
A Solanesyl-diphosphate Synthase Localizes in Glycosomes of Trypanosoma cruzi

Pbp2x localizes separately from Pbp2b and otherpeptidoglycan synthesis proteins during later stages of celldivision of Streptococcus pneumoniae D39
Ho-Ching T. Tsui,1† Michael J. Boersma,1†
Stephen A. Vella,1 Ozden Kocaoglu,2 Erkin Kuru,2
Julia K. Peceny,1 Erin E. Carlson,2,3
Michael S. VanNieuwenhze,2,3 Yves V. Brun,1,2
Sidney L. Shaw1 and Malcolm E. Winkler1,2*1Department of Biology, 2Department of Molecular andCellular Biochemistry, and 3Department of Chemistry,Indiana University Bloomington, Bloomington, IN 47405,USA.
Summary
The relative localization patterns of class B penicillin-binding proteins Pbp2x and Pbp2b were used as posi-tional indicators of septal and peripheral (side-wall-like) peptidoglycan (PG) synthesis, respectively, in themid-cell regions of Streptococcus pneumoniae cells atdifferent stages of division. We confirm that Pbp2x andPbp2b are essential in the strain D39 genetic back-ground, which differs from that of laboratory strains.We show that Pbp2b, like Pbp2x and class A Pbp1a,follows a different localization pattern than FtsZ andremains at division septa after FtsZ reappears at theequators of daughter cells. Pulse-experiments withfluorescent D-amino acids (FDAAs) were performed inwild-type cells and in cells in which Pbp2x activity waspreferentially inhibited by methicillin or Pbp2x amountwas depleted. These experiments show that Pbp2xactivity separates from that of other PBPs to thecentres of constricting septa in mid-to-late divisionalcells resolved by high-resolution 3D-SIM microscopy.Dual-protein and protein-fluorescent vancomycin 2Dand 3D-SIM immunofluorescence microscopy (IFM) ofcells at different division stages corroborate thatPbp2x separates to the centres of septa surrounded byan adjacent constricting ring containing Pbp2b, Pbp1aand regulators, StkP and MreC. The separate localiza-tion of Pbp2x suggests distinctive roles in completingseptal PG synthesis and remodelling.
Introduction
Streptococcus pneumoniae (pneumococcus) is a humancommensal bacterium that colonizes the nasopharynx andcauses a number of serious respiratory and invasive dis-eases (Donkor, 2013; Henriques-Normark and Tuomanen,2013; Vernatter and Pirofski, 2013). Drug-resistant S.pneumoniae was recently listed by the CDC as a seriousthreat to public health in the USA (CDC, 2013). S. pneu-moniae cells are shaped liked prolate-ellipsoids referred toas ovococci that divide perpendicularly to their long axisand often form chains of cells, especially when capsule ispresent (Fig. 1A; Zapun et al., 2008b; Barendt et al., 2009;Massidda et al., 2013; Pinho et al., 2013). Pneumococcalcell shape and chaining play important roles during colo-nization and infection (Dalia and Weiser, 2011; Rodriguezet al., 2012). The ellipsoid shape of S. pneumoniae andother ovococcus bacteria is largely determined by thepeptidoglycan (PG) cell wall that surrounds these Gram (+)bacteria (Sham et al., 2012; Massidda et al., 2013; Pinhoet al., 2013). PG is composed of glycan chains that arecross-linked by PG peptides (Lovering et al., 2012; Typaset al., 2012; Egan and Vollmer, 2013). PG provides themajor resilient scaffolding for the covalent attachment ofsurface structures, including wall-teichoic acid (WTA),capsule, and surface proteins linked by sortases, many ofwhich are virulence factors (Tomasz and Fischer, 2006;Denapaite et al., 2012; Eberhardt et al., 2012; Schneewindand Missiakas, 2013). Because of its surface exposure andprokaryotic-specific mechanism of biosynthesis, PG is thetarget for many clinically important antibiotics used to treatS. pneumoniae and other bacterial infections, includingβ-lactam (e.g. penicillins and cephalosporins) and glyco-peptide (e.g. vancomycin) antibiotics (Zapun et al., 2008a;Hakenbeck et al., 2012; Sham et al., 2012; den Blaauwenet al., 2014; Nikolaidis et al., 2014).
Formation of prolate-ellipsoid-shaped cells requires twomodes of PG synthesis by penicillin-binding proteins(PBPs) accompanied by PG remodelling by PG hydrolases(Fig. 1A; Higgins and Shockman, 1970; Zapun et al.,2008b; Sham et al., 2012; Massidda et al., 2013; Pinhoet al., 2013). In ovococcus cells, equators of daughtercells become septa during division (Fig. 1A). As in other
Accepted 1 August, 2014. *For correspondence. E-mail [email protected]; Tel. (+1) 812 856 1318; Fax (+1) 812 855 6705. †Con-tributed equally to this work.
Molecular Microbiology (2014) ■ doi:10.1111/mmi.12745
© 2014 John Wiley & Sons Ltd

eubacteria, FtsZ-mediated septal PG synthesis occurs atthe mid-cell and leads to cell division (Fig. 1A; Typas et al.,2012; Egan and Vollmer, 2013; Massidda et al., 2013;Pinho et al., 2013). Unlike rod-shaped cells, ovococcuscells carry out a form of peripheral PG elongation atmid-cells instead of at side-walls (light blue, Fig. 1A).Peripheral PG synthesis accounts for the elongation ofovococci that distinguishes them from spherical-shapedcocci (Zapun et al., 2008b; Pinho et al., 2013). S. pneumo-niae and other ovococci lack MreB homologues that movein circumferential patterns around the bodies of rod-shaped bacteria and mediate lateral side-wall PG synthe-sis (Garner et al., 2011; Lee et al., 2014). Therefore,pneumococcal peripheral PG synthesis resembles prese-ptal PG side-wall synthesis that occurs at mid-cells early in
the division cycle of E. coli and other rod-shaped bacteria(see Potluri et al., 2012; van der Ploeg et al., 2013).
Septal and peripheral PG synthesis can separately beinhibited in S. pneumoniae cells (Fig. 1B). Pbp2x andPbp2b are class B (transpeptidase only) PBPs thatmediate septal and peripheral PG synthesis in S. pneumo-niae (Berg et al., 2013; Massidda et al., 2013; Fleurieet al., 2014; Peters et al., 2014) and correspond to Pbp3and Pbp2 in E. coli respectively (see Egan and Vollmer,2013; van der Ploeg et al., 2013). Specific inhibition ofPbp2x with antibiotics results in cell elongation, consistentwith inhibition of septal PG synthesis (Fig. 1B; Perez-Nunez et al., 2011; Land et al., 2013), whereas depletion ofPbp2x results in a mixture of round cells and elongatedcells, often with pointed ends (Fig. 1B; Results; Berg et al.,
GpsBdepletion
Methicillin treatment
MreCdepletion
23% 30% 12% 35%
Normal Elongated Enlarged Mixed
Pbp2x depletion
Pbp2bdepletion
50min
3h
3h
3h
4h
A B
Fig. 1. Introductory model of PG biosynthesis in ovococcus bacteria, such as S. pneumoniae, and summary of phenotypes caused bydisruption of septal and peripheral PG synthesis.A. Top. Ovococci divide perpendicularly to their long axis (Zapun et al., 2008b). Unencapsulated derivatives of wild-type S. pneumoniae formmostly diplococci and chains of two cells, whereas capsulated strains form short chains of 8–10 cells (Barendt et al., 2009). Bottom. Formationof prolate-ellipsoid-shaped bacteria requires two modes of PG synthesis, peripheral (side-wall-like) and septal PG synthesis, that occur in themid-cell regions of dividing S. pneumoniae cells (Zapun et al., 2008b; Sham et al., 2012; Massidda et al., 2013; Pinho et al., 2013). At the startof a division cycle, components of both peripheral PG synthesis complexes (orange ovals) and septal synthesis complexes (green rectangles)locate to the equators of cells (bottom). Peripheral PG synthesis (light blue; top) occurs between the future equator and septum of dividingcells and may commence before septal synthesis (Zapun et al., 2008b; Wheeler et al., 2011; Massidda et al., 2013). At some point, septal PGsynthesis (medium blue) commences to divide the cell in two. Data presented in this paper show that the complexes that carry out peripheraland septal PG synthesis locate to a large constricting ring throughout the division cycle, with the exception of Pbp2x, which moves to thecentres of septa in mid-to-late divisional cells. The grey Pac-Man corresponds to PG hydrolases that remodel the PG and allow septalseparation.B. Comparison of morphological changes of cells of an unencapsulated derivative of S. pneumoniae D39 when septal (top) or peripheral PGsynthesis (bottom) is blocked. Septal PG synthesis and ring closure is blocked by selective inhibition of Pbp2x by methicillin (Land et al.,2013), depletion of Pbp2x (Results; Berg et al., 2013; Peters et al., 2014), or GpsB depletion (Land et al., 2013), causing cell elongation.Peripheral PG synthesis is blocked by depletion of Pbp2b (Results; Berg et al., 2013), or MreC (Land and Winkler, 2011), causing formation ofsimilar-looking chains of nearly spherical cells. See text for additional details. All micrographs are at the same magnification, with scalebar = 1 μm.
2 H.-C. T. Tsui et al. ■
© 2014 John Wiley & Sons Ltd, Molecular Microbiology

2013; Peters et al., 2014). By contrast, depletion of Pbp2bleads to the formation of rounded cells in chains, consistentwith inhibition of peripheral PG synthesis (Fig. 1B; Results;Berg et al., 2013). These depletion experiments suggestthat Pbp2x and Pbp2b are essential in S. pneumoniae(Results; Berg et al., 2013; 2014; Fleurie et al., 2014;Peters et al., 2014).
Regulatory proteins can also be assigned to septal orperipheral PG synthesis based on cell-shape defectscaused by depletion and on the functions of homologues inrod-shaped bacteria (Zapun et al., 2008b; Massidda et al.,2013). Depletion of the essential MreC or MreD protein inthe D39 genetic background of S. pneumoniae results information of chains of spherical cells remarkably similar tothose formed by blocking peripheral PG synthesis byPbp2b depletion (Fig. 1B; Land and Winkler, 2011; Berget al., 2013). In the D39 genetic background, GpsB isessential, and depletion of GpsB causes formation ofenlarged, elongated cells unable to close division septa(Fig. 1B; Land et al., 2013). However, bypass suppressormutations have become a major complication in determin-ing primary phenotypes of regulatory proteins in pneumo-coccal cell division. Strain D39 is the encapsulated virulentserotype 2 progenitor strain of most unencapsulated labo-ratory strains, which have accumulated over 80 mutationsthat change metabolism and PG composition (Lanie et al.,2007; Carvalho et al., 2013). Neither MreCD nor GpsB isessential in laboratory strains (Land and Winkler, 2011;Fleurie et al., 2014), which contain suppressors that mayalter pathways of cell division and PG biosynthesis. Inother instances, bypass suppressors likely have accumu-lated during mutant construction. For example, the StkPserine/threonine kinase certainly regulates some stage ofcell division of S. pneumoniae (Beilharz et al., 2012;Fleurie et al., 2012), but ΔstkP mutants exhibit a variety ofmorphology defects, due in part to suppressor accumula-tion (see Massidda et al., 2013).
Despite complications from suppressor accumulation,Pbp2x, Pbp2b, and some regulatory proteins can beassigned separable functions in septal or peripheral PGsynthesis. Therefore, it is likely that separate complexes ofsome components are present at the mid-cells of dividingpneumococcal cells (Fig. 1A). However, it is unclearwhether some components, such as class A (dual trans-glycosylase and transpeptidase) Pbp1a, which has beenimplicated in peripheral PG synthesis (Land and Winkler,2011), also participate in septal PG synthesis (Massiddaet al., 2013; Peters et al., 2014) and whether septal andperipheral PG synthesis complexes are intermixed, adja-cent, or physically separate during different stages ofdivision (Zapun et al., 2008b; Land et al., 2013; Massiddaet al., 2013; Fleurie et al., 2014). Moreover, measure-ments of dimensions of dividing pneumococcal cells indi-cated that peripheral and septal PG synthesis occur
simultaneously over much of the pneumococcal cell cycle,although there may be a short period of preseptal PGperipheral synthesis before the start of septal PG synthesis(Wheeler et al., 2011). However, the extent of switchingand co-ordination between peripheral and septal PG syn-thesis remain largely unknown.
Most localization studies of Pbp2x, Pbp2b, and Pbp1ahave been performed using 2D immunofluorescencemicroscopy (IFM) or epifluorescence microscopy of livecells containing fluorescent-protein fusions (resolution≈ 250 nm; Morlot et al., 2003; Zapun et al., 2008b; Fleurieet al., 2014; Peters et al., 2014). In these studies, numbersof cells showing localization patterns were counted. Bythese criteria, Pbp2x, Pbp2b, Pbp1a and other PG synthe-sis proteins appear to colocalize at all stages of cell divi-sion. In a previous study, we used an image analysisgraphical user interface (IMA-GUI) to quantify averagefluorescence intensities of immunolabelled proteins alongthe long axis of images of cells at different division stagesobtained by dual-protein 2D IFM (Land et al., 2013). Thisanalysis revealed that FtsZ and GpsB have overlapping,but not identical, patterns of localization. Notably, FtsZ andGpsB migrated from mid-cell septa to equators of daughtercells at an earlier stage of division than Pbp2x and Pbp1a,consistent with other reports (Morlot et al., 2003; Zapunet al., 2008b). IMA-GUI analysis also revealed that theaverage diameter of Pbp2x rings appeared to be smallerthan that of Pbp1a rings in mid-divisional cells (Land et al.,2013). Results from 2D analyses of FtsZ, GpsB, Pbp2x andPbp1a localization were confirmed and extended by high-resolution, dual-protein 3D-SIM (structured illuminationmicroscopy) IFM (resolution XY ≈ 100 nm; Z ≈ 250 nm;Land et al., 2013). 3D-SIM images showed that Pbp2x andPbp1a colocalize to an equatorial ring in early-to-mid divi-sional cells, but Pbp2x separates from the ring containingPbp1a and locates as a disk at the centre of the septum inmid-to-late divisional cells (Land et al., 2013).
In this paper, we report the use of fluorescent D-aminoacid (FDAA) probes (Kuru et al., 2012) to independentlydemonstrate that Pbp2x locates separately from otherPBPs in mid-to-late stages of pneumococcal cell division.An additional tool was added to quantify dual-protein 2DIFM images to show that Pbp2x localizes separately fromPbp1a and other PG synthesis proteins, including Pbp2b,MreC, and StkP, at a later stage of division. 3D-SIM IFMcorroborated that PG synthesis proteins remain together ina ring structure until a later stage of cell division, after whichPbp2x locates to the centre of division septa, wherePG-pentapeptides labelled by fluorescent-vancomycin(FL-V) concentrate. These results show that pneumococ-cal PG synthesis proteins colocalize contiguously in aconstricting ring through the early and middle stages ofpneumococcal cell division; but, separate protein com-plexes in these constricting rings could not be resolved at
Separation of pneumococcal Pbp2x from Pbp2b at septa 3
© 2014 John Wiley & Sons Ltd, Molecular Microbiology

the resolution of 3D-SIM. The exceptional localization ofPbp2x to the centres of septa in mid-to-late divisional cellssuggests that the PG transpeptidase cross-linking activityof Pbp2x functions not only in septal PG synthesis, but mayhave other roles in PG remodelling.
Results
pbp2x and pbp2b are essential and mediate septaland peripheral PG synthesis, respectively, inS. pneumoniae D39
Tracking the localization patterns of the primary class BPBPs has been an effective strategy to study preseptal(PBP2) and septal (PBP3) PG synthesis in E. coli (van derPloeg et al., 2013). Based on this precedent, we performedstudies of Pbp2x (septal) and Pbp2b (peripheral) localiza-tion in S. pneumoniae. Recent studies have shown thatPbp2x and Pbp2b are essential in laboratory strains of S.pneumoniae and their depletion causes distinctive cellmorphologies (Berg et al., 2013; Peters et al., 2014). Weperformed these studies in the strain D39 genetic back-ground that lacks suppressors found in laboratory strains(Introduction; Land and Winkler, 2011). We first needed toconfirm the essentiality of Pbp2x and Pbp2b and theirdepletion phenotypes in the D39 background. The capsuleof strain D39 causes the formation of short chains of cells,whereas unencapsulated (Δcps) derivatives of D39 formsingle and diplococcal cells that are highly amenable tobinning retrospectively into cell division stages (Fig. 1A;Tsui et al., 2011; Land et al., 2013). Hence, this study wasperformed using an isogenic Δcps derivative of strain D39(Table S1). A merodiploid D39 Δcps (Δpbp2x//PfcsK-pbp2x+)strain was constructed to control Pbp2x amount from anectopic fucose-inducible promoter (Chan et al., 2003; Landet al., 2013; Table S1). Depletion of Pbp2x upon removal offucose led to cessation of growth (Fig. S1A) and formationof cells with a variety of abnormal shapes, including elon-gated cells with pointed ends (3 h, Fig. S1B; below). Asdepletion continued, Pbp2x-depleted cells enlarged,rounded up and lysed. This pointed-cell phenotype in theD39 strain is similar to that reported for pneumococcallaboratory strains (Berg et al., 2013; Peters et al., 2014),and contrasts with the formation of elongated rod-shapedcells when Pbp2x activity is inhibited preferentially bymethicillin (below), as noted before (Land et al., 2013).
A D39 Δcps (Δpbp2b//PfcsK-pbp2b+) merodiploid strainwas also constructed for depletion of Pbp2b (Table S1).Depletion of Pbp2b upon removal of fucose caused ces-sation of growth of the D39 strain (Fig. S2A) and formationof chains of rounded cells (see below; Fig. S2B), similar tothose reported for pneumococcal laboratory strains (Berget al., 2013). Whisker-plot analysis showed that the celllength of Pbp2b cells remains the same, but the cell widths
increase until the cells become nearly spherical (Fig. S2C).Following prolonged incubation, D39 cells depleted ofPbp2b assume irregular shapes and lyse (data not shown),similar to the laboratory strains. We conclude that Pbp2xand Pbp2b are essential in the D39 genetic background,with depletion causing similar cell defects in septal andperipheral PG synthesis, respectively, and autolysis, aswas reported in laboratory strains (Berg et al., 2013; Peterset al., 2014). Depletion of Pbp2x or Pb2b is used in thepulse-labelling experiments with FDAAs described below.
Pbp2b and MreC, like Pbp2x and Pbp1a, localizedifferently from FtsZ at later division stages
An early conclusion that Pbp2b localized exclusively topneumococcal cell equators (Morlot et al., 2003) was latermodified based on additional 2D IFM data to colocalizationof Pbp2b with Pbp2x and other PBPs at division equatorsand septa (Zapun et al., 2008b). A recent study concludesthat FtsZ-RFP colocalizes at mid-cells with GFP-Pbp2xand GFP-Pbp2b (Fleurie et al., 2014). Our previous work(Land et al., 2013) extended earlier work (Morlot et al.,2003) to show that Pbp2x and Pbp1a follow a differentlocalization pattern than FtsZ and GpsB at later stages ofdivision. To distinguish among these patterns of localiza-tion of Pbp2b, we performed dual-protein 2D IFM and3D-SIM IFM to track Pbp2b and FtsZ in the same cells atdifferent stages of pneumococcal cell division (Wayneet al., 2010; Tsui et al., 2011; Land et al., 2013). In parallel,we performed similar studies on MreC, which like Pbp2b,mediates pneumococcal peripheral PG synthesis (Landand Winkler, 2011; Massidda et al., 2013). Proteins weretagged with short epitope peptides (FLAG, Myc or HA),expressed from single-copy genes in native chromosomalloci, and strains were checked for absence of tag cleavage,lack of cell morphology and growth defects, and normalPBP activity by staining with fluorescent bocillin (Boc-FL;Fig. S3; Land et al., 2013). Localization patterns wereconfirmed using different epitope tags (Experimental pro-cedures). Because S. pneumoniae cells divide perpen-dicularly to their long axis and remain initially attached asdiplococci (Fig. 1A), IFM images of cells can be sortedretrospectively into division stages (Figs 2A and S4; Tsuiet al., 2011; Land et al., 2013). Protein intensities wereaveraged in 2D IFM images of cells at different divisionstages and compared using the IMA-GUI reported previ-ously (Fig. 2A; Land et al., 2013). Similar to Pbp2x andPbp1a (Fig. S4; Land et al., 2013), Pbp2b and MreClocalize differently from FtsZ in mid-to-late divisional cells.In pre- and early-divisional cells (stages 1 and 2, Figs 2Aand S5A), Pbp2b, Pbp2x, Pbp1a and MreC colocalize as aring with FtsZ. In mid-to-late divisional cells (stage 3), a ringof Pbp2b, Pbp2x, Pbp1a, MreC, and some FtsZ constrictsat division septa, while most FtsZ appears to migrate to the
4 H.-C. T. Tsui et al. ■
© 2014 John Wiley & Sons Ltd, Molecular Microbiology
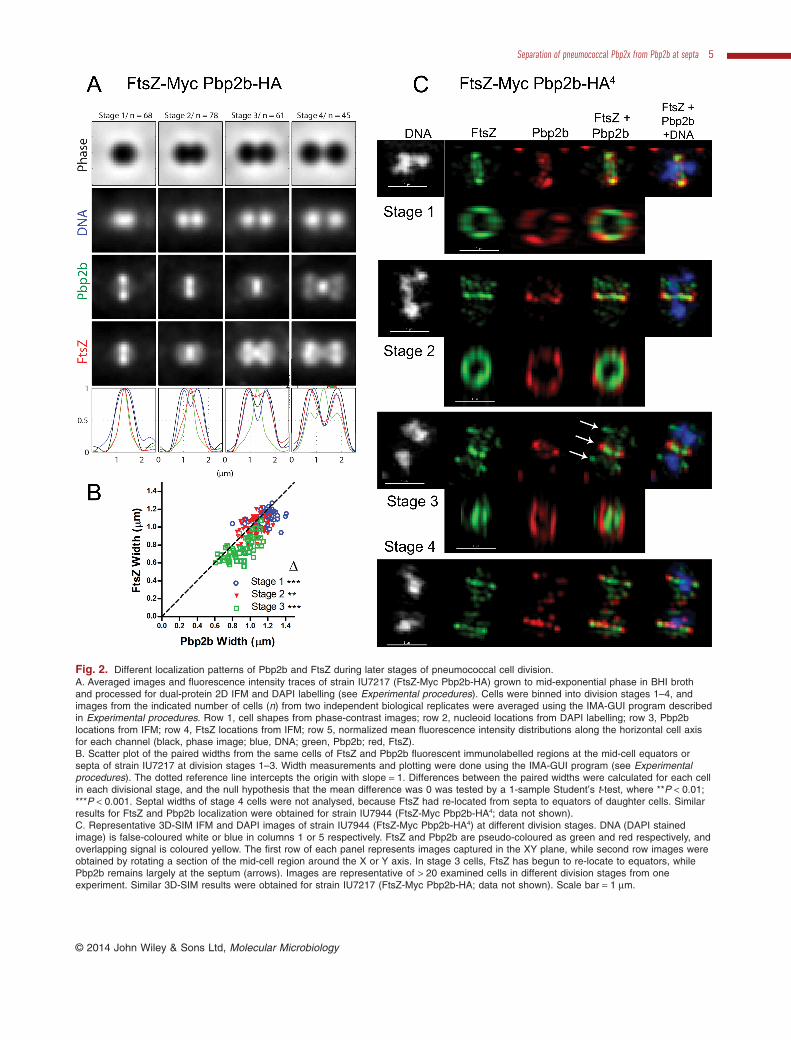
Fig. 2. Different localization patterns of Pbp2b and FtsZ during later stages of pneumococcal cell division.A. Averaged images and fluorescence intensity traces of strain IU7217 (FtsZ-Myc Pbp2b-HA) grown to mid-exponential phase in BHI brothand processed for dual-protein 2D IFM and DAPI labelling (see Experimental procedures). Cells were binned into division stages 1–4, andimages from the indicated number of cells (n) from two independent biological replicates were averaged using the IMA-GUI program describedin Experimental procedures. Row 1, cell shapes from phase-contrast images; row 2, nucleoid locations from DAPI labelling; row 3, Pbp2blocations from IFM; row 4, FtsZ locations from IFM; row 5, normalized mean fluorescence intensity distributions along the horizontal cell axisfor each channel (black, phase image; blue, DNA; green, Pbp2b; red, FtsZ).B. Scatter plot of the paired widths from the same cells of FtsZ and Pbp2b fluorescent immunolabelled regions at the mid-cell equators orsepta of strain IU7217 at division stages 1–3. Width measurements and plotting were done using the IMA-GUI program (see Experimentalprocedures). The dotted reference line intercepts the origin with slope = 1. Differences between the paired widths were calculated for each cellin each divisional stage, and the null hypothesis that the mean difference was 0 was tested by a 1-sample Student’s t-test, where **P < 0.01;***P < 0.001. Septal widths of stage 4 cells were not analysed, because FtsZ had re-located from septa to equators of daughter cells. Similarresults for FtsZ and Pbp2b localization were obtained for strain IU7944 (FtsZ-Myc Pbp2b-HA4; data not shown).C. Representative 3D-SIM IFM and DAPI images of strain IU7944 (FtsZ-Myc Pbp2b-HA4) at different division stages. DNA (DAPI stainedimage) is false-coloured white or blue in columns 1 or 5 respectively. FtsZ and Pbp2b are pseudo-coloured as green and red respectively, andoverlapping signal is coloured yellow. The first row of each panel represents images captured in the XY plane, while second row images wereobtained by rotating a section of the mid-cell region around the X or Y axis. In stage 3 cells, FtsZ has begun to re-locate to equators, whilePbp2b remains largely at the septum (arrows). Images are representative of > 20 examined cells in different division stages from oneexperiment. Similar 3D-SIM results were obtained for strain IU7217 (FtsZ-Myc Pbp2b-HA; data not shown). Scale bar = 1 μm.
Separation of pneumococcal Pbp2x from Pbp2b at septa 5
© 2014 John Wiley & Sons Ltd, Molecular Microbiology
equators of daughter cells. In late-divisional cells (stage 4),Pbp2b, Pbp2x, Pbp1a, and MreC remain at highly con-stricted septa, but the majority of these proteins locate withFtsZ at equators. 3D-SIM IFM confirmed that Pbp2b andMreC localize differently than FtsZ in mid-to-late divisionalcells (Figs 2C and S5C).
The widths of Pbp2b, Pbp2x, Pbp1a and MreC septalrings appeared to be larger than that of FtsZ in averaged2D IFM images of cells at different division stages (Figs 2A,S4 and S5A; Land et al., 2013). We added a quantificationfunction to the IMA-GUI program to measure the widths ofseptal rings in 2D IFM images of cells (see Experimentalprocedures). Septal widths of different proteins rangedfrom ≈ 1.2 μm in stage 1 cells to ≈ 0.66 μm in stage 4 cells.Paired septal widths of two different proteins detected inthe same cell were plotted against each other (Figs 2B, S4and S5B; Experimental procedures). The points on scatterplots of FtsZ versus Pbp2b or MreC septal ring widthsmostly fall below the diagonal line, indicating that diam-eters of FtsZ septal rings are smaller than those of Pbp2bor MreC septal rings at division stages 1–3. To statisticallyanalyse these data, we took the average differences of theFtsZ and Pbp2b or MreC septal ring widths and performeda Student’s t-test for the null hypothesis that the differenceis zero. For cells at division stages 1–3, the Pbp2b andMreC septal rings are significantly larger than the FtsZ ring(Fig. 2B). This graphical and statistical method is usedthroughout this paper. A difference in ring diameters wasconfirmed by 3D-SIM IFM (Figs 2C and S5C), where theapparent separation of FtsZ and these other proteinsacross the membrane in rotated mid-cell images is likelyenhanced by the width of primary and fluorescent second-ary antibodies (≈ 10 nm each) bound to each protein.Taken together, the 2D and 3D-SIM IFM results are con-sistent with the cytoplasmic location of FtsZ compared tothe extracellular location of the epitope tags at theC-terminus of the PBPs and MreC (Land and Winkler,2011; Land et al., 2013).
Pulse-labelling with FDAAs demonstrates that Pbp2xactivity separates from that of other PBPs in mid-to-latedivisional cells
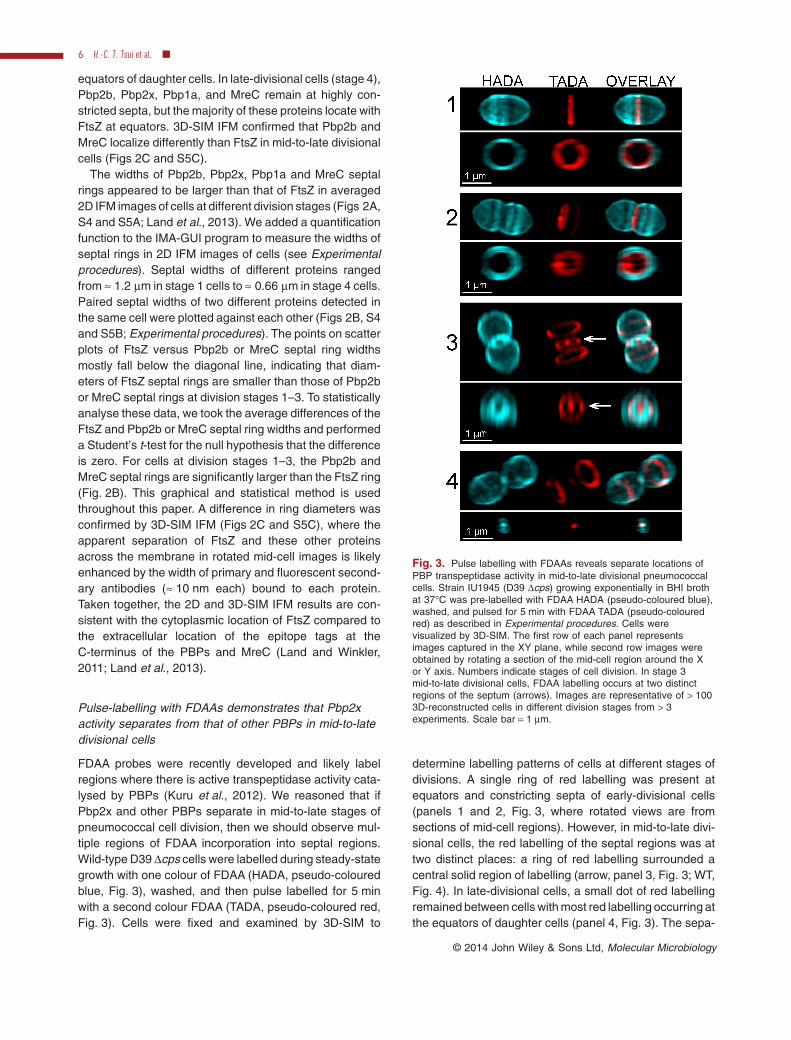
FDAA probes were recently developed and likely labelregions where there is active transpeptidase activity cata-lysed by PBPs (Kuru et al., 2012). We reasoned that ifPbp2x and other PBPs separate in mid-to-late stages ofpneumococcal cell division, then we should observe mul-tiple regions of FDAA incorporation into septal regions.Wild-type D39 Δcps cells were labelled during steady-stategrowth with one colour of FDAA (HADA, pseudo-colouredblue, Fig. 3), washed, and then pulse labelled for 5 minwith a second colour FDAA (TADA, pseudo-coloured red,Fig. 3). Cells were fixed and examined by 3D-SIM to
determine labelling patterns of cells at different stages ofdivisions. A single ring of red labelling was present atequators and constricting septa of early-divisional cells(panels 1 and 2, Fig. 3, where rotated views are fromsections of mid-cell regions). However, in mid-to-late divi-sional cells, the red labelling of the septal regions was attwo distinct places: a ring of red labelling surrounded acentral solid region of labelling (arrow, panel 3, Fig. 3; WT,Fig. 4). In late-divisional cells, a small dot of red labellingremained between cells with most red labelling occurring atthe equators of daughter cells (panel 4, Fig. 3). The sepa-
Fig. 3. Pulse labelling with FDAAs reveals separate locations ofPBP transpeptidase activity in mid-to-late divisional pneumococcalcells. Strain IU1945 (D39 Δcps) growing exponentially in BHI brothat 37°C was pre-labelled with FDAA HADA (pseudo-coloured blue),washed, and pulsed for 5 min with FDAA TADA (pseudo-colouredred) as described in Experimental procedures. Cells werevisualized by 3D-SIM. The first row of each panel representsimages captured in the XY plane, while second row images wereobtained by rotating a section of the mid-cell region around the Xor Y axis. Numbers indicate stages of cell division. In stage 3mid-to-late divisional cells, FDAA labelling occurs at two distinctregions of the septum (arrows). Images are representative of > 1003D-reconstructed cells in different division stages from > 3experiments. Scale bar = 1 μm.
6 H.-C. T. Tsui et al. ■
© 2014 John Wiley & Sons Ltd, Molecular Microbiology

ration of the FDAA labelling into two distinct regions at thesepta of mid-to-late divisional cells could not be resolvedby standard 2D fluorescence microscopy without imagerotation (data not shown), as was tried previously (Fleurieet al., 2014). We conclude that the PBP transpeptidaseactivities separate to two locations in the septa of mid-to-late divisional pneumococcal cells.
Two experiments demonstrate that the central septallabelling with FDAAis due to Pbp2x transpeptidase activity.Previously we showed that addition of 0.1 μg ml−1 methi-cillin preferentially inhibited > 80% of Pbp2x transpepti-dase activity [with < 20% inhibition of Pbp3 (DacA)transpeptidase activity] and caused cell elongation(Fig. 1B; Land et al., 2013). Therefore, we treated wild-type cells with methicillin before pulse labelling with TADAFDAA (Fig. 4). Untreated mid-to-late divisional cellsshowed central septal labelling with TADA surrounded by aring of labelling in > 80% of ≈ 60 examined cells (Fig. 4, left,where the rotated views are from ends of cells and alsoshow labelled equatorial rings). In contrast, methicillintreatment allowed labelling of rings, but abolished centralseptal labelling with TADA in > 98% of ≈ 60 examined cells
(Fig. 4, right). Since Pbp2x is preferentially inhibited inthese cells (Land et al., 2013) and Pbp3 (DacA) does notlocalize to septa (Morlot et al., 2004; Barendt et al., 2011),these results support the conclusion that central septallabelling is catalysed by the transpeptidase activity of classB Pbp2x.
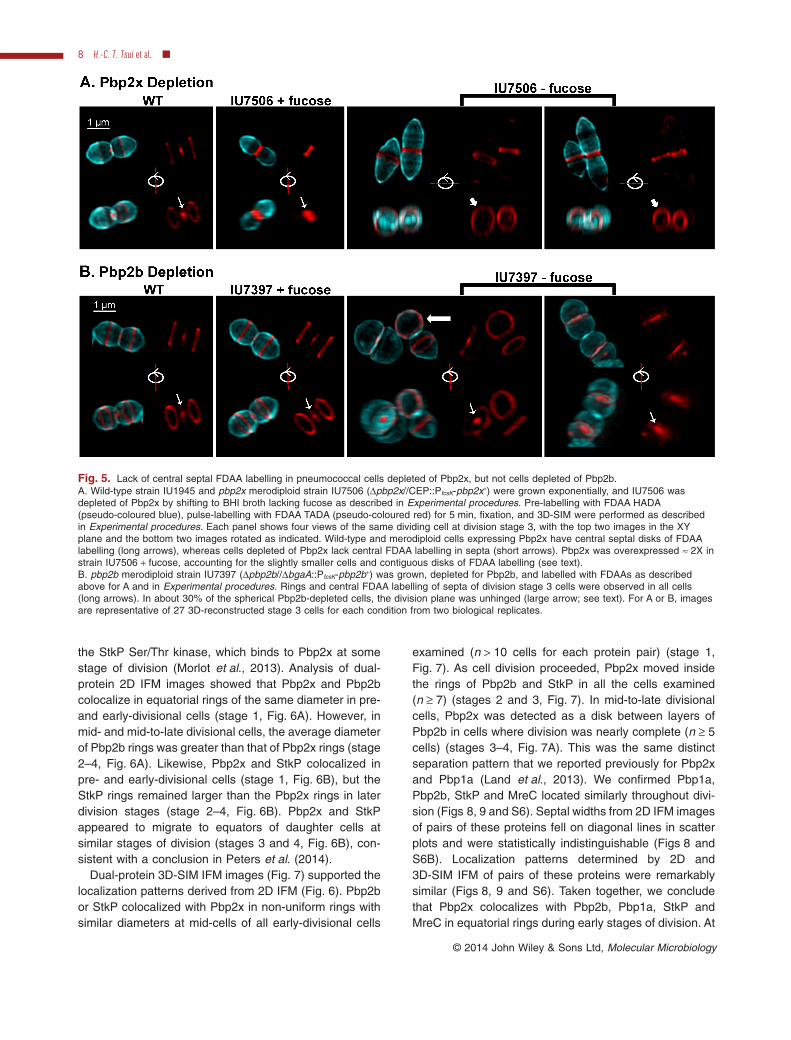
In a second experiment, we depleted Pbp2x beforepulse labelling with the TADA FDAA (Fig. 5). pbp2x mer-odiploid cells (Δpbp2x//PfcsK-pbp2x+) grown in 1% (w/v)fucose-containing medium were somewhat smaller thanwild-type cells, and more irregular cells were observed(Figs 5A and S1). PBPs were labelled to saturation withBoc-FL (Experimental procedures), which revealed thatPbp2x was overexpressed by ≈ 2-fold in this merodiploidstrain (data not shown). In contrast to the two-positionlabelling in most (88%) wild-type cells (Figs 3 and 4),overexpression of Pbp2x resulted in TADA labelling(pseudo-coloured red) as contiguous disks (78%) ornearly contiguous thickened rings and dots (22%) at thesepta of mid-to-late divisional merodiploid cells (Fig. 5A,panel IU7506 + fucose). These results were confirmedby overexpression of Pbp2x in a pbp2x+ merodiploidstrain (pbp2x+//PfcsK-pbp2x+) grown in 1% (w/v) fucose-containing medium (data not shown). Removal of fucosefrom the pbp2x merodiploid strain (Δpbp2x//PfcsK-pbp2x+)for ≈ 3 h depleted the relative Pbp2x amount to < 20%based on Boc-FL labelling (data not shown). Pbp2xdepletion led to the formation of enlarged, pointed cellscontaining septal bands without significant central septalTADA labelling in > 90% of 27 examined cells (Fig. 5A,right panels). As a control experiment, TADA pulse label-ling was performed in pbp2b merodiploid cells (Δpbp2b//PfcsK-pbp2b+) depleted of Pbp2b (Fig. 5B). In this case,pbp2b merodiploid cells grown in fucose resembled wild-type cells. Spherical cells depleted of Pbp2b still showedcentral septal labelling, consistent with active Pbp2xactivity (Fig. 5B, right). Unexpectedly, in ≈ 30% of ≈ 100examined spherical Pbp2b-depleted cells, the relativeorientation of the division planes in sister cells wasrotated instead of being parallel (Fig. 5B, third panel).Together, these results indicate that inhibition of Pbp2xactivity by methicillin or depletion of Pbp2x abrogatesthe FDAA labelling at the centres of septa, supportingthe conclusion that this labelling is due to the transpepti-dase activity of Pbp2x, which locates to septal centresin mid-to-late divisional cells (see below; Land et al.,2013).
Pbp2x protein separates from other PG synthesisproteins besides Pbp1a in mid-to-late division cells
Because of their distinct roles in septal and peripheral PGsynthesis, we localized Pbp2x and Pbp2b in the samecells. We also tracked the localization of Pbp2x relative to
Fig. 4. Lack of central septal FDAA labelling in cells treated witha concentration of methicillin that preferentially inhibits thetranspeptidase activity of pneumococcal Pbp2x. Cultures ofexponentially growing wild-type strain IU1945 (D39 Δcps) weresplit, duplicate cultures were treated with methicillin (0.1 μg ml−1) for40 min, and treated and untreated cultures were pulse labelled for5 min with FDAA TADA (pseudo-coloured red; see Experimentalprocedures). Two views of 3D-SIM images of untreated (left) andmethicillin-treated (right) cells are shown. The first column of eachcondition is in the XY plane, and the second column is a rotation ofthe entire cell around X or Y axis as indicated by the line inside thecircular arrows. Methicillin-treated cells are elongated and ringslack the central septal disks of FDAA labelling seen in untreatedcells (arrows). Images are representative of > 60 3D-reconstructedstage-3 cells for each condition from two biological replicates.Scale bar = 1 μm.
Separation of pneumococcal Pbp2x from Pbp2b at septa 7
© 2014 John Wiley & Sons Ltd, Molecular Microbiology
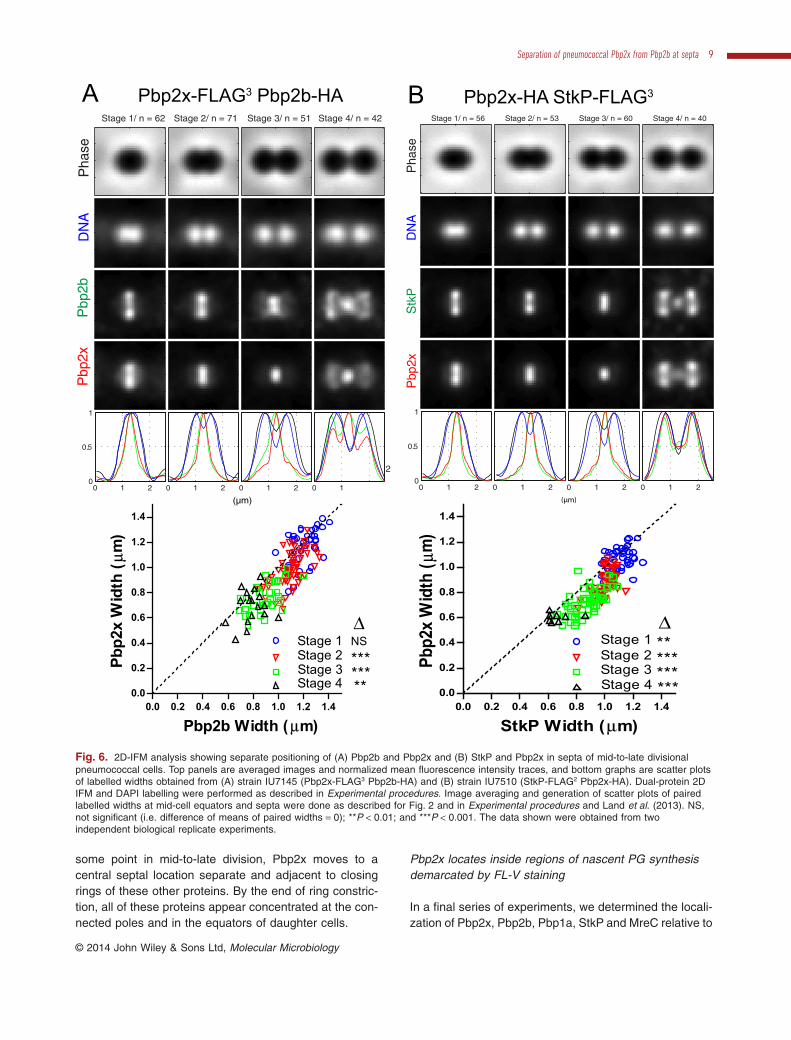
the StkP Ser/Thr kinase, which binds to Pbp2x at somestage of division (Morlot et al., 2013). Analysis of dual-protein 2D IFM images showed that Pbp2x and Pbp2bcolocalize in equatorial rings of the same diameter in pre-and early-divisional cells (stage 1, Fig. 6A). However, inmid- and mid-to-late divisional cells, the average diameterof Pbp2b rings was greater than that of Pbp2x rings (stage2–4, Fig. 6A). Likewise, Pbp2x and StkP colocalized inpre- and early-divisional cells (stage 1, Fig. 6B), but theStkP rings remained larger than the Pbp2x rings in laterdivision stages (stage 2–4, Fig. 6B). Pbp2x and StkPappeared to migrate to equators of daughter cells atsimilar stages of division (stages 3 and 4, Fig. 6B), con-sistent with a conclusion in Peters et al. (2014).
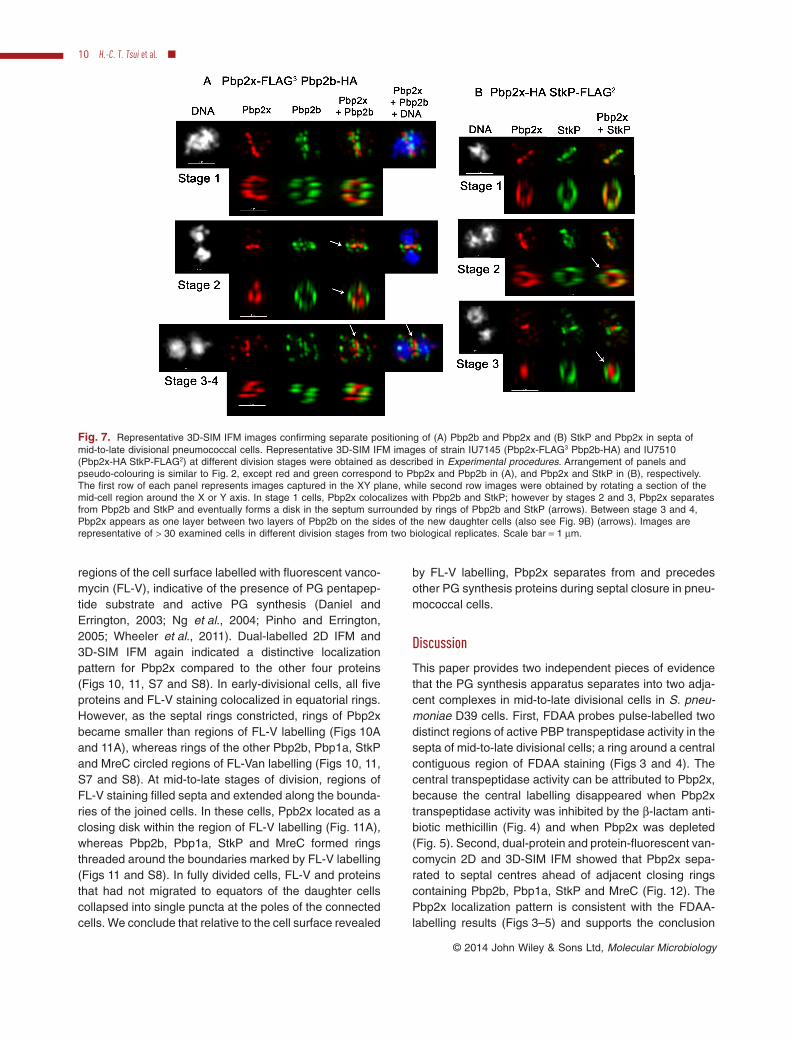
Dual-protein 3D-SIM IFM images (Fig. 7) supported thelocalization patterns derived from 2D IFM (Fig. 6). Pbp2bor StkP colocalized with Pbp2x in non-uniform rings withsimilar diameters at mid-cells of all early-divisional cells
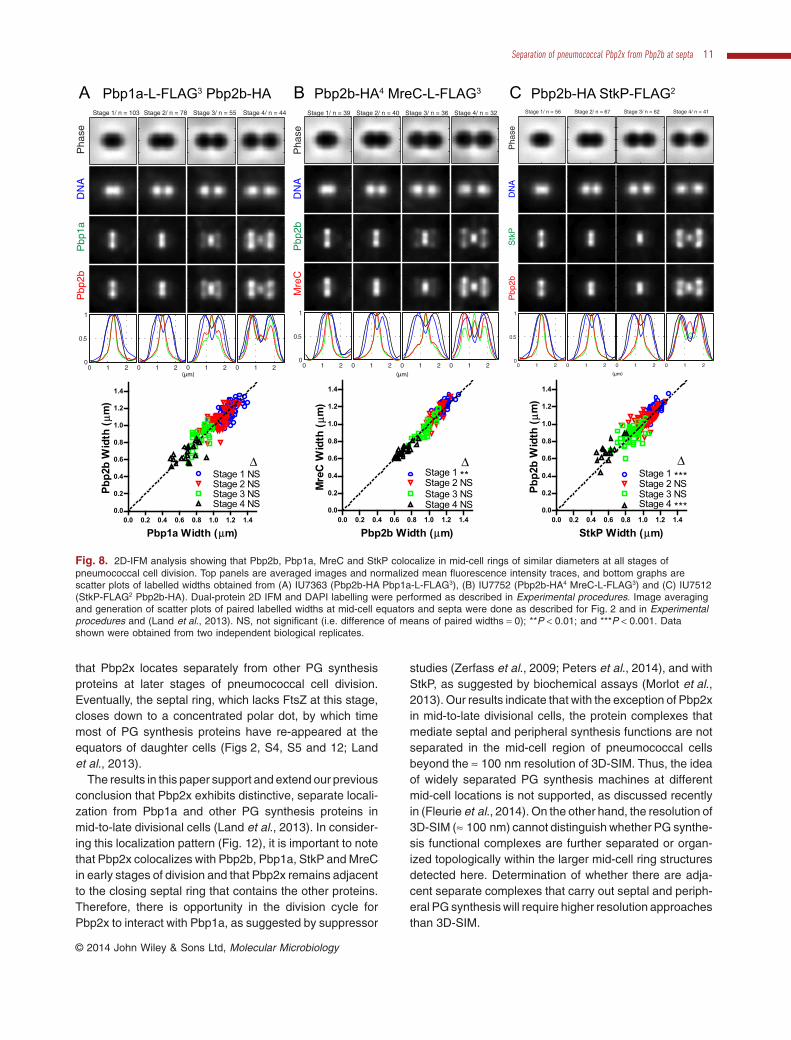
examined (n > 10 cells for each protein pair) (stage 1,Fig. 7). As cell division proceeded, Pbp2x moved insidethe rings of Pbp2b and StkP in all the cells examined(n ≥ 7) (stages 2 and 3, Fig. 7). In mid-to-late divisionalcells, Pbp2x was detected as a disk between layers ofPbp2b in cells where division was nearly complete (n ≥ 5cells) (stages 3–4, Fig. 7A). This was the same distinctseparation pattern that we reported previously for Pbp2xand Pbp1a (Land et al., 2013). We confirmed Pbp1a,Pbp2b, StkP and MreC located similarly throughout divi-sion (Figs 8, 9 and S6). Septal widths from 2D IFM imagesof pairs of these proteins fell on diagonal lines in scatterplots and were statistically indistinguishable (Figs 8 andS6B). Localization patterns determined by 2D and3D-SIM IFM of pairs of these proteins were remarkablysimilar (Figs 8, 9 and S6). Taken together, we concludethat Pbp2x colocalizes with Pbp2b, Pbp1a, StkP andMreC in equatorial rings during early stages of division. At
Fig. 5. Lack of central septal FDAA labelling in pneumococcal cells depleted of Pbp2x, but not cells depleted of Pbp2b.A. Wild-type strain IU1945 and pbp2x merodiploid strain IU7506 (Δpbp2x//CEP::PfcsK-pbp2x+) were grown exponentially, and IU7506 wasdepleted of Pbp2x by shifting to BHI broth lacking fucose as described in Experimental procedures. Pre-labelling with FDAA HADA(pseudo-coloured blue), pulse-labelling with FDAA TADA (pseudo-coloured red) for 5 min, fixation, and 3D-SIM were performed as describedin Experimental procedures. Each panel shows four views of the same dividing cell at division stage 3, with the top two images in the XYplane and the bottom two images rotated as indicated. Wild-type and merodiploid cells expressing Pbp2x have central septal disks of FDAAlabelling (long arrows), whereas cells depleted of Pbp2x lack central FDAA labelling in septa (short arrows). Pbp2x was overexpressed ≈ 2X instrain IU7506 + fucose, accounting for the slightly smaller cells and contiguous disks of FDAA labelling (see text).B. pbp2b merodiploid strain IU7397 (Δpbp2b//ΔbgaA::PfcsK-pbp2b+) was grown, depleted for Pbp2b, and labelled with FDAAs as describedabove for A and in Experimental procedures. Rings and central FDAA labelling of septa of division stage 3 cells were observed in all cells(long arrows). In about 30% of the spherical Pbp2b-depleted cells, the division plane was unhinged (large arrow; see text). For A or B, imagesare representative of 27 3D-reconstructed stage 3 cells for each condition from two biological replicates.
8 H.-C. T. Tsui et al. ■
© 2014 John Wiley & Sons Ltd, Molecular Microbiology
some point in mid-to-late division, Pbp2x moves to acentral septal location separate and adjacent to closingrings of these other proteins. By the end of ring constric-tion, all of these proteins appear concentrated at the con-nected poles and in the equators of daughter cells.
Pbp2x locates inside regions of nascent PG synthesisdemarcated by FL-V staining
In a final series of experiments, we determined the locali-zation of Pbp2x, Pbp2b, Pbp1a, StkP and MreC relative to
Stage 1/ n = 56
Pha
seD
NA
Stk
PP
bp2x
0 1 20
0.5
1
Stage 2/ n = 53
0 1 2
Stage 3/ n = 60
0 1 2
Stage 4/ n = 40
0 1 2
(μm)
Stage 1/ n = 62
Pha
seD
NA
Pbp
2bP
bp2x
0 1 20
0.5
1
Stage 2/ n = 71
0 1 2
Stage 3/ n = 51
0 1 2
Stage 4/ n = 42
0 1
2
(µm)
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.40.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Stage 1 NSStage 2 ***Stage 3 ***Stage 4 **
Δ
Pbp2b Width ( μm)
Pbp2
x W
idth
(μm
)
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.40.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Stage 1 **Stage 2 ***Stage 3 ***Stage 4 ***
Δ
StkP Width (μm)
Pbp2
x W
idth
( μm
)
A Pbp2x-FLAG3 Pbp2b-HA B Pbp2x-HA StkP-FLAG3
Fig. 6. 2D-IFM analysis showing separate positioning of (A) Pbp2b and Pbp2x and (B) StkP and Pbp2x in septa of mid-to-late divisionalpneumococcal cells. Top panels are averaged images and normalized mean fluorescence intensity traces, and bottom graphs are scatter plotsof labelled widths obtained from (A) strain IU7145 (Pbp2x-FLAG3 Pbp2b-HA) and (B) strain IU7510 (StkP-FLAG2 Pbp2x-HA). Dual-protein 2DIFM and DAPI labelling were performed as described in Experimental procedures. Image averaging and generation of scatter plots of pairedlabelled widths at mid-cell equators and septa were done as described for Fig. 2 and in Experimental procedures and Land et al. (2013). NS,not significant (i.e. difference of means of paired widths = 0); **P < 0.01; and ***P < 0.001. The data shown were obtained from twoindependent biological replicate experiments.
Separation of pneumococcal Pbp2x from Pbp2b at septa 9
© 2014 John Wiley & Sons Ltd, Molecular Microbiology
regions of the cell surface labelled with fluorescent vanco-mycin (FL-V), indicative of the presence of PG pentapep-tide substrate and active PG synthesis (Daniel andErrington, 2003; Ng et al., 2004; Pinho and Errington,2005; Wheeler et al., 2011). Dual-labelled 2D IFM and3D-SIM IFM again indicated a distinctive localizationpattern for Pbp2x compared to the other four proteins(Figs 10, 11, S7 and S8). In early-divisional cells, all fiveproteins and FL-V staining colocalized in equatorial rings.However, as the septal rings constricted, rings of Pbp2xbecame smaller than regions of FL-V labelling (Figs 10Aand 11A), whereas rings of the other Pbp2b, Pbp1a, StkPand MreC circled regions of FL-Van labelling (Figs 10, 11,S7 and S8). At mid-to-late stages of division, regions ofFL-V staining filled septa and extended along the bounda-ries of the joined cells. In these cells, Ppb2x located as aclosing disk within the region of FL-V labelling (Fig. 11A),whereas Pbp2b, Pbp1a, StkP and MreC formed ringsthreaded around the boundaries marked by FL-V labelling(Figs 11 and S8). In fully divided cells, FL-V and proteinsthat had not migrated to equators of the daughter cellscollapsed into single puncta at the poles of the connectedcells. We conclude that relative to the cell surface revealed
by FL-V labelling, Pbp2x separates from and precedesother PG synthesis proteins during septal closure in pneu-mococcal cells.
Discussion
This paper provides two independent pieces of evidencethat the PG synthesis apparatus separates into two adja-cent complexes in mid-to-late divisional cells in S. pneu-moniae D39 cells. First, FDAA probes pulse-labelled twodistinct regions of active PBP transpeptidase activity in thesepta of mid-to-late divisional cells; a ring around a centralcontiguous region of FDAA staining (Figs 3 and 4). Thecentral transpeptidase activity can be attributed to Pbp2x,because the central labelling disappeared when Pbp2xtranspeptidase activity was inhibited by the β-lactam anti-biotic methicillin (Fig. 4) and when Pbp2x was depleted(Fig. 5). Second, dual-protein and protein-fluorescent van-comycin 2D and 3D-SIM IFM showed that Pbp2x sepa-rated to septal centres ahead of adjacent closing ringscontaining Pbp2b, Pbp1a, StkP and MreC (Fig. 12). ThePbp2x localization pattern is consistent with the FDAA-labelling results (Figs 3–5) and supports the conclusion
Fig. 7. Representative 3D-SIM IFM images confirming separate positioning of (A) Pbp2b and Pbp2x and (B) StkP and Pbp2x in septa ofmid-to-late divisional pneumococcal cells. Representative 3D-SIM IFM images of strain IU7145 (Pbp2x-FLAG3 Pbp2b-HA) and IU7510(Pbp2x-HA StkP-FLAG2) at different division stages were obtained as described in Experimental procedures. Arrangement of panels andpseudo-colouring is similar to Fig. 2, except red and green correspond to Pbp2x and Pbp2b in (A), and Pbp2x and StkP in (B), respectively.The first row of each panel represents images captured in the XY plane, while second row images were obtained by rotating a section of themid-cell region around the X or Y axis. In stage 1 cells, Pbp2x colocalizes with Pbp2b and StkP; however by stages 2 and 3, Pbp2x separatesfrom Pbp2b and StkP and eventually forms a disk in the septum surrounded by rings of Pbp2b and StkP (arrows). Between stage 3 and 4,Pbp2x appears as one layer between two layers of Pbp2b on the sides of the new daughter cells (also see Fig. 9B) (arrows). Images arerepresentative of > 30 examined cells in different division stages from two biological replicates. Scale bar = 1 μm.
10 H.-C. T. Tsui et al. ■
© 2014 John Wiley & Sons Ltd, Molecular Microbiology
that Pbp2x locates separately from other PG synthesisproteins at later stages of pneumococcal cell division.Eventually, the septal ring, which lacks FtsZ at this stage,closes down to a concentrated polar dot, by which timemost of PG synthesis proteins have re-appeared at theequators of daughter cells (Figs 2, S4, S5 and 12; Landet al., 2013).
The results in this paper support and extend our previousconclusion that Pbp2x exhibits distinctive, separate locali-zation from Pbp1a and other PG synthesis proteins inmid-to-late divisional cells (Land et al., 2013). In consider-ing this localization pattern (Fig. 12), it is important to notethat Pbp2x colocalizes with Pbp2b, Pbp1a, StkP and MreCin early stages of division and that Pbp2x remains adjacentto the closing septal ring that contains the other proteins.Therefore, there is opportunity in the division cycle forPbp2x to interact with Pbp1a, as suggested by suppressor
studies (Zerfass et al., 2009; Peters et al., 2014), and withStkP, as suggested by biochemical assays (Morlot et al.,2013). Our results indicate that with the exception of Pbp2xin mid-to-late divisional cells, the protein complexes thatmediate septal and peripheral synthesis functions are notseparated in the mid-cell region of pneumococcal cellsbeyond the ≈ 100 nm resolution of 3D-SIM. Thus, the ideaof widely separated PG synthesis machines at differentmid-cell locations is not supported, as discussed recentlyin (Fleurie et al., 2014). On the other hand, the resolution of3D-SIM (≈ 100 nm) cannot distinguish whether PG synthe-sis functional complexes are further separated or organ-ized topologically within the larger mid-cell ring structuresdetected here. Determination of whether there are adja-cent separate complexes that carry out septal and periph-eral PG synthesis will require higher resolution approachesthan 3D-SIM.
Stage 1/ n = 56
Pha
seD
NA
Stk
PP
bp2b
0 1 20
0.5
1
Stage 2/ n = 67
0 1 2
Stage 3/ n = 62
0 1 2
Stage 4/ n = 41
0 1 2
(μm)
Stage 1/ n = 103
Pha
seD
NA
Pbp
1aP
bp2b
0 1 20
0.5
1
Stage 2/ n = 78
0 1 2
Stage 3/ n = 55
0 1 2
Stage 4/ n = 44
0 1 2(μm)
Stage 1/ n = 39
Pha
seD
NA
Pbp
2bM
reC
0 1 20
0.5
1
Stage 2/ n = 40
0 1 2
Stage 3/ n = 36
0 1 2
Stage 4/ n = 32
0 1 2(μm)
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.40.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Stage 1 ***Stage 2 NSStage 3 NSStage 4 ***
Δ
StkP Width (μm)Pb
p2b
Wid
th (
μm)
A Pbp1a-L-FLAG3 Pbp2b-HA
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.40.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Stage 2 NSStage 3 NSStage 4 NS
Stage 1 **Δ
Pbp2b Width (μm)
Mre
C W
idth
(μm
)
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.40.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Stage 1 NSStage 2 NSStage 3 NSStage 4 NS
Δ
Pbp1a Width (μm)
Pbp2
b W
idth
(μm
)B Pbp2b-HA4 MreC-L-FLAG3 C Pbp2b-HA StkP-FLAG2
Fig. 8. 2D-IFM analysis showing that Pbp2b, Pbp1a, MreC and StkP colocalize in mid-cell rings of similar diameters at all stages ofpneumococcal cell division. Top panels are averaged images and normalized mean fluorescence intensity traces, and bottom graphs arescatter plots of labelled widths obtained from (A) IU7363 (Pbp2b-HA Pbp1a-L-FLAG3), (B) IU7752 (Pbp2b-HA4 MreC-L-FLAG3) and (C) IU7512(StkP-FLAG2 Pbp2b-HA). Dual-protein 2D IFM and DAPI labelling were performed as described in Experimental procedures. Image averagingand generation of scatter plots of paired labelled widths at mid-cell equators and septa were done as described for Fig. 2 and in Experimentalprocedures and (Land et al., 2013). NS, not significant (i.e. difference of means of paired widths = 0); **P < 0.01; and ***P < 0.001. Datashown were obtained from two independent biological replicates.
Separation of pneumococcal Pbp2x from Pbp2b at septa 11
© 2014 John Wiley & Sons Ltd, Molecular Microbiology

Our results add an important qualification to the gener-alization from recent studies that pneumococcal Pbp2xlocalizes similarly to Pbp2b, Pbp1a and StkP throughoutthe cell cycle (Beilharz et al., 2012; Morlot et al., 2013;Fleurie et al., 2014; Peters et al., 2014). In one recentstudy (Fleurie et al., 2014), FDAA labelling failed to revealseparate regions of transpeptidase activity in the septa ofmid-to-late divisional cells, as this labelling pattern wouldnot be observable in transversely displayed cells at con-ventional resolution. Here, the separation of FDAA label-ling was readily detected by high-resolution 3D-SIM(Figs 3 and 4). Moreover, separation of Pbp2x from otherPG synthesis proteins occurs in mid-to-late, but not in earlystages, of division (Fig. 12). Reliable detection of Pbp2xseparation required averaging of 2D images of populationsof cells at different stages of division (Figs 2, 6, 8 and 10),combined with analysis of 3D-SIM images (Figs 3, 7, 9 and11). These combined approaches were not used in previ-ous studies. Finally, conclusions about PG synthesisprotein colocalization in one study partly relied on patternsobserved in elongated ΔgpsB mutant cells of a laboratorystrain that likely contains a suppressor (Fleurie et al., 2014)compared to the strain D39 background used here, inwhich gpsB is essential (Land et al., 2013).
The mechanisms that target Pbp2x to the centres ofsepta in later stages of pneumococcal division arecurrently unknown. Recent results establish that the extra-cellular C-terminal PASTA domains, but not the transpepti-dase domain or its activity, are required for normallocalization of pneumococcal Pbp2x (Peters et al., 2014).This conclusion was further supported in a new papershowing that the cytoplasmic and transmembranedomains of pneumococcal Pbp2x are essential for func-tion, but not for normal localization (Berg et al., 2014). Inanother recent study, the extracellular PASTA domains ofthe StkP Ser/Thr kinase were implicated in normal locali-zation of Pbp2x, possibly through a direct interaction(Morlot et al., 2013). However, the relationship betweenStkP and Pbp2x localization seems to have broken down inΔgpsB mutants, which showed mislocalization of StkP, butnot Pbp2x (Fleurie et al., 2014). The roles of PASTA andother domains of Pbp2x and StkP in early-stage equatorialand mid-to-late-stage septal localization remain to bedetermined.
Its exceptional localization in later divisional cells sug-gests that Pbp2x may play additional roles in septal com-pletion. We tested whether Pbp2x and Pbp1a localizationpatterns changed in a mutant that was deleted for the other
Fig. 9. Representative 3D-SIM images confirming that Pbp2b, Pbp1a, MreC and StkP colocalize in a constricting mid-cell ring duringpneumococcal cell division. Representative 3D-SIM IFM images were obtained from (A) IU7363 (Pbp1a-L-FLAG3 Pbp2b-HA), (B) IU7752(Pbp2b-HA4 MreC-L-FLAG3) and (C) IU7512 (Pbp2b-HA StkP-FLAG2) at different division stages. Arrangements of panels are similar toFigs 2C and 7. Images shown are representative of > 30 examined cells in different division stages from two biological replicates. Scalebar = 1 μm.
12 H.-C. T. Tsui et al. ■
© 2014 John Wiley & Sons Ltd, Molecular Microbiology

class A PBPs, Pbp2a and Pbp1b. In a Δpbp2a Δpbp1bmutant (IU8447, Table S1), Pbp1a is the only enzyme thatcatalyses PG transglycosylase activity that makes glycanchains (see Massidda et al., 2013). We found that Pbp1aand Pbp2x localization was the same in the Δpbp2aΔpbp1b mutant as in the wild-type parent (data not shown).Thus, in this mutant, the Pbp1a PG transglycosylase activ-ity does not localize with Pbp2x to the centres of septa inmid-to-late divisional cells. Pbp1a and Pbp2x colocalizeagain only in the last stages of division, when the septalring condenses to a dot, and in the next division cycle at theequators of predivisional cells (Fig. 12). Together theseresults suggest the following model. In early stages ofdivision, Pbp2x is likely required for cross-link formation inseptal PG synthesis. In later stages of division after Pbp2xseparates to septal centres, this role in septal PG synthesis
may continue by using glycan strands synthesized byadjacent Pbp1a and other class A PBPs. In addition, sepa-rated Pbp2x may assume a role in PG remodelling byattaching new glycan strands to old glycan strands, whosecross-links have been cleaved by remodelling PG endo-peptidases (Singh et al., 2012). Current studies are aimedat comparing the localization patterns of septal remodellingPG hydrolases, such as the PcsB : FtsEX complex (Shamet al., 2011; 2012; 2013; Bartual et al., 2014), to those ofPbp2x and other PG synthesis proteins in septa of mid-to-late divisional cells (Fig. 12).
Finally, the results presented here show that FDAAlabelling patterns are strongly correlated with theD,D-transpeptidase activities of pneumococcal PBPs(Figs 3–5). In wild-type pneumococcal cells, FDAA label-ling appears as a single ring at equators of early divisional
Stage 1/ n = 56
Pha
seD
NA
FL−
VP
bp1a
0 1 20
0.5
1
Stage 2/ n = 56
0 1 2
Stage 3/ n = 52
0 1 2
Stage 4/ n = 56
0 1 2(μm)
Stage 1/ n = 63
Pha
seD
NA
FL−
VP
bp2b
0 1 20
0.5
1
Stage 2/ n = 60
0 1 2
Stage 3/ n = 52
0 1 2
Stage 4/ n = 49
0 1 2(μm)
Stage 1/ n = 44
Pha
seD
NA
FL−
VP
bp2x
0 1 20
0.5
1
Stage 2/ n = 44
0 1 2
Stage 3/ n = 55
0 1 2
Stage 4/ n = 41
0 1 2
(μm)
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.40.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Stage 2 **Stage 3 ***Stage 4 ***
Stage 1 NSΔ
FL-V Width (μm)
Pbp2
x W
idth
(μm
)
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.40.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Stage 1 ***
Stage 4 ***
Stage 2 ***Stage 3 ***
Δ
FL-V Width (μm)
Pbp2
b W
idth
(μm
)
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.40.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Stage 2 ***Stage 3 ***Stage 4 ***
Stage 1 NSΔ
FL-V Width (μm)Pb
p1a
Wid
th (
μm)
A Pbp2x-HA B Pbp2b-HA4 C Pbp1a-L-FLAG3
Fig. 10. 2D-IFM analysis showing smaller septal width of Pbp2x, but larger septal widths of other PG synthesis proteins compared to FL-Vstaining at different stages of pneumococcal cell division. Dual FL-V staining and protein IFM was performed as described in Experimentalprocedures. Top panels are averaged images and normalized mean fluorescence intensity traces, and bottom graphs are scatter plots oflabelled widths for FL-V staining and immunolabelled proteins for (A) IU6929 (Pbp2x-HA), (B) IU7426 (Pbp2b-HA4) and (C) IU5544(Pbp1a-L-FLAG3). Image averaging and generation of scatter plots of paired labelled widths at mid-cell equators and septa were done asdescribed for Fig. 2 and in Experimental procedures and (Land et al., 2013). NS, not significant (i.e. difference of means of paired widths = 0);**P < 0.01; and ***P < 0.001. Data shown were obtained from two independent biological replicates.
Separation of pneumococcal Pbp2x from Pbp2b at septa 13
© 2014 John Wiley & Sons Ltd, Molecular Microbiology

cells and as a ring surrounded by a central disk at the septaof mid-to-late divisional cells (Figs 3 and 4). This patternmatches the localization of the Pbp2x protein at differentdivision stages (Fig. 12). As a cautionary note, evenmodest overexpression (≈ 2-fold) of Pbp2x obscures thetwo-position FDAA labelling pattern at septa, and FDAAlabelling now occurs as thickened disks over entire septa(Fig. 5A). This modest overexpression of Pbp2x is accom-panied by moderate size and shape changes of cells (Fig.S1B). Inhibition of Pbp2x transpeptidase activity or deple-tion of Pbp2x amount results in FDAA labelling of equato-rial rings, but not central septal disks (Figs 4 and 5A).HPLC analysis of pneumococcal PG peptides showed thatFDAAs predominantly label PG pentapeptide monomersand PG peptide dimers containing a pentapeptide (datanot shown). S. pneumoniae lacks homologues of L,D-
transpeptidase that are found in E. coli and other bacteria(see Sanders and Pavelka, 2013). Moreover, FDAA label-ling of equators and septa is abrogated by prior treatmentof cells with penicillin or vancomycin that generally inhibitPBP transpeptidation; after antibiotic addition, FDAA dis-tributes throughout pneumococcal cells (data not shown).Together, these results indicate that FDAA labelling reportsregions on cells where PBP transpeptidases are active atdifferent stages of the pneumococcal cell cycle.
Experimental procedures
Bacterial strains and growth conditions
Bacterial strains used in this study were derived from strainIU1945, an unencapsulated derivative of serotype 2 S. pneu-moniae strain D39 (Lanie et al., 2007; Table S1). Strains
Fig. 11. Representative 3D-SIM images showing different distributions of the PBPs with respect to FL-V staining of (A) IU6929 (Pbp2x-HA),(B) IU7426 (Pbp2b-HA4) and (C) IU5544 (Pbp1a-L-FLAG3). Dual FL-V staining and protein IFM was performed as described in Experimentalprocedures. FL-V stain is pseudo-coloured green, and Pbp2x (A), Pbp2b (B) and Pbp1a (C) are pseudo-coloured as red, and overlappingFL-V and PBP signal is represented by yellow. Numbers to the left of each panel indicate division stages. The first row of each panel showsimages captured in the XY plane, while second row images were obtained by rotating a section of the mid-cell region around the X or Y axis.In mid-to-late stage 3 divisional cells, FL-V staining is splitting between the daughter cells, Pbp2x (A) is present in septal centres, and Pbp2b(B) and Pbp1a (C) are present in the outer rims of FL-V labelling (arrows). Images shown are representatives of > 20 examined cells indifferent division stages from two biological replicates. Scale bar = 1 μm.
14 H.-C. T. Tsui et al. ■
© 2014 John Wiley & Sons Ltd, Molecular Microbiology

containing antibiotic markers were constructed by transform-ing linear DNA amplicons synthesized by overlapping fusionPCR into competent pneumococcal cells as described previ-ously (Ramos-Montanez et al., 2008; Tsui et al., 2010; 2011).Primers used for the generation of amplicons are listed inTable S2. All constructs were confirmed by DNA sequencingof chromosomal regions corresponding to the ampliconregion used for transformation. Bacteria were grown onplates containing trypticase soy agar II (modified; Becton-Dickinson) and 5% (v/v) defibrinated sheep blood (TSAII-BA).Plates were incubated at 37°C in an atmosphere of 5%CO2. For antibiotic selections, TSAII BA plates contained250 μg kanamycin ml−1, 150 μg spectinomycin ml−1, 0.3 μgerythromycin ml−1, 250 μg streptomycin ml−1, or 2.5 μgchloramphenicol ml−1. Strains were cultured statically inBecton-Dickinson brain heart infusion (BHI) broth at 37°C inan atmosphere of 5% CO2, and growth was monitored byOD620 as described before (Tsui et al., 2010; 2011; Landet al., 2013). Bacteria were inoculated into BHI broth fromfrozen cultures or colonies, serially diluted into the samemedium, and propagated overnight. For growth experiments,overnight cultures that were still in exponential phase(OD620 = 0.1–0.4) were diluted back to OD620 ≈ 0.003 to startfinal cultures, which lacked antibiotics.
Transformations of Δpbp2x< >aad9 amplicon into merodip-loid strain IU7470 (pbp2x+//CEP::PfcsK-pbp2x+) to obtain strainIU7506 or Δpbp2b< >aad9 amplicon into merodiploid strainIU7337 (pbp2b+//ΔbgaA::PfcsK-pbp2b+) to obtain strain IU7397,were carried out in the presence of 1% (w/v) L-fucose (Sigma,F2252) in the transformation mix and in all subsequent steps.To deplete cells of Pbp2x or Pbp2b, strains IU7506 (Δpbp2x//CEP::PfcsK-pbp2x+) or IU7397 (Δpbp2b//ΔbgaA::PfcsK-pbp2b+)were grown overnight in BHI broth containing 1.0% (w/v)L-fucose. Cells from overnight cultures were collected bycentrifugation (10 min at 16 000 g at 25°C) and resuspendedto OD620 ≈ 0.005 in BHI broth containing 1.0% (w/v) L-fucose orno fucose.
Cell length and width measurements
Cell lengths and widths of parent strain IU1945 and strainIU7397 (Δpbp2b//ΔbgaA::PfcsK-pbp2b+) grown in BHI brothwith or without fucose were measured from phase-contrastimages by using Nikon NIS-Element AR software asdescribed before (Barendt et al., 2009). For IU1945, sepa-rated stage 1 cells or daughter cells of stage 4 cells weremeasured. For strain IU7397 grown with or without fucose,measurements included cells in chains whose widths at con-striction sites were < 50% of cell widths.
Boc-FL labelling of PBPs in pbp2x merodiploid andHA-tagged pbp2b strains
Quantitative labelling with fluorescent bocillin (Boc-FL;Molecular Probes) to determine PBP activity and/or amountwas performed for strains expressing Pbp2b-HA (IU6933), orPbp2b-HA4 (IU7426), and for a pbp2x merodiploid strain(IU7506) as described previously (Zhao et al., 1999;Kocaoglu et al., 2012; Land et al., 2013).
Western blotting
Strains were grown exponentially in BHI broth toOD620 ≈ 0.15. Lysates were prepared as described previously(Wayne et al., 2010), and polypeptides were separated using4–15% mini-protean TGX pre-cast gradient gels (Bio-Rad,456–1083). A Benchmark pre-stained protein standard (LifeTechnologies, 10748-010) calibrated against a Novex sharpunstained protein standard (Life Technologies, LC5801) wasused to determine polypeptide molecular masses. FLAG-,HA-, and Myc-tagged proteins were detected by Westernblotting using a 1:1000 dilution of primary anti-FLAG poly-clonal antibody (Sigma, F7425), anti-HA rabbit polyclonalantibody (Invitrogen, 71–5500) at 1 μg ml−1, or a 1:1000 dilu-
Pbp2x, Pbp1aPbp2b, StkP,MreC, FL-V
Pre-divisionalcell
Mid-divisionalcell
Pbp2b, Pbp1a,StkP, MreC
Pbp2xFL-V
Pbp2xFL-V
Pbp2b, Pbp1aStkP, MreC
Mid-to-late-divisional cell
Pbp2xFL-VPbp2bPbp1aStkPMreC
FtsZ
Late-divisionalcell
remainingFtsZ
FtsZ
Fig. 12. Summary of localization patterns of S. pneumoniae PBPs, PG synthesis regulators StkP and MreC, and regions of fluorescent-vancomycin (FL-V) staining at different stages of cell division. Sections of mid-cells looking down the long axes of cells are illustrated. Inpre-divisional cells, all proteins and FtsZ are located in an equatorial ring that will become the septum. The FtsZ and PG synthesis proteinrings likely coincide, although FtsZ rings appear to be smaller than PG synthesis protein rings, because the FtsZ epitope tag is cytoplasmic,whereas the epitope tags on PG synthesis proteins are extracellular (see text). In mid-divisional cells, Pbp2x has started to form a ring insideof the constricting septal ring that contains the other proteins and surrounds regions of FL-V staining; FtsZ has started to disappear from theseptum and reorganize at the equators of daughter cells. In mid-to-late division, FtsZ has left septa and is assembling into new equatorialrings, Pbp2x has moved to septal centres overlapping regions of FL-V staining, while the other proteins remain in a constricting septal ringsurrounding regions of FL-V staining. In the last stages of division, cells have or are about to fully separate and the remaining proteins andFL-V staining condense into a polar dot; most of these proteins have started to reassemble at the equatorial rings or daughter cells. See textfor additional details.
Separation of pneumococcal Pbp2x from Pbp2b at septa 15
© 2014 John Wiley & Sons Ltd, Molecular Microbiology

tion of primary anti-Myc polyclonal rabbit antibody (Sigma,C3956), followed by a 1:10 000 dilution of HRP-conjugateddonkey anti-rabbit antibody (GE Healthcare, NA934), andECL detection reagent. Chemiluminescent signal from poly-peptide bands was detected by using an IVIS imaging systemas described in Wayne et al. (2010).
2D immunofluorescence microscopy
Localization of FLAG-, Myc- and HA-tagged proteins by IFMwas performed for exponentially growing cells as describedbefore (Wayne et al., 2010; Tsui et al., 2011; Land et al.,2013). Primary antibodies used were anti-FLAG rabbit poly-clonal antibody (Sigma, F7425, 1:100 dilution), anti-FLAGmouse monoclonal M2 antibody (Sigma, F1804, 1:100), anti-c-Myc rabbit polyclonal antibody (Sigma, C3956, 1:100dilution), anti-c-Myc mouse monoclonal antibody (LifeTechnologies, R950-25, 1:100 dilution), or anti-HA rabbitpolyclonal antibody (Invitrogen, 71–5500, 1:50 dilution). Sec-ondary antibodies used were Alexa Fluor 488 or Alexa Fluor568 highly cross-adsorbed goat anti-rabbit IgG (Life Tech-nologies; A11034 or A11036, respectively, 1:100 dilution) andAlexa Fluor 488 or Alexa Fluor 568 (Life Technologies;A11029 or A11031, respectively, 1:100 dilution) highly cross-adsorbed goat anti-mouse IgG. For-single-protein labellingexperiments, polyclonal rabbit antibodies were used followedby Alexa Fluor 488 or Alexa Fluor 568 goat anti-rabbit IgG.For dual labelling of FLAG- and Myc-tagged proteins[(IU6978 (pbp2x-FLAG3 ftsZ-Myc); IU7074 (mreC-L-FLAG3
ftsZ-Myc)], primary antibodies were anti-FLAG rabbit poly-clonal antibody and mouse monoclonal antibody againstc-Myc, and secondary antibodies were Alexa Fluor 488 goatanti-rabbit and Alexa Fluor 568 goat anti-mouse. For duallabelling of HA- and Myc-tagged proteins [IU7217 (pbp2b-HAftsZ-Myc)], primary antibodies were anti-HA rabbit polyclonalantibody and anti-Myc mouse monoclonal antibody, and sec-ondary antibodies were Alexa Fluor 488 goat anti-rabbit andAlexa Fluor 568 goat anti-mouse. For dual labelling of HA-and FLAG-tagged proteins [(IU7145 (pbp2b-HA pbp2x-FLAG3); IU7590 (stkP-HA pbp1a-L-FLAG3); IU7752 (pbp2b-HA4 mreC-L-FLAG3); IU7363 (pbp2b-HA pbp1a-L-FLAG3);IU7365 (pbp2x-HA pbp1a-L-FLAG3); IU7510 (pbp2x-HA stkP-FLAG2); IU7512 (pbp2b-HA stkP-FLAG2)], primary antibodieswere anti-HA rabbit polyclonal antibody and anti-FLAGmouse monoclonal antibody. Secondary antibodies wereAlexa Fluor 488 goat anti-rabbit and Alexa Fluor 568 goatanti-mouse for strains IU7145, IU7590 and IU7752, andAlexa Fluor 568 goat anti-rabbit and Alexa Fluor 488 goatanti-mouse for strains IU7363, IU7365, IU7510, and IU7512.Primary antibody incubations were for 1h at 24°C for singletagged strains, and FLAG-Myc and Myc-HA double taggedstrains and for 2 h at 37°C for FLAG-HA double taggedstrains. Secondary antibody incubation was for 1 h at 24°Cfor all strains. DNA in nucleoids was stained by using mount-ing media SlowFade gold antifade reagent with DAPI (LifeTechnologies, S36936).
Control experiments showed similar localization patternsfor each protein when tracked in single or dual-protein label-ling experiments (data not shown). Similar localization pat-terns were observed for each protein fused to different tags[Pbp2x fused to HA (IU6929), FLAG3(IU6819), or HA3
(IU7722); Pbp2b fused to HA (IU6933) or HA4 (IU7426);StkP fused to FLAG2 (IU7434) or HA (IU7438)]. Similarlocalization patterns were observed when Alexa Fluor 488or Alexa Fluor 568 combinations were swapped in dual-protein experiments [IU7217 (pbp2b-HA ftsZ-Myc); IU7590(pbp1a-L-FLAG3 stkP-HA)] (data not shown). Controlexperiments showed no detectable labelling in cells notexpressing tagged proteins (IU1945) with the combinationsof antibodies used (data not shown). Labelling of strainscontaining single-tagged proteins tested with the doublelabelling procedure that contained both sets of primary andsecondary antibodies produced signal only in the expectedfluorescence channel (data not shown).
Image analysis of 2D IFM images
2D IFM images of dividing pneumococcal cells were alignedand manually binned into four division stages (pre-, early-,mid- and late-divisional) using a point-and-click IMA-GUIorganized in MATLAB (The Mathworks) as reported before(Land et al., 2013). Mean normalized cell outlines fromphase-contrast images and mean normalized fluorescentintensities of DNA and proteins from all aligned cells at agiven stage were calculated to graphically represent therelative distribution of molecules along the longitudinal cellaxis during the division process (Land et al., 2013).Numbers of cells averaged for each stage are indicated (n)and were taken from at least two fields from each of twoindependent biological replicate experiments, unless speci-fied otherwise.
To quantitatively evaluate the relative positions of indi-vidual proteins through the process of septation, we extendedthe IMA-GUI to include methods for independently markingthe septal edge positions of different proteins in the samedouble-labelled cell. The distance between each edge repre-sents the extent to which the protein is distributed within theseptal ring (i.e. its septal width). Widths were determined forprotein septal rings with sharp edges in images of cells aver-aged at different division stages. Using the cell morphologyas an indicator of cell cycle phase, we reasoned that if twoproteins were discretely colocalized, the mean difference inseptal width for all cells in a particular phase would becentred at zero with a normal distribution across the cellpopulation. For statistical analyses, paired widths from indi-vidual cells were output to a data sheet in GraphPad Prism 5.Some proteins were largely absent from septa of stage 4 cells(e.g. FtsZ, Fig. 2A) or did not form septal protein rings withdistinct edges and were not included in width comparisons.For pairs of different proteins, septal widths were comparedfor 63–100%, 78–99%, 67–100% and 20–100% of binnedcells at stages 1, 2, 3 and 4 respectively. Differences betweenpaired widths were calculated for cells in each division stage.One sample Student’s t-tests were performed to determinewhether mean differences in widths were significantly differ-ent from the null hypothesis value of zero (NS, not significant;**P < 0.01; and ***P < 0.001 in graphs). To visualize theextent to which each protein pair changes relative widththrough different cell divisional stages, paired data from eachcell were plotted as a scatterplot with a superimposed diago-nal line to indicate the mean position for the assumption thatthe two proteins are strictly colocalized through the cell cycle.
16 H.-C. T. Tsui et al. ■
© 2014 John Wiley & Sons Ltd, Molecular Microbiology

3D-SIM IFM
Samples were prepared as described previously (Land et al.,2013), and 3D-SIM was performed using the OMX 3D-SIMsuper-resolution system located in the Indiana UniversityBloomington Light Microscopy Imaging Center (http://www.indiana.edu/~lmic/microscopes/index.html#OMX). Thesystem used is equipped with four Photometrics Cascade IIEMCCD cameras that allow simultaneous four colourimaging, and is controlled by DV-OMX software, with imageprocessing by Applied Precision softWoRx 6.0.0 software.Exposure times and %T settings for DAPI, Alexa 488, andAlexa 568 images were 50–100 ms and 50%, 50 ms and1–10%, and 50 ms and 10–50%, respectively.
3D-SIM of FDAA pre-labelled and pulse-labelled cells
FDAAs HADA (7-hydroxycoumarin-3-carboxylic acid 3-amino-D-alanine) and TADA (tetramethylrhodamine 3-amino-D-alanine) were synthesized as reported in Kuru et al. (2012),with the following change: TADA was synthesized asreported for TDL, except that Boc-D-DAP-OH (N-alpha-t-butyloxycarbonyl-D-2,3-diaminopropionic acid) was used inplace of Boc-D-Lys-OH (N-alpha-t-butyloxycarbonyl-D-lysine).Working solutions of HADAand TADA in BHI were diluted from500 mM stocks in DMSO, which were stored at −70°C. ForHADA pre-labelling, bacterial cells from overnight cultureswere diluted to OD620 ≈ 0.01 in 2 ml of fresh BHI broth (37°C)and incubated at 37°C in an atmosphere of 5% CO2. AtOD620 ≈ 0.02, 1 μl of 500 mM HADA in DMSO was added to afinal concentration of 250 μM. At OD620 ≈ 0.2–0.25 (≈ 3.5 dou-blings), cultures were transferred to a 2 ml microfuge tube,placed in an ice bath for 1 min, and then centrifuged for 5 minat 16 000 g at 4°C. Supernates were discarded, and pelletswere resuspended in 1 ml of cold (4°C) phosphate-bufferedsaline (PBS; Ambion; AM9625). Cells were centrifuged for2.5 min at 16 000 g at 4°C, resuspended in cold PBS, andcentrifuged again. After the second wash and centrifugation,cells were pulse-labelled with TADA. Pellets were resus-pended in 1 ml of warm (37°C) BHI broth containing 500 μMTADA. Cells were incubated at 37°C in an atmosphere of 5%CO2 for 5 min (14% of doubling time) and then chilled on ice for1 min. Cells were centrifuged for 2.5 min at 16 000 g at 4°C,and pellets were resuspended in 1 ml of cold (4°C) PBS. Afterthe second wash and centrifugation, pellets were resus-pended in 1 ml of 4% paraformaldehyde (EMS; 157-4) andincubated for 15 min at room temperature, followed by 45 minon ice. Fixed cells were centrifuged, and pellets were washedthree times with PBS at 4°C as described above.After the thirdwash and centrifugation, cells were resuspended in 0.3 ml ofcold (4°C) GTE buffer (50 mM glucose, 1 mM EDTA, 20 mMTris-HCl, pH 7.5), and stored in the dark at 4°C for up to 20 h.Fifty microlitres of cell suspension was deposited on amethanol-treated coverslip and incubated for 5 min at roomtemperature. Unattached cells were aspirated, and attachedcells were treated with 50 μl of 0.2% Triton X-100 (Mallinck-rodt; H282) in PBS (PBS-T) for 10 s. PBS-T was aspiratedfrom the wells, and the coverslip was air dried completelybefore immersion in cold (−20°C) methanol and kept at −20°Cfor 10 min. The coverslip was then removed from methanoland air dried, after which the coverslip was incubated with
50 μl of PBS-T for 5 min, followed by two washes with 50 μlPBS and aspirated immediately, and once with 50 μl PBS for5 min at room temperature before aspiration. After the cover-slip was air dried, 7 μl of Slowfade Gold antifade reagent(Invitrogen; S36936) was applied to the coverslip, which wasthen inverted onto a cleaned and methanol-treated slide, andsealed. 3D-SIM was performed using the OMX 3D-SIM super-resolution system described above. Laser lines used forexamination of samples were 405 with emission filters of419–465 for HADA, and 561 with emission filters of 609–705for TADA. Exposure times and %T settings for both HADA andTADA were 50 ms and 30%. Images in Fig. 3 are representa-tive of > 100 3D-reconstructed cells in different division stagesfrom > 3 independent experiments.
For TADA labelling after methicillin treatment of strainIU1945 (D39 Δcps, WT), duplicate 5 ml cultures of IU1945were grown to exponential phase (OD620 ≈ 0.1) at 37°C in anatmosphere of 5% CO2. Methicillin was added to a finalconcentration of 0.1 μg ml−1 to one culture (Land et al., 2013).After 40 min, 250 μl samples of the treated and untreatedcultures were added to a microfuge tubes. TADA was addedto a final concentration of 500 μM, and incubation was con-tinued for 5 min at 37°C. Samples were prepared for 3D-SIMas described above. Images in Fig. 4 are representative of> 60 3D reconstructed stage-3 cells from two independentexperiments.
FDAA labelling and 3D-SIM microscopy of merodiploidstrains IU7506 (Δpbp2x//CEP::PfcsK-pbp2x+) and IU7397(Δpbp2b//ΔbgaA::PfcsK-pbp2b+) were performed as describedabove, with the following changes. Overnight cultures ofstrains IU7506 and IU7397 in BHI broth containing 1.0% (w/v)L-fucose were centrifuged, and pellets were resuspended into5 ml of warm (37°C) BHI broth containing or lacking 1.0% (w/v)L-fucose and diluted to a starting OD ≈ 0.01. To conserveFDAA usage, HADA was added to a final concentration of250 μM to a 500 μl aliquot of each starting culture in amicrofuge tube, which was covered by perforated parafilm andincubated at 37°C in an atmosphere of 5% CO2. Parallel OD620
readings were monitored for the remaining 4.5 ml of cultures.At OD620 ≈ 0.1 (≈ 2.8 h after the start of the cultures), sampleswere washed and resuspended in 250 μl BHI broth containingTADA at a final concentration of 500 μM. The cultures wereincubated at 37°C for 5 min, washed, and prepared for 3D-SIMas described above. Images shown in Fig. 5 are representa-tive of > 24 3D-reconstructed stage-3 cells for each conditionfrom two independent biological replicates.
Dual FL-V staining and protein IFM
One hundred microgram of Bodipy-FL-conjugated vancomy-cin (FL-V; Molecular Probes, V-34850) was dissolved in 10 μlof DMSO, and 90 μl of sterile water was added to make a1 μg μl−1 solution. This solution was mixed with equal volumeof unlabelled vancomycin at the same concentration. Threemicrolitres of this mixture was added to 1.5 ml of exponentiallygrowing cells (OD620 ≈ 0.15) in BHI broth to give a final con-centration of 2 μg ml−1 total vancomycin. Samples were incu-bated at room temperature for 5 min, placed on ice for 2 min,collected by centrifugation, and washed twice with cold PBS,followed by fixation with 4% paraformaldehyde and subse-quent steps used for IFM (Wayne et al., 2010; Land et al.,
Separation of pneumococcal Pbp2x from Pbp2b at septa 17
© 2014 John Wiley & Sons Ltd, Molecular Microbiology

2013). FLAG-tag strains [IU5544 (pbp1a-L-FLAG3), IU7434(stkP-FLAG2); IU4970 (mreC-L-FLAG3)] and HA-taggedstrains [IU6929 (pbp2x-HA); IU7426 (pbp2b-HA4)] werelabelled with primary anti-FLAG rabbit polyclonal antibody oranti-HA rabbit polyclonal antibody, respectively, followed byAlexa Fluor 568 highly cross-absorbed goat anti-rabbit IgG.DNA in nucleoids was stained by using mounting media Slow-Fade gold antifade reagent with DAPI. Similar cell morphologyand protein localization were observed in samples labelled ornot labelled with FL-V, indicating that the FL-V labelling pro-cedure did not alter localization of Pbp1a, Pbp2b, Pbp2x,MreC and StkP (data not shown).
Acknowledgements
We thank Amilcar Perez, Britta Rued, Kevin Bruce and DanKearns for helpful discussions. We thank the Indiana Univer-sity Light Microscopy Imaging Center (3D-SIM supported byNIH S10RR028697-01) and Jim Powers for technical supportand discussions. This work was supported by NIAIDgrants AI097289 and AI107075 (M.E.W.), by NIH grantDP2OD008592 (E.E.C), by a Pew Biomedical Scholar Award(E.E.C.), by NIGMS grant GM051986 (Y.V.B.), and by NSFgrant MCB-1157982 (S.L.S.). The contents of this paper aresolely the responsibility of the authors and do not necessarilyrepresent official views of the funding agencies.
References
Barendt, S.M., Land, A.D., Sham, L.T., Ng, W.L., Tsui, H.C.,Arnold, R.J., and Winkler, M.E. (2009) Influences ofcapsule on cell shape and chain formation of wild-type andpcsB mutants of serotype 2 Streptococcus pneumoniae. JBacteriol 191: 3024–3040.
Barendt, S.M., Sham, L.T., and Winkler, M.E. (2011) Charac-terization of mutants deficient in the L,D-carboxypeptidase(DacB) and WalRK (VicRK) regulon, involved in peptidog-lycan maturation of Streptococcus pneumoniae serotype 2strain D39. J Bacteriol 193: 2290–2300.
Bartual, S.G., Straume, D., Stamsas, G.A., Munoz, I.G.,Alfonso, C., Martinez-Ripoll, M., et al. (2014) Structuralbasis of PcsB-mediated cell separation in Streptococcuspneumoniae. Nat Commun 5: 3842.
Beilharz, K., Novakova, L., Fadda, D., Branny, P., Massidda,O., and Veening, J.W. (2012) Control of cell division inStreptococcus pneumoniae by the conserved Ser/Thrprotein kinase StkP. Proc Natl Acad Sci USA 109: E905–E913.
Berg, K.H., Stamsas, G.A., Straume, D., and Havarstein, L.S.(2013) Effects of low PBP2b levels on cell morphology andpeptidoglycan composition in Streptococcus pneumoniaeR6. J Bacteriol 195: 4342–4354.
Berg, K.H., Straume, D., and Havarstein, L.S. (2014) Thefunction of the transmembrane and cytoplasmic domains ofpneumococcal penicillin-binding proteins 2x and 2bextends beyond that of simple anchoring devices. Microbi-ology 160 (Pt 8): 1585–1598. doi: 10.1099/mic.0.078535-0
den Blaauwen, T., Andreu, J.M., and Monasterio, O. (2014)Bacterial cell division proteins as antibiotic targets. BioorgChem 55: 27–38.
Carvalho, S.M., Kuipers, O.P., and Neves, A.R. (2013) Envi-
ronmental and nutritional factors that affect growth andmetabolism of the pneumococcal serotype 2 strain D39and its nonencapsulated derivative strain R6. PLoS ONE8: e58492.
CDC (2013) Antibiotic resistance threats in the UnitedStates, 2013. http://www.cdc.gov/drugresistance/threat-report-2013/
Chan, P.F., O’Dwyer, K.M., Palmer, L.M., Ambrad, J.D.,Ingraham, K.A., So, C., et al. (2003) Characterization of anovel fucose-regulated promoter (PfcsK) suitable for geneessentiality and antibacterial mode-of-action studies inStreptococcus pneumoniae. J Bacteriol 185: 2051–2058.
Dalia, A.B., and Weiser, J.N. (2011) Minimization of bacterialsize allows for complement evasion and is overcome bythe agglutinating effect of antibody. Cell Host Microbe 10:486–496.
Daniel, R.A., and Errington, J. (2003) Control of cell morpho-genesis in bacteria: two distinct ways to make a rod-shaped cell. Cell 113: 767–776.
Denapaite, D., Bruckner, R., Hakenbeck, R., and Vollmer, W.(2012) Biosynthesis of teichoic acids in Streptococcuspneumoniae and closely related species: lessons fromgenomes. Microb Drug Resist 18: 344–358.
Donkor, E.S. (2013) Understanding the pneumococcus:transmission and evolution. Front Cell Infect Microbiol 3: 7.doi: 10.3389/fcimb.2013.00007
Eberhardt, A., Hoyland, C.N., Vollmer, D., Bisle, S.,Cleverley, R.M., Johnsborg, O., et al. (2012) Attachment ofcapsular polysaccharide to the cell wall in Streptococcuspneumoniae. Microb Drug Resist 18: 240–255.
Egan, A.J., and Vollmer, W. (2013) The physiology of bacte-rial cell division. Ann NY Acad Sci 1277: 8–28.
Fleurie, A., Cluzel, C., Guiral, S., Freton, C., Galisson, F.,Zanella-Cleon, I., et al. (2012) Mutational dissection of theS/T-kinase StkP reveals crucial roles in cell division ofStreptococcus pneumoniae. Mol Microbiol 83: 746–758.
Fleurie, A., Manuse, S., Zhao, C., Campo, N., Cluzel, C.,Lavergne, J.P., et al. (2014) Interplay of the serine/threonine-kinase StkP and the paralogs DivIVA and GpsBin pneumococcal cell elongation and division. PLoS Genet10: e1004275.
Garner, E.C., Bernard, R., Wang, W., Zhuang, X., Rudner,D.Z., and Mitchison, T. (2011) Coupled, circumferentialmotions of the cell wall synthesis machinery and MreBfilaments in B. subtilis. Science 333: 222–225.
Hakenbeck, R., Bruckner, R., Denapaite, D., and Maurer, P.(2012) Molecular mechanisms of beta-lactam resistance inStreptococcus pneumoniae. Future Microbiol 7: 395–410.
Henriques-Normark, B., and Tuomanen, E.I. (2013) Thepneumococcus: epidemiology, microbiology, and patho-genesis. Cold Spring Harb Perspect Med 3: a010215. doi:10.1101/cshperspect.a010215
Higgins, M.L., and Shockman, G.D. (1970) Model for cell wallgrowth of Streptococcus faecalis. J Bacteriol 101: 643–648.
Kocaoglu, O., Calvo, R.A., Sham, L.T., Cozy, L.M., Lanning,B.R., Francis, S., et al. (2012) Selective penicillin-bindingprotein imaging probes reveal substructure in bacterial celldivision. ACS Chem Biol 7: 1746–1753.
Kuru, E., Hughes, H.V., Brown, P.J., Hall, E., Tekkam, S.,Cava, F., et al. (2012) In Situ probing of newly synthesized
18 H.-C. T. Tsui et al. ■
© 2014 John Wiley & Sons Ltd, Molecular Microbiology

peptidoglycan in live bacteria with fluorescent D-aminoacids. Angew Chem Int Ed Engl 51: 12519–12523.
Land, A.D., and Winkler, M.E. (2011) The requirement forpneumococcal MreC and MreD is relieved by inactivationof the gene encoding PBP1a. J Bacteriol 193: 4166–4179.
Land, A.D., Tsui, H.C., Kocaoglu, O., Vella, S.A., Shaw, S.L.,Keen, S.K., et al. (2013) Requirement of essential Pbp2xand GpsB for septal ring closure in Streptococcus pneu-moniae D39. Mol Microbiol 90: 939–955.
Lanie, J.A., Ng, W.L., Kazmierczak, K.M., Andrzejewski, T.M.,Davidsen, T.M., Wayne, K.J., et al. (2007) Genomesequence of Avery’s virulent serotype 2 strain D39 of Strep-tococcus pneumoniae and comparison with that of unen-capsulated laboratory strain R6. J Bacteriol 189: 38–51.
Lee, T.K., Tropini, C., Hsin, J., Desmarais, S.M., Ursell, T.S.,Gong, E., et al. (2014) A dynamically assembled cell wallsynthesis machinery buffers cell growth. Proc Natl AcadSci USA 111: 4554–4559.
Lovering, A.L., Safadi, S.S., and Strynadka, N.C. (2012)Structural perspective of peptidoglycan biosynthesis andassembly. Annu Rev Biochem 81: 451–478.
Massidda, O., Novakova, L., and Vollmer, W. (2013) Frommodels to pathogens: how much have we learned aboutStreptococcus pneumoniae cell division? Environ Microbiol15: 3133–3157.
Morlot, C., Zapun, A., Dideberg, O., and Vernet, T. (2003)Growth and division of Streptococcus pneumoniae: locali-zation of the high molecular weight penicillin-binding pro-teins during the cell cycle. Mol Microbiol 50: 845–855.
Morlot, C., Noirclerc-Savoye, M., Zapun, A., Dideberg, O.,and Vernet, T. (2004) The D,D-carboxypeptidase PBP3organizes the division process of Streptococcus pneumo-niae. Mol Microbiol 51: 1641–1648.
Morlot, C., Bayle, L., Jacq, M., Fleurie, A., Tourcier, G.,Galisson, F., et al. (2013) Interaction of penicillin-bindingprotein 2x and Ser/Thr protein kinase StkP, two key playersin Streptococcus pneumoniae R6 morphogenesis. MolMicrobiol 90: 88–102.
Ng, W.L., Kazmierczak, K.M., and Winkler, M.E. (2004)Defective cell wall synthesis in Streptococcus pneumoniaeR6 depleted for the essential PcsB putative murein hydro-lase or the VicR (YycF) response regulator. Mol Microbiol53: 1161–1175.
Nikolaidis, I., Favini-Stabile, S., and Dessen, A. (2014)Resistance to antibiotics targeted to the bacterial cell wall.Protein Sci 23: 243–259.
Perez-Nunez, D., Briandet, R., David, B., Gautier, C.,Renault, P., Hallet, B., et al. (2011) A new morphogenesispathway in bacteria: unbalanced activity of cell wall syn-thesis machineries leads to coccus-to-rod transition andfilamentation in ovococci. Mol Microbiol 79: 759–771.
Peters, K., Schweizer, I., Beilharz, K., Stahlmann, C.,Veening, J.W., Hakenbeck, R., and Denapaite, D. (2014)Streptococcus pneumoniae PBP2x mid-cell localizationrequires the C-terminal PASTA domains and is essential forcell shape maintenance. Mol Microbiol 92: 733–755.
Pinho, M.G., and Errington, J. (2005) Recruitment ofpenicillin-binding protein PBP2 to the division site ofStaphylococcus aureus is dependent on its transpeptida-tion substrates. Mol Microbiol 55: 799–807.
Pinho, M.G., Kjos, M., and Veening, J.W. (2013) How to get
(a)round: mechanisms controlling growth and division ofcoccoid bacteria. Nat Rev Microbiol 11: 601–614.
van der Ploeg, R., Verheul, J., Vischer, N.O., Alexeeva, S.,Hoogendoorn, E., Postma, M., et al. (2013) Colocalizationand interaction between elongasome and divisome duringa preparative cell division phase in Escherichia coli. MolMicrobiol 87: 1074–1087.
Potluri, L.P., Kannan, S., and Young, K.D. (2012) ZipA isrequired for FtsZ-dependent preseptal peptidoglycan syn-thesis prior to invagination during cell division. J Bacteriol194: 5334–5342.
Ramos-Montanez, S., Tsui, H.C., Wayne, K.J., Morris, J.L.,Peters, L.E., Zhang, F., et al. (2008) Polymorphism andregulation of the spxB (pyruvate oxidase) virulence factorgene by a CBS-HotDog domain protein (SpxR) in serotype2 Streptococcus pneumoniae. Mol Microbiol 67: 729–746.
Rodriguez, J.L., Dalia, A.B., and Weiser, J.N. (2012)Increased chain length promotes pneumococcal adher-ence and colonization. Infect Immun 80: 3454–3459.
Sanders, A.N., and Pavelka, M.S. (2013) Phenotypic analysisof Eschericia coli mutants lacking L,D-transpeptidases.Microbiology 159: 1842–1852.
Schneewind, O., and Missiakas, D. (2013) Sec-secretion andsortase-mediated anchoring of proteins in Gram-positivebacteria. Biochim Biophys Acta 1843: 1687–1697. doi:10.1016/j.bbamcr.2013.11.009
Sham, L.T., Barendt, S.M., Kopecky, K.E., and Winkler, M.E.(2011) Essential PcsB putative peptidoglycan hydrolaseinteracts with the essential FtsXSpn cell division protein inStreptococcus pneumoniae D39. Proc Natl Acad Sci USA108: E1061–E1069.
Sham, L.T., Tsui, H.C., Land, A.D., Barendt, S.M., andWinkler, M.E. (2012) Recent advances in pneumococcalpeptidoglycan biosynthesis suggest new vaccine and anti-microbial targets. Curr Opin Microbiol 15: 194–203.
Sham, L.T., Jensen, K.R., Bruce, K.E., and Winkler, M.E.(2013) Involvement of FtsE ATPase and FtsX extracellularloops 1 and 2 in FtsEX-PcsB complex function in celldivision of Streptococcus pneumoniae D39. mBio 4:e00431-13. doi: 10.1128/mBio.00431-13
Singh, S.K., SaiSree, L., Amrutha, R.N., and Reddy, M.(2012) Three redundant murein endopeptidases catalysean essential cleavage step in peptidoglycan synthesis ofEscherichia coli K12. Mol Microbiol 86: 1036–1051.
Tomasz, A., and Fischer, W. (2006) The cell wall of Strepto-coccus pneumonaie. In Gram-postive Pathogens. Fischetti,V.A., Novick, R.P., Ferretti, J.J., Portnoy, D.A., and Rood,J.I. (eds). Washington, DC: ASM Press, pp. 230–240.
Tsui, H.C., Mukherjee, D., Ray, V.A., Sham, L.T., Feig, A.L.,and Winkler, M.E. (2010) Identification and characteriza-tion of noncoding small RNAs in Streptococcus pneumo-niae serotype 2 strain D39. J Bacteriol 192: 264–279.
Tsui, H.C., Keen, S.K., Sham, L.T., Wayne, K.J., and Winkler,M.E. (2011) Dynamic distribution of the SecA and SecYtranslocase subunits and septal localization of the HtrAsurface chaperone/protease during Streptococcus pneu-moniae D39 cell division. mBio 2: e00202-11. doi: 10.1128/mBio.00202-11
Typas, A., Banzhaf, M., Gross, C.A., and Vollmer, W. (2012)From the regulation of peptidoglycan synthesis to bacterialgrowth and morphology. Nat Rev Microbiol 10: 123–136.
Separation of pneumococcal Pbp2x from Pbp2b at septa 19
© 2014 John Wiley & Sons Ltd, Molecular Microbiology

Vernatter, J., and Pirofski, L.A. (2013) Current concepts inhost-microbe interaction leading to pneumococcal pneu-monia. Curr Opin Infect Dis 26: 277–283.
Wayne, K.J., Sham, L.T., Tsui, H.C., Gutu, A.D., Barendt,S.M., Keen, S.K., and Winkler, M.E. (2010) Localizationand cellular amounts of the WalRKJ (VicRKX) two-component regulatory system proteins in serotype 2 Strep-tococcus pneumoniae. J Bacteriol 192: 4388–4394.
Wheeler, R., Mesnage, S., Boneca, I.G., Hobbs, J.K., andFoster, S.J. (2011) Super-resolution microscopy revealscell wall dynamics and peptidoglycan architecture in ovo-coccal bacteria. Mol Microbiol 82: 1096–1109.
Zapun, A., Contreras-Martel, C., and Vernet, T. (2008a)Penicillin-binding proteins and beta-lactam resistance.FEMS Microbiol Rev 32: 361–385.
Zapun, A., Vernet, T., and Pinho, M.G. (2008b) The differentshapes of cocci. FEMS Microbiol Rev 32: 345–360.
Zerfass, I., Hakenbeck, R., and Denapaite, D. (2009) Animportant site in PBP2x of penicillin-resistant clinical iso-lates of Streptococcus pneumoniae: mutational analysis ofThr338. Antimicrob Agents Chemother 53: 1107–1115.
Zhao, G., Meier, T.I., Kahl, S.D., Gee, K.R., and Blaszczak,L.C. (1999) BOCILLIN FL, a sensitive and commerciallyavailable reagent for detection of penicillin-binding pro-teins. Antimicrob Agents Chemother 43: 1124–1128.
Supporting information
Additional supporting information may be found in the onlineversion of this article at the publisher’s web-site.
20 H.-C. T. Tsui et al. ■
© 2014 John Wiley & Sons Ltd, Molecular Microbiology
Copyright © 2022 FDOKUMEN




















