
PECTIC ENZYMES SECRETED BY VERTICILLIUM DAHLIAE AND THEIR ROLE IN THE DEVELOPMENT OF THE WILT...
19
KAMAL, M. & WOOD, R. K. S. (1956). Ann. appl. Biol. 44 (z), 322-340. PECTIC ENZYMES SECRETED BY VERTICILLIUM MENT OF THE WILT DISEASE OF COTTON DAHLIAE AND THEIR ROLE IN THE DEVELOP- BY M. KAMAL AND R. K. S. WOOD Botany Department, Imperial College, London, S. W. 7 (With z Text-figures) An isolate of Verticiiliurn dahliae obtained from Uganda was highly virulent to young cotton plants under greenhouse conditions. A hyaline variant which often appeared in culture was as virulent as the parent isolate, but preliminary experiments indicated that it did not survive as long in unsterile soil. The parent isolate grew rapidly in cotton plants after root inoculation and was isolated from stems and leaves well before the appearance of disease symptoms visible to the naked eye. Protopectinase was produced in the absence of pectic materials, but more active preparations were obtained when media contained pectic substances. In general, there was a close correspondence between the protopectinase activity of culture filtrates and the toxicity of these filtrates to parenchymatous cells. Some separation of the two activities was obtained by heating enzyme solutions or by plasmolysing the test tissue. Protopectinase solutions had little pectinesterase activity but rapidly reduced the viscosity of solutions of pectic substances. In general, the properties of proto- pectinase and the viscosity-reducing enzyme were similar. Young cotton shoots wilted rapidly when placed in cell-free filtrates from cultures of the pathogen. Wilting was delayed under conditions unfavourable for transpiration. Evidence was obtained which showed that wilting was caused by the uptake of thermostable compounds of high molecular weight which impeded the upward flow of the vascular sap. Pronounced vascular browning was obtained only when solutions containing protopectinase were used. Wilting and vascular browning were obtained with solutions having little pectinesterase activity; in contrast, a solution having high pectinesterase activity produced relatively little vascular browning. INTRODUCTION Species of Verticillium are responsible for diseases of a wide range of higher plants; they are soil-borne pathogens which invade roots and become established in the vascular system. The symptoms of disease which develop depend to some extent upon the host plant and conditions of growth, but infection is generally followed by wilting and death of a part or the whole of the shoot system. The mechanisms by which these effects are produced have not been as well studied as those involved in the wilt diseases caused by certain Fusaria where the pattern of symptom development is very similar. The wilt of tomato plants caused by Fusarium oxysporum f. lycopersici has been investigated by many workers, particularly in
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of PECTIC ENZYMES SECRETED BY VERTICILLIUM DAHLIAE AND THEIR ROLE IN THE DEVELOPMENT OF THE WILT...

KAMAL, M. & WOOD, R. K. S . (1956). Ann. appl. Biol. 44 (z), 322-340.
PECTIC ENZYMES SECRETED BY VERTICILLIUM
MENT OF THE WILT DISEASE OF COTTON DAHLIAE AND THEIR ROLE IN THE DEVELOP-
BY M. KAMAL AND R. K. S. WOOD Botany Department, Imperial College, London, S. W. 7
(With z Text-figures)
An isolate of Verticiiliurn dahliae obtained from Uganda was highly virulent to young cotton plants under greenhouse conditions. A hyaline variant which often appeared in culture was as virulent as the parent isolate, but preliminary experiments indicated that it did not survive as long in unsterile soil. The parent isolate grew rapidly in cotton plants after root inoculation and was isolated from stems and leaves well before the appearance of disease symptoms visible to the naked eye.
Protopectinase was produced in the absence of pectic materials, but more active preparations were obtained when media contained pectic substances. In general, there was a close correspondence between the protopectinase activity of culture filtrates and the toxicity of these filtrates to parenchymatous cells. Some separation of the two activities was obtained by heating enzyme solutions or by plasmolysing the test tissue.
Protopectinase solutions had little pectinesterase activity but rapidly reduced the viscosity of solutions of pectic substances. In general, the properties of proto- pectinase and the viscosity-reducing enzyme were similar.
Young cotton shoots wilted rapidly when placed in cell-free filtrates from cultures of the pathogen. Wilting was delayed under conditions unfavourable for transpiration. Evidence was obtained which showed that wilting was caused by the uptake of thermostable compounds of high molecular weight which impeded the upward flow of the vascular sap. Pronounced vascular browning was obtained only when solutions containing protopectinase were used. Wilting and vascular browning were obtained with solutions having little pectinesterase activity; in contrast, a solution having high pectinesterase activity produced relatively little vascular browning.
INTRODUCTION Species of Verticillium are responsible for diseases of a wide range of higher plants; they are soil-borne pathogens which invade roots and become established in the vascular system. The symptoms of disease which develop depend to some extent upon the host plant and conditions of growth, but infection is generally followed by wilting and death of a part or the whole of the shoot system. The mechanisms by which these effects are produced have not been as well studied as those involved in the wilt diseases caused by certain Fusaria where the pattern of symptom development is very similar. The wilt of tomato plants caused by Fusarium oxysporum f . lycopersici has been investigated by many workers, particularly in

Pectic enzymes secreted by Verticillium dahliae 323 recent years, but in spite of the accumulation of much information on the action on the host plant of culture filtrates and specific metabolic products of the pathogen, there is no satisfactory explanation of the way in which the growth of the fungus in the vascular elements causes the plant to wilt and die, When the present study was started there was relatively little information on the effect of filtrates from cultures of Verticillium species on host plants. Accordingly, in 1952, following a suggestion from Dr C. M. Wickens, of the Empire Cotton Growing Corporation, Namulonge, Uganda, a study was begun of the wilt disease of cotton and of the causal pathogen, V . dahliae. The work to be reported in this paper deals primarily with the pectic enzymes secreted by this pathogen in liquid culture and the effects of culture filtrates on certain parenchymatous tissue and on cotton plants.
MATERIALS AND GENERAL METHODS
Origin and identity of the pathogen A culture of the pathogen, identified as V . dahliae Kleb. (isolate number W52/1)
and isolated from a diseased cotton plant was supplied by Dr C. M. Wickens. Cultures were obtained from single spores and maintained on sucrose-nitrate agar slants. Stock master cultures on this agar were stored at - 16" C. and remained viable for at least 2 years. The fungus grew well on this medium and produced spores and microsclerotia abundantly. From time to time, a variant appeared which spored less profusely and did not produce microsclerotia; it was therefore readily distinguished from the parent isolate by its white appearance and was easily avoided when subcultures were made.
Host plants Cotton seed of a susceptible (U4/8161) and a resistant (B 181) variety were
supplied by Dr G. M. Wickens. Seedlings were grown in a roof greenhouse at 20-25O C., using John Innes potting compost contained in seed boxes; they were transplanted to 5 in. pots on the appearance of the first true leaf; during the winter months, additional illumination of 160 foot-candles was provided to ensure a reasonably sturdy growth. Healthy seedlings were obtained from some 80% of the seed sown.
Preparation of culture filtrates Various liquid media were used in the course of the investigation; 40 ml. of
media were placed in a medicine bottle measuring 13 x 6 x 3-5 cm. and sterilized by autoclaving at 120O C. for 15 min. ; media were inoculated with 1.0 ml. of a spore suspension prepared by adding 40 ml. of sterile distilled water to the surface of a 7-day culture on sucrose-nitrate agar contained in medicine bottles. After incubation at 22.5' C. cultures were passed through muslin before centrifuging at 10,000 r.p.m. for 10 min. The clear cell-free supernatant fluid was stored in 15 ml. quantities at - 16" C. until required.

324 M. K A M A L A N D R. K. S. WOOD
Measurement of growth, enzyme activity and toxicity (u) Growth. Mycelium weights were obtained by collecting the hyphal mat on
stretched muslin and washing with distilled water; the mycelium was then trans- ferred to a weighed aluminium foil container and the dry weight determined after 24 hr. at 70' C. Preliminary experiments had shown that the weights obtained in this way were only I-2% less than those obtained by methods which retained spores and the smaller hyphal fragments.
(b ) Protopectinase or macerating activity was estimated with medullary tissue of potato tubers and turnip roots and mesocarp tissue of cucumber fruit. Cylinders, 8 mm. in diameter, were injected with water under vacuum for 30 min. and disks of the following thicknesses were cut with a sliding microtome: turnip 0.3 cm., potato 0.4 cm. and cucumber 0.5 cm. The disks were washed, dried between filter-papers and five were placed in test solutions at 25' C. The time taken for the disks to lose coherence under slight tension was noted as the reaction time (R.T.) and protopectinase activity was expressed as IOOO/R.T. (min.).
( c ) Toxicity. The technique used for estimating the toxicity of solutions to parenchymatous cells was similar to that described by Tribe (1955) and was based on the assumption that a cell was dead when it did not plasmolyse in a non-toxic hypertonic solution. Twenty disks of the test tissue, generally the medulla of potato tuber, were immersed in the test solution; at appropriate intervals, two disks were removed, washed quickly and transferred to a plasmolysing solution con- taining a vital stain. The protoplasts which were plasmolysed appeared as dark red spots after 10-15 min. and, with practice, the number of plasmolysed cells in a disk could be estimated with some accuracy ; it was therefore possible, by transferring disks from the test solution to the plasmolysing solution at suitable intervals, to obtain an estimate of the time taken for half the cells of the disks to be killed. This time will be indicated by the term D5o and the toxicity of a solution expressed as 1ooo/D50 (min.).
The plasmolysing solution had the following composition: M - K N O ~ 8.5 ml. (0.5~ when cucumber was used), 0'1 % neutral redchloride 1.0 ml., M/Ig-phosphate buffer at pH 7.6, 0.5 ml.
( d ) Wilting activity. Seedlings, 3-4 weeks old, were removed from the soil and the stems cut under water just above soil level; they were left in water for 30 min., and any plants showing signs of wilting after this treatment were discarded. The seedlings were transferred to test solutions contained in small vials and placed at 25' C. under diffuse illumination of 250 foot-candles. Two seedlings were used for each test, and the time taken for irreversible wilting to occur was taken as a measure of the wilting activity of the solution. Wilting was considered irreversible when the seedlings did not recover on transfer to water.
In some experiments the ability of solutions to cause browning of the vascular elements was also observed. Seedlings were kept in test solutions for 48 hr. and

Pectic enzymes secreted by Verticillium dahliae 325 left for a further 24 hr. in water; 2-3 mm. of the stem ends were removed daily and the test solutions were changed twice daily. The epidermis and cortex were removed from the main stem and the degree of vascular browning was recorded using the following scale: 0, no browning; I , slight browning at base of stem; 2, browning of lower third of stem; 3, browning of lower two-thirds of stem; 4, browning of whole stem.
Details of the method used for measuring the activity of the enzyme reducing the viscosity of solutions of pectic substances will be given later.
PATHOGENICITY EXPERIMENTS
The infection of cotton plants by the original isolate and by the hyaline variant was studied in a number of experiments, the results of which are summarized below. Both fungi rapidly infected plants when they were added as sand-maize meal cultures to soil, when they were injected as spore suspensions into stems just above soil level or when the roots of plants were dipped into spore suspensions before transplanting. Definite symptoms of disease appeared some 3-5 weeks after addition of the fungi and SO-IOO% of plants became infected. The parent strain, with abundant microsclerotia, persisted in a virulent condition in artificially con- taminated soil for a considerably longer period than did the hyaline variant which did not form microsclerotia; similar results were obtained by Isaac (1946) with the Verticillium wilt of sainfoin.
The spread of both strains was followed in plants which had become infected after their roots had been dipped in spore suspensions before they were transplanted in soil. Small fragments of plants were surface-sterilized, placed on plain agar and examined later for the pathogens. Both strains behaved similarly and were isolated from the stem 3 in. above soil level I week after inoculation, and from the apex (some 6 in. above the soil level), petiole and lamina of the last fully expanded leaf after z weeks. The fungi were distributed through infected plants well before disease symptoms visible to the naked eye had appeared.
PRODUCTION OF PECTIC ENZYMES AND TOXINS
Many studies have shown that the production of pectic enzymes by a number of fungi is greatly affected by the nutrients in the culture medium, the amount and nature of the carbon and nitrogen sources being particularly important. Enzyme production by some organisms has also been found to be adaptive in that it was stimulated by the addition of pectic materials, while other organisms produce pectic enzymes without such stimulation, i.e. in the absence of pectic materials.
In a preliminary investigation of protopectinase and toxin production by V . duhliae, a basal medium consisting of KH,P04 0-1 yo, MgS04.7H20 o*os%, KCl 0.05 yo, yeast extract 0-02%, minor elements (Fe, Cu, Zn, Mn) and CaCO, 0.75% was used with 0.276 NaNO,, or 0.16% (NH4),S04 as a nitrogen source; glucose, fructose, galactose, sucrose or cellulose at concentrations equivalent to

326 M. KAMAL A N D R. K. S. WOOD I yo glucose (for carbon) were used as sources of carbon in one series; in a second series, pectin was used either as the sole carbon source or in combination with each of the above carbon sources in turn. The media were sown with spores, and after incubation for 10 days at 25' C., cell-free filtrates were prepared; protopectinase activity was estimated with potato and cucumber disks and toxicity with cucumber disks. The results obtained may be summarized as follows:
(i) Growth occurred in all media and was between 85 mg. in the medium con- taining pectin and (NH,),SO, and 330 mg. in the medium containing fructose, pectin and NaNO,.
(ii) There were no substantial differences in the growth obtained with the different N sources.
(iii) The addition of pectin increased growth, generally by about 25 yo. (iv) The final pH value of media was always higher than the initial value which
was close to neutrality in each case; when NaNO, was used, the final values were in the range 7.8-8-4, but with (NH4),S0, the range was rather lower, 7'2-7.5.
(v) The protopectinase activity and also the toxicity of culture filtrates were considerably different and, in general, were not closely related to the amount of mycelium produced.
(vi) There was in all cases a close relation between protopectinase activity and toxicity.
(vii) More active filtrates were obtained with (NH,),SO, than with NaNO, as the N source; the addition of pectin also increased the activity of the filtrates.
Further experiments with three of the media used above showed that filtrates from I-, 2- or 3-week cultures had similar protopectinase activity and toxicity but that 4-week culture filtrates were considerably less active. The wilt-inducing power of these filtrates increased slightly with the age of the culture and was unaffected by autoclaving which almost completely destroyed protopectinase activity and toxicity. The experiments also showed that filtrates from cultures on media based on glucose, pectin and (NH,),SO, had greater protopectinase activity and toxicity but similar wilt-inducing power to those based on sucrose and NaNO,.
I n later work, the first type of medium was used for the production of enzyme solutions and the second for studies of the mechanism of wilting where it was necessary to avoid complications which might have arisen from the presence of pectin or its breakdown products.
In a final series of experiments, the effect of different pectic substances, different concentrations of pectin and different (NH,),SO,: glucose ratios on protopectinase activity and toxicity were studied. A protopectin prepared from turnip roots and containing c. 33 yo pectic substances and low and high methoxyl citrus pectins all supported good growth and gave very active filtrates when added to the basal medium containing glucose and (NH,),S04; the most active filtrates macerated potato tuber disks in 8-10 min.
The effect of different (NH,),SO,:glucose ratios in the presence of 2% low

Pectic enzymes secreted by Verticillium dahliae 327 methoxyl pectin is illustrated by the results shown in Table I. The following points emerge from these data. At each nitrogen level, increasing the carbohydrate concentration increased growth but did not always increase enzyme secretion; at high and medium nitrogen levels, the highest concentration of carbohydrate reduced activity. Again, therefore, secretion of protopectinase was not always correlated with growth.
TABLE I . Effect oj' dzgerent (NH,),SO,:gZucose ratios on enxyme production Mean R.T.
Mean dry (min.) Yo(NH4)pS04 :h glucose wt. (mg.) potato disks
0.05 0.25 270 22 0.05 1'5 410 25 0 0 5 3 '0 5 80 22
0'15 0 2 5 280 I 2 0 1 5 1'5 450 8 0.15 3 '0 610 20
0.60 0.25 270 15 060 1'5 420 8 060 3'0 5 80 20
Very active preparations were obtained with intermediate concentrations of nitrogen and carbohydrate, i.e. 0.15% (NH4) ,S04 and 1.5% glucose. These con- centrations were therefore used with 2 yo low methoxyl pectin in the basal medium for preparing relatively large quantities of active protopectinase solutions. Filtrates from cultures on this medium macerated potato disks in 8-10 min. and had a D-50 value of 20 rnin.; they were stored in 15 ml. quantities at - 16" C. until required; filtrates retained their activity for many months at this temperature.
PROPERTIES OF PROTOPECTINASE SOLUTIONS
In the experiments to be summarized in this section, the cell-free filtrates described in the last paragraph were used to determine the effect of various treatments on protopectinase activity and toxicity; because of their ready availability throughout the year, potato disks were used in most tests.
(a) Storage. Samples were stored under toluene at - 16,4, 15 and 25' C. After 5 days at 4 and C. activity was reduced by about half and at 25" C. by about two-thirds; there was little further loss of activity after storage at these tempera- tures for a further 5 days. No loss of activity was detected after storage at - 16" C.
(b) Dilution. The protopectinase activity and toxicity of solutions containing different proportions of untreated and autoclaved filtrates were determined with the results shown in Table 2. Protopectinase activity and toxicity were similarly affected by diluting, and it is clear that the relations between enzyme concentration and activity expressed as IOOO/R.T. (min.) or rooo/D50 (min.) are non-linear. Thus, diluting the standard preparation four times only led to a 50% reduction in
21 App. Riol. 44

328 M. KAMAL A N D R. K. S. WOOD protopectinase activity, while diluting it ten times gave only a 75 "/o reduction in activity. Moreover, the filtrate had to be considerably diluted before any marked reduction in activity was readily measured.
These facts must be borne in mind in the interpretation of data in which activity is expressed as the reciprocal of the R.T. or D50 values.
TABLE 2. Egect of dilution on protopectinase activity and toxicity yo untreated enzyme Protopectinase
solution activity Toxicity I 0 0
75 5 0 25
5 I 0
I 0 0 1 0 0
65 5 0 25 I 0
(c ) Temperature. Samples were brought to temperatures of 5 , 10, 15, 20, 25, 30 and 50' C. and protopectinase activity and toxicity measured. The results are given in Table 3. As was to be expected, protopectinase activity increased considerably with temperature; the increase in toxicity with temperature paralleled the increase in protopectinase activity. In view of the complexity of the reactions which are involved, no attempt will be made to interpret further these data; it will be noted, however, that the increase in activity was quite marked between 15 and 25' C. which were, approximately, the extreme temperatures in the laboratory during the year. Protopectinase activity and toxicity were therefore measured in a constant temperature room at 25' C.
TABLE 3 . Effect of temperature on protopectinase activity and toxicity Temp. (" C.) Protopectinase activity Toxicity
I 0
25 30 40 5 0 55 I 0 0
( d ) pH. Samples of enzyme solutions were brought to a series of pH values with 0-1 N - H C ~ or NaOH and then adjusted to the same final volume with water. One volume of veronal buffer (Michaelis, 1930) at an appropriate pH value was then added to four volumes of the enzyme solution; preliminary experiments had shown that at this concentration the buffer did not reduce enzyme activity. Similarly adjusted autoclaved enzyme solutions were kept as controls. The pH values were determined at the beginning and end of each test; it was found that the pH value did not change by more than 0.5 unit during the tests, and that changes of this
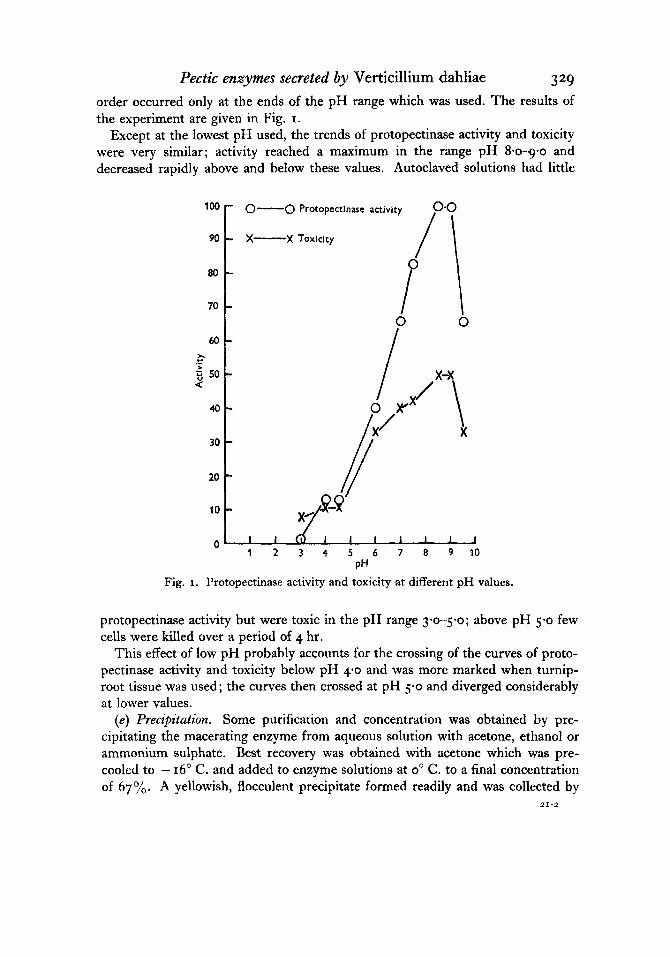
Pectic enzymes secreted by Verticillium dahliae 329 order occurred only at the ends of the pH range which was used. The results of the experiment are given in Fig. I.
Except at the lowest pH used, the trends of protopectinase activity and toxicity were very similar; activity reached a maximum in the range pH 8-0-g-o and decreased rapidly above and below these values. Autoclaved solutions had little
100
90
80
70
60 h
> Y -
50 a
40-
30
20
10
O ?
Fig.
- 0-0 Protopectinase activity
- X-X Toxicity
0
- -
0 - -
- - -
I ' 1 1 1 1 1 1 J 1 2 3 4 5 6 7 8 9 10
protopectinase activity but were toxic in the pH range 360-5.0; above pH 5.0 few cells were killed over a period of 4 hr.
This effect of low pH probably accounts for the crossing of the curves of proto- pectinase activity and toxicity below pH 40 and was more marked when turnip- root tissue was used; the curves then crossed at pH 5-0 and diverged considerably at lower values.
(e) Precipitation. Some purification and concentration was obtained by pre- cipitating the macerating enzyme from aqueous solution with acetone, ethanol or ammonium sulphate. Best recovery was obtained with acetone which was pre- cooled to - 16" C. and added to enzyme solutions at 0" C. to a final concentration of 67 yo. A yellowish, flocculent precipitate formed readily and was collected by
21-2

3 3 0 M. K A M A L AND R. K. S . WOOD centrifuging. The supernatant liquid was decanted and residual acetone removed under vacuum. The precipitate was taken up in water and the insoluble residue removed by centrifuging; the final solutions contained some 50% of the original activity. The precipitates which formed at lower concentrations of acetone were also collected and, together with the above, were tested for protopectinase activity and toxicity. No separation of the two activities was obtained. (f) Dialysis. A 20.0 ml. sample of the crude enzyme solution was placed in a
cellophane tube and dialysed in 1000 ml. of distilled water at 4" C. for 48 hr. ; the water was changed five times during the process. The p H of the enzyme solution fell from 8.2 to 5-2 during dialysis. T h e activity of the dialysed solution was tested at pH 8.0 with and without the addition of an autoclaved undialysed solution at a final concentration of 25%. There was a considerable loss of protopectinase activity and toxicity following dialysis which was, however, largely restored by the addition of the autoclaved solution. It may be inferred therefore that certain salts present in the crude solution were required for full activity; their identity was not determined.
( g ) Thermal inactivation. Small samples of a crude enzyme solution were trans- ferred to thin-walled tubes and maintained at a series of different temperatures for 5 min.; the tubes were then placed first in ice-water and then at - 16" C. until all samples were ready; protopectinase activity and toxicity were measured using separately potato and turnip disks ; essentially similar results were obtained with both types of tissue; the results with potato disks are shown in Fig. 2.
There was a steady loss of protopectinase activity as the temperature of treatment increased from 20 to 60" C., but above 60" C. there was no further loss of activity, so that even after 5 min. at 100" C. a considerable part of the original activity remained. The enzyme responsible for maceration was therefore remarkably thermostable. The toxicity of solutions followed closely that of protopectinase activity up to 40" C. At higher temperatures, the two curves diverged; above 40" C., toxicity decreased to a low value at 60" C. and remained at this value after treatment at higher temperatures. Heat treatment produced, therefore, a distinct separation of the factors responsible for maceration and killing. This effect was studied further by treating samples at IOOO C. for different periods before measuring protopectinase activity and toxicity. After 5 min. treatments, results similar to those of the previous experiments were obtained ; appreciable protopectinase activity remained even after 15 min. at 100" C., but the solutions were then only very slightly toxic. Both activities were lost in solutions kept at 100' C. for 60 min.
Similar experiments were carried out with dialysed and partially purified enzyme solutions with results agreeing with the above.
A comparison was also made between the toxicity of a heat-treated enzyme solution and diluted but untreated solutions of very similar protopectinase activity (R.T. of 35 min.); D50 values of 45 and 250 min. respectively were obtained. Disks of tissue which had macerated in a heated enzyme solution retained their suscepti-
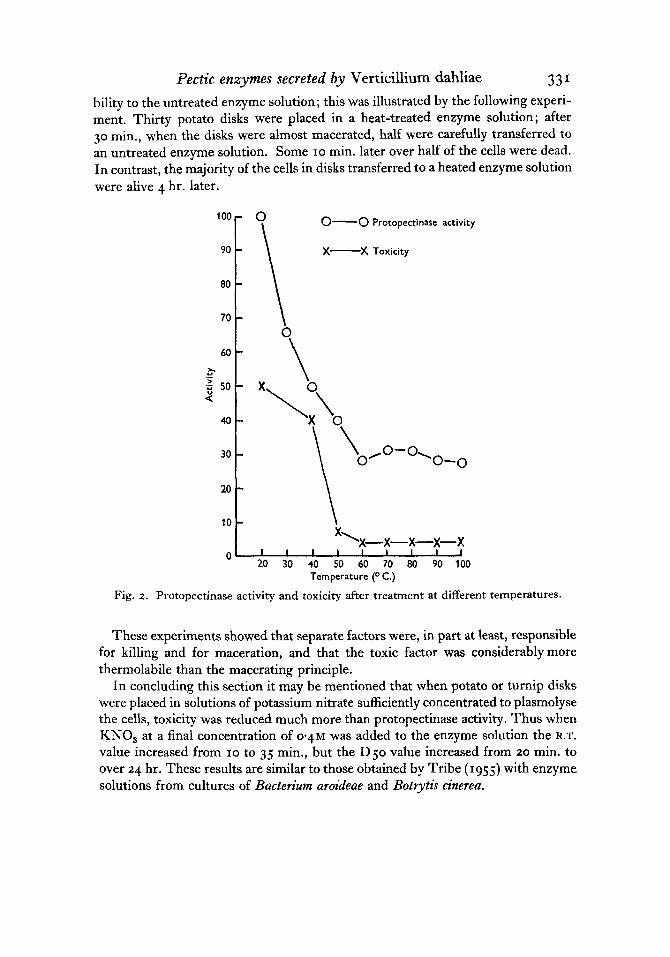
Tectic enzymes secreted by Verticillium dahliae 331 bility to the untreated enzyme solution; this was illustrated by the following experi- ment. Thirty potato disks were placed in a heat-treated enzyme solution; after 30 min., when the disks were almost macerated, half were carefully transferred to an untreated enzyme solution. Some 10 min. later over half of the cells were dead. In contrast, the majority of the cells in disks transferred to a heated enzyme solution were alive 4 hr. later.
0-0 Protopectinase activity
X-% Toxicity
80
70
60 -
40
30 - 20 - 10 -
Temperature (" C.)
Fig. 2. Protopectinase activity and toxicity after treatment at different temperatures.
These experiments showed that separate factors were, in part at least, responsible for killing and for maceration, and that the toxic factor was considerablymore thermolabile than the macerating principle.
In concluding this section it may be mentioned that when potato or turnip disks were placed in solutions of potassium nitrate sufficiently concentrated to plasmolyse the cells, toxicity was reduced much more than protopectinase activity. Thus when KNO, at a final concentration of 0.4~ was added to the enzyme solution the R.T. value increased from 10 to 35 min., but the D5o value increased from 20 min. to over 24 hr. These results are similar to those obtained by Tribe (1955) with enzyme solutions from cultures of Bacterium aroideae and Botrytis cinerea.

332 M. KAMAL AND R. K. S. WOOD
ACTION OF PROTOPECTINASE SOLUTIONS ON PECTIC SUBSTANCES
It is likely that maceration involves, inter alia, the degradation of pectic substances of the middle lamella and cell wall. The action of the protopectinase solutions used in experiments described in the preceding section on a number of pectic substances in vitro was, therefore, investigated.
Pectinesterase has recently been reported to be one of the factors responsible for the vascular browning which is a characteristic symptom of tomato wilt caused by Fusarium oxysporurn f. lycopersici (Gothoskar, Scheffer, Walker & Stahmann, 1953 ; Winstead & Walker, 1954). Vascular browning is also a feature of Verticillium wilt of cotton, so that it was of interest to determine the pectinesterase activity of proto- pectinase solutions which, it may be added here, produce this symptom in young cotton plants.
5 ml. of the protopectinase solution was added to 25 ml. of a I - ~ O / ~ solution of a high methoxyl citrus pectin (63% methoxylated by number) to which had been added sufficient N-NaOH to bring the pH of the solution to 7.5; the pH of the mixture was kept as close as possible to pH 7-5 over a period of 3 hr. by adding 0.1 N-NaOH. The experiment was repeated with an autoclaved enzyme solution. The difference in the amount of NaOH added to the first and second mixtures was taken to represent the COOH groups liberated by the action of the pectinesterase contained in the untreated protopectinase solution. The results, expressed as the percentage of the methoxyl groups initially present which were removed per hour, were as follows: crude enzyme 0.3, precipitated enzyme 0.6, dialysed enzyme 0.6. The experiment was repeated a number of times with similar results and showed that culture filtrates of high protopectinase activity and which also caused vascular browning had a very low pectinesterase activity.
The reduction of the viscosity of solutions of pectic substances has been much used for estimating the activity of pectic enzymes which attack the main structure of the molecule or the molecular aggregates which may be present when the pectic substance is in a highly polymerized state. The following substances were used for viscometric studies of the action of protopectinase solutions: (i) citrus pectin, 63 yo methoxylated, which was washed before use with acidified ethanol (95 ml. ethanol, 5 ml. conc. HCl) and then with 90% ethanol until there was no trace of HCl and finally dried at 50' C.; (ii) sodium pectate prepared by de-esterifying a 1.2% solution of (i) at pH 7.0 with pectinesterase prepared from orange peel.
Dialysed enzyme solutions were used because the Ca2+ present in undialysed solutions formed a precipitate when added to solutions of sodium pectate.
The substrates used were as follows: (i) 1.0% pectin 5.0 ml., 0.7 ml. 0.1 N-NaOH to bring the pH of the solution to 8.0 which was near the optimum for activity (see later); veronal buffer at pH 8.0, 2.0 ml., water to 9.0 ml.; (ii) 0.95% Na pectate 5-0 ml., veronal buffer at pH 7.0, the optimum for activity (see later), 2.0 ml., water to 9.0 ml.

Pectic enzymes secreted by Verticillium dahliae 333 The substrate was placed in a water-bath at 25' C., and after a few minutes
1.0 ml. of the enzyme solution was added. One minute later, 5.0 ml. of the mixture was transferred to an Ostwald viscometer at 25' C. and flow times were measured at appropriate intervals over a 2-30 min. period. Under these conditions it was sufficiently accurate to assume that the viscosity was directly proportional to the time of flow of the mixture through the viscometer. The effect of various treatments on the activity of the viscosity reducing enzymes is given below.
(Q) Dilution. Appropriate dilutions of a dialyzed enzyme solution were made by adding suitable volumes of an autoclaved solution. A series of viscosity/time curves was obtained for the substrates described above, and from these curves, the time taken for the enzyme, in different concentrations, to produce a certain change of viscosity of the mixtures was calculated. The end-point chosen was that corre- sponding to 75"/0 of the total possible loss. Activity was then expressed as the inverse of the time taken for a particular enzyme solution to produce this change and was plotted against enzyme concentration in arbitrary units. Linear curves were obtained with both substrates; it was therefore assumed that under the condi- tions described above, the activity of viscosity-reducing enzymes in enzyme- substrate mixtures similar to the above was inversely proportional to the time taken for these mixtures to show a given loss of viscosity.
(b) pH. Activity was determined over a range of pH values with both substrates; these were first brought to the required pH with 0.1 N-NaOH or HC1 and then buffered with 2.0 ml. of a Verona1 buffer at that pH. Activity was then determined in the way described above with the results shown in Table 4. The pH values of
TABLE 4 . Effect of pH on activity of viscosity reducing enzymes Enzyme activity (arbitrary units)
P H 5 '0 6.0 7'0 7'5 8.0 8.5 9.0 9'5
Pectin
7
15 26 62
I2
I 0 0 I 0 0
85
Na pectate
32 56 100
42 6 4 6
13
solutions were determined immediately before and after the viscosity measure- ments; no material changes were observed at any value within the range investi- gated. The results show that with pectin the optimum pH was in the range 8.5-9-0 with considerable activity at pH 8.0 and 9.5. With sodium pectate, however, the optimum was clearly at or near pH 7.0, and activity fell very sharply on both sides of this value; indeed, activity was lowest at pH 8-5 which was close to the optimum when pectin was the substrate. Above pH 8.5, activity increased slightly;

334 M. K A M A L A N D R. K. S. WOOD no measurements were made at higher values. This experiment was repeated a number of times with essentially similar results.
(c) Thermal inactivation. Tests were made with samples of a crude enzyme solu- tion which had been heated at IOOO C. for 5 , 10 and 15 min.; a o*5yo solution of pectin at pH 8.0 was used. The activity of the untreated enzyme solution was taken to be 100 units. The results are shown in Table 5 .
TABLE 5 . Effect of heat treatment on the activity of the viscosity reducing enzjme Treatment Activity
Untreated solution I 0 0 Solution at 100' C. for j min. Solution at 100' C. for 10 min. Solution at 100' C . for I j min. Autoclaved solution 0
I 3.0 1 r . j
1 0 . 5
Approximately 90% of the activity was lost after each of the heat treatments. The viscosity reducing enzymes therefore have a thermostability comparable with that of protopectinase.
MECHANISM OF WILTING
Comparative studies of the properties of culture fluids obtained in various ways showed that, while there were considerable differences in protopectinase activity and toxicity, the majority rapidly caused wilting of cotton shoots. Some of the factors involved in this process were investigated with filtrates from cultures on the sucrose-nitrate (SN) and glucose-(NH,),SO,-pectin (GNP) media described earlier; these had low and high protopectinase activity and toxicity respectively. The SN filtrate was used in most experiments because the uninoculated GNP medium itself caused wilting, thus making it difficult to obtain suitable controls. It was confirmed that the relatively high pH of culture filtrates (about 8.0) did not cause wilt.
Young cotton plants with the cut ends of the stems placed in a SN culture filtrate wilted irreversibly in 8-10 hr. under laboratory conditions (in front of an east-facing window at approximately 20' C.); the filtrate had to be diluted con- siderably to reduce this period substantially. Even after four-fold dilution plants wilted in under 24 hr., and after ten-fold dilution in less than 40 hr. Plants, with the cut ends of the stem in a SN culture filtrate, did not wilt even after 72 hr. when placed in the dark; the leaves remained turgid but were readily detached from the main stem. I n a saturated atmosphere, plants wilted after 54 hr. Autoclaved and untreated solutions behaved similarly in these experiments which were repeated a number of times with similar results.
In another experiment, plants were placed in SN filtrates in a saturated atmo- sphere; some were transferred after 20, 40 and 48 hr. to water under laboratory conditions. These plants wilted irreversibly 60-66 hr. from the beginning of the experiment. The plants which were left in the saturated atmosphere wilted in

Pectic enzymes secreted by Verticillium dahliae 335 55-60 hr. This indicated that in 20 hr. a sufficient quantity of the factor responsible for wilting had been taken up to cause the plants to wilt under conditions permitting loss of water.
The effect of removing parts of the stem of wilted plants was also studied. Shoots having a stem 6 cm. long were placed in SN filtrates for 16 hr.; all leaves were then completely wilted. Different lengths of stem were removed and the rest of the shoot transferred to water. The results 24 hr. later may be summarized as follows: (i) bottom I or z cm. of stem removed-no recovery; (ii) bottom 3, 4 or 5 cm. of stem removed-basal portion of leaves regained turgidity; (iii) leaves detached and placed in water regained full turgidity. This suggested that wilting was caused by a blocking of the vascular system of the main stem. The water loss from shoots of 4-week-old plants placed in SN culture filtrates or in SN uninoculated media was therefore followed gravimetrically.
TABLE 6. Water loss from plants in culture solutions Total loss of weight
b i g . dry wt.) Loss of fresh Water Wilting wt. (mg.) uptake time
Treatment 1st 6 hr. 2nd 6 hr. 12 hr. (mg.1 (hr.) SN culture filtrate 5’3 2.9 240 360 7-8 SN medium 5’5 5 ’4 40 800 0
S N medium 3 4 2.8 480 0 3-1 (uninoculated)
(uninoculated) Blocked ‘Vaseline’
Plants were transferred from pots and the stems cut under water. Ten were placed in glass vials (3 x 4 in. diameter) containing SN culture filtrate, ten were placed in SN uninoculated medium as were another ten the stem ends of which had been sealed with ‘Vaseline’. The vials were weighed at the beginning of the experi- ment, 6 and 12 hr. later. The fresh weights of the plants were determined at the beginning of the experiment and 12 hr. later and the final dry weight of the plants determined at the end of the experiment by drying at 70’ C. for 24 hr. The results obtained are summarized in Table 6. The figures in the third column represent, essentially, the loss of water from the plants themselves which was negligible in the controls and twice as high in sealed plants as in those placed in the culture filtrate. The values in columns I and 2 give a measure of the total loss of water through transpiration and from the plant as a proportion of the dry weight of the plant. After 6 hr. this loss was much lower from sealed plants than from the others. During a second 6 hr. the losses from sealed plants and plants in untreated filtrates were very similar and only about half of those from the control plants. These results indicate that the effect of the culture filtrate over a period of 12 hr. was to reduce the water uptake to a value intermediate between that of the control and sealed plants. This is borne out by the figures shown in column 4 which show that

336 M. KAMAL A N D R. K. S. WOOD the water uptake by the control plants was twice that of the plants placed in culture filtrates.
'Vaseline'-treated shoots wilted quite rapidly, followed some 3-4 hr. later by the shoots in culture filtrates; the control plants did not wilt during the experiment.
Plants which had wilted in the above experiments did not show certain other symptoms which appear in naturally infected plants, particularly browning of the vascular system. In a number of preliminary experiments the ability of various solutions to produce vascular browning was studied. Untreated and autoclaved filtrates from cultures on sucrose-nitrate media were tested at a number of dilutions. The time taken for shoots to wilt irreversibly increased from 10 to 48 hr. as the concentration of the filtrate was reduced from IOO to 5 % ; autoclaved and untreated solutions behaved similarly. There was a little vascular browning ( I on the scale given earlier) in the untreated filtrates and none in autoclaved filtrates. In contrast, when filtrates from cultures on glucose-ammonium sulphate-pectin media were used, vascular browning was pronounced (scale reading 3-4) in unheated filtrates even after a ten-fold dilution but vascular browning was virtually absent in the autoclaved filtrates. Both untreated and autoclaved filtrates had similar wilting activity which was somewhat higher than that of the SN culture filtrates.
Both SN and GNP cultures behaved in the way described above after dialysis for 48 hr. at 4' C.
It was reported by Winstead & Walker (1954) that in Fusarium wilt of tomato, the vascular browning of culture filtrates was always associated with pectinesterase activity. It has already been stated that filtrates from cultures of V . dahliae had little pectinesterase activity; nevertheless, some of these filtrates were able to produce conspicuous browning. I t was therefore of interest to see if filtrates from cultures of other pathogens were able to produce vascular browning. The following were used : cell-free preparations from Bacterium aroideae or Sclerotium rolfsii cultures with an R.T. of 25 and 10 min. respectively; GNP filtrates of V. dahliae and a pectinesterase preparation from orange peel. The pectinesterase activity of each solution was estimated by measuring the acid liberated from a I "/o pectin solution. The pH of the enzyme/pectin mixture was brought to the value shown in the table and kept as close as possible to this value by the periodic addition of alkali. Activity is expressed as ml. 0.1 N-NaOH/hr./So ml. I % pectin solution. The results of the experiment are given in Table 7. The results showed quite clearly that the ability
TABLE 7. Activity of dzflerent enzyme solutions Proto-
pectinase Pectinesterase activity Vascular Wilting
Enzyme solution PH activity (R.T. min.) browning time (hr.) Bacterium aroideae 8.0 0 25 I 16 Sclerotium rolfsii 3'5 0 I 1 4 8 Verticillium dahliae 8.0 0.04 15 4 8 Orange peel 7'0 4'4 < 1440 2 6

Pectic enzymes secreted by Verticillium dahliae 337 of a solution to cause vascular browning was not related to high pectinesterase activity. This is best illustrated by the results obtained with the Sclerotium rolfsi filtrate and the orange-peel preparation; the former had no pectinesterase activity but caused much more vascular browning than the latter which had a high pectinesterase activity.
This experiment was repeated three times with similar results. The effect of GNP filtrates on shoots of varieties susceptible and resistant to
infection by V . dahliae was similar; shoots from both plants wilted irreversibly in 8 hr. and showed pronounced vascular browning.
DISCUSSION The pectic enzymes produced by species of Verticillium have been little investigated in the past. In an early paper, Bewley (1922) stated that culture filtrates of V. albo- atrum had no cytase (i.e. protopectinase) activity, but as the ends of tomato shoots were stated to be softened after immersion in some filtrates which had been heated at 100' C. it seems that these filtrates might have had some protopectinase activity. The present study has shown that the secretion of pectic enzymes by V . dahliae depended, to a large extent, on the presence in the media of pectic substances. Protopectinase was secreted in response to a number of fairly well-defined pectic substances; this provides circumstantial evidence that the substances degraded during maceration are similar to those which were added to. media to stimulate production of the macerating enzymes.
In view of the importance ascribed to toxins in certain other wilt diseases, the toxicity of filtrates from cultures of V . dahliae was studied in some detail. Cells of tissues were considered to have been killed by culture filtrates when they had lost the ability to plasmolyse; this is a generally accepted criterion of death. The toxicity of filtrates derived or treated in various ways was, in general, closely related to their protopectinase activity; this result was also obtained by Brown ( I ~ I S ) , Tribe (1955) and Fushtey (1953) and supports the view that the enzymes responsible for maceration also kill cells, directly or indirectly. Toxicity and macerating activity could, however, be separated in two ways. The first, which produced the greatest separation, was originally noted by Tribe (1955), who found that plasmolysis of a tissue only slightly increased its resistance to maceration but greatly increased its resistance to killing. In the second treatment filtrates were kept at temperatures within the range 60--100~ C. for 5 min. or longer; toxicity was then reduced much more than macerating activity. This effect needs further study in view of the fact that it does not occur with Bacterium aroideae and Botrytis n'nerea preparations which otherwise behave similarly to V. dahliae filtrates. The results with the latter suggest that more than one enzyme is responsible for macera- tion and killing of cells, and in this connexion the behaviour of the enzymes which reduce the viscosity of solutions of pectic substances is, perhaps, significant.

338 M. K A M A L A N D R. K. S. WOOD Different pH optima were obtained when pectin and sodium pectate were used as substrates; an explanation of this result is that these substances are hydrolysed by different enzymes. Similar results have been obtained by Ayres, Dingle, Phipps, Reid & Solomons (1952) with preparations from cultures of Aspergillus foetidus. It is also now established that some organisms produce enzymes which degrade pectin more rapidly than pectates (Seegmiller & Jansen, 1952; Gupta, 1953), while others do just the reverse (Jansen & MacDonnell, 1945). If V . dahliae produces more than one pectic enzyme, it may well be that these enzymes play different parts in maceration and killing of cells and differ in their thermolability.
Bewley (1922) showed that filtrates from cultures of If. albo-atrum produced many of the symptoms observed in tomato plants infected with the pathogen. In the present investigation, the symptoms produced in cotton shoots differed when different culture media were used. Filtrates from cultures on media not containing pectic substances readily caused shoots to wilt irreversibly, and this activity was unaffected by autoclaving or dialysis ; these solutions were relatively non-toxic to cells of potato tubers. A more complete set of symptoms which included vascular browning was obtained when cotton shoots were treated with filtrates from cultures on a medium containing pectin. These solutions readily macerated potato-tuber tissue and were highly toxic, but these properties and the ability to cause vascular browning were much reduced after autoclaving ; in contrast, the wilting activity was little affected by this treatment. These results do not agree with those reported recently by Green (1954), who studied the effect on tomato shoots of filtrates from cultures of V. albo-atrum on Czapek-Dox solution. I t was shown that proteinaceous material was responsible for host responses other than vascular discoloration, and that this fraction was not present in cultures until autolysis had begun; on removal of the protein, solutions caused an atypical vascular discoloration but shoots did not wilt. With V. dahliae, however, wilting was caused by thermostable substances of high molecular weight, while vascular browning was associated with thermolabile substances, presumably proteins. Whether or not vascular browning is caused by pectic or other enzymes remains to be determined.
Some evidence was obtained which suggested that wilting of shoots placed in culture filtrates was caused by the uptake of substances which restricted the flow of the vascular solution. This effect could be produced by the uptake of compounds of relatively high molecular weight already present in the culture filtrate (cf. Hodgson, Peterson & Riker, 1949) or by the action of enzymes on the vascular elements. Recently, it has been claimed that pectinesterase plays an important part in the production of disease symptoms in tomato plants by filtrates from cultures of Fusarium oxysporum f. lycopersici (Gothoskar et al. 1953 ; Winstead 8t Walker, 1954). This enzyme can have played only a minor role in the production ofdisease symptoms in cotton shoots because V. dahliae culture filtrates had little or no pectinesterase activity. Moreover, I/. dahliae culture filtrates produced much more vascular browning than a preparation from orange peel having high pectinesterase

Pectic enzymes secreted by Verticillium dahliae 339 activity. It is, in fact, not easy to see how the action of pectinesterase alone could produce disease symptoms ; the action of this enzyme on pectic materials contained in the walls of vascular elements is likely to, make them even less soluble and less likely to be released into the vascular stream. The combined action of a pectic enzyme able to hydrolyse compounds of high molecular weight, e.g. polygalacturo- nase, and pectinesterase might, however, lead to the formation of insoluble pectinates or pectates which, if persisting, might impede the flow of the vascular solution. These precipitates are likely to be composed of units of relatively low molecular weight because it seems probable that the familiar pectinate and pectate gels would be slowly broken down by polygalacturonase-type enzymes which might be present. It is of course also possible that the action of one or more pectic enzymes on cell walls might release substances which act as substrates for other enzymes produced by the pathogen in vivo and that it is the end-products of these reactions which are the immediate cause of wilt symptoms.
The work described above in which culture filtrates have been used can do no more than suggest how pathogens living in vascular systems of plants produce symptoms of disease. Ultimately, any activity of a filtrate which is related to a disease symptom must be demonstrated in a diseased plant and, in the case of wilt diseases, this is likely to be a technically difficult matter. However, a start has been made by Waggoner & Dimond (1954), who have demonstrated that exudates from the stems of tomato plants infected with Fusarium oxysporum f. lycopersici have some pectinesterase and polygalacturonase activity.
REFERENCES
AYRES, A., DINGLE, J., PHIPPS, A,, REID, W. W. & SOLOMONS, S. L. (1952). Enzymic degradation of pectic acid and the complex nature of polygalacturonase. Nature, Lond., 170, 834.
BEWLEY, W. F. (1922). 'Sleepy disease' of the tomato. Ann. appl. Biol. 9, 116-34. BROWN, W. (1915). Studies in the physiology of parasitism. I . The action of Botrytis
cinerea. Ann. Bot. , Lond., 29, 313-48. FLJSHTEY, S. G. (1953). Studies in the cell-killing action of extracts derived from various
fungi and bacteria. Ph.D. Thesis, University of London. GREEN, R. J. (1954). A preliminary investigation of toxins produced in vitro by Verticillium
albo-atrum. Phytopathology, 4, 433-7. GOTHOSKAR, S. S., SCHEFFER, R. P., WALKER, J. c. & STAHMANN, M. A. (1953). The role of
pectic enzymes in Fusarium wilt of tomato. Phytopathology, 43, 535. GUPTA, S. C. (1953). The production and properties of pectolytic enzymes secreted by
Pythium debaryanum Hesse. Ph.D. Thesis, University of London. HODGSON, R., PETERSON, W. H. & RIKER, A. J. (1949). The toxicity of polysaccharides and
other large molecules to tomato cuttings. Phytopathology, 39, 47-62. ISAAC, I. (1946). Verticillium wilt of sainfoin. Ann. appl. Biol. 33, 28-34. JANSEN, E. F. & MACDONNELL, L. R. (1945). The influence of methoxyl content of pectic
MICHAELIS, I. (1930). Diethylbarbiturate buffer. J. biol. Chem. 87, 33-5. SEEGMILLER, C. G. & JANSEN, E. F. (1952). Polymethylgalacturonase, an enzyme causing
substances on the action of polygalacturonase. Arch. Biochem. 8, 97-1 I 2.
the glycosidic hydrolysis of esterified pectic substances. J . biol. C h m . 195, 327-36.

340 M. KAMAL AND R. K. S. WOOD TRIBE, H. T. (1955). Studies in the physiology of parasitism. XIX. On the killing of plant
cells by enzymes from Botrytis cinerea and Bacterium aroideae. Ann. Bat., Lond., N.S.,
WAGGONER, P. E. & DIMOND, A. E. (1954). Pectic enzymes produced by Fusarium oxy-
WINSTEAD, N. N. & WALKER, J . C . (1954). Production of vascular browning by metabolites
19, 351-68.
sporum f. lycopersici in infected plants. Phytopathology, 4, 509.
from several pathogens. Phytopathology, 44, I 53-8.
(Received 5 November 1955)




















