
Acyl-Phosphates Initiate Membrane Phospholipid Synthesis in Gram-Positive Pathogens
8
Molecular Cell 23, 765–772, September 1, 2006 ª2006 Elsevier Inc. DOI 10.1016/j.molcel.2006.06.030 Short Article Acyl-Phosphates Initiate Membrane Phospholipid Synthesis in Gram-Positive Pathogens Ying-Jie Lu, 1 Yong-Mei Zhang, 1 Kimberly D. Grimes, 2 Jianjun Qi, 2 Richard E. Lee, 2 and Charles O. Rock 1, * 1 Department of Infectious Diseases St. Jude Children’s Research Hospital 332 North Lauderdale Memphis, Tennessee 38105 2 Department of Pharmaceutical Sciences University of Tennessee Health Science Center 847 Monroe Avenue Memphis, Tennessee 38163 Summary It is not known how Gram-positive bacterial pathogens carry out glycerol-3-phosphate (G3P) acylation, which is the first step in the formation of phosphatidic acid, the key intermediate in membrane phospholipid syn- thesis. In Escherichia coli, acylation of the 1-position of G3P is carried out by PlsB; however, the majority of bacteria lack a plsB gene and in others it is not es- sential. We describe a two-step pathway that utilizes a new fatty acid intermediate for the initiation of phos- pholipid formation. First, PlsX produces a unique acti- vated fatty acid by catalyzing the synthesis of fatty acyl-phosphate from acyl-acyl carrier protein, and then PlsY transfers the fatty acid from acyl-phosphate to the 1-position of G3P. The PlsX/Y pathway defines the most widely distributed pathway for the initiation of phos- pholipid formation in bacteria and represents a new tar- get for the development of antibacterial therapeutics. Introduction Phosphatidic acid (PtdOH) is a universal intermediate in the biosynthesis of membrane phospholipids in eubac- teria. In the Escherichia coli model, PtdOH formation is initiated by the acylation of sn-glycerol-3-phosphate (G3P) by PlsB, the G3P acyltransferase (Cronan and Rock, 1996). PlsB of E. coli has been extensively charac- terized and utilizes either acyl-acyl carrier protein (ACP) or acyl-CoA thioesters to acylate the 1-position of G3P (Green et al., 1981; Lightner et al., 1980). PlsB is respon- sible for the selection of fatty acids incorporated into membrane phospholipids and is a key regulatory point in the pathway (Cronan and Rock, 1996; Heath et al., 1994; Rock et al., 1981b). However, most bacteria, in- cluding important human Gram-positive pathogens such as Streptococcus pneumoniae and Staphylococcus aureus, lack a plsB gene, and the pathway for the acyla- tion of G3P is unknown. In some Gram-negative bacte- ria, such as Pseudomonas aeruginosa, the plsB gene is not essential (Jacobs et al., 2003), demonstrating the presence of an alternate pathway to acyl-G3P. The plsB gene was identified in 1974 with the isolation of E. coli strain BB26, a G3P auxotroph that has a mis- sense mutation that results in an elevated K M for G3P (Bell, 1974, 1975; Heath and Rock, 1999). Subsequently, it was discovered that this growth phenotype required a second mutation that inactivated another gene, plsX (Larson et al., 1984). These data indicated that plsX either encoded a second acyltransferase or was in- volved in G3P metabolism. However, only the K M -defec- tive acyltransferase activity was detected in membranes prepared from cells possessing a wild-type plsX and a mutant plsB gene, suggesting that PlsX is not an alter- nate G3P acyltransferase. A second acyltransferase, PlsC (Coleman, 1992), is universally expressed in bacte- ria and completes the synthesis of PtdOH by transfer- ring a fatty acid to the 2-position of acyl-G3P via acyl- ACP or acyl-CoA. This work describes the discovery of the missing acyltransferase system that utilizes an acyl-phosphate (acyl-PO 4 ) intermediate derived from acyl-ACP by PlsX and transferred to G3P by PlsY. The PlsX/Y system is the most widely distributed pathway for membrane phospholipid formation in bacteria. Results and Discussion We selected S. pneumoniae as a model Gram-positive organism to investigate PtdOH formation. This bacte- rium has an EcPlsX homolog (SPR0037) that was cloned, expressed in E. coli, and purified as a dimer (Fig- ure 1A). SpPlsX is 33.2% identical and 55.0% similar to EcPlsX. The bioinformatic analysis of the PlsX pri- mary sequences shows that the PlsX group of proteins (pfam02504/COG0416) is related to the protein family that catalyzes the transfer of either an acetyl or butyryl group from CoA to orthophosphate (pfam01515/ COG0280). Likewise, the structures of the PlsX protein from Bacillus subtilis (accession number 1VI1; Badger et al., 2005) and the B. subtilis phosphotransacetylase (accession number 1TD9; Xu et al., 2005) are related with a Z score of 24 and a root mean square deviation in the Ca chains of 3.0–3.1 A ˚ based on a comparison using DaliLite (Holm and Park, 2000). Phosphotransace- tylase catalyzes the reversible interconversion of acetyl- phosphate + CoA with acetyl-CoA + phosphate; there- fore, biochemical assays were developed to determine if purified SpPlsX had a similar activity. SpPlsX cata- lyzed the conversion of [ 14 C]16:0-ACP to a 14 C-labeled lipid with an R f of 0.21 (Figure 1B). This conversion re- quired the presence of orthophosphate (Figure 1B), and [ 32 P]orthophosphate was also incorporated into the product (Figure 1C). PlsX did not have intrinsic G3P acyl- transferase or fatty acid kinase activity (Figure 1B). PlsX was specific for acyl-ACP, and acyl-CoA and free fatty acid were not substrates (Figure 1C). Negative-ion mass spectroscopy showed the PlsX reaction product had a molecular weight of 336 (Figure 1D). These data showed that SpPlsX catalyzed the formation of palmi- toyl-phosphate (16:0-PO 4 ) from 16:0-ACP and ortho- phosphate. The activity of SpPlsX was modestly enhanced by Mg 2+ but did not require a divalent cation (Figure 1B). The apparent K M for acyl-ACP was 300 mM, and the K M for orthophosphate was 140 mM(Figures 1F and 1G). Traces of labeled free fatty acid were detected in the *Correspondence: [email protected]
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Acyl-Phosphates Initiate Membrane Phospholipid Synthesis in Gram-Positive Pathogens

Molecular Cell 23, 765–772, September 1, 2006 ª2006 Elsevier Inc. DOI 10.1016/j.molcel.2006.06.030
Short ArticleAcyl-Phosphates Initiate MembranePhospholipid Synthesisin Gram-Positive Pathogens
Ying-Jie Lu,1 Yong-Mei Zhang,1 Kimberly D. Grimes,2
Jianjun Qi,2 Richard E. Lee,2 and Charles O. Rock1,*1Department of Infectious DiseasesSt. Jude Children’s Research Hospital332 North LauderdaleMemphis, Tennessee 381052Department of Pharmaceutical SciencesUniversity of Tennessee Health Science Center847 Monroe AvenueMemphis, Tennessee 38163
Summary
It is not known how Gram-positive bacterial pathogenscarry out glycerol-3-phosphate (G3P) acylation, which
is the first step in the formation of phosphatidic acid,
the key intermediate in membrane phospholipid syn-thesis. In Escherichia coli, acylation of the 1-position
of G3P is carried out by PlsB; however, the majorityof bacteria lack a plsB gene and in others it is not es-
sential. We describe a two-step pathway that utilizesa new fatty acid intermediate for the initiation of phos-
pholipid formation. First, PlsX produces a unique acti-vated fatty acid by catalyzing the synthesis of fatty
acyl-phosphate from acyl-acyl carrier protein, and thenPlsY transfers the fatty acid from acyl-phosphate to the
1-positionofG3P. The PlsX/Y pathwaydefines the mostwidely distributed pathway for the initiation of phos-
pholipid formation in bacteria and represents a new tar-get for the development of antibacterial therapeutics.
Introduction
Phosphatidic acid (PtdOH) is a universal intermediate inthe biosynthesis of membrane phospholipids in eubac-teria. In the Escherichia coli model, PtdOH formation isinitiated by the acylation of sn-glycerol-3-phosphate(G3P) by PlsB, the G3P acyltransferase (Cronan andRock, 1996). PlsB of E. coli has been extensively charac-terized and utilizes either acyl-acyl carrier protein (ACP)or acyl-CoA thioesters to acylate the 1-position of G3P(Green et al., 1981; Lightner et al., 1980). PlsB is respon-sible for the selection of fatty acids incorporated intomembrane phospholipids and is a key regulatory pointin the pathway (Cronan and Rock, 1996; Heath et al.,1994; Rock et al., 1981b). However, most bacteria, in-cluding important human Gram-positive pathogens suchas Streptococcus pneumoniae and Staphylococcusaureus, lack a plsB gene, and the pathway for the acyla-tion of G3P is unknown. In some Gram-negative bacte-ria, such as Pseudomonas aeruginosa, the plsB gene isnot essential (Jacobs et al., 2003), demonstrating thepresence of an alternate pathway to acyl-G3P.
The plsB gene was identified in 1974 with the isolationof E. coli strain BB26, a G3P auxotroph that has a mis-sense mutation that results in an elevated KM for G3P(Bell, 1974, 1975; Heath and Rock, 1999). Subsequently,
*Correspondence: [email protected]
it was discovered that this growth phenotype requireda second mutation that inactivated another gene, plsX(Larson et al., 1984). These data indicated that plsXeither encoded a second acyltransferase or was in-volved in G3P metabolism. However, only the KM-defec-tive acyltransferase activity was detected in membranesprepared from cells possessing a wild-type plsX anda mutant plsB gene, suggesting that PlsX is not an alter-nate G3P acyltransferase. A second acyltransferase,PlsC (Coleman, 1992), is universally expressed in bacte-ria and completes the synthesis of PtdOH by transfer-ring a fatty acid to the 2-position of acyl-G3P via acyl-ACP or acyl-CoA. This work describes the discoveryof the missing acyltransferase system that utilizes anacyl-phosphate (acyl-PO4) intermediate derived fromacyl-ACP by PlsX and transferred to G3P by PlsY. ThePlsX/Y system is the most widely distributed pathwayfor membrane phospholipid formation in bacteria.
Results and Discussion
We selected S. pneumoniae as a model Gram-positiveorganism to investigate PtdOH formation. This bacte-rium has an EcPlsX homolog (SPR0037) that wascloned, expressed in E. coli, and purified as a dimer (Fig-ure 1A). SpPlsX is 33.2% identical and 55.0% similarto EcPlsX. The bioinformatic analysis of the PlsX pri-mary sequences shows that the PlsX group of proteins(pfam02504/COG0416) is related to the protein familythat catalyzes the transfer of either an acetyl or butyrylgroup from CoA to orthophosphate (pfam01515/COG0280). Likewise, the structures of the PlsX proteinfrom Bacillus subtilis (accession number 1VI1; Badgeret al., 2005) and the B. subtilis phosphotransacetylase(accession number 1TD9; Xu et al., 2005) are relatedwith a Z score of 24 and a root mean square deviationin the Ca chains of 3.0–3.1 A based on a comparisonusing DaliLite (Holm and Park, 2000). Phosphotransace-tylase catalyzes the reversible interconversion of acetyl-phosphate + CoA with acetyl-CoA + phosphate; there-fore, biochemical assays were developed to determineif purified SpPlsX had a similar activity. SpPlsX cata-lyzed the conversion of [14C]16:0-ACP to a 14C-labeledlipid with an Rf of 0.21 (Figure 1B). This conversion re-quired the presence of orthophosphate (Figure 1B),and [32P]orthophosphate was also incorporated into theproduct (Figure 1C). PlsX did not have intrinsic G3P acyl-transferase or fatty acid kinase activity (Figure 1B). PlsXwas specific for acyl-ACP, and acyl-CoA and free fattyacid were not substrates (Figure 1C). Negative-ionmass spectroscopy showed the PlsX reaction producthad a molecular weight of 336 (Figure 1D). These datashowed that SpPlsX catalyzed the formation of palmi-toyl-phosphate (16:0-PO4) from 16:0-ACP and ortho-phosphate.
The activity of SpPlsX was modestly enhanced byMg2+ but did not require a divalent cation (Figure 1B).The apparent KM for acyl-ACP was 300 mM, and the KM
for orthophosphate was 140 mM (Figures 1F and 1G).Traces of labeled free fatty acid were detected in the

Molecular Cell766
Figure 1. PlsX Is a Phosphate:acyl-ACP Acyltransferase
(A) Purified SpPlsX was a soluble protein with a Stoke’s radius determined by gel filtration chromatography consistent with its existence as a
dimer in solution. The insets show the calibration of the column and an SDS gel of the purified SpPlsX preparation.
(B) The activity requirements for the formation of acyl-PO4 from [14C]16:0-ACP by SpPlsX were assessed by thin-layer chromatography of the
reaction products on Silica Gel G layers developed with chloroform:methanol:acetic acid (90/10/10).
(C) Acyl-ACP, but not fatty acid or 16:0-CoA, was a substrate for SpPlsX, which also incorporated [32P]orthophosphate into the product as
determined by thin-layer chromatography on Silica Gel G layers developed with chloroform:methanol:acetic acid (90/10/10). The asterisk (*)
indicates the radiolabeled substrate used in the assay.
(D) Negative ion electrospray precursor ion scanning MS of the product of the SpPlsX reaction using 16:0-ACP and phosphate.
(E) The PlsX reaction was reversible, catalyzing the synthesis of acyl-ACP from 16:0-PO4 as determined by conformationally sensitive gel elec-
trophoresis and staining with Coomassie blue.
(F and G) The apparent SpPlsX KM values for acyl-ACP (F) and orthophosphate (G) were 300 mM and 140 mM, respectively. The data shown are the
means of three independent experiments with the standard errors.
experiments with SpPlsX, but only in the presence of or-thophosphate. This result indicated that the fatty acidsarose from the hydrolysis of the 16:0-PO4 anhydrideand that SpPlsX did not have acyl-ACP thioesteraseactivity. Authentic 16:0-PO4 was chemically synthesized
and characterized, and the synthetic 16:0-PO4 hadthe same Rf as the product of the SpPlsX reaction.Phosphotransacetylase reactions are reversible, andaccordingly, SpPlsX was capable of forming 16:0-ACPfrom ACP and synthetic 16:0-PO4 as well as ACP from
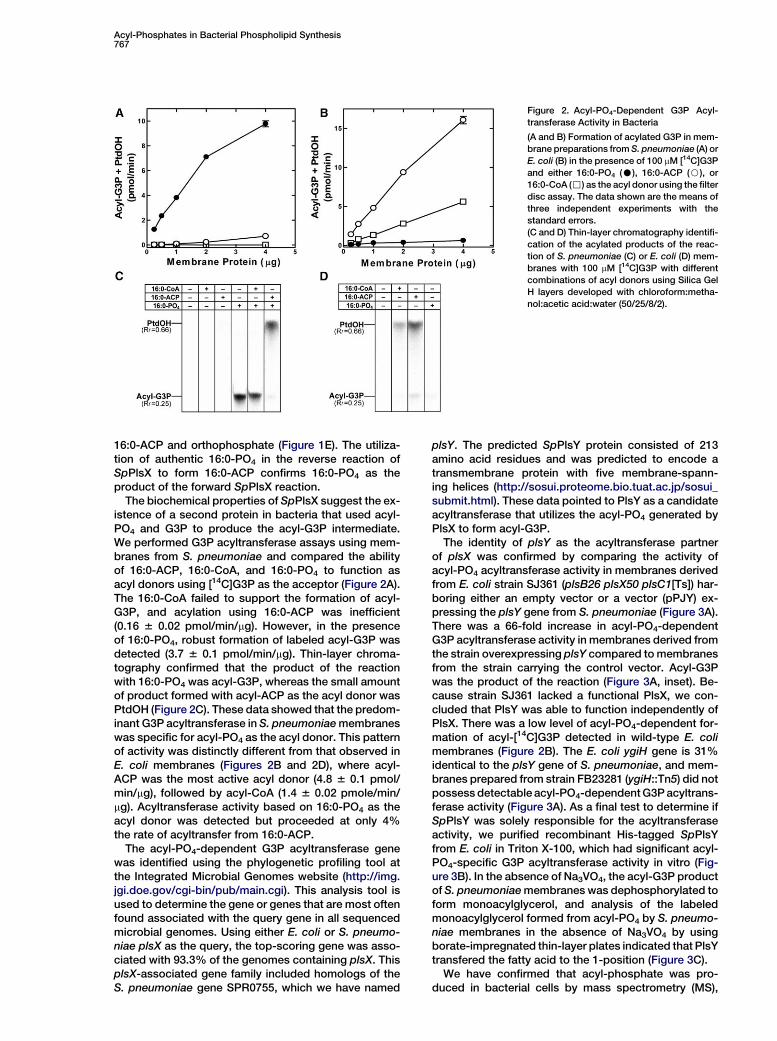
Acyl-Phosphates in Bacterial Phospholipid Synthesis767
Figure 2. Acyl-PO4-Dependent G3P Acyl-
transferase Activity in Bacteria
(A and B) Formation of acylated G3P in mem-
brane preparations from S. pneumoniae (A) or
E. coli (B) in the presence of 100 mM [14C]G3P
and either 16:0-PO4 (C), 16:0-ACP (B), or
16:0-CoA (,) as the acyl donor using the filter
disc assay. The data shown are the means of
three independent experiments with the
standard errors.
(C and D) Thin-layer chromatography identifi-
cation of the acylated products of the reac-
tion of S. pneumoniae (C) or E. coli (D) mem-
branes with 100 mM [14C]G3P with different
combinations of acyl donors using Silica Gel
H layers developed with chloroform:metha-
nol:acetic acid:water (50/25/8/2).
16:0-ACP and orthophosphate (Figure 1E). The utiliza-tion of authentic 16:0-PO4 in the reverse reaction ofSpPlsX to form 16:0-ACP confirms 16:0-PO4 as theproduct of the forward SpPlsX reaction.
The biochemical properties of SpPlsX suggest the ex-istence of a second protein in bacteria that used acyl-PO4 and G3P to produce the acyl-G3P intermediate.We performed G3P acyltransferase assays using mem-branes from S. pneumoniae and compared the abilityof 16:0-ACP, 16:0-CoA, and 16:0-PO4 to function asacyl donors using [14C]G3P as the acceptor (Figure 2A).The 16:0-CoA failed to support the formation of acyl-G3P, and acylation using 16:0-ACP was inefficient(0.16 6 0.02 pmol/min/mg). However, in the presenceof 16:0-PO4, robust formation of labeled acyl-G3P wasdetected (3.7 6 0.1 pmol/min/mg). Thin-layer chroma-tography confirmed that the product of the reactionwith 16:0-PO4 was acyl-G3P, whereas the small amountof product formed with acyl-ACP as the acyl donor wasPtdOH (Figure 2C). These data showed that the predom-inant G3P acyltransferase in S. pneumoniae membraneswas specific for acyl-PO4 as the acyl donor. This patternof activity was distinctly different from that observed inE. coli membranes (Figures 2B and 2D), where acyl-ACP was the most active acyl donor (4.8 6 0.1 pmol/min/mg), followed by acyl-CoA (1.4 6 0.02 pmole/min/mg). Acyltransferase activity based on 16:0-PO4 as theacyl donor was detected but proceeded at only 4%the rate of acyltransfer from 16:0-ACP.
The acyl-PO4-dependent G3P acyltransferase genewas identified using the phylogenetic profiling tool atthe Integrated Microbial Genomes website (http://img.jgi.doe.gov/cgi-bin/pub/main.cgi). This analysis tool isused to determine the gene or genes that are most oftenfound associated with the query gene in all sequencedmicrobial genomes. Using either E. coli or S. pneumo-niae plsX as the query, the top-scoring gene was asso-ciated with 93.3% of the genomes containing plsX. ThisplsX-associated gene family included homologs of theS. pneumoniae gene SPR0755, which we have named
plsY. The predicted SpPlsY protein consisted of 213amino acid residues and was predicted to encode atransmembrane protein with five membrane-spann-ing helices (http://sosui.proteome.bio.tuat.ac.jp/sosui_submit.html). These data pointed to PlsY as a candidateacyltransferase that utilizes the acyl-PO4 generated byPlsX to form acyl-G3P.
The identity of plsY as the acyltransferase partnerof plsX was confirmed by comparing the activity ofacyl-PO4 acyltransferase activity in membranes derivedfrom E. coli strain SJ361 (plsB26 plsX50 plsC1[Ts]) har-boring either an empty vector or a vector (pPJY) ex-pressing the plsY gene from S. pneumoniae (Figure 3A).There was a 66-fold increase in acyl-PO4-dependentG3P acyltransferase activity in membranes derived fromthe strain overexpressing plsY compared to membranesfrom the strain carrying the control vector. Acyl-G3Pwas the product of the reaction (Figure 3A, inset). Be-cause strain SJ361 lacked a functional PlsX, we con-cluded that PlsY was able to function independently ofPlsX. There was a low level of acyl-PO4-dependent for-mation of acyl-[14C]G3P detected in wild-type E. colimembranes (Figure 2B). The E. coli ygiH gene is 31%identical to the plsY gene of S. pneumoniae, and mem-branes prepared from strain FB23281 (ygiH::Tn5) did notpossess detectable acyl-PO4-dependent G3P acyltrans-ferase activity (Figure 3A). As a final test to determine ifSpPlsY was solely responsible for the acyltransferaseactivity, we purified recombinant His-tagged SpPlsYfrom E. coli in Triton X-100, which had significant acyl-PO4-specific G3P acyltransferase activity in vitro (Fig-ure 3B). In the absence of Na3VO4, the acyl-G3P productof S. pneumoniae membranes was dephosphorylated toform monoacylglycerol, and analysis of the labeledmonoacylglycerol formed from acyl-PO4 by S. pneumo-niae membranes in the absence of Na3VO4 by usingborate-impregnated thin-layer plates indicated that PlsYtransfered the fatty acid to the 1-position (Figure 3C).
We have confirmed that acyl-phosphate was pro-duced in bacterial cells by mass spectrometry (MS),

Molecular Cell768
Figure 3. PlsY Is an Acyl-PO4-Dependent G3P Acyltransferase
(A) 16:0-PO4 G3P acyltransferase activity in membranes prepared from E. coli strain SJ361 (plsB26 plsX50 plsC1[Ts]) harboring either an empty
vector (pPJ131, ,) or a plasmid directing the production of SpPlsY (pPJY, -), and E. coli strain FB23281 (ygiH::Tn5) with the E. coli plsY homolog
inactivated by insertion of the Tn5 element (C) using the filter disc assay. Acyl-G3P was the only product detected, and the inset shows the acyl-
G3P region of a thin-layer chromatogram of the products of the reactions using 4 mg of membrane protein. The data shown are the means of three
independent experiments with the standard errors.
(B) Purified SpPlsY was assayed for G3P acyltransferase activity in the presence of 100 mM [14C]G3P and either 16:0-PO4 (C), 16:0-ACP (B), or
16:0-CoA (,). Acyl-G3P was the only product detected by thin-layer chromatography (data not shown). The inset shows an SDS gel of purified
SpPlsY. SpPlsY has a predicted molecular weight of 23 kDa but migrates as a 19 kDa protein in SDS gel electrophoresis. The data shown are the
means of three independent experiments with the standard errors.
(C) Position of the acyl chain in the PlsY reaction product. Acyl-glycerol was prepared as described in the methods section. The location of
the labeled monoacylglycerol (1-MG, Rf = 0.16) was visualized using a PhosphoImager screen. Standards were commercial 1-MG (Rf = 0.16)
and 2-monoacylglycerol (2-MG, Rf = 0.38) prepared by the action of Rhizomucor miehei lipase on diacylglycerol. The only labeled species
detected in the reactions was the 1-acyl isomer.
(D) Negative ion electrospray precursor ion scanning MS of acyl-phosphates isolated from E. coli strain FB23281 harboring plasmid pBlue-
spPlsX.
(E) Product formation in the presence of a mixture of 16:0-PO4 plus 16:0-ACP. Membranes were prepared from strain SJ361 (plsB26 plsX50
plsC1[Ts]) harboring either an empty expression vector (pPJ131), a SpPlsY expression vector (pPJY), or a SpPlsC expression vector (pYL12).
providing in vivo evidence for the acyl-PO4 intermediate.Analysis of lipid extracts from the wild-type S. pneumo-niae strain D39 did not reveal detectable levels of acyl-PO4 by negative ion electrospray precursor ion scanningMS, suggesting that acyl-PO4 is an ephemeral interme-diate that is rapidly used by SpPlsY. Thus, we generatedan E. coli strain to overexpress SpPlsX in a plsY nullbackground by transforming plasmid pBlue-spPlsX intostrain FB23281 (ygiH::Tn5). Precursor ion scanning MSof the lipid extract from the resultant strain demon-strated the presence of two major acyl-PO4 intermedi-ates, 16:0-PO4 and 18:1-PO4 (Figure 3D). These twopeaks correspond to the primary fatty acids found in
the 1-position of bacterial phospholipids and indicatedthat SpPlsX was active with both saturated and unsatu-rated acyl-ACPs in vivo.
The second step in PtdOH formation is catalyzed byPlsC in E. coli, which, like PlsB, utilizes either acyl-ACPor acyl-CoA to acylate 1-acyl-G3P. S. pneumoniae con-tains a PlsC homolog (SPR1465) that is 31.4% identicalto E. coli PlsC. The introduction of a plasmid (pYL12)expressing SpPlsC into E. coli strain SM2-1 (plsC1[Ts])corrected the temperature-sensitive growth phenotypeof the bacterium, illustrating that SpPlsC was function-ally interchangeable with EcPlsC and was capable ofutilizing acyl-ACP as the acyl donor. This conclusion

Acyl-Phosphates in Bacterial Phospholipid Synthesis769
Figure 4. The PlsX/Y Pathway for PtdOH For-
mation Is the Predominant Route to PtdOH in
Bacteria
(A) The most widely distributed pathway
starts with the conversion of a long-chain
acyl-ACP end product of fatty acid synthesis
to an acyl-PO4 by PlsX. PlsY transfers the
fatty acid from the acyl-PO4 to G3P. Acyl-
G3P is then converted to PtdOH by PlsC
using acyl-ACP as the acyl donor.
(B) The alternate pathway proceeds by the
PlsB-catalyzed transfer of a fatty acid to
G3P from acyl-ACP (or acyl-CoA) followed
by the acylation of acyl-G3P by PlsC.
(C) Distribution of the PlsB (red), PlsX/Y
(green), and PlsC (blue) acyltransferase genes
in bacteria. PlsX/Y occurred in 314 genomes,
PlsC occurred in 350 genomes, and PlsB
occurred in 102 genomes in the Integrated
Microbial Genomes database (a total of 271
finished/124 draft sequences were interro-
gated).
was confirmed in vitro using membranes isolated fromS. pneumoniae with both acyl-ACP and acyl-PO4 asacyl donors and [14C]G3P as the acceptor (Figure 2C).When acyl-ACP alone was added, there was little prod-uct formed from G3P, and in the presence of 16:0-PO4,acyl-G3P was detected. In the presence of 16:0-PO4
plus 16:0-ACP donors, PtdOH was the primary product,indicating that acyl-ACP was the substrate for SpPlsCand that acyl-PO4 was not. However, 16:0-PO4 plus16:0-CoA did not yield PtdOH, indicating that SpPlsCdid not use acyl-CoA as an acyl donor (Figure 2C). Inthe E. coli system, 16:0-PO4 formed only traces ofacyl-G3P, whereas either acyl-CoA or acyl-ACP led tothe formation of both acyl-G3P and PtdOH (Figure 2D).The products of SpPlsY and SpPlsC were confirmedusing membranes prepared from E. coli strain SJ361transformed with either a SpPlsY or a SpPlsC expres-sion vector and assayed at 42�C with a mixture of acyl-PO4 and acyl-ACP. Membranes containing only SpPlsYproduced exclusively acyl-G3P, whereas the additionof membranes expressing SpPlsC produced primarilyPtdOH (Figure 3E).
This work describes the most prevalent pathway forthe acylation of G3P in bacteria that utilizes a new acti-vated fatty acid intermediate, acyl-PO4 (Figure 4A). Thispathway begins by the conversion of the acyl-ACP endproducts of type II fatty acid synthesis to acyl-PO4 inter-
mediates by the soluble PlsX enzyme. The acyl-PO4 isthen utilized by the membrane-associated PlsY G3Pacyltransferase to form 1-acyl-G3P. PlsC completesthe synthesis of the PtdOH intermediate by using acyl-ACP to transfer a second fatty acid to acyl-G3P. ThePlsX protein family (pfam02504/COG0416) should bedefined as phosphate:acyl-ACP acyltransferase. Simi-larly, the PlsY protein family (pfam02660/COG0344) en-compassing the domain of unknown function, DUF205,should be defined as acyl-PO4 G3P acyltransferase.The first PtdOH pathway discovered in the E. coli modelsystem involves two membrane-associated enzymesthat sequentially acylate G3P at the 1-position (PlsB) fol-lowed by the 2-position (PlsC) with both acyltrans-ferases utilizing acyl-ACP (or acyl-CoA) (Figure 4B).The PlsX/Y pathway is widely distributed in 314 bacterialgenomes out of 271 finished and 124 draft bacterial se-quences deposited in the Integrated Microbial Genomesdatabase (Figure 4C). The only group that did not con-tain PlsX/Y was the Xanthomonadales in the g-proteo-bacteria, which have PlsB only. Archaea and eukaryado not have a recognizable PlsX/Y pathway. The PlsBpathway is found in 102 of the same genomes and is re-stricted primarily to the g-proteobacteria (Figure 4C).The existence of PlsB in these bacteria provides themwith an acyltransferase capable of utilizing acyl-CoA,which allows the incorporation of exogenous fatty acids

Molecular Cell770
into membrane phospholipids. The plsB gene may notbe essential in many of these bacteria, most of whichalso contain the PlsX/Y system; for example, P. aerugi-nosa (Jacobs et al., 2003). On the other hand, B. subtilislacks plsB, and both the plsX and plsY genes are essen-tial (Kobayashi et al., 2003). PlsC is universally distrib-uted in bacteria (Figure 4C).
The regulatory properties of the acyl donors for phos-pholipid synthesis in controlling bacterial lipid metabo-lism suggest that acyl-PO4 may also play a regulatoryrole in addition to its function as a biosynthetic interme-diate. Acyl-CoA is also a ligand for the FadR transcrip-tional regulator that controls fabA (Cronan and Subrah-manyam, 1998) and fabB gene expression (Campbelland Cronan, 2001). Acyl-ACP is an allosteric regulator ofthe initiating steps in fatty acid synthesis, acetyl-CoAcarboxylase (Davis and Cronan, 2001), and b-ketoacyl-ACP synthase III (Heath and Rock, 1996). A regulatoryfunction for acyl-PO4 may explain the retention of PlsX/Yin bacteria that possess PlsB, and there are transcrip-tional regulators that control the expression of the genesof fatty acid biosynthesis whose regulatory ligands haveyet to be positively identified (Lu and Rock, 2006; Schuj-man et al., 2003; Zhang et al., 2002). It will be important todetermine if acyl-PO4 signals through these factors orperhaps has a role in the coordination of membrane for-mation with macromolecular synthesis, which remainslargely experimentally undefined (Cronan and Rock,1996; Heath et al., 1994; Rock and Jackowski, 2002;Seyfzadeh et al., 1993). Another important distinctionbetween the PlsX/Y and PlsB pathways is that PlsB iscapable of using either acyl-ACP or acyl-CoA as theacyl donor. Many g-proteobacteria, like E. coli, are capa-ble of transporting exogenous fatty acids and convertingthem to acyl-CoAs (Black and DiRusso, 2003). The exis-tence of PlsB is an advantage in these bacteria that useexogenous fatty acids to produce membrane phospho-lipid in lieu of the energy-expensive fatty acid synthesispathway (Cronan and Rock, 1996; Polacco and Cronan,1977). The discovery of the essential role of PlsX/Y in theFirmicutes also has potential medical relevance. Thisgroup of bacteria contains the major human Gram-posi-tive pathogens and uses the PlsX/Y pathway exclusively(Figure 4C). These essential enzymes are unique to bac-teria and thus represent attractive targets for the devel-opment of antibacterial agents to combat the growingproblem of resistance to current therapies.
Experimental Procedures
Materials
Sources of supplies were the following: Avanti Polar Lipids, Inc., pal-
mitoyl-CoA and phospholipids standards; Sigma Chemical Co., G3P
and fatty acid free BSA. Acyl-ACPs were prepared enzymatically by
using theacyl-ACPsynthetasemethod (Rock andCronan,1981; Rock
and Garwin, 1979). Protein was measured by the Bradford method
(Bradford, 1976). All other chemicals were of reagent grade or better.
Strain SJ361 harbors plsB26, plsX50, and plsC1(Ts) mutations
(Heath and Rock, 1999) and has an auxotrophic requirement for
G3P and a temperature-sensitive growth phenotype. Strain
FB23281 (ygiH::Tn5) was obtained from E. coli genome project (Uni-
versity of Wisconsin, Madison).
Cloning the plsX, plsY, and plsC Genes from S. pneumoniae
SPR0037 (plsX) was amplified and inserted into pET15b within NdeI
and BamHI sites to generate pET-plsX. SPR0755 (plsY) was ampli-
fied and inserted within NcoI and XhoI sites of pPJ131 (derived
from pBluescript II KS+ containing the XbaI-BamHI fragment from
pET15b) to generate plasmid pPJY, which expressed SpPlsY with
a C-terminal His tag. SPR1465 (plsC) was amplified and cloned
into plasmid pET28a within NdeI and XhoI sites and then transferred
to pBluescript II KS+ using XbaI and XhoI enzymes to generate plas-
mid pYL12. All plasmids were sequenced.
Expression and Purification of SpPlsX and SpPlsY
Vectors pET-plsX and pPJY were used to transform E. coli Rosetta
competent cells for protein expression. SpPlsX was induced with
1 mM IPTG for 3 hr at 37�C, and SpPlsY was induced with 0.2 mM
IPTG at 16�C overnight. Cells were collected by centrifugation
(6000 rpm, 4�C, 15 min), and cell pellets were lysed with a french
press. Membranes from SpPlsY cultures were isolated (see below)
and solubilized with 0.5% Triton X-100. Soluble proteins were ap-
plied to a Ni2+-agarose column and washed with MCAC-40 buffer
(with 0.5% Triton X-100 for PlsY). His-tagged proteins were eluted
with MCAC-200 buffer (with 0.1% Triton X-100 for SpPlsY). Purified
proteins were dialyzed against 20 mM Tris-HCl (pH 7.4), 5 mM EDTA,
100 mM NaCl, and 1 mM DTT (with 0.1% Triton X-100 for SpPlsY).
The proteins were stored at 220�C.
Synthesis of Palmitoyl-PO4
The synthesis of 16:0-PO4 was achieved using the method of Leh-
ninger for the synthesis of acylphosphates (Lehninger, 1945). Silver
phosphate (376 mg, 0.90 mmol) was added to anhydrous phospho-
ric acid (199 mg, 2.03 mmol) in diethyl ether (10 ml), and the mixture
was stirred at room temperature for 16 hr. Palmitoyl chloride
(558 mg, 2.03 mmol) was dissolved in diethyl ether (5 ml) and added
dropwise to the resulting mixture. After the addition was complete,
the mixture was stirred for an additional hour. The resulting mixture
was filtered, and the retained solid was washed with diethyl ether
(2 3 5 ml). The filtrate and washings were combined and concen-
trated in vacuo. Recrystallization of the resulting residue from warm
benzene yielded 16:0-PO4, a white powder (165 mg, 24%). 1HNMR
(CD3OD, 300 MHz), d 0.92 (t, 3H, J = 6.0 Hz), 1.31 (bs, 26 H), 1.65
(m, 2H), 2.46 (t, 2H, J = 7.0 Hz). 13CNMR (CD3OD, 300MHz), d 12.5,
21.8, 23.6, 28.6, 28.7, 28.8, 31.1, 34.0, 168.3. MS(ESI), m/z = 359
[M + Na]+; MS(ESI), m/z = 335 [M 2 H]2.
SpPlsX Assay
The reaction mix for acyl chain transfer to orthophophate contained
0.1 M HEPES (pH 7.4), 0.1 M sodium phosphate, 2 mM MgCl2, and
200 mM 16:0-ACP. The reaction for acyl chain transfer from 16:0-
PO4 to ACP contained 0.1 M HEPES (pH 7.4), 1 mM DTT, 2 mM
MgCl2, 200 mM 16:0-PO4, and 200 mM ACP. Reaction was started
by adding SpPlsX and was incubated at 37�C for 20 min. [32P]phos-
phate incorporation into 16:0-PO4 was measured in assays contain-
ing 4 ml of [32P]phosphate (4000 Ci/mmol, GE Healthcare). A G-50
spin column (GE Healthcare) was used to remove the unreacted
[32P]phosphate. Samples were analyzed by TLC using preadsorbent
G layers (Analtech) in chloroform/methanol/acetic acid (90/10/10,
v/v/v) solvent system or by conformationally sensitive gel electro-
phoresis in 13% polyacrylamide gels containing 0.5 M urea (Cronan,
1982; Rock et al., 1981a).
Extraction of Acyl-PO4, Acyl-G3P, and PtdOH
Acyl-PO4, acyl-G3P, and PtdOH were extracted from reaction mix-
tures using the method for phosphoinositides extraction as de-
scribed (Jackowski et al., 1986). Briefly, 50 ml reaction mixture was
mixed with 180 ml of chloroform:methanol:concentrated HCl
(1:2:0.02, v/v/v) followed by 60 ml of chloroform and 60 ml of 2 M
KCl. Samples were vortexed and centrifuged for 2 min to separate
the phases. The lower phase was transferred to a new tube, and
the upper phase was extracted again with 100 ml chloroform and
combined with the first extraction. The organic phase was evapo-
rated under a stream of nitrogen and redissolved in chloroform:
methanol (1:1, v/v).
Mass Spectrometry Detection of Acyl-PO4
MS of 16:0-PO4 was performed by the Hartwell Center for Bioinfor-
matics and Biotechnology at St. Jude Children’s Research Hospital.
A SpPlsX reaction was extracted using chloroform:methanol:

Acyl-Phosphates in Bacterial Phospholipid Synthesis771
concentrated HCl and dissolved in 50% methanol:water. MS
analysis was performed using a Finnigan TSQ Quantum (Thermo
Electron, San Jose, California) triple quadrupole mass spectrometer
equipped with the nanospray ion source. The instrument was oper-
ated in the negative ion mode using single MS (Q1) scanning and/or
precursor ion scanning corresponding to the loss of the phosphate
group. Ion source parameters were as follows: spray voltage 1600 V,
capillary temperature 270�C, and capillary offset 235 V, and tube
lens offset was set by infusion of the polytyrosine tuning and calibra-
tion solution (Thermo Electron, San Jose, California) in electrospray
mode. MS acquisition parameters for Q1 scanning were as follows:
scan range 225–500 m/z, scan time 0.1 s, and peak width Q1 0.7
FWHM. MS acquisition parameters for parent ion scanning were
the following: scan range 225–500 m/z, scan time 0.1 s, product
mass 79 m/z, collision energy 15 V, peak width Q1 and Q3 0.7
FWHM, and Q2 CID gas (argon) 0.5 mTorr. Instrument control and
data acquisition were performed with the Finnigan Xcalibur (version
1.4 SR1) software (Thermo Electron, San Jose, California).
Acyl-PO4 was detected in vivo by cloning SpPlsX into pBluescript
II KS+ to generate plasmid pBlue-spPlsX, which was used to trans-
form E. coli strain FB23281 (ygiH::Tn5). The resulting strain was
grown in LB medium at 37�C. Cells were collected by centrifugation
(6000 rpm, 4�C, 10 min) when OD600 reached 1.0, and total lipids in-
cluding acyl-PO4 intermediates were extracted using chloroform:
methanol:concentrated HCl as described above. The dried sample
was dissolved in 50% methanol:water and analyzed by negative
ion electrospray precursor ion scanning MS as described above.
Preparation of Membranes and Assay of Acyltransferase
Activity
Membranes were purified from 500 ml early stationary phase E. coli
or S. pneumoniae using the sucrose gradient centrifugation method
(Ito et al., 1977; Rock et al., 1981b). In the experiments using SJ361
cells, cultures were shifted to 45�C for 30 min prior to harvest to in-
activate PlsC. Membranes in 10% glycerol were used within 2 days
when stored at 4�C or were stored at 280�C for up to 2 weeks.
Acyltransferase assays contained 0.1 M Tris-HCl (pH 9), 0.1 M so-
dium phosphate, 1 mg/ml bovine serum albumin, 2 mM MgCl2, 5 mM
Na3VO4 (to inhibit phosphatases), and 100 mM [U-14C]G3P (8 mCi/
mmol, American Radiolabeled Chemicals Inc.). Reactions were
started by adding either 40 mM 16:0-CoA, 200 mM 16:0-ACP, or
200 mM 16:0-PO4 and were terminated after incubation at 37�C for
30 min by pipetting 20 ml of the reaction mixture onto a Whatman
No. 3 mm filter disc. Filter discs were subsequently washed in
10%, 5%, and 1% ice-cold trichloroacetic acid (20 min, 20 ml/disc)
prior to scintillation counting (Heath and Rock, 1998).
Position of the Acyl Chain in the PlsY Reaction Product
PlsY acyltransferase reactions were carried out as described
above using membranes from either S. pneumoniae strain D39 or
E. coli strain SJ361/pPJY, except Na3VO4 was omitted leading to
the formation of monoacylglycerol by dephosphorylation of acyl-
G3P. Products were extracted and spotted onto a Silica G plate
impregnated with 5% boric acid (Analtech). The plate was devel-
oped in chloroform:acetone (80:20, v/v), and the location of the
labeled monoacylglycerol was visualized using a PhosphoImager
screen.
Acknowledgments
We thank Fan Zhang, Amy Sullivan, Matthew Frank, and Pam Jack-
son for their technical assistance; Athanasios Lykidis for his insight-
ful bioinformatic analysis; Phil Poston and the St. Jude Hartwell Cen-
ter for mass spectra; and Suzanne Jackowski for strain SJ361 and
her review of the manuscript. This work was supported by National
Institutes of Health Grant GM 34496, Cancer Center (CORE) Support
Grant CA 21765, and the American Lebanese and Syrian Associated
Charities.
Received: April 28, 2006
Revised: June 16, 2006
Accepted: June 30, 2006
Published: August 31, 2006
References
Badger, J., Sauder, J.M., Adams, J.M., Antonysamy, S., Bain, K.,
Bergseid, M.G., Buchanan, S.G., Buchanan, M.D., Batiyenko, Y.,
Christopher, J.A., et al. (2005). Structural analysis of a set of
proteins resulting from a bacterial genomics project. Proteins 60,
787–796.
Bell, R.M. (1974). Mutants of Escherichia coli defective in membrane
phospholipid synthesis: macromolecular synthesis in an sn-glycerol
3-phosphate acyltransferase Km mutant. J. Bacteriol. 117, 1065–
1076.
Bell, R.M. (1975). Mutants of Escherichia coli defective in membrane
phospholipid synthesis: properties of wild type and Km defective
sn-glycerol-3-phosphate acyltransfersae activities. J. Biol. Chem.
250, 7147–7152.
Black, P.N., and DiRusso, C.C. (2003). Transmembrane movement
of exogenous long-chain fatty acids: proteins, enzymes, and vecto-
rial esterification. Microbiol. Mol. Biol. Rev. 67, 454–472.
Bradford, M.M. (1976). A rapid and sensitive method for quantitation
of microgram quantities of protein utilizing the principle of protein-
dye binding. Anal. Biochem. 72, 248–254.
Campbell, J.W., and Cronan, J.E., Jr. (2001). Escherichia coli FadR
positively regulates transcription of the fabB fatty acid biosynthetic
gene. J. Bacteriol. 183, 5982–5990.
Coleman, J. (1992). Characterization of the Escherichia coli gene for
1-acyl-sn-glycerol-3-phosphate acyltransferase (plsC). Mol. Gen.
Genet. 232, 295–303.
Cronan, J.E., Jr. (1982). Molecular properties of short chain acyl thio-
esters of acyl carrier protein. J. Biol. Chem. 257, 5013–5017.
Cronan, J.E., Jr., and Rock, C.O. (1996). Biosynthesis of membrane
lipids. In Escherichia coli and Salmonella typhimurium: Cellular and
Molecular Biology, F.C. Neidhardt, R. Curtis, C.A. Gross, J.L. Ingra-
ham, E.C.C. Lin, K.B. Low, B. Magasanik, W. Reznikoff, M. Riley, M.
Schaechter, and H.E. Umbarger, eds. (Washington, DC: American
Society for Microbiology), pp. 612–636.
Cronan, J.E., Jr., and Subrahmanyam, S. (1998). FadR, transcrip-
tional co-ordination of metabolic expediency. Mol. Microbiol. 29,
937–943.
Davis, M.S., and Cronan, J.E., Jr. (2001). Inhibition of Eschericia coli
acetyl coenzyme A carboxylase by acyl-acyl carrier protein. J. Bac-
teriol. 183, 1499–1503.
Green, P.R., Merrill, A.H., Jr., and Bell, R.M. (1981). Membrane phos-
pholipid synthesis in Escherichia coli: purification, reconstitution,
and characterization of sn-glycerol-3-phosphate acyltransferase.
J. Biol. Chem. 256, 11151–11159.
Heath, R.J., and Rock, C.O. (1996). Inhibition of b-ketoacyl-acyl car-
rier protein synthase III (FabH) by acyl-acyl carrier protein in Escher-
ichia coli. J. Biol. Chem. 271, 10996–11000.
Heath, R.J., and Rock, C.O. (1998). A conserved histidine is essential
for glycerolipid acyltransferase catalysis. J. Bacteriol. 180, 1425–
1430.
Heath, R.J., and Rock, C.O. (1999). A missense mutation accounts
for the defect in the glycerol-3-phosphate acyltransferase ex-
pressed in the plsB26 mutant. J. Bacteriol. 181, 1944–1946.
Heath, R.J., Jackowski, S., and Rock, C.O. (1994). Guanosine tetra-
phosphate inhibition of fatty acid and phospholipid synthesis in
Escherichia coli is relieved by overexpression of glycerol-3-phos-
phate acyltransferase (plsB). J. Biol. Chem. 269, 26584–26590.
Holm, L., and Park, J. (2000). DaliLite workbench for protein struc-
ture comparison. Bioinformatics 16, 566–567.
Ito, K., Sato, T., and Yura, T. (1977). Synthesis and assembly of the
membrane proteins in E. coli. Cell 11, 551–559.
Jackowski, S., Rettenmier, C.W., Sherr, C.J., and Rock, C.O. (1986).
A guanine nucleotide-dependent phosphatidylinositol 4,5- diphos-
phate phospholipase C in cells transformed by the v-fms and v-fes
oncogenes. J. Biol. Chem. 261, 4978–4985.
Jacobs, M.A., Alwood, A., Thaipisuttikul, I., Spencer, D., Haugen, E.,
Ernst, S., Will, O., Kaul, R., Raymond, C., Levy, R., et al. (2003). Com-
prehensive transposon mutant library of Pseudomonas aeruginosa.
Proc. Natl. Acad. Sci. USA 100, 14339–14344.

Molecular Cell772
Kobayashi, K., Ehrlich, S.D., Albertini, A., Amati, G., Andersen, K.K.,
Arnaud, M., Asai, K., Ashikaga, S., Aymerich, S., Bessieres, P., et al.
(2003). Essential Bacillus subtilis genes. Proc. Natl. Acad. Sci. USA
100, 4678–4683.
Larson, T.J., Ludtke, D.N., and Bell, R.M. (1984). sn-Glycerol-3-
phosphate auxotrophy of plsB strains of Escherichia coli: evidence
that a second mutation, plsX, is required. J. Bacteriol. 160, 711–717.
Lehninger, A.L. (1945). Synthesis and properties of the acyl phos-
phates of some higher fatty acids. J. Biol. Chem. 162, 333–342.
Lightner, V.A., Larson, T.J., Tailleur, P., Kantor, G.D., Raetz, C.R.H.,
Bell, R.M., and Modrich, P. (1980). Membrane phospholipid synthe-
sis in Escherichia coli: cloning of a structural gene (plsB) of the
sn-glycerol-3-phosphate acyltransferase. J. Biol. Chem. 255,
9413–9420.
Lu, Y.-J., and Rock, C.O. (2006). Transcriptional regulation of fatty
acid biosynthesis in Streptococcus pneumoniae. Mol. Microbiol.
59, 551–566.
Polacco, M.L., and Cronan, J.E., Jr. (1977). Mechanism of the appar-
ent regulation of Escherichia coli unsaturated fatty acid synthesis by
exogenous oleic acid. J. Biol. Chem. 252, 5488–5490.
Rock, C.O., and Cronan, J.E., Jr. (1981). Acyl-acyl carrier protein
synthetase from Escherichia coli. Methods Enzymol. 71, 163–168.
Rock, C.O., and Garwin, J.L. (1979). Preparative enzymatic synthesis
and hydrophobic chromatography of acyl-acyl carrier protein.
J. Biol. Chem. 254, 7123–7128.
Rock, C.O., and Jackowski, S. (2002). Forty years of fatty acid bio-
synthesis. Biochem. Biophys. Res. Commun. 292, 1155–1166.
Rock, C.O., Cronan, J.E., Jr., and Armitage, I.M. (1981a). Molecular
properties of acyl carrier protein derivatives. J. Biol. Chem. 256,
2669–2674.
Rock, C.O., Goelz, S.E., and Cronan, J.E., Jr. (1981b). Phospholipid
synthesis in Escherichia coli. Characteristics of fatty acid transfer
from acyl-acyl carrier protein to sn-glycerol-3-phosphate. J. Biol.
Chem. 256, 736–742.
Schujman, G.E., Paoletti, L., Grossman, A.D., and de Mendoza, D.
(2003). FapR, a bacterial transcription factor involved in global reg-
ulation of membrane lipid biosynthesis. Dev. Cell 4, 663–672.
Seyfzadeh, M., Keener, J., and Nomura, M. (1993). spoT-dependent
accumulation of guanosine tetraphosphate in response to fatty acid
starvation in Escherichia coli. Proc. Natl. Acad. Sci. USA 90, 11004–
11008.
Xu, Q.S., Jancarik, J., Lou, Y., Kuznetsova, K., Yakunin, A.F., Yokota,
H., Adams, P., Kim, R., and Kim, S.-H. (2005). Crystal structures of
a phosphotransacetylase from Bacillus subtilis and its complex
with acetyl phosphate. J. Struct. Funct. Genomics 6, 269–279.
Zhang, Y.-M., Marrakchi, H., and Rock, C.O. (2002). The FabR (YijC)
transcription factor regulates unsaturated fatty acid biosynthesis in
Escherichia coli. J. Biol. Chem. 277, 15558–15565.




















