
Elite Upland Cotton Germplasm-Pool Assessment of Fusarium Wilt Resistance in California
10
Agronomy Journal • Volume 105, Issue 6 • 2013 1635 Agronomy, Soils & Environmental Quality Elite Upland Cotton Germplasm-Pool Assessment of Fusarium Wilt Resistance in California Robert B. Hutmacher, Mauricio Ulloa,* Steven D. Wright, B. Todd Campbell, Richard Percy, Ted Wallace, Gerald Myers, Fred Bourland, David Weaver, Peng Chee, Peggy Thaxton, Jinfa Zhang, Wayne Smith, Jane Dever, Vasu Kuraparthy, Daryl Bowman, Don Jones, and John Burke Published in Agron. J. 105:1635–1644 (2013) doi:10.2134/agronj2013.0264 Copyright © 2013 by the American Society of Agronomy, 5585 Guilford Road, Madison, WI 53711. All rights reserved. No part of this periodical may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. C otton is an economically important crop and provides the world’s leading natural fiber for the textile industry (Smith et al., 1999; Ulloa et al., 2007). e future of cotton improvement depends on plant characteristics confer- ring not only improved water-use efficiency and heat tolerance but also disease resistance. Integrating disease resistance into high-yielding, high-fiber-quality cultivars is one of the most important objectives in cotton breeding programs (Smith et al., 1999; Ulloa et al., 2009). Host-plant resistance can be a highly effective and economical approach for dealing with disease threats such as FOV and limiting yield loss in cotton (Hutm- acher et al., 2011; Ulloa et al., 2010, 2011, 2013). Eight genotypes of FOV, called races, have been described (Skovgaard et al., 2001; Kim et al., 2005). Before 1986, only Races 1 and 2 were known to occur on significant acreage in the United States (DeVay, 1986). Since FOV disease in cotton was first reported in California in 1959 (Garber and Paxman, 1963), FOV Races 1 or 2 were recognized as the causal organism and were typically found in sandy soils with root-knot nematode (RKN) [Meloidogyne incognita (Kofoid and White) Chitwood] (Garber et al., 1979; Veech, 1984; Bell, 1984). e FOV fungal pathogen is a soil inhabitant that can survive for long periods in the absence of a host. Consequently, it is nearly impossible to eradicate from a field. A marked increased in the susceptibility of cultivars to FOV Race 1 was noted in the presence of RKN (Garber et al., 1979). Cotton developed for resistance to FOV on soils infested with RKN usually maintained their resistance when simultaneously challenged by both organisms, FOV Race 1 and RKN (Sasser, 1972; Heald and Orr, 1982). In 2005, University of California-Davis scientists (Kim et al., 2005) identified additional FOV races (1, 3, 4, and 8) in California soils. During the past 9 yr, FOV Race 4 has increas- ingly impacted cotton fields in the San Joaquin Valley (SJV) of California (Hutmacher et al., 2011). In field soils with rela- tively high levels of FOV Race 4, extensive disease symptoms ABSTRACT During the past 9 yr, a new race of Fusarium (Fusarium oxysporum f. sp. vasinfectum [FOV Race 4]) has increasingly impacted cot- ton ( Gossypium spp.) in the San Joaquin Valley of California. To assess the vulnerability of upland cotton ( G. hirsutum L.) in Cali- fornia to FOV disease, elite upland germplasm lines from 13 U.S. public breeding programs across the Cotton Belt and commercial cultivars were evaluated for disease resistance to FOV Races 1 and 4. Ten independent replicated field trials were conducted: three in 2008, four in 2010, and three in 2011. Significant differences (P £ 0.05) were observed for disease severity index of leaves, vascular root staining, and plant survival values among the elite germplasm lines in all 3 yr for the levels of resistance–response to FOV Races 1 and 4. Also, significant interactions among germplasm lines, FOV races (1 and 4), and evaluation sites indicated that germplasm lines differed in mechanisms of plant-defense response for the two FOV races. Selected lines from programs in the states of Alabama, Arkansas, Louisiana, and Mississippi showed at least a moderate level of tolerance to both FOV races; however, several of these lines produced weak and coarse fibers. Based on these evaluations, many of the entries in public breeders’ current elite upland germplasm pools may be more susceptible than expected to some FOV races, and sources of acceptable levels of resistance may be limited when tested under infestation levels that resulted in only 5 to 35% plant survival in susceptible check cultivars. R.B. Hutmacher, Dep. of Plant Science, Univ. of California-Davis, West Side Research and Extension Center, Five Points, CA 93624; M. Ulloa and J. Burke, USDA-ARS, Cropping Systems Research Lab., Plant Stress and Germplasm Development Unit, Lubbock, TX 79415; S.D. Wright, Univ. of California, Tulare, CA 93274; B.T. Campbell, USDA-ARS, Coastal Plains Soil and Water Conservation Research Center, Florence, SC 29501; R. Percy, USDA-ARS, Southern Plains Agricultural Research Center, Crop Germplasm Unit, College Station, TX 77845; T. Wallace, Mississippi State Univ., MS 39762; G. Myers, Agriculture Center, Louisiana State Univ., Baton Rouge, LA 70803; F. Bourland, Northeast Research and Extension Center, Univ. of Arkansas, Keiser, AR 72351; D. Weaver, Dep. of Agronomy and Soils, Auburn Univ., Auburn, AL 36849; P. Chee, Univ. of Georgia, National Environmentally Sound Production Agriculture Laboratory, Tiſton, GA 31793; P. axton, Delta Research and Extension Center, Stoneville, MS 38776; J. Zhang, Dep. of Plant and Environmental Sciences, New Mexico State Univ., Las Cruces, NM 88003; W. Smith, Dep. Crop and Soil Sciences, Texas A&M Univ., College Station, TX 77843; J. Dever, Texas A&M AgriLife Research and Extension Center, Lubbock, TX 79403; V. Kuraparthy and D. Bowman, Dep. of Crop Science, North Carolina State Univ., Raleigh, NC 27695; and D. Jones, Cotton Incorporated, Cary, NC 27513. Received 3 June 2013. *Corresponding author ([email protected]). Abbreviations: DSI, disease severity index of leaves; FOV, Fusarium wilt; PS, plant survival; RBTN, Regional Breeders Testing Network; RKN, root- knot nematode; SJV, San Joaquin Valley; VRS, disease index of vascular root staining. Published September 6, 2013
-
Upload
lsuagcenter -
Category
Documents
-
view
0 -
download
0
Transcript of Elite Upland Cotton Germplasm-Pool Assessment of Fusarium Wilt Resistance in California

Agronomy Journa l • Volume 105 , I s sue 6 • 2013 1635
Agronomy, Soils & Environmental Quality
Elite Upland Cotton Germplasm-Pool Assessment of Fusarium Wilt Resistance in California
Robert B. Hutmacher, Mauricio Ulloa,* Steven D. Wright, B. Todd Campbell, Richard Percy, Ted Wallace, Gerald Myers, Fred Bourland, David Weaver, Peng Chee, Peggy Thaxton, Jinfa Zhang, Wayne Smith, Jane Dever, Vasu Kuraparthy, Daryl Bowman, Don Jones, and John Burke
Published in Agron. J. 105:1635–1644 (2013)doi:10.2134/agronj2013.0264Copyright © 2013 by the American Society of Agronomy, 5585 Guilford Road, Madison, WI 53711. All rights reserved. No part of this periodical may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher.
Cotton is an economically important crop and provides the world’s leading natural fiber for the textile
industry (Smith et al., 1999; Ulloa et al., 2007). The future of cotton improvement depends on plant characteristics confer-ring not only improved water-use efficiency and heat tolerance but also disease resistance. Integrating disease resistance into high-yielding, high-fiber-quality cultivars is one of the most important objectives in cotton breeding programs (Smith et al.,
1999; Ulloa et al., 2009). Host-plant resistance can be a highly effective and economical approach for dealing with disease threats such as FOV and limiting yield loss in cotton (Hutm-acher et al., 2011; Ulloa et al., 2010, 2011, 2013).
Eight genotypes of FOV, called races, have been described (Skovgaard et al., 2001; Kim et al., 2005). Before 1986, only Races 1 and 2 were known to occur on significant acreage in the United States (DeVay, 1986). Since FOV disease in cotton was first reported in California in 1959 (Garber and Paxman, 1963), FOV Races 1 or 2 were recognized as the causal organism and were typically found in sandy soils with root-knot nematode (RKN) [Meloidogyne incognita (Kofoid and White) Chitwood] (Garber et al., 1979; Veech, 1984; Bell, 1984). The FOV fungal pathogen is a soil inhabitant that can survive for long periods in the absence of a host. Consequently, it is nearly impossible to eradicate from a field. A marked increased in the susceptibility of cultivars to FOV Race 1 was noted in the presence of RKN (Garber et al., 1979). Cotton developed for resistance to FOV on soils infested with RKN usually maintained their resistance when simultaneously challenged by both organisms, FOV Race 1 and RKN (Sasser, 1972; Heald and Orr, 1982).
In 2005, University of California-Davis scientists (Kim et al., 2005) identified additional FOV races (1, 3, 4, and 8) in California soils. During the past 9 yr, FOV Race 4 has increas-ingly impacted cotton fields in the San Joaquin Valley (SJV) of California (Hutmacher et al., 2011). In field soils with rela-tively high levels of FOV Race 4, extensive disease symptoms
AbStrActDuring the past 9 yr, a new race of Fusarium (Fusarium oxysporum f. sp. vasinfectum [FOV Race 4]) has increasingly impacted cot-ton (Gossypium spp.) in the San Joaquin Valley of California. To assess the vulnerability of upland cotton (G. hirsutum L.) in Cali-fornia to FOV disease, elite upland germplasm lines from 13 U.S. public breeding programs across the Cotton Belt and commercial cultivars were evaluated for disease resistance to FOV Races 1 and 4. Ten independent replicated field trials were conducted: three in 2008, four in 2010, and three in 2011. Significant differences (P £ 0.05) were observed for disease severity index of leaves, vascular root staining, and plant survival values among the elite germplasm lines in all 3 yr for the levels of resistance–response to FOV Races 1 and 4. Also, significant interactions among germplasm lines, FOV races (1 and 4), and evaluation sites indicated that germplasm lines differed in mechanisms of plant-defense response for the two FOV races. Selected lines from programs in the states of Alabama, Arkansas, Louisiana, and Mississippi showed at least a moderate level of tolerance to both FOV races; however, several of these lines produced weak and coarse fibers. Based on these evaluations, many of the entries in public breeders’ current elite upland germplasm pools may be more susceptible than expected to some FOV races, and sources of acceptable levels of resistance may be limited when tested under infestation levels that resulted in only 5 to 35% plant survival in susceptible check cultivars.
R.B. Hutmacher, Dep. of Plant Science, Univ. of California-Davis, West Side Research and Extension Center, Five Points, CA 93624; M. Ulloa and J. Burke, USDA-ARS, Cropping Systems Research Lab., Plant Stress and Germplasm Development Unit, Lubbock, TX 79415; S.D. Wright, Univ. of California, Tulare, CA 93274; B.T. Campbell, USDA-ARS, Coastal Plains Soil and Water Conservation Research Center, Florence, SC 29501; R. Percy, USDA-ARS, Southern Plains Agricultural Research Center, Crop Germplasm Unit, College Station, TX 77845; T. Wallace, Mississippi State Univ., MS 39762; G. Myers, Agriculture Center, Louisiana State Univ., Baton Rouge, LA 70803; F. Bourland, Northeast Research and Extension Center, Univ. of Arkansas, Keiser, AR 72351; D. Weaver, Dep. of Agronomy and Soils, Auburn Univ., Auburn, AL 36849; P. Chee, Univ. of Georgia, National Environmentally Sound Production Agriculture Laboratory, Tifton, GA 31793; P. Thaxton, Delta Research and Extension Center, Stoneville, MS 38776; J. Zhang, Dep. of Plant and Environmental Sciences, New Mexico State Univ., Las Cruces, NM 88003; W. Smith, Dep. Crop and Soil Sciences, Texas A&M Univ., College Station, TX 77843; J. Dever, Texas A&M AgriLife Research and Extension Center, Lubbock, TX 79403; V. Kuraparthy and D. Bowman, Dep. of Crop Science, North Carolina State Univ., Raleigh, NC 27695; and D. Jones, Cotton Incorporated, Cary, NC 27513. Received 3 June 2013. *Corresponding author ([email protected]).
Abbreviations: DSI, disease severity index of leaves; FOV, Fusarium wilt; PS, plant survival; RBTN, Regional Breeders Testing Network; RKN, root-knot nematode; SJV, San Joaquin Valley; VRS, disease index of vascular root staining.
Published September 6, 2013

1636 Agronomy Journa l • Volume 105, Issue 6 • 2013
and, in some cases, large reductions in plant survival were observed in clay loam and loam soils even when RKN popula-tions and root damage from nematodes were nonexistent or extremely low. Major plant damage and losses with FOV Race 4 in the absence of RKN is in marked contrast to observations with FOV Race 1. Acala and non-Acala upland germplasm tested for susceptibility to FOV Race 4 were infected by FOV Race 4 at field sites or in greenhouse artificial inoculation tri-als, showing mild to severe symptoms (Hutmacher et al., 2011; Ulloa et al., 2006, 2010, 2013).
The four FOV races identified in California cotton by Uni-versity of California-Davis scientists (Kim et al., 2005) were originally classified on the basis of pathogenicity tests on dif-ferent cotton species—G. hirsutum, G. barbadense L., and G. arboreum L. (Armstrong and Armstrong, 1958, 1960, 1978; Ibrahim, 1966)—and by their pathogenicity on alfalfa (Medi-cago sativa), soybean [Glycine max (L.) Merr.], and tobacco (Nicotiana tabacum L.) (Armstrong and Armstrong, 1978). Presently, these races are categorized according to a number of genetic markers. Based on sequence differences in the transla-tional elongation factor (EF-1a), phosphate permase (PHO), and b-tubulin (BT) genes and intergenic spacer (IGS) regions with restriction enzyme digests, worldwide strains of FOV can be classified into five major lineages (Kim et al., 2005). Recently, analyses with partial sequences from the above genes of 61 FOV isolates across the United States (Holmes et al., 2009) revealed four additional FOV genotypes, and for the first time, Races 3 and 8 were reported outside of California. The majority of these novel genotypes were from the state of Arkan-sas and were pathogenic to at least one cotton cultivar used in the study (Holmes et al., 2009).
For breeders, it is important to understand the genetic inheritance of FOV based on host-plant resistance (Ulloa et al., 2013). In early genetic analyses, FOV resistance was reported to be determined by one or two major genes with complete to incomplete dominance and possibly additional minor genes (Mohamed, 1963; Smith and Dick, 1960; Ulloa et al., 2006). In recent genetic studies (Wang et al., 2009; Ulloa et al., 2011; Becerra et al., 2012), resistance to Races 1, 7, and the Australian FOV races was reported to be inherited through gene interac-tions detected in more than one chromosome. In addition, a major dominant gene was identified that conferred resistance to FOV Race 4 in the Pima S-6 germplasm line. The Fov4 gene appears to be located near a genome region on chromosome 14 marked with a quantitative trait locus Fov41-C14 and made the biggest contribution to the FOV Race 4 resistance of the gener-ated F2 progeny (Ulloa et al., 2013). Collectively, these studies (Wang et al., 2009; Ulloa et al., 2011, 2013; Becerra et al., 2012) suggest a different gene specificity of FOV resistance in cotton (Fov1 [Race 1], chromosome 16; Fov4 [Race 4], chromosome 14; FWR [Race 7], chromosome 17; and Australian race Fov [AUS], chromosomes 6, 22, and 25) (Ulloa et al., 2013).
A key component to improving breeding lines and germplasm is the ability to test their improvements in an extensive network of sites. The Regional Breeders Testing Network (RBTN) is a multienvironment trial (MET) supported by Cotton Incor-porated (www.cottonrbtn.com or www.cottoninc.com/fiber/AgriculturalDisciplines/Variety-Improvement/RBTN/). Uni-versity and USDA-ARS cotton geneticists and breeders conduct
a cooperative MET across the major cotton growing regions of the United States. At these MET sites, elite upland germplasm lines are tested for yield and fiber quality performance, biotic and abiotic stresses such as nematodes, tarnished plant bug, heat, and FOV and Verticillium wilt (Verticillium dahlia Kleb) resistance. In 2008, 2010, and 2011, the USDA-ARS Shafter Cotton Research Station and the University of California were involved in FOV host-plant resistance trials, evaluating advanced breeding lines from the RBTN group.
Observations reported in recent years (Holmes et al., 2009; Kim et al., 2005; Kochman et al., 2002; Ulloa et al., 2006; Wang et al., 2004) have confirmed that FOV is becoming a recurring and potentially expanding threat to cotton production. Recent analyses with partial sequences of marker genes of 61 FOV isolates across the United States revealed four additional new FOV genotypes. The majority of these novel genotypes were from Arkansas and were pathogenic to at least one tested cot-ton cultivar used in the study (Holmes et al., 2009; Davis and Nichols, personal communication, 2010). In addition, little is known about resistance in U.S. upland and wild species of cot-ton to these FOV races or how soil type, soil pH, natural antago-nists, and interactions with other pathogens (e.g., Verticillium and reniform nematode [Rotylenchulus reniformis] Linford and Oliveira) could affect this pathogen. Their complete host range is not well understood. Based on our current knowledge of the eco-logical factors that favor the strains of FOV, cotton production in broad areas of the United States could be at risk.
The objective of the research reported here was to assess the vulnerability of upland cotton in the SJV of California to FOV disease through a series of field trial evaluations conducted at field sites infested with FOV Race 1 and RKN, and at separate sites found to be infested with FOV Race 4, in both cases using elite upland germplasm lines from 13 U.S. public breeding pro-grams across the Cotton Belt and commercial cultivars. Results from our studies can be utilized in further genetic evaluations to identify sources of host plant resistance useful to breeders and of potential economic value to growers. Long-term man-agement of Fusarium wilt in cotton will be enhanced by the development of resistant cultivars.
MAtEriAlS And MEthodSGermplasm and cultivars
Elite upland germplasm lines from the RBTN representing 13 public breeding/genetics programs from 10 states and check cultivars were used in this study (Table 1). The elite upland germplasm lines from the RBTN were evaluated for resistance to FOV Race 1 in the presence of RKN, for FOV Race 4 resis-tance, and for yield potential and fiber quality in 2008, 2010, and 2011 (Table 2). Upland and Pima (G. barbadense) cotton cultivars from private seed companies were included as addi-tional comparison genotypes (Tables 1 and 2). Cultivars DP 393 (Monsanto Co., PVP no. 200400266), Fiber Max (FM) 958, (Bayer CropScience Co., PVP no. 200100208), SureGrow (SG) 105 (SureGrow Co., PVP no. 9900190), and Phytogen 72 (Phytogen Seed Co., PVP no. 200100115) cultivars were used as checks for yield and fiber quality characteristics. Upland cul-tivar Shorty (Ulloa et al., 2013) was used as a susceptible check for both races (1 and 4). Pima S-7 (Turcotte et al., 1992), Pima 3-79 (GB 1585), and Phytogen 72 were used as resistant checks

Agronomy Journa l • Volume 105, Issue 6 • 2013 1637
table 1. Elite germplasm lines from the regional breeders testing network representing 13 public breeding/genetics programs from 10 U.S. states.
U.S. state institution breeder 2008 2010 2011 Alabama Auburn Univ. Weaver AU-5491 AU3202 AU3202
AU-5367 AU6001 AU3111AU-6103 AU3111 AU3223AU-1107 AU6202 AU3095
Arkansas Univ. of Arkansas Bourland Ark0012-03-08 Ark 0219-15 Ark 0305-07Ark0015-06-11 Ark 0203-11 Ark M222-07Ark0002-03-02 Ark 0222-12 Ark 0316-36Ark0001-01-03 Ark 0232-24 Ark 0309-31Ark008-22-10 Ark 0304-23
Georgia Univ. of Georgia Chee GA2004230 GA 2004143 GA 2004143GA2004089 GA 2006106 GA 2008083GA2004303 GA 2006053 GA 2008057GA2006128 GA 2007095
Louisiana Louisiana State Univ.
Myers LA05307029 LA06307025 LA06307025LA05307061 LA07307111 LA07307106LA05307083 LA07307122 LA00405034LA05307094 LA07307106
Mississippi USDA-ARS Meredith MD25ne MD25-26neMD25Y MD 25-87Y
MD 25-27YMississippi State Univ.
Wallace 0149-17 ne 0033-60147-22 ne0020-31 ne0028-16 ne
8921-2-2-14-13-11Thaxton 04PST-250
04-PST-27504PST-246
N orth Carolina North Carolina State Univ.
Kuraparthy NC09AZ09 NC08AZ21NC05AZ06
N ew Mexico New Mexico State Univ.
Zhang Acala 1517-99 NM W1218 NM08N1562NM 03012 NM 06N1166 NM08N1564
NM 03012 NM08N1084NM 06N1104 Acala 1517-08
Acala 1517-99
S outh Carolina USDA-ARS Campbell PD99041 PX03201-66-1 PD 05069PD04012 PD05041 PD 05070PD03025 PD05035 PD 06001PD03011 PX03202-65-1
Texas Texas A&M Univ. Dever (seed provided) LBB-07-21-311Smith TAM B182-34 TAM 03WZ-37 Tamcot 73
United States private seed company
obsolete cultivars
DP 393 DP 393 DP 393FM 958 FM 958 FM 958SG 105 SG 105 SG 105
Phytogen Co. Phytogen 72 Phytogen 72 Phytogen 72Phytogen 800
California USDA-ARS USDA-ARS Pima S-7 Pima S-7 Pima S-7Pima 3-79 Pima S-6
TM-1Ulloa SJ-07P-FR01 TM-1 SJ-10P10
SJ-07P-FR02 Pima S-6 SJ-10P13SJ-07P-FR03 Upland Shorty SJ-10P17SJ-07P-FR04 096 Nem SJ-10P19
1638 Agronomy Journa l • Volume 105, Issue 6 • 2013
for FOV Race 1 and as susceptible checks for FOV Race 4. The Pima S-6 (Ulloa et al., 2013; CV-81 NSL183011), SJ-07-FR01 through SJ-07-FR04 (Ulloa et al., 2009), SJ-10P10, SJ-10P13, SJ-10P17, and SJ-10P19 germplasm lines (Ulloa et al., unpub-lished data, 2010) and Phytogen 800 (Phytogen Seed Co., S. Patent no. 7332.657) were used as resistant checks for both FOV races (1 and 4).
Fusarium Wilt Evaluations
Ten independent replicated field evaluations were con-ducted: three in 2008, four in 2010, and three in 2011 (Table 2). Additional information about the different trials conducted using the RBTN germplasm is presented in Table 2. In 2008, 2010, and 2011, we conducted evaluations at a sandy loam soil field site infested with FOV Race 1 + RKN (Field SCA08- 11_FOVR1 in Table 2). Entries were grown in one-row plots, 5 by 1 m, in a randomized complete block design with three rep-lications. Five plants were randomly chosen per plot to be used as subsamples to obtain estimates of disease severity indices of leaves (DSI) and vascular root staining (VRS) at 81 d (2008), 92 d (2010), and 87 d (2011) after planting (Table 2). The num-ber of main stem nodes (excluding the cotyledonary node) and plant height (measured from the cotyledon node to the upper-most node) were measured on each plant. Plant survival (PS) in each plot was recorded at 81 d (2008, Field SCA08_FOVR1), 92 d (2010, Field SCA10_FOVR1), and 87 d (2011, Field SCA11_FOVR1) after planting. The PS (expressed as a per-centage) was calculated by dividing the total number of surviv-ing plants on each sample date by the initial plant count made 12 to 14 d after planting and multiplying by 100.
Evaluations were also conducted at clay loam soil field sites infested with FOV Race 4 in Fresno (Field FCA08_FOVR4) and Kern (Field KCA08- 11_FOVR4) counties in 2008, 2010, and 2011 (Table 2). Plants at these sites consistently developed severe FOV Race 4 symptoms, and infection of plants were confirmed to be by Race 4. Entries were grown in one-row plots, 5 by 1 m, in a randomized complete block or incom-plete complete block design with three replications. The DSI, VRS, number of nodes, plant height, and PS were determined at 91 d (2008, Fresno County, Field FCA08_FOVR4), 92 d (2008, Kern County, Field KCA08_FOVR4), 48 d (2010,
Kern County, Field KCA10_FOVR4), and 68 d (2011, Kern County, Field KFCA11_FOVR4) after planting (Table 2).
Individual plants were rated for DSI based on the following scale: 0 = no symptoms, 1 = epinasty and slight dwarfing, 2 = 1 to 30% of leaves chlorotic, 3 = 31 to 80% of leaves chlorotic and severe stunting, 4 = 81 to 100% of leaves chlorotic, and 5 = plant death. Stems and the upper part of the primary root of the same plants were cut longitudinally and evaluated for VRS. The following scale was used for VRS: 0 = no vascular root staining evident, 1 = light vascular root staining evident as spotty areas, 2 = root staining more continuous, but light-colored staining covering an area between one quarter and one half of the stem cross-section, 3 = moderate brown to black staining evident in a band encircling most of the stem cross-section, 4 = brown to black staining evident across most vascu-lar tissue in stem cross-section, and 5 = plant severely damaged or plant dead, with staining evident throughout a cross-section of root tissue (Ulloa et al., 2013).
To evaluate the performance of the elite germplasm lines of the RBTN group under SJV environments on non- FOV-infested fields and to compare them with check cultivars in 2010 and 2011, replicated field evaluations were conducted for yield potential and fiber quality characteristics at the Shafter Cotton Research Station, Shafter, CA (Field SCA10_YnF) and at the University of California West Side Research and Extension Center, Five Points, CA (Fields WSCA10_YnF and WSCA11_YnF) (Table 2). All tests were replicated four times in a randomized complete block design. Entries were planted in one-row plots varying from 9 to 14 by 1 m, depending on location. Fiber samples were obtained by hand picking 25-boll samples from each plot. All plots were machine harvested with a modified spindle picker to determine seed cotton yield. Fiber samples were subjected to high-volume instrument (HVI) fiber quality testing at Cotton Incorporated. The following agro-nomic data were included in the analysis of performance and comparisons of elite germplasm lines and check cultivars under SJV environments: seed cotton yield per plot (g), lint yield (kg/ha), and lint percentage (%). The following fiber quality characteristics were included: fiber length (upper half mean), fiber strength, fiber elongation, fineness (micronaire), and short fiber content (%). In addition, in 2011 from the WSCA11_YnF
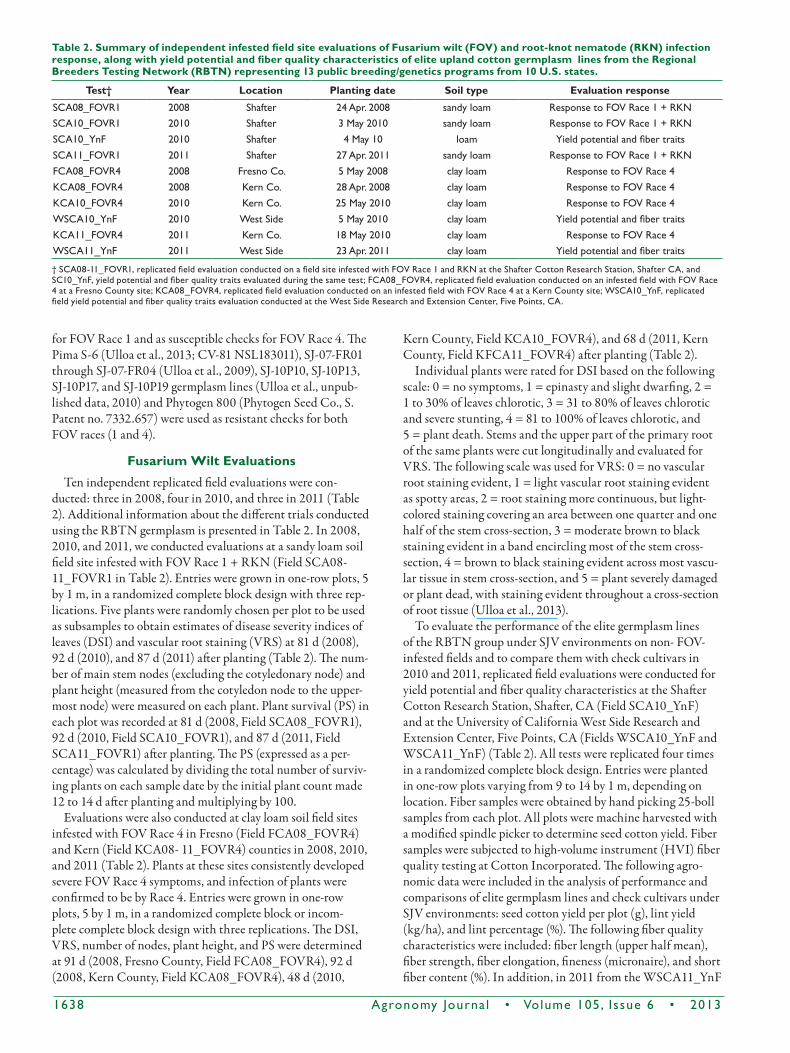
Table 2. Summary of independent infested field site evaluations of Fusarium wilt (FOV) and root-knot nematode (RKN) infection response, along with yield potential and fiber quality characteristics of elite upland cotton germplasm lines from the Regional Breeders Testing Network (RBTN) representing 13 public breeding/genetics programs from 10 U.S. states.
test† Year location Planting date Soil type Evaluation response
SCA08_FOVR1 2008 Shafter 24 Apr. 2008 sandy loam Response to FOV Race 1 + RKNSCA10_FOVR1 2010 Shafter 3 May 2010 sandy loam Response to FOV Race 1 + RKNSCA10_YnF 2010 Shafter 4 May 10 loam Yield potential and fiber traitsSCA11_FOVR1 2011 Shafter 27 Apr. 2011 sandy loam Response to FOV Race 1 + RKNFCA08_FOVR4 2008 Fresno Co. 5 May 2008 clay loam Response to FOV Race 4KCA08_FOVR4 2008 Kern Co. 28 Apr. 2008 clay loam Response to FOV Race 4KCA10_FOVR4 2010 Kern Co. 25 May 2010 clay loam Response to FOV Race 4WSCA10_YnF 2010 West Side 5 May 2010 clay loam Yield potential and fiber traitsKCA11_FOVR4 2011 Kern Co. 18 May 2010 clay loam Response to FOV Race 4WSCA11_YnF 2011 West Side 23 Apr. 2011 clay loam Yield potential and fiber traits
† SCA08-11_FOVR1, replicated field evaluation conducted on a field site infested with FOV Race 1 and RKN at the Shafter Cotton Research Station, Shafter CA, and SC10_YnF, yield potential and fiber quality traits evaluated during the same test; FCA08_FOVR4, replicated field evaluation conducted on an infested field with FOV Race 4 at a Fresno County site; KCA08_FOVR4, replicated field evaluation conducted on an infested field with FOV Race 4 at a Kern County site; WSCA10_YnF, replicated field yield potential and fiber quality traits evaluation conducted at the West Side Research and Extension Center, Five Points, CA.

Agronomy Journa l • Volume 105, Issue 6 • 2013 1639
site, fiber samples were subjected to the advanced fiber infor-mation system (AFIS) instrument for analyzing fiber quality traits. Eight fiber traits were included: short fiber content (g/kg), number of nep size by weight (no./g), upper quartile of fiber length (mm), fiber span length (mm), seed coat fragment size (g), fiber fineness (millitex), immature fiber content by weight (g/kg), and maturity ratio (unit).
data Analyses
Data were analyzed with PROC GLM (SAS version 9.2, SAS Institute) for different values of response (DSI, VRS, number of nodes, plant height, and PS) in 2008, 2010, and 2011. Mean separation among germplasm lines and check cultivars was conducted using the Waller–Duncan k-ratio procedure (Ott, 1988). In addition, genotype ´ sites and genotype ´ FOV race interactions were examined using PROC GLIMMIX and mean separations such as LSD and adjust = simulate (data not presented). Correlation analyses were performed to examine the similarity of response of the elite germplasm lines and check cultivars from the various evaluations of the different trials as indicated by FOV Races 1 and 4 plant infection using DSI, VRS, number of nodes, plant height, and PS. Similarity in plant response of the FOV Races 1 and 4 infection among germplasm lines and check cultivars was assessed by correlation analyses of VRS corresponding to each evaluation with DSI, VRS, number of nodes, plant height, and PS. All correlations were performed using PROC CORR in SAS (version 9.2, SAS Institute).
rESUltSFusarium Wilt resistance Evaluations
Significant variation (P £ 0.05) in resistance response to FOV Races 1 and 4 was observed for the elite germplasm lines and check cultivars based on DSI and VRS values in the 2008, 2010, and 2011 evaluations. In 2008, at the SCA08_FOVR1 field site, VRS values of infection for FOV Race 1 + RKN on tolerant and resistant check cultivars such as upland Phytogen 72, Pima S-7, Phytogen 800, and the SJ-07P-FR01-04 lines
ranged from 0.27 to 1.27 (Fig. 1). At the SCA10_FOVR1 field site in 2010, VRS values on Phytogen 72, Pima S-6, and Pima S-7 ranged from 0.5 to 1.0. In 2011, similar responses for FOV Race 1 + RKN infection was observed on tolerant (VRS £ 2.0) and susceptible (VRS > 2.0) check cultivars at the Shafter Cotton Research Station field site (SCA11_FOVR1, data not presented). Significant variation (P £ 0.05) in plant height and PS was observed for the check cultivars when they were assessed for FOV Races 1 and 4 resistance (Fig. 2). Also at the SCA10_FOVR1 field site, PS at 92 d after planting for upland FOV Race 1 + RKN susceptible Shorty was 52.5%, while toler-ant upland Phytogen 72 was 96.5% at the conclusion of the trial. The VRS and PS values were negatively correlated, ranging from r = –0.17 (2008) to r = –0.68 (2010) at P £ 0.05 within each race and evaluation site, indicating that a high VRS value for FOV Race 1 + RKN infection resulted in low PS.
When we compared infested field evaluation sites (FOV Race 1 + RKN [Shafter site] and FOV Race 4 [Kern and Fresno counties]) (Table 2), significant differences (2008, P £ 0.05; 2010, P £ 0.06; 2011, P £ 0.05) were observed between FOV Race 1 + RKN and FOV Race 4 based on recorded VRS values for the elite germplasm and check cultivars. Overall, the results revealed interactions in disease response between assayed elite germplasm lines and check cultivars, FOV races (1 and 4), and evaluation sites (data not presented). For example, we observed that Pima S-7 and Acala Phytogen 72 responded differently to the two FOV races. Both entries were more susceptible to Race 4 at the FCA08_FOVR4 Fresno and KCA_FOVR4 Kern counties field sites than to FOV Race 1 + RKN at the SCA08_FOVR1 Shafter field site (Fig. 1 and 2). At the SCA08_FOVR1 field site in 2008, the VRS and PS values of FOV Race 1 + RKN were 0.53 and 93.9%, respectively, for resistant check Pima S-7 and 0.27 and 78.7%, respectively, for resistant check Phytogen 72 (Fig. 1 and 2). In 2010, VRS and PS values were 1.0 and 88.6%, respectively, for Pima S-7 and 1.3 and 96.5%, respec-tively, for Phytogen 72 for FOV Race 1 + RKN.
Fig. 1. Distribution of disease index of vascular root staining of Fusarium wilt (FOV) Races 1 and 4 (y axis scale from 0 to 5: 0 = no symptoms and 5 = plant dead) on check cultivars and elite germplasm lines from 10 public breeding/genetics programs from nine U.S. states: Arkansas, South carolina, Georgia, louisiana, texas, Mississippi, Alabama, new Mexico, and california. three independent replicated field evaluations were conducted in California in 2008: FOV Race 1 + root-knot nematode at Shafter Cotton Research Sta-tion, planted 24 Apr. 2008; FOV Race 4 at the Kern County site, planted 28 Apr. 2008; and FOV Race 4 at Fresno County site, planted 5 May 2008.

1640 Agronomy Journa l • Volume 105, Issue 6 • 2013
In 2008, the VRS and PS values for FOV Race 4 of suscepti-ble check Pima S-7 were 4.4 and 5.1% for the FCA08_FOVR4 Fresno County field site and 3.0 and 6% for the KCA08_FOVR4 Kern County field site. For the susceptible check Phy-togen 72, VRS and PS values were 2.6 and 60% for the Fresno County and 2.1 and 26% for the Kern County sites (Fig. 1 and 2). At the KCA10_FOVR4 field site in 2010, the VRS and PS values of FOV Race 4 were 2.9 and 45.1%, respectively, for Pima S-7 and 1.9 and 84.7%, respectively, for Phytogen 72. In addition, no correlation or association was observed for VRS values of infection between FOV Race 1 + RKN and FOV Race 4 in either 2008 or 2010 evaluations. In 2011, a similar response for FOV Race 1 + RKN and FOV Race 4 (Fig. 3 and 4) infection was observed on tolerant or resistant (VRS £ 2.0 and PS ³ 80%) and susceptible (VRS > 2.0 and PS £ 80%) check cultivars.
In 2008 at the FCA08_FOVR4 and KCA08_FOVR4 field sites, VRS values of FOV Race 4 infection were significantly
correlated between the two infested evaluation field sites, with r = 0.58 at P £ 0.05. A correlation was also observed between the two sites for PS (r = 0.37) at P £ 0.05, indicating overall similarity in the responses of the evaluated lines at both FOV Race 4 sites. Averages for VRS and PS were 2.10 and 55% (FCA08_FOVR4) and 2.0 and 61% (KCA08_FOVR4). In 2008, VRS and PS values of infection for FOV Race 4 at the two sites were also negatively correlated, ranging from r = –0.34 (KCA08_FOVR4) to r = –0.68 (FCA08_FOVR4). In 2010, VRS and PS were also negatively correlated, with r = –0.64 (KCA10_FOVR4) at P £ 0.05. These correlations also indicated that a high VRS value resulted in low PS.
Similar results were obtained for elite germplasm lines when comparing infested field evaluation sites (FOV Race 1 + RKN [Shafter site] and FOV Race 4 [Kern and Fresno counties]) (Table 2). For example, in 2008, we observed that AU-5491 (Alabama), Ark0008-22-10 (Arkansas), and LA05307061 (Louisiana) responded differently to the two races based on
Fig. 3. Distribution of disease index of vascular root staining of Fusarium wilt (FOV) Race 4 (y axis scale from 0 to 5: 0 = no symptoms and 5 = plant dead) on check cultivars and elite germplasm lines from 13 public breeding/genetics programs from 10 U.S. states: Ar-kansas, South carolina, Georgia, Mississippi, louisiana, Alabama, new Mexico, texas, north carolina, and california. one indepen-dent replicated field evaluation was conducted in California in 2011 for FOV Race 4 at the Kern County site, planted 27 Apr. 2011.
Fig. 2. Distribution of field plant survival (y axis scale from 0 to 100%) of check cultivars and elite germplasm lines from 10 public breeding/genetics programs from nine U.S. states: Arkansas, South carolina, Georgia, louisiana, texas, Mississippi, Alabama, new Mexico, and California. Plant death was assumed to be due to disease infection of Fusarium wilt (FOV) Races 1 and 4. Three indepen-dent replicated field evaluations were conducted in California in 2008: FOV Race 1 + root-knot nematode at Shafter Cotton Research Station, planted 24 Apr. 2008; FOV Race 4 at the Kern County site, planted 28 Apr. 2008; and FOV Race 4 at Fresno County site, planted 5 May 2008.

Agronomy Journa l • Volume 105, Issue 6 • 2013 1641
VRS values. Each was more susceptible to Race 4 than Race 1 + RKN (Fig. 1 and 2). In addition, in 2010, we observed that GA2006053 (Georgia), MD51ne (Mississippi), and 0033-6 (Mississippi) were more susceptible to FOV Race 1 + RKN than to FOV Race 4 (data not presented). In 2010, VRS and PS values on elite germplasm for FOV Races 1 and 4 infections were lower (possibly due to different inoculum levels in the field) than in 2008; however, cultivar resistance and susceptible checks for the two races responded as expected in 2010, with high VRS and low PS values similar to 2008 observations.
Entries AU-6103 (Alabama), Ark0015-06-11 (Arkansas), LA05307029 (Louisiana), and 0149-17ne and 04-PST-275 (Mississippi) appeared tolerant to both FOV races (1 and 4) based on VRS and PS values from the 2008 trials. Similar results were obtained in 2010, when some elite germplasm lines from Alabama (AU6001), Arkansas (Ark0203-11), Louisiana (LA07307106), Mississippi (MD25Y), and South Carolina (PX03201-66-1) also appeared to be tolerant to both FOV races (data not presented). Other germplasm lines appeared tolerant in terms of good growth rates, lack of visual foliar symptoms, and high PS but still showed high infection rates as evidenced by high VRS values (Fig. 1 and 2).
In 2008, >50% of the germplasm lines had a VRS for FOV Race 1 + RKN ³ 1.4 and a PS £ 85%, while for FOV Race 4, >50% of the germplasm lines had a VRS of ³ 2.0 and a PS £ 65% at least at one site (Fig. 1 and 2). In 2010, >30% of the germplasm lines had a VRS for FOV Race 4 ³ 1.5 and a PS £ 92%, while for FOV Race 1 + RKN they had a VRS ³ 1.0 and a PS £ 87% (data not presented). In 2011, many upland lines were severely affected by FOV at the field test site (KCA11_FOVR4). Fusarium wilt Race 4 has been the more devastating race for cotton in California; VRS values ranged from 1.1 to 2.5 for the elite germplasm lines, with resistant Pima S-6 = 0.6, susceptible Pima S-7 = 4.5, and Phytogen 72 = 2.6 (Fig. 3). Plant survival ranged from 58 to 91% for the elite germplasm lines, with resistant Pima S-6 = 95%, susceptible Pima S-7 = 8%, and Phytogen 72 = 46% (Fig. 4).
Based on these evaluations to date, many of the entries in the current RBTN public elite upland germplasm pool may be more susceptible than expected to some FOV races, and sources of acceptable levels of resistance may be limited when tested under infestation levels that resulted in only 5 to 35% plant survival in susceptible check cultivars.
Yield and Fiber Quality Evaluations
When the performance of the elite germplasm lines was evaluated under SJV environments on non-FOV-infested fields for yield potential and fiber quality characteristics, significant variation (P £ 0.05) was observed among elite germplasm lines and check cultivars for agronomic (seed cotton yield, lint yield, and lint percentage) and fiber quality traits (fiber length (upper half mean), fiber strength, fiber elongation, micronaire, short fiber content, short fibers by weight, nep size, upper quartile fiber length, fiber length, seed coat fragment size, fiber fineness, immature fiber content, and maturity ratio) during 2 yr (2010 and 2011) at the University of California West Side Research and Extension Center, Five Points, CA (WSCA10- 11_YnF) and Shafter Cotton Research Station, Shafter, CA (SCA11_YnF) (Table 2). Genotype ´ site interaction was significant (P £ 0.05) for agronomic and fiber quality traits, except for lint percentage and fiber strength.
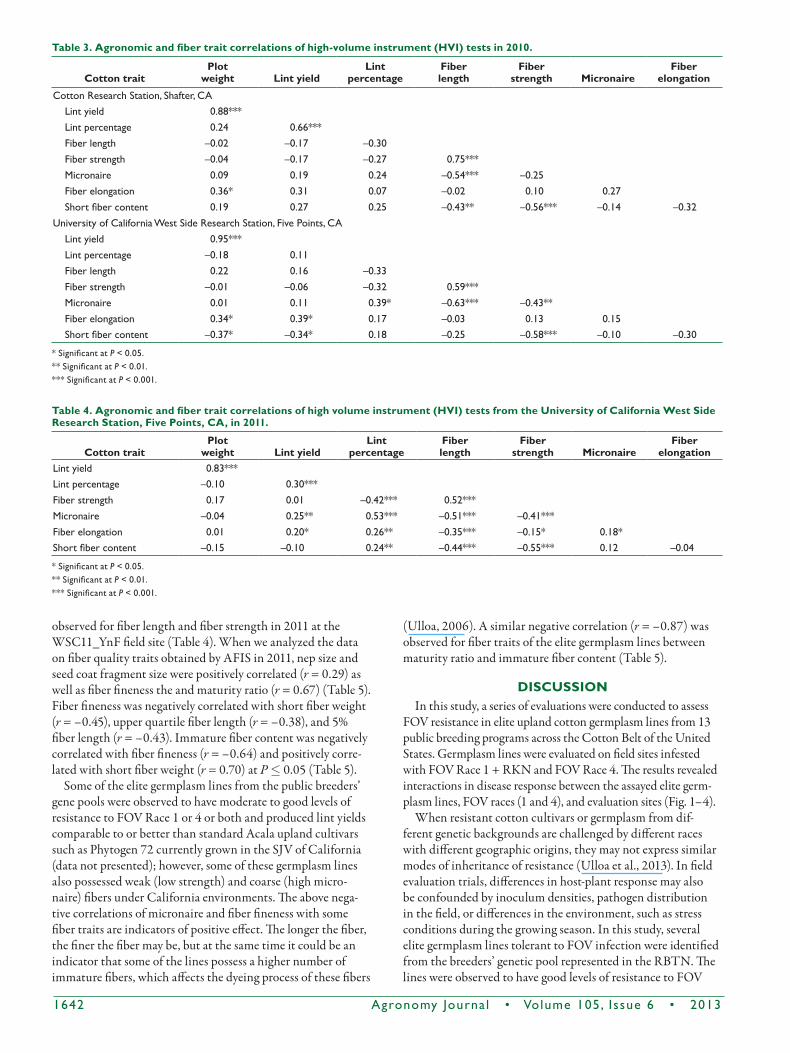
In the test at the SCA10_YnF Shafter field site in 2010, cor-relations between agronomic traits on a mean basis were only significant between lint yield and lint percentage (r = 0.66) (Table 3). For fiber quality traits, fiber length was negatively correlated with micronaire (r = –0.54) and short fiber content (r = –0.43) and positively correlated with fiber strength (r = 0.75) at P £ 0.05. Fiber strength was negatively correlated with short fiber content (r = –0.56) (Table 3). In the test at the WSCA10_YnF West Side field site in 2010, lint percent-age was positively correlated with micronaire (r = 0.39) (Table 3); fiber length was also negatively correlated with micronaire (r = –0.63) and positively correlated with fiber strength (r = 0.59). Fiber strength was also negatively correlated with short fiber content (r = –0.58) (Table 3). Similar correlations were
Fig. 4. Distribution of field plant survival (y axis scale from 0 to 100%) of check cultivars and elite germplasm lines from 13 public breeding/genetics programs from 10 U.S. states: : Arkansas, South carolina, Georgia, Mississippi, louisiana, Alabama, new Mexico, Texas, North Carolina, and California. Plant death was assumed to be due to disease infection of Fusarium wilt (FOV) Race 4. One independent replicated field evaluation conducted in California in 2011 for FOV Race 4 at the Kern County site, planted 27 Apr. 2011.
1642 Agronomy Journa l • Volume 105, Issue 6 • 2013
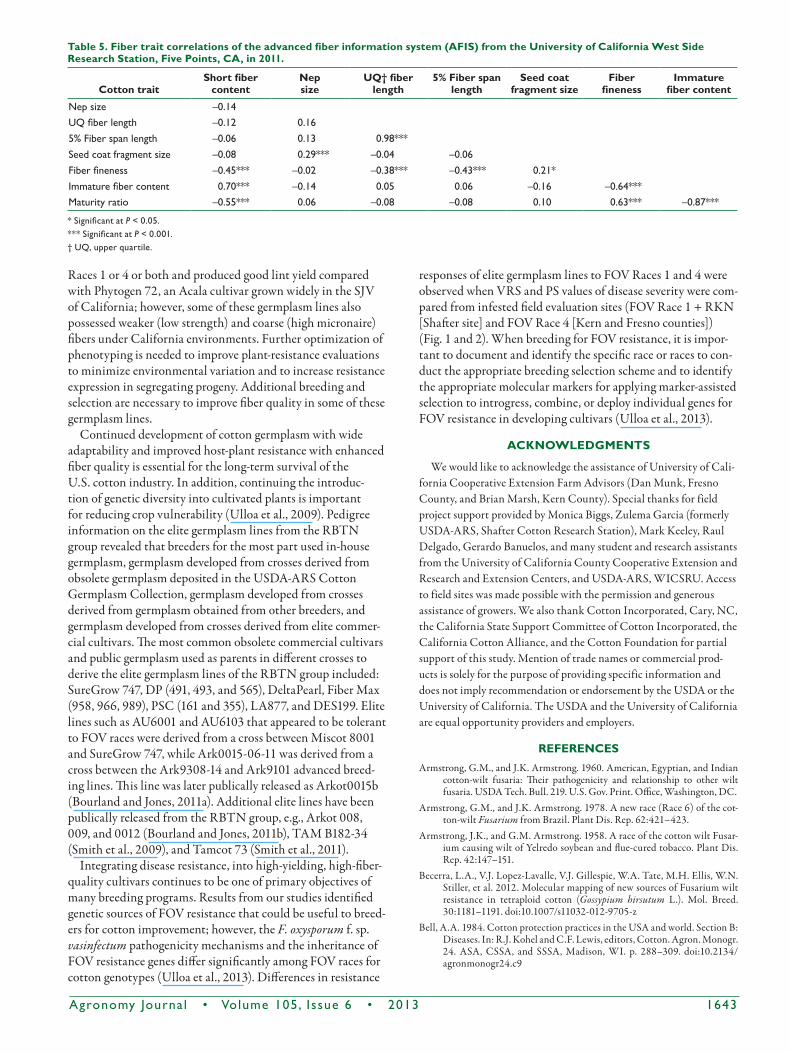
observed for fiber length and fiber strength in 2011 at the WSC11_YnF field site (Table 4). When we analyzed the data on fiber quality traits obtained by AFIS in 2011, nep size and seed coat fragment size were positively correlated (r = 0.29) as well as fiber fineness the and maturity ratio (r = 0.67) (Table 5). Fiber fineness was negatively correlated with short fiber weight (r = –0.45), upper quartile fiber length (r = –0.38), and 5% fiber length (r = –0.43). Immature fiber content was negatively correlated with fiber fineness (r = –0.64) and positively corre-lated with short fiber weight (r = 0.70) at P £ 0.05 (Table 5).
Some of the elite germplasm lines from the public breeders’ gene pools were observed to have moderate to good levels of resistance to FOV Race 1 or 4 or both and produced lint yields comparable to or better than standard Acala upland cultivars such as Phytogen 72 currently grown in the SJV of California (data not presented); however, some of these germplasm lines also possessed weak (low strength) and coarse (high micro-naire) fibers under California environments. The above nega-tive correlations of micronaire and fiber fineness with some fiber traits are indicators of positive effect. The longer the fiber, the finer the fiber may be, but at the same time it could be an indicator that some of the lines possess a higher number of immature fibers, which affects the dyeing process of these fibers
(Ulloa, 2006). A similar negative correlation (r = –0.87) was observed for fiber traits of the elite germplasm lines between maturity ratio and immature fiber content (Table 5).
diScUSSionIn this study, a series of evaluations were conducted to assess
FOV resistance in elite upland cotton germplasm lines from 13 public breeding programs across the Cotton Belt of the United States. Germplasm lines were evaluated on field sites infested with FOV Race 1 + RKN and FOV Race 4. The results revealed interactions in disease response between the assayed elite germ-plasm lines, FOV races (1 and 4), and evaluation sites (Fig. 1–4).
When resistant cotton cultivars or germplasm from dif-ferent genetic backgrounds are challenged by different races with different geographic origins, they may not express similar modes of inheritance of resistance (Ulloa et al., 2013). In field evaluation trials, differences in host-plant response may also be confounded by inoculum densities, pathogen distribution in the field, or differences in the environment, such as stress conditions during the growing season. In this study, several elite germplasm lines tolerant to FOV infection were identified from the breeders’ genetic pool represented in the RBTN. The lines were observed to have good levels of resistance to FOV
Table 3. Agronomic and fiber trait correlations of high-volume instrument (HVI) tests in 2010.
cotton traitPlot
weight lint yieldlint
percentageFiber length
Fiber strength Micronaire
Fiber elongation
Cotton Research Station, Shafter, CALint yield 0.88***Lint percentage 0.24 0.66***Fiber length –0.02 –0.17 –0.30Fiber strength –0.04 –0.17 –0.27 0.75***Micronaire 0.09 0.19 0.24 –0.54*** –0.25Fiber elongation 0.36* 0.31 0.07 –0.02 0.10 0.27Short fiber content 0.19 0.27 0.25 –0.43** –0.56*** –0.14 –0.32
University of California West Side Research Station, Five Points, CALint yield 0.95***Lint percentage –0.18 0.11Fiber length 0.22 0.16 –0.33Fiber strength –0.01 –0.06 –0.32 0.59***Micronaire 0.01 0.11 0.39* –0.63*** –0.43**Fiber elongation 0.34* 0.39* 0.17 –0.03 0.13 0.15Short fiber content –0.37* –0.34* 0.18 –0.25 –0.58*** –0.10 –0.30
* Significant at P < 0.05.** Significant at P < 0.01.*** Significant at P < 0.001.
Table 4. Agronomic and fiber trait correlations of high volume instrument (HVI) tests from the University of California West Side research Station, Five Points, cA, in 2011.
cotton traitPlot
weight lint yieldlint
percentageFiber length
Fiber strength Micronaire
Fiber elongation
Lint yield 0.83***Lint percentage –0.10 0.30***Fiber strength 0.17 0.01 –0.42*** 0.52***Micronaire –0.04 0.25** 0.53*** –0.51*** –0.41***Fiber elongation 0.01 0.20* 0.26** –0.35*** –0.15* 0.18*Short fiber content –0.15 –0.10 0.24** –0.44*** –0.55*** 0.12 –0.04
* Significant at P < 0.05.** Significant at P < 0.01.*** Significant at P < 0.001.
Agronomy Journa l • Volume 105, Issue 6 • 2013 1643
Races 1 or 4 or both and produced good lint yield compared with Phytogen 72, an Acala cultivar grown widely in the SJV of California; however, some of these germplasm lines also possessed weaker (low strength) and coarse (high micronaire) fibers under California environments. Further optimization of phenotyping is needed to improve plant-resistance evaluations to minimize environmental variation and to increase resistance expression in segregating progeny. Additional breeding and selection are necessary to improve fiber quality in some of these germplasm lines.
Continued development of cotton germplasm with wide adaptability and improved host-plant resistance with enhanced fiber quality is essential for the long-term survival of the U.S. cotton industry. In addition, continuing the introduc-tion of genetic diversity into cultivated plants is important for reducing crop vulnerability (Ulloa et al., 2009). Pedigree information on the elite germplasm lines from the RBTN group revealed that breeders for the most part used in-house germplasm, germplasm developed from crosses derived from obsolete germplasm deposited in the USDA-ARS Cotton Germplasm Collection, germplasm developed from crosses derived from germplasm obtained from other breeders, and germplasm developed from crosses derived from elite commer-cial cultivars. The most common obsolete commercial cultivars and public germplasm used as parents in different crosses to derive the elite germplasm lines of the RBTN group included: SureGrow 747, DP (491, 493, and 565), DeltaPearl, Fiber Max (958, 966, 989), PSC (161 and 355), LA877, and DES199. Elite lines such as AU6001 and AU6103 that appeared to be tolerant to FOV races were derived from a cross between Miscot 8001 and SureGrow 747, while Ark0015-06-11 was derived from a cross between the Ark9308-14 and Ark9101 advanced breed-ing lines. This line was later publically released as Arkot0015b (Bourland and Jones, 2011a). Additional elite lines have been publically released from the RBTN group, e.g., Arkot 008, 009, and 0012 (Bourland and Jones, 2011b), TAM B182-34 (Smith et al., 2009), and Tamcot 73 (Smith et al., 2011).
Integrating disease resistance, into high-yielding, high-fiber-quality cultivars continues to be one of primary objectives of many breeding programs. Results from our studies identified genetic sources of FOV resistance that could be useful to breed-ers for cotton improvement; however, the F. oxysporum f. sp. vasinfectum pathogenicity mechanisms and the inheritance of FOV resistance genes differ significantly among FOV races for cotton genotypes (Ulloa et al., 2013). Differences in resistance
responses of elite germplasm lines to FOV Races 1 and 4 were observed when VRS and PS values of disease severity were com-pared from infested field evaluation sites (FOV Race 1 + RKN [Shafter site] and FOV Race 4 [Kern and Fresno counties]) (Fig. 1 and 2). When breeding for FOV resistance, it is impor-tant to document and identify the specific race or races to con-duct the appropriate breeding selection scheme and to identify the appropriate molecular markers for applying marker-assisted selection to introgress, combine, or deploy individual genes for FOV resistance in developing cultivars (Ulloa et al., 2013).
ACKNOWleDgMeNTS
We would like to acknowledge the assistance of University of Cali-fornia Cooperative Extension Farm Advisors (Dan Munk, Fresno County, and Brian Marsh, Kern County). Special thanks for field project support provided by Monica Biggs, Zulema Garcia (formerly USDA-ARS, Shafter Cotton Research Station), Mark Keeley, Raul Delgado, Gerardo Banuelos, and many student and research assistants from the University of California County Cooperative Extension and Research and Extension Centers, and USDA-ARS, WICSRU. Access to field sites was made possible with the permission and generous assistance of growers. We also thank Cotton Incorporated, Cary, NC, the California State Support Committee of Cotton Incorporated, the California Cotton Alliance, and the Cotton Foundation for partial support of this study. Mention of trade names or commercial prod-ucts is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the USDA or the University of California. The USDA and the University of California are equal opportunity providers and employers.
rEFErEncESArmstrong, G.M., and J.K. Armstrong. 1960. American, Egyptian, and Indian
cotton-wilt fusaria: Their pathogenicity and relationship to other wilt fusaria. USDA Tech. Bull. 219. U.S. Gov. Print. Office, Washington, DC.
Armstrong, G.M., and J.K. Armstrong. 1978. A new race (Race 6) of the cot-ton-wilt Fusarium from Brazil. Plant Dis. Rep. 62:421–423.
Armstrong, J.K., and G.M. Armstrong. 1958. A race of the cotton wilt Fusar-ium causing wilt of Yelredo soybean and flue-cured tobacco. Plant Dis. Rep. 42:147–151.
Becerra, L.A., V.J. Lopez-Lavalle, V.J. Gillespie, W.A. Tate, M.H. Ellis, W.N. Stiller, et al. 2012. Molecular mapping of new sources of Fusarium wilt resistance in tetraploid cotton (Gossypium hirsutum L.). Mol. Breed. 30:1181–1191. doi:10.1007/s11032-012-9705-z
Bell, A.A. 1984. Cotton protection practices in the USA and world. Section B: Diseases. In: R.J. Kohel and C.F. Lewis, editors, Cotton. Agron. Monogr. 24. ASA, CSSA, and SSSA, Madison, WI. p. 288–309. doi:10.2134/agronmonogr24.c9
Table 5. Fiber trait correlations of the advanced fiber information system (AFIS) from the University of California West Side research Station, Five Points, cA, in 2011.
cotton traitShort fiber
contentnepsize
UQ† fiber length
5% Fiber span length
Seed coat fragment size
Fiber fineness
immature fiber content
Nep size –0.14UQ fiber length –0.12 0.165% Fiber span length –0.06 0.13 0.98***Seed coat fragment size –0.08 0.29*** –0.04 –0.06Fiber fineness –0.45*** –0.02 –0.38*** –0.43*** 0.21*Immature fiber content 0.70*** –0.14 0.05 0.06 –0.16 –0.64***Maturity ratio –0.55*** 0.06 –0.08 –0.08 0.10 0.63*** –0.87***
* Significant at P < 0.05.*** Significant at P < 0.001.† UQ, upper quartile.

1644 Agronomy Journa l • Volume 105, Issue 6 • 2013
Bourland, F.M., and D.C. Jones. 2011a. Registration of Arkot 0015a, Arkot 0015b, and Arkot 0016 germplasm lines of cotton. J. Plant Regist. 5:379–383. doi:10.3198/jpr2010.12.0704crg
Bourland, F.M., and D.C. Jones. 2011b. Registration of Arkot 0008, Arkot 0009, and Arkot 0012 germplasm lines of cotton. J. Plant Regist. 5:379–383. doi:10.3198/jpr2010.12.0704crg
DeVay, J.E. 1986. Half a century dynamics and control of cotton diseases: Fusarium and Verticillium wilts. In: Proceedings of the Beltwide Cotton Production Research Conferences, Las Vegas, NV. 4–9 Jan. 1986. Natl. Cotton Counc. Am., Memphis, TN. p. 35–41.
Garber, R.H., E.C.C. Jorgenson, S. Smith, and A.H. Hyer. 1979. Interaction of population levels of Fusarium oxysporum f.sp. vasinfectum and Meloido-gyne incognita on cotton. J. Nematol. 11:33–37.
Garber, R.H., and G.A. Paxman. 1963. Fusarium wilt of cotton in California. Plant Dis. Rep. 47:398–400.
Heald, C.M., and C.C. Orr. 1982. Nematode parasites of cotton. In: W.R. Nickle, editor, Plant and insect nematodes. Marcel Dekker, New York.
Holmes, E.A., R.S. Bennett, D.W. Spurgeon, P.D. Colyer, and R.M. Davis. 2009. New genotypes of Fusarium oxysporum f. sp. vasinfectum from the southeastern United States. Plant Dis. 93:1298–1304. doi:10.1094/PDIS-93-12-1298
Hutmacher, R.B., M. Ulloa, S.D. Wright, R.M. Davis, M.P. Keeley, and R. Delgado, et al. 2011. Fusarium Race 4: Management recommenda-tions for growers. In: Proceedings of the Beltwide Cotton Conferences, Atlanta, GA. 4–7 Jan. 2011. Natl. Cotton Counc. Am., Memphis, TN. p. 188–192.
Ibrahim, F.M. 1966. A new race of the cotton-wilt Fusarium in the Sudan Gezira. Emp. Cotton Grow. Rev. 43:296–299.
Kim, Y., R.B. Hutmacher, and R.M. Davis. 2005. Characterization of Califor-nia isolates of Fusarium oxysporum f. sp. vasinfectum. Plant Dis. 89:366–372. doi:10.1094/PD-89-0366
Kochman, J., L. Swan, N. Moore, S. Bentley, W. O’Neill, A. Mitchell, et al. 2002. The Fusarium threat: Are we making the progress? In: Proceedings of the 11th Australian Cotton Conference, Brisbane, QLD, Australia. 13–15 Aug. 2002. Aust. Cotton Growers’ Res. Assoc., Wee Waa, NSW, Australia. p. 643–652.
Mohamed, H.A. 1963. Inheritance of resistance to Fusarium wilt in some Egyptian cottons. Emp. Cotton Grow. Rev. 40:292–295.
Ott, L. 1988. An introduction to statistical methods and data analysis. PWS-Kent Publ. Co., Boston.
Sasser, J.N. 1972. Nematode disease of cotton. In: J.M. Webster, editor, Eco-nomic nematology. Academic Press, New York. p. 187–214.
Skovgaard, K., H.I. Nirenberg, K. O’Donnell, and S. Rosendahl. 2001. Evo-lution of Fusarium oxysporum f. sp vasinfectum races inferred from multigene genealogies. Phytopathology 91:1231–1237. doi:10.1094/PHYTO.2001.91.12.1231
Smith, A.L., and J.B. Dick. 1960. Inheritance of resistance to Fusarium wilt in upland and Sea Island cotton as complicated by nematodes under field conditions. Phytopathology 50:44–48.
Smith, C.W., R.G. Cantrell, H.S. Moser, and S.R. Oakley. 1999. History of cultivar development in the United States. In: C.W. Smith and J. Cothren, editors, Cotton: Origin, history, technology, and production. John Wiley & Sons, New York. p. 99–171.
Smith, C.W., S. Hague, and D. Jones. 2011. Registration of ‘Tamcot 73’ upland cotton cultivar. J. Plant Regist. 5:273–278. doi:10.3198/jpr2010.12.0723crc
Smith, C.W., S. Hague, P.S. Thaxton, E. Hequet, and D. Jones. 2009. Regis-tration of eight extra-long staple upland cotton germplasm lines. J. Plant Regist. 3:81–85. doi:10.3198/jpr2008.02.0114crg
Turcotte, E.L., R.G. Percy, and C.F. Feaster. 1992. Registration of ‘Pima S-7’ American Pima cotton. Crop Sci. 32:1291. doi:10.2135/cropsci1992.0011183X003200050047x
Ulloa, M. 2006. Heritability and correlations of agronomic and fiber traits in an okra-leaf upland cotton population. Crop Sci. 46:1508–1514. doi:10.2135/cropsci2005.08-0271
Ulloa, M., C. Brubaker, and P. Chee. 2007. Cotton. In: C. Kole, editor, Genome mapping and molecular breeding. Vol. 6. Technical crops. Springer-Ver-lag, Heidelberg, Germany. p. 1–49. doi:10.1007/978-3-540-34538-1_1
Ulloa, M., R.B. Hutmacher, R.M. Davis, S.D. Wright, R. Percy, and B. Marsh. 2006. Breeding for Fusarium wilt Race 4 resistance in cotton under field and greenhouse conditions. J. Cotton Sci. 10:114–127.
Ulloa, M., R.B. Hutmacher, P.A. Roberts, S.D. Wright, R.L. Nichols, and R.M. Davis. 2013. Inheritance and QTL mapping of Fusarium wilt Race 4 resistance in cotton. Theor. Appl. Genet. 126:1405–1418. doi:10.1007/s00122-013-2061-5
Ulloa, M., R.B. Hutmacher, S.D. Wright, B. Campbell, T. Wallace, G. Myers, et al. 2010. Beltwides’ elite upland germplasm pool assessment of Fusar-ium wilt (FOV) Races 1 and 4 in California. In: Proceedings of the Belt-wide Cotton Conferences, New Orleans, LA. 4–7 Jan. 2010. Natl. Cot-ton Counc. Am, Memphis, TN. p. 765–766.
Ulloa, M., R. Percy, J. Zhang, R.B. Hutmacher, S.D. Wright, and R.M. Davis. 2009. Registration of four Pima cotton germplasm lines having good levels of Fusarium wilt Race 4 resistance with moderate yields and good fibers. J. Plant Regist. 3:198–202. doi:10.3198/jpr2008.09.0530crg
Ulloa, M., C. Wang, R.B. Hutmacher, S.D. Wright, R.M. Davis, C.A. Saski, and P.A. Roberts. 2011. Mapping Fusarium wilt Race 1 resistance genes in cotton by inheritance, QTL, and sequencing composition. Mol. Genet. Genomics 286:21–36. doi:10.1007/s00438-011-0616-1
Veech, J.A. 1984. Cotton protection practices in the USA and world. Sec-tion C: Nematodes. In: R.J. Kohel and C.F. Lewis, editors, Cotton. Agron. Monogr. 24. ASA, CSSA, and SSSA, Madison, WI. p. 309–329. doi:10.2134/agronmonogr24.c9
Wang, B., C.L. Brubaker, and J. Burdon. 2004. Fusarium species and Fusarium wilt pathogens associated with native Gossypium populations in Austra-lia. Mycol. Res. 108:35–44. doi:10.1017/S0953756203008803
Wang, P., L. Su, L. Qin, B. Hu, W. Guo, and T. Zhang. 2009. Identification and molecular mapping of Fusarium wilt resistant gene in upland cot-ton. Theor. Appl. Genet. 119:733–739. doi:10.1007/s00122-009-1084-4




















