
In search of decoy/guardee to R genes: deciphering the role of sugars in defense against Fusarium...
7
www.landesbioscience.com Plant Signaling & Behavior 1081 Plant Signaling & Behavior 5:9, 1081-1087; September 2010; © 2010 Landes Bioscience REVIEW REVIEW ‘Survival for existence’ is the dictum followed by the entire living world. Similarly ‘survival of the fittest’ is nature’s preference. Owing to the extensive surveillance system of higher organ- isms resistance becomes the natural rule while susceptibility the exception. 1 All living entities are being exposed to a plethora of interactions ranging from mutualism to antagonism. 2 However the adaptive strategies opted by the plants are unique, versatile and still grossly unknown which have attracted the research- ers since decades towards looking into the varied responses and diversification of plant adaptation. Plants are hosts to a large number of organisms such as symbi- otic/pathogenic bacteria, phytopathogenic fungi, harmful viruses and nematodes. All have their own stratagem to gain over their host. 3 However only the plant-fungal interaction with Chickpea- Fusarium case study in particular, shall be the focal area of the present review. Fungi are classified as necrotrophic and biotro- phic according to their nutritional requirements. 4 Necrotrophs apply ‘brute force’ by killing host cells and thriving on their dead remains while biotrophs prefer subtler ‘modus operandi’—the *Correspondence to: Sampa Das; Email: [email protected] Submitted: 04/29/10; Accepted: 04/29/10 Previously published online: www.landesbioscience.com/journals/psb/article/12234 DOI:10.4161/psb.5.9.12234 Plant responses are coordinately controlled by both external and internal signals. Apt perception of pathogen attack and its appropriate conversion to internal signals ultimately determine the outcome of innate immunity. The present review predicts the involvement of unconventional ‘guard/decoy model’ in chickpea-Fusarium encounter. Rapid alkalinization factor is predicted to act as initial ‘Gatekeeper decoy’ counteracting fungal entry. Phospholipases and cystatins probably function as ‘Guardees’ being shielded by R gene(s). Serine Threonine Kinases decodes external pathogenic signals to in planta defense alarms. 14.3.3 provides clues to the wilt mechanism. The versatile sugars serve as signal generators and transmitters maintaining intra and inter cellular connectivity during stress. In search of decoy/guardee to R genes Deciphering the role of sugars in defense against Fusarium wilt in chickpea Sumanti Gupta, 1 Dipankar Chakraborti, 1,2 Debabrata Basu 1 and Sampa Das 1, * 1 Division of Plant Biology; Bose Institute; Centenary Campus; Kankurgachi, Kolkata India; 2 P.G. Department of Biotechnology; St. Xavier’s College; Kolkata, India Key words: R gene, decoy, guardee, RALF, ROS, STK, 14.3.3, sugar, defense Abbreviations: RALF, rapid alkalinization factor; ROS, reactive oxygen species; HR, hypersensitive response; STK, serine threonine kinase stealth mechanism used to derive nutrients from live host cells. 5 Irrespective of the pathogen type and their mode of nutrition procurement, perception of attack lies central to effective induc- tion of innate immunity in plants. Arms Race vs. Trench Warfare: Explaining the Coevolution in R-avr Plant’s perception of pathogen attack was well explained by Gene-for-Gene (GFG) hypothesis at the population level. The Gene-for-Gene hypothesis (GFG) characterizing the plant’s R gene and the pathogen’s cognate avr gene, interacting in a recep- tor-ligand fashion was supported by extensive experimental data since 1940s to recent. 6-10 The ‘theory of coevolution’ of plant and pathogen had given rise to two different schools of thought among the scientific community. One believed that both the plant and pathogen undergoes antagonistic coevolution support- ing the ‘Arms Race Model’ while the other school emphasized the ‘Trench Warfare Model’. 11 The proponents of ‘Arms Race Model’ predicted the transient variation in disease resistance to be the outcome of short-term ecological dynamics. This school also supported the R loci in host population to be monomorphic in nature. 11 On the other hand, the followers of ‘trench warfare’ believed that the advances and retreats of R gene allele frequency maintained variation for resistance as an age-old dynamic poly- morphic process. 12 Nevertheless, both the followers coalesced in accepting the ‘Geographical Mosaic of Coevolution Theory’ which explicitly brought into account the inherent complexity of local and distant interacting communities, temporal and spatial species variation and the heterogeneity in magnitude and direction of reciprocal selection across localities. 13,14 From Guard to Decoy Model: Molecular Basis of Pathogen Perception by Host Recent studies have proposed the ‘Guard Model’ opposing the classical GFG hypothesis. The supporters of ‘Guard Model’ showed the involvement of more than two genes in plant-pathogen recognition. 15 The ‘Guard Model’ emphasizes the interaction
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of In search of decoy/guardee to R genes: deciphering the role of sugars in defense against Fusarium...

www.landesbioscience.com Plant Signaling & Behavior 1081
Plant Signaling & Behavior 5:9, 1081-1087; September 2010; © 2010 Landes Bioscience
REVIEW REVIEW
‘Survival for existence’ is the dictum followed by the entire living world. Similarly ‘survival of the fittest’ is nature’s preference. Owing to the extensive surveillance system of higher organ-isms resistance becomes the natural rule while susceptibility the exception.1 All living entities are being exposed to a plethora of interactions ranging from mutualism to antagonism.2 However the adaptive strategies opted by the plants are unique, versatile and still grossly unknown which have attracted the research-ers since decades towards looking into the varied responses and diversification of plant adaptation.
Plants are hosts to a large number of organisms such as symbi-otic/pathogenic bacteria, phytopathogenic fungi, harmful viruses and nematodes. All have their own stratagem to gain over their host.3 However only the plant-fungal interaction with Chickpea-Fusarium case study in particular, shall be the focal area of the present review. Fungi are classified as necrotrophic and biotro-phic according to their nutritional requirements.4 Necrotrophs apply ‘brute force’ by killing host cells and thriving on their dead remains while biotrophs prefer subtler ‘modus operandi’—the
*Correspondence to: Sampa Das; Email: [email protected]: 04/29/10; Accepted: 04/29/10Previously published online:www.landesbioscience.com/journals/psb/article/12234DOI:10.4161/psb.5.9.12234
Plant responses are coordinately controlled by both external and internal signals. Apt perception of pathogen attack and its appropriate conversion to internal signals ultimately determine the outcome of innate immunity. The present review predicts the involvement of unconventional ‘guard/decoy model’ in chickpea-Fusarium encounter. Rapid alkalinization factor is predicted to act as initial ‘Gatekeeper decoy’ counteracting fungal entry. Phospholipases and cystatins probably function as ‘Guardees’ being shielded by R gene(s). Serine Threonine Kinases decodes external pathogenic signals to in planta defense alarms. 14.3.3 provides clues to the wilt mechanism. The versatile sugars serve as signal generators and transmitters maintaining intra and inter cellular connectivity during stress.
In search of decoy/guardee to R genesDeciphering the role of sugars in defense
against Fusarium wilt in chickpeaSumanti Gupta,1 Dipankar Chakraborti,1,2 Debabrata Basu1 and Sampa Das1,*
1Division of Plant Biology; Bose Institute; Centenary Campus; Kankurgachi, Kolkata India; 2P.G. Department of Biotechnology; St. Xavier’s College; Kolkata, India
Key words: R gene, decoy, guardee, RALF, ROS, STK, 14.3.3, sugar, defense
Abbreviations: RALF, rapid alkalinization factor; ROS, reactive oxygen species; HR, hypersensitive response; STK, serine threonine kinase
stealth mechanism used to derive nutrients from live host cells.5 Irrespective of the pathogen type and their mode of nutrition procurement, perception of attack lies central to effective induc-tion of innate immunity in plants.
Arms Race vs. Trench Warfare: Explaining the Coevolution in R-avr
Plant’s perception of pathogen attack was well explained by Gene-for-Gene (GFG) hypothesis at the population level. The Gene-for-Gene hypothesis (GFG) characterizing the plant’s R gene and the pathogen’s cognate avr gene, interacting in a recep-tor-ligand fashion was supported by extensive experimental data since 1940s to recent.6-10 The ‘theory of coevolution’ of plant and pathogen had given rise to two different schools of thought among the scientific community. One believed that both the plant and pathogen undergoes antagonistic coevolution support-ing the ‘Arms Race Model’ while the other school emphasized the ‘Trench Warfare Model’.11 The proponents of ‘Arms Race Model’ predicted the transient variation in disease resistance to be the outcome of short-term ecological dynamics. This school also supported the R loci in host population to be monomorphic in nature.11 On the other hand, the followers of ‘trench warfare’ believed that the advances and retreats of R gene allele frequency maintained variation for resistance as an age-old dynamic poly-morphic process.12
Nevertheless, both the followers coalesced in accepting the ‘Geographical Mosaic of Coevolution Theory’ which explicitly brought into account the inherent complexity of local and distant interacting communities, temporal and spatial species variation and the heterogeneity in magnitude and direction of reciprocal selection across localities.13,14
From Guard to Decoy Model: Molecular Basis of Pathogen Perception by Host
Recent studies have proposed the ‘Guard Model’ opposing the classical GFG hypothesis. The supporters of ‘Guard Model’ showed the involvement of more than two genes in plant-pathogen recognition.15 The ‘Guard Model’ emphasizes the interaction

1082 Plant Signaling & Behavior Volume 5 Issue 9
of non-cannonical transcripts with known metabolic functions but unknown stress or immune relations. Such unexpected out-come of root transcriptome was attributed to be somehow con-trolled by cellular and epigenetic environment.27 Aiming to solve the puzzle behind the exceptions of Chickpea-Fusarium inter-action we identified and explained the role of several molecular factors and the networks in governing pathogen recognition and downstream defense signaling in our previous reports.28,29 The present study shall step ahead in deciphering the role of probable guardees, decoys, kinases and sugars in imparting defense.
Initial ‘No Entry’ Board for Pathogen; RALF vs. ROS
Successful penetration of host cell is referred to as a milestone achievement for an invading fungus. Besides, in case of plant biotrophic interaction host cell penetration is considered to be a ‘remarkable biological phenomenon’ since the pathogen repro-grams the host metabolism in such a fashion that it disrupts the cell wall integrity but poses no threat to the cell viability.1 Further colonization depends on the ability of the entering fungus to optimize its growth and propagation within the host interior.30 Thus, appropriate and timely pathogen recognition becomes crucial from the host’s perspective since it ultimately decides the outcome of resistance/susceptibility.
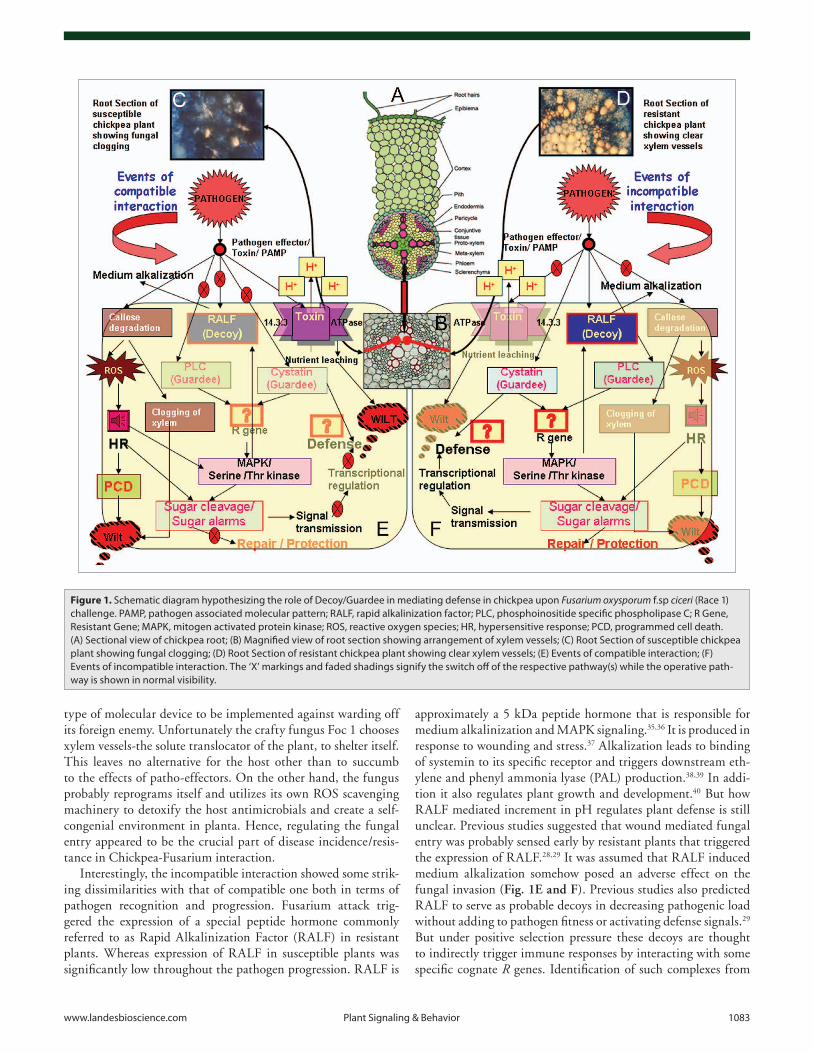
It is well documented that Pathogen Associated Molecular Patterns (PAMPs) triggered immunity (PTI) leads to genera-tion of Reactive Oxygen Species (ROS) in host plants at the site of attempted penetration.31 The role of ROS in mediating compatibility/resistance is controversial, albeit it’s known rela-tion to downstream Mitogen Activated Protein Kinase (MAPK) signaling post inoculation.32 In Chickpea-Fusarium interactive study the function of ROS further adds to the controversy of ROS being a universal resistant marker. In compatible inter-action between susceptible chickpea plant and Foc1, the ROS generation coupled with Hypersensitive Response (HR) failed to prevent fungal colonization or reproduction within the xylem tissues (Fig. 1C) while the resistant plants showed the reverse phenomenon (Fig. 1D). Previous reports suggested a few possible explanations for such a phenomenon.28,29 Firstly, single load of pathotoxins or patho-effectors often fail to alter host’s normal homeostasis. Constant flow of patho-elements is needed to opti-mize in planta pathogen establishment.33 Similarly single flush of ROS fails to impart host resistance. Conversely it makes the host vulnerable and prone to more powerful attacks.34 Thus, ROS density has direct proportionality with the extent of disease/resis-tance. Secondly, successful pathogens redirect host machinery in a direction that fails to produce ROS scavengers.32 As a result ROS mediated HR proves to be suicidal for the host itself. This assumption was well supported in our previous studies where ROS generation occurred but failed to impart resistance probably due to the insufficient or defunct ROS scavenging machinery of the susceptible host. Our future work is focused on analyzing the ROS density and its relevance in pathogen susceptibility. Lastly, ROS generation followed by HR is utilized as an option by the plant to prevent further pathogen invasion. But, the site of colo-nization is important depending on which the host decides the
between ‘guardee’ (guarded by R gene) and its corresponding avr/avr induced effectors which adds to pathogen fitness in absence of the ‘guarding R gene’ in susceptible plants or triggers innate immunity in presence of R genes in resistant plants. But the ‘guardee’ is always subjected to opposing selection forces of pro-moting or escaping interaction that ultimately controls suscepti-bility/resistance. This leads to an evolutionary unstable situation, which questions the rationale of the hypothesis. However, the lat-est proposition of ‘Decoy Model’ has provided satisfactory justi-fication from evolutionary point of view.16 ‘Decoys’ are mimics of real pathogen effector targets or ‘guardees’ preferentially evolved for enhanced pathogen effector binding. They compete with the guardee in effector binding thus lowering the pathogenic load without altering the pathogenic fitness irrespective of presence or absence of R genes.17,18 However often in the presence of R genes innate immunity is triggered following the interaction between pathogen effectors and decoys. But decoys are mainly known to restrict the pathogen rather than participate in immune signal-ing. This hypothesis was well supported by the expression of Pto in tomato following Pseudomonas syringae infestation,19 pBS3 in pepper upon Xanthomonas campestris pv vesicatoria infection,20 RCR3 in tomato upon Cladosporium fulvum attack17 and RIN4 in Arabidopsis by Psuedomonas syringae induction.21 Previous reports suggested gene duplication of actual host targets, with selective evolution of those duplicated genes to be the probable cause of origin of the ‘decoys’.19 Even though logically placed, but extensive analyses with experimental support of genetic variation of these ‘target mimics’/decoys at population level is required to strengthen the plausibility of the model.
In the present study the interaction between chickpea and wilt causing biotroph Fusarium oxysporum f.sp. ciceri (Race 1) (Foc 1) shall be discussed putting forth the involvement of prob-able decoys/guardees and emphasizing the role of sugar signaling in modulating defense.
Chickpea-Fusarium Encounter: An Overview
Unconventions ignite inquisition. Maintaining terms with the notion the unconventions of Chickpea-Fusarium interaction have triggered searchers since long. Moreover annual losses ranging from 10–15% to total, caused by wilt of this third most important pulse crop has made the management of this crop a matter of deep concern for the scientists as well as agriculturists.22 Although Foc the vascular tissue colonizing pathogen has two different pathotypes namely yellowing pathotype and wilt caus-ing pathotype, the later is known to cause wide devastation.23 Among eight pathovarieties of Foc, Race 1 has drawn the atten-tion of scientists and agronomists for its diverse geographical distribution, ecological niche and extensive destructive ability.24 Several previous reports have highlighted the limitations of clas-sical resistance breeding programs for Fusarium management.25 Additionally, transcript analyses supported by biochemical assays in Chickpea-Fusarium casesudy have shown deviation from sali-cylic acid dependent defense responses usually associated with other well studied plant-biotrophic interaction.23,25-27 Besides experimental data also suggested the expression of large number
www.landesbioscience.com Plant Signaling & Behavior 1083
approximately a 5 kDa peptide hormone that is responsible for medium alkalinization and MAPK signaling.35,36 It is produced in response to wounding and stress.37 Alkalization leads to binding of systemin to its specific receptor and triggers downstream eth-ylene and phenyl ammonia lyase (PAL) production.38,39 In addi-tion it also regulates plant growth and development.40 But how RALF mediated increment in pH regulates plant defense is still unclear. Previous studies suggested that wound mediated fungal entry was probably sensed early by resistant plants that triggered the expression of RALF.28,29 It was assumed that RALF induced medium alkalization somehow posed an adverse effect on the fungal invasion (Fig. 1E and F). Previous studies also predicted RALF to serve as probable decoys in decreasing pathogenic load without adding to pathogen fitness or activating defense signals.29 But under positive selection pressure these decoys are thought to indirectly trigger immune responses by interacting with some specific cognate R genes. Identification of such complexes from
type of molecular device to be implemented against warding off its foreign enemy. Unfortunately the crafty fungus Foc 1 chooses xylem vessels-the solute translocator of the plant, to shelter itself. This leaves no alternative for the host other than to succumb to the effects of patho-effectors. On the other hand, the fungus probably reprograms itself and utilizes its own ROS scavenging machinery to detoxify the host antimicrobials and create a self-congenial environment in planta. Hence, regulating the fungal entry appeared to be the crucial part of disease incidence/resis-tance in Chickpea-Fusarium interaction.
Interestingly, the incompatible interaction showed some strik-ing dissimilarities with that of compatible one both in terms of pathogen recognition and progression. Fusarium attack trig-gered the expression of a special peptide hormone commonly referred to as Rapid Alkalinization Factor (RALF) in resistant plants. Whereas expression of RALF in susceptible plants was significantly low throughout the pathogen progression. RALF is
Figure 1. Schematic diagram hypothesizing the role of Decoy/Guardee in mediating defense in chickpea upon Fusarium oxysporum f.sp ciceri (Race 1) challenge. PAMP, pathogen associated molecular pattern; RALF, rapid alkalinization factor; PLC, phosphoinositide specific phospholipase C; R Gene, Resistant Gene; MAPK, mitogen activated protein kinase; ROS, reactive oxygen species; HR, hypersensitive response; PCD, programmed cell death. (A) Sectional view of chickpea root; (B) Magnified view of root section showing arrangement of xylem vessels; (C) Root Section of susceptible chickpea plant showing fungal clogging; (D) Root Section of resistant chickpea plant showing clear xylem vessels; (E) Events of compatible interaction; (F) Events of incompatible interaction. The ‘X’ markings and faded shadings signify the switch off of the respective pathway(s) while the operative path-way is shown in normal visibility.

1084 Plant Signaling & Behavior Volume 5 Issue 9
Serine Threonine Kinases (STK); Receptors and Down Stream Signal Generators
Defense responses include combinations of appropriate signal per-ception, transmission and activation of cadres of genes regulating downstream reactions.61 The Serine threonine kinases (STKs) act as receptors along with other external and internal protein partners affecting wide array of processes ranging from self ver-sus non self recognition to disease resistance and developmental regulation.62 In other words they are the R genes regulating both external receptions and internal communication.63 The STK are known to regulate phosphorylation at serine and threonine resis-dues.64 They are triggered by wounding, biotic and abiotic stress factors.65,66 The well studied receptor protein of Arabidopsis FLS2 upon interaction with bacterial flagellin induces phosphorylation and activation of its intracellular kinase domain that eventually leads to a cascade of cellular events including phytoalexin, eth-ylene and salicylic acid production, ROS generation and activa-tion of pathogenesis related proteins (PR proteins).67,68 Similar responses are also documented in parsley upon elicitation.69,70 STKs are also known to regulate the sugar metabolism by revers-ibly catalyzing the generation or hexose sugars from sucrose or starch molecules.71 However the activation of STK is often medi-ated by ‘guardees’ instead of direct GFG interaction.15 Therefore these STK receptors are prone to extensive diversification prob-ably to manage the variable opposing selection pressure of the evolving pathogen and its destructive devices.72
The STKs function as ‘central processing units’ in Chickpea-Fusarium interaction. They are assumed to percept external pathogenic signals and process them into internal alarms thus triggering downstream cellular responses.29 The expression of STK in both susceptible and resistant plants presumes their role in signal conversion and transmission probably by activat-ing the sugar cleaving machinery thus generating sugar alarms in both cases (Fig. 1E and F). However enhanced levels of STK in incompatible interaction could probably explain the readiness of the resistant plants in sensing pathogen ingress and efficiently transmitting the “Danger” alarms to the cell interior.29 These STKs regulate the expression of many sugar cleaving enzymes, which predict the conditions of carbon stress as a consequence of pathogen attack.29 However identification of the interacting partners of STKs would probably help delineate their role as ‘Key Communicators’ of external and internal signals.
14.3.3s: Providing Clues to the Mechanism of Nutrient Leaching
There is no specific role attributed to the enigmatic 14.3.3s. They are associated with many diverse functions indicating their rapid evolving propensity best suited according to situation’s demand.73 They are widely involved in regulation of apoptosis, cell cycle and gene transcription.74 They are known to function as adapt-ers influencing the localization of transcription factors or expres-sion of stress inducible genes.75 These 14.3.3s are soluble acidic homo or heterodimeric proteins with each monomer having binding affinity for a specific target.76 They undergo N terminal
Fusarium induced chickpea would possibly concretize the role of RALFs as ‘True Decoys’. Thus it is presumable that RALFs served as ‘No Entry’ signal generators and transmitters for the invading fungus and somehow facilitated the priming of host plants against consecutive flushes of patho-effectors. However, only experimental proof can support the hypothesis.
Phospholipases and Cystatins; the Defense Mediators or the Victim of Attack
Phospholipase C (PLC) exists both in cytosolic and secreted forms. Several secreted PLCs have affinity for GPI anchored proteins known to have key roles in regulating viral and para-site infection.41 Upon elicitation phosphoinositide specific PLC hydrolyses the PI(4,5)P
2 into two secondary messengers IP
3 and
Diacylgycerol (DAG).42 Stimulated IP3 triggers the release of Ca2+
from intracellular stores whereas DAG concomitantly converts to phosphatidic acid (PA).43,44 PA in turn acts as a plant signal and activates MAPK/Serine threonine kinase pathway.44 PA is also known to activate plasma membrane ion channels probably for signal transmission in Cladosporium infested tomato.45 PLCs are also reported to be induced upon elicitor driven oxidative burst in soybean.46
Cystatins are proteinaceous inhibitors of cysteine proteases in animals,47 dicot and monocot plants.48,49 Most plant cystatins are 12–16 kDa protein sharing sequence homology with fam-ily 2 of animal cystatins.50 Cystatins are reported to regulate plant defense by inhibiting exogenous proteases of attacking insects and nematodes.51 Plant cystatins isolated from barley,52 soybean,53 tomato,54 sugarcane,55 wheat,56 strawberry57 and Colocasia58 are also reported to have shown antifungal activ-ity. This inspired plant scientists to exploit plant cystatins as promising targets for development of fungicides.59 Though researches indicated chestnut cystatin to be a possible inhibi-tor of fungal cell wall development,60 nonetheless the basis of interaction and the mechanism of inhibition of fungal proteases by cystatins remain elusive. Additionally researches on tomato cystatins induced upon Cladosporium fulvum attack showed the interaction between apoplastic cystatin with Cf resistant gene, providing evidence to the ‘Guard Model’ for plant pathogen communication.17
Both PLC and cystatin transcripts showed elevated expres-sion in chickpea during incompatible interaction with Foc1 predicting pathogen incited lipid and MAPK kinase signaling in resistant plants. Earlier studies suggested the involvement of more than two partners in pathogen recognition and its associ-ated downstream signaling. PLC and cystatin are presumed to be the probable ‘Guardees’ under the supervision of some still unidentified R gene controlling defense in resistant plants (Fig. 1E and F). On the other hand, these genes though expressed in suboptimal concentration in susceptible plants probably add to pathogen fitness in the absence of any monitoring/guarding R genes thus culminating in disease. Identification of specific R genes along with their interacting complexes would prob-ably help in deciphering the mode of action of guardees more convincingly.

www.landesbioscience.com Plant Signaling & Behavior 1085
invertase.93 The SAR pathway is reported to be upregulated by the overexpression of sugar cleaving enzymes.94 However only sugar production is insufficient to control signal transduction. Appropriate and uniform in planta transmission of signals is also a prerequisite for signaling. This role is performed by the sugar transporters. Monosaccharide transporters are induced in Arabidopsis upon Alternaria brassicicola and Fusarium oxysporum attack particularly to support the demand of increased carbohy-drates.95 Erysiphe induced Arabidopsis shows similar expression of sugar transporters.96 Reports suggest that the membrane recep-tors sense external signals and activate these transporters.97
The enhanced expression of sugar cleaving enzymes in incom-patible interaction between chickpea and Fusarium oxysporum f.sp ciceri suggested a vital role of sugars in mediating defense against fungal wilt. The sugars possibly helped in generating alarm sig-nals and efficiently transmitted them throughout with the aid of sugar transporters. Besides, they were assumed to activate the wound repair machinery by promoting the synthesis of cell wall polysaccharides (Fig. 1E and F). But how the entire defense sig-nal is perceived by the sugar sensing system and how the sugar alarms are uniformly transmitted throughout the stressed area ultimately triggering defense is still unanswered. Probably the localization studies of sugar cleavers and transporters could pro-vide answers to such questions.
Future Challenges: Far Way to Go
Our previous studies along with early reports of Chickpea-Fusarium interaction pointed out the involvement of a large number of known and unknown, common and unique candidate genes.27 But how these candidates probably orchestrated the entire metabolic network still remains unresolved. Aiming to solve the queries of defense responsive metabolic network we have tried to hypothesize the probable roles of ‘guardees’ and ‘decoys’ in defense mediation. We have also hypothesized the role of ‘RALF’ to operate as ‘gatekeepers’ generating the ‘no entry’ message to the fungal intruders. Besides we have also tried to focus on the function of STKs as ‘Interpreters’ translating foreign signals into internal alarms. Additionally, we have emphasized the role of sugars in signal perception, transmission and defense modula-tion (Fig. 1E and F). Nevertheless, many questions regarding the basis and mechanism of signal recognition and its appropri-ate translation into cellular responses remains to be answered. Identification and characterization of the still unidentified R genes could possibly solve the defense network puzzle.
Acknowledgements
The authors are thankful to Bose Institute, Kolkata, India for providing the infrastructural facilities. Authors are thankful to Dr. S.C. Pande and Dr. K.K. Sharma of ICRISAT, Patancheru, Andhra Pradesh, India for providing the fungal culture and chickpea seeds used for experimentations.
dimerization.77 14.3.3s are reported to regulate sugar metabolism by activating sucrose phosphate synthase.78 Alternatively they are known to regulate nitrogen metabolism by inactivating nitrate reductase.79 Besides 14.3.3s have specialized role in plant defense. 14.3.3 accumulation occurs in barley following Erysiphe graminis attack,80 in soybean after Pseudomonas syringae pv glycinea infec-tion81 and in maize upon Puccinia recondita f.sp. tritici-repentis infestation.82 Defense enzymes viz ascorbate peroxidase and caffeate-o-methyl transferase are also known to bind 14.3.3.83 Additionally they bind to the C terminal region of plasma mem-brane H+ ATPase and regulate proton pumping.84 Binding occurs following the phosphorylation of the Thr947 the ‘Gatekeeper resi-due’ of the C terminal ends of ATPase.74,85 Fusicoccin, a diter-pene wilt inducing toxin isolated from Fusicoccum amygdali was known to induce irreversible binding between 14.3.3 and H+ ATPase.74 Such interaction provided a satisfactory explanation to the irreversible opening of the stomatal pore resulting in constant nutrient leaching resulting in wilt.86
In Chickpea-Fusarium interaction enhanced expression of 14.3.3 gene during compatible interaction provided a clue to the probable mechanism of wilt (Fig. 1E and F). Although there has been no evidence of any toxin secreted by Fusarium oxysporum f.sp ciceri, identification of probable toxin(s) and its association with 14.3.3 bound ATPase shall help in deducing the mechanism of wilt and the role of 14.3.3s in chickpea defense.
Sugars: The Dynamic Players Mediating and Modulating Defense
Sugars are fuel generators essential for carbon and energy metab-olism of the cell. They also play pivotal role as signaling mol-ecules.87 Sugar metabolism is a dynamic process with metabolic fluxes and sugar concentration alterating every now and then with even slightest change in development or environment.88 Although sucrose is the major photosynthetic as well as transport product of plants, many signaling effects regulating growth and metabolism are also attributed to the hydrolytic hexose products such as glucose and fructose.87,89 The present review shall focus on the role of sugar molecules as dynamic defense signal gen-erators and transmitters. Stress management includes protection, excess production facilitating repair, uniform distribution of metabolites and preventing and priming self for facing next bout of attack. Sugars are known to efficiently regulate both biotic and abiotic stresses. Sucrose production is regulated by sugar cleav-ing enzymes like sucrose synthase, beta amylase, invertase, gly-coside hydrolases etc.89,90 Sucrose responsive element is reported to influence the expression of WRKY transcription factor during defense.91 Sucrose synthase plays a crucial role in nitrogen fixa-tion as its downregulation following stress leads to impairment in glycolytic carbon flux resulting in carbon substrate depletion.92 Besides, sugars are reported to accumulate at sites of infection or wounding particularly by the induction of cell wall and vacuolar

1086 Plant Signaling & Behavior Volume 5 Issue 9
43. Kiegle E, Moore CA, Haseloff J, Tester MA, Knight MR. Cell-type-specific calcium responses to drought, salt and cold in the Arabidopsis root. Plant J 2000; 23:267-78.
44. Munnik T, Meijer HJG, ter Reit B, van Himbergen JAJ, Hirt H, Frank W, et al. Hyperosmotic stress stimu-lates phospholipase D activity and elevates the levels of phosphatidic acid and diacylglycerol pyrophosphate. Plant J 2000; 22:147-54.
45. Legendre L, Yueh GY, Crain R, Haddock N, Heinstein PF, Low PS. Phospholipase C activation during elicita-tion of the oxidative burst in cultured plant cells. J Biol Chem 1993; 268:24559-63.
46. Joosten MHAJ, de-Wit PJGM. The tomato-Cladospo-rium fulvum interaction: a versatile experimental sys-tem to study plant-pathogen interactions. Annu Rev Phytopathol 1999; 37:335-67.
47. Anastasi A, Brown MA, Kembhavi AA, Nicklin M, Sayers C, Sunter D, et al. Cystatin, a protein inhibitor of cysteine proteinases. Biochem J 1983; 211:129-38.
48. Abe M, Abe K, Kuroda M, Arai S. Corn kernel cysteine proteinase inhibitor as a novel cystatin superfamily mem-ber of plant origin. Eur J Biochem 1992; 209:933-7.
49. Misaka T, Kuroda M, Iwabuchi K, Abe K, Arai S. Soya-cystatin, a novel cysteine proteinase inhibitor in soybean, is distinct in protein structure and gene orga-nization from other cystatins of animal or plant origin. Eur J Biochem 1996; 240:609-14.
50. Barret A. The cystatins: A new class of peptidase inhibi-tors. Trends Biochem Sci 1987; 12:193-6.
51. Haq SK, Atif SM, Khan RH. Protein proteinase inhibitor genes in combat against insects, pests and pathogens: natural and engineered phytoprotection. Arch Biochem Biophys 2004; 431:145-59.
52. Martınez M, Lopez-Solanilla E, Rodrıguez-Palenzuela P, Carbonero P, Dıaz I. Inhibition of plant-pathogenic Fungi by the barley cystatin Hv-CPI (Gene Icy) is not associated with its cysteine-proteinase inhibitory prop-erties. Mol Plant Microbe Int 2003; 16:876-83.
53. Solomon M, Belenghi B, Delledonne M, Menachem E, Levine A. The involvement of cysteine proteases and protease inhibitor genes in the regulation of programmed cell death in plants. Plant Cell 1999; 11:431-43.
54. Wu J, Haard NF. Purification and characterization of a cys-tatin from the leaves of methyl jasmonate treated tomato plants. Comp Biochem Physiol 2000; 127:209-20.
55. Soares-Costa A, Beltramini LM, Thiemann OH, Henrique-Silva F. A sugarcane cystatin: recombi-nant expression, purification and antifungal activity. Biochem Biophysical Res Comm 2002; 296:1194-9.
56. Christova PK, Christov NK, Imai RA. Cold inducible multidomain cystatin from wheat inhibits growth of the snow mold fungus, Microdochium nivale. Planta 1999; 40:69-79.
57. Martinez M, Abraham Z, Gamberdella M, Echaide M, Carbonero P, Diaz I. The strawberry gene Cyf1 encodes a phytocystatin with antifungal properties. J Exp Bot 2005; 56:1821-9.
58. Yang AH, Yeh KW. Molecular cloning, recombinant gene expression, and antifungal activity of cystatin from taro (Colocasia esculenta cv. Kaosiung no. 1). Planta 2005; 221:493-501.
59. Santino A, Poltroniere P, Mita G. Advances on plant products with potential to control toxigenic fungi—a review. Food Addit Contam 2005; 22:389-95.
60. Pernas M, Löpez-Solanilla E, Sànchez-Monge R, Salcedo G. Antifungal activity of plant cystatin. Mol Plant Microbe Int 1999; 12:624-7.
61. Gachomo EW, Shonukan OO, Kotchoni SO. The molecular initiation and subsequent acquisition of dis-ease resistance in plants. Afr J Biotech 2003; 2:26-32.
62. Goring DR, Walker JC. Plant sciences: Self rejection, a new kinase connection. Science 2004; 303:1474-5.
63. Afzal AJ, Wood AJ, Lightfoot DA. Plant receptor-like serine threonine kinases: roles in signaling and plant defense. Mol Plant Microbe Int 2008; 21:507-17.
23. Cho S, Muehlbauer FJ. Genetic effect of differentially regulated fungal response genes on resistance to necro-trophic fungal pathogens in chickpea (Cicer arietinum L.). Physiol Mol Plant Pathol 2004; 64:57-66.
24. Jimènez-Gasco MM, Navas-Cortes JA, Jimènez-Diaz RM. The Foc/Cicer a pathosystem: a case study of the evolution of plant-pathogenic fungi into races and pathotypes. Internat Microbiol 2004; 7:95-104.
25. Nimalkar SB, Harsulkar AM, Giri AP, Sainani MN, Franceshi V, Gupta VS. Differentially expressesd gene transcripts in roots of resistant and susceptible chickpea plant (Cicer arietinum L.) upon Fusarium oxysporum infection. Physiol Mol Plant Pathol 2006; 68:176-88.
26. Giri AP, Harsulkar AM, Patankar AG, Gupta VS, Sainani MN, Despande VV, et al. Association of induc-tion of protease and chitinase in chickpea roots with resistance to Fusarium oxysporum. f.sp. ciceris. Plant Pathol 1998; 47:693-9.
27. Ashraf N, Ghai D, Barman P, Basu S, Gangisetty N, Mandal MK, et al. Comparative analyses of genotype dependent expressed sequence tags and stress-respon-sive transcriptome of chickpea wilt illustrate predicted and unexpected genes and novel regulators of plant immunity. BMC Genomics 2009; 10:415.
28. Gupta S, Chakraborti D, Rangi RK, Basu D, Das S. A molecular insight into the early events of chickpea (Cicer arietinum L.) and Fusarium oxysporum f.sp ciceri (Race 1) interaction through cDNA-AFLP analysis. Phytopathol 2009; 99:1245-57.
29. Gupta S, Chakraborti D, Sengupta A, Basu D, Das S. Primary metabolism of Chickpea is the initial target of wound inducing early sensed Fusarium oxysporum f sp ciceri race 1. Plos One 2010; 5:9030.
30. Knogge W. Fungal infection of plants. Plant Cell 1996; 8:1711-22.
31. Apel K. Hirt H. Reactive oxygen species: Metabolism, oxidative stress and signal transduction. Annu Rev Plant Biol 2004; 55:373-401.
32. Hückelhoven R, Kogel KH. Reactive oxygen species in plant microbe interaction: Who is who in powdery mildew resistance? Planta 2003; 216:891-902.
33. Stanghellini ME, Rasmussen SL, Vandemark GJ. Relationship of callose deposition to resistance of let-tuce to Plasmopara lactucae-radicis. Phytopathology 1993; 83:1498-501.
34. Baker MA, Orlandi EW. Active oxygen in plant patho-genesis. Ann Rev Phytopathol 1995; 33:299-321.
35. Franssen HJ, Bisseling T. Peptide signaling in plants. Proc Natl Acad Sci USA 2001; 98:12855-6.
36. Ryan CA, Pearce G, Scheer J, Moura DS. Polypeptide hormones. Plant Cell 2002; 14:251-64.
37. Takahashi H, Ishikawa T, Kaido M, Takita K, Hayakawa T, Okazaki K, et al. Plasmodiophora brassicae-induced Cell Death and Medium Alkalization in Clubroot-resistant cultured roots of Brassica rapa. J Phytopathol 2006; 154:156-62.
38. Schaller A, Oecking C. Modulation of plasma mem-brane H+ ATPase activity differentially activates wound and pathogen defense responses in tomato plants. Plant Cell 1999; 11:263-72.
39. Haruta M, Constabel CP. Rapid alkalization factors in Popular Cell Cultures. Peptide isolation, cDNA cloning and differential expression in leaves and methyl jas-monate treated cells. Plant physiol 2003; 131:814-23.
40. Pearce G, Moura DS, Stratmann J, Ryan CA. RALF, a 5-kDa ubiquitous polypeptide in plants, arrests root growth and development. Proc Nat Acad Sci USA 2001; 98:12843-7.
41. Roberts MF. Phospholipases: structural and functional motifs for working at an interface. FASEB J 1996; 10:1159-72.
42. Munnik T, Meijer HJG. Osmotic stress activates dis-tinct lipid and MAPK signaling pathways in plants. FEBS Lett 2001; 498:172-8.
References1. Richard J, Panstruga R. Tête à tête inside a plant cell:
establishing compatibility between plants and biotrophic fungi and oomycetes. New Phytol 2006; 171:699-718.
2. Pandey SP, Somssich IE. The role of WRKY transcrip-tion factors in plant immunity. Plant Physiol 2009; 150:1648-55.
3. Baker B, Zambryski P, Staskawicz B, Kumar SPD. Signaling in plant-microbe interactions. Science 1997; 276:726-33.
4. Döehlemann G, Ramon W, Horst RJ, Voll LM, Usadel B, Poree F, et al. Reprogramming a maize plant: tran-scriptional and metabolic changes induced by the fun-gal biotroph Ustilago maydis. Plant J 2008; 56:181-95.
5. Panstruga R, Schulze-Lefert P. Live and let live: insights into powdery mildery mildew disease and resistance. Mol Plant Pathol 2002; 3:495-502.
6. Flor HH. Genetics of pathogenecity in Melampspora lini. J Agric Res 1946; 73:335-57.
7. Dangl JL, Jones JDG. Plant pathogens and integrated defense responses to infection. Nature 2001; 411:826-33.
8. Deslandes L, Olivier J, Peeters N, Feng DX, Khounlotham M, Boucher C, et al. Physical interac-tion between RRS1-R, a protein conferring resistance to bacterial wilt, and PopP2, a type III effector targeted to the plant nucleus. Proc Natl Acad Sci USA 2003; 100:8024-9.
9. Dodds PN, Lawrence GJ, Catanzariti AM, Teh T, Wang CI, Ayliffe MA, et al. Direct protein interaction underlies gene for gene specificity and coevolution of the flax resistance genes and flax rust avirulent genes. Proc Natl Acad Sci USA 2006; 103:8888-93.
10. Ueda H, Yamaguchi Y, Sano H. Direct interaction between the tobacco mosaic virus helicase domain and the ATP-bound resistance protein, N factor, during the hypersensitive response in tobacco plants. Plant Mol Biol 2006; 61:31-45.
11. Stahl EA, Dwyer G, Mauricio R, Kreitman M, Bergelson J. Dynamics of disease resistance polymor-phism at the Rpm 1 locus of Arabidopsis. Nature 1999; 400:667-71.
12. Jayakar SD. A mathematical model for interaction of gene frequencies in a parasite and its host. Theor Popul Biol 1940; 1:140-64.
13. Thompson JN. The geographic mosaic of coevolution. Chicago: University of Chicago Press 2005; 443.
14. Tellier A, Brown JKM. Stability of genetic polymor-phism in host-parasite interactions. Proc R Soc B 2007; 274:809-17.
15. van der Biezen EA, Jones JDG. Plant disease resis-tance proteins and the gene-for-gene concept. Trends Biochem Sci 1998; 30:27340-6.
16. van der Hoorn RAL, Kamoun S. From guard to decoy: A new model for perception of plant pathogen effec-tors. Plant Cell 2008; 20:2009-17.
17. Shabab M, Shindo T, Gu C, Kaschani F, Pansuriya T, Chintha R, et al. Fungal effector protein AVR2 targets diversifying defense related Cys proteases of tomato. Plant Cell 2008; 20:1169-83.
18. Zhou JM, Chai J. Plant pathogenic bacterial type III effectors subdue host responses. Curr Opin Microbiol 2008; 11:179-85.
19. Xiang T, Zhong N, Zou Y, Wu Y, Zhang J, Xing W, et al. Pseudomonas syringae effector Avr Pto blocks innate immunity by targeting receptor kinases. Curr Biol 2008; 18:74-80.
20. Kay S, Hahn S, Marois E, Hause G, Bonas U. A bac-terial effector acts as a plant transcription factor and induces a cell size regulator. Science 2007; 318:648-51.
21. Kim MG, da Cunha L, McFAll AJ, Belkhadir Y, Debroy S, Dangl JL, et al. Two Pseudomonas syringae type III effectors inhibit RIN4-regulated basal defense in Arabidopsis. Cell 2005; 121:749-59.
22. Haware MP, Nene YL. Races of Fusarium oxysporum. Plant Dis 1982; 66:809-10.

www.landesbioscience.com Plant Signaling & Behavior 1087
88. Smith AM, Zeeman SC, Smith SM. Starch degrada-tion. Annu Rev Plant Biol 2005; 56:73-98.
89. Roitsch T, Balibrea ME, Hofmann M, Proels R, Sinha AK. Extracellular invertase: key metabolic enzyme and PR protein. J Exp Bot 2003; 54:513-24.
90. Koch KE. Sucrose metabolism: regulatory mechanisms and pivotal roles in sugar sensing and plant develop-ment. Curr Opin Plant Biol 2004; 7:235-46.
91. Sun C, Palmqvist S, Olsson H, Boren M, Ahlandsberg S, Jansson C. A novel WRKY transcription factor, SUSIBA2, participates in sugar signaling in barley by binding to the sugar-responsive elements of the Iso 1 promoter. Plant Cell 2003; 15:2076-92.
92. Marino D, Hohnjec N, Kuster H, Gonzalez EM, Aresse-Igor C. Evidence for transcriptional and post translational Regulation of sucrose synthase in pea nodules by the cellular redox state. Mol Plant Microbe Int 2008; 21:622-30.
93. Herbers K, Meuwly P, Frommer WB, Mètraux JP, Sozzenwald U. Systemic acquired resistance mediated by ectopic expression of invertase: Possible Hexose sensing in the secretory pathway. Plant Cell 1996; 8:793-803.
94. Rowland LJ, Chen YC, Chourey PS. Anaerobic treat-ment alters the cell specific expression of Adh-1, Sh and Sus genes in roots of maize seedlings. Mol Gen Genet 1989; 218:33-40.
95. Truernit E, Schmid J, Epple P, Illig J, Sauer N. The sink-specific and stress regulated Arabidopsis STP4 gene: enhanced expression of a gene encoding a monosaccharide transporter by wounding, elicitors and pathogen challenge. Plant Cell 1996; 8:2169-82.
96. Hall JL, Williams LE. Assimilate transport and parti-tioning in fungal biotrophic interactions. Aust J Plant Physiol 2000; 27:549-60.
97. Özcan S, Dover J. Johnston M. Glucose sensing and signaling by two glucose receptors in the yeast Saccharomyces cerevisiae. EMBO J 1998; 17:2566-73.
77. Wu K, Lu G, Sehnke P, Ferl RJ. The heterologous inter-actions among plant 14.3.3 proteins and identification of regions that is important for dimerization. Arch Biochem Biophys 1997; 339:2-8.
78. Bachmann M, Huber JL, Athwal GS, Wu K, Ferl RJ, Huber SC. 14.3.3 proteins associate with the regulatory phosphorylation site of spinach leaf nitrate reductase in an isoform specific manner and reduce dephosphoryla-tion of Ser-543 by endogenous protein phosphatases. FEBS Lett 1996; 398:26-30.
79. Moorhead G, Douglas P, Cotelle V, Harthill J, Morrice N, Meek S, et al. Phosphorylation dependent interac-tions between enzymes of plant metabolism and 14.3.3 proteins. Plant J 1999; 18:1-12.
80. Gregerson PL, Thordal-Christensen H, Förster H, Collinge DB. Differential gene transcript accumulation in barley leaf epidermis and mesophyll in response to attack by Blumeria graminis f. sp. hordei. Physiol Mol Plant Path 1997; 51:85-97.
81. Seehaus K, Tenhaken R. Cloning of genes by mRNA differential display induced during the hypersensitive reaction of soybean after inoculation with Pseudomonas syringae pv. glycinea. Plant Mol Biol 1998; 38:1225-34.
82. Faris JD, Li WL, Liu DJ, Chen PD, Gill BS. Candidate gene analysis of quantitative disease resistance in wheat. Theor Appl Genet 1999; 98:219-25.
83. Zhang H, Wang J, Goodman HM. An Arabidopsis gene encoding a putative 14-3-3-interacting protein, caffeic acid/5-hydroxyferulic acid O-methyltransferase. Biochim Biophys Acta 1997; 1353:199-202.
84. Jahn T, Fuglsang AT, Olsson A, Brüntrup IM, Collinge DB, Volkmann D, et al. The 14.3.3 protein interacts directly with the C-terminal region of the plant plasma membrane H+ATPase. Plant Cell 1997; 9:1805-14.
85. Yaffe MB. How do 14.3.3 proteins work?-gatekeeper phosphorylation and the molecular anvil hypothesis. FEBS Lett 2002; 513:53-7.
86. Wuertele M, Ottmann CJ, Wittinghofer A, Oecking C. Structural view of a fungal toxin acting on a 14-3-3 regulatory complex. EMBO J 2003; 22:987-94.
87. Rolland F, Moore B, Sheen J. Sugar sensing and signal-ing in plants. Plant Cell 2002; 14:185-205.
64. Becraft PW. Receptor kinase signaling in plant develop-ment. Ann Rev Cell Dev Biol 2002; 18:163-92.
65. Song WJ, Yang GL, Chen LL, Kim HS, Pi LY, Holsten T, et al. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995; 270:1804-6.
66. Gomez-Gomez L, Boller T. Flagellin perception: a paradigm for innate immunity. Trends Plant Sci 2002; 7:251-6.
67. Chinchilla D, Bauer Z, Regenass M, Boller T, Felix G. The Arabidopsis receptor kinase FLS2 binds flg22 and determines the specificity of flagellin perception. Plant Cell 2006; 18:465-76.
68. Debarry M, Marten I, Ngezahayo A, Kolb HA. Differential defense responses in sweet potato suspen-sion culture. Plant Sci 2005; 168:1171-9.
69. Somssich IE. MAP kinases and plant defence. Trends Plant Sci 1997; 2:406-7.
70. Hirt H. Multiple roles of MAP kinases in plant signal transduction. Trends Plant Sci 1997; 2:11-5.
71. Purcell PC, Smith AM. Halford NG. Antisense expres-sion of a sucrose non-fermenting-1-related protein kinase sequence in potato results in decreased expres-sion of sucrose synthase in tubers and loss of sucrose-inducibility of sucrose synthase transcripts in leaves. Plant J 1998; 14:195-202.
72. Fluhr R. Sentinels of disease: Plant resistance genes. Plant Physiol 2001; 127:1367-74.
73. Ferl RJ. 14-3-3 proteins and signal transduction. Annu Rev Plant Physiol 1996; 47:49-73.
74. Fuglsang AT, Visconti S, Drumm K, Jahn T, Stensballe A, Mattei B, et al. Binding of 14.3.3 protein to the plasma membrane H+ ATPase AHA2 involves the three C terminal Residues Tyr946-Thr-Val and requires phos-phorylation of Thr947. J Biol Chem 1999; 274:36774-80.
75. Muslin AJ, Xing H. 14-3-3 proteins: regulation of subcellular localization by molecular interference. Cell Signal 2000; 12:703-9.
76. Finnie C, Andersen CH, Borch J, Gjetting S, Christensen AB, de Boer AH, et al. Do 14-3-3 proteins and plasma membrane H+-ATPases interact in the bar-ley epidermis in response to the barley powdery mildew fungus? Plant Mol Biol 2002; 49:137-47.




















