balatonis (as Cyclotella balatonis) inturn has been assigned to meso-eutrophic lakes and rivers consistently with thetrophic character of its type locality ndash lake Balaton Our results correspond with theautecology of P balatonis (as Cyclotella balatonis) proposed by Houk et al (2010)widening even slightly the preferences of the species for the trophic conditionsdetermined by Houk et al (2010) as it was also able to develop a large populationin an eutrophic to hypereutrophic water body
The highest numbers of P balatonis in Rusałka Lake were observed in the winter-spring period The diatom bloom in the spring plankton is a well-known phenomenon(Sommer et al 1986) ndash sometimes also occurring during the winter period (Kiss ampGenkal 1993) ndash and is usually associated with a changing silica andor phosphorusavailability during that period (Lampert amp Sommer 2001) Sinking was also provedto be an essential factor hampering diatom success during the thermal stratificationperiod (Jewson et al 1981 Ferris amp Lehman 2007) explaining probably the seasonalityof the P balatonis growth in Rusałka Lake as its high development finished withthe stratification onset Relatively high numbers of frustules in deeper water layersmight also indicate a significant role of the sedimentation process
522
Conclusions
The morphology of Puncticulata balatonis in Rusałka Lake corresponds well toother species around P comta with the exception of the location of marginalfultoportula openings and the length of the shortened striae (notae) Analysis of themorphological characters enabled the transfer of Cyclotella balatonis into the genusPuncticulata A large population of the species in the eutrophic to hypereutrophicRusałka Lake remains congruent with its tolerance ranges with respect to the trophicstate proposed by Houk et al (2010) The variation in the cell number and averagedimension of frustules indicated the species preference to develop large populationsin spring
Acknowledgements
We thank Prof Ryszard Gołdyn (AMickiewicz University in Poznań) for his inspiration researchsupervision and useful comments to the manuscript We are also grateful to Dr Bart Van de Vijver andtwo anonymous reviewers for their valuable suggestions and to Dr Krisztina Buczko for enabling theanalysis of the type material of Cyclotella balatonis from the Pantocsekrsquos collection at HungarianNatural History Museum Sampling on Rusałka Lake was part of a project financially supported by theDistrict and Municipial Funds of Enviromental Protection and Water Management in Poznań (Poland)
References
CARLSON RE (1977) A trophic state index for lakes ndash Limnol Oceanogr 22 (2) 361ndash369
CREMER H amp H KOOLMEES (2010) Common diatoms (Centrales and Fragilariaceae Bacillario-phyta) of modern and fossil freshwater environments in the Netherlands ndash Nova Hedwigia 90(3ndash4)343ndash381
FERRIS JA amp JT LEHMAN (2007) Interannual variation in diatom bloom dynamics Roles ofhydrology nutrient limitation sinking and whole lake manipulation ndash Water Res 41 2551ndash2562
GENKAL SI (1977) On the method of calculation of some taxonomicaly significant structuralelements of the valve in the diatoms of the family Thalassiosiraceae Lebour emend Hasle (Bacillariophyta)ndash Bot Zhurn 62 848ndash851 (in Russian)
GENKAL SI amp KT KISS (1993) Morphological variability of the diatom Cyclotella atomusHustedt var atomus and C atomus var gracilis var nov ndash Hydrobiologia 269270 39ndash47
GOŁDYN R S PODSIADŁOWSKI K KOWALCZEWSKA-MADURA R DONDAJEWSKAE SZELĄG-WASIELEWSKA A BUDZYŃSKA P DOMEK amp W ROMANOWICZ (2010)Functioning of the Rusałka Lake ecosystem in Poznań (west Poland) ndash Ocean Hydrobiol Stud39(3) 65ndash80
GOŁDYN R amp E SZELĄG-WASIELEWSKA (2004) Changes in the phytoseston of a river-lakesystem in Drawieński National Park ndash Ocean Hydrobiol Stud 33(2) 17ndash28
GUumlTTINGER W amp F STRAUB (1998) Diatoms of Lake Cadagno ndash In PEDUZZI RR BACHOFEN amp M TONOLLA (eds) Lake Cadagno a meromictic Alpine lake ndash Doc Ist ItalIdrobiol 63 57ndash64
HANDMANN R (1913) Die Diatomeenflora des Almfeegebiets ndash In MANZ GJ (ed) Mitteilungendes Mikrologischen Vereins Linz ndash Mikrolog Ver Regensburg (in German)
HAUSMANN S amp AF LOTTER (2001) Morphological variation within the diatom taxon Cyclotellacomensis and its importance for quantitative temperature reconstructions ndash Freshw Biol 46 1323ndash1333
523
HAYASHI T (2007) Puncticulata versiformis sp nov and Cyclotella kathmanduensis sp nov(Bacillariophyta) new fossil species from middle Pleistocene lacustrine sediments Kathmandu NepalHilalaya ndash J Phycol 43(2) 304ndash318
HAringKANSSON H (2002) A compilation and evaluation of species in the genera StephanodiscusCyclostephanos and Cylotella with a new genus in the family Stephanodiscaceae ndash Diatom Res 17(1)1ndash139
HINDAacuteK F amp A HINDAacuteKOVAacute (2003) Diversity of cyanobacteria and algae of urban gravel pitlakes in Bratislava Slovakia a survey ndash Hydrobiologia 506ndash509 155ndash162
HINDAacuteK F amp A HINDAacuteKOVAacute (2008) Diversity of cyanobacteria and algae of the NationalNature Reserve Čiacutečovskeacute mŕtve rameno oxbow (Southern Slovakia) ndash Bulletin SBS 30 (1) 11ndash19 (inSlovak with an English abstract)
HOUK V R KLEE amp H TANAKA (2010) Atlas of freshwater centric diatoms with a brief key anddescriptions Part III Stephanodiscaceae A Cyclotella Tertiarius Discostella ndash Fottea 10 (Supplement)1ndash498
JEWSON DH BH RIPPEY amp WJK GILMORE (1981) Loss rates from sedimentation parasitismand grazing during the growth nutrient limitation and dormancy of a diatom crop ndash Limnol Oceanogr26 1045ndash1056
KISS KT amp SI GENKAL (1993) Winter blooms of centric diatoms in the River Danube and in itsside arms near Budapest ndash Hydrobiologia 269270 317ndash325
KISS KT amp G PAJĄK (1994) Seasonal change of diatom s in the plankton of the Vistula riverabove and below the Goczalkowice reservoir Poland ndash In KOCIOLEK JP (ed) Proceedings ofthe 11th International Diatom Symposium San Francisco Aug 12ndash17 1990 Memoirs of the CaliforniaAcademy of Sciences 17 517ndash524 San Francisco California Academy of Sciences
KOBAYASI H M IDEI S MAYAMA T NAGUMO amp K OSADA (2006) H KobayasirsquosAtlas of Japanese diatoms based on electron microscopy 1 1ndash531 Uchida Rokakuho Publishing CoLtd Tokyo
KOTLIŃSKA E (1976) Fitoplankton Jeziora Rusałka In Fitoplankton sztucznych jezior położonychna terenie Poznania 1ndash54 Poznańskie Tow Przyjacioacuteł Nauk Prace Komisji Biologicznej (in Polish)
KRAMMER K amp H LANGE-BERTALOT (1991) Bacillariophyceae 3 Teil Centrales Fragilaria-ceae Eunotiaceae ndash In ETTL H J GERLOFF H HEYNIGH amp D MOLLENHAUER (eds)Suumlsswasserflora von Mitteleuropa 2(3) 1ndash576 Gustav Fischer Verlag Stuttgart Jena
LAMPERT W amp U SOMMER (2001) Ekologia woacuted śroacutedlądowych ndash PWN Warszawa
LANGE-BERTALOT H amp D METZELTIN (1996) Indicators of oligotrophy 800 representativeof three ecologically distinct lake types Carbonate buffered-Oligotrophic-Weakly buffered soft waterndash Iconogr Diatomol 2 1ndash390
LUND JWG (1951) Contribution to our knowledge of British algae ndash Hydrobiologia 3(1) 93ndash100
ORGANIZATION FOR ECONOMIC COOPERATION AND DEVELOPMENT (1982)Eutrophication of Waters ndash Monitoring Assessment and Control Final Report OECD Paris
PANTOCSEK J (1901) Die Kieselalgen oder Bacillarien des Balaton 1ndash112 XVI Tafeln ndash Druckder K und K Hofbuchdruckerei des Victor Hornyaacutenszky Budapest
RAKOWSKA B (1996) Preliminary elaboration of the diatom communities occuring in the benthosof the Kamionka River and Lake Czarne at the Pojezierze Suwalskie (1993ndash1994) ndash Fragm FloristGeobot Polon 3 221ndash238
RIMET F JC DRUART amp O ANNEVILLE (2009) Exploring the dynamics of plankton diatomcommunities in Lake Geneva using emergent self-organizing maps (1974ndash2007) ndash Ecol Inform4 99ndash110
524
SCHMIDT R R PSENNER J MUumlLLER P INDINGER amp C KAMENIK (2002) Impact of thelate glacial climate variations on stratification and trophic state of the meromictic lake Laumlngsee (Austria)validation of a conceptual model by multiproxy studies ndash J Limnol 61(1) 49ndash60
SIEMIŃSKA J amp K WOŁOWSKI (2003) Catalogue of Polish prokaryotic and eukaryotic algaendash W Szafer Institute of Botany Polish Academy of Sciences Krakoacutew
SIVER PA (1999) Development of paleolimnological inference models for pH total nitrogen andspecific conductivity based on planktonic diatoms ndash J Paleolimnol 21 45ndash59
SOMMER U ZM GLIWICZ W LAMPERT amp A DUNCAN (1986) The PEG-model of seasonalsuccession of planktonic events in fresh waters ndash Arch Hydrobiol 106 433ndash471
TANAKA H (2007) Taxonomic studies of the genera Cyclotella (Kuumltzing) Breacutebisson DiscostellaHouk et Klee and Puncticulata Haringkansson in the family Stepanodiscaceae Glezer et Makarova(Bacillariophyta) in Japan ndash Biblioth Diatomol 53 1ndash205
TOLOTTI M D CALLIARI amp F CORRADINI (2004) Interannual variability of the phytoplanktonof Tovel lake (Trentino Italy) ndash Studi Trent di Scienze Naturali Acta Biol 81 Suppl 2 327ndash340 (inItalian)
WETZEL RG amp GE LIKENS (1991) Limnological analyses 1ndash391 ndash Springer-Verlag NewYork Berlin Heidelberg
WOJTAL AZ amp J KWANDRANS (2006) Diatoms of the Wyżyna Krakowsko-Częstochowskaupland (S Poland) ndash Coscinodiscophyceae (Thalassiosirophycidae) ndash Polish Bot J 51(2) 177ndash207
WUNSAM S R SCHMIDT amp R KLEE (1995) Cyclotella-taxa (Bacillariophyceae) in lakes of theAlpine region and their relationship to environmental variables ndash Aquat Sci 57(4) 360ndash381
Received 25 March 2010 accepted in revised form 31 March 2011

516
Tab
le 3
Com
pari
son
of P
unct
icul
ata
bala
toni
s an
d m
orph
olog
ical
ly s
imila
r sp
ecie
s C
F ndash
cent
ral
fulto
port
ulae
MF
ndash m
argi
nal
fulto
port
ulae
o
urob
serv
atio
n ty
pe m
ater
ial S
iofoacute
k s
ampl
e no
152
our
obse
rvat
ion
base
d on
the
fig
62
11
12
in K
ram
mer
amp L
ange
-Ber
talo
t 199
1 w
hich
sho
ws
spec
imen
s fr
om th
e ty
pe m
ater
ial o
f Cyc
lote
lla
bala
toni
s Pa
ntoc
sek
(as
C r
adio
sa)
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
Ref
eren
ces
Pant
ocse
kH
ouk
et a
lL
und
1951
H
aringkan
sson
H
ouk
et a
lH
aringkan
sson
H
ouk
et a
lH
aringkan
sson
H
ouk
et a
l19
01 p
104
20
10 p
39
p 9
820
02 p
116
20
10 p
41
2002
p 1
142
010
p 3
720
02 p
113
20
10 p
36
Plat
e X
V
Plat
es 2
69
Figs
1 2
Figs
422
ndashPl
ates
280
Fi
gs 4
15ndash
Plat
es 2
61
Figs
410
ndash P
late
s 25
7 1
ndashFi
gs 3
19
1ndash11
270
42
61ndash
11 2
81
421
1ndash11
262
41
423
258
1ndash1
133
21ndash
6 2
71
1ndash6
1ndash6
1ndash12
263
25
9 1
ndash12
1ndash5
264
1ndash6
260
1ndash8
Col
onie
sno
dat
a2ndash
82ndash
7 ce
lls2ndash
8 ce
lls
shor
tC
ells
solit
ary
no d
ata
mos
tlyce
lls s
olita
ryno
dat
alo
osel
ylo
osel
ylo
osel
yor
in s
hort
solit
ary
rare
ly in
unite
dun
ited
unite
d ch
ains
loos
ely
unite
dsh
ort c
hain
sch
ains
2(8
)ce
llsD
iam
eter
16ndash2
016
ndash22
87ndash
210
8ndash25
8ndash25
8ndash30
7ndash35
7ndash25
8ndash50
5ndash32
(microm
)St
riae
20 p
er 1
0 microm
ca
16
per
15ndash1
85
per
13ndash1
9 pe
r13
ndash15
per
15ndash1
8 pe
r 10
no d
ata
15ndash1
8 pe
r 10
no d
ata
14ndash1
8 pe
r10
microm
of
10 micro
m10
microm
10 micro
mmicrom
of n
earl
ymicrom
of n
early
10 micro
mne
arly
equ
aleq
ual l
engt
heq
ual l
engt
hle
ngth
occ
upy
occu
py 1
2ndash1
4ta
king
abo
ut1
3ndash1
2 of
of th
e va
lvae
12
of th
e va
lve
valv
e fa
cedi
amet
er fa
ce d
iam
eter
diam
eter
517
Cen
tral
are
a
flat
tofla
t to
slig
htly
flat t
o sl
ight
lyce
ntra
l are
aco
ncen
trica
llyfla
t or s
light
lysl
ight
ly c
on-
dist
inct
ly c
on-f
lat t
o co
n-fl
at to
con
-sl
ight
ly c
on-
conc
entri
cally
conc
entri
cally
with
mor
e or
undu
late
co
ncen
trica
llyce
ntri
cally
cent
rica
llyce
ntri
cally
cent
rica
llyce
ntric
ally
un-
undu
late
un
dula
te
less
regu
larl
yw
ith sc
atte
red
undu
late
un
dula
te
undu
late
and
undu
late
un
dula
te
dula
te w
ithw
ith C
F an
dw
ith n
ume-
arra
nged
or ra
dial
lyce
ntra
lra
dial
lyra
dial
lysm
ooth
smoo
th C
Fnu
mer
ous C
Far
eola
ero
us C
F an
dra
dial
arra
nged
CF
areo
lae
and
wrin
kled
C
Fw
rinkl
ed C
Fan
d ar
eola
ean
d ar
eola
ear
rang
ed in
areo
lae
ope-
punc
taan
d ar
eola
eC
F sc
atte
red
and
areo
lae
and
areo
lae
scat
tere
d or
open
ings
mor
e or
less
ning
s sca
ttere
dor
arr
ange
dar
rang
ed in
alw
ays
arra
nged
insc
atte
red
orra
dial
row
sor
occ
asio
nal-
in ra
dial
radi
al p
atte
rnar
rang
ed in
radi
al ro
ws
arra
nged
inly
arr
ange
d in
row
sra
dial
row
sra
dial
pat
tern
radi
al p
atte
rn(F
igs
8 amp
9)
Rim
o-2
ndash3 in
the
(1)2
ndash5 in
the
1ndash2(
3) in
the
no d
ata
at le
ast o
ne
1ndash5
in th
eat
leas
t one
at
leas
t one
in
at le
ast o
ne
at le
ast o
ne
port
ulae
mar
gina
lm
argi
nal
mar
gina
l
Inte
rnal
lyon
the
valv
aem
argi
nal a
rea
on th
e va
lve
the
mar
gina
lon
the v
alva
eon
the
valv
aear
ea n
ear t
hear
ea n
ear t
hear
ea n
ear t
hea
sess
ile la
bi-
face
at t
hecl
ose
to li
mit
face
at t
hear
ea c
lose
tofa
ce a
t the
face
at t
heva
lve
mar
gin
valv
e mar
gin
valv
e mar
gin
um in
obl
ique
end
of s
hor-
of th
e ce
ntra
len
d of
sho
r-lim
it of
the
end
of s
hor-
end
of s
light
lyE
xter
nal o
pe-
Ext
erna
lE
xter
nal
posi
tion
lo-
tene
d st
ria
area
Ext
erna
lte
ned
stri
ace
ntra
l are
ate
ned
stri
ash
orte
ned
stri
ani
ngs
at th
eop
enin
gs a
top
enin
gs a
tca
ted
betw
een
Inte
rnal
ly a
open
ings
on
Inte
rnal
ly a
Ext
erna
l ope
-In
tern
ally
aIn
tern
ally
a la
rge
end
of s
hor-
the
end
ofth
e en
d of
alve
olar
sess
ile la
bi-
a hy
alin
e ar
eare
lativ
ely
ning
s on
ala
rge
sess
ilese
ssile
labi
umte
ned
stri
ash
orte
ned
shor
tene
dop
enin
gsum
in ra
dial
at th
e en
d of
larg
e se
ssile
hyal
ine
area
labi
um in
in o
bliq
ue o
rIn
tern
ally
ast
ria
Inte
r-st
ria
Inte
r-an
d ce
ntra
lpo
sitio
n lo
-a
slig
htly
labi
um in
slit
at th
e en
dob
lique
po-
circ
umfe
rent
ial
smal
l ses
sile
nally
a se
ssile
nally
a sm
all
area
cate
d m
id-
shor
tene
dpo
sitio
n lo
-of
a s
light
lysi
tion
loca
-po
sitio
n lo
cate
dla
bium
clo
sela
bium
ver
yse
ssile
labi
-w
ay b
etw
een
stria
Int
erna
l-ca
ted
betw
een
shor
tene
dte
d be
twee
nin
the
mar
gina
lto
inte
rnal
clos
e to
inte
r-um
clo
se to
alve
olar
ope
-ly
a s
essi
leal
veol
ar o
pe-
stri
a In
ter-
alve
olar
ope
-ar
ea n
ear t
o th
eal
veol
ar o
pe-
nal a
lveo
lar
inte
rnal
alv
e-ni
ngs
and
labi
um in
ning
s an
dna
lly a
ses
-ni
ngs
and
cent
ral a
rea
ning
s w
ithop
enin
gs
olar
ope
ning
sce
ntra
l are
aob
lique
to c
ir-ce
ntra
l are
asi
le la
bium
cent
ral a
rea
radi
ally
or
with
radi
ally
with
radi
ally
cum
fere
ntia
lw
ith a
slit
ori-
obliq
uely
po-
or o
bliq
uely
or o
bliq
uely
slit
posi
tion
ente
d ob
lique
-si
tione
d sl
itpo
sitio
ned
posi
tione
d sl
itly
to n
earl
ysl
itci
rcum
fere
n-tia
llyIn
tern
alsm
all
cost
aesm
all
cost
aesm
all
cost
ae
radi
ally
radi
ally
elon
-ra
dial
ly e
lon-
smal
l co
stae
smal
l co
stae
smal
l co
stae
smal
l co
stae
alve
olar
bear
ing
MF
bear
ing
MF
bear
ing
MF
elon
gate
dga
ted
cos
tae
gate
d c
osta
ebe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fop
enin
gscl
early
larg
ercl
early
larg
ercl
early
larg
erco
stae
bea
-be
arin
g M
Fbe
arin
g M
Fcl
earl
y la
rger
clea
rly la
rger
clea
rly la
rger
clea
rly
larg
erth
an th
in ri
bsth
an th
in ri
bsth
an th
in ri
bsri
ng M
Fcl
early
larg
ersl
ight
ly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
than
thin
ribs
clea
rly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
518
Lak
e B
alat
onM
esot
roph
ic
Eut
roph
ic
Plan
kton
of a
no d
ata
Plan
kton
or
no d
ata
Plan
kton
of
no d
ata
foss
il di
atom
near
Sioacute
fok
eutr
ophi
chy
pere
utro
-nu
mbe
r of
litto
ral o
ligo
olig
om
eso-
lake
s re
ser
ph
ic re
serv
oir
lake
s po
orm
esot
roph
ictr
ophi
csl
owly
run
-in
org
anic
lake
sm
ainl
y su
b-ni
ng w
ater
sm
atte
r and
alpi
ne la
kes
One
of t
heso
me
quar
rym
ost c
omm
onpo
ols
inE
urop
ean
Eng
lish
Lak
esp
ecie
s of
the
Dis
tric
tC
-rad
iosa
com
plex
Tab
le 3
con
tinue
d
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
519
samples declined in subsequent winter-spring months which is a typical consequenceof diatom cell division The number of striae in 10 microm was found to be lower (11 to17 striae in 10 microm) in some specimens from Rusałka Lake than has been originallystated by Pantocsek (1901) It might be related to the striae counting method becauseeven at original Pantocseks drawings (Plate XV figs 319 332) number of striae in10 microm is lower when the Genkalrsquos method (1977) is applied It also remains inaccordance with our observations of the type material (Table 3) and the observationsin Houk et al (2010)
Puncticulata balatonis is morphologically similar to several species of the complexmaking it difficult for a non-specialist to distinguish them The species resemblesP radiosa but differs from it in valve face ultrastructure such as the location of theinternal rimoportula openings The valves of P radiosa have a more coarse andradially wrinkled central area in the valve face whereas P balatonis has a smootherand finely silicified valve face Additionally the radial pattern of the valve facefultoportulae and areolae is very distinct in P radiosa The external rimoportulaopenings of P balatonis are situated at the end of shortened stria well discernible inLM as gaps (notae) whereas according to Houk et al (2010) the external openingsof P radiosa are situated at the end of only slightly shortened striae The differencesare clearly visible from the internal valvae view because P balatonis has valve facerimoportula openings located close to the internal alveolar openings whereas the
Fig 23 The variation in Puncticulata balatonis valve size in Rusałka Lake in subsequent months in2007
520
rimoportula openings in P radiosa are located more close to the central area ofthe valves Such a close location of the internal rimoportula openings to the valvemargin is typical only for P balatonis within the discussed group (Table 3)
Another diatom showing a similar morphology to Puncticulata balatonis is P prae-termissa (Lund) Haringkansson Most of the morphological features are similar forboth species (Table 3) but the later possesses radially elongated relatively largeinternal alveolar openings Moreover the costae bearing marginal fultoportulaeare only slightly thicker than the rest and the internal rimoportula openings arecloser to the central part of the valve face (Table 3) The last feature as well as themarginal stria length enable also the morphological distinction between P balatonisand P comta
Ecology and distribution
The distinction between the species of the Puncticulata radiosa-complex includingPuncticulata balatonis is not easy for a plankton ecologist or other researcher whois not a centric diatom specialist The correct identification requires detailedmorphological studies with SEM andor specialized knowledge That is why apartfrom the long-lasting taxonomical confusion the real distribution of the speciesfrom the complex is uncertain According to Houk et al (2010) Puncticulata balatonis(as Cyclotella balatonis) is one of the most common members of the P radiosa (as
Fig 24 The number of Puncticulata balatonis frustules in the water column of Rusałka Lake in 2007
521
Cyclotella radiosa) complex in Europe found in numerous mesotrophic and eutrophiclakes and slowly running waters in Hungary Germany and Czech Republic Howeverreports of the species are not numerous Till now P balatonis has also never beenreported from Poland
The limited number of reports on the occurrence of P balatonis may be related to itsmisidentificationmisinterpretation as Cyclotella comta (Ehrenb) Kuumltz (Kiss amp Pająk1994 fig 13) found in the plankton of the eutrophic Vistula River as Cyclotellapraetermissa quadrijuncta (Wunsam et al 1995) or as Puncticulata radiosa (Wojtalamp Kwandrans 2006) found in the eutrophic Rudawa River We are convinced that apart of other records of P comta the most commonly reported species of the complexin Poland (Siemińska amp Wołowski 2003) in fact might concern P balatonis InRusałka Lake the species may have also been mistaken for P comta (asC comta) reported from the lake by Kotlińska (1976) It is also likely that P bala-tonis in Poland may have been misidentified as P praetermissa although the latterspecies was only reported several times including findings in the benthos of CzarneLake and the Kamionka River NE Poland (Rakowska 1996) in the plankton ofseveral lakes of the Drawa National Park NW Poland (Gołdyn amp Szeląg-Wasielewska2004) in a sand-pit lake in Krakoacutew and a spring fed pond in Krztynia in S Poland(Wojtal unpublished data)
Up to now there has also been much confusion around the ecology of the speciesfrom the complex For instance P praetermissa described by Lund (1951) fromlakes with a low organic matter content was later found both in the plankton and thebenthos in a wide variety of natural and man-made lakes including alpine andsubalpine lakes (Wunsam et al 1995 Lange-Bertalot amp Metzeltin 1996 Guumlttingeramp Straub 1998 Tolotti et al 2004) as well as in more eutrophic habitats (egSchmidt et al 2002 Hindaacutek amp Hindaacutekovaacute 2003 Tanaka 2007) Houk et al (2010)cleared up the ambiguities ascribing P praetermissa and P radiosa to oligo-mesotrophic habitats According to the authors P comta (as Cyclotella comta) is tillnow known only from fossil materials P balatonis (as Cyclotella balatonis) inturn has been assigned to meso-eutrophic lakes and rivers consistently with thetrophic character of its type locality ndash lake Balaton Our results correspond with theautecology of P balatonis (as Cyclotella balatonis) proposed by Houk et al (2010)widening even slightly the preferences of the species for the trophic conditionsdetermined by Houk et al (2010) as it was also able to develop a large populationin an eutrophic to hypereutrophic water body
The highest numbers of P balatonis in Rusałka Lake were observed in the winter-spring period The diatom bloom in the spring plankton is a well-known phenomenon(Sommer et al 1986) ndash sometimes also occurring during the winter period (Kiss ampGenkal 1993) ndash and is usually associated with a changing silica andor phosphorusavailability during that period (Lampert amp Sommer 2001) Sinking was also provedto be an essential factor hampering diatom success during the thermal stratificationperiod (Jewson et al 1981 Ferris amp Lehman 2007) explaining probably the seasonalityof the P balatonis growth in Rusałka Lake as its high development finished withthe stratification onset Relatively high numbers of frustules in deeper water layersmight also indicate a significant role of the sedimentation process
522
Conclusions
The morphology of Puncticulata balatonis in Rusałka Lake corresponds well toother species around P comta with the exception of the location of marginalfultoportula openings and the length of the shortened striae (notae) Analysis of themorphological characters enabled the transfer of Cyclotella balatonis into the genusPuncticulata A large population of the species in the eutrophic to hypereutrophicRusałka Lake remains congruent with its tolerance ranges with respect to the trophicstate proposed by Houk et al (2010) The variation in the cell number and averagedimension of frustules indicated the species preference to develop large populationsin spring
Acknowledgements
We thank Prof Ryszard Gołdyn (AMickiewicz University in Poznań) for his inspiration researchsupervision and useful comments to the manuscript We are also grateful to Dr Bart Van de Vijver andtwo anonymous reviewers for their valuable suggestions and to Dr Krisztina Buczko for enabling theanalysis of the type material of Cyclotella balatonis from the Pantocsekrsquos collection at HungarianNatural History Museum Sampling on Rusałka Lake was part of a project financially supported by theDistrict and Municipial Funds of Enviromental Protection and Water Management in Poznań (Poland)
References
CARLSON RE (1977) A trophic state index for lakes ndash Limnol Oceanogr 22 (2) 361ndash369
CREMER H amp H KOOLMEES (2010) Common diatoms (Centrales and Fragilariaceae Bacillario-phyta) of modern and fossil freshwater environments in the Netherlands ndash Nova Hedwigia 90(3ndash4)343ndash381
FERRIS JA amp JT LEHMAN (2007) Interannual variation in diatom bloom dynamics Roles ofhydrology nutrient limitation sinking and whole lake manipulation ndash Water Res 41 2551ndash2562
GENKAL SI (1977) On the method of calculation of some taxonomicaly significant structuralelements of the valve in the diatoms of the family Thalassiosiraceae Lebour emend Hasle (Bacillariophyta)ndash Bot Zhurn 62 848ndash851 (in Russian)
GENKAL SI amp KT KISS (1993) Morphological variability of the diatom Cyclotella atomusHustedt var atomus and C atomus var gracilis var nov ndash Hydrobiologia 269270 39ndash47
GOŁDYN R S PODSIADŁOWSKI K KOWALCZEWSKA-MADURA R DONDAJEWSKAE SZELĄG-WASIELEWSKA A BUDZYŃSKA P DOMEK amp W ROMANOWICZ (2010)Functioning of the Rusałka Lake ecosystem in Poznań (west Poland) ndash Ocean Hydrobiol Stud39(3) 65ndash80
GOŁDYN R amp E SZELĄG-WASIELEWSKA (2004) Changes in the phytoseston of a river-lakesystem in Drawieński National Park ndash Ocean Hydrobiol Stud 33(2) 17ndash28
GUumlTTINGER W amp F STRAUB (1998) Diatoms of Lake Cadagno ndash In PEDUZZI RR BACHOFEN amp M TONOLLA (eds) Lake Cadagno a meromictic Alpine lake ndash Doc Ist ItalIdrobiol 63 57ndash64
HANDMANN R (1913) Die Diatomeenflora des Almfeegebiets ndash In MANZ GJ (ed) Mitteilungendes Mikrologischen Vereins Linz ndash Mikrolog Ver Regensburg (in German)
HAUSMANN S amp AF LOTTER (2001) Morphological variation within the diatom taxon Cyclotellacomensis and its importance for quantitative temperature reconstructions ndash Freshw Biol 46 1323ndash1333
523
HAYASHI T (2007) Puncticulata versiformis sp nov and Cyclotella kathmanduensis sp nov(Bacillariophyta) new fossil species from middle Pleistocene lacustrine sediments Kathmandu NepalHilalaya ndash J Phycol 43(2) 304ndash318
HAringKANSSON H (2002) A compilation and evaluation of species in the genera StephanodiscusCyclostephanos and Cylotella with a new genus in the family Stephanodiscaceae ndash Diatom Res 17(1)1ndash139
HINDAacuteK F amp A HINDAacuteKOVAacute (2003) Diversity of cyanobacteria and algae of urban gravel pitlakes in Bratislava Slovakia a survey ndash Hydrobiologia 506ndash509 155ndash162
HINDAacuteK F amp A HINDAacuteKOVAacute (2008) Diversity of cyanobacteria and algae of the NationalNature Reserve Čiacutečovskeacute mŕtve rameno oxbow (Southern Slovakia) ndash Bulletin SBS 30 (1) 11ndash19 (inSlovak with an English abstract)
HOUK V R KLEE amp H TANAKA (2010) Atlas of freshwater centric diatoms with a brief key anddescriptions Part III Stephanodiscaceae A Cyclotella Tertiarius Discostella ndash Fottea 10 (Supplement)1ndash498
JEWSON DH BH RIPPEY amp WJK GILMORE (1981) Loss rates from sedimentation parasitismand grazing during the growth nutrient limitation and dormancy of a diatom crop ndash Limnol Oceanogr26 1045ndash1056
KISS KT amp SI GENKAL (1993) Winter blooms of centric diatoms in the River Danube and in itsside arms near Budapest ndash Hydrobiologia 269270 317ndash325
KISS KT amp G PAJĄK (1994) Seasonal change of diatom s in the plankton of the Vistula riverabove and below the Goczalkowice reservoir Poland ndash In KOCIOLEK JP (ed) Proceedings ofthe 11th International Diatom Symposium San Francisco Aug 12ndash17 1990 Memoirs of the CaliforniaAcademy of Sciences 17 517ndash524 San Francisco California Academy of Sciences
KOBAYASI H M IDEI S MAYAMA T NAGUMO amp K OSADA (2006) H KobayasirsquosAtlas of Japanese diatoms based on electron microscopy 1 1ndash531 Uchida Rokakuho Publishing CoLtd Tokyo
KOTLIŃSKA E (1976) Fitoplankton Jeziora Rusałka In Fitoplankton sztucznych jezior położonychna terenie Poznania 1ndash54 Poznańskie Tow Przyjacioacuteł Nauk Prace Komisji Biologicznej (in Polish)
KRAMMER K amp H LANGE-BERTALOT (1991) Bacillariophyceae 3 Teil Centrales Fragilaria-ceae Eunotiaceae ndash In ETTL H J GERLOFF H HEYNIGH amp D MOLLENHAUER (eds)Suumlsswasserflora von Mitteleuropa 2(3) 1ndash576 Gustav Fischer Verlag Stuttgart Jena
LAMPERT W amp U SOMMER (2001) Ekologia woacuted śroacutedlądowych ndash PWN Warszawa
LANGE-BERTALOT H amp D METZELTIN (1996) Indicators of oligotrophy 800 representativeof three ecologically distinct lake types Carbonate buffered-Oligotrophic-Weakly buffered soft waterndash Iconogr Diatomol 2 1ndash390
LUND JWG (1951) Contribution to our knowledge of British algae ndash Hydrobiologia 3(1) 93ndash100
ORGANIZATION FOR ECONOMIC COOPERATION AND DEVELOPMENT (1982)Eutrophication of Waters ndash Monitoring Assessment and Control Final Report OECD Paris
PANTOCSEK J (1901) Die Kieselalgen oder Bacillarien des Balaton 1ndash112 XVI Tafeln ndash Druckder K und K Hofbuchdruckerei des Victor Hornyaacutenszky Budapest
RAKOWSKA B (1996) Preliminary elaboration of the diatom communities occuring in the benthosof the Kamionka River and Lake Czarne at the Pojezierze Suwalskie (1993ndash1994) ndash Fragm FloristGeobot Polon 3 221ndash238
RIMET F JC DRUART amp O ANNEVILLE (2009) Exploring the dynamics of plankton diatomcommunities in Lake Geneva using emergent self-organizing maps (1974ndash2007) ndash Ecol Inform4 99ndash110
524
SCHMIDT R R PSENNER J MUumlLLER P INDINGER amp C KAMENIK (2002) Impact of thelate glacial climate variations on stratification and trophic state of the meromictic lake Laumlngsee (Austria)validation of a conceptual model by multiproxy studies ndash J Limnol 61(1) 49ndash60
SIEMIŃSKA J amp K WOŁOWSKI (2003) Catalogue of Polish prokaryotic and eukaryotic algaendash W Szafer Institute of Botany Polish Academy of Sciences Krakoacutew
SIVER PA (1999) Development of paleolimnological inference models for pH total nitrogen andspecific conductivity based on planktonic diatoms ndash J Paleolimnol 21 45ndash59
SOMMER U ZM GLIWICZ W LAMPERT amp A DUNCAN (1986) The PEG-model of seasonalsuccession of planktonic events in fresh waters ndash Arch Hydrobiol 106 433ndash471
TANAKA H (2007) Taxonomic studies of the genera Cyclotella (Kuumltzing) Breacutebisson DiscostellaHouk et Klee and Puncticulata Haringkansson in the family Stepanodiscaceae Glezer et Makarova(Bacillariophyta) in Japan ndash Biblioth Diatomol 53 1ndash205
TOLOTTI M D CALLIARI amp F CORRADINI (2004) Interannual variability of the phytoplanktonof Tovel lake (Trentino Italy) ndash Studi Trent di Scienze Naturali Acta Biol 81 Suppl 2 327ndash340 (inItalian)
WETZEL RG amp GE LIKENS (1991) Limnological analyses 1ndash391 ndash Springer-Verlag NewYork Berlin Heidelberg
WOJTAL AZ amp J KWANDRANS (2006) Diatoms of the Wyżyna Krakowsko-Częstochowskaupland (S Poland) ndash Coscinodiscophyceae (Thalassiosirophycidae) ndash Polish Bot J 51(2) 177ndash207
WUNSAM S R SCHMIDT amp R KLEE (1995) Cyclotella-taxa (Bacillariophyceae) in lakes of theAlpine region and their relationship to environmental variables ndash Aquat Sci 57(4) 360ndash381
Received 25 March 2010 accepted in revised form 31 March 2011

517
Cen
tral
are
a
flat
tofla
t to
slig
htly
flat t
o sl
ight
lyce
ntra
l are
aco
ncen
trica
llyfla
t or s
light
lysl
ight
ly c
on-
dist
inct
ly c
on-f
lat t
o co
n-fl
at to
con
-sl
ight
ly c
on-
conc
entri
cally
conc
entri
cally
with
mor
e or
undu
late
co
ncen
trica
llyce
ntri
cally
cent
rica
llyce
ntri
cally
cent
rica
llyce
ntric
ally
un-
undu
late
un
dula
te
less
regu
larl
yw
ith sc
atte
red
undu
late
un
dula
te
undu
late
and
undu
late
un
dula
te
dula
te w
ithw
ith C
F an
dw
ith n
ume-
arra
nged
or ra
dial
lyce
ntra
lra
dial
lyra
dial
lysm
ooth
smoo
th C
Fnu
mer
ous C
Far
eola
ero
us C
F an
dra
dial
arra
nged
CF
areo
lae
and
wrin
kled
C
Fw
rinkl
ed C
Fan
d ar
eola
ean
d ar
eola
ear
rang
ed in
areo
lae
ope-
punc
taan
d ar
eola
eC
F sc
atte
red
and
areo
lae
and
areo
lae
scat
tere
d or
open
ings
mor
e or
less
ning
s sca
ttere
dor
arr
ange
dar
rang
ed in
alw
ays
arra
nged
insc
atte
red
orra
dial
row
sor
occ
asio
nal-
in ra
dial
radi
al p
atte
rnar
rang
ed in
radi
al ro
ws
arra
nged
inly
arr
ange
d in
row
sra
dial
row
sra
dial
pat
tern
radi
al p
atte
rn(F
igs
8 amp
9)
Rim
o-2
ndash3 in
the
(1)2
ndash5 in
the
1ndash2(
3) in
the
no d
ata
at le
ast o
ne
1ndash5
in th
eat
leas
t one
at
leas
t one
in
at le
ast o
ne
at le
ast o
ne
port
ulae
mar
gina
lm
argi
nal
mar
gina
l
Inte
rnal
lyon
the
valv
aem
argi
nal a
rea
on th
e va
lve
the
mar
gina
lon
the v
alva
eon
the
valv
aear
ea n
ear t
hear
ea n
ear t
hear
ea n
ear t
hea
sess
ile la
bi-
face
at t
hecl
ose
to li
mit
face
at t
hear
ea c
lose
tofa
ce a
t the
face
at t
heva
lve
mar
gin
valv
e mar
gin
valv
e mar
gin
um in
obl
ique
end
of s
hor-
of th
e ce
ntra
len
d of
sho
r-lim
it of
the
end
of s
hor-
end
of s
light
lyE
xter
nal o
pe-
Ext
erna
lE
xter
nal
posi
tion
lo-
tene
d st
ria
area
Ext
erna
lte
ned
stri
ace
ntra
l are
ate
ned
stri
ash
orte
ned
stri
ani
ngs
at th
eop
enin
gs a
top
enin
gs a
tca
ted
betw
een
Inte
rnal
ly a
open
ings
on
Inte
rnal
ly a
Ext
erna
l ope
-In
tern
ally
aIn
tern
ally
a la
rge
end
of s
hor-
the
end
ofth
e en
d of
alve
olar
sess
ile la
bi-
a hy
alin
e ar
eare
lativ
ely
ning
s on
ala
rge
sess
ilese
ssile
labi
umte
ned
stri
ash
orte
ned
shor
tene
dop
enin
gsum
in ra
dial
at th
e en
d of
larg
e se
ssile
hyal
ine
area
labi
um in
in o
bliq
ue o
rIn
tern
ally
ast
ria
Inte
r-st
ria
Inte
r-an
d ce
ntra
lpo
sitio
n lo
-a
slig
htly
labi
um in
slit
at th
e en
dob
lique
po-
circ
umfe
rent
ial
smal
l ses
sile
nally
a se
ssile
nally
a sm
all
area
cate
d m
id-
shor
tene
dpo
sitio
n lo
-of
a s
light
lysi
tion
loca
-po
sitio
n lo
cate
dla
bium
clo
sela
bium
ver
yse
ssile
labi
-w
ay b
etw
een
stria
Int
erna
l-ca
ted
betw
een
shor
tene
dte
d be
twee
nin
the
mar
gina
lto
inte
rnal
clos
e to
inte
r-um
clo
se to
alve
olar
ope
-ly
a s
essi
leal
veol
ar o
pe-
stri
a In
ter-
alve
olar
ope
-ar
ea n
ear t
o th
eal
veol
ar o
pe-
nal a
lveo
lar
inte
rnal
alv
e-ni
ngs
and
labi
um in
ning
s an
dna
lly a
ses
-ni
ngs
and
cent
ral a
rea
ning
s w
ithop
enin
gs
olar
ope
ning
sce
ntra
l are
aob
lique
to c
ir-ce
ntra
l are
asi
le la
bium
cent
ral a
rea
radi
ally
or
with
radi
ally
with
radi
ally
cum
fere
ntia
lw
ith a
slit
ori-
obliq
uely
po-
or o
bliq
uely
or o
bliq
uely
slit
posi
tion
ente
d ob
lique
-si
tione
d sl
itpo
sitio
ned
posi
tione
d sl
itly
to n
earl
ysl
itci
rcum
fere
n-tia
llyIn
tern
alsm
all
cost
aesm
all
cost
aesm
all
cost
ae
radi
ally
radi
ally
elon
-ra
dial
ly e
lon-
smal
l co
stae
smal
l co
stae
smal
l co
stae
smal
l co
stae
alve
olar
bear
ing
MF
bear
ing
MF
bear
ing
MF
elon
gate
dga
ted
cos
tae
gate
d c
osta
ebe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fbe
arin
g M
Fop
enin
gscl
early
larg
ercl
early
larg
ercl
early
larg
erco
stae
bea
-be
arin
g M
Fbe
arin
g M
Fcl
earl
y la
rger
clea
rly la
rger
clea
rly la
rger
clea
rly
larg
erth
an th
in ri
bsth
an th
in ri
bsth
an th
in ri
bsri
ng M
Fcl
early
larg
ersl
ight
ly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
than
thin
ribs
clea
rly la
rger
than
thin
ribs
than
thin
ribs
than
thin
ribs
518
Lak
e B
alat
onM
esot
roph
ic
Eut
roph
ic
Plan
kton
of a
no d
ata
Plan
kton
or
no d
ata
Plan
kton
of
no d
ata
foss
il di
atom
near
Sioacute
fok
eutr
ophi
chy
pere
utro
-nu
mbe
r of
litto
ral o
ligo
olig
om
eso-
lake
s re
ser
ph
ic re
serv
oir
lake
s po
orm
esot
roph
ictr
ophi
csl
owly
run
-in
org
anic
lake
sm
ainl
y su
b-ni
ng w
ater
sm
atte
r and
alpi
ne la
kes
One
of t
heso
me
quar
rym
ost c
omm
onpo
ols
inE
urop
ean
Eng
lish
Lak
esp
ecie
s of
the
Dis
tric
tC
-rad
iosa
com
plex
Tab
le 3
con
tinue
d
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
519
samples declined in subsequent winter-spring months which is a typical consequenceof diatom cell division The number of striae in 10 microm was found to be lower (11 to17 striae in 10 microm) in some specimens from Rusałka Lake than has been originallystated by Pantocsek (1901) It might be related to the striae counting method becauseeven at original Pantocseks drawings (Plate XV figs 319 332) number of striae in10 microm is lower when the Genkalrsquos method (1977) is applied It also remains inaccordance with our observations of the type material (Table 3) and the observationsin Houk et al (2010)
Puncticulata balatonis is morphologically similar to several species of the complexmaking it difficult for a non-specialist to distinguish them The species resemblesP radiosa but differs from it in valve face ultrastructure such as the location of theinternal rimoportula openings The valves of P radiosa have a more coarse andradially wrinkled central area in the valve face whereas P balatonis has a smootherand finely silicified valve face Additionally the radial pattern of the valve facefultoportulae and areolae is very distinct in P radiosa The external rimoportulaopenings of P balatonis are situated at the end of shortened stria well discernible inLM as gaps (notae) whereas according to Houk et al (2010) the external openingsof P radiosa are situated at the end of only slightly shortened striae The differencesare clearly visible from the internal valvae view because P balatonis has valve facerimoportula openings located close to the internal alveolar openings whereas the
Fig 23 The variation in Puncticulata balatonis valve size in Rusałka Lake in subsequent months in2007
520
rimoportula openings in P radiosa are located more close to the central area ofthe valves Such a close location of the internal rimoportula openings to the valvemargin is typical only for P balatonis within the discussed group (Table 3)
Another diatom showing a similar morphology to Puncticulata balatonis is P prae-termissa (Lund) Haringkansson Most of the morphological features are similar forboth species (Table 3) but the later possesses radially elongated relatively largeinternal alveolar openings Moreover the costae bearing marginal fultoportulaeare only slightly thicker than the rest and the internal rimoportula openings arecloser to the central part of the valve face (Table 3) The last feature as well as themarginal stria length enable also the morphological distinction between P balatonisand P comta
Ecology and distribution
The distinction between the species of the Puncticulata radiosa-complex includingPuncticulata balatonis is not easy for a plankton ecologist or other researcher whois not a centric diatom specialist The correct identification requires detailedmorphological studies with SEM andor specialized knowledge That is why apartfrom the long-lasting taxonomical confusion the real distribution of the speciesfrom the complex is uncertain According to Houk et al (2010) Puncticulata balatonis(as Cyclotella balatonis) is one of the most common members of the P radiosa (as
Fig 24 The number of Puncticulata balatonis frustules in the water column of Rusałka Lake in 2007
521
Cyclotella radiosa) complex in Europe found in numerous mesotrophic and eutrophiclakes and slowly running waters in Hungary Germany and Czech Republic Howeverreports of the species are not numerous Till now P balatonis has also never beenreported from Poland
The limited number of reports on the occurrence of P balatonis may be related to itsmisidentificationmisinterpretation as Cyclotella comta (Ehrenb) Kuumltz (Kiss amp Pająk1994 fig 13) found in the plankton of the eutrophic Vistula River as Cyclotellapraetermissa quadrijuncta (Wunsam et al 1995) or as Puncticulata radiosa (Wojtalamp Kwandrans 2006) found in the eutrophic Rudawa River We are convinced that apart of other records of P comta the most commonly reported species of the complexin Poland (Siemińska amp Wołowski 2003) in fact might concern P balatonis InRusałka Lake the species may have also been mistaken for P comta (asC comta) reported from the lake by Kotlińska (1976) It is also likely that P bala-tonis in Poland may have been misidentified as P praetermissa although the latterspecies was only reported several times including findings in the benthos of CzarneLake and the Kamionka River NE Poland (Rakowska 1996) in the plankton ofseveral lakes of the Drawa National Park NW Poland (Gołdyn amp Szeląg-Wasielewska2004) in a sand-pit lake in Krakoacutew and a spring fed pond in Krztynia in S Poland(Wojtal unpublished data)
Up to now there has also been much confusion around the ecology of the speciesfrom the complex For instance P praetermissa described by Lund (1951) fromlakes with a low organic matter content was later found both in the plankton and thebenthos in a wide variety of natural and man-made lakes including alpine andsubalpine lakes (Wunsam et al 1995 Lange-Bertalot amp Metzeltin 1996 Guumlttingeramp Straub 1998 Tolotti et al 2004) as well as in more eutrophic habitats (egSchmidt et al 2002 Hindaacutek amp Hindaacutekovaacute 2003 Tanaka 2007) Houk et al (2010)cleared up the ambiguities ascribing P praetermissa and P radiosa to oligo-mesotrophic habitats According to the authors P comta (as Cyclotella comta) is tillnow known only from fossil materials P balatonis (as Cyclotella balatonis) inturn has been assigned to meso-eutrophic lakes and rivers consistently with thetrophic character of its type locality ndash lake Balaton Our results correspond with theautecology of P balatonis (as Cyclotella balatonis) proposed by Houk et al (2010)widening even slightly the preferences of the species for the trophic conditionsdetermined by Houk et al (2010) as it was also able to develop a large populationin an eutrophic to hypereutrophic water body
The highest numbers of P balatonis in Rusałka Lake were observed in the winter-spring period The diatom bloom in the spring plankton is a well-known phenomenon(Sommer et al 1986) ndash sometimes also occurring during the winter period (Kiss ampGenkal 1993) ndash and is usually associated with a changing silica andor phosphorusavailability during that period (Lampert amp Sommer 2001) Sinking was also provedto be an essential factor hampering diatom success during the thermal stratificationperiod (Jewson et al 1981 Ferris amp Lehman 2007) explaining probably the seasonalityof the P balatonis growth in Rusałka Lake as its high development finished withthe stratification onset Relatively high numbers of frustules in deeper water layersmight also indicate a significant role of the sedimentation process
522
Conclusions
The morphology of Puncticulata balatonis in Rusałka Lake corresponds well toother species around P comta with the exception of the location of marginalfultoportula openings and the length of the shortened striae (notae) Analysis of themorphological characters enabled the transfer of Cyclotella balatonis into the genusPuncticulata A large population of the species in the eutrophic to hypereutrophicRusałka Lake remains congruent with its tolerance ranges with respect to the trophicstate proposed by Houk et al (2010) The variation in the cell number and averagedimension of frustules indicated the species preference to develop large populationsin spring
Acknowledgements
We thank Prof Ryszard Gołdyn (AMickiewicz University in Poznań) for his inspiration researchsupervision and useful comments to the manuscript We are also grateful to Dr Bart Van de Vijver andtwo anonymous reviewers for their valuable suggestions and to Dr Krisztina Buczko for enabling theanalysis of the type material of Cyclotella balatonis from the Pantocsekrsquos collection at HungarianNatural History Museum Sampling on Rusałka Lake was part of a project financially supported by theDistrict and Municipial Funds of Enviromental Protection and Water Management in Poznań (Poland)
References
CARLSON RE (1977) A trophic state index for lakes ndash Limnol Oceanogr 22 (2) 361ndash369
CREMER H amp H KOOLMEES (2010) Common diatoms (Centrales and Fragilariaceae Bacillario-phyta) of modern and fossil freshwater environments in the Netherlands ndash Nova Hedwigia 90(3ndash4)343ndash381
FERRIS JA amp JT LEHMAN (2007) Interannual variation in diatom bloom dynamics Roles ofhydrology nutrient limitation sinking and whole lake manipulation ndash Water Res 41 2551ndash2562
GENKAL SI (1977) On the method of calculation of some taxonomicaly significant structuralelements of the valve in the diatoms of the family Thalassiosiraceae Lebour emend Hasle (Bacillariophyta)ndash Bot Zhurn 62 848ndash851 (in Russian)
GENKAL SI amp KT KISS (1993) Morphological variability of the diatom Cyclotella atomusHustedt var atomus and C atomus var gracilis var nov ndash Hydrobiologia 269270 39ndash47
GOŁDYN R S PODSIADŁOWSKI K KOWALCZEWSKA-MADURA R DONDAJEWSKAE SZELĄG-WASIELEWSKA A BUDZYŃSKA P DOMEK amp W ROMANOWICZ (2010)Functioning of the Rusałka Lake ecosystem in Poznań (west Poland) ndash Ocean Hydrobiol Stud39(3) 65ndash80
GOŁDYN R amp E SZELĄG-WASIELEWSKA (2004) Changes in the phytoseston of a river-lakesystem in Drawieński National Park ndash Ocean Hydrobiol Stud 33(2) 17ndash28
GUumlTTINGER W amp F STRAUB (1998) Diatoms of Lake Cadagno ndash In PEDUZZI RR BACHOFEN amp M TONOLLA (eds) Lake Cadagno a meromictic Alpine lake ndash Doc Ist ItalIdrobiol 63 57ndash64
HANDMANN R (1913) Die Diatomeenflora des Almfeegebiets ndash In MANZ GJ (ed) Mitteilungendes Mikrologischen Vereins Linz ndash Mikrolog Ver Regensburg (in German)
HAUSMANN S amp AF LOTTER (2001) Morphological variation within the diatom taxon Cyclotellacomensis and its importance for quantitative temperature reconstructions ndash Freshw Biol 46 1323ndash1333
523
HAYASHI T (2007) Puncticulata versiformis sp nov and Cyclotella kathmanduensis sp nov(Bacillariophyta) new fossil species from middle Pleistocene lacustrine sediments Kathmandu NepalHilalaya ndash J Phycol 43(2) 304ndash318
HAringKANSSON H (2002) A compilation and evaluation of species in the genera StephanodiscusCyclostephanos and Cylotella with a new genus in the family Stephanodiscaceae ndash Diatom Res 17(1)1ndash139
HINDAacuteK F amp A HINDAacuteKOVAacute (2003) Diversity of cyanobacteria and algae of urban gravel pitlakes in Bratislava Slovakia a survey ndash Hydrobiologia 506ndash509 155ndash162
HINDAacuteK F amp A HINDAacuteKOVAacute (2008) Diversity of cyanobacteria and algae of the NationalNature Reserve Čiacutečovskeacute mŕtve rameno oxbow (Southern Slovakia) ndash Bulletin SBS 30 (1) 11ndash19 (inSlovak with an English abstract)
HOUK V R KLEE amp H TANAKA (2010) Atlas of freshwater centric diatoms with a brief key anddescriptions Part III Stephanodiscaceae A Cyclotella Tertiarius Discostella ndash Fottea 10 (Supplement)1ndash498
JEWSON DH BH RIPPEY amp WJK GILMORE (1981) Loss rates from sedimentation parasitismand grazing during the growth nutrient limitation and dormancy of a diatom crop ndash Limnol Oceanogr26 1045ndash1056
KISS KT amp SI GENKAL (1993) Winter blooms of centric diatoms in the River Danube and in itsside arms near Budapest ndash Hydrobiologia 269270 317ndash325
KISS KT amp G PAJĄK (1994) Seasonal change of diatom s in the plankton of the Vistula riverabove and below the Goczalkowice reservoir Poland ndash In KOCIOLEK JP (ed) Proceedings ofthe 11th International Diatom Symposium San Francisco Aug 12ndash17 1990 Memoirs of the CaliforniaAcademy of Sciences 17 517ndash524 San Francisco California Academy of Sciences
KOBAYASI H M IDEI S MAYAMA T NAGUMO amp K OSADA (2006) H KobayasirsquosAtlas of Japanese diatoms based on electron microscopy 1 1ndash531 Uchida Rokakuho Publishing CoLtd Tokyo
KOTLIŃSKA E (1976) Fitoplankton Jeziora Rusałka In Fitoplankton sztucznych jezior położonychna terenie Poznania 1ndash54 Poznańskie Tow Przyjacioacuteł Nauk Prace Komisji Biologicznej (in Polish)
KRAMMER K amp H LANGE-BERTALOT (1991) Bacillariophyceae 3 Teil Centrales Fragilaria-ceae Eunotiaceae ndash In ETTL H J GERLOFF H HEYNIGH amp D MOLLENHAUER (eds)Suumlsswasserflora von Mitteleuropa 2(3) 1ndash576 Gustav Fischer Verlag Stuttgart Jena
LAMPERT W amp U SOMMER (2001) Ekologia woacuted śroacutedlądowych ndash PWN Warszawa
LANGE-BERTALOT H amp D METZELTIN (1996) Indicators of oligotrophy 800 representativeof three ecologically distinct lake types Carbonate buffered-Oligotrophic-Weakly buffered soft waterndash Iconogr Diatomol 2 1ndash390
LUND JWG (1951) Contribution to our knowledge of British algae ndash Hydrobiologia 3(1) 93ndash100
ORGANIZATION FOR ECONOMIC COOPERATION AND DEVELOPMENT (1982)Eutrophication of Waters ndash Monitoring Assessment and Control Final Report OECD Paris
PANTOCSEK J (1901) Die Kieselalgen oder Bacillarien des Balaton 1ndash112 XVI Tafeln ndash Druckder K und K Hofbuchdruckerei des Victor Hornyaacutenszky Budapest
RAKOWSKA B (1996) Preliminary elaboration of the diatom communities occuring in the benthosof the Kamionka River and Lake Czarne at the Pojezierze Suwalskie (1993ndash1994) ndash Fragm FloristGeobot Polon 3 221ndash238
RIMET F JC DRUART amp O ANNEVILLE (2009) Exploring the dynamics of plankton diatomcommunities in Lake Geneva using emergent self-organizing maps (1974ndash2007) ndash Ecol Inform4 99ndash110
524
SCHMIDT R R PSENNER J MUumlLLER P INDINGER amp C KAMENIK (2002) Impact of thelate glacial climate variations on stratification and trophic state of the meromictic lake Laumlngsee (Austria)validation of a conceptual model by multiproxy studies ndash J Limnol 61(1) 49ndash60
SIEMIŃSKA J amp K WOŁOWSKI (2003) Catalogue of Polish prokaryotic and eukaryotic algaendash W Szafer Institute of Botany Polish Academy of Sciences Krakoacutew
SIVER PA (1999) Development of paleolimnological inference models for pH total nitrogen andspecific conductivity based on planktonic diatoms ndash J Paleolimnol 21 45ndash59
SOMMER U ZM GLIWICZ W LAMPERT amp A DUNCAN (1986) The PEG-model of seasonalsuccession of planktonic events in fresh waters ndash Arch Hydrobiol 106 433ndash471
TANAKA H (2007) Taxonomic studies of the genera Cyclotella (Kuumltzing) Breacutebisson DiscostellaHouk et Klee and Puncticulata Haringkansson in the family Stepanodiscaceae Glezer et Makarova(Bacillariophyta) in Japan ndash Biblioth Diatomol 53 1ndash205
TOLOTTI M D CALLIARI amp F CORRADINI (2004) Interannual variability of the phytoplanktonof Tovel lake (Trentino Italy) ndash Studi Trent di Scienze Naturali Acta Biol 81 Suppl 2 327ndash340 (inItalian)
WETZEL RG amp GE LIKENS (1991) Limnological analyses 1ndash391 ndash Springer-Verlag NewYork Berlin Heidelberg
WOJTAL AZ amp J KWANDRANS (2006) Diatoms of the Wyżyna Krakowsko-Częstochowskaupland (S Poland) ndash Coscinodiscophyceae (Thalassiosirophycidae) ndash Polish Bot J 51(2) 177ndash207
WUNSAM S R SCHMIDT amp R KLEE (1995) Cyclotella-taxa (Bacillariophyceae) in lakes of theAlpine region and their relationship to environmental variables ndash Aquat Sci 57(4) 360ndash381
Received 25 March 2010 accepted in revised form 31 March 2011
518
Lak
e B
alat
onM
esot
roph
ic
Eut
roph
ic
Plan
kton
of a
no d
ata
Plan
kton
or
no d
ata
Plan
kton
of
no d
ata
foss
il di
atom
near
Sioacute
fok
eutr
ophi
chy
pere
utro
-nu
mbe
r of
litto
ral o
ligo
olig
om
eso-
lake
s re
ser
ph
ic re
serv
oir
lake
s po
orm
esot
roph
ictr
ophi
csl
owly
run
-in
org
anic
lake
sm
ainl
y su
b-ni
ng w
ater
sm
atte
r and
alpi
ne la
kes
One
of t
heso
me
quar
rym
ost c
omm
onpo
ols
inE
urop
ean
Eng
lish
Lak
esp
ecie
s of
the
Dis
tric
tC
-rad
iosa
com
plex
Tab
le 3
con
tinue
d
Cyc
lote
llaP
unct
icul
ata
Pun
ctic
ulat
aC
yclo
tella
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
Pun
ctic
ulat
aP
unct
icul
ata
bala
toni
sba
lato
nis
bala
toni
spr
aete
rmis
sapr
aete
rmis
sapr
aete
rmis
sara
dios
ara
dios
aco
mta
com
taPa
ntoc
sek
(Pan
tocs
ek)
(Pan
tocs
ek)
Lun
d(L
und)
(Lun
d)(L
emm
er-
(Lem
mer
-(E
hren
berg
)(E
hren
berg
)W
ojta
l et
Woj
tal e
tH
aringkan
sson
Haringk
anss
onm
ann)
man
n)H
aringkan
sson
Haringk
anss
onB
udzy
ńska
Bud
zyńs
kaas
Cyc
lote
llaH
aringkan
sson
Haringk
anss
on
as C
yclo
tella
as C
yclo
tella
our
prae
term
issa
as C
yclo
tella
com
taba
lato
nis
obse
rvat
ion
Lun
dra
dios
a(E
hren
berg
)Pa
ntoc
sek
(Gru
now
)K
uumltzi
ngL
emm
erm
ann
519
samples declined in subsequent winter-spring months which is a typical consequenceof diatom cell division The number of striae in 10 microm was found to be lower (11 to17 striae in 10 microm) in some specimens from Rusałka Lake than has been originallystated by Pantocsek (1901) It might be related to the striae counting method becauseeven at original Pantocseks drawings (Plate XV figs 319 332) number of striae in10 microm is lower when the Genkalrsquos method (1977) is applied It also remains inaccordance with our observations of the type material (Table 3) and the observationsin Houk et al (2010)
Puncticulata balatonis is morphologically similar to several species of the complexmaking it difficult for a non-specialist to distinguish them The species resemblesP radiosa but differs from it in valve face ultrastructure such as the location of theinternal rimoportula openings The valves of P radiosa have a more coarse andradially wrinkled central area in the valve face whereas P balatonis has a smootherand finely silicified valve face Additionally the radial pattern of the valve facefultoportulae and areolae is very distinct in P radiosa The external rimoportulaopenings of P balatonis are situated at the end of shortened stria well discernible inLM as gaps (notae) whereas according to Houk et al (2010) the external openingsof P radiosa are situated at the end of only slightly shortened striae The differencesare clearly visible from the internal valvae view because P balatonis has valve facerimoportula openings located close to the internal alveolar openings whereas the
Fig 23 The variation in Puncticulata balatonis valve size in Rusałka Lake in subsequent months in2007
520
rimoportula openings in P radiosa are located more close to the central area ofthe valves Such a close location of the internal rimoportula openings to the valvemargin is typical only for P balatonis within the discussed group (Table 3)
Another diatom showing a similar morphology to Puncticulata balatonis is P prae-termissa (Lund) Haringkansson Most of the morphological features are similar forboth species (Table 3) but the later possesses radially elongated relatively largeinternal alveolar openings Moreover the costae bearing marginal fultoportulaeare only slightly thicker than the rest and the internal rimoportula openings arecloser to the central part of the valve face (Table 3) The last feature as well as themarginal stria length enable also the morphological distinction between P balatonisand P comta
Ecology and distribution
The distinction between the species of the Puncticulata radiosa-complex includingPuncticulata balatonis is not easy for a plankton ecologist or other researcher whois not a centric diatom specialist The correct identification requires detailedmorphological studies with SEM andor specialized knowledge That is why apartfrom the long-lasting taxonomical confusion the real distribution of the speciesfrom the complex is uncertain According to Houk et al (2010) Puncticulata balatonis(as Cyclotella balatonis) is one of the most common members of the P radiosa (as
Fig 24 The number of Puncticulata balatonis frustules in the water column of Rusałka Lake in 2007
521
Cyclotella radiosa) complex in Europe found in numerous mesotrophic and eutrophiclakes and slowly running waters in Hungary Germany and Czech Republic Howeverreports of the species are not numerous Till now P balatonis has also never beenreported from Poland
The limited number of reports on the occurrence of P balatonis may be related to itsmisidentificationmisinterpretation as Cyclotella comta (Ehrenb) Kuumltz (Kiss amp Pająk1994 fig 13) found in the plankton of the eutrophic Vistula River as Cyclotellapraetermissa quadrijuncta (Wunsam et al 1995) or as Puncticulata radiosa (Wojtalamp Kwandrans 2006) found in the eutrophic Rudawa River We are convinced that apart of other records of P comta the most commonly reported species of the complexin Poland (Siemińska amp Wołowski 2003) in fact might concern P balatonis InRusałka Lake the species may have also been mistaken for P comta (asC comta) reported from the lake by Kotlińska (1976) It is also likely that P bala-tonis in Poland may have been misidentified as P praetermissa although the latterspecies was only reported several times including findings in the benthos of CzarneLake and the Kamionka River NE Poland (Rakowska 1996) in the plankton ofseveral lakes of the Drawa National Park NW Poland (Gołdyn amp Szeląg-Wasielewska2004) in a sand-pit lake in Krakoacutew and a spring fed pond in Krztynia in S Poland(Wojtal unpublished data)
Up to now there has also been much confusion around the ecology of the speciesfrom the complex For instance P praetermissa described by Lund (1951) fromlakes with a low organic matter content was later found both in the plankton and thebenthos in a wide variety of natural and man-made lakes including alpine andsubalpine lakes (Wunsam et al 1995 Lange-Bertalot amp Metzeltin 1996 Guumlttingeramp Straub 1998 Tolotti et al 2004) as well as in more eutrophic habitats (egSchmidt et al 2002 Hindaacutek amp Hindaacutekovaacute 2003 Tanaka 2007) Houk et al (2010)cleared up the ambiguities ascribing P praetermissa and P radiosa to oligo-mesotrophic habitats According to the authors P comta (as Cyclotella comta) is tillnow known only from fossil materials P balatonis (as Cyclotella balatonis) inturn has been assigned to meso-eutrophic lakes and rivers consistently with thetrophic character of its type locality ndash lake Balaton Our results correspond with theautecology of P balatonis (as Cyclotella balatonis) proposed by Houk et al (2010)widening even slightly the preferences of the species for the trophic conditionsdetermined by Houk et al (2010) as it was also able to develop a large populationin an eutrophic to hypereutrophic water body
The highest numbers of P balatonis in Rusałka Lake were observed in the winter-spring period The diatom bloom in the spring plankton is a well-known phenomenon(Sommer et al 1986) ndash sometimes also occurring during the winter period (Kiss ampGenkal 1993) ndash and is usually associated with a changing silica andor phosphorusavailability during that period (Lampert amp Sommer 2001) Sinking was also provedto be an essential factor hampering diatom success during the thermal stratificationperiod (Jewson et al 1981 Ferris amp Lehman 2007) explaining probably the seasonalityof the P balatonis growth in Rusałka Lake as its high development finished withthe stratification onset Relatively high numbers of frustules in deeper water layersmight also indicate a significant role of the sedimentation process
522
Conclusions
The morphology of Puncticulata balatonis in Rusałka Lake corresponds well toother species around P comta with the exception of the location of marginalfultoportula openings and the length of the shortened striae (notae) Analysis of themorphological characters enabled the transfer of Cyclotella balatonis into the genusPuncticulata A large population of the species in the eutrophic to hypereutrophicRusałka Lake remains congruent with its tolerance ranges with respect to the trophicstate proposed by Houk et al (2010) The variation in the cell number and averagedimension of frustules indicated the species preference to develop large populationsin spring
Acknowledgements
We thank Prof Ryszard Gołdyn (AMickiewicz University in Poznań) for his inspiration researchsupervision and useful comments to the manuscript We are also grateful to Dr Bart Van de Vijver andtwo anonymous reviewers for their valuable suggestions and to Dr Krisztina Buczko for enabling theanalysis of the type material of Cyclotella balatonis from the Pantocsekrsquos collection at HungarianNatural History Museum Sampling on Rusałka Lake was part of a project financially supported by theDistrict and Municipial Funds of Enviromental Protection and Water Management in Poznań (Poland)
References
CARLSON RE (1977) A trophic state index for lakes ndash Limnol Oceanogr 22 (2) 361ndash369
CREMER H amp H KOOLMEES (2010) Common diatoms (Centrales and Fragilariaceae Bacillario-phyta) of modern and fossil freshwater environments in the Netherlands ndash Nova Hedwigia 90(3ndash4)343ndash381
FERRIS JA amp JT LEHMAN (2007) Interannual variation in diatom bloom dynamics Roles ofhydrology nutrient limitation sinking and whole lake manipulation ndash Water Res 41 2551ndash2562
GENKAL SI (1977) On the method of calculation of some taxonomicaly significant structuralelements of the valve in the diatoms of the family Thalassiosiraceae Lebour emend Hasle (Bacillariophyta)ndash Bot Zhurn 62 848ndash851 (in Russian)
GENKAL SI amp KT KISS (1993) Morphological variability of the diatom Cyclotella atomusHustedt var atomus and C atomus var gracilis var nov ndash Hydrobiologia 269270 39ndash47
GOŁDYN R S PODSIADŁOWSKI K KOWALCZEWSKA-MADURA R DONDAJEWSKAE SZELĄG-WASIELEWSKA A BUDZYŃSKA P DOMEK amp W ROMANOWICZ (2010)Functioning of the Rusałka Lake ecosystem in Poznań (west Poland) ndash Ocean Hydrobiol Stud39(3) 65ndash80
GOŁDYN R amp E SZELĄG-WASIELEWSKA (2004) Changes in the phytoseston of a river-lakesystem in Drawieński National Park ndash Ocean Hydrobiol Stud 33(2) 17ndash28
GUumlTTINGER W amp F STRAUB (1998) Diatoms of Lake Cadagno ndash In PEDUZZI RR BACHOFEN amp M TONOLLA (eds) Lake Cadagno a meromictic Alpine lake ndash Doc Ist ItalIdrobiol 63 57ndash64
HANDMANN R (1913) Die Diatomeenflora des Almfeegebiets ndash In MANZ GJ (ed) Mitteilungendes Mikrologischen Vereins Linz ndash Mikrolog Ver Regensburg (in German)
HAUSMANN S amp AF LOTTER (2001) Morphological variation within the diatom taxon Cyclotellacomensis and its importance for quantitative temperature reconstructions ndash Freshw Biol 46 1323ndash1333
523
HAYASHI T (2007) Puncticulata versiformis sp nov and Cyclotella kathmanduensis sp nov(Bacillariophyta) new fossil species from middle Pleistocene lacustrine sediments Kathmandu NepalHilalaya ndash J Phycol 43(2) 304ndash318
HAringKANSSON H (2002) A compilation and evaluation of species in the genera StephanodiscusCyclostephanos and Cylotella with a new genus in the family Stephanodiscaceae ndash Diatom Res 17(1)1ndash139
HINDAacuteK F amp A HINDAacuteKOVAacute (2003) Diversity of cyanobacteria and algae of urban gravel pitlakes in Bratislava Slovakia a survey ndash Hydrobiologia 506ndash509 155ndash162
HINDAacuteK F amp A HINDAacuteKOVAacute (2008) Diversity of cyanobacteria and algae of the NationalNature Reserve Čiacutečovskeacute mŕtve rameno oxbow (Southern Slovakia) ndash Bulletin SBS 30 (1) 11ndash19 (inSlovak with an English abstract)
HOUK V R KLEE amp H TANAKA (2010) Atlas of freshwater centric diatoms with a brief key anddescriptions Part III Stephanodiscaceae A Cyclotella Tertiarius Discostella ndash Fottea 10 (Supplement)1ndash498
JEWSON DH BH RIPPEY amp WJK GILMORE (1981) Loss rates from sedimentation parasitismand grazing during the growth nutrient limitation and dormancy of a diatom crop ndash Limnol Oceanogr26 1045ndash1056
KISS KT amp SI GENKAL (1993) Winter blooms of centric diatoms in the River Danube and in itsside arms near Budapest ndash Hydrobiologia 269270 317ndash325
KISS KT amp G PAJĄK (1994) Seasonal change of diatom s in the plankton of the Vistula riverabove and below the Goczalkowice reservoir Poland ndash In KOCIOLEK JP (ed) Proceedings ofthe 11th International Diatom Symposium San Francisco Aug 12ndash17 1990 Memoirs of the CaliforniaAcademy of Sciences 17 517ndash524 San Francisco California Academy of Sciences
KOBAYASI H M IDEI S MAYAMA T NAGUMO amp K OSADA (2006) H KobayasirsquosAtlas of Japanese diatoms based on electron microscopy 1 1ndash531 Uchida Rokakuho Publishing CoLtd Tokyo
KOTLIŃSKA E (1976) Fitoplankton Jeziora Rusałka In Fitoplankton sztucznych jezior położonychna terenie Poznania 1ndash54 Poznańskie Tow Przyjacioacuteł Nauk Prace Komisji Biologicznej (in Polish)
KRAMMER K amp H LANGE-BERTALOT (1991) Bacillariophyceae 3 Teil Centrales Fragilaria-ceae Eunotiaceae ndash In ETTL H J GERLOFF H HEYNIGH amp D MOLLENHAUER (eds)Suumlsswasserflora von Mitteleuropa 2(3) 1ndash576 Gustav Fischer Verlag Stuttgart Jena
LAMPERT W amp U SOMMER (2001) Ekologia woacuted śroacutedlądowych ndash PWN Warszawa
LANGE-BERTALOT H amp D METZELTIN (1996) Indicators of oligotrophy 800 representativeof three ecologically distinct lake types Carbonate buffered-Oligotrophic-Weakly buffered soft waterndash Iconogr Diatomol 2 1ndash390
LUND JWG (1951) Contribution to our knowledge of British algae ndash Hydrobiologia 3(1) 93ndash100
ORGANIZATION FOR ECONOMIC COOPERATION AND DEVELOPMENT (1982)Eutrophication of Waters ndash Monitoring Assessment and Control Final Report OECD Paris
PANTOCSEK J (1901) Die Kieselalgen oder Bacillarien des Balaton 1ndash112 XVI Tafeln ndash Druckder K und K Hofbuchdruckerei des Victor Hornyaacutenszky Budapest
RAKOWSKA B (1996) Preliminary elaboration of the diatom communities occuring in the benthosof the Kamionka River and Lake Czarne at the Pojezierze Suwalskie (1993ndash1994) ndash Fragm FloristGeobot Polon 3 221ndash238
RIMET F JC DRUART amp O ANNEVILLE (2009) Exploring the dynamics of plankton diatomcommunities in Lake Geneva using emergent self-organizing maps (1974ndash2007) ndash Ecol Inform4 99ndash110
524
SCHMIDT R R PSENNER J MUumlLLER P INDINGER amp C KAMENIK (2002) Impact of thelate glacial climate variations on stratification and trophic state of the meromictic lake Laumlngsee (Austria)validation of a conceptual model by multiproxy studies ndash J Limnol 61(1) 49ndash60
SIEMIŃSKA J amp K WOŁOWSKI (2003) Catalogue of Polish prokaryotic and eukaryotic algaendash W Szafer Institute of Botany Polish Academy of Sciences Krakoacutew
SIVER PA (1999) Development of paleolimnological inference models for pH total nitrogen andspecific conductivity based on planktonic diatoms ndash J Paleolimnol 21 45ndash59
SOMMER U ZM GLIWICZ W LAMPERT amp A DUNCAN (1986) The PEG-model of seasonalsuccession of planktonic events in fresh waters ndash Arch Hydrobiol 106 433ndash471
TANAKA H (2007) Taxonomic studies of the genera Cyclotella (Kuumltzing) Breacutebisson DiscostellaHouk et Klee and Puncticulata Haringkansson in the family Stepanodiscaceae Glezer et Makarova(Bacillariophyta) in Japan ndash Biblioth Diatomol 53 1ndash205
TOLOTTI M D CALLIARI amp F CORRADINI (2004) Interannual variability of the phytoplanktonof Tovel lake (Trentino Italy) ndash Studi Trent di Scienze Naturali Acta Biol 81 Suppl 2 327ndash340 (inItalian)
WETZEL RG amp GE LIKENS (1991) Limnological analyses 1ndash391 ndash Springer-Verlag NewYork Berlin Heidelberg
WOJTAL AZ amp J KWANDRANS (2006) Diatoms of the Wyżyna Krakowsko-Częstochowskaupland (S Poland) ndash Coscinodiscophyceae (Thalassiosirophycidae) ndash Polish Bot J 51(2) 177ndash207
WUNSAM S R SCHMIDT amp R KLEE (1995) Cyclotella-taxa (Bacillariophyceae) in lakes of theAlpine region and their relationship to environmental variables ndash Aquat Sci 57(4) 360ndash381
Received 25 March 2010 accepted in revised form 31 March 2011

519
samples declined in subsequent winter-spring months which is a typical consequenceof diatom cell division The number of striae in 10 microm was found to be lower (11 to17 striae in 10 microm) in some specimens from Rusałka Lake than has been originallystated by Pantocsek (1901) It might be related to the striae counting method becauseeven at original Pantocseks drawings (Plate XV figs 319 332) number of striae in10 microm is lower when the Genkalrsquos method (1977) is applied It also remains inaccordance with our observations of the type material (Table 3) and the observationsin Houk et al (2010)
Puncticulata balatonis is morphologically similar to several species of the complexmaking it difficult for a non-specialist to distinguish them The species resemblesP radiosa but differs from it in valve face ultrastructure such as the location of theinternal rimoportula openings The valves of P radiosa have a more coarse andradially wrinkled central area in the valve face whereas P balatonis has a smootherand finely silicified valve face Additionally the radial pattern of the valve facefultoportulae and areolae is very distinct in P radiosa The external rimoportulaopenings of P balatonis are situated at the end of shortened stria well discernible inLM as gaps (notae) whereas according to Houk et al (2010) the external openingsof P radiosa are situated at the end of only slightly shortened striae The differencesare clearly visible from the internal valvae view because P balatonis has valve facerimoportula openings located close to the internal alveolar openings whereas the
Fig 23 The variation in Puncticulata balatonis valve size in Rusałka Lake in subsequent months in2007
520
rimoportula openings in P radiosa are located more close to the central area ofthe valves Such a close location of the internal rimoportula openings to the valvemargin is typical only for P balatonis within the discussed group (Table 3)
Another diatom showing a similar morphology to Puncticulata balatonis is P prae-termissa (Lund) Haringkansson Most of the morphological features are similar forboth species (Table 3) but the later possesses radially elongated relatively largeinternal alveolar openings Moreover the costae bearing marginal fultoportulaeare only slightly thicker than the rest and the internal rimoportula openings arecloser to the central part of the valve face (Table 3) The last feature as well as themarginal stria length enable also the morphological distinction between P balatonisand P comta
Ecology and distribution
The distinction between the species of the Puncticulata radiosa-complex includingPuncticulata balatonis is not easy for a plankton ecologist or other researcher whois not a centric diatom specialist The correct identification requires detailedmorphological studies with SEM andor specialized knowledge That is why apartfrom the long-lasting taxonomical confusion the real distribution of the speciesfrom the complex is uncertain According to Houk et al (2010) Puncticulata balatonis(as Cyclotella balatonis) is one of the most common members of the P radiosa (as
Fig 24 The number of Puncticulata balatonis frustules in the water column of Rusałka Lake in 2007
521
Cyclotella radiosa) complex in Europe found in numerous mesotrophic and eutrophiclakes and slowly running waters in Hungary Germany and Czech Republic Howeverreports of the species are not numerous Till now P balatonis has also never beenreported from Poland
The limited number of reports on the occurrence of P balatonis may be related to itsmisidentificationmisinterpretation as Cyclotella comta (Ehrenb) Kuumltz (Kiss amp Pająk1994 fig 13) found in the plankton of the eutrophic Vistula River as Cyclotellapraetermissa quadrijuncta (Wunsam et al 1995) or as Puncticulata radiosa (Wojtalamp Kwandrans 2006) found in the eutrophic Rudawa River We are convinced that apart of other records of P comta the most commonly reported species of the complexin Poland (Siemińska amp Wołowski 2003) in fact might concern P balatonis InRusałka Lake the species may have also been mistaken for P comta (asC comta) reported from the lake by Kotlińska (1976) It is also likely that P bala-tonis in Poland may have been misidentified as P praetermissa although the latterspecies was only reported several times including findings in the benthos of CzarneLake and the Kamionka River NE Poland (Rakowska 1996) in the plankton ofseveral lakes of the Drawa National Park NW Poland (Gołdyn amp Szeląg-Wasielewska2004) in a sand-pit lake in Krakoacutew and a spring fed pond in Krztynia in S Poland(Wojtal unpublished data)
Up to now there has also been much confusion around the ecology of the speciesfrom the complex For instance P praetermissa described by Lund (1951) fromlakes with a low organic matter content was later found both in the plankton and thebenthos in a wide variety of natural and man-made lakes including alpine andsubalpine lakes (Wunsam et al 1995 Lange-Bertalot amp Metzeltin 1996 Guumlttingeramp Straub 1998 Tolotti et al 2004) as well as in more eutrophic habitats (egSchmidt et al 2002 Hindaacutek amp Hindaacutekovaacute 2003 Tanaka 2007) Houk et al (2010)cleared up the ambiguities ascribing P praetermissa and P radiosa to oligo-mesotrophic habitats According to the authors P comta (as Cyclotella comta) is tillnow known only from fossil materials P balatonis (as Cyclotella balatonis) inturn has been assigned to meso-eutrophic lakes and rivers consistently with thetrophic character of its type locality ndash lake Balaton Our results correspond with theautecology of P balatonis (as Cyclotella balatonis) proposed by Houk et al (2010)widening even slightly the preferences of the species for the trophic conditionsdetermined by Houk et al (2010) as it was also able to develop a large populationin an eutrophic to hypereutrophic water body
The highest numbers of P balatonis in Rusałka Lake were observed in the winter-spring period The diatom bloom in the spring plankton is a well-known phenomenon(Sommer et al 1986) ndash sometimes also occurring during the winter period (Kiss ampGenkal 1993) ndash and is usually associated with a changing silica andor phosphorusavailability during that period (Lampert amp Sommer 2001) Sinking was also provedto be an essential factor hampering diatom success during the thermal stratificationperiod (Jewson et al 1981 Ferris amp Lehman 2007) explaining probably the seasonalityof the P balatonis growth in Rusałka Lake as its high development finished withthe stratification onset Relatively high numbers of frustules in deeper water layersmight also indicate a significant role of the sedimentation process
522
Conclusions
The morphology of Puncticulata balatonis in Rusałka Lake corresponds well toother species around P comta with the exception of the location of marginalfultoportula openings and the length of the shortened striae (notae) Analysis of themorphological characters enabled the transfer of Cyclotella balatonis into the genusPuncticulata A large population of the species in the eutrophic to hypereutrophicRusałka Lake remains congruent with its tolerance ranges with respect to the trophicstate proposed by Houk et al (2010) The variation in the cell number and averagedimension of frustules indicated the species preference to develop large populationsin spring
Acknowledgements
We thank Prof Ryszard Gołdyn (AMickiewicz University in Poznań) for his inspiration researchsupervision and useful comments to the manuscript We are also grateful to Dr Bart Van de Vijver andtwo anonymous reviewers for their valuable suggestions and to Dr Krisztina Buczko for enabling theanalysis of the type material of Cyclotella balatonis from the Pantocsekrsquos collection at HungarianNatural History Museum Sampling on Rusałka Lake was part of a project financially supported by theDistrict and Municipial Funds of Enviromental Protection and Water Management in Poznań (Poland)
References
CARLSON RE (1977) A trophic state index for lakes ndash Limnol Oceanogr 22 (2) 361ndash369
CREMER H amp H KOOLMEES (2010) Common diatoms (Centrales and Fragilariaceae Bacillario-phyta) of modern and fossil freshwater environments in the Netherlands ndash Nova Hedwigia 90(3ndash4)343ndash381
FERRIS JA amp JT LEHMAN (2007) Interannual variation in diatom bloom dynamics Roles ofhydrology nutrient limitation sinking and whole lake manipulation ndash Water Res 41 2551ndash2562
GENKAL SI (1977) On the method of calculation of some taxonomicaly significant structuralelements of the valve in the diatoms of the family Thalassiosiraceae Lebour emend Hasle (Bacillariophyta)ndash Bot Zhurn 62 848ndash851 (in Russian)
GENKAL SI amp KT KISS (1993) Morphological variability of the diatom Cyclotella atomusHustedt var atomus and C atomus var gracilis var nov ndash Hydrobiologia 269270 39ndash47
GOŁDYN R S PODSIADŁOWSKI K KOWALCZEWSKA-MADURA R DONDAJEWSKAE SZELĄG-WASIELEWSKA A BUDZYŃSKA P DOMEK amp W ROMANOWICZ (2010)Functioning of the Rusałka Lake ecosystem in Poznań (west Poland) ndash Ocean Hydrobiol Stud39(3) 65ndash80
GOŁDYN R amp E SZELĄG-WASIELEWSKA (2004) Changes in the phytoseston of a river-lakesystem in Drawieński National Park ndash Ocean Hydrobiol Stud 33(2) 17ndash28
GUumlTTINGER W amp F STRAUB (1998) Diatoms of Lake Cadagno ndash In PEDUZZI RR BACHOFEN amp M TONOLLA (eds) Lake Cadagno a meromictic Alpine lake ndash Doc Ist ItalIdrobiol 63 57ndash64
HANDMANN R (1913) Die Diatomeenflora des Almfeegebiets ndash In MANZ GJ (ed) Mitteilungendes Mikrologischen Vereins Linz ndash Mikrolog Ver Regensburg (in German)
HAUSMANN S amp AF LOTTER (2001) Morphological variation within the diatom taxon Cyclotellacomensis and its importance for quantitative temperature reconstructions ndash Freshw Biol 46 1323ndash1333
523
HAYASHI T (2007) Puncticulata versiformis sp nov and Cyclotella kathmanduensis sp nov(Bacillariophyta) new fossil species from middle Pleistocene lacustrine sediments Kathmandu NepalHilalaya ndash J Phycol 43(2) 304ndash318
HAringKANSSON H (2002) A compilation and evaluation of species in the genera StephanodiscusCyclostephanos and Cylotella with a new genus in the family Stephanodiscaceae ndash Diatom Res 17(1)1ndash139
HINDAacuteK F amp A HINDAacuteKOVAacute (2003) Diversity of cyanobacteria and algae of urban gravel pitlakes in Bratislava Slovakia a survey ndash Hydrobiologia 506ndash509 155ndash162
HINDAacuteK F amp A HINDAacuteKOVAacute (2008) Diversity of cyanobacteria and algae of the NationalNature Reserve Čiacutečovskeacute mŕtve rameno oxbow (Southern Slovakia) ndash Bulletin SBS 30 (1) 11ndash19 (inSlovak with an English abstract)
HOUK V R KLEE amp H TANAKA (2010) Atlas of freshwater centric diatoms with a brief key anddescriptions Part III Stephanodiscaceae A Cyclotella Tertiarius Discostella ndash Fottea 10 (Supplement)1ndash498
JEWSON DH BH RIPPEY amp WJK GILMORE (1981) Loss rates from sedimentation parasitismand grazing during the growth nutrient limitation and dormancy of a diatom crop ndash Limnol Oceanogr26 1045ndash1056
KISS KT amp SI GENKAL (1993) Winter blooms of centric diatoms in the River Danube and in itsside arms near Budapest ndash Hydrobiologia 269270 317ndash325
KISS KT amp G PAJĄK (1994) Seasonal change of diatom s in the plankton of the Vistula riverabove and below the Goczalkowice reservoir Poland ndash In KOCIOLEK JP (ed) Proceedings ofthe 11th International Diatom Symposium San Francisco Aug 12ndash17 1990 Memoirs of the CaliforniaAcademy of Sciences 17 517ndash524 San Francisco California Academy of Sciences
KOBAYASI H M IDEI S MAYAMA T NAGUMO amp K OSADA (2006) H KobayasirsquosAtlas of Japanese diatoms based on electron microscopy 1 1ndash531 Uchida Rokakuho Publishing CoLtd Tokyo
KOTLIŃSKA E (1976) Fitoplankton Jeziora Rusałka In Fitoplankton sztucznych jezior położonychna terenie Poznania 1ndash54 Poznańskie Tow Przyjacioacuteł Nauk Prace Komisji Biologicznej (in Polish)
KRAMMER K amp H LANGE-BERTALOT (1991) Bacillariophyceae 3 Teil Centrales Fragilaria-ceae Eunotiaceae ndash In ETTL H J GERLOFF H HEYNIGH amp D MOLLENHAUER (eds)Suumlsswasserflora von Mitteleuropa 2(3) 1ndash576 Gustav Fischer Verlag Stuttgart Jena
LAMPERT W amp U SOMMER (2001) Ekologia woacuted śroacutedlądowych ndash PWN Warszawa
LANGE-BERTALOT H amp D METZELTIN (1996) Indicators of oligotrophy 800 representativeof three ecologically distinct lake types Carbonate buffered-Oligotrophic-Weakly buffered soft waterndash Iconogr Diatomol 2 1ndash390
LUND JWG (1951) Contribution to our knowledge of British algae ndash Hydrobiologia 3(1) 93ndash100
ORGANIZATION FOR ECONOMIC COOPERATION AND DEVELOPMENT (1982)Eutrophication of Waters ndash Monitoring Assessment and Control Final Report OECD Paris
PANTOCSEK J (1901) Die Kieselalgen oder Bacillarien des Balaton 1ndash112 XVI Tafeln ndash Druckder K und K Hofbuchdruckerei des Victor Hornyaacutenszky Budapest
RAKOWSKA B (1996) Preliminary elaboration of the diatom communities occuring in the benthosof the Kamionka River and Lake Czarne at the Pojezierze Suwalskie (1993ndash1994) ndash Fragm FloristGeobot Polon 3 221ndash238
RIMET F JC DRUART amp O ANNEVILLE (2009) Exploring the dynamics of plankton diatomcommunities in Lake Geneva using emergent self-organizing maps (1974ndash2007) ndash Ecol Inform4 99ndash110
524
SCHMIDT R R PSENNER J MUumlLLER P INDINGER amp C KAMENIK (2002) Impact of thelate glacial climate variations on stratification and trophic state of the meromictic lake Laumlngsee (Austria)validation of a conceptual model by multiproxy studies ndash J Limnol 61(1) 49ndash60
SIEMIŃSKA J amp K WOŁOWSKI (2003) Catalogue of Polish prokaryotic and eukaryotic algaendash W Szafer Institute of Botany Polish Academy of Sciences Krakoacutew
SIVER PA (1999) Development of paleolimnological inference models for pH total nitrogen andspecific conductivity based on planktonic diatoms ndash J Paleolimnol 21 45ndash59
SOMMER U ZM GLIWICZ W LAMPERT amp A DUNCAN (1986) The PEG-model of seasonalsuccession of planktonic events in fresh waters ndash Arch Hydrobiol 106 433ndash471
TANAKA H (2007) Taxonomic studies of the genera Cyclotella (Kuumltzing) Breacutebisson DiscostellaHouk et Klee and Puncticulata Haringkansson in the family Stepanodiscaceae Glezer et Makarova(Bacillariophyta) in Japan ndash Biblioth Diatomol 53 1ndash205
TOLOTTI M D CALLIARI amp F CORRADINI (2004) Interannual variability of the phytoplanktonof Tovel lake (Trentino Italy) ndash Studi Trent di Scienze Naturali Acta Biol 81 Suppl 2 327ndash340 (inItalian)
WETZEL RG amp GE LIKENS (1991) Limnological analyses 1ndash391 ndash Springer-Verlag NewYork Berlin Heidelberg
WOJTAL AZ amp J KWANDRANS (2006) Diatoms of the Wyżyna Krakowsko-Częstochowskaupland (S Poland) ndash Coscinodiscophyceae (Thalassiosirophycidae) ndash Polish Bot J 51(2) 177ndash207
WUNSAM S R SCHMIDT amp R KLEE (1995) Cyclotella-taxa (Bacillariophyceae) in lakes of theAlpine region and their relationship to environmental variables ndash Aquat Sci 57(4) 360ndash381
Received 25 March 2010 accepted in revised form 31 March 2011
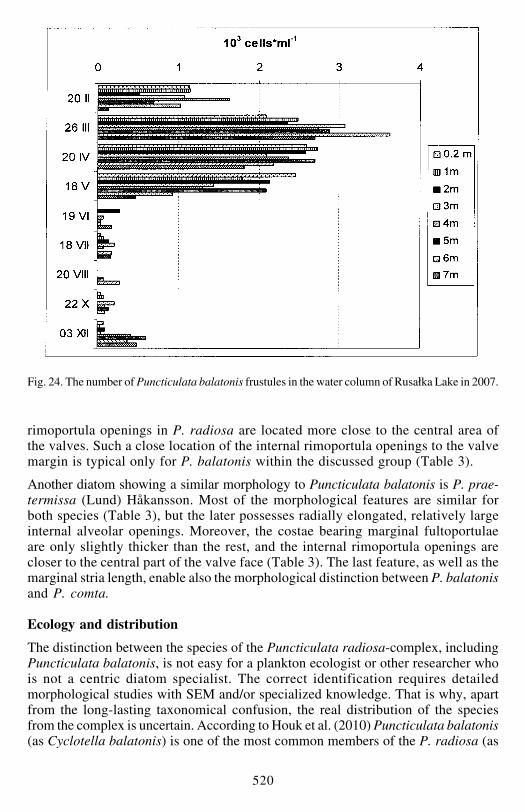
520
rimoportula openings in P radiosa are located more close to the central area ofthe valves Such a close location of the internal rimoportula openings to the valvemargin is typical only for P balatonis within the discussed group (Table 3)
Another diatom showing a similar morphology to Puncticulata balatonis is P prae-termissa (Lund) Haringkansson Most of the morphological features are similar forboth species (Table 3) but the later possesses radially elongated relatively largeinternal alveolar openings Moreover the costae bearing marginal fultoportulaeare only slightly thicker than the rest and the internal rimoportula openings arecloser to the central part of the valve face (Table 3) The last feature as well as themarginal stria length enable also the morphological distinction between P balatonisand P comta
Ecology and distribution
The distinction between the species of the Puncticulata radiosa-complex includingPuncticulata balatonis is not easy for a plankton ecologist or other researcher whois not a centric diatom specialist The correct identification requires detailedmorphological studies with SEM andor specialized knowledge That is why apartfrom the long-lasting taxonomical confusion the real distribution of the speciesfrom the complex is uncertain According to Houk et al (2010) Puncticulata balatonis(as Cyclotella balatonis) is one of the most common members of the P radiosa (as
Fig 24 The number of Puncticulata balatonis frustules in the water column of Rusałka Lake in 2007
521
Cyclotella radiosa) complex in Europe found in numerous mesotrophic and eutrophiclakes and slowly running waters in Hungary Germany and Czech Republic Howeverreports of the species are not numerous Till now P balatonis has also never beenreported from Poland
The limited number of reports on the occurrence of P balatonis may be related to itsmisidentificationmisinterpretation as Cyclotella comta (Ehrenb) Kuumltz (Kiss amp Pająk1994 fig 13) found in the plankton of the eutrophic Vistula River as Cyclotellapraetermissa quadrijuncta (Wunsam et al 1995) or as Puncticulata radiosa (Wojtalamp Kwandrans 2006) found in the eutrophic Rudawa River We are convinced that apart of other records of P comta the most commonly reported species of the complexin Poland (Siemińska amp Wołowski 2003) in fact might concern P balatonis InRusałka Lake the species may have also been mistaken for P comta (asC comta) reported from the lake by Kotlińska (1976) It is also likely that P bala-tonis in Poland may have been misidentified as P praetermissa although the latterspecies was only reported several times including findings in the benthos of CzarneLake and the Kamionka River NE Poland (Rakowska 1996) in the plankton ofseveral lakes of the Drawa National Park NW Poland (Gołdyn amp Szeląg-Wasielewska2004) in a sand-pit lake in Krakoacutew and a spring fed pond in Krztynia in S Poland(Wojtal unpublished data)
Up to now there has also been much confusion around the ecology of the speciesfrom the complex For instance P praetermissa described by Lund (1951) fromlakes with a low organic matter content was later found both in the plankton and thebenthos in a wide variety of natural and man-made lakes including alpine andsubalpine lakes (Wunsam et al 1995 Lange-Bertalot amp Metzeltin 1996 Guumlttingeramp Straub 1998 Tolotti et al 2004) as well as in more eutrophic habitats (egSchmidt et al 2002 Hindaacutek amp Hindaacutekovaacute 2003 Tanaka 2007) Houk et al (2010)cleared up the ambiguities ascribing P praetermissa and P radiosa to oligo-mesotrophic habitats According to the authors P comta (as Cyclotella comta) is tillnow known only from fossil materials P balatonis (as Cyclotella balatonis) inturn has been assigned to meso-eutrophic lakes and rivers consistently with thetrophic character of its type locality ndash lake Balaton Our results correspond with theautecology of P balatonis (as Cyclotella balatonis) proposed by Houk et al (2010)widening even slightly the preferences of the species for the trophic conditionsdetermined by Houk et al (2010) as it was also able to develop a large populationin an eutrophic to hypereutrophic water body
The highest numbers of P balatonis in Rusałka Lake were observed in the winter-spring period The diatom bloom in the spring plankton is a well-known phenomenon(Sommer et al 1986) ndash sometimes also occurring during the winter period (Kiss ampGenkal 1993) ndash and is usually associated with a changing silica andor phosphorusavailability during that period (Lampert amp Sommer 2001) Sinking was also provedto be an essential factor hampering diatom success during the thermal stratificationperiod (Jewson et al 1981 Ferris amp Lehman 2007) explaining probably the seasonalityof the P balatonis growth in Rusałka Lake as its high development finished withthe stratification onset Relatively high numbers of frustules in deeper water layersmight also indicate a significant role of the sedimentation process
522
Conclusions
The morphology of Puncticulata balatonis in Rusałka Lake corresponds well toother species around P comta with the exception of the location of marginalfultoportula openings and the length of the shortened striae (notae) Analysis of themorphological characters enabled the transfer of Cyclotella balatonis into the genusPuncticulata A large population of the species in the eutrophic to hypereutrophicRusałka Lake remains congruent with its tolerance ranges with respect to the trophicstate proposed by Houk et al (2010) The variation in the cell number and averagedimension of frustules indicated the species preference to develop large populationsin spring
Acknowledgements
We thank Prof Ryszard Gołdyn (AMickiewicz University in Poznań) for his inspiration researchsupervision and useful comments to the manuscript We are also grateful to Dr Bart Van de Vijver andtwo anonymous reviewers for their valuable suggestions and to Dr Krisztina Buczko for enabling theanalysis of the type material of Cyclotella balatonis from the Pantocsekrsquos collection at HungarianNatural History Museum Sampling on Rusałka Lake was part of a project financially supported by theDistrict and Municipial Funds of Enviromental Protection and Water Management in Poznań (Poland)
References
CARLSON RE (1977) A trophic state index for lakes ndash Limnol Oceanogr 22 (2) 361ndash369
CREMER H amp H KOOLMEES (2010) Common diatoms (Centrales and Fragilariaceae Bacillario-phyta) of modern and fossil freshwater environments in the Netherlands ndash Nova Hedwigia 90(3ndash4)343ndash381
FERRIS JA amp JT LEHMAN (2007) Interannual variation in diatom bloom dynamics Roles ofhydrology nutrient limitation sinking and whole lake manipulation ndash Water Res 41 2551ndash2562
GENKAL SI (1977) On the method of calculation of some taxonomicaly significant structuralelements of the valve in the diatoms of the family Thalassiosiraceae Lebour emend Hasle (Bacillariophyta)ndash Bot Zhurn 62 848ndash851 (in Russian)
GENKAL SI amp KT KISS (1993) Morphological variability of the diatom Cyclotella atomusHustedt var atomus and C atomus var gracilis var nov ndash Hydrobiologia 269270 39ndash47
GOŁDYN R S PODSIADŁOWSKI K KOWALCZEWSKA-MADURA R DONDAJEWSKAE SZELĄG-WASIELEWSKA A BUDZYŃSKA P DOMEK amp W ROMANOWICZ (2010)Functioning of the Rusałka Lake ecosystem in Poznań (west Poland) ndash Ocean Hydrobiol Stud39(3) 65ndash80
GOŁDYN R amp E SZELĄG-WASIELEWSKA (2004) Changes in the phytoseston of a river-lakesystem in Drawieński National Park ndash Ocean Hydrobiol Stud 33(2) 17ndash28
GUumlTTINGER W amp F STRAUB (1998) Diatoms of Lake Cadagno ndash In PEDUZZI RR BACHOFEN amp M TONOLLA (eds) Lake Cadagno a meromictic Alpine lake ndash Doc Ist ItalIdrobiol 63 57ndash64
HANDMANN R (1913) Die Diatomeenflora des Almfeegebiets ndash In MANZ GJ (ed) Mitteilungendes Mikrologischen Vereins Linz ndash Mikrolog Ver Regensburg (in German)
HAUSMANN S amp AF LOTTER (2001) Morphological variation within the diatom taxon Cyclotellacomensis and its importance for quantitative temperature reconstructions ndash Freshw Biol 46 1323ndash1333
523
HAYASHI T (2007) Puncticulata versiformis sp nov and Cyclotella kathmanduensis sp nov(Bacillariophyta) new fossil species from middle Pleistocene lacustrine sediments Kathmandu NepalHilalaya ndash J Phycol 43(2) 304ndash318
HAringKANSSON H (2002) A compilation and evaluation of species in the genera StephanodiscusCyclostephanos and Cylotella with a new genus in the family Stephanodiscaceae ndash Diatom Res 17(1)1ndash139
HINDAacuteK F amp A HINDAacuteKOVAacute (2003) Diversity of cyanobacteria and algae of urban gravel pitlakes in Bratislava Slovakia a survey ndash Hydrobiologia 506ndash509 155ndash162
HINDAacuteK F amp A HINDAacuteKOVAacute (2008) Diversity of cyanobacteria and algae of the NationalNature Reserve Čiacutečovskeacute mŕtve rameno oxbow (Southern Slovakia) ndash Bulletin SBS 30 (1) 11ndash19 (inSlovak with an English abstract)
HOUK V R KLEE amp H TANAKA (2010) Atlas of freshwater centric diatoms with a brief key anddescriptions Part III Stephanodiscaceae A Cyclotella Tertiarius Discostella ndash Fottea 10 (Supplement)1ndash498
JEWSON DH BH RIPPEY amp WJK GILMORE (1981) Loss rates from sedimentation parasitismand grazing during the growth nutrient limitation and dormancy of a diatom crop ndash Limnol Oceanogr26 1045ndash1056
KISS KT amp SI GENKAL (1993) Winter blooms of centric diatoms in the River Danube and in itsside arms near Budapest ndash Hydrobiologia 269270 317ndash325
KISS KT amp G PAJĄK (1994) Seasonal change of diatom s in the plankton of the Vistula riverabove and below the Goczalkowice reservoir Poland ndash In KOCIOLEK JP (ed) Proceedings ofthe 11th International Diatom Symposium San Francisco Aug 12ndash17 1990 Memoirs of the CaliforniaAcademy of Sciences 17 517ndash524 San Francisco California Academy of Sciences
KOBAYASI H M IDEI S MAYAMA T NAGUMO amp K OSADA (2006) H KobayasirsquosAtlas of Japanese diatoms based on electron microscopy 1 1ndash531 Uchida Rokakuho Publishing CoLtd Tokyo
KOTLIŃSKA E (1976) Fitoplankton Jeziora Rusałka In Fitoplankton sztucznych jezior położonychna terenie Poznania 1ndash54 Poznańskie Tow Przyjacioacuteł Nauk Prace Komisji Biologicznej (in Polish)
KRAMMER K amp H LANGE-BERTALOT (1991) Bacillariophyceae 3 Teil Centrales Fragilaria-ceae Eunotiaceae ndash In ETTL H J GERLOFF H HEYNIGH amp D MOLLENHAUER (eds)Suumlsswasserflora von Mitteleuropa 2(3) 1ndash576 Gustav Fischer Verlag Stuttgart Jena
LAMPERT W amp U SOMMER (2001) Ekologia woacuted śroacutedlądowych ndash PWN Warszawa
LANGE-BERTALOT H amp D METZELTIN (1996) Indicators of oligotrophy 800 representativeof three ecologically distinct lake types Carbonate buffered-Oligotrophic-Weakly buffered soft waterndash Iconogr Diatomol 2 1ndash390
LUND JWG (1951) Contribution to our knowledge of British algae ndash Hydrobiologia 3(1) 93ndash100
ORGANIZATION FOR ECONOMIC COOPERATION AND DEVELOPMENT (1982)Eutrophication of Waters ndash Monitoring Assessment and Control Final Report OECD Paris
PANTOCSEK J (1901) Die Kieselalgen oder Bacillarien des Balaton 1ndash112 XVI Tafeln ndash Druckder K und K Hofbuchdruckerei des Victor Hornyaacutenszky Budapest
RAKOWSKA B (1996) Preliminary elaboration of the diatom communities occuring in the benthosof the Kamionka River and Lake Czarne at the Pojezierze Suwalskie (1993ndash1994) ndash Fragm FloristGeobot Polon 3 221ndash238
RIMET F JC DRUART amp O ANNEVILLE (2009) Exploring the dynamics of plankton diatomcommunities in Lake Geneva using emergent self-organizing maps (1974ndash2007) ndash Ecol Inform4 99ndash110
524
SCHMIDT R R PSENNER J MUumlLLER P INDINGER amp C KAMENIK (2002) Impact of thelate glacial climate variations on stratification and trophic state of the meromictic lake Laumlngsee (Austria)validation of a conceptual model by multiproxy studies ndash J Limnol 61(1) 49ndash60
SIEMIŃSKA J amp K WOŁOWSKI (2003) Catalogue of Polish prokaryotic and eukaryotic algaendash W Szafer Institute of Botany Polish Academy of Sciences Krakoacutew
SIVER PA (1999) Development of paleolimnological inference models for pH total nitrogen andspecific conductivity based on planktonic diatoms ndash J Paleolimnol 21 45ndash59
SOMMER U ZM GLIWICZ W LAMPERT amp A DUNCAN (1986) The PEG-model of seasonalsuccession of planktonic events in fresh waters ndash Arch Hydrobiol 106 433ndash471
TANAKA H (2007) Taxonomic studies of the genera Cyclotella (Kuumltzing) Breacutebisson DiscostellaHouk et Klee and Puncticulata Haringkansson in the family Stepanodiscaceae Glezer et Makarova(Bacillariophyta) in Japan ndash Biblioth Diatomol 53 1ndash205
TOLOTTI M D CALLIARI amp F CORRADINI (2004) Interannual variability of the phytoplanktonof Tovel lake (Trentino Italy) ndash Studi Trent di Scienze Naturali Acta Biol 81 Suppl 2 327ndash340 (inItalian)
WETZEL RG amp GE LIKENS (1991) Limnological analyses 1ndash391 ndash Springer-Verlag NewYork Berlin Heidelberg
WOJTAL AZ amp J KWANDRANS (2006) Diatoms of the Wyżyna Krakowsko-Częstochowskaupland (S Poland) ndash Coscinodiscophyceae (Thalassiosirophycidae) ndash Polish Bot J 51(2) 177ndash207
WUNSAM S R SCHMIDT amp R KLEE (1995) Cyclotella-taxa (Bacillariophyceae) in lakes of theAlpine region and their relationship to environmental variables ndash Aquat Sci 57(4) 360ndash381
Received 25 March 2010 accepted in revised form 31 March 2011

521
Cyclotella radiosa) complex in Europe found in numerous mesotrophic and eutrophiclakes and slowly running waters in Hungary Germany and Czech Republic Howeverreports of the species are not numerous Till now P balatonis has also never beenreported from Poland
The limited number of reports on the occurrence of P balatonis may be related to itsmisidentificationmisinterpretation as Cyclotella comta (Ehrenb) Kuumltz (Kiss amp Pająk1994 fig 13) found in the plankton of the eutrophic Vistula River as Cyclotellapraetermissa quadrijuncta (Wunsam et al 1995) or as Puncticulata radiosa (Wojtalamp Kwandrans 2006) found in the eutrophic Rudawa River We are convinced that apart of other records of P comta the most commonly reported species of the complexin Poland (Siemińska amp Wołowski 2003) in fact might concern P balatonis InRusałka Lake the species may have also been mistaken for P comta (asC comta) reported from the lake by Kotlińska (1976) It is also likely that P bala-tonis in Poland may have been misidentified as P praetermissa although the latterspecies was only reported several times including findings in the benthos of CzarneLake and the Kamionka River NE Poland (Rakowska 1996) in the plankton ofseveral lakes of the Drawa National Park NW Poland (Gołdyn amp Szeląg-Wasielewska2004) in a sand-pit lake in Krakoacutew and a spring fed pond in Krztynia in S Poland(Wojtal unpublished data)
Up to now there has also been much confusion around the ecology of the speciesfrom the complex For instance P praetermissa described by Lund (1951) fromlakes with a low organic matter content was later found both in the plankton and thebenthos in a wide variety of natural and man-made lakes including alpine andsubalpine lakes (Wunsam et al 1995 Lange-Bertalot amp Metzeltin 1996 Guumlttingeramp Straub 1998 Tolotti et al 2004) as well as in more eutrophic habitats (egSchmidt et al 2002 Hindaacutek amp Hindaacutekovaacute 2003 Tanaka 2007) Houk et al (2010)cleared up the ambiguities ascribing P praetermissa and P radiosa to oligo-mesotrophic habitats According to the authors P comta (as Cyclotella comta) is tillnow known only from fossil materials P balatonis (as Cyclotella balatonis) inturn has been assigned to meso-eutrophic lakes and rivers consistently with thetrophic character of its type locality ndash lake Balaton Our results correspond with theautecology of P balatonis (as Cyclotella balatonis) proposed by Houk et al (2010)widening even slightly the preferences of the species for the trophic conditionsdetermined by Houk et al (2010) as it was also able to develop a large populationin an eutrophic to hypereutrophic water body
The highest numbers of P balatonis in Rusałka Lake were observed in the winter-spring period The diatom bloom in the spring plankton is a well-known phenomenon(Sommer et al 1986) ndash sometimes also occurring during the winter period (Kiss ampGenkal 1993) ndash and is usually associated with a changing silica andor phosphorusavailability during that period (Lampert amp Sommer 2001) Sinking was also provedto be an essential factor hampering diatom success during the thermal stratificationperiod (Jewson et al 1981 Ferris amp Lehman 2007) explaining probably the seasonalityof the P balatonis growth in Rusałka Lake as its high development finished withthe stratification onset Relatively high numbers of frustules in deeper water layersmight also indicate a significant role of the sedimentation process
522
Conclusions
The morphology of Puncticulata balatonis in Rusałka Lake corresponds well toother species around P comta with the exception of the location of marginalfultoportula openings and the length of the shortened striae (notae) Analysis of themorphological characters enabled the transfer of Cyclotella balatonis into the genusPuncticulata A large population of the species in the eutrophic to hypereutrophicRusałka Lake remains congruent with its tolerance ranges with respect to the trophicstate proposed by Houk et al (2010) The variation in the cell number and averagedimension of frustules indicated the species preference to develop large populationsin spring
Acknowledgements
We thank Prof Ryszard Gołdyn (AMickiewicz University in Poznań) for his inspiration researchsupervision and useful comments to the manuscript We are also grateful to Dr Bart Van de Vijver andtwo anonymous reviewers for their valuable suggestions and to Dr Krisztina Buczko for enabling theanalysis of the type material of Cyclotella balatonis from the Pantocsekrsquos collection at HungarianNatural History Museum Sampling on Rusałka Lake was part of a project financially supported by theDistrict and Municipial Funds of Enviromental Protection and Water Management in Poznań (Poland)
References
CARLSON RE (1977) A trophic state index for lakes ndash Limnol Oceanogr 22 (2) 361ndash369
CREMER H amp H KOOLMEES (2010) Common diatoms (Centrales and Fragilariaceae Bacillario-phyta) of modern and fossil freshwater environments in the Netherlands ndash Nova Hedwigia 90(3ndash4)343ndash381
FERRIS JA amp JT LEHMAN (2007) Interannual variation in diatom bloom dynamics Roles ofhydrology nutrient limitation sinking and whole lake manipulation ndash Water Res 41 2551ndash2562
GENKAL SI (1977) On the method of calculation of some taxonomicaly significant structuralelements of the valve in the diatoms of the family Thalassiosiraceae Lebour emend Hasle (Bacillariophyta)ndash Bot Zhurn 62 848ndash851 (in Russian)
GENKAL SI amp KT KISS (1993) Morphological variability of the diatom Cyclotella atomusHustedt var atomus and C atomus var gracilis var nov ndash Hydrobiologia 269270 39ndash47
GOŁDYN R S PODSIADŁOWSKI K KOWALCZEWSKA-MADURA R DONDAJEWSKAE SZELĄG-WASIELEWSKA A BUDZYŃSKA P DOMEK amp W ROMANOWICZ (2010)Functioning of the Rusałka Lake ecosystem in Poznań (west Poland) ndash Ocean Hydrobiol Stud39(3) 65ndash80
GOŁDYN R amp E SZELĄG-WASIELEWSKA (2004) Changes in the phytoseston of a river-lakesystem in Drawieński National Park ndash Ocean Hydrobiol Stud 33(2) 17ndash28
GUumlTTINGER W amp F STRAUB (1998) Diatoms of Lake Cadagno ndash In PEDUZZI RR BACHOFEN amp M TONOLLA (eds) Lake Cadagno a meromictic Alpine lake ndash Doc Ist ItalIdrobiol 63 57ndash64
HANDMANN R (1913) Die Diatomeenflora des Almfeegebiets ndash In MANZ GJ (ed) Mitteilungendes Mikrologischen Vereins Linz ndash Mikrolog Ver Regensburg (in German)
HAUSMANN S amp AF LOTTER (2001) Morphological variation within the diatom taxon Cyclotellacomensis and its importance for quantitative temperature reconstructions ndash Freshw Biol 46 1323ndash1333
523
HAYASHI T (2007) Puncticulata versiformis sp nov and Cyclotella kathmanduensis sp nov(Bacillariophyta) new fossil species from middle Pleistocene lacustrine sediments Kathmandu NepalHilalaya ndash J Phycol 43(2) 304ndash318
HAringKANSSON H (2002) A compilation and evaluation of species in the genera StephanodiscusCyclostephanos and Cylotella with a new genus in the family Stephanodiscaceae ndash Diatom Res 17(1)1ndash139
HINDAacuteK F amp A HINDAacuteKOVAacute (2003) Diversity of cyanobacteria and algae of urban gravel pitlakes in Bratislava Slovakia a survey ndash Hydrobiologia 506ndash509 155ndash162
HINDAacuteK F amp A HINDAacuteKOVAacute (2008) Diversity of cyanobacteria and algae of the NationalNature Reserve Čiacutečovskeacute mŕtve rameno oxbow (Southern Slovakia) ndash Bulletin SBS 30 (1) 11ndash19 (inSlovak with an English abstract)
HOUK V R KLEE amp H TANAKA (2010) Atlas of freshwater centric diatoms with a brief key anddescriptions Part III Stephanodiscaceae A Cyclotella Tertiarius Discostella ndash Fottea 10 (Supplement)1ndash498
JEWSON DH BH RIPPEY amp WJK GILMORE (1981) Loss rates from sedimentation parasitismand grazing during the growth nutrient limitation and dormancy of a diatom crop ndash Limnol Oceanogr26 1045ndash1056
KISS KT amp SI GENKAL (1993) Winter blooms of centric diatoms in the River Danube and in itsside arms near Budapest ndash Hydrobiologia 269270 317ndash325
KISS KT amp G PAJĄK (1994) Seasonal change of diatom s in the plankton of the Vistula riverabove and below the Goczalkowice reservoir Poland ndash In KOCIOLEK JP (ed) Proceedings ofthe 11th International Diatom Symposium San Francisco Aug 12ndash17 1990 Memoirs of the CaliforniaAcademy of Sciences 17 517ndash524 San Francisco California Academy of Sciences
KOBAYASI H M IDEI S MAYAMA T NAGUMO amp K OSADA (2006) H KobayasirsquosAtlas of Japanese diatoms based on electron microscopy 1 1ndash531 Uchida Rokakuho Publishing CoLtd Tokyo
KOTLIŃSKA E (1976) Fitoplankton Jeziora Rusałka In Fitoplankton sztucznych jezior położonychna terenie Poznania 1ndash54 Poznańskie Tow Przyjacioacuteł Nauk Prace Komisji Biologicznej (in Polish)
KRAMMER K amp H LANGE-BERTALOT (1991) Bacillariophyceae 3 Teil Centrales Fragilaria-ceae Eunotiaceae ndash In ETTL H J GERLOFF H HEYNIGH amp D MOLLENHAUER (eds)Suumlsswasserflora von Mitteleuropa 2(3) 1ndash576 Gustav Fischer Verlag Stuttgart Jena
LAMPERT W amp U SOMMER (2001) Ekologia woacuted śroacutedlądowych ndash PWN Warszawa
LANGE-BERTALOT H amp D METZELTIN (1996) Indicators of oligotrophy 800 representativeof three ecologically distinct lake types Carbonate buffered-Oligotrophic-Weakly buffered soft waterndash Iconogr Diatomol 2 1ndash390
LUND JWG (1951) Contribution to our knowledge of British algae ndash Hydrobiologia 3(1) 93ndash100
ORGANIZATION FOR ECONOMIC COOPERATION AND DEVELOPMENT (1982)Eutrophication of Waters ndash Monitoring Assessment and Control Final Report OECD Paris
PANTOCSEK J (1901) Die Kieselalgen oder Bacillarien des Balaton 1ndash112 XVI Tafeln ndash Druckder K und K Hofbuchdruckerei des Victor Hornyaacutenszky Budapest
RAKOWSKA B (1996) Preliminary elaboration of the diatom communities occuring in the benthosof the Kamionka River and Lake Czarne at the Pojezierze Suwalskie (1993ndash1994) ndash Fragm FloristGeobot Polon 3 221ndash238
RIMET F JC DRUART amp O ANNEVILLE (2009) Exploring the dynamics of plankton diatomcommunities in Lake Geneva using emergent self-organizing maps (1974ndash2007) ndash Ecol Inform4 99ndash110
524
SCHMIDT R R PSENNER J MUumlLLER P INDINGER amp C KAMENIK (2002) Impact of thelate glacial climate variations on stratification and trophic state of the meromictic lake Laumlngsee (Austria)validation of a conceptual model by multiproxy studies ndash J Limnol 61(1) 49ndash60
SIEMIŃSKA J amp K WOŁOWSKI (2003) Catalogue of Polish prokaryotic and eukaryotic algaendash W Szafer Institute of Botany Polish Academy of Sciences Krakoacutew
SIVER PA (1999) Development of paleolimnological inference models for pH total nitrogen andspecific conductivity based on planktonic diatoms ndash J Paleolimnol 21 45ndash59
SOMMER U ZM GLIWICZ W LAMPERT amp A DUNCAN (1986) The PEG-model of seasonalsuccession of planktonic events in fresh waters ndash Arch Hydrobiol 106 433ndash471
TANAKA H (2007) Taxonomic studies of the genera Cyclotella (Kuumltzing) Breacutebisson DiscostellaHouk et Klee and Puncticulata Haringkansson in the family Stepanodiscaceae Glezer et Makarova(Bacillariophyta) in Japan ndash Biblioth Diatomol 53 1ndash205
TOLOTTI M D CALLIARI amp F CORRADINI (2004) Interannual variability of the phytoplanktonof Tovel lake (Trentino Italy) ndash Studi Trent di Scienze Naturali Acta Biol 81 Suppl 2 327ndash340 (inItalian)
WETZEL RG amp GE LIKENS (1991) Limnological analyses 1ndash391 ndash Springer-Verlag NewYork Berlin Heidelberg
WOJTAL AZ amp J KWANDRANS (2006) Diatoms of the Wyżyna Krakowsko-Częstochowskaupland (S Poland) ndash Coscinodiscophyceae (Thalassiosirophycidae) ndash Polish Bot J 51(2) 177ndash207
WUNSAM S R SCHMIDT amp R KLEE (1995) Cyclotella-taxa (Bacillariophyceae) in lakes of theAlpine region and their relationship to environmental variables ndash Aquat Sci 57(4) 360ndash381
Received 25 March 2010 accepted in revised form 31 March 2011

522
Conclusions
The morphology of Puncticulata balatonis in Rusałka Lake corresponds well toother species around P comta with the exception of the location of marginalfultoportula openings and the length of the shortened striae (notae) Analysis of themorphological characters enabled the transfer of Cyclotella balatonis into the genusPuncticulata A large population of the species in the eutrophic to hypereutrophicRusałka Lake remains congruent with its tolerance ranges with respect to the trophicstate proposed by Houk et al (2010) The variation in the cell number and averagedimension of frustules indicated the species preference to develop large populationsin spring
Acknowledgements
We thank Prof Ryszard Gołdyn (AMickiewicz University in Poznań) for his inspiration researchsupervision and useful comments to the manuscript We are also grateful to Dr Bart Van de Vijver andtwo anonymous reviewers for their valuable suggestions and to Dr Krisztina Buczko for enabling theanalysis of the type material of Cyclotella balatonis from the Pantocsekrsquos collection at HungarianNatural History Museum Sampling on Rusałka Lake was part of a project financially supported by theDistrict and Municipial Funds of Enviromental Protection and Water Management in Poznań (Poland)
References
CARLSON RE (1977) A trophic state index for lakes ndash Limnol Oceanogr 22 (2) 361ndash369
CREMER H amp H KOOLMEES (2010) Common diatoms (Centrales and Fragilariaceae Bacillario-phyta) of modern and fossil freshwater environments in the Netherlands ndash Nova Hedwigia 90(3ndash4)343ndash381
FERRIS JA amp JT LEHMAN (2007) Interannual variation in diatom bloom dynamics Roles ofhydrology nutrient limitation sinking and whole lake manipulation ndash Water Res 41 2551ndash2562
GENKAL SI (1977) On the method of calculation of some taxonomicaly significant structuralelements of the valve in the diatoms of the family Thalassiosiraceae Lebour emend Hasle (Bacillariophyta)ndash Bot Zhurn 62 848ndash851 (in Russian)
GENKAL SI amp KT KISS (1993) Morphological variability of the diatom Cyclotella atomusHustedt var atomus and C atomus var gracilis var nov ndash Hydrobiologia 269270 39ndash47
GOŁDYN R S PODSIADŁOWSKI K KOWALCZEWSKA-MADURA R DONDAJEWSKAE SZELĄG-WASIELEWSKA A BUDZYŃSKA P DOMEK amp W ROMANOWICZ (2010)Functioning of the Rusałka Lake ecosystem in Poznań (west Poland) ndash Ocean Hydrobiol Stud39(3) 65ndash80
GOŁDYN R amp E SZELĄG-WASIELEWSKA (2004) Changes in the phytoseston of a river-lakesystem in Drawieński National Park ndash Ocean Hydrobiol Stud 33(2) 17ndash28
GUumlTTINGER W amp F STRAUB (1998) Diatoms of Lake Cadagno ndash In PEDUZZI RR BACHOFEN amp M TONOLLA (eds) Lake Cadagno a meromictic Alpine lake ndash Doc Ist ItalIdrobiol 63 57ndash64
HANDMANN R (1913) Die Diatomeenflora des Almfeegebiets ndash In MANZ GJ (ed) Mitteilungendes Mikrologischen Vereins Linz ndash Mikrolog Ver Regensburg (in German)
HAUSMANN S amp AF LOTTER (2001) Morphological variation within the diatom taxon Cyclotellacomensis and its importance for quantitative temperature reconstructions ndash Freshw Biol 46 1323ndash1333
523
HAYASHI T (2007) Puncticulata versiformis sp nov and Cyclotella kathmanduensis sp nov(Bacillariophyta) new fossil species from middle Pleistocene lacustrine sediments Kathmandu NepalHilalaya ndash J Phycol 43(2) 304ndash318
HAringKANSSON H (2002) A compilation and evaluation of species in the genera StephanodiscusCyclostephanos and Cylotella with a new genus in the family Stephanodiscaceae ndash Diatom Res 17(1)1ndash139
HINDAacuteK F amp A HINDAacuteKOVAacute (2003) Diversity of cyanobacteria and algae of urban gravel pitlakes in Bratislava Slovakia a survey ndash Hydrobiologia 506ndash509 155ndash162
HINDAacuteK F amp A HINDAacuteKOVAacute (2008) Diversity of cyanobacteria and algae of the NationalNature Reserve Čiacutečovskeacute mŕtve rameno oxbow (Southern Slovakia) ndash Bulletin SBS 30 (1) 11ndash19 (inSlovak with an English abstract)
HOUK V R KLEE amp H TANAKA (2010) Atlas of freshwater centric diatoms with a brief key anddescriptions Part III Stephanodiscaceae A Cyclotella Tertiarius Discostella ndash Fottea 10 (Supplement)1ndash498
JEWSON DH BH RIPPEY amp WJK GILMORE (1981) Loss rates from sedimentation parasitismand grazing during the growth nutrient limitation and dormancy of a diatom crop ndash Limnol Oceanogr26 1045ndash1056
KISS KT amp SI GENKAL (1993) Winter blooms of centric diatoms in the River Danube and in itsside arms near Budapest ndash Hydrobiologia 269270 317ndash325
KISS KT amp G PAJĄK (1994) Seasonal change of diatom s in the plankton of the Vistula riverabove and below the Goczalkowice reservoir Poland ndash In KOCIOLEK JP (ed) Proceedings ofthe 11th International Diatom Symposium San Francisco Aug 12ndash17 1990 Memoirs of the CaliforniaAcademy of Sciences 17 517ndash524 San Francisco California Academy of Sciences
KOBAYASI H M IDEI S MAYAMA T NAGUMO amp K OSADA (2006) H KobayasirsquosAtlas of Japanese diatoms based on electron microscopy 1 1ndash531 Uchida Rokakuho Publishing CoLtd Tokyo
KOTLIŃSKA E (1976) Fitoplankton Jeziora Rusałka In Fitoplankton sztucznych jezior położonychna terenie Poznania 1ndash54 Poznańskie Tow Przyjacioacuteł Nauk Prace Komisji Biologicznej (in Polish)
KRAMMER K amp H LANGE-BERTALOT (1991) Bacillariophyceae 3 Teil Centrales Fragilaria-ceae Eunotiaceae ndash In ETTL H J GERLOFF H HEYNIGH amp D MOLLENHAUER (eds)Suumlsswasserflora von Mitteleuropa 2(3) 1ndash576 Gustav Fischer Verlag Stuttgart Jena
LAMPERT W amp U SOMMER (2001) Ekologia woacuted śroacutedlądowych ndash PWN Warszawa
LANGE-BERTALOT H amp D METZELTIN (1996) Indicators of oligotrophy 800 representativeof three ecologically distinct lake types Carbonate buffered-Oligotrophic-Weakly buffered soft waterndash Iconogr Diatomol 2 1ndash390
LUND JWG (1951) Contribution to our knowledge of British algae ndash Hydrobiologia 3(1) 93ndash100
ORGANIZATION FOR ECONOMIC COOPERATION AND DEVELOPMENT (1982)Eutrophication of Waters ndash Monitoring Assessment and Control Final Report OECD Paris
PANTOCSEK J (1901) Die Kieselalgen oder Bacillarien des Balaton 1ndash112 XVI Tafeln ndash Druckder K und K Hofbuchdruckerei des Victor Hornyaacutenszky Budapest
RAKOWSKA B (1996) Preliminary elaboration of the diatom communities occuring in the benthosof the Kamionka River and Lake Czarne at the Pojezierze Suwalskie (1993ndash1994) ndash Fragm FloristGeobot Polon 3 221ndash238
RIMET F JC DRUART amp O ANNEVILLE (2009) Exploring the dynamics of plankton diatomcommunities in Lake Geneva using emergent self-organizing maps (1974ndash2007) ndash Ecol Inform4 99ndash110
524
SCHMIDT R R PSENNER J MUumlLLER P INDINGER amp C KAMENIK (2002) Impact of thelate glacial climate variations on stratification and trophic state of the meromictic lake Laumlngsee (Austria)validation of a conceptual model by multiproxy studies ndash J Limnol 61(1) 49ndash60
SIEMIŃSKA J amp K WOŁOWSKI (2003) Catalogue of Polish prokaryotic and eukaryotic algaendash W Szafer Institute of Botany Polish Academy of Sciences Krakoacutew
SIVER PA (1999) Development of paleolimnological inference models for pH total nitrogen andspecific conductivity based on planktonic diatoms ndash J Paleolimnol 21 45ndash59
SOMMER U ZM GLIWICZ W LAMPERT amp A DUNCAN (1986) The PEG-model of seasonalsuccession of planktonic events in fresh waters ndash Arch Hydrobiol 106 433ndash471
TANAKA H (2007) Taxonomic studies of the genera Cyclotella (Kuumltzing) Breacutebisson DiscostellaHouk et Klee and Puncticulata Haringkansson in the family Stepanodiscaceae Glezer et Makarova(Bacillariophyta) in Japan ndash Biblioth Diatomol 53 1ndash205
TOLOTTI M D CALLIARI amp F CORRADINI (2004) Interannual variability of the phytoplanktonof Tovel lake (Trentino Italy) ndash Studi Trent di Scienze Naturali Acta Biol 81 Suppl 2 327ndash340 (inItalian)
WETZEL RG amp GE LIKENS (1991) Limnological analyses 1ndash391 ndash Springer-Verlag NewYork Berlin Heidelberg
WOJTAL AZ amp J KWANDRANS (2006) Diatoms of the Wyżyna Krakowsko-Częstochowskaupland (S Poland) ndash Coscinodiscophyceae (Thalassiosirophycidae) ndash Polish Bot J 51(2) 177ndash207
WUNSAM S R SCHMIDT amp R KLEE (1995) Cyclotella-taxa (Bacillariophyceae) in lakes of theAlpine region and their relationship to environmental variables ndash Aquat Sci 57(4) 360ndash381
Received 25 March 2010 accepted in revised form 31 March 2011

523
HAYASHI T (2007) Puncticulata versiformis sp nov and Cyclotella kathmanduensis sp nov(Bacillariophyta) new fossil species from middle Pleistocene lacustrine sediments Kathmandu NepalHilalaya ndash J Phycol 43(2) 304ndash318
HAringKANSSON H (2002) A compilation and evaluation of species in the genera StephanodiscusCyclostephanos and Cylotella with a new genus in the family Stephanodiscaceae ndash Diatom Res 17(1)1ndash139
HINDAacuteK F amp A HINDAacuteKOVAacute (2003) Diversity of cyanobacteria and algae of urban gravel pitlakes in Bratislava Slovakia a survey ndash Hydrobiologia 506ndash509 155ndash162
HINDAacuteK F amp A HINDAacuteKOVAacute (2008) Diversity of cyanobacteria and algae of the NationalNature Reserve Čiacutečovskeacute mŕtve rameno oxbow (Southern Slovakia) ndash Bulletin SBS 30 (1) 11ndash19 (inSlovak with an English abstract)
HOUK V R KLEE amp H TANAKA (2010) Atlas of freshwater centric diatoms with a brief key anddescriptions Part III Stephanodiscaceae A Cyclotella Tertiarius Discostella ndash Fottea 10 (Supplement)1ndash498
JEWSON DH BH RIPPEY amp WJK GILMORE (1981) Loss rates from sedimentation parasitismand grazing during the growth nutrient limitation and dormancy of a diatom crop ndash Limnol Oceanogr26 1045ndash1056
KISS KT amp SI GENKAL (1993) Winter blooms of centric diatoms in the River Danube and in itsside arms near Budapest ndash Hydrobiologia 269270 317ndash325
KISS KT amp G PAJĄK (1994) Seasonal change of diatom s in the plankton of the Vistula riverabove and below the Goczalkowice reservoir Poland ndash In KOCIOLEK JP (ed) Proceedings ofthe 11th International Diatom Symposium San Francisco Aug 12ndash17 1990 Memoirs of the CaliforniaAcademy of Sciences 17 517ndash524 San Francisco California Academy of Sciences
KOBAYASI H M IDEI S MAYAMA T NAGUMO amp K OSADA (2006) H KobayasirsquosAtlas of Japanese diatoms based on electron microscopy 1 1ndash531 Uchida Rokakuho Publishing CoLtd Tokyo
KOTLIŃSKA E (1976) Fitoplankton Jeziora Rusałka In Fitoplankton sztucznych jezior położonychna terenie Poznania 1ndash54 Poznańskie Tow Przyjacioacuteł Nauk Prace Komisji Biologicznej (in Polish)
KRAMMER K amp H LANGE-BERTALOT (1991) Bacillariophyceae 3 Teil Centrales Fragilaria-ceae Eunotiaceae ndash In ETTL H J GERLOFF H HEYNIGH amp D MOLLENHAUER (eds)Suumlsswasserflora von Mitteleuropa 2(3) 1ndash576 Gustav Fischer Verlag Stuttgart Jena
LAMPERT W amp U SOMMER (2001) Ekologia woacuted śroacutedlądowych ndash PWN Warszawa
LANGE-BERTALOT H amp D METZELTIN (1996) Indicators of oligotrophy 800 representativeof three ecologically distinct lake types Carbonate buffered-Oligotrophic-Weakly buffered soft waterndash Iconogr Diatomol 2 1ndash390
LUND JWG (1951) Contribution to our knowledge of British algae ndash Hydrobiologia 3(1) 93ndash100
ORGANIZATION FOR ECONOMIC COOPERATION AND DEVELOPMENT (1982)Eutrophication of Waters ndash Monitoring Assessment and Control Final Report OECD Paris
PANTOCSEK J (1901) Die Kieselalgen oder Bacillarien des Balaton 1ndash112 XVI Tafeln ndash Druckder K und K Hofbuchdruckerei des Victor Hornyaacutenszky Budapest
RAKOWSKA B (1996) Preliminary elaboration of the diatom communities occuring in the benthosof the Kamionka River and Lake Czarne at the Pojezierze Suwalskie (1993ndash1994) ndash Fragm FloristGeobot Polon 3 221ndash238
RIMET F JC DRUART amp O ANNEVILLE (2009) Exploring the dynamics of plankton diatomcommunities in Lake Geneva using emergent self-organizing maps (1974ndash2007) ndash Ecol Inform4 99ndash110
524
SCHMIDT R R PSENNER J MUumlLLER P INDINGER amp C KAMENIK (2002) Impact of thelate glacial climate variations on stratification and trophic state of the meromictic lake Laumlngsee (Austria)validation of a conceptual model by multiproxy studies ndash J Limnol 61(1) 49ndash60
SIEMIŃSKA J amp K WOŁOWSKI (2003) Catalogue of Polish prokaryotic and eukaryotic algaendash W Szafer Institute of Botany Polish Academy of Sciences Krakoacutew
SIVER PA (1999) Development of paleolimnological inference models for pH total nitrogen andspecific conductivity based on planktonic diatoms ndash J Paleolimnol 21 45ndash59
SOMMER U ZM GLIWICZ W LAMPERT amp A DUNCAN (1986) The PEG-model of seasonalsuccession of planktonic events in fresh waters ndash Arch Hydrobiol 106 433ndash471
TANAKA H (2007) Taxonomic studies of the genera Cyclotella (Kuumltzing) Breacutebisson DiscostellaHouk et Klee and Puncticulata Haringkansson in the family Stepanodiscaceae Glezer et Makarova(Bacillariophyta) in Japan ndash Biblioth Diatomol 53 1ndash205
TOLOTTI M D CALLIARI amp F CORRADINI (2004) Interannual variability of the phytoplanktonof Tovel lake (Trentino Italy) ndash Studi Trent di Scienze Naturali Acta Biol 81 Suppl 2 327ndash340 (inItalian)
WETZEL RG amp GE LIKENS (1991) Limnological analyses 1ndash391 ndash Springer-Verlag NewYork Berlin Heidelberg
WOJTAL AZ amp J KWANDRANS (2006) Diatoms of the Wyżyna Krakowsko-Częstochowskaupland (S Poland) ndash Coscinodiscophyceae (Thalassiosirophycidae) ndash Polish Bot J 51(2) 177ndash207
WUNSAM S R SCHMIDT amp R KLEE (1995) Cyclotella-taxa (Bacillariophyceae) in lakes of theAlpine region and their relationship to environmental variables ndash Aquat Sci 57(4) 360ndash381
Received 25 March 2010 accepted in revised form 31 March 2011

524
SCHMIDT R R PSENNER J MUumlLLER P INDINGER amp C KAMENIK (2002) Impact of thelate glacial climate variations on stratification and trophic state of the meromictic lake Laumlngsee (Austria)validation of a conceptual model by multiproxy studies ndash J Limnol 61(1) 49ndash60
SIEMIŃSKA J amp K WOŁOWSKI (2003) Catalogue of Polish prokaryotic and eukaryotic algaendash W Szafer Institute of Botany Polish Academy of Sciences Krakoacutew
SIVER PA (1999) Development of paleolimnological inference models for pH total nitrogen andspecific conductivity based on planktonic diatoms ndash J Paleolimnol 21 45ndash59
SOMMER U ZM GLIWICZ W LAMPERT amp A DUNCAN (1986) The PEG-model of seasonalsuccession of planktonic events in fresh waters ndash Arch Hydrobiol 106 433ndash471
TANAKA H (2007) Taxonomic studies of the genera Cyclotella (Kuumltzing) Breacutebisson DiscostellaHouk et Klee and Puncticulata Haringkansson in the family Stepanodiscaceae Glezer et Makarova(Bacillariophyta) in Japan ndash Biblioth Diatomol 53 1ndash205
TOLOTTI M D CALLIARI amp F CORRADINI (2004) Interannual variability of the phytoplanktonof Tovel lake (Trentino Italy) ndash Studi Trent di Scienze Naturali Acta Biol 81 Suppl 2 327ndash340 (inItalian)
WETZEL RG amp GE LIKENS (1991) Limnological analyses 1ndash391 ndash Springer-Verlag NewYork Berlin Heidelberg
WOJTAL AZ amp J KWANDRANS (2006) Diatoms of the Wyżyna Krakowsko-Częstochowskaupland (S Poland) ndash Coscinodiscophyceae (Thalassiosirophycidae) ndash Polish Bot J 51(2) 177ndash207
WUNSAM S R SCHMIDT amp R KLEE (1995) Cyclotella-taxa (Bacillariophyceae) in lakes of theAlpine region and their relationship to environmental variables ndash Aquat Sci 57(4) 360ndash381
Received 25 March 2010 accepted in revised form 31 March 2011



















