
Photosynthesis, water respiration and growth performance of Litopenaeus vannamei in a...
8
Photosynthesis, water respiration and growth performance of Litopenaeus vannamei in a super-intensive raceway culture with zero water exchange: Interaction of water quality variables Luis Vinatea a, *, Alfredo O. Ga ´ lvez b , Craig L. Browdy c , Al Stokes c , Jesus Venero c , Jason Haveman c , Beth L. Lewis c , Alisha Lawson c , Andrew Shuler c , John W. Leffler c a Universidade Federal de Santa Catarina, Depto. de Aquicultura, Laborato ´rio de Camaro˜es Marinhos, Floriano ´polis, SC, CEP 88040-900, Brazil b Universidade Federal Rural de Pernambuco, Depto. de Pesca e Aquicultura, Laborato ´rio de Maricultura Sustenta ´vel, Recife, PE, CEP 52171-900, Brazil c South Carolina Department of Natural Resources, Waddell Mariculture Center, Bluffton, SC, SC 29910, USA 1. Introduction Aquaculture in general and shrimp culture in particular have recently been developing strategies of super-intensive cultures without water exchange. This approach addresses environmental questions raised by both society and the scientific community regarding sustainable development concepts which demand a convergence of ecological prudence, economic efficiency and social equity in all human activities (Bailey, 1988; Brown, 1989; Pruder, 1992; Macintosh and Phillips, 1992; Kinkelin and Michel, 1992; Pe ´ rez, 1993; Currie, 1994; Primavera, 1994; Rosenthal, 1994; Larsson et al., 1995; Kestemont, 1995). Pioneer work such as Reid and Arnold (1992) and Williams et al. (1996) demonstrated that it is possible to produce shrimp at high density in raceway systems that use water recirculation. The recent progress made in the area of super-dense cultures, also known as ‘‘suspended-growth systems’’ (Hargreaves, 2006) and ‘‘active suspension ponds’’ (Avnimelech, 2006), reinforces the idea that it is possible to produce aquatic organisms in an intensive and especially bio-safe way. The absence of effluents, the reduction of utilized space, and the dramatic reduction of the introduction of infectious diseases, are the main criteria for justifying its development (McNeil, 2000; McAbee et al., 2003; Burford et al., 2003, 2004; Pruder, 2004; Sowers et al., 2005; Wasielesky et al., 2006; Azim and Little, 2008; De Schryver et al., 2008; Schveitzer et al., 2008). Although this relatively new aquaculture technology is still developing (De Schryver et al., 2008), important research efforts have been made with the objective of comprehending the physical, chemical and biological phenomena present in the water that dictate the dynamic of the cultures (Hopkins et al., 1993; Moss and Pruder, 1995; Sandifer and Hopkins, 1996; Davis and Arnold, 1998; Avnimelech, 1999; Browdy et al., 2001). Some studies suggest that shrimp culture in a closed system (without water exchange) can maintain water quality considered acceptable for these organisms (Thakur and Lin, 2003). Others suggest that the water quality, in spite of being ‘‘poor’’ due to the large quantity of nutrients that are accumulated over time, is sufficient to guarantee the growth and Aquacultural Engineering 42 (2010) 17–24 ARTICLE INFO Article history: Received 12 November 2008 Accepted 8 September 2009 Keywords: Water quality Photosynthesis Litopenaeus vannamei Super-intensive culture ABSTRACT Gross and net photosynthesis, and water column respiration of a super-intensive Litopenaeus vannamei raceway culture with zero water exchange were measured over 21 weeks using the light and dark bottle method. Spearman correlation analysis was used to explore the relationship of these parameters with other water quality parameters (O 2 , temperature, pH, alkalinity, TAN, NO 2 , NO 3 , dissolved orthopho- sphate, turbidity, TSS and VSS) and of shrimp performance based on growth rate and feed conversion rate. Net photosynthesis generally had negative values, indicating that the system was predominantly heterotrophic. Gross photosynthesis was significantly related only with VSS, and water column respiration was related only with turbidity. Shrimp growth rate increased with higher temperature and dropped with higher pH. Nitrite had a strong inverse relationship with shrimp growth rate (R 2 = 0.8044, p < 0.001) and a weak relationship with FCR (R 2 = 0.3319, p = 0.0284). VSS has a significant inverse relationship with shrimp growth rate (R 2 = 0.4989, p = 0.0102) and a direct relationship with FCR (R 2 = 0.5356, p = 0.0162). Photosynthesis and the water column respiration did not seem to have a significant impact on shrimp productivity. ß 2009 Elsevier B.V. All rights reserved. * Corresponding author. E-mail address: [email protected] (L. Vinatea). Contents lists available at ScienceDirect Aquacultural Engineering journal homepage: www.elsevier.com/locate/aqua-online 0144-8609/$ – see front matter ß 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.aquaeng.2009.09.001
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Photosynthesis, water respiration and growth performance of Litopenaeus vannamei in a...

Photosynthesis, water respiration and growth performance of Litopenaeusvannamei in a super-intensive raceway culture with zero water exchange:Interaction of water quality variables
Luis Vinatea a,*, Alfredo O. Galvez b, Craig L. Browdy c, Al Stokes c, Jesus Venero c, Jason Haveman c,Beth L. Lewis c, Alisha Lawson c, Andrew Shuler c, John W. Leffler c
a Universidade Federal de Santa Catarina, Depto. de Aquicultura, Laboratorio de Camaroes Marinhos, Florianopolis, SC, CEP 88040-900, Brazilb Universidade Federal Rural de Pernambuco, Depto. de Pesca e Aquicultura, Laboratorio de Maricultura Sustentavel, Recife, PE, CEP 52171-900, Brazilc South Carolina Department of Natural Resources, Waddell Mariculture Center, Bluffton, SC, SC 29910, USA
Aquacultural Engineering 42 (2010) 17–24
A R T I C L E I N F O
Article history:
Received 12 November 2008
Accepted 8 September 2009
Keywords:
Water quality
Photosynthesis
Litopenaeus vannamei
Super-intensive culture
A B S T R A C T
Gross and net photosynthesis, and water column respiration of a super-intensive Litopenaeus vannamei
raceway culture with zero water exchange were measured over 21 weeks using the light and dark bottle
method. Spearman correlation analysis was used to explore the relationship of these parameters with
other water quality parameters (O2, temperature, pH, alkalinity, TAN, NO2, NO3, dissolved orthopho-
sphate, turbidity, TSS and VSS) and of shrimp performance based on growth rate and feed conversion
rate. Net photosynthesis generally had negative values, indicating that the system was predominantly
heterotrophic. Gross photosynthesis was significantly related only with VSS, and water column
respiration was related only with turbidity. Shrimp growth rate increased with higher temperature and
dropped with higher pH. Nitrite had a strong inverse relationship with shrimp growth rate (R2 = 0.8044,
p < 0.001) and a weak relationship with FCR (R2 = 0.3319, p = 0.0284). VSS has a significant inverse
relationship with shrimp growth rate (R2 = 0.4989, p = 0.0102) and a direct relationship with FCR
(R2 = 0.5356, p = 0.0162). Photosynthesis and the water column respiration did not seem to have a
significant impact on shrimp productivity.
� 2009 Elsevier B.V. All rights reserved.
Contents lists available at ScienceDirect
Aquacultural Engineering
journa l homepage: www.e lsev ier .com/ locate /aqua-onl ine
1. Introduction
Aquaculture in general and shrimp culture in particular haverecently been developing strategies of super-intensive cultureswithout water exchange. This approach addresses environmentalquestions raised by both society and the scientific communityregarding sustainable development concepts which demand aconvergence of ecological prudence, economic efficiency and socialequity in all human activities (Bailey, 1988; Brown, 1989; Pruder,1992; Macintosh and Phillips, 1992; Kinkelin and Michel, 1992;Perez, 1993; Currie, 1994; Primavera, 1994; Rosenthal, 1994;Larsson et al., 1995; Kestemont, 1995).
Pioneer work such as Reid and Arnold (1992) and Williams et al.(1996) demonstrated that it is possible to produce shrimp at highdensity in raceway systems that use water recirculation. Therecent progress made in the area of super-dense cultures, alsoknown as ‘‘suspended-growth systems’’ (Hargreaves, 2006) and
* Corresponding author.
E-mail address: [email protected] (L. Vinatea).
0144-8609/$ – see front matter � 2009 Elsevier B.V. All rights reserved.
doi:10.1016/j.aquaeng.2009.09.001
‘‘active suspension ponds’’ (Avnimelech, 2006), reinforces the ideathat it is possible to produce aquatic organisms in an intensive andespecially bio-safe way. The absence of effluents, the reduction ofutilized space, and the dramatic reduction of the introduction ofinfectious diseases, are the main criteria for justifying itsdevelopment (McNeil, 2000; McAbee et al., 2003; Burford et al.,2003, 2004; Pruder, 2004; Sowers et al., 2005; Wasielesky et al.,2006; Azim and Little, 2008; De Schryver et al., 2008; Schveitzeret al., 2008).
Although this relatively new aquaculture technology is stilldeveloping (De Schryver et al., 2008), important research effortshave been made with the objective of comprehending the physical,chemical and biological phenomena present in the water thatdictate the dynamic of the cultures (Hopkins et al., 1993; Moss andPruder, 1995; Sandifer and Hopkins, 1996; Davis and Arnold, 1998;Avnimelech, 1999; Browdy et al., 2001). Some studies suggest thatshrimp culture in a closed system (without water exchange) canmaintain water quality considered acceptable for these organisms(Thakur and Lin, 2003). Others suggest that the water quality, inspite of being ‘‘poor’’ due to the large quantity of nutrients that areaccumulated over time, is sufficient to guarantee the growth and

L. Vinatea et al. / Aquacultural Engineering 42 (2010) 17–2418
survival of the animals (Burford et al., 2003). Hargreaves (2006)criticizes the majority of studies regarding these so-called‘‘suspended-growth systems’’, because they suggest that the roleof the bacteria in reference to the maintenance and theimprovement of the water quality is more important than thephytoplankton metabolism.
The importance that primary productivity has in the cultures(extensive, semi-intensive, polycultures, etc.) of different organ-isms is well-documented (Noriega-Curtis, 1979; Laws andMalecha, 1981; Chang and Ouyang, 1988; Yusoff and McNabb,1989; Kadir et al., 2006). However there still exist many doubtswith respect to the role that photosynthesis plays in super-intensive cultures, regarding both level of water quality and thegrowth and well-being of the organisms that are being cultivated.This study contributes to the better understanding of thephenomenon by proposing possible relationships between photo-synthesis, water respiration, shrimp growth and other parametersof water quality present in a zero exchange, super-intensiveshrimp culture.
2. Materials and methods
The present study was conducted at the Waddell MaricultureCenter (South Carolina, Department of Natural Resources, Bluffton,SC, USA) from March to August 2008. A 271 m2 raceway containedwithin a transparent, high-density plastic greenhouse was stockedwith 254,250 PL10 SPF Litopenaeus vannamei (Shrimp Improve-ments Systems Inc., Islamorda, FL, USA) on March 13. Following a43-day nursery period, the growout period began on April 25 atwhich time the population of 1.48 g animals was estimated at165,250 for a density of 610/m2. The shrimp were fed a complete,balanced 35% protein commercial feed (Ziegler Hyper IntensiveRaceway, Ziegler Brothers, Inc., Gardners, PA, USA). The racewayaeration was supplemented by pure oxygen though diffusersstarting with the eighth week of culture. Some solids wereremoved from the water column by an experiment with externalsettling tanks, but this did not significantly alter the total amountof suspended solids (TSS) in the system.
The raceway aeration system consisted of one 5 HP blowerconnected to 60 air stones, and two oxygen generators (40 and140 L) paired with venturi injectors and one 250 L Sweetwater1
oxygen saturation cone. The raceway was also provided with three
Table 1Water quality parameters determined during the experiment and methods used for
the analyses.
Parameter (units) Method
Dissolved oxygen (mg O2/L)a Multiparameter YSI 556 MPS
Temperature (8C)a Multiparameter YSI 556 MPS
Salinity (%)a Multiparameter YSI 556 MPS
Light (mmol photons/m2 s)a Light meter LI COR (LI-1400)b
pHc Multiparameter YSI 556 MPS
Alkalinity (mg CaCO3/L)d Titration method (HCl 0.1 N)
Turbidity (NTU)d La Motte Turbidity Meter 2020
Total ammonium nitrogen-TAN (mg/L)c,e HACH method 8155 (salicylate)
Nitrite nitrogen (mg N-NO2/L)d HACH method 8507
(diazotization)
Nitrate nitrogen (mg N-NO3/L)d HACH method 8039
(cadmium reduction)
Dissolved orthophosphate (mg PO4/L)d HACH method 8114
(ascorbic acid)
Total suspended solids: TSS (mg/L)d EPA method 340.2
Volatile suspended solids: VSS (mg/L)d EPA method 340.2
a Twice daily (morning and afternoon).b Underwater sensor, located 50 cm below water surface.c Once daily.d Once weekly.e NH3 + NH4
+.
O2 cylinders as a backup system. The pure oxygen was suppliedwhen the dissolved oxygen concentration could not be maintainedabove 3 mg/L with simple mechanical aeration.
The water quality control system consisted in measuringdissolved oxygen, temperature, salinity and light twice a day, everyday; pH once a day, every day, and alkalinity, turbidity, TAN, nitrite,nitrate, dissolved orthophosphate, TSS and VSS once a week(Table 1). When the alkalinity was below 60 mg/L, pure sodiumbicarbonate was applied at the rate of 0.12 g/L (a total of 24 kg forthe run), and when the TAN was above 2.0 mg/L, dextrose wasadded at the rate of 0.05 g/L (a total of 22 kg for the run).
Water column respiration and photosynthesis were recordedstarting from the third week of culture, and most physical andchemical parameters were recorded starting from the eighth week.Weekly microscopic examinations identified the main groups ofmicroorganisms present in the water column.
Gross and net photosynthesis and water column respirationwere measured by the classic dark and light bottle method(Strickland, 1960), using the following formulas:
Gross photosynthesis ðmg O2=L hÞ
¼ final O2 of light bottle� final O2 of dark bottle
time ðhÞ
� �
Net photosynthesis ðmg O2=L hÞ
¼ final O2 of light bottle� initial O2 of light bottle
time ðhÞ
� �
Respiration ðmg O2=L hÞ
¼ initial O2 of dark bottle� final O2 of dark bottle
time ðhÞ
� �
Once weekly water from the raceway was collected at a depth of20 cm and placed in triplicate sets of 300 mL glass BOD bottles thatwere either clear or black. Initial and final oxygen concentrations
Fig. 1. Polycarbonate water bath used to measure photosynthesis and water
respiration of a super-intensive Litopenaeus vannamei raceway culture with zero
water exchange by the light and dark bottle method. (A) Water bath with rotating
wheel. (B) 300 mL BOD bottles used in the experiment.
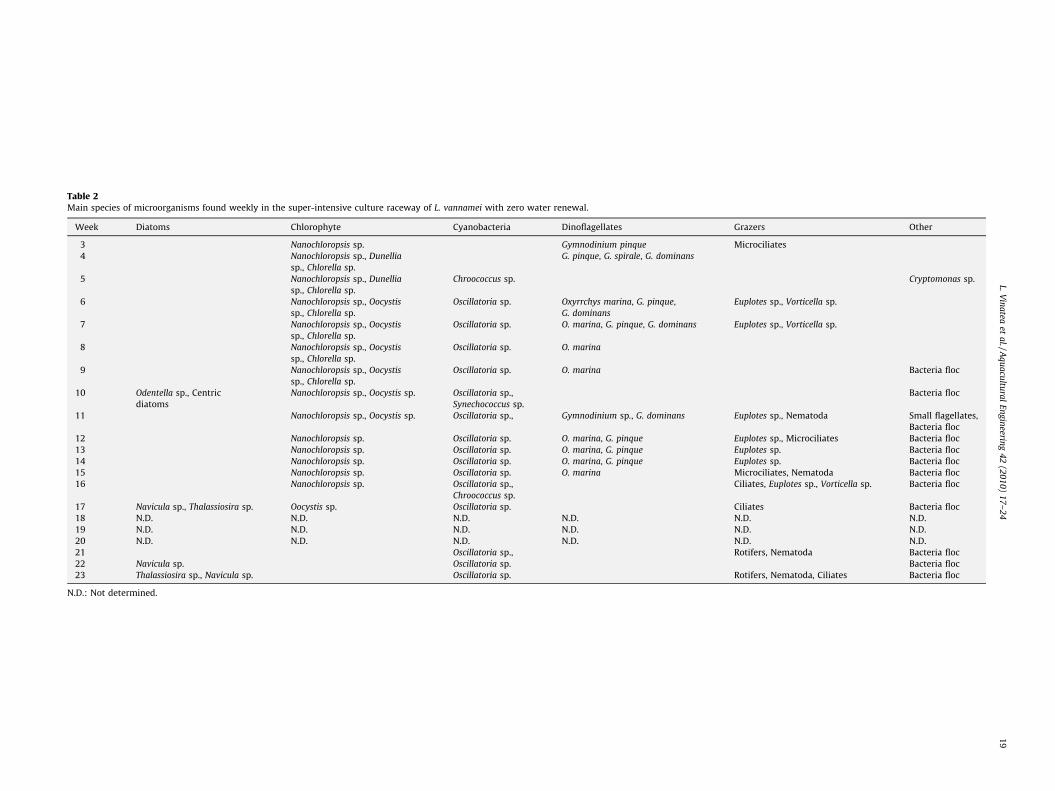
Table 2Main species of microorganisms found weekly in the super-intensive culture raceway of L. vannamei with zero water renewal.
Week Diatoms Chlorophyte Cyanobacteria Dinoflagellates Grazers Other
3 Nanochloropsis sp. Gymnodinium pinque Microciliates
4 Nanochloropsis sp., Dunellia
sp., Chlorella sp.
G. pinque, G. spirale, G. dominans
5 Nanochloropsis sp., Dunellia
sp., Chlorella sp.
Chroococcus sp. Cryptomonas sp.
6 Nanochloropsis sp., Oocystis
sp., Chlorella sp.
Oscillatoria sp. Oxyrrchys marina, G. pinque,
G. dominans
Euplotes sp., Vorticella sp.
7 Nanochloropsis sp., Oocystis
sp., Chlorella sp.
Oscillatoria sp. O. marina, G. pinque, G. dominans Euplotes sp., Vorticella sp.
8 Nanochloropsis sp., Oocystis
sp., Chlorella sp.
Oscillatoria sp. O. marina
9 Nanochloropsis sp., Oocystis
sp., Chlorella sp.
Oscillatoria sp. O. marina Bacteria floc
10 Odentella sp., Centric
diatoms
Nanochloropsis sp., Oocystis sp. Oscillatoria sp.,
Synechococcus sp.
Bacteria floc
11 Nanochloropsis sp., Oocystis sp. Oscillatoria sp., Gymnodinium sp., G. dominans Euplotes sp., Nematoda Small flagellates,
Bacteria floc
12 Nanochloropsis sp. Oscillatoria sp. O. marina, G. pinque Euplotes sp., Microciliates Bacteria floc
13 Nanochloropsis sp. Oscillatoria sp. O. marina, G. pinque Euplotes sp. Bacteria floc
14 Nanochloropsis sp. Oscillatoria sp. O. marina, G. pinque Euplotes sp. Bacteria floc
15 Nanochloropsis sp. Oscillatoria sp. O. marina Microciliates, Nematoda Bacteria floc
16 Nanochloropsis sp. Oscillatoria sp.,
Chroococcus sp.
Ciliates, Euplotes sp., Vorticella sp. Bacteria floc
17 Navicula sp., Thalassiosira sp. Oocystis sp. Oscillatoria sp. Ciliates Bacteria floc
18 N.D. N.D. N.D. N.D. N.D. N.D.
19 N.D. N.D. N.D. N.D. N.D. N.D.
20 N.D. N.D. N.D. N.D. N.D. N.D.
21 Oscillatoria sp., Rotifers, Nematoda Bacteria floc
22 Navicula sp. Oscillatoria sp. Bacteria floc
23 Thalassiosira sp., Navicula sp. Oscillatoria sp. Rotifers, Nematoda, Ciliates Bacteria floc
N.D.: Not determined.
L.V
ina
teaet
al./A
qu
acu
ltura
lE
ng
ineerin
g4
2(2
01
0)
17
–2
41
9

L. Vinatea et al. / Aquacultural Engineering 42 (2010) 17–2420
were measured with an YSI 5000 digital oxygen meter equippedwith a stirring BOD probe (Yellow Spring Instruments, YellowSprings, OH, USA). Once initial oxygen was recorded, the bottleswere sealed with glass stoppers held in place by plastic lids of thesame color as the bottle. The bottles were attached to a rotatingwheel contained within a 250 L polycarbonate aquarium andincubated for 2 h (Fig. 1). The movement of the rotating wheelsimulated the turbulence within the raceway and insured that themicrobial floc particles in the water column remained insuspension throughout the incubation. One such incubator washoused within the laboratory with artificial metal halide light andelectric heaters in order to create standard conditions of150 mmol photons/m2 s and 28 8C. An identical incubator washoused next to the raceway within the greenhouse with racewaywater pumped through it continuously to maintain the same lightand temperature regime experienced within the raceway. Netphotosynthesis data were useful for verifying the predominanttrophic condition of the raceway’s water column community overtime.
The shrimp growth rate (GR) was established using a randomsample of 100 shrimp, measuring individual weight andsubtracting the mean weight found from the mean weightregistered one week earlier. Feed conversion rate (FCR) wascalculated by dividing the amount of feed offered during a week bythe increase in total biomass from the previous week calculatedfrom growth rate and estimated mortality. The GR and FCR werecalculated, respectively, starting from the eighth and eleventhweek of the culture.
Gross photosynthesis and water column respiration measuredunder raceway and laboratory conditions were compared using a t-test analysis (p < 0.05). This analysis was performed using JMP 5.0statistical software (SAS Institute Inc., Cary, NC, USA). Nonpara-metric Spearman correlation coefficients were calculated to assessthe strength of the association between gross photosynthesis,water column respiration, shrimp growth rate, FCR and each of theother water quality parameters using SigmaStat 3.1 statisticalsoftware (Systat Software Inc., San Jose, CA, USA). The discussion is
Table 3Gross photosynthesis (mg O2/L h), net photosynthesis (mg O2/L h) and trophic condition
and food conversion rate (FCR) found in a super-intensive raceway of Litopenaeus vannam
of equations and not means, so they don’t have a standard deviation).
Week Gross photosynthesis
(mg O2/L h)a
Net photosynthesis
(mg O2/L h)a
Wat
(mg
3 1.51 0.54 (A) 0.94
4 0.84 �0.40 (H) 1.25
5 0.87 0.36 (A) 0.47
6 0.51 0.19 (A) 0.32
7 0.65 �0.29 (H) 0.94
8 0.35 �1.26 (H) 1.48
9 0.37 �1.65 (H) 1.94
10 0.34 �2.29 (H) 2.60
11 1.13 �0.43 (H) 1.49
12 0.73 �0.44 (H) 1.16
13 1.32 0.02 (A) 1.29
14 0.76 �1.10 (H) 1.86
15 1.28 �0.01 (H) 1.25
16 0.49 �0.76 (H) 1.19
17 0.02 �1.06 (H) 1.09
18 0.36 �1.15 (H) 1.51
19 0.19 �0.82 (H) 1.06
20 0.33 �0.93 (H) 1.28
21 0.24 �0.97 (H) 1.25
22 0.38 �0.85 (H) 1.28
23 0.33 �0.56 (H) 0.92
(A) Autotrophic, (H) heterotrophic.a Initial and final oxygen concentrations in the bottles were measured in triplicate
equations.b The same procedure was adopted for these parameters, only differing in the fact t
based only on those variables that showed statistically significantand biologically meaningful relationships. Simple linear regressionanalyses were performed on those pairs of parameters thatresulted in high correlation coefficients. The resulting relationshipswere nitrite concentration versus growth rate and FCR, and nitriteconcentration versus VSS.
3. Results
By August 8 (growout day 105) shrimp had grown an average of1.19 g/week, had a mean individual weight of 19.3 g, and survivalappeared high. A failure of the oxygen delivery system on August 9and 10 resulted in a major mortality event and forced anemergency harvest on August 14. This resulted in a final meanindividual weight of 18.1 g and a survival rate of 40% based on theestimated density at the start of the growout period. Despite thismisfortune, none of the results presented in this paper wereimpacted by the oxygen failure because all measurements,including shrimp growth rates and FCRs, being reported wereconcluded at least a week prior to the event.
The main taxonomic groups of microorganisms found in thewater column are listed in Table 2. Table 3 shows the measuredvalues of gross and net photosynthesis, water respiration, meanweekly weight, growth rate, and food conversion rate of theshrimp.
Gross photosynthesis data did not show significant differencesbetween measurements from the raceway or from the laboratory(t-test, p < 0.05). Similarly, water respiration data were notsignificantly different when measured in the raceway or in thelaboratory.
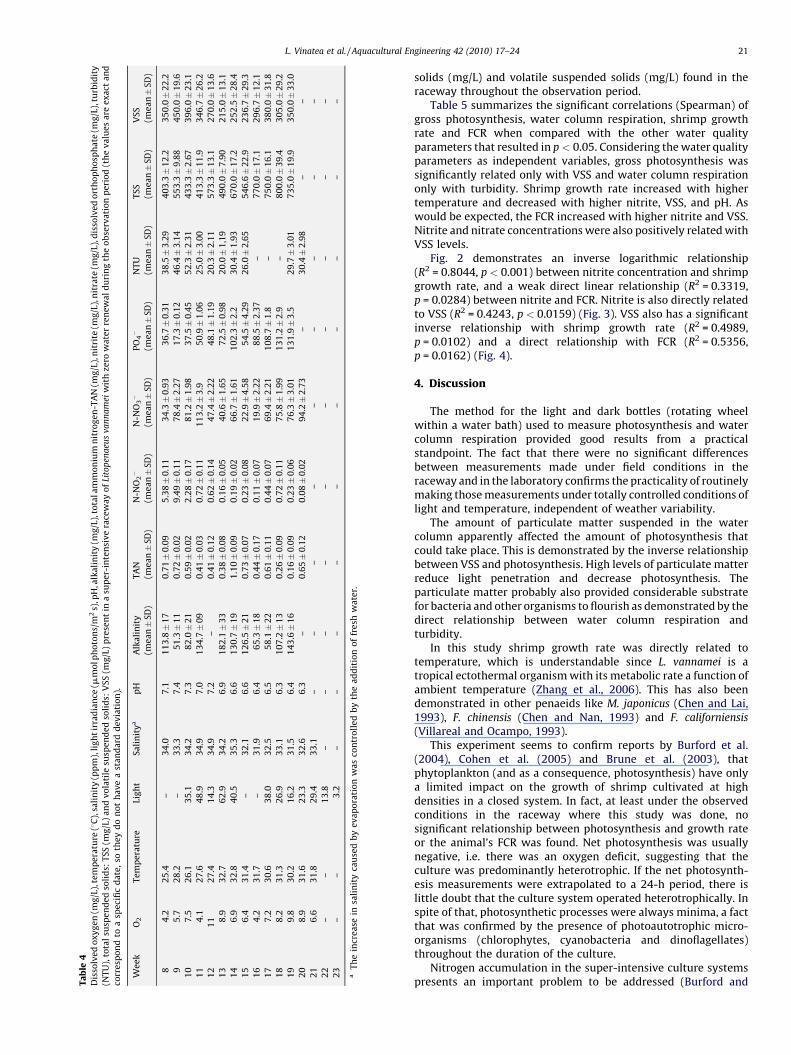
With regard to net photosynthesis, of the 21 weeks ofobservation, 4 presented positive values (oxygen surplus) and17 presented negative values (oxygen deficit). Table 4 shows thevalues of dissolved oxygen (mg/L), temperature (8C), salinity (ppt),light irradiance (mmol photons/m2 s), pH, alkalinity (mg/L), totalammonium nitrogen-TAN (mg/L), nitrite (mg/L), nitrate (mg/L),dissolved orthophosphate (mg/L), turbidity (NTU), total suspended
(autotrophic or heterotrophic), water respiration (mg O2/L h), growth rate (g/week)
ei with zero water renewal (all the values, except the animals’ weight, are a product
er respiration
O2/L h)a
Weight (g� SD) Growth rate
(g/week)b
FCRb
– – –
– – –
– – –
– – –
– – –
1.87� 0.08 0.39 –
2.06� 0.07 0.18 –
2.60� 0.20 0.54 1.78
3.27� 0.21 0.68 1.84
4.19� 0.27 0.80 1.39
5.61� 0.28 1.42 1.18
8.51� 0.49 2.90 0.55
10.92� 0.55 2.41 0.68
12.12� 0.52 1.20 1.46
13.24� 0.65 1.13 1.49
14.12� 0.37 1.03 1.47
15.47�1.08 1.34 1.35
17.13� 0.92 1.66 1.09
18.13� 0.41 1.00 2.20
19.32�1.45 1.19 1.19
– – –
but only mean values were used in the respective photosynthesis and respiration
hat each mean value used in the equations resulted from 25 measurements.
Ta
ble
4D
isso
lve
do
xy
ge
n(m
g/L
),te
mp
era
ture
(8C
),sa
lin
ity
(pp
m),
lig
ht
irra
dia
nce
( mm
olp
ho
ton
s/m
2s)
,pH
,alk
ali
nit
y(m
g/L
),to
tala
mm
on
ium
nit
rog
en
-TA
N(m
g/L
),n
itri
te(m
g/L
),n
itra
te(m
g/L
),d
isso
lve
do
rth
op
ho
sph
ate
(mg
/L),
turb
idit
y
(NT
U),
tota
lsu
spe
nd
ed
soli
ds:
TS
S(m
g/L
)a
nd
vo
lati
lesu
spe
nd
ed
soli
ds:
VS
S(m
g/L
)p
rese
nt
ina
sup
er-
inte
nsi
ve
race
wa
yo
fLi
top
ena
eus
va
nn
am
eiw
ith
zero
wa
ter
ren
ew
al
du
rin
gth
eo
bse
rva
tio
np
eri
od
(th
ev
alu
es
are
ex
act
an
d
corr
esp
on
dto
asp
eci
fic
da
te,
soth
ey
do
no
th
av
ea
sta
nd
ard
de
via
tio
n).
We
ek
O2
Te
mp
era
ture
Lig
ht
Sa
lin
ity
ap
HA
lka
lin
ity
(me
an�
SD)
TA
N
(me
an�
SD)
N-N
O2�
(me
an�
SD)
N-N
O3�
(me
an�
SD)
PO
4�
(me
an�
SD)
NT
U
(me
an�
SD)
TS
S
(me
an�
SD)
VS
S
(me
an�
SD)
84
.22
5.4
–3
4.0
7.1
11
3.8�
17
0.7
1�
0.0
95
.38�
0.1
13
4.3�
0.9
33
6.7�
0.3
13
8.5�
3.2
94
03
.3�
12
.23
50
.0�
22
.2
95
.72
8.2
–3
3.3
7.4
51
.3�
11
0.7
2�
0.0
29
.49�
0.1
17
8.4�
2.2
71
7.3�
0.1
24
6.4�
3.1
45
53
.3�
9.8
84
50
.0�
19
.6
10
7.5
26
.13
5.1
34
.27
.38
2.0�
21
0.5
9�
0.0
22
.28�
0.1
78
1.2�
1.9
83
7.5�
0.4
55
2.3�
2.3
14
33
.3�
2.6
73
96
.0�
23
.1
11
4.1
27
.64
8.9
34
.97
.01
34
.7�
09
0.4
1�
0.0
30
.72�
0.1
11
13
.2�
3.9
50
.9�
1.0
62
5.0�
3.0
04
13
.3�
11
.93
46
.7�
26
.2
12
11
27
.41
4.3
34
.97
.2–
0.4
1�
0.1
20
.62�
0.1
44
7.4�
2.2
24
8.1�
1.1
92
0.3�
2.1
15
73
.3�
13
.12
70
.0�
13
.6
13
8.9
32
.76
2.9
34
.26
.91
82
.1�
33
0.3
8�
0.0
80
.16�
0.0
54
0.6�
1.6
57
2.5�
0.9
82
0.0�
1,1
94
90
.0�
7.9
02
15
.0�
13
.1
14
6.9
32
.84
0.5
35
.36
.61
30
.7�
19
1.1
0�
0.0
90
.19�
0.0
26
6.7�
1.6
11
02
.3�
2.2
30
.4�
1.9
36
70
.0�
17
.22
52
.5�
28
.4
15
6.4
31
.4–
32
.16
.61
26
.5�
21
0.7
3�
0.0
70
.23�
0.0
82
2.9�
4.5
85
4.5�
4.2
92
6.0�
2,6
55
46
.6�
22
.92
36
.7�
29
.3
16
4.2
31
.7–
31
.96
.46
5.3�
18
0.4
4�
0.1
70
.11�
0.0
71
9.9�
2.2
28
8.5�
2.3
7–
77
0.0�
17
.12
96
.7�
12
.1
17
7.2
30
.63
8.0
32
.56
.55
8.1�
22
0.6
1�
0.1
10
.44�
0.0
76
9.4�
2.2
11
08
.7�
1.8
–7
50
.0�
16
.13
80
.0�
31
.8
18
8.2
31
.32
6.9
33
.16
.31
07
.2�
13
0.2
6�
0.0
90
.72�
0.1
17
5.8�
1.9
91
31
.2�
2.9
–8
00
.0�
39
.43
05
.0�
29
.2
19
9.8
30
.21
6.2
31
.56
.41
43
.6�
16
0.1
6�
0.0
90
.23�
0.0
67
6.3�
3.0
11
31
.9�
3.5
29
.7�
3.0
17
35
.0�
19
.93
50
.0�
33
.0
20
8.9
31
.62
3.3
32
.66
.3–
0.6
5�
0.1
20
.08�
0.0
29
4.2�
2.7
3–
30
.4�
2.9
8–
–
21
6.6
31
.82
9.4
33
.1–
––
––
––
––
22
––
13
.8–
––
––
––
––
–
23
––
3.2
––
––
––
––
––
aT
he
incr
ea
sein
sali
nit
yca
use
db
ye
va
po
rati
on
wa
sco
ntr
oll
ed
by
the
ad
dit
ion
of
fre
shw
ate
r.
L. Vinatea et al. / Aquacultural Engineering 42 (2010) 17–24 21
solids (mg/L) and volatile suspended solids (mg/L) found in theraceway throughout the observation period.
Table 5 summarizes the significant correlations (Spearman) ofgross photosynthesis, water column respiration, shrimp growthrate and FCR when compared with the other water qualityparameters that resulted in p < 0.05. Considering the water qualityparameters as independent variables, gross photosynthesis wassignificantly related only with VSS and water column respirationonly with turbidity. Shrimp growth rate increased with highertemperature and decreased with higher nitrite, VSS, and pH. Aswould be expected, the FCR increased with higher nitrite and VSS.Nitrite and nitrate concentrations were also positively related withVSS levels.
Fig. 2 demonstrates an inverse logarithmic relationship(R2 = 0.8044, p < 0.001) between nitrite concentration and shrimpgrowth rate, and a weak direct linear relationship (R2 = 0.3319,p = 0.0284) between nitrite and FCR. Nitrite is also directly relatedto VSS (R2 = 0.4243, p < 0.0159) (Fig. 3). VSS also has a significantinverse relationship with shrimp growth rate (R2 = 0.4989,p = 0.0102) and a direct relationship with FCR (R2 = 0.5356,p = 0.0162) (Fig. 4).
4. Discussion
The method for the light and dark bottles (rotating wheelwithin a water bath) used to measure photosynthesis and watercolumn respiration provided good results from a practicalstandpoint. The fact that there were no significant differencesbetween measurements made under field conditions in theraceway and in the laboratory confirms the practicality of routinelymaking those measurements under totally controlled conditions oflight and temperature, independent of weather variability.
The amount of particulate matter suspended in the watercolumn apparently affected the amount of photosynthesis thatcould take place. This is demonstrated by the inverse relationshipbetween VSS and photosynthesis. High levels of particulate matterreduce light penetration and decrease photosynthesis. Theparticulate matter probably also provided considerable substratefor bacteria and other organisms to flourish as demonstrated by thedirect relationship between water column respiration andturbidity.
In this study shrimp growth rate was directly related totemperature, which is understandable since L. vannamei is atropical ectothermal organism with its metabolic rate a function ofambient temperature (Zhang et al., 2006). This has also beendemonstrated in other penaeids like M. japonicus (Chen and Lai,1993), F. chinensis (Chen and Nan, 1993) and F. californiensis
(Villareal and Ocampo, 1993).This experiment seems to confirm reports by Burford et al.
(2004), Cohen et al. (2005) and Brune et al. (2003), thatphytoplankton (and as a consequence, photosynthesis) have onlya limited impact on the growth of shrimp cultivated at highdensities in a closed system. In fact, at least under the observedconditions in the raceway where this study was done, nosignificant relationship between photosynthesis and growth rateor the animal’s FCR was found. Net photosynthesis was usuallynegative, i.e. there was an oxygen deficit, suggesting that theculture was predominantly heterotrophic. If the net photosynth-esis measurements were extrapolated to a 24-h period, there islittle doubt that the culture system operated heterotrophically. Inspite of that, photosynthetic processes were always minima, a factthat was confirmed by the presence of photoautotrophic micro-organisms (chlorophytes, cyanobacteria and dinoflagellates)throughout the duration of the culture.
Nitrogen accumulation in the super-intensive culture systemspresents an important problem to be addressed (Burford and
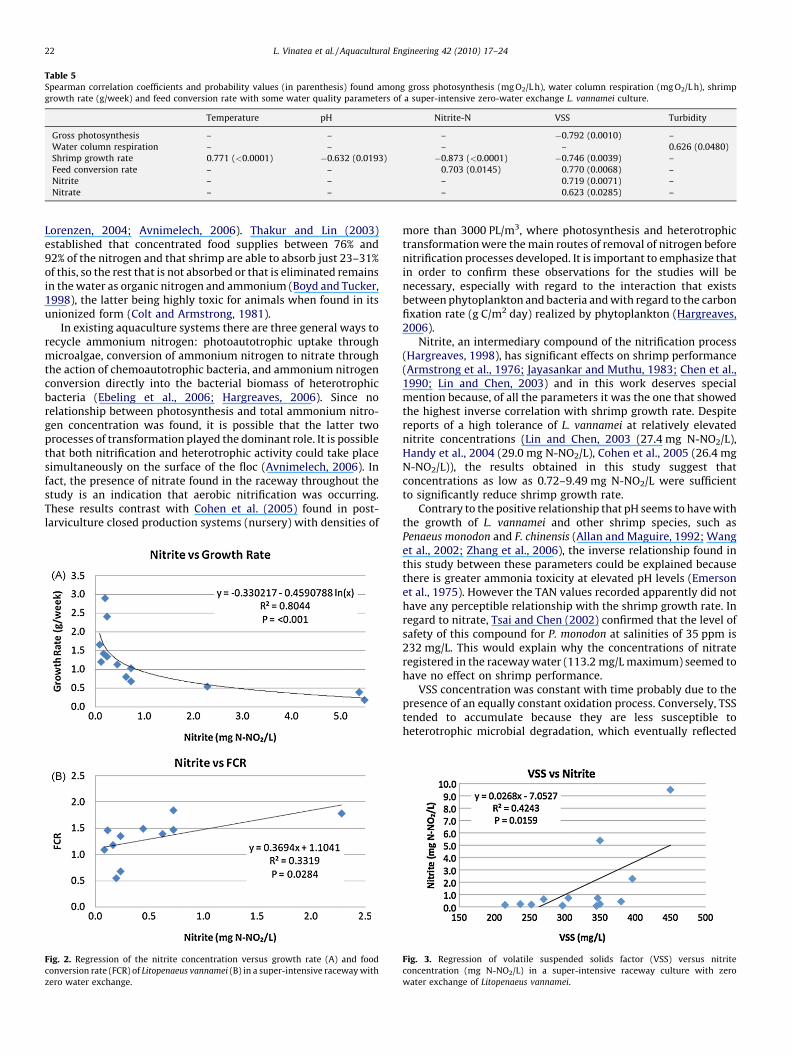
Table 5Spearman correlation coefficients and probability values (in parenthesis) found among gross photosynthesis (mg O2/L h), water column respiration (mg O2/L h), shrimp
growth rate (g/week) and feed conversion rate with some water quality parameters of a super-intensive zero-water exchange L. vannamei culture.
Temperature pH Nitrite-N VSS Turbidity
Gross photosynthesis – – – �0.792 (0.0010) –
Water column respiration – – – – 0.626 (0.0480)
Shrimp growth rate 0.771 (<0.0001) �0.632 (0.0193) �0.873 (<0.0001) �0.746 (0.0039) –
Feed conversion rate – – 0.703 (0.0145) 0.770 (0.0068) –
Nitrite – – – 0.719 (0.0071) –
Nitrate – – – 0.623 (0.0285) –
L. Vinatea et al. / Aquacultural Engineering 42 (2010) 17–2422
Lorenzen, 2004; Avnimelech, 2006). Thakur and Lin (2003)established that concentrated food supplies between 76% and92% of the nitrogen and that shrimp are able to absorb just 23–31%of this, so the rest that is not absorbed or that is eliminated remainsin the water as organic nitrogen and ammonium (Boyd and Tucker,1998), the latter being highly toxic for animals when found in itsunionized form (Colt and Armstrong, 1981).
In existing aquaculture systems there are three general ways torecycle ammonium nitrogen: photoautotrophic uptake throughmicroalgae, conversion of ammonium nitrogen to nitrate throughthe action of chemoautotrophic bacteria, and ammonium nitrogenconversion directly into the bacterial biomass of heterotrophicbacteria (Ebeling et al., 2006; Hargreaves, 2006). Since norelationship between photosynthesis and total ammonium nitro-gen concentration was found, it is possible that the latter twoprocesses of transformation played the dominant role. It is possiblethat both nitrification and heterotrophic activity could take placesimultaneously on the surface of the floc (Avnimelech, 2006). Infact, the presence of nitrate found in the raceway throughout thestudy is an indication that aerobic nitrification was occurring.These results contrast with Cohen et al. (2005) found in post-larviculture closed production systems (nursery) with densities of
Fig. 2. Regression of the nitrite concentration versus growth rate (A) and food
conversion rate (FCR) of Litopenaeus vannamei (B) in a super-intensive raceway with
zero water exchange.
more than 3000 PL/m3, where photosynthesis and heterotrophictransformation were the main routes of removal of nitrogen beforenitrification processes developed. It is important to emphasize thatin order to confirm these observations for the studies will benecessary, especially with regard to the interaction that existsbetween phytoplankton and bacteria and with regard to the carbonfixation rate (g C/m2 day) realized by phytoplankton (Hargreaves,2006).
Nitrite, an intermediary compound of the nitrification process(Hargreaves, 1998), has significant effects on shrimp performance(Armstrong et al., 1976; Jayasankar and Muthu, 1983; Chen et al.,1990; Lin and Chen, 2003) and in this work deserves specialmention because, of all the parameters it was the one that showedthe highest inverse correlation with shrimp growth rate. Despitereports of a high tolerance of L. vannamei at relatively elevatednitrite concentrations (Lin and Chen, 2003 (27.4 mg N-NO2/L),Handy et al., 2004 (29.0 mg N-NO2/L), Cohen et al., 2005 (26.4 mgN-NO2/L)), the results obtained in this study suggest thatconcentrations as low as 0.72–9.49 mg N-NO2/L were sufficientto significantly reduce shrimp growth rate.
Contrary to the positive relationship that pH seems to have withthe growth of L. vannamei and other shrimp species, such asPenaeus monodon and F. chinensis (Allan and Maguire, 1992; Wanget al., 2002; Zhang et al., 2006), the inverse relationship found inthis study between these parameters could be explained becausethere is greater ammonia toxicity at elevated pH levels (Emersonet al., 1975). However the TAN values recorded apparently did nothave any perceptible relationship with the shrimp growth rate. Inregard to nitrate, Tsai and Chen (2002) confirmed that the level ofsafety of this compound for P. monodon at salinities of 35 ppm is232 mg/L. This would explain why the concentrations of nitrateregistered in the raceway water (113.2 mg/L maximum) seemed tohave no effect on shrimp performance.
VSS concentration was constant with time probably due to thepresence of an equally constant oxidation process. Conversely, TSStended to accumulate because they are less susceptible toheterotrophic microbial degradation, which eventually reflected
Fig. 3. Regression of volatile suspended solids factor (VSS) versus nitrite
concentration (mg N-NO2/L) in a super-intensive raceway culture with zero
water exchange of Litopenaeus vannamei.

Fig. 4. Regression of volatile suspended solids factor (VSS) versus growth rate (A)
and food conversion rate (FCR) of Litopenaeus vannamei (B) in a super-intensive
raceway culture with zero water exchange.
L. Vinatea et al. / Aquacultural Engineering 42 (2010) 17–24 23
in the increase in NTU. However, the dynamics of organic materialin high-density cultures is complex and poorly understood(Hargreaves, 2006). De Schryver et al. (2008) stated that there isno established range with respect to the optimum level of volatilesuspended solids for bio-floc technology culture systems. In thisstudy VSS, with concentrations of 215–450 mg/L, had an inverserelationship with photosynthesis and shrimp growth rate, and adirect relationship with FCR. That is, the larger the concentrationsof VSS the lower the photosynthetic rate of the system, the lowerthe weekly weight gain of the animals and the greater the FCR.With respect to the inverse relationship between VSS and shrimpgrowth rate, it is possible to conclude that in this case theconcentrations of VSS were directly detrimental to the develop-ment of the shrimp. According to Burford and Lorenzen (2004),organic suspended solids affect the animals in both a direct waythrough the demand for oxygen, and an indirect way through theanaerobic formation of toxic metabolites, mainly ammonium andnitrite. This final observation seems to be corroborated by thedirect relationship found in this study between VSS and bothnitrite and nitrate. At least nitrite is a parameter that issignificantly influenced by other factors, including the densityand the type of substrate of the culture (Thakur and Lin, 2003). Thiscalls to attention the fact that no significant relationship of eitherturbidity (20–52.3 NTU) or TSS (403.3–800 mg/L) with growth rateand FCR was observed. Although these parameters have been usedin managing cultivated fish (Avnimelech, 2006), both havesignificant logistical problems, compared to VSS, which limit theirutility in saltwater bio-floc systems.
For the conditions of this study, it is possible to conclude thatphotosynthesis was reduced by increased levels of volatilesuspended solids (VSS), while water respiration increased indirect proportion to turbidity. The final productivity of L. vannamei
during the course of the culture was influenced directly only bytemperature and inversely by pH, nitrite and volatile suspendedsolids. Photosynthesis and water column respiration, in spite ofsignificant relationships with some water quality parameters, didnot have a significant impact on shrimp productivity. It is stronglyrecommended that the same analyses be repeated in future trialsto confirm these observations and to establish possible character-istics shrimp performance and water quality dynamics present inzero water exchange super-intensive shrimp culture.
Acknowledgements
The first and second authors would like to thank the ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico (CNPq-MCT) and to Coordenacao de Aperfeicoamento de Pessoal de NıvelSuperior (CAPES-MEC), both of the Brazilian government, by thepost-doctorate’s grants. Our thanks go to Waddell MaricultureCenter (South Carolina Department of Natural Resources) staff, forall the help offered.
References
Allan, G., Maguire, G., 1992. Lethal levels of low dissolved oxygen and effects ofshort-term oxygen stress on subsequent growth of juvenile Penaeus monodon.Aquaculture 94, 27–37.
Armstrong, D., Stephenson, M., Knight, A., 1976. Acute toxicity of nitrite to larvae ofthe giant Malayasian prawn, Macrobrachium rosembergii. Aquaculture 9, 39–46.
Avnimelech, Y., 1999. Carbon/nitrogen ratio as a control element in aquaculturesystems. Aquaculture 176, 227–235.
Avnimelech, Y., 2006. Bio-filters: the need for a new comprehensive approach.Aquacult. Eng. 34, 172–178.
Azim, K., Little, D., 2008. The biofloc technology (BFT) in indoor tanks: water quality,biofloc composition, and growth and welfare of Nile tilapia (Oreochromisniloticus). Aquaculture 283, 29–35.
Bailey, C., 1988. The social consequences of tropical shrimp mariculture develop-ment. Ocean Shoreline Manage., Lond. (11), 31–44.
Boyd, C., Tucker, C., 1998. Pond Aquaculture Water Quality Management. KluwerAcademic Publishers, Boston, MA.
Browdy, C., Bratvold, D., Stokes, A., McIntosh, R., 2001. Perspectives on the applica-tion of closed shrimp culture systems. In: Browdy, C., Jory, D. (Eds.), The NewWave, Proceedings of the Special Session on Sustainable Shrimp Culture,Aquaculture 2001. The World Aquaculture Society, Baton Rouge, pp. 20–34.
Brown, J., 1989. Antibiotics: their use and abuse in aquaculture. World Aquacult.,Baton Rouge 20 (2), 34–43.
Brune, D., Schwartz, G., Eversole, A.G., Collier, J.A., Schwedler, T.E., 2003. Intensifica-tion of pond aquaculture and high rate photosynthetic system. Aquacult. Eng.28 (1–2), 65–86.
Burford, M., Lorenzen, K., 2004. Modeling nitrogen dynamics in intensive shrimpponds: the role of sediment remineralization. Aquaculture 229, 129–145.
Burford, M., Thompson, P., McIntosh, R., Bauman, R., Pearson, D., 2003. Nutrient andmicrobial dynamics in high-intensity, zero-exchange shrimp ponds in Belize.Aquaculture 219, 393–411.
Burford, M., Thompson, P., McIntosh, R., Bauman, R., Pearson, D., 2004. The con-tribution of flocculated material to shrimp (Litopenaeus vannamei) nutrition in ahigh-intensity zero-exchange system. Aquaculture 232, 525–537.
Chang, W., Ouyang, H., 1988. Dynamics of dissolved oxygen and vertical circulationin fish ponds. Aquaculture 74 (3–4), 263–276.
Chen, J., Lai, S., 1993. Effects of temperature and salinity on oxygen consumptionand ammonia-N excretion of juvenile Penaeus japonicus Bate. J. Exp. Mar. Biol.Ecol. 165, 161–170.
Chen, J., Nan, F., 1993. Effects of temperature, salinity and ambient ammonia onlethal dissolved oxygen of Penaeus chinensis. Comp. Biochem. Physiol. 101C,459–461.
Chen, J., Liu, P., Lei, S., 1990. Toxicities of ammonia and nitrite to Penaeus monodonadolescents. Aquaculture 89, 127–137.
Cohen, J., Samocha, T., Fox, J., Gandy, R., Lawrence, A., 2005. Characterization ofwater quality factors during intensive raceway production of juvenile Litope-naeus vannamei using limited discharge and biosecure management tools.Aquacult. Eng. 32, 425–442.
Colt, J., Armstrong, D., 1981. Nitrogen toxicity to crustaceans, fish and mollusks. In:Allen, L., Kinney, E. (Eds.), Proceedings of the Bioengineering Symposium forFish Culture. Fish Culture Section of the American Fisheries Society, Bethesda,MD, USA, pp. 34–47.
Currie, D., 1994. Sustainable aquaculture in developing countries. World Aquacult.,Baton Rouge 25 (4), 20–25.
Davis, D., Arnold, C., 1998. The design, management and production of a recirculat-ing raceway system for the production of marine shrimp. Aquacult. Eng. 17,193–211.
De Schryver, P., Crab, R., Defoirdt, T., Boon, N., Verstraete, W., 2008. The basics of bio-flocs technology: the added value for aquaculture. Aquaculture 277, 125–137.

L. Vinatea et al. / Aquacultural Engineering 42 (2010) 17–2424
Ebeling, J., Timmons, M., Bisogni, J., 2006. Engineering analysis of the stoichiometryof photoautotrophic, autotrophic, and heterotrophic removal of ammonia-nitrogen in aquaculture systems. Aquaculture 257, 346–358.
Emerson, K., Russo, R., Lund, R., Thurston, R., 1975. Aqueous ammonia equilibriumcalculations: effects of pH and temperature. J. Fish. Res. Board Can. 32, 2379–2383.
Handy, M., Samocha, T., Patnaik, S., Gandy, R., McKee, D., 2004. Nursery trialcompares filtration system performance in intensive raceways. Global Aqua-cult. Advoc. 8, 77–79.
Hargreaves, J., 1998. Nitrogen biogeochemistry of aquaculture ponds. Aquaculture166, 181–212.
Hargreaves, J., 2006. Photosynthetic suspended-growth systems in aquaculture.Aquacult. Eng. 34, 344–363.
Hopkins, J., Hamilton II, R., Sandifer, P., Browdy, C., Stokes, A., 1993. Effects of waterexchange rate on production, water quality, effluent characteristics and nitro-gen budgets of intensive shrimp ponds. J. World Aquacult. Soc. 24, 304–320.
Jayasankar, P., Muthu, M., 1983. Toxicity of nitrite to the larvae of Penaeus indicus.Indian J. Fisheries 30 (2), 231–240.
Kadir, A., Kundu, R., Milstein, A., Wahab, M., 2006. Effects of silver carp and smallindigenous species on pond ecology and carp polycultures in Bangladesh.Aquaculture 261 (2006), 1065–1076.
Kestemont, P., 1995. Different systems of carp production and their impacts on theenvironment. Aquaculture 129, 347–372.
Kinkelin, P., Michel, C., 1992. The use of drugs in aquaculture. Infofish Int., KualaLumpur 4 (92), 45–49.
Larsson, J., Folke, C., Kautsky, N., 1995. Ecological limitations of shrimp farming inColombia. In: Calderon, J., Sorgeloos, P. (Eds.), Memorias del Segundo CongresoEcuatoriano de Acuicultura. Escuela Superior Politecnica del Litoral, Guayaquil,pp. 175–188.
Laws, E., Malecha, S., 1981. Application of a nutrient-saturated growth model tophytoplankton management in freshwater prawn (Macrobrachium rosenbergii)ponds in Hawaii. Aquaculture 24, 91–101.
Lin, Y., Chen, J., 2003. Acute toxicity of nitrite on Litopenaeus vannamei (Boone)juveniles at different salinity levels. Aquaculture 224, 193–201.
Macintosh, D., Phillips, M., 1992. Environmental issues in shrimp farming. In:Saram, H., Singh, T. (Eds.), Shrimp 92, Proceedings of the 3th Global Conferenceon the Shrimp Industry, Kuala Lumpur, INFOFISH, pp. 118–145.
McAbee, B., Browdy, C., Rhodes, R., Stokes, A., 2003. The use of greenhouse-enclosed raceway systems for the super-intensive production of pacific whiteshrimp Litopenaeus vannamei in the United States. Global Aquacult. Advoc. 6(3), 40–43.
McNeil, R., 2000. Zero exchange, aerobic, heterotrophic systems: key considera-tions. Global Aquacult. Advoc. 3 (3), 72–76.
Moss, S., Pruder, G., 1995. Characterization of organic particles associated withrapid growth in juvenile white shrimp, Penaeus vannamei Boone, reared underintensive culture conditions. J. Exp. Mar. Biol. Ecol. 187 (2), 175–191.
Noriega-Curtis, P., 1979. Primary productivity and related fish yield in intenselymanured fishponds. Aquaculture 17 (4), 335–344.
Perez, J., 1993. La conservacion de los recursos geneticos de organismos acuaticos.Interciencia 18 (4), 190–194.
Primavera, H., 1994. Environmental and socioeconomic effects of shrimp farming:the Philippine experience. Infofish Int., Kuala Lumpur (1/94), 44–48.
Pruder, G., 1992. Marine shrimp pond effluent: characterization and environmentalimpact. In: Wyban, J. (Ed.), Proceedings of Special Session on Shrimp Farming ofWAS ‘92. The World Aquaculture Society, Baton Rouge, pp. 187–190.
Pruder, G., 2004. Biosecurity: application in aquaculture. Aquacult. Eng. 32, 3–10.Reid, B., Arnold, C., 1992. The intensive culture of the Penaeid shrimp Penaeus
vannamei Boone in a recirculating raceway system. J. World Aquacult. Soc. 23,146–153.
Rosenthal, H., 1994. Aquaculture and the environment. World Aquacult., BatonRouge 25 (2), 4–11.
Sandifer, P., Hopkins, J., 1996. Conceptual design of a sustainable pond-basedshrimp culture system. Aquacult. Eng. 15, 41–52.
Schveitzer, R., Andreatta, E., Souza, J., Arantes, R., Seiffert, W., 2008. O cultivo combioflocos: engorda e formacao de matrizes de Litopenaeus vannamei. Panoramada Aquicultura 107, 38–43.
Sowers, A., Gatlin, D., Young, S., Isely, J., Browdy, C., Tomasso, J., 2005. Responses ofLitopenaeus vannamei (Boone) in water containing low concentrations of totaldissolved solids. Aquacult. Res. 36, 819–823.
Strickland, J., 1960. Measuring the production of marine phytoplankton. Bull. Fish.Res. Bd. Canada 122 172 pp.
Thakur, D., Lin, C., 2003. Water quality and nutrient budget in closed shrimp(Penaeus monodon) culture systems. Aquacult. Eng. 27, 159–176.
Tsai, S., Chen, J., 2002. Acute toxicity of nitrate on Penaeus monodon juveniles atdifferent salinity levels. Aquaculture 213, 163–170.
Villareal, H., Ocampo, L., 1993. Effect of size and temperature on the oxygenconsumption of the brown shrimp Penaeus californiensis (Holmes, 1900). Comp.Biochem. Physiol. 106A (1), 97–101.
Wang, W., Wang, A., Chen, L., Liu, Y., Sun, R., 2002. Effects of pH on survival,phosphorus concentration, adenylate energy charge and Na+-K+ ATPase activ-ities of Penaeus chinensis Osbeck juveniles. Aquat. Toxicol. 60, 75–83.
Wasielesky, W., Atwood, H., Stokes, A., Browdy, C., 2006. Effect of natural productionin a zero exchange suspended microbial floc based super-intensive system forwhite shrimp Litopenaeus vannamei. Aquaculture 258, 396–408.
Williams, A., Davis, D., Arnold, C., 1996. Density-dependent growth and survival ofPenaeus setiferus and Penaeus vannamei in a semi-closed recirculating system. J.World Aquac. Soc. 27, 107–112.
Yusoff, F., McNabb, C., 1989. Effects of nutrient availability on primary productivityand fish production in fertilized tropical ponds. Aquaculture 78 (3–4), 303–319.
Zhang, P., Zhang, X., Li, J., Huang, G., 2006. The effects of body weight, temperature,salinity, pH, light intensity and feeding condition on lethal DO levels of whitelegshrimp, Litopenaeus vannamei (Boone, 1931). Aquaculture 256, 579–587.




















