
Phospholipid oxidation, non-enzymatic browning development and volatile compounds generation in...
11
Eur Food Res Technol (2007) 225:665–675 DOI 10.1007/s00217-006-0462-2 ORIGINAL PAPER Phospholipid oxidation, non-enzymatic browning development and volatile compounds generation in model systems containing liposomes from porcine Longissimus dorsi and selected amino acids Sonia Ventanas · Mario Est´ evez · Carmen L. Delgado · Jorge Ruiz Received: 16 January 2006 / Revised: 25 July 2006 / Accepted: 9 August 2006 / Published online: 15 September 2006 C Springer-Verlag 2006 Abstract Lipid oxidation, non-enzymatic browning devel- opment and volatile compounds generation were studied in sterile meat-model systems containing selected amino acids and/or liposomes during 35 days at 25 ◦ C under pro-oxidative conditions, in order to simulate the ripening conditions of dry-cured meat products. Liposomes were prepared with polar lipids (PL) from Longissimus dorsi muscle of out- doors pigs fed on grass and acorns (M) or indoors ones fed on concentrates (C). Thiobarbituric acid reactive sub- stances (TBA-RS) in systems containing both amino acids and liposomes were higher than in those containing only liposomes. The higher susceptibility to lipid oxidation of liposomes from C animal was reflected in higher TBA-RS throughout the experiment. All model systems containing li- posomes and/or amino acids showed non-enzymatic brown- ing development, as measured by the yellowness index. Ma- jor volatile compounds were fatty acid oxidation products. Strecker-derived volatile compounds were predominantly detected in model systems containing both amino acids and liposomes. The chromatographic areas of Strecker-derived volatiles increased over time whereas those of lipid-derived volatiles decreased. The occurrence of lipid oxidation, non- enzymatic browning and Strecker-type degradation of amino acids were shown in sterile meat-model systems at low temperatures. Keywords Lipid oxidation . Non-enzymatic browning . Strecker-type degradation . Liposomes . Model system S. Ventanas · M. Est´ evez · C. L. Delgado · J. Ruiz () Food Science, School of Veterinary Sciences, University of Extremadura, C´ aceres, Spain e-mail: [email protected] Introduction Ripening conditions (processing time and temperature) and fatty acid compositions of polar lipids (PL) of raw meat, are two of the most important factors influencing the profile of volatile compounds of dry-cured products and particularly of Iberian dry-cured hams. The particular processing conditions (temperatures from 15 to 25 ◦ C, pH near 6, water activities around 0.85 and a processing time up to 30 months) during the processing of Iberian dry-cured hams lead to the occurrence of lipid oxi- dation phenomena [1–3] and could favour the development of Maillard reactions, including the oxidative-deamination- decarboxylation of amino acids via Strecker degradation [4]. The enormous amount of free amino acids released during the drying phase [5], together with the formation of reactive carbonyls from lipid oxidation [3], could enable the subse- quent development of these type of Maillard reactions, and thus the generation of certain branched aldehydes, such as 3-methylbutanal, that have been described as major contribu- tors to the desirable flavour of Iberian ham [6]. Nevertheless, other authors [7, 8] have claimed that Strecker degradation of amino acids is not a possible mechanism for methyl- branched aldehydes formation in dry-cured hams, since the temperatures reached during ripening are not high enough. Consequently, these authors pointed out that the only possi- ble source of these aldehydes in dry-cured ham is the micro- bial metabolism. The role of PL in the development of the flavour of meat and meat products has been discussed extensively [9]. PL are considered the main precursors of lipid-oxidation products in meat [10] due to the high degree of unsaturation of their fatty acids, together with the fact that, as components of cells membranes, they are exposed to proteins and other catalysts of lipid oxidative reactions such as metals [11]. Aldehydes, Springer
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Phospholipid oxidation, non-enzymatic browning development and volatile compounds generation in...

Eur Food Res Technol (2007) 225:665–675DOI 10.1007/s00217-006-0462-2
ORIGINAL PAPER
Phospholipid oxidation, non-enzymatic browning developmentand volatile compounds generation in model systems containingliposomes from porcine Longissimus dorsi and selected amino acidsSonia Ventanas · Mario Estevez · Carmen L. Delgado ·Jorge Ruiz
Received: 16 January 2006 / Revised: 25 July 2006 / Accepted: 9 August 2006 / Published online: 15 September 2006C© Springer-Verlag 2006
Abstract Lipid oxidation, non-enzymatic browning devel-opment and volatile compounds generation were studied insterile meat-model systems containing selected amino acidsand/or liposomes during 35 days at 25 ◦C under pro-oxidativeconditions, in order to simulate the ripening conditions ofdry-cured meat products. Liposomes were prepared withpolar lipids (PL) from Longissimus dorsi muscle of out-doors pigs fed on grass and acorns (M) or indoors onesfed on concentrates (C). Thiobarbituric acid reactive sub-stances (TBA-RS) in systems containing both amino acidsand liposomes were higher than in those containing onlyliposomes. The higher susceptibility to lipid oxidation ofliposomes from C animal was reflected in higher TBA-RSthroughout the experiment. All model systems containing li-posomes and/or amino acids showed non-enzymatic brown-ing development, as measured by the yellowness index. Ma-jor volatile compounds were fatty acid oxidation products.Strecker-derived volatile compounds were predominantlydetected in model systems containing both amino acids andliposomes. The chromatographic areas of Strecker-derivedvolatiles increased over time whereas those of lipid-derivedvolatiles decreased. The occurrence of lipid oxidation, non-enzymatic browning and Strecker-type degradation of aminoacids were shown in sterile meat-model systems at lowtemperatures.
Keywords Lipid oxidation . Non-enzymatic browning .
Strecker-type degradation . Liposomes . Model system
S. Ventanas · M. Estevez · C. L. Delgado · J. Ruiz (�)Food Science, School of Veterinary Sciences,University of Extremadura,Caceres, Spaine-mail: [email protected]
Introduction
Ripening conditions (processing time and temperature) andfatty acid compositions of polar lipids (PL) of raw meat, aretwo of the most important factors influencing the profile ofvolatile compounds of dry-cured products and particularlyof Iberian dry-cured hams.
The particular processing conditions (temperatures from15 to 25 ◦C, pH near 6, water activities around 0.85 and aprocessing time up to 30 months) during the processing ofIberian dry-cured hams lead to the occurrence of lipid oxi-dation phenomena [1–3] and could favour the developmentof Maillard reactions, including the oxidative-deamination-decarboxylation of amino acids via Strecker degradation [4].The enormous amount of free amino acids released duringthe drying phase [5], together with the formation of reactivecarbonyls from lipid oxidation [3], could enable the subse-quent development of these type of Maillard reactions, andthus the generation of certain branched aldehydes, such as3-methylbutanal, that have been described as major contribu-tors to the desirable flavour of Iberian ham [6]. Nevertheless,other authors [7, 8] have claimed that Strecker degradationof amino acids is not a possible mechanism for methyl-branched aldehydes formation in dry-cured hams, since thetemperatures reached during ripening are not high enough.Consequently, these authors pointed out that the only possi-ble source of these aldehydes in dry-cured ham is the micro-bial metabolism.
The role of PL in the development of the flavour of meatand meat products has been discussed extensively [9]. PL areconsidered the main precursors of lipid-oxidation productsin meat [10] due to the high degree of unsaturation of theirfatty acids, together with the fact that, as components of cellsmembranes, they are exposed to proteins and other catalystsof lipid oxidative reactions such as metals [11]. Aldehydes,
Springer

666 Eur Food Res Technol (2007) 225:665–675
the main products of lipid oxidation, seem to be the maincontributors to Iberian ham flavour, since they show lowolfaction thresholds and distinctive odour characters (e.g.rancid, sweet, floral notes) [12, 13]. The fatty acid compo-sition of dietary fat influences the fatty acid composition ofPL [14] and thus, the volatile profile in Iberian dry-curedham [1]. The particular PL profile (high monounsaturated(MUFA) content) of Iberian hams from pigs reared outdoorsand fed on grass and acorns, partly explains the high sensoryquality of this type of hams compared to those from pigsreared indoors and fed on mixed concentrates [15].
Several studies have been carried out in order to elu-cidate the influence of the modifications of the fatty acidcomposition of PL through dietary means, on the volatileprofile in different meat-model systems [16, 17]. Moreover,numerous works have studied the occurrence of Maillardreactions and the interactions between Maillard and lipid-derived products in model systems containing amino acidsor proteins and carbohydrates [18, 19] and/or PL [20, 21].Nevertheless, most of these studies were performed at hightemperatures (above 100 ◦C), since they were focused on thestudy of generation of volatile compounds in cooked meatproducts. In addition, most meat-model systems includedsugars in order to study the volatiles generation via the clas-sical Maillard reaction between reducing sugars and aminoacids.
The objective of the present work was to study theformation of volatile compounds, particularly those fromStrecker degradation of amino acids, at low tempera-tures in model systems mimicking dry-cured productsand containing selected amino acids and liposomes fromL. dorsi with different fatty acids profiles. Lipid oxida-tion and non-enzymatic browning development were alsoevaluated.
Materials and methods
Reagents
l-Leucine (minimum 98% TLC), l-methionine (>99%TLC) and l-phenylalanine (minimum 98% TLC) werepurchased from Sigma-Aldrich Chemie (Steinheim, Ger-many). Piperazine-N-N′-bis-(2-ethanosulfonate) solutions(10 mM, 0.15 M NaCl, pH 6.0 adjusted with NaOH 0.1 N)(PIPES buffer) were prepared with Piperacin-1,4-bis-(2-ethanosulfonic) acid supplied by Panreac Quimica S.A.(Barcelona, Spain) and Milli-Q ultra-pure water (18 M�).Sodium L( + )-ascorbate was supplied by Panreac QuimicaS.A. (Barcelona, Spain), whereas extra pure iron(III) chlo-ride hexahydrate (FeCl3·6H2O) was purchased from Schar-lau Chemie (La Jota, Barcelona, Spain).
Table 1 Studied model systems and their composition
LPS-M LPS-C Phe Met Leu Fe(III)/ascorbate PIPES
M ∗ ∗ ∗
M-AA ∗ ∗ ∗ ∗ ∗ ∗
C ∗ ∗ ∗
C-AA ∗ ∗ ∗ ∗ ∗ ∗
AA ∗ ∗ ∗ ∗ ∗
PIPES ∗ ∗
LPS-M, liposomes from PL of L. dorsi from Iberian pigs reared in“montanera” systems and fed on acorn and grass. LPS-C, liposomesfrom PL of L. dorsi from Iberian pigs reared indoors and fed onconcentrates.
The rest of reagents and solvents were of analytical gradeand purchased from Sigma-Aldrich Chemie (Steinheim, Ger-many).
Experimental design
Different meat-model systems were prepared: (i) AA, con-taining three selected amino acids (Phe, Met and Leu); (ii)M and C, containing liposomes made with PL from M andC pigs, respectively; (iii) M-AA and C-AA, containing bothliposomes and amino acids; and (iv) PIPES, containing onlythe PIPES buffer as blank model system. Liposomes wereprepared from L. dorsi muscles from Iberian pigs reared indifferent systems and fed on different diets: free-range rearedand exclusively fed on acorn and grass according to the tra-ditional “montanera” feeding systems (M) or reared indoorsand fed on concentrates (C). The final composition of the sixstudied model systems is presented in Table 1. The formula-tion of the model systems was chosen in order to simulate,in a simplified way, the main chemical components of dry-cured meat products and taking into account that the majorprecursors of Iberian ham flavour are amino acids and fattyacids, especially those from muscle cell membranes PL [1].
The experiment was carried out at 25 ◦C for 35 daysin a pro-oxidant environment (Fe(III)/ascorbate) in order tosimulate the conditions occurring during certain phases ofIberian hams ripening process in which the most relevantchemical reactions lead to volatile compounds generationare taken place.
The experimental analyses: Volatiles from the headspace,TBA-RS and yellowness indices, were carried out at day 0,7, 21, 28 and 35 of the experiment in each studied modelsystems.
Liposome preparation and characterization
Intramuscular total lipids were extracted according to themethod described by Bligh and Dyer [22]. The PL fractionwas separated from total lipid by using NH2-aminopropyl
Springer

Eur Food Res Technol (2007) 225:665–675 667
minicolumns following the method described by Muriel et al.[23]. To eliminate all traces of solvent (methanol), the PLfraction was maintained under a nitrogen stream for 90 minuntil dryness. Polypropylene cone tubes containing PL wereflushed with nitrogen, capped with Teflon-lined screw capsand stored at frozen conditions prior to liposomes preparationin order to avoid oxidative reactions.
To prepare liposome suspensions, 30 ml of PIPES bufferwere added to tubes containing the PL fraction. The mixturewas vigorously stirred for 2 min, maintained in a thermosta-tized bath at 37 ◦C for 1 h and stirred again for 2 min each10 min in order to achieve the distribution of PL as mul-tilamellar vesicles (MLV). Large unilamellar vesicles wereprepared by extrusion of MLV 10 times through sterilizedcellulose acetated membrane filters (0.20 µm diameter ofpore). Final liposome suspensions were stored in sterilizedpropylene cone tubes at 4 ◦C for 12 h, until model systempreparation.
The PL concentration of liposomes suspensions wasquantified by measuring organic phosphorous following themethod described by Barlett [24]. The PL content was cal-culated considering that their average phosphorous contentwas 4% [25].
The fatty acid composition of PL was determined bypreparing their corresponding fatty acid methyl esters(FAMEs) by acidic trans-esterification in presence of sul-phuric acid (5% sulphuric acid in methanol) [26] prior toanalysis by gas chromatography using a Hewlett-PackardHP-5890A gas chromatograph coupled to a flame ionisationdetector (FID). Separation was carried out on a polyethy-lene glycol-TPA modified fused silica semicapillary column(30 m long, 0.53 mm i.d., 1 µm film thickness) maintainedat 225 ◦C. Injector and detector temperatures were 230 ◦C.The carrier gas was nitrogen at a flow rate of 1.8 ml min−1.Individual FAME peaks were identified by comparing theirretention times with those of standards (Sigma, St. Louis).
The peroxidisability index (PI) reflects the relative ratesof peroxidation of each fatty acids and was calculated fromthe following equation as described by Meynier et al. [16]:PI = (% monoenoic × 0.025) + (% dienoic × 1) + (%trienoic × 2) + (% tetranoic × 4) + (% pentaneoic × 6)+ (% hexanenoic × 8).
Model systems preparation
Three amino acid stock solutions (6.5 mg ml−1) (phenylala-nine, methionine and l-leucine) were prepared with PIPESbuffer and stored at 4 ◦C for 12 h until model systems prepa-ration. Three amino acids with different chemical character-istics were chosen: an aliphatic (L-Leu), an aromatic (Phe)and a sulphur (Met) amino acid. Sodium ascorbate and FeCl3stock solutions in ultra-pure Milli-Q water were daily pre-
pared and mixed in equimolar concentrations just beforebeing used.
All the stock solutions (amino acids, PIPES, FeCl3, ascor-bate and liposome suspensions) were filtered through ster-ilized of cellulose acetate membranes (0.2 µm pore diame-ter) before model system preparation, in order to guaranteethe absence of microbial contamination that could interferewith experimental results. All the laboratory material usedto prepare and store liposome suspensions and model sys-tems was sterilized in an autoclave (SELECTA, presoclave75). Moreover, in order to keep sterile conditions, model sys-tems preparation was carried out in a laminar flow chamber(TELSTAR, AV-30/70). All model systems (final volume of6 ml) were prepared in triplicate, in sterilized glass vials(12 ml) sealed with a teflon/silicone septum and contain-ing a final concentration of 0.5 mg PL/ml of liposomes,0.5 mg/ml of each amino acid and 45 µM of pro-oxidant so-lution (Fe(III)/ascorbate). The different components of themodel systems were poured into sterilized glass vials fromthe corresponding stock solutions in the following order:PIPES, liposomes, amino acids and finally the pro-oxidantsolution. All the mixed model systems were kept in an or-bital shaker device (Innova 4000, PACISA, New Bruswickscientific) at 25 ◦C in constant agitation (125 rpm) for 35days.
TBA reactive substances (TBA-RS)
Thiobarbituric acid reactive substances (TBA-RS) were mea-sured according to the method described by Buege and Aust[27] from aliquots of 0.250 ml of each model system. Briefly,2 ml of the TBA reagent [trichloroacetic acid (TCA) 15%,TBA 0.375%, hydrochloric acid (HCL) 0.25 M] and 750 µlof ultra-pure water (Milli-Q) were added to TBA tubes con-taining the aliquot. The reaction mixture was heated at 100 ◦Cfor 15 min. After cooling, the samples were centrifuged at2000 rpm for 15 min. The absorbance of supernatant wasmeasured at 532 nm, against a blank with a spectropho-tometer (Hitachi, U-2000). The results were expressed innmoles equivalent malonaldehyde per milligram PL (nmoleeq. MDA mg−1 PL) using the molar extinction coefficient ofthe TBA–MDA adducts at 532 nm (1.56 × 105 M−1 cm−1).
Browning measurement
The browning of the solutions (1 ml) was determined by us-ing the weighted-ordinate method [28]. Tristimulus values(X, Y, Z) were calculated from the transmittances (T) obtainedby spectrophotometry from 400 to 700 nm at constants inter-vals of 10 nm. Transmittance values were then converted intothe corresponding Tristimulus and into the CIELAB L∗, a∗
and b∗ colour values [29]. The yellowness index at different
Springer

668 Eur Food Res Technol (2007) 225:665–675
times was calculated according to Francis and Clydeslade[30] as follow: yellowness index = 142.86b∗/L∗.
Analysis of volatile compounds from the headspaceof model systems by SPME-GC-MS
Volatile compounds were analysed by HS-SPME and GC–MS (gas chromatograph Hewlett-Packard 5890 series II cou-pled to a mass selective detector Hewlett-Packard HP-5791A). One millimetre from each model system was poured intoa vial (4 ml) and sealed with a teflon/silicone septum. A mag-netic stirrer was placed into the model systems for stirringduring the extraction. An SPME fibre (85 µm carboxen-polidimethylsiloxane coating) was inserted throughthe septum and exposed to the headspace of thevial.
Vials were preconditioned for 30 min at 37 ◦C. Extractionwas carried out at 37 ◦C for 30 min in a water bath. Af-ter extraction, the SPME fibre was immediately transferredto the injector of the chromatograph (gas chromatographHewlett-Packard 5890 series II) which was in splitless modeat 280 ◦C. The fibre was desorbed for 30 min. The separa-tion of volatile compounds was performed on a 5% phenyl-methyl silicone (HP-5) bonded phase fused silica capillarycolumn (Hewlett-Packard, 50 m × 0.32 mm i.d., film thick-ness 1.05 µm), with helium as carrier gas operating at 6 psiof column head pressure, resulting in a flow of 1.3 ml min−1
at 40 ◦C. Oven program was 40 ◦C for 10 min, 5 ◦C min−1 to200 ◦C, 20 ◦C min−1 to 250 ◦C, and held 250 ◦C for 5 min.Transfer line to the mass spectrometer was maintained at280 ◦C. The mass spectra were obtained using a mass se-lective detector (Hewlett-Packard HP-5791 A) by electronicimpact at 70 eV, a multiplier voltage of 1756 V and col-lecting data at a rate of 1 scan s−1 over the m/z range 30–500.
Identification of volatile compounds
Eighteen volatile compounds were positively identified bycomparing their linear retention indices (LRI) and mass spec-tra with those of standard compounds (Sigma, St. Louis,USA) run in the same conditions. The rest were tentativelyidentified by comparing their mass spectra with those re-ported in the Wiley library and by comparing their LRI withthose reported in the literature [31] and in the NIST database[32]. Results from the volatiles analysis are provided in areaunits (AU).
Microbial analysis
An aliquot of 100 µl was taken from the model systemsthe last day of the experiment (day 35) and inoculated inPetri plates containing Standard plate count agar (PCA) in
Table 2 Fatty acid profile and peroxidisability index (PI) of polarlipids from liposomes included in the studied model systems
∑SFA
∑MUFA
∑PUFA
∑n6
∑n3 PI
LPS-C 33.03 15.44 51.53 48.79 2.32 97.09LPS-M 44.24 16.60 38.21 35.74 2.10 76.55
Results of total SFA, MUFA, PUFA, n6 and n3 are expressed in per-centage of total FAME. LPS-M, liposomes from PL of L. dorsi fromIberian pigs reared in “montanera” systems and fed on acorn and grass.LPS-C, liposomes from PL of L. dorsi from Iberian pigs reared indoorsand fed on concentrates.
order to confirm the absence of microbial growth in themodel systems during the experiment. Plates were examinedafter 48 h of incubation at 30 ◦C for microbial counts. Allthe model systems that presented microbial growth werediscarded.
Results and discussion
Fatty acid profile of PL
Table 2 shows the proportion of total saturated fatty acids(SFA), monounsaturated fatty acids (MUFA), polyunsatu-rated fatty acids (PUFA), n − 6 and n − 3-fatty acids, and thePI of liposomes made with PL from M and C pigs. Overall,the fatty acid composition of PL isolated from L. dorsi mus-cle tended to reflect that of the diets. C liposomes showed ahigher proportion of PUFA, primarily n − 6 fatty acids, and alower of MUFA and SFA than liposomes from M pigs. Con-sistently, liposomes from C pigs had higher values of PI thanthose from M animals (97.09 vs. 76.55) as a consequence ofa higher PUFA content that could affect their oxidative sus-ceptibility. The influence of the feeding systems on the fattyacid composition of PL of L. dorsi muscle will be profuselydiscussed elsewhere.
Liposome oxidation
The evolution of TBA-RS expressed as nmoles of MDA/mgPL from studied model system is shown in Fig. 1. TBA-RSvalues found in this work are similar to those reported inoxidized liposomes from PL of turkey breast [16] and fromPL of porcine muscle [33]. In the present study, the highestTBA-RS detected were those recorded at day 0 whateverthe model system, and thereafter, there was a slight decreaseuntil the end of the experiment. This evolution of TBA-RSagrees with the tendency observed during the Iberian hamprocess, in which an increase in TBA-RS and a subsequentdecrease and stabilization have been described [2]. Neverthe-less, Meynier et al. [16] and Kansci et al. [33], working withoxidized liposomes, reported an increase in TBA-RS num-bers throughout all the experiment. However, these studies
Springer
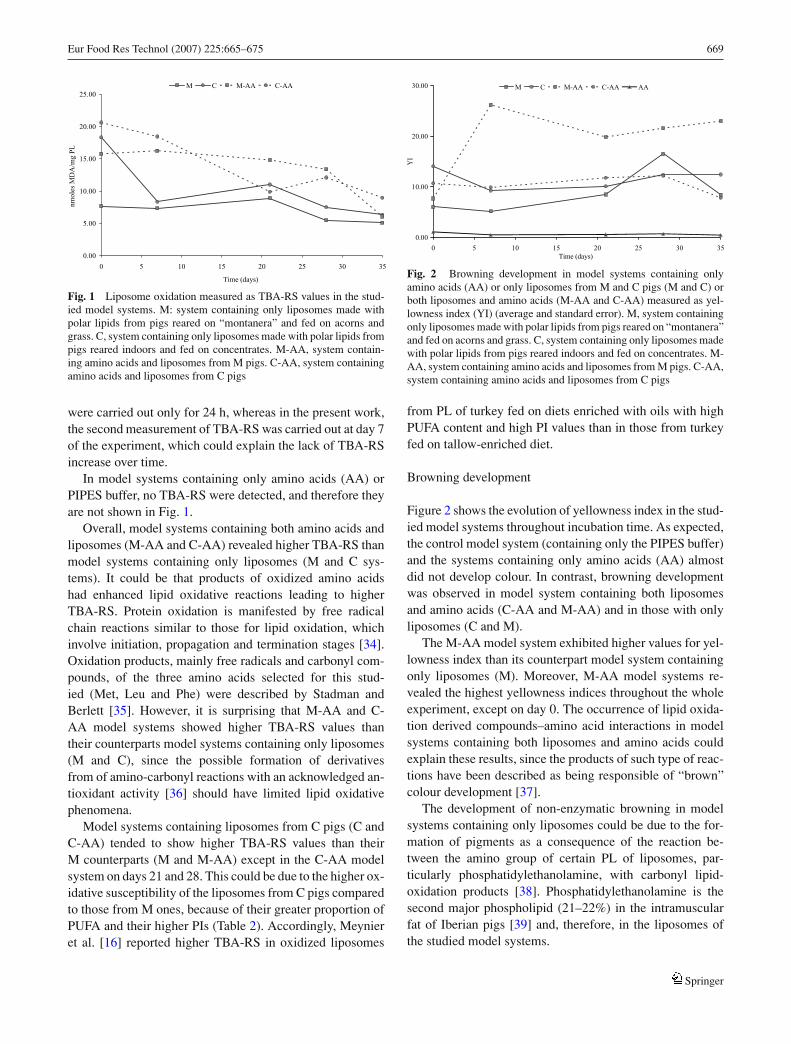
Eur Food Res Technol (2007) 225:665–675 669
0.00
5.00
10.00
15.00
20.00
25.00
0 5 10 15 20 25 30 35
Time (days)
nmol
es M
DA
/mg
PL
M C M-AA C-AA
Fig. 1 Liposome oxidation measured as TBA-RS values in the stud-ied model systems. M: system containing only liposomes made withpolar lipids from pigs reared on “montanera” and fed on acorns andgrass. C, system containing only liposomes made with polar lipids frompigs reared indoors and fed on concentrates. M-AA, system contain-ing amino acids and liposomes from M pigs. C-AA, system containingamino acids and liposomes from C pigs
were carried out only for 24 h, whereas in the present work,the second measurement of TBA-RS was carried out at day 7of the experiment, which could explain the lack of TBA-RSincrease over time.
In model systems containing only amino acids (AA) orPIPES buffer, no TBA-RS were detected, and therefore theyare not shown in Fig. 1.
Overall, model systems containing both amino acids andliposomes (M-AA and C-AA) revealed higher TBA-RS thanmodel systems containing only liposomes (M and C sys-tems). It could be that products of oxidized amino acidshad enhanced lipid oxidative reactions leading to higherTBA-RS. Protein oxidation is manifested by free radicalchain reactions similar to those for lipid oxidation, whichinvolve initiation, propagation and termination stages [34].Oxidation products, mainly free radicals and carbonyl com-pounds, of the three amino acids selected for this stud-ied (Met, Leu and Phe) were described by Stadman andBerlett [35]. However, it is surprising that M-AA and C-AA model systems showed higher TBA-RS values thantheir counterparts model systems containing only liposomes(M and C), since the possible formation of derivativesfrom of amino-carbonyl reactions with an acknowledged an-tioxidant activity [36] should have limited lipid oxidativephenomena.
Model systems containing liposomes from C pigs (C andC-AA) tended to show higher TBA-RS values than theirM counterparts (M and M-AA) except in the C-AA modelsystem on days 21 and 28. This could be due to the higher ox-idative susceptibility of the liposomes from C pigs comparedto those from M ones, because of their greater proportion ofPUFA and their higher PIs (Table 2). Accordingly, Meynieret al. [16] reported higher TBA-RS in oxidized liposomes
0.00
10.00
20.00
30.00
0 5 10 15 20 25 30 35Time (days)
YI
M C M-AA C-AA AA
Fig. 2 Browning development in model systems containing onlyamino acids (AA) or only liposomes from M and C pigs (M and C) orboth liposomes and amino acids (M-AA and C-AA) measured as yel-lowness index (YI) (average and standard error). M, system containingonly liposomes made with polar lipids from pigs reared on “montanera”and fed on acorns and grass. C, system containing only liposomes madewith polar lipids from pigs reared indoors and fed on concentrates. M-AA, system containing amino acids and liposomes from M pigs. C-AA,system containing amino acids and liposomes from C pigs
from PL of turkey fed on diets enriched with oils with highPUFA content and high PI values than in those from turkeyfed on tallow-enriched diet.
Browning development
Figure 2 shows the evolution of yellowness index in the stud-ied model systems throughout incubation time. As expected,the control model system (containing only the PIPES buffer)and the systems containing only amino acids (AA) almostdid not develop colour. In contrast, browning developmentwas observed in model system containing both liposomesand amino acids (C-AA and M-AA) and in those with onlyliposomes (C and M).
The M-AA model system exhibited higher values for yel-lowness index than its counterpart model system containingonly liposomes (M). Moreover, M-AA model systems re-vealed the highest yellowness indices throughout the wholeexperiment, except on day 0. The occurrence of lipid oxida-tion derived compounds–amino acid interactions in modelsystems containing both liposomes and amino acids couldexplain these results, since the products of such type of reac-tions have been described as being responsible of “brown”colour development [37].
The development of non-enzymatic browning in modelsystems containing only liposomes could be due to the for-mation of pigments as a consequence of the reaction be-tween the amino group of certain PL of liposomes, par-ticularly phosphatidylethanolamine, with carbonyl lipid-oxidation products [38]. Phosphatidylethanolamine is thesecond major phospholipid (21–22%) in the intramuscularfat of Iberian pigs [39] and, therefore, in the liposomes ofthe studied model systems.
Springer

670 Eur Food Res Technol (2007) 225:665–675
Volatile compounds
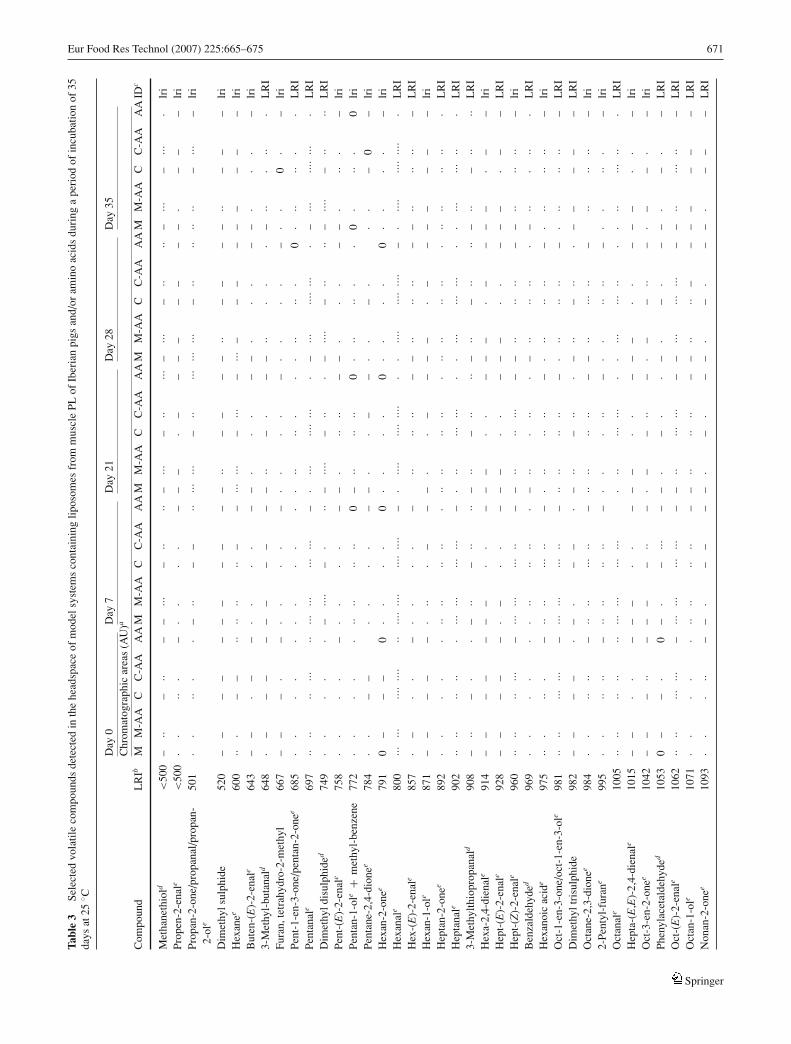
Forty-seven out of 180 volatile compounds detected in theheadspace of M, M-AA, C, C-AA and AA model systems onday 0, 7, 21, 28 and 35 of the experiment were selected andare shown in Table 3 in accordance with their most probableorigin. These compounds were chosen on the basis of theirpresence in most model systems and on their proportionof chromatographic area in the headspace of the studiedmodel systems. Selected compounds included 27 aldehydes,7 ketones, 5 alcohols, 4 hydrocarbons, 4 sulphur compounds,2 furans, 1 acid and 1 thiazole.
Major compounds detected in the headspace of the stud-ied model systems were typical fatty acids oxidation products[9]. Among these, the most abundant lipid-derived volatiles(above 108 AU of chromatographic area at day 7) in M, M-AA, C and C-AA model systems were saturated aldehydes(pentanal, hexanal, heptanal, octanal and nonanal), unsat-urated aldehydes (hept-(Z)-2-enal, oct-(E)-2-enal) and theco-elution of oct-1-en-3-one/oct-1-en-3-ol. Volatiles fromStrecker degradation of the selected amino acids [40–42] (methanethiol, 3-methylbutanal, dimethyl disulphide,dimethyl sulphide, trimethyl trisulphide, methional, ben-zaldehyde and phenylacetaldehyde) were mainly isolatedfrom the headspace of model systems containing both li-posomes and amino acids.
Volatile profiles described from the headspace of the Mand C model systems (Table 3) are similar to those previouslyreported in the headspace of model systems containing lipo-somes from PL from turkey breast [16] and from porcineL. dorsi [33] under similar pro-oxidative conditions. In ad-dition, compounds listed in Table 3 have been previouslyreported as volatile components of Iberian dry-cured prod-ucts, including Iberian hams [6, 23] and other dry-cured meatproducts [43]. Moreover, some of them have been previouslydescribed as odour active compounds in Iberian ham withdifferent odour descriptors [12, 13] such as methanethiol,3-methylbutanal, pent-1-en-3-one, hexanal, hex-(E)-2-enal,heptan-2-one, oct-1-en-3-one/ oct-1-en-3-ol and octanal.
Regarding to lipid-derived volatiles, diet affected volatilecompounds formed in model systems containing liposomes(Table 3). Thus, systems containing liposomes from C pigs,with high levels of PUFA and n − 6 fatty acids (C and C-AA), showed greater chromatographic areas of lipid-derivedvolatiles, particularly of those derived from n − 6 fatty acidssuch as hexanal, deca-(E,E)-2,4-dienal and 2-pentyl-furan,which arise mainly from linoleic acid (C18:2, n − 6) [17, 43].Hexanal was the major volatile whatever the studied modelsystems. Hexanal and deca-(E,E)-2,4-dienal have been as-sociated with the development of warmed over flavour incooked meat [9] and hexanal content has shown good cor-relations with rancid notes in dry-cured ham [1]. Modelsystems containing liposomes from C pigs exhibited higher
chromatographic areas of deca-(E,E)-2,4-dienal than thosecontaining liposomes from M animals, in accordance withstudies in Iberian hams where this volatile compound hasbeen largely reported in hams from pigs fed on concentrates(high PUFA content) than in those from pigs fed on acornsin “montanera” system.
All Strecker-derived volatile compounds were detected inthe headspace of model system containing liposomes and theselected amino acids Leu, Met and Phe. These compounds in-cluded methyl branched aldehydes such as 3-methylbutanaland 3-methylthiopropanal derived from Leu and Met, re-spectively, sulphur compounds such as methanethiol anddimethyl disulphide, derived from Met, and aromatic com-pounds derived from Phe such as benzaldehyde and pheny-lacetaldehyde [4, 40–42]. Dimethyl disulphide is an oxida-tion product of methanethiol and can further react to formdimethyl trisulphide and dimethyl sulphide [43], which havebeen also isolated from the headspace of studied model sys-tems.
The Strecker degradation of amino acids, which is a minorpathway of the Maillard reactions [44], involves the oxidativedeamination and decarboxilation of α-amino acids in thepresence of compounds such as reducing sugars or somelipid-oxidation products [40]. The temperature, pH and pro-oxidative experimental conditions established for the studiedmodel systems for 35 days, similar to those present during theprocessing of dry-cured meat products, seem to encouragethe generation of a great number of lipid-derived carbonylcompounds, particularly volatile aldehydes, that could havereacted with the amino acids from the model systems, leadingto the formation of aforementioned Strecker compounds.Several volatile compounds such as malondialdehyde [45],deca-2,4-dienal [46] and hexanal [47] have been reportedas lipid-oxidation products that can react with the reactivegroups of the amino acids and proteins.
Ventanas et al. [4] have previously hypothesized that re-actions between free amino acids and carbonyls could bean important source of flavour compounds in Iberian dry-cured hams. However, Hinrichsen et al. [7, 8] establishedthat methyl branched aldehydes including 3-methylbutanalcan be generated by secondary metabolism of microorganismand also claimed that Maillard and Strecker reactions onlytake place at higher temperatures than those reached dur-ing dry-cured ham processing. Nevertheless, the detectionof Strecker-derived volatiles in the studied model systemssupports the occurrence of this type of reaction under the es-tablished environmental conditions, similar to those reachedduring dry-cured ham process. Moreover, our results are inagreement with those reported by other authors who havedescribed the occurrence of Strecker reactions at low temper-atures in different model systems [41, 42]. Previous studieshave revealed low microbial counts inside dry-cured hams[48]. Thus, microbial metabolism cannot be considered the
Springer
Eur Food Res Technol (2007) 225:665–675 671
Tabl
e3
Sele
cted
vola
tile
com
poun
dsde
tect
edin
the
head
spac
eof
mod
elsy
stem
sco
ntai
ning
lipos
omes
from
mus
cle
PLof
Iber
ian
pigs
and/
oram
ino
acid
sdu
ring
ape
riod
ofin
cuba
tion
of35
days
at25
◦ C
Day
0D
ay7
Day
21D
ay28
Day
35C
hrom
atog
raph
icar
eas
(AU
)a
Com
poun
dL
RIb
MM
-AA
CC
-AA
AA
MM
-AA
CC
-AA
AA
MM
-AA
CC
-AA
AA
MM
-AA
CC
-AA
AA
MM
-AA
CC
-AA
AA
IDc
Met
hane
thio
ld<
500
–··
–··
––
···–
····
–···
–··
···–
···–
····
–···
–···
·lr
iPr
open
-2-e
nale
<50
0·
···
·–
··
··
––
–·
––
––
––
––
·–
––
lri
Prop
an-2
-one
/pro
pana
l/pro
pan-
2-ol
e50
1·
···
··
–··
––
·····
···–
·····
······
–··
····
··–
···–
lri
Dim
ethy
lsul
phid
e52
0–
––
––
––
––
––
··–
––
–··
––
––
··–
––
lri
Hex
anee
600
···
––
····
····
––
······
–···
–···
––
––
––
––
–lr
iB
uten
-(E
)-2-
enal
e64
3–
–·
––
··
··
––
··
·–
–·
··
––
··
·–
lri
3-M
ethy
l-bu
tana
ld64
8·
––
––
––
––
––
··–
·–
–··
··
·–
···
···
LR
IFu
ran,
tetr
ahyd
ro-2
-met
hyl
667
––
–·
–·
··
·–
··
··
–·
··
·–
··
0·
–lr
iPe
nt-1
-en-
3-on
e/pe
ntan
-2-o
nee
685
··
··
··
··
··
···
···
··
····
·0
···
···
·L
RI
Pent
anal
e69
7··
····
·····
······
······
–·
·······
····
–···
·······
·–
·······
····
LR
ID
imet
hyld
isul
phid
ed74
9·
··
··
–····
–·
··–
····–
···
–····
–··
··–
····–
····
LR
IPe
nt-(
E)-
2-en
ale
758
··
··
–·
··
·–
–·
····
––
··
·–
–·
···
–lr
iPe
ntan
-1-o
le+
met
hyl-
benz
ene
772
··
··
···
····
··0
–··
····
0·
····
··
0·
···
0lr
iPe
ntan
e-2,
4-di
onee
784
··
––
··
··
––
··
––
··
–·
··
–0
–lr
iH
exan
-2-o
nee
791
0–
––
0·
··
·0
··
··
0·
··
·0
··
··
–lr
iH
exan
ale
800
······
········
······
········
····–
·····
········
··
········
····–
·····
········
·L
RI
Hex
-(E
)-2-
enal
e85
7·
–·
·–
··
··
–··
····
––
····
··–
–··
····
–L
RI
Hex
an-1
-ole
871
––
––
–·
···
––
–·
·–
––
–·
––
––
––
–lr
iH
epta
n-2-
onee
892
··
··
···
····
···
····
····
···
····
···
····
····
·L
RI
Hep
tana
le90
2··
····
···
······
······
–·
·····
····
····
······
··
······
···
LR
I3-
Met
hylth
iopr
opan
ald
908
–··
–·
·–
··–
····
–··
–··
··–
··–
····
–··
–··
··L
RI
Hex
a-2,
4-di
enal
e91
4–
––
––
––
··
––
–·
·–
––
·–
––
–·
––
lri
Hep
t-(E
)-2-
enal
e92
8–
––
––
·–
··
––
–·
·–
––
··
––
–·
––
LR
IH
ept-
(Z)-
2-en
ale
960
····
·····
–···
······
··–
–··
·····
––
····
··–
–··
····
–lr
iB
enza
ldeh
yded
969
··
··
··
····
···
–··
···
·–
····
···
–··
···
·L
RI
Hex
anoi
cac
ide
975
···
···
–··
·····
··–
···
····
–·
····
··–
···
····
–lr
iO
ct-1
-en-
3-on
e/oc
t-1-
en-3
-ole
981
····
······
–···
······
··–
····
····
–·
····
··–
···
····
–L
RI
Dim
ethy
ltri
sulp
hide
982
––
––
·–
·–
–·
–··
–··
·–
··–
···
––
––
–L
RI
Oct
ane-
2,3-
dion
ee98
4·
···
··–
····
·····
–··
····
··–
–··
·····
–··
····
–lr
i2-
Pent
yl-f
uran
e99
5·
···
··–
····
····
–·
··
··–
··
····
–·
···
·–
lri
Oct
anal
e10
05··
····
····
······
······
···
······
··
······
···
···
·····
·L
RI
Hep
ta-(
E,E
)-2,
4-di
enal
e10
15–
–·
·–
––
··
––
–·
·–
––
··
––
–·
·–
lri
Oct
-3-e
n-2-
onee
1042
––
–··
––
––
··–
·–
–··
–·
––
··–
·–
–·
–lr
iPh
enyl
acet
alde
hyde
d10
530
––
·0
–·
–···
––
·–
··
–·
–·
––
·–
·–
LR
IO
ct-(
E)-
2-en
ale
1062
····
······
–···
······
···–
–··
······
––
······
···–
–··
·····
–L
RI
Oct
an-1
-ole
1071
··
··
···
····
··–
–··
····
––
····
––
––
––
–L
RI
Non
an-2
-one
e10
93·
··
··–
–·
––
––
·–
·–
–·
–·
––
·–
––
LR
I
Springer

672 Eur Food Res Technol (2007) 225:665–675
Tabl
e3
Con
tinue
d.
Day
0D
ay7
Day
21D
ay28
Day
35C
hrom
atog
raph
icar
eas
(AU
)a
Com
poun
dL
RIb
MM
-AA
CC
-AA
AA
MM
-AA
CC
-AA
AA
MM
-AA
CC
-AA
AA
MM
-AA
CC
-AA
AA
MM
-AA
CC
-AA
AA
IDc
Non
anal
e11
06··
····
·····
······
······
···
····
····
···
····
––
····
···
lri
Oct
a-(E
,E)-
2,4-
dien
ale
1115
··
··
–·
··
·–
–·
···
––
··
·–
–·
··
–lr
iN
on-(
E)-
2-en
ale
1163
··
····
-··
····
··–
–··
····
––
····
·–
–·
··
–lr
iN
ona-
2,4-
dien
ale
1198
––
··
–··
––
·–
––
··
––
··
·–
––
––
–lr
iD
ecan
ale
1207
··
··
···
···
··–
··
··
··
···
·–
–·
···
·lr
iN
ona-
(E,E
)-2,
4-di
enal
e12
19·
···
··–
···
··
––
··
··–
–·
···
––
––
·–
lri
Ben
zoth
iazo
le12
50–
––
–·
––
–·
–·
··
··
––
––
––
––
––
lri
Dec
-(E
)-2-
enal
e12
67··
····
···
····
····
––
··
·–
–·
··
––
··
·–
lri
Dec
a-(E
,E)-
2,4-
dien
ale
1325
··
····
–·
··
·–
–0
–·
––
·–
––
––
––
–lr
iU
ndec
-2-e
nale
1370
····
····
···
···
––
––
––
––
––
––
––
––
–lr
i
M,s
yste
mco
ntai
ning
only
lipos
omes
mad
ew
ithpo
lar
lipid
sfr
ompi
gsfe
don
acor
nsan
dgr
ass.
C,s
yste
mco
ntai
ning
only
lipos
omes
mad
ew
ithpo
lar
lipid
sfr
ompi
gsfe
dco
ncen
trat
es.M
-AA
,sy
stem
cont
aini
ngam
ino
acid
san
dlip
osom
esfr
omM
pigs
.C-A
A,s
yste
mco
ntai
ning
amin
oac
ids
and
lipos
omes
from
Cpi
gs.
aC
hrom
atog
raph
icar
eas
(AU
).N
otde
tect
ed(–
),ab
ove
105
(0),
abov
e106
(·),a
bove
107
(··),
abov
e108
(···),
abov
e10
9(··
··).
bL
inea
rre
tent
ion
inde
x.c M
etho
dof
iden
tifica
tion.
LR
I,m
ass
spec
trum
and
LR
Iagr
eew
ithth
ose
ofst
anda
rdco
mpo
unds
;lri
,mas
ssp
ectr
aag
ree
with
thos
ere
port
edin
the
Wile
yli
brar
yan
dL
RIa
gree
sw
ithth
ose
repo
rted
inth
elit
erat
ure
(29)
and
inth
eN
IST
data
base
(30)
.dV
olat
iles
incl
uded
inth
eSt
reck
er-d
eriv
edvo
latil
eco
mpo
unds
grou
pin
Fig.
3.e V
olat
iles
incl
uded
inth
elip
id-d
eriv
edvo
latil
eco
mpo
unds
grou
pin
Fig.
3.
Springer
Eur Food Res Technol (2007) 225:665–675 673
only possible source of generation branched aldehydes indry-cured products.
Numerous studies have been aimed to study Strecker-derived volatiles formation in model systems containingamino acids in the presence of ribose [18, 19], glucose [44]epoxy derivates compounds [38, 42] and other carbonyl com-pounds such us diacetyl o pentan-2,3-dione [41]. In otherstudies, PL [21] or lipidic systems such as cocoa butter[40] were included into amino acid–sugar model systems.However, most of these studies were performed at high tem-peratures to simulate the conditions during meat cooking.In some studies, experimental temperatures were low, 25 ◦C[41] or 37 ◦C [38, 42]. Nevertheless, in these papers, the gen-eration of Strecker-derived compounds was achieved usingsynthetic carbonyl compounds and was performed for a shortperiod of incubation (overnight) [41]. To our knowledge, thisis the first study in which the generation of Strecker-derivedvolatile compounds in model systems mimicking meat prod-ucts containing amino acids and lipids without any reducingsugar is shown.
Overall, M-AA systems exhibited greater chromato-graphic area than C-AA ones for methanethiol, dimethyldisulphide and 3-methyl-butanal (Table 3). The majorStrecker-derived volatile compound in M-AA model sys-tems was dimethyl disulphide, whereas in C-AA systems themajor compound was phenylacetaldehyde.
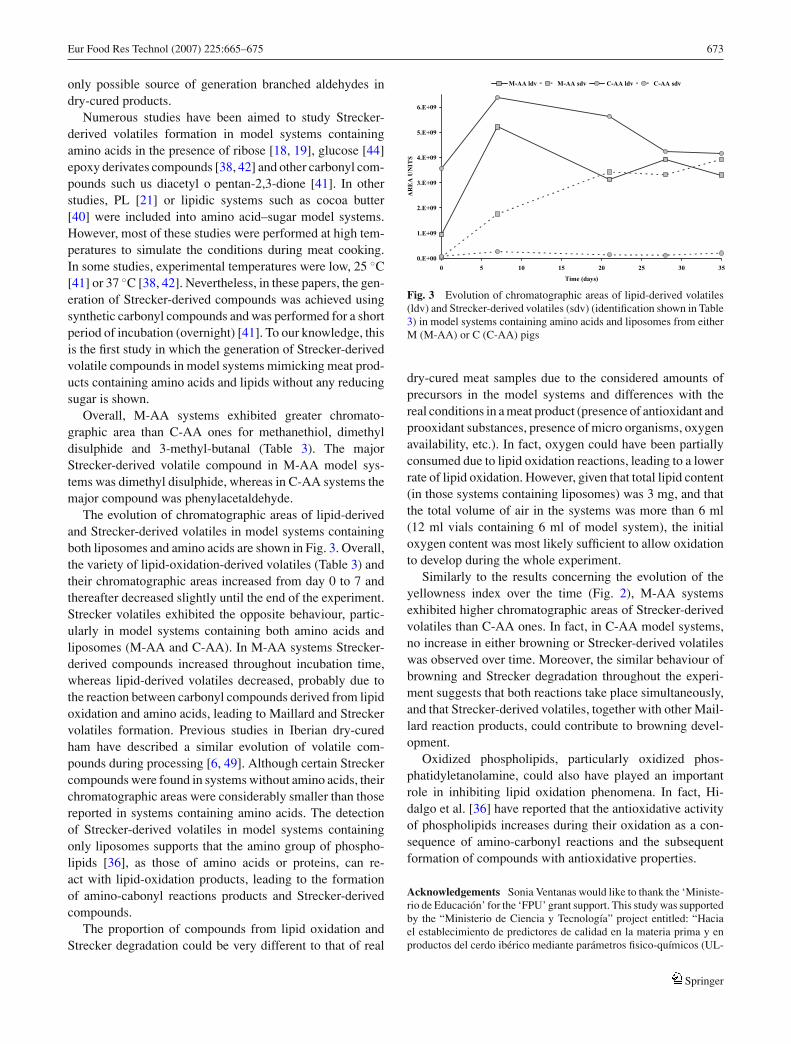
The evolution of chromatographic areas of lipid-derivedand Strecker-derived volatiles in model systems containingboth liposomes and amino acids are shown in Fig. 3. Overall,the variety of lipid-oxidation-derived volatiles (Table 3) andtheir chromatographic areas increased from day 0 to 7 andthereafter decreased slightly until the end of the experiment.Strecker volatiles exhibited the opposite behaviour, partic-ularly in model systems containing both amino acids andliposomes (M-AA and C-AA). In M-AA systems Strecker-derived compounds increased throughout incubation time,whereas lipid-derived volatiles decreased, probably due tothe reaction between carbonyl compounds derived from lipidoxidation and amino acids, leading to Maillard and Streckervolatiles formation. Previous studies in Iberian dry-curedham have described a similar evolution of volatile com-pounds during processing [6, 49]. Although certain Streckercompounds were found in systems without amino acids, theirchromatographic areas were considerably smaller than thosereported in systems containing amino acids. The detectionof Strecker-derived volatiles in model systems containingonly liposomes supports that the amino group of phospho-lipids [36], as those of amino acids or proteins, can re-act with lipid-oxidation products, leading to the formationof amino-cabonyl reactions products and Strecker-derivedcompounds.
The proportion of compounds from lipid oxidation andStrecker degradation could be very different to that of real
0.E+00
1.E+09
2.E+09
3.E+09
4.E+09
5.E+09
6.E+09
0 5 10 15 20 25 30 35
Time (days)
AR
EA
UN
ITS
M-AA ldv M-AA sdv C-AA ldv C-AA sdv
Fig. 3 Evolution of chromatographic areas of lipid-derived volatiles(ldv) and Strecker-derived volatiles (sdv) (identification shown in Table3) in model systems containing amino acids and liposomes from eitherM (M-AA) or C (C-AA) pigs
dry-cured meat samples due to the considered amounts ofprecursors in the model systems and differences with thereal conditions in a meat product (presence of antioxidant andprooxidant substances, presence of micro organisms, oxygenavailability, etc.). In fact, oxygen could have been partiallyconsumed due to lipid oxidation reactions, leading to a lowerrate of lipid oxidation. However, given that total lipid content(in those systems containing liposomes) was 3 mg, and thatthe total volume of air in the systems was more than 6 ml(12 ml vials containing 6 ml of model system), the initialoxygen content was most likely sufficient to allow oxidationto develop during the whole experiment.
Similarly to the results concerning the evolution of theyellowness index over the time (Fig. 2), M-AA systemsexhibited higher chromatographic areas of Strecker-derivedvolatiles than C-AA ones. In fact, in C-AA model systems,no increase in either browning or Strecker-derived volatileswas observed over time. Moreover, the similar behaviour ofbrowning and Strecker degradation throughout the experi-ment suggests that both reactions take place simultaneously,and that Strecker-derived volatiles, together with other Mail-lard reaction products, could contribute to browning devel-opment.
Oxidized phospholipids, particularly oxidized phos-phatidyletanolamine, could also have played an importantrole in inhibiting lipid oxidation phenomena. In fact, Hi-dalgo et al. [36] have reported that the antioxidative activityof phospholipids increases during their oxidation as a con-sequence of amino-carbonyl reactions and the subsequentformation of compounds with antioxidative properties.
Acknowledgements Sonia Ventanas would like to thank the ‘Ministe-rio de Educacion’ for the ‘FPU’ grant support. This study was supportedby the “Ministerio de Ciencia y Tecnologıa” project entitled: “Haciael establecimiento de predictores de calidad en la materia prima y enproductos del cerdo iberico mediante parametros fisico-quımicos (UL-
Springer

674 Eur Food Res Technol (2007) 225:665–675
TRAFAT)” (Project AGL 2001-6932-01). Authors are grateful to Dr.Francisco J. Hidalgo (Instituto de la Grasa, Sevilla, SPAIN) for hisscientific advices during the development of this study
References
1. Ruiz J, Muriel E, Ventanas J (2002) The flavour of Iberian ham.In: Toldra F (ed) Research advances in the quality of meat andmeat products. Research Singpost, Trivandrum, Kerala, India, pp290–309
2. Andres AI, Cava R, Ventanas J, Ruiz J (2004) Lipid oxidativechanges throughout the ripening of dry-cured Iberian hams withdifferent salt contents and processing conditions. Food Chem84:375–381
3. Antequera T, Lopez-Bote C, Cordoba JJ, Garcıa C, Asensio MA,Ventanas J (1994) Lipid oxidative changes in the processing ofIberian pig hams. Food Chem 45:105–110
4. Ventanas J, Cordoba JJ, Antequera T, Garcıa C, Lopez-Bote C,Asensio MA (1992) Hydrolysis and maillard reactions duringripening of Iberian ham. J Food Sci 57:803–815
5. Cordoba JJ, Antequera T, Garcıa C, Ventanas J, Lopez-Bote C,Asensio MA (1994) Evolution of free amino acids and aminesduring ripening of Iberian cured ham. J Agric Food Chem 42:2296–2301
6. Ruiz J, Ventanas J, Cava R, Andres AI, Garcıa C (1999) Iberianham as affected by the length of the curing process. Meat Sci52:19–27
7. Hinrichsen LL, Andersen HJ (1994) Volatile compounds and chem-ical changes in cured pork: role of three halotolerant bacteria. JAgric Food Chem 42:1537–1542
8. Hinrichsen LL, Pedersen SB (1995) Relationship among flavour,volatile compounds, chemical changes and microflora in Italian-type dry-cured hams during processing. J Agric Food Chem43:2932–2940
9. Mottram DS (1998) Flavour formation in meat and meat products:a review. Food Chem 62:415–424
10. Pikul J, Leszczynski DE, Kummerow FA (1984) Relative role ofphospholipids, triacylglycerols and cholesterol esters on malon-aldehyde formation in fat extracted from chicken meat. J Food Sci49:704–708
11. Boylston TD, Morgan SA, Johnson KA, Wright RW, Busboom JR,Reeves JJ (1996) Volatile lipid oxidation products of Wagyu anddomestic breeds of beef. J Agric Food Chem 44:1091–1095
12. Carrapiso AI, Ventanas J, Garcıa C (2002) Characterization of themost odor-active compounds of Iberian ham headspace. J AgricFood Chem 50:1996–2000
13. Carrapiso AI, Jurado A, Timon ML, Garcıa C (2002) Odor-activecompounds of Iberian hams with different aroma characteristics. JAgric Food Chem 50:6453–6458
14. Cava R, Ruiz J, Lopez-Bote C, Martın L, Garcıa C, Ventanas J,Antequera T (1997) Influence of finishing diet on fatty acid profilesof intramuscular lipids, triglycerides and phospholipids in musclesof the Iberian pig. Meat Sci 2:263–270
15. Cava R, Ruiz J, Ventanas J, Antequera T (1999) Oxidative andlipolityc changes during ripening of Iberian hams as affected byfeeding regime: extensive feeding and alpha-tocopheryl acetatesupplementation. Meat Sci 52:165–172
16. Meynier A, Genot C, Gandemer G (1999) Oxidation of musclephospholipids in relation to their fatty acid composition with em-phasis on volatile compounds. J Sci Food Agric 79:797–804
17. Elmore JS, Mottram DS, Enser M, Wood JD (1999) Effect ofpolyunsaturated fatty acid composition of beef muscle on the pro-file of aroma volatiles. J Agric Food Chem 47:1619–1625
18. Meynier A, Mottram SD (1995) The effect of pH on the formationof volatile compounds in meat-related model systems. Food Chem52:361–366
19. Cerny C, Davidek T (2003) Formation of aroma compounds fromribose and cysteine during the maillard reaction. J Agric FoodChem 51:2714–2721
20. Elmore JS, Campo MM, Enser M, Mottram DS (2002) Effectof lipid composition on meat-like model systems containing cys-teine, ribose and polyunsaturated fatty acids. J Agric Food Chem50:1126–1132
21. Mottram DS, Whitfield FB (1995) Volatile compounds from thereaction of cysteine, ribose, and phospholipids in low-moisturesystems. J Agric Food Chem 43:984–988
22. Bligh EG, Dyer WJ (1959) A rapid method of total lipid extractionand purification. Can J Biochem Phys 37:911–917
23. Muriel E, Ruiz J, Ventanas J, Petron MJ, Antequera T (2004) Meatquality characteristics in different lines of Iberian pigs. Meat Sci67:299–307
24. Barlett GR (1959) Phosphorus assay in column chromatography. JBiol Chem 234:466–468
25. Wangen RM, Marion WW, Hotchkiss DK (1971) Influence of ageon total lipids and phospholipids of turkey muscle. J Food Sci36:560–562
26. Sandler SR, Karo W (1992) Source book of advances organiclaboratory preparations. Academic Press, San Diego
27. Buege JA, Aust SD (1978) Microsomal lipid peroxidation. In:Parker L, Fleischer S (eds) Methods in enzymology. AcademyPress, New York, pp 302–312
28. Hunter RS (1973) The measurement of appearance. Hunter Asso-ciates Laboratory, Fairfax, VA
29. CIE (Commission International de l’Eclariage) (1978) Recommen-dations on Uniform Color Spaces, Color-Differences Equations,Psychometric Color Terms; CIE Publications 15 (Supplement 2);Bureau Central de la Commission Interantionale de l’Eclariage,Paris
30. Francis FJ, Clydeslade FH (1975) Food colorimetry: theory andapplications. AVI Publishing, Westport, CT
31. Kondjoyan N, Berdague JL (1996). In Compilation of RelativeRetention Indices of the Analysis of Aromatic Compounds. Labo-ratoire Flaveur, station de rescherches sur la viande, INRA, Theix,France
32. NIST (National Institute of Standards and Technology) database.http://webbook.nist.gov
33. Kansci G, Genot C, Meynier A, Gandemer G (1997) The antiox-idant activity of carnosine and its consequences on the volatileprofile of liposomes during iron/ascorbate induced phospholipidsoxidation. Food Chem 60:165–175
34. Schaich KM (1980) Free radical initiation in proteins and aminoacids by ionizing and ultraviolet radiations and lipid oxidation. III:Free radical transfer from oxidizing lipids. CRC Crit Rev Food SciNutr 13:189–244
35. Stadman ER, Berlett BS (1997) Reactive oxygen-mediated pro-tein oxidation in aging and disease. Chem Res Toxicol 10:484–494
36. Hidalgo FJ, Nogales F, Zamora R (2005) Changes produced in theantioxidative activity of phospholipids as a consequence of theiroxidation. J Agric Food Chem 53:659–662
37. Mastrocola D, Munari M, Cioroi M, Lerici CR (2000) In-teraction between Maillard reaction products and lipid oxida-tion in starch-based model systems. J Sci Food Agric 80:684–690
38. Zamora R, Nogales F, Hidalgo FJ (2005) Phospholipid ox-idation and nonenzymatic browning development in phos-phatidylethanolamine/ribose/lysine model systems. Eur Food ResTechnol 220:459–465
Springer

Eur Food Res Technol (2007) 225:665–675 675
39. Tejeda JL (1999) Influence of breed and diet on the intramuscularlipid fraction in Iberian pigs (Estudio de la influencia de la razay de la alimentacion sobre la fraccion lipıdica intramuscular delcerdo iberico). Doctoral Thesis, Universidad de Extremadura
40. Arnoldi A, Arnoldi C, Baldi O, Griffini A (1987) Strecker degrada-tion of leucine and valine in a lipidic model system. J Agric FoodChem 35:1035–1038
41. Pripis-Nicolau L, de Revel G, Bertrand A, Maujean A (2000)Formation of flavour components by the reaction of amino acidand carbonyl compounds in mild conditions. J Agric Food Chem48:3761–3766
42. Hidalgo FJ, Zamora R (2004) Strecker-type degradation by thelipid oxidation products 4,5-epoxy-2-alkenals. J Agric Food Chem52:7126–7131
43. Flores M, Spanier AM, Toldra F (1998) Flavour analysis of dry-cured ham. In: Shahidi F (ed) Flavour of meat and meat productsand seafoods, 2nd edn. Blackie Academic and Professional, Lon-don, UK, pp 320–341
44. Cremer DR, Eichner K (2000) The reaction kinetics for the forma-tion of Strecker aldehydes in low moisture model systems and inplant powders. Food Chem 71:37–43
45. Itakura K, Uchida K, Osawa T (1996) A novel fluorescentmalondialdehyde-lysine adduct. Chem Phys Lipids 84:75–79
46. Leake L, Karel M (1985) Nature of fluorescent compounds gen-erated by exposure of protein to oxidizing lipids. J Food Biochem9:117–136
47. Kaneko S, Okitani A, Hayase F, Kato H (1991) Identificationof an intermediate product and formation mechanisms of cross-linking compounds from Nα-acetyltryptophan and hexanal. AgricBiol Chem 55:723–730
48. Molina I, Toldra F (1992) Detection of proteolityc activity in mi-croorganism isolated from dry-cured ham. J Food Sci 57:1308–1310
49. Jurado A (2005) Evolution of taste and aroma compounds dur-ing the ripening of Iberian dry cured hams of different qualities(Evolucion de compuestos relacionados con el sabor y el aromadel jamon iberico de distintas calidades durante su procesado).Doctoral Thesis, Universidad de Extremadura, Spain
Springer













![Fusion of liposomes containing a novel cationic lipid, N-[2,3-(dioleyloxy)propyl]-N,N,N-trimethylammonium: induction by multivalent anions and asymmetric fusion with acidic phospholipid](https://static.fdokumen.com/doc/165x107/63139fc43ed465f0570ac29a/fusion-of-liposomes-containing-a-novel-cationic-lipid-n-23-dioleyloxypropyl-nnn-trimethylammonium.jpg)






