
Stages of Child Development : –Prenatal; and –Birth to Three ...

B
PugES
Otc
Scihrm
Ra
Cg
Ep(tp
FDo(GDHM
R2
RoNCd
TBRF
0©d
Research www.AJOG.orgARTICLE IN PRESS
ASIC SCIENCE: OBSTETRICS
artial contributions of developmental hypoxia andndernutrition to prenatal alterations in somaticrowth and cardiovascular structure and function
mily J. Camm, PhD; Jeremy A. Hansell, PhD; Andrew D. Kane, BA; Emilio A. Herrera, PhD; Cara Lewis, BA;amuel Wong, BA; Nicholas W. Morrell, MD, FRCP; Dino A. Giussani, PhD
bvtcf
Ctf
K
BJECTIVE: The objective of the study was to compare and contrasthe effects of developmental hypoxia vs undernutrition on fetal growth,ardiovascular morphology, and function.
TUDY DESIGN: On day 15 of gestation, Wistar dams were divided intoontrol, hypoxic (10% O2), or undernourished (35% reduction in foodntake) pregnancy. On day 20, fetal thoraces were fixed, and the fetaleart and aorta underwent quantitative histological analysis. In a sepa-ate group, fetal aortic vascular reactivity was determined via wireyography.
ESULTS: Both hypoxic and undernourished pregnancy was associ-
rowth and cardiovascular structure and function. Am J Obstet Gynecol 2010;203:xx
mIhtctfidtsIdiwda
cfgtnaornrdelivery to the fetus bu
oi: 10.1016/j.ajog.2010.06.046
y hypoxia promoted fetal aortic thickening without changes in cardiacolumes when expressed as a percentage of total heart volume. In con-rast, maternal undernutrition affected fetal cardiac morphology withouthanges in aortic structure. Fetal aortic vascular reactivity was also dif-erentially affected by hypoxia or undernutrition.
ONCLUSION: Developmental hypoxia or undernutrition in late gesta-ion has differential effects on fetal cardiovascular morphology andunction.
ey words: cardiovascular, hypoxia, placental insufficiency,
ted with asymmetric fetal growth restriction. Pregnancy complicated preeclampsia, undernutritionite this article as: Camm EJ, Hansell JA, Kane AD, et al. Partial contributions of developmental hypoxia and undernutrition to prenatal alterations in somatic
-xx.ptc
omibItiectgifu
matocpcc
pidemiological and experimental evi-dence suggests that, in complicated
regnancy, intrauterine growth restrictionIUGR) is associated with early changes inhe developing cardiovascular system. Re-orted abnormalities in cardiovascular
rom the Department of Physiology,evelopment, and Neuroscience, Universityf Cambridge, Cambridge, United KingdomDrs Camm, Hansell, Kane, Herrera, andiussani and Miss Lewis and Mr Wong); andepartment of Medicine, Addenbrooke’sospital, Cambridge, United Kingdom (Drorrell).
eceived March 2, 2010; revised May 28,010; accepted June 17, 2010.
eprints: Dino A. Giussani, PhD, Departmentf Physiology, Development, andeuroscience, University of Cambridge,ambridge CB2 3EG, United [email protected].
his study was supported in part by theiotechnology and Biological Sciencesesearch Council and the British Heartoundation.
002-9378/$36.002010 Mosby, Inc. All rights reserved.
orphology and function of the humanUGR fetus include an increase in relativeeart weight and ventricular wall hyper-rophy,1 a decrease in ventricle and myo-yte volume,2 and compromised biven-ricular ejection force3 and diastoliclling.4 Atheromatous changes in the ab-ominal aorta5 and thickening of the aor-ic wall, even when adjusted for small bodyize,6,7 have also been documented inUGR offspring in early childhood. In ad-ition, studies have reported a significant
nverse relationship between low birth-eightandendothelialdysfunction inchil-ren in the first decade of life8 and in earlydulthood.9
In complicated pregnancy, the mostommon challenges to the developingetus are reductions in nutrient and oxy-en delivery. However, the partial con-ributions of prenatal hypoxia vs under-utrition to slow fetal growth andlterations in the structure and functionf the developing cardiovascular systememain uncertain. For instance, preg-ancies complicated by undernutritionesult in not only a decrease in nutrient
t also a decrease in l
MONTH 2010 Ame
lacental blood flow,10-12 which will fur-her compound nutrition as well as de-rease oxygen delivery to the fetus.
Similarly, the natural hypobaric hyp-xia of pregnancy at high altitude in hu-ans13,14 and experimental maternal
sobaric hypoxia in rats15-20 have botheen extensively reported to promoteUGR. However, because most high-al-itude populations are also impover-shed and because maternal hypoxia inxperimental rats can lead to a signifi-ant decrease in maternal food intake,21
he extent to which any effects on fetalrowth and cardiovascular developments governed by fetal undernutrition oretal underoxygenation again remainsncertain.Using the chick embryo as an animalodel, a cluster of studies have been
ble to isolate the effects on growth andhe developing cardiovascular systemf chronic hypoxia, independent ofhanges in maternal nutrition and of thehysiology of the mother and the pla-enta.22-29 However, no study to date hasompared and contrasted in a mamma-
ian species the partial contributions ofrican Journal of Obstetrics & Gynecology 1.e1

ptcd
iptgtsponflaa
MAAcP(h(p(lcRarpWTc�pham
EAmctnnpcamt
lumouiMbc
fodiupbfinsmp�
TOfodtAxMUmpsptwfaws
cstTdmitq
ttibw
HPihLsvsessfiOac
C(BbvcmesdtaefAc
cptomcoromcotca
Research Basic Science: Obstetrics www.AJOG.org
1
ARTICLE IN PRESS
renatal hypoxia vs prenatal undernutri-ion on alterations in fetal growth andardiovascular development already evi-ent prior to birth.This study tested the hypothesis that,
n mammalian complicated pregnancy,renatal hypoxia and prenatal undernu-rition have differential effects on fetalrowth and on the anatomy and func-ion of the developing cardiovascularystem. The hypothesis was tested inregnant rats by investigating the effectsf maternal hypoxia vs maternal under-utrition in the last third of gestation on
etal biometry, stereological and histo-ogical changes in the fetal heart andorta, and functional changes in fetalortic vascular reactivity.
ATERIALS AND METHODSnimals, breeding, and housingll procedures involving animals werearried out under the Animals (Scientificrocedures) Act of 1986. Wistar ratsCharles River Ltd, Margate, UK) wereoused in individually ventilated cagesIVC units, 21% O2, 70-80 air changeser hour) under standard conditions60% humidity, 21°C and a 12 houright, 12 hour dark cycle), with free ac-ess to food (maintenance diet; Charlesiver Ltd) and water. After 10 days ofcclimatization, virgin female Wistarats (n � 39, 10-12 weeks of age) wereaired individually with fertile maleistar rats (minimum 12 weeks of age).
he presence of a copulatory plug wasonsidered day 0 of pregnancy (term22 days). Upon the confirmation of
regnancy, the female was weighed andoused individually. Maternal weightnd food and water consumption wereonitored daily.
xperimental designt day 15 of gestation, 14 pregnant fe-ales were placed inside a hypoxic
hamber, which combined a PVC isola-or (PFI Plastics Ltd, Keynes, UK) with aitrogen generator (N2MID60, Dom-ick Hunter Ltd, Warwick, UK). Theercentage of oxygen in the chamber wasontrolled by altering the inflow of airnd nitrogen. Oxygen concentration wasonitored continuously throughout the
reatment period with an oxygen ana- t
.e2 American Journal of Obstetrics & Gynecology
yzer (ICA, London, UK). Pregnanciesndergoing maternal hypoxia wereaintained at an inspired fraction of
xygen of 10% from day 15 of gestationntil day 20. The chamber was housed
n the same room as the IVC units.aternal 10% isobaric hypoxia has
een shown to decrease maternal foodonsumption.19,21,30
To discriminate between the direct ef-ects on development of prenatal hyp-xia and those induced by hypoxia-in-uced reductions in maternal food
ntake, 11 dams underwent pregnancynder normoxic conditions but wereair fed to the daily amount consumedy hypoxic dams from day 15 onward. Anal group of 14 dams underwent preg-ancy under normoxic conditions toerve as controls. This yielded 3 experi-
ental groups: control (n � 14), hy-oxic (n � 14), and undernourished (n
11) pregnancy.
issue collectionn day 20 of gestation, some of the dams
rom each group (control: n � 7; hyp-xia: n � 7; undernutrition: n � 5) wereeeply anesthetized with ketamine (in-raperitoneally, 40 mg/kg–1, Fort Dodgenimal Health, Southampton, UK) andylazine (intraperitoneally, 5 mg/kg–1;illpledge Veterinary, Gainsborough,K). The uterus was then exposed via aidline incision, and the anesthetized
ups isolated and killed via spinal tran-ection. In dams that had been in the hy-oxic chamber from days 15 to 20 of ges-ation, this procedure was carried outhile ventilated with 10% oxygen via a
ace mask. All fetuses and their associ-ted placentae were isolated andeighed. The fetal sex was noted by mea-
urement of the anogenital distance.To assess the symmetry of growth,
rown-rump length (CRL) was mea-ured using digital callipers. Weights ofhe brain and liver were also recorded.o assess alterations in cardiovascularevelopment, the fetal thorax from 1ale pup per litter was immersion fixed
n 4% paraformaldehyde for 5 days andhen processed into paraffin for subse-uent stereological analysis.The remaining dams were assigned to
he in vitro wire myography study (con- t
MONTH 2010
rol: n � 7; hypoxia: n � 7; undernutri-ion: n � 6) and killed via carbon dioxidenhalation. Fetuses were removed andody and organs weights, as well as CRL,ere recorded.
istologyaraffin-embedded fetal thoraces, which
ncluded the heart and aorta, were ex-austively sectioned at 5 �m using aeica RM 2235 microtome (Leica Micro-ystems, Heidelberg, Germany). Trans-erse sections were stained with Mas-on’s Trichrome and hematoxylin andosin to identify cardiac muscle andmooth muscle cell (SMC) nuclei, re-pectively. All quantitative analyses ofxed tissue were performed using anlympus BX-50 microscope, fitted withmotorized specimen stage and micro-
ator (Olympus, Tokyo, Japan).All analyses were performed using the
omputer Assisted Stereology ToolboxCAST) version 2.0 program (Olympus,allerup, Denmark), with the observerlind to the treatment groups. A meanalue of the variable being analyzed wasalculated for each animal, and a mean ofeans � SEM was then obtained for
ach treatment group. To account forhrinkage due to paraffin processing, theiameter of erythrocytes present in sec-ions of fetal fixed tissue was measurednd compared with the diameter ofrythrocytes present in fresh blood frometal rats of the same gestational age.31
ll measurements were subsequentlyorrected using this factor.
The volume of the fetal heart and itsompartments was determined using aoint grid, which was superimposed onhe sections and viewed using a �1.25bjective. To ensure that a precise esti-ate of the volumes of the different
ompartments of the heart and the areasf the wall and lumen of the aorta waseached, which was unaffected by inter-r intraobserver variability, approxi-ately 200 points on 10-15 sections were
ounted for each animal.32 Points fallingn the left ventricular wall plus interven-ricular septum, left lumen, right ventri-le wall, and right lumen were counted,nd the Cavalieri Principle32 was applied
o calculate estimated volumes:
wttst�c
lpvawuepwe
wt�r
nwmsfnm
talowaeefc
IFoidtsit4
V(PC
www.AJOG.org Basic Science: Obstetrics ResearchARTICLE IN PRESS
V�obj� � t x a�p� � � P
here V(obj) is the estimated volume ofhe heart compartment, t is the totalhickness of the heart (t � number ofections � sections thickness), a(p) ishe area associated with each point, andP is the sum of points for that
ompartment.To determine the areas of the wall and
umen of the aortae, a point grid was su-erimposed on each section and wasiewed using a �10 objective. The aver-ge area and wall thickness of the aortaas measured from the base of the heartntil the start of the aortic arch. Thisquated to approximately 10-11 sectionser animal. Points falling on either theall or lumen were counted and the ar-
as were calculated as:
A�obj� � a�p� x � P
here A(obj)is the estimated area, a(p)ishe area associated with each point, andP is the sum of points falling on the
elevant area, averaged over the sections.Using the CAST program, the thick-
ess for the left and right free ventricularalls and aortic walls were also esti-ated. A line grid was superimposed on
ections to establish random start pointsor measuring distances between the in-er and the outer membrane using theethod of orthogonal intercepts.33
The density of SMC nuclei in the aor-ic wall was assessed in 5 hematoxylinnd eosin–stained sections spanning theength of the thoracic aorta. Using a �60bjective, the number of SMC nucleiithin a counting frame (617 �m2) was
ssessed in 10 fields per section. The av-rage density was then calculated forach animal and then a mean of meansor each group. Approximately 200 nu-lei were counted per animal.
n vitro wire myographyetal thoracic aortas were dissected freef adipose and connective tissue and cut
nto 2 � 2 mm sections. Two 40 �miameter stainless steel wires werehreaded through the lumen of the aorticections, maintaining the endotheliumntact. The wires were then placed be-ween the mounting support jaws of a
-chamber small-vessel wire myographFIGURE 1Maternal food intake during pregnancy
alues are mean � SEM for maternal food consumption in control (n � 14, white diamonds ), hypoxicn � 14, black squares ), and undernourished (n � 11, gray triangles ) pregnancies. Asterisk indicates� .05 vs control (2-way repeated-measures ANOVA with post-hoc Tukey test).
amm. Fetal cardiovascular morphology and function following complicated pregnancy. Am J Obstet Gynecol 2010.
TABLE 1Fetal data at day 20 of gestationVariable Control Hypoxic Undernourished
Fetus, g 3.48 � 0.07 3.00 � 0.06a 3.10 � 0.05a
..............................................................................................................................................................................................................................................
CRL, mm 36.0 � 0.4 34.2 � 0.3a 34.5 � 0.4a
..............................................................................................................................................................................................................................................
BMI, g/mm2 2.72 � 0.04 2.57 � 0.04a 2.62 � 0.04a
..............................................................................................................................................................................................................................................
Placenta, g 0.503 � 0.008 0.521 � 0.008 0.488 � 0.010b
..............................................................................................................................................................................................................................................
BW/PW 7.03 � 0.15 5.80 � 0.10a 6.43 � 0.11a,b
..............................................................................................................................................................................................................................................
Heart.....................................................................................................................................................................................................................................
Absolute, g 0.027 � 0.001 0.025 � 0.001 0.025 � 0.001.....................................................................................................................................................................................................................................
Relative, % 0.75 � 0.02 0.85 � 0.04a 0.81 � 0.04..............................................................................................................................................................................................................................................
Brain.....................................................................................................................................................................................................................................
Absolute, g 0.159 � 0.004 0.154 � 0.004 0.150 � 0.004.....................................................................................................................................................................................................................................
Relative, % 4.53 � 0.11 5.16 � 0.18a 4.89 � 0.16a
..............................................................................................................................................................................................................................................
Liver.....................................................................................................................................................................................................................................
Absolute, g 0.291 � 0.009 0.224 � 0.007a 0.243 � 0.009a
.....................................................................................................................................................................................................................................
Relative, % 8.28 � 0.15 7.42 � 0.18a 7.80 � 0.19a
..............................................................................................................................................................................................................................................
Brain/liver 0.55 � 0.01 0.71 � 0.04a 0.64 � 0.03a
..............................................................................................................................................................................................................................................
Values are mean � SEM in fetal rats from control (n � 14 litters), hypoxic (n � 14 litters), and undernourished (n � 11 litters)offspring.BMI, body mass index; BW, body weight; CRL, crown-rump length; PW, placental weight.a P � .05 vs control; b P � .05 vs hypoxic (1-way ANOVA with post-hoc Tukey test).
Camm. Fetal cardiovascular morphology and function following complicated pregnancy. Am J Obstet Gynecol 2010.
MONTH 2010 American Journal of Obstetrics & Gynecology 1.e3

(DwOmcU
c8atetpost
fvcC(PMatds1Aw(mnwllc
DTit1olaamtfS
ptttaCaaE5
gapt(Scs
REcIm
spa(miceda
ETs0.(uf�.mwn(
wHhctncnwbbpscCiia
HFwpulasdo
lowi
Research Basic Science: Obstetrics www.AJOG.org
1
ARTICLE IN PRESS
Multi Wire Myograph System 610M;MT, Aarhus, Denmark) containingarmed oxygenated Kreb’s buffer (95%
2/5% CO2 mix, 37°C). The vessel di-ensions were measured using a digital
amera (Olympus, Southend-on-Sea,K).Force data from the myograph were re-
orded at 4 Hz (Labchart 6.0, Powerlab/30; AD Instruments, Chalgrove, UK),nd each vessel was standardized to an op-imal working tension.34 To our knowl-dge, the arterial pressure is unknown inhe fetal rat; therefore, different workingressures were tested. Fifteen millimetersf mercury yielded optimal vascular re-ponses and were thereby used throughouthe experiment.
Both vasoconstrictor and vasodilatorunction was assessed in the vessels. Allessels were contracted with increasingoncentrations of KCl (0-125 mM).onstrictor function to phenylephrine
PE) (10–9 to 10–5 M; Sigma Aldrich,oole, UK) and U46619 (10–14 to 10– 6
; Cayman Chemicals, Tallinn, Estonia)nd vasodilator responses to sodium ni-roprusside (10–9 to 10– 4 M; Sigma Al-rich) and endothelium-dependent va-odilatation with acetylcholine (10–9 to0– 4.5 M; Sigma Aldrich) were assessed.ll vasodilator responses were assessedith the vessels precontracted with PE
10– 6 M). Responses were recorded for 2inutes after each dose, whereupon the
ext cumulative dose was given. Vesselsere repeatedly washed with Kreb’s so-
ution and allowed to equilibrate for ateast 20 minutes between different con-
TABLE 2Fetal aortic measurementsVariable Control
Wall area, mm2 0.057 � 0.0...................................................................................................................
Lumen area, mm2 0.120 � 0.0...................................................................................................................
Whole vessel area, mm2 0.177 � 0.0...................................................................................................................
Wall to lumen ratio 0.525 � 0.0...................................................................................................................
Media wall thickness, �m 37.5 � 1.8...................................................................................................................
Values are mean � SEM for aortic measurements in fetal rats� 5) offspring.a P � .05 vs control (1-way ANOVA with post-hoc Tukey test)
Camm. Fetal cardiovascular morphology and function fol
entration-response curves. d
.e4 American Journal of Obstetrics & Gynecology
ata and statistical analyseso account for sex differences and with-
n-litter variation, data from no morehan 2 male pups from any 1 litter for anyvariable were taken, with the exceptionf any biometric variables. For the histo-
ogical studies, values given as percent-ges or ratios were first transformed torcsin to equalize variance. For the wireyography studies, cumulative concen-
ration-response curves were generatedor each vessel and plotted as mean �EM.Vasoconstrictor function was ex-
ressed relative to the maximal responseo KCl (125 mM) and vasodilator func-ion expressed relative to the precon-racted state. Individual curves were thennalyzed (GraphPad Prism, San Diego,A) using a sigmoidal fit curve with vari-ble slope for maximal response and pD2
s a measure of sensitivity (pD2 � –log10
C50, where EC50 is the dose required for0% of the maximal effect).Comparisons between experimental
roups were made using either 1-waynalysis of variance (ANOVA) or re-eated-measures ANOVA followed byhe Tukey post-hoc test as appropriateSigmaStat for Windows, version 2.0;PSS Inc, Chicago, IL). For all statisticalomparisons, P � .05 was consideredtatistically significant.
ESULTSffects on maternal food and wateronsumption and weight gainn all pregnancies, irrespective of treat-
ent, maternal food intake between
Hypoxic Undernourished
0.065 � 0.004 0.070 � 0.004..................................................................................................................
0.079 � 0.009a 0.114 � 0.009..................................................................................................................
0.145 � 0.011 0.185 � 0.013..................................................................................................................
0.901 � 0.115a 0.623 � 0.035..................................................................................................................
43.2 � 1.4a 37.9 � 1.9..................................................................................................................
control (n � 7), hypoxic (n � 7), and undernourished (n
ng complicated pregnancy. Am J Obstet Gynecol 2010.
ays 0 and 15 of gestation did not vary p
MONTH 2010
ignificantly and averaged 22.7 � 0.2 ger day (Figure 1). This equated to anverage daily caloric intake of 277 � 3 kJ66.3 � 0.6 kcal). During the 5 day treat-ent period, hypoxic and undernour-
shed dams ate significantly less thanontrols (Figure 1). For both groups, thisquated to a similar and approximate re-uction in maternal food consumptionnd caloric intake by 35% of baseline.
ffects on fetal biometryhere was no effect of treatment on litter
ize (control: 12.4 � 0.7; hypoxia: 12.2 �.8; undernourished: 11.6 � 0.8; P �
05) or on the number per litter of malescontrol: 6.3 � 0.4; hypoxia: 6.2 � 0.5;ndernourished: 6.2 � 0.6; P � .05) or
emales (control: 6.1 � 0.6; hypoxia: 6.00.6; undernourished: 5.5 � 0.5; P �
05). Fetal body weight, CRL, and bodyass index (BMI) at day 20 of gestationere reduced in hypoxic and under-ourished fetuses relative to controlsTable 1).
Undernutrition reduced placentaleight relative to hypoxic pregnancies.ypoxic and undernourished fetuses
ad a decreased fetal body weight to pla-ental weight ratio compared with con-rol fetuses. However, this ratio was sig-ificantly lower in fetuses from hypoxicompared with undernourished preg-ancies. Although relative heart weightas significantly increased in hypoxicut not undernourished fetuses, relativerain weight was increased in both hy-oxic and undernourished fetuses. Ab-olute and relative liver weights were de-reased in both experimental cohorts.onsequently, hypoxic and undernour-
shed pregnancies resulted in an increasen the fetal brain to liver weight ratio rel-tive to control pregnancies.
istological analysisetal aorta: the absolute areas of the aorticall and whole vessel were not affected byregnancy compromised by hypoxia orndernutrition (Table 2). However, abso-
ute lumen area and media wall thickness,s well as the wall to lumen area ratio, wereignificantly altered in hypoxic but not un-ernourished pregnancies. When the areasf the lumen and wall were expressed as a
04.........
09.........
12.........
85.........
.........
from
.
ercentage of the total vessel area (Figure

2ssc
thb
pan
AeAC
www.AJOG.org Basic Science: Obstetrics ResearchARTICLE IN PRESS
, B), both the wall and lumen areas wereignificantly increased and decreased, re-pectively, in hypoxic fetuses. Relative to
FIGURE 2Photomicrographs and summary o
, Photomicrographs of fetal aortae stained with hexpressed as a percentage of total vessel area and Csterisk indicates P � .05 vs control (1-way ANOVamm. Fetal cardiovascular morphology and function follow
ontrols, the density of SMC nuclei in 2
he media wall (used as an indicator ofyperplasia) was increased in hypoxic,ut not undernourished, fetuses (Figure
tal aortic morphometry
oxylin and eosin (H&E). B, Histograms are mean �nsity of smooth muscle cell nuclei in the media walith post-hoc Tukey test). Scale bar, 5 �m. Controlomplicated pregnancy. Am J Obstet Gynecol 2010.
, C). Conversely, the profile area occu- p
MONTH 2010 Ame
ied per smooth muscle nuclei (used asn indicator of hypertrophy) was not sig-ificantly different from that in control
M for the areas of the wall and lumen of the aortaecontrol, hypoxic, and undernourished pregnancies.
7; hypoxia: n � 7; undernutrition: n � 5.
f fe
mat SE, de l, inA w : n �ing c
regnancies (control: 8.42 � 0.62 � 10– 6
rican Journal of Obstetrics & Gynecology 1.e5

mum
twpflcluwnvvdnuta
tetsswp
ITlwg45u�sap
tbtsgfst
nmpHdr(
CMddtitgpsatpaatiwrrstI
mwchnpagrmear
lowi
Research Basic Science: Obstetrics www.AJOG.org
1
ARTICLE IN PRESS
m2; hypoxia: 7.58 � 0.54 � 10– 6 mm2;ndernourished: 9.74 � 0.50 � 10– 6
m2; P � .05).Fetal heart: the relative volumes of the
otal heart and the left and right ventricularalls were significantly increased in hy-oxic fetuses when compared with control
etuses (Table 3). In contrast, the absoluteeft ventricular wall volume was signifi-antly reduced and relative right ventricu-ar wall volume significantly increased inndernourished fetuses when comparedith control fetuses. In addition, in under-ourished fetuses, the relative total heartolume and the absolute and relative leftentricular wall volumes were significantlyifferent from hypoxic fetuses. There wereo significant alterations in lumen vol-mes or ventricular wall thickness of either
he left or right sides of the fetal heart by
TABLE 3Fetal cardiac measurementsVariable Cont
Total heart volume..........................................................................................................
Absolute, mm3 13.9..........................................................................................................
Relative, mm3/g 3.7...................................................................................................................
Ventricle wall volume..........................................................................................................
LV absolute, mm3 7.7..........................................................................................................
LV relative, mm3/g 2.1..........................................................................................................
RV absolute, mm3 4.4..........................................................................................................
RV relative, mm3/g 1.2...................................................................................................................
Lumen volume..........................................................................................................
LV absolute, mm3 1.0..........................................................................................................
LV relative, mm3/g 0.2..........................................................................................................
RV absolute, mm3 0.7..........................................................................................................
RV relative, mm3/g 0.1...................................................................................................................
Wall thickness..........................................................................................................
LV absolute, �m 693.5..........................................................................................................
LV relative, �m/g 187.3..........................................................................................................
LV relative heart volume, �m/mm3 13.6..........................................................................................................
RV absolute, �m 544.7..........................................................................................................
RV relative, �m/g 147.6..........................................................................................................
RV relative heart volume, �m/mm3 10.7...................................................................................................................
Values are mean � SEM absolute and relative to body weight� 7), hypoxic (n � 7), and undernourished (n � 5) offspringLV, left ventricle; RV, right ventricle.a P � .05 vs control; b P � .05 vs hypoxic (1-way ANOVA wi
Camm. Fetal cardiovascular morphology and function fol
ny treatment during pregnancy. When u
.e6 American Journal of Obstetrics & Gynecology
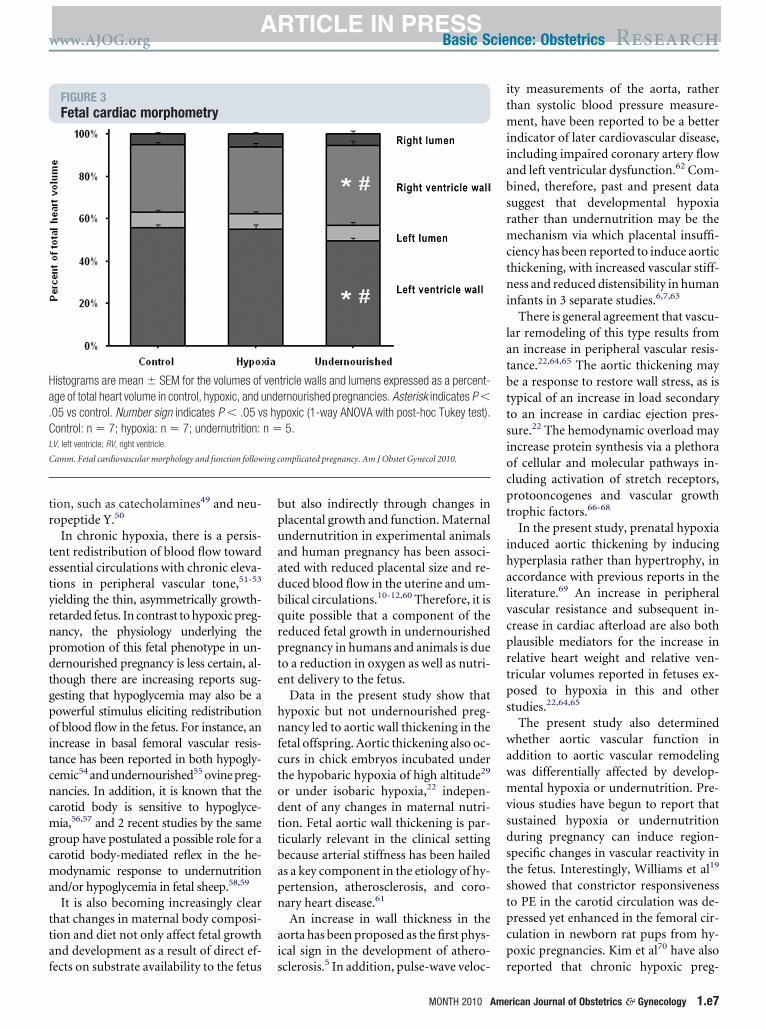
he partial components of the heart werexpressed relative to total heart volume,he left and right ventricular volumes wereignificantly increased and decreased, re-pectively, in undernourished fetuseshen compared with both control and hy-oxic fetuses (Figure 3).
n vitro wire myographyhe internal and external diameter and
ength of aortic segment being testedere similar for each experimentalroup (normoxia: 491 � 38 �m, 645 �1 �m, 1.81 � 0.08 mm; hypoxia: 509 �1 �m, 653 � 53 �m, 1.72 � 0.12 mm;ndernourished: 470 � 21 �m, 602 � 25m, 1.72 � 0.08 mm). Maximal con-
triction to KCl was depressed in fetalortas from undernourished but not hy-oxic offspring relative to controls (Fig-
Hypoxic Undernourished
..................................................................................................................
0.70 14.63 � 0.54 12.82 � 0.58..................................................................................................................
0.22 5.04 � 0.24a 4.30 � 0.09b
..................................................................................................................
..................................................................................................................
0.30 8.10 � 0.53 6.33 � 0.37a,b
..................................................................................................................
0.11 2.80 � 0.22a 2.12 � 0.06b
..................................................................................................................
0.25 4.60 � 0.18 4.87 � 0.36..................................................................................................................
0.08 1.58 � 0.06a 1.64 � 0.11a
..................................................................................................................
..................................................................................................................
0.16 1.06 � 0.16 0.94 � 0.22..................................................................................................................
0.05 0.37 � 0.06 0.32 � 0.07..................................................................................................................
0.13 0.88 � 0.13 0.68 � 0.22..................................................................................................................
0.04 0.30 � 0.04 0.23 � 0.07..................................................................................................................
..................................................................................................................
39.5 680.7 � 39.7 690.5 � 58.1..................................................................................................................
11.0 235.1 � 17.2 232.3 � 19.5..................................................................................................................
1.1 16.0 � 0.8 18.4 � 2.0..................................................................................................................
26.7 545.7 � 32.6 549.7 � 61.8..................................................................................................................
8.7 188.5 � 13.9 184.6 � 20.0..................................................................................................................
0.7 12.9 � 0.9 14.6 � 2.0..................................................................................................................
es for cardiac measurements in fetal rats, from control (n
st-hoc Tukey test).
ng complicated pregnancy. Am J Obstet Gynecol 2010.
re 4, A; P � .05). Conversely, the sensi- t
MONTH 2010
ivity to KCl was increased in hypoxicut not undernourished offspring rela-ive to controls. Vasoconstrictor re-ponses to PE were not different betweenroups (Figure 4, B; P � .05). Fetal aortasrom undernourished offspring alsohowed reduced vasoconstrictor sensi-ivity to U46619 (Figure 4, C; P � .05).
Fetal aortas from undernourished, butot hypoxic, offspring showed enhancedaximal relaxation to sodium nitro-
russide (SNP) (Figure 5, A; P � .05).ypoxic or undernourished pregnancy
id not alter the sensitivity or maximalesponse of fetal aortas to methacholineMetCh) (Figure 5, B; P � .05).
OMMENTaternal isobaric hypoxia-induced re-
uctions in fetal growth have been wellocumented in rats.19,20 It is also knownhat birthweight decreases with increas-ng high altitude in human popula-ions.14,35-37 Similarly, a decrease in fetalrowth following undernutrition duringregnancy has previously been demon-trated in the rat,38 guinea pig,39 sheep,40
nd human.41 In the present study, fe-uses from hypoxic or undernourishedregnancies were not only lighter butlso thin for their length, as evidenced bysignificant fall in their BMI. In addi-
ion, in both hypoxic and undernour-shed pregnancy, the fetal relative braineight and the fetal brain to liver weight
atio were increased. Thinness at birth, aelative decrease in liver weight and con-ervation of brain growth are all indica-ors of brain sparing and asymmetricUGR.42,43
These observations have many com-onalities with the findings of Barker, 44
ho related thinness at birth with in-reased rates of cardiovascular disease inuman populations. In hypoxic preg-ancy, the redistribution of cardiac out-ut in the fetus away from peripheralnd towards essential circulations is trig-ered by a carotid chemoreflex.45,46 Theedistribution of blood flow is thenaintained via local vasodilator influ-
nces in essential circulations, such asdenosine47 and nitric oxide,48 and theelease of agents into the fetal circulation
rol
.........
6 �.........
8 �.........
.........
3 �.........
0 �.........
6 �.........
1 �.........
.........
6 �.........
9 �.........
0 �.........
9 �.........
.........
�.........
�.........
�.........
�.........
�.........
�.........
volum.
th po
hat promote peripheral vasoconstric-
tr
tetyrnpdtgpoitcncmgcma
ttaf
bpuaadbqrpte
hnfctodttbapn
ai
itmiiabsrmctni
latbttsiocpt
ihalvcprtps
wawmvsdststpcp
Ha.CL
C
www.AJOG.org Basic Science: Obstetrics ResearchARTICLE IN PRESS
ion, such as catecholamines49 and neu-opeptide Y.50
In chronic hypoxia, there is a persis-ent redistribution of blood flow towardssential circulations with chronic eleva-ions in peripheral vascular tone,51-53
ielding the thin, asymmetrically growth-etarded fetus. In contrast to hypoxic preg-ancy, the physiology underlying theromotion of this fetal phenotype in un-ernourished pregnancy is less certain, al-hough there are increasing reports sug-esting that hypoglycemia may also be aowerful stimulus eliciting redistributionf blood flow in the fetus. For instance, an
ncrease in basal femoral vascular resis-ance has been reported in both hypogly-emic54 and undernourished55 ovine preg-ancies. In addition, it is known that thearotid body is sensitive to hypoglyce-ia,56,57 and 2 recent studies by the same
roup have postulated a possible role for aarotid body-mediated reflex in the he-odynamic response to undernutrition
nd/or hypoglycemia in fetal sheep.58,59
It is also becoming increasingly clearhat changes in maternal body composi-ion and diet not only affect fetal growthnd development as a result of direct ef-
FIGURE 3Fetal cardiac morphometry
istograms are mean � SEM for the volumes ofge of total heart volume in control, hypoxic, and u05 vs control. Number sign indicates P � .05 vontrol: n � 7; hypoxia: n � 7; undernutrition:
V, left ventricle; RV, right ventricle.
amm. Fetal cardiovascular morphology and function follow
ects on substrate availability to the fetus s
ut also indirectly through changes inlacental growth and function. Maternalndernutrition in experimental animalsnd human pregnancy has been associ-ted with reduced placental size and re-uced blood flow in the uterine and um-ilical circulations.10-12,60 Therefore, it isuite possible that a component of theeduced fetal growth in undernourishedregnancy in humans and animals is dueo a reduction in oxygen as well as nutri-nt delivery to the fetus.
Data in the present study show thatypoxic but not undernourished preg-ancy led to aortic wall thickening in the
etal offspring. Aortic thickening also oc-urs in chick embryos incubated underhe hypobaric hypoxia of high altitude29
r under isobaric hypoxia,22 indepen-ent of any changes in maternal nutri-ion. Fetal aortic wall thickening is par-icularly relevant in the clinical settingecause arterial stiffness has been haileds a key component in the etiology of hy-ertension, atherosclerosis, and coro-ary heart disease.61
An increase in wall thickness in theorta has been proposed as the first phys-cal sign in the development of athero-
tricle walls and lumens expressed as a percent-rnourished pregnancies. Asterisk indicates P �poxic (1-way ANOVA with post-hoc Tukey test).5.
omplicated pregnancy. Am J Obstet Gynecol 2010.
clerosis.5 In addition, pulse-wave veloc- r
MONTH 2010 Ame
ty measurements of the aorta, ratherhan systolic blood pressure measure-
ent, have been reported to be a betterndicator of later cardiovascular disease,ncluding impaired coronary artery flownd left ventricular dysfunction.62 Com-ined, therefore, past and present datauggest that developmental hypoxiaather than undernutrition may be theechanism via which placental insuffi-
iency has been reported to induce aortichickening, with increased vascular stiff-ess and reduced distensibility in human
nfants in 3 separate studies.6,7,63
There is general agreement that vascu-ar remodeling of this type results fromn increase in peripheral vascular resis-ance.22,64,65 The aortic thickening maye a response to restore wall stress, as isypical of an increase in load secondaryo an increase in cardiac ejection pres-ure.22 The hemodynamic overload mayncrease protein synthesis via a plethoraf cellular and molecular pathways in-luding activation of stretch receptors,rotooncogenes and vascular growthrophic factors.66-68
In the present study, prenatal hypoxianduced aortic thickening by inducingyperplasia rather than hypertrophy, inccordance with previous reports in theiterature.69 An increase in peripheralascular resistance and subsequent in-rease in cardiac afterload are also bothlausible mediators for the increase inelative heart weight and relative ven-ricular volumes reported in fetuses ex-osed to hypoxia in this and othertudies.22,64,65
The present study also determinedhether aortic vascular function in
ddition to aortic vascular remodelingas differentially affected by develop-ental hypoxia or undernutrition. Pre-
ious studies have begun to report thatustained hypoxia or undernutritionuring pregnancy can induce region-pecific changes in vascular reactivity inhe fetus. Interestingly, Williams et al19
howed that constrictor responsivenesso PE in the carotid circulation was de-ressed yet enhanced in the femoral cir-ulation in newborn rat pups from hy-oxic pregnancies. Kim et al70 have also
vennde
s hyn �
ing c
eported that chronic hypoxic preg-
rican Journal of Obstetrics & Gynecology 1.e7

VpupP
C
Research Basic Science: Obstetrics www.AJOG.org
1
ARTICLE IN PRESS
FIGURE 4Vasoconstrictor function in fetal rats
0 50 100 1500.0
0.2
0.4
0.6
-7 -6 -50.0
0.2
0.4
0.6
0.0
0.5
1.0
1.5
A
B
C
-
alues are the mean � SEM for the concentration-response curves, the maximal response (Emax or percent Kmax), and the sensitivity (pD2 or EC50) tootassium (K�), PE, and the thromboxane mimetic agent U46619 in fetal rats from control (white circles, n � 7), hypoxic (black circles, n � 7), andndernourished pregnancies (gray circles, n � 6). Asterisk indicates P � .05 vs control. Number sign indicates P � .05 vs hypoxic (1-way ANOVA withost-hoc Tukey test).E, phenylephrine.
amm. Fetal cardiovascular morphology and function following complicated pregnancy. Am J Obstet Gynecol 2010.
.e8 American Journal of Obstetrics & Gynecology MONTH 2010

nl
otilltwvcpetr
tssaacev
atsfe(
otacnrahtnptf
v
B
V(sM
C
www.AJOG.org Basic Science: Obstetrics ResearchARTICLE IN PRESS
ancy enhances the fetal femoral vascu-ar reactivity to PE in sheep.
These findings, consistent with reportsf enhanced sympathetic innervation ofhe femoral circulation in chick embryosncubated under chronic hypoxia,71 il-ustrate how developing regional circu-ations can modify their function, aidinghe redistribution of cardiac output to-ard essential and away from peripheralascular beds during adverse prenatalonditions. In the present study, thehysiology of the aortic rings was notvaluated as an index of vascular resis-ance because it is clearly a conduit
FIGURE 5Vasodilator function in fetal rats
A
alues are the mean � SEM for the concentrationwhite circles, n � 7), hypoxic (black circles, n �ign indicates P � .05 vs hypoxic (1-way ANOVetCh, methacholine; SNP, sodium nitroprusside.
amm. Fetal cardiovascular morphology and function follow
ather than a resistance vessel. However, n
he functions of the aorta are to bufferystolic blood pressure and to use thistored energy to help propel blood flowlong the systemic circulation. Theseortic functions will be influenced byhanges in contractile and relaxant prop-rties as well as structural changes of theessel.Data in the present study show that
ortic constrictor responsiveness tohe thromboxane mimetic U46619 wasignificantly diminished in fetusesrom undernourished pregnancy, anffect that tended toward significanceP � .06) in fetuses from hypoxic preg-
sponse curves, the maximal relaxation, and the sand undernourished pregnancies (gray circles, nith post-hoc Tukey test).
omplicated pregnancy. Am J Obstet Gynecol 2010.
ancy. These data concur with reports s
MONTH 2010 Ame
f reduced constrictor responsivenesso U46619 in other vascular beds, suchs the coronary72 and the carotid73 cir-ulations in fetuses from hypoxic preg-ancy. Because vasomotor responsesepresent the balance between dilatornd constrictor influences, the en-anced dilator response to the endo-helium-independent nitric oxide do-or sodium nitroprusside may be, inart, responsible for offsetting some ofhe constrictor mechanisms in fetusesrom complicated pregnancy.
Stereological investigation of cardio-ascular morphology in the present
itivity to SNP and MetCh in fetal rats from control). Asterisk indicates P � .05 vs control. Number
-re ens7), � 6A w
ing c
tudy also revealed that developmental
rican Journal of Obstetrics & Gynecology 1.e9

uwtwmpanmsiocwgprtiemppri
sdwHhioevcdcvauphhcowrw
R1Bun
2utcg13RgG4Lg15aoaGeB6Jbc7ctg8Fdf9Act11cfl11rtr21Elc51h1DoR1ocI1Loa
1Rsi21EdAH1DorPR2Sga22gh2Hlpe2Mpc2Catb22Ci92mmN2SfiM2ocle2Ddt3Zg
Research Basic Science: Obstetrics www.AJOG.org
1
ARTICLE IN PRESS
ndernutrition caused left ventricularall thinning without alterations in aor-
ic structure in the fetus. Left ventricularall thinning and an increase in cardio-yocyte apoptosis has also been re-
orted in neonatal rats from mothers fedlow protein diet throughout preg-
ancy.74 Structural remodeling of theyocardium occurs in an attempt to re-
tore myocardial wall stress.75 Depend-ng on the cause, duration, and severityf the increase in myocardial stress, theardiomyocytes undergo hypertrophy,hereas the extracellular matrix under-oes alterations in its interstitial collagenroperties.75 If the remodeling processesults in near-normal diastolic and sys-olic function, then the stressed ventricles considered to be compensated. How-ver, should the sustained increase inyocardial wall stress exceed the com-
ensatory ability of the heart, the mor-hology and function of the heart mayeflect a state of decompensation, show-ng early signs of cardiac failure.
In conclusion, the data in the presenttudy show that both hypoxic and un-ernourished pregnancy was associatedith asymmetric fetal growth restriction.owever, pregnancy complicated by
ypoxia promoted fetal aortic thicken-ng because of cellular hyperplasia with-ut changes in cardiac volumes, whenxpressed as a percentage of total heartolume. In contrast, pregnancy compli-ated by undernutrition showed evi-ence of cardiac wall thinning withouthanges in aortic structure. Fetal aorticascular reactivity was also differentiallyffected by developmental hypoxia orndernutrition. Therefore, the data sup-ort the hypothesis tested that prenatalypoxia and prenatal undernutritionave differential effects on cardiovas-ular morphology and function in theffspring prior to birth. Elucidatinghether these cardiovascular alterations
esolve, persist, or amplify later in lifeill be the focus of future research. f
EFERENCES. Veille JC, Hanson R, Sivakoff M, Hoen H,en-Ami M. Fetal cardiac size in normal, intra-terine growth retarded, and diabetic preg-
ancies. Am J Perinatol 1993;10:275-9. A.e10 American Journal of Obstetrics & Gynecolog
. Mayhew TM, Gregson C, Fagan DG. Ventric-lar myocardium in control and growth-re-arded human fetuses: growth in different tissueompartments and variation with fetal weight,estational age, and ventricle size. Hum Pathol999;30:655-60.. Rizzo G, Capponi A, Rinaldo D, Arduini D,omanini C. Ventricular ejection force inrowth-retarded fetuses. Ultrasound Obstetynecol 1995;5:247-55.. Miyague NI, Ghidini A, Fromberg R, MiyagueL. Alterations in ventricular filling in small-for-estational-age fetuses. Fetal Diagn Ther997;12:332-5.. McGill HC Jr, McMahan CA, Herderick EE, etl. Effects of coronary heart disease risk factorsn atherosclerosis of selected regions of theorta and right coronary artery. PDAY Researchroup. Pathobiological Determinants of Ath-rosclerosis in Youth. Arterioscler Thromb Vasciol 2000;20:836-45.. Skilton MR, Evans N, Griffiths KA, HarmerA, Celermajer DS. Aortic wall thickness in new-orns with intrauterine growth restriction. Lan-et 2005;365:1484-6.. Koklu E, Kurtoglu S, Akcakus M, et al. In-reased aortic intima-media thickness is relatedo lipid profile in newborns with intrauterinerowth restriction. Horm Res 2006;65:269-75.. Leeson CP, Whincup PH, Cook DG, et al.low-mediated dilation in 9- to 11-year-old chil-ren: the influence of intrauterine and childhood
actors. Circulation 1997;96:2233-8.. Leeson CP, Kattenhorn M, Morley R, Lucas, Deanfield JE. Impact of low birth weight andardiovascular risk factors on endothelial func-ion in early adult life. Circulation 2001;103:264-8.0. Morriss FH, Rosenfeld CR, Crandell SS, Ad-ock EW 3rd. Effects of fasting on uterine bloodow and substrate uptake in sheep. J Nutr980;110:2433-43.1. Gilbert M, Leturque A. Fetal weight and itselationship to placental blood flow and placen-al weight in experimental intrauterine growthetardation in the rat. J Dev Physiol 1982;4:37-46.2. Chandler KD, Leury BJ, Bird AR, Bell AW.ffects of undernutrition and exercise during
ate pregnancy on uterine, fetal and uteropla-ental metabolism in the ewe. Br J Nutr 1985;3:625-35.3. Moore LG. Human genetic adaptation toigh altitude. High Alt Med Biol 2001;2:257-79.4. Giussani DA, Phillips PS, Anstee S, BarkerJ. Effects of altitude versus economic statusn birth weight and body shape at birth. Pediatres 2001;49:490-4.5. Xiao D, Ducsay CA, Zhang L. Chronic hyp-xia and developmental regulation of cyto-hrome c expression in rats. J Soc Gynecol
nvestig 2000;7:279-83.6. Peyronnet J, Dalmaz Y, Ehrstrom M, et al.ong-lasting adverse effects of prenatal hyp-xia on developing autonomic nervous systemnd cardiovascular parameters in rats. Pflugers
rch 2002;443:858-65. Ay MONTH 2010
7. Li G, Xiao Y, Estrella JL, Ducsay CA, GilbertD, Zhang L. Effect of fetal hypoxia on heartusceptibility to ischemia and reperfusion injury
n the adult rat. J Soc Gynecol Investig003;10:265-74.8. Bae S, Xiao Y, Li G, Casiano CA, Zhang L.ffect of maternal chronic hypoxic exposureuring gestation on apoptosis in fetal rat heart.m J Physiol Heart Circ Physiol 2003;285:983-90.9. Williams SJ, Campbell ME, McMillen IC,avidge ST. Differential effects of maternal hyp-xia or nutrient restriction on carotid and femo-al vascular function in neonatal rats. Am Jhysiol Regul Integr Comp Physiol 2005;288:360-7.0. Rueda-Clausen CF, Morton JS, DavidgeT. Effects of hypoxia-induced intrauterinerowth restriction on cardiopulmonary structurend function during adulthood. Cardiovasc Res009;81:713-22.1. de Grauw TJ, Myers RE, Scott WJ. Fetalrowth retardation in rats from different levels ofypoxia. Biol Neonate 1986;49:85-9.2. Rouwet EV, Tintu AN, Schellings MW, et al.ypoxia induces aortic hypertrophic growth,
eft ventricular dysfunction, and sympathetic hy-erinnervation of peripheral arteries in the chickmbryo. Circulation 2002;105:2791-6.3. Miller SL, Green LR, Peebles DM, HansonA, Blanco CE. Effects of chronic hypoxia androtein malnutrition on growth in the developinghick. Am J Obstet Gynecol 2002;186:261-7.4. Villamor E, Kessels CG, Ruijtenbeek K, et al.hronic in ovo hypoxia decreases pulmonaryrterial contractile reactivity and induces biven-ricular cardiac enlargement in the chicken em-ryo. Am J Physiol Regul Integr Comp Physiol004;287:R642-51.5. Giussani DA, Salinas CE, Villena M, BlancoE. The role of oxygen in prenatal growth: stud-
es in the chick embryo. J Physiol 2007;585:11-7.6. Mortola JP, Cooney E. Cost of growth andaintenance in chicken embryos during nor-oxic or hypoxic conditions. Respir Physioleurobiol 2008;162:223-9.7. Ghatpande SK, Billington CJ Jr, RivkeesA, Wendler CC. Hypoxia induces cardiac mal-
ormations via A1 adenosine receptor activationn chicken embryos. Birth Defects Res A Clin
ol Teratol 2008;82:121-30.8. Tintu A, Rouwet E, Verlohren S, et al. Hyp-xia induces dilated cardiomyopathy in thehick embryo: mechanism, intervention, and
ong-term consequences. PLoS One 2009;4:5155.9. Salinas CE, Blanco CE Jr, Villena M, et al.evelopmental origin of cardiac and vascularisease in chick embryos incubated at high at-itude. J DOHaD 2010;1:60-6.0. Van Geijn HP, Kaylor WM Jr, Nicola KR,uspan FP. Induction of severe intrauterinerowth retardation in the Sprague-Dawley rat.
m J Obstet Gynecol 1980;137:43-7.
3th3Sma3tf13esr3Ka23ml13Ert23Cfao3ssp4naa24eaRs14p24t14d4Aah54npt1
4Aapt4BotaJ4e15Fsdn5deG5kgA5Lvpfl5Fiubo5Crstp5taN5tsr5cmt5lht56Mso
6sE6Wwdi16fa16Gac16Cl16nfpaS6Ptmd6AlM6Hgv7hfS7avP7lo27hip27tmd7Tl
www.AJOG.org Basic Science: Obstetrics ResearchARTICLE IN PRESS
1. Burton GJ, Palmer ME. Eradicating fetoma-ernal fluid shift during perfusion fixation of theuman placenta. Placenta 1988;9:327-32.2. Gundersen HJ, Bendtsen TF, Korbo L, et al.ome new, simple and efficient stereologicalethods and their use in pathological research
nd diagnosis. APMIS 1988;96:379-94.3. Jensen EB, Gundersen HJ, Osterby R. De-ermination of membrane thickness distributionrom orthogonal intercepts. J Microsc 1979;15:19-33.4. Mulvany MJ, Halpern W. Contractile prop-rties of small arterial resistance vessels inpontaneously hypertensive and normotensiveats. Circ Res 1977;41:19-26.5. Unger C, Weiser JK, McCullough RE,eefer S, Moore LG. Altitude, low birth weight,nd infant mortality in Colorado. JAMA 1988;59:3427-32.6. Moore LG, Niermeyer S, Zamudio S. Hu-an adaptation to high altitude: regional and
ife-cycle perspectives. Am J Phys Anthropol998;Suppl 27:25-64.7. Keyes LE, Armaza JF, Niermeyer S, Vargas, Young DA, Moore LG. Intrauterine growthestriction, preeclampsia, and intrauterine mor-ality at high altitude in Bolivia. Pediatr Res003;54:20-5.8. Holemans K, Gerber R, Meurrens K, Delerck F, Poston L, Van Assche FA. Maternal
ood restriction in the second half of pregnancyffects vascular function but not blood pressuref rat female offspring. Br J Nutr 1999;81:73-9.9. Kind KL, Simonetta G, Clifton PM, Robin-on JS, Owens JA. Effect of maternal feed re-triction on blood pressure in the adult guineaig. Exp Physiol 2002;87:469-77.0. Kwon H, Ford SP, Bazer FW, et al. Maternalutrient restriction reduces concentrations ofmino acids and polyamines in ovine maternalnd fetal plasma and fetal fluids. Biol Reprod004;71:901-8.1. Neumann CG, Harrison GG. Onset andvolution of stunting in infants and children. Ex-mples from the Human Nutrition Collaborativeesearch Support Program. Kenya and Egypttudies. Eur J Clin Nutr 1994;48(Suppl 1):S90-02.2. Gruenwald P. Chronic fetal distress andlacental insufficiency. Biol Neonat 1963;5:15-65.3. Wigglesworth JS. Experimental growth re-ardation in the foetal rat. J Pathol Bacteriol964;88:1-13.4. Barker DJ. In utero programming of chronicisease. Clin Sci (Lond) 1998;95:115-28.5. Bartelds B, van Bel F, Teitel DF, RudolphM. Carotid, not aortic, chemoreceptors medi-te the fetal cardiovascular response to acuteypoxemia in lambs. Pediatr Res 1993;34:1-5.6. Giussani DA, Spencer JA, Moore PJ, Ben-et L, Hanson MA. Afferent and efferent com-onents of the cardiovascular reflex responseso acute hypoxia in term fetal sheep. J Physiol
993;461:431-49. 27. Giussani DA, Gardner DS, Cox DT, FletcherJ. Purinergic contribution to circulatory, met-bolic, and adrenergic responses to acute hy-oxemia in fetal sheep. Am J Physiol Regul In-egr Comp Physiol 2001;280:R678-85.8. Morrison S, Gardner DS, Fletcher AJ,loomfield MR, Giussani DA. Enhanced nitricxide activity offsets peripheral vasoconstric-ion during acute hypoxaemia via chemoreflexnd adrenomedullary actions in the sheep fetus.Physiol 2003;547:283-91.9. Jones CT, Robinson RO. Plasma cat-cholamines in foetal and adult sheep. J Physiol975;248:15-33.0. Fletchert AJ, Edwards CM, Gardner DS,owden AL, Giussani DA. Neuropeptide Y in theheep fetus: effects of acute hypoxemia andexamethasone during late gestation. Endocri-ology 2000;141:3976-82.1. Cohn HE, Sacks EJ, Heymann MA, Ru-olph AM. Cardiovascular responses to hypox-mia and acidemia in fetal lambs. Am J Obstetynecol 1974;120:817-24.2. Peeters LL, Sheldon RE, Jones MD Jr, Ma-owski EL, Meschia G. Blood flow to fetal or-ans as a function of arterial oxygen content.m J Obstet Gynecol 1979;135:637-46.3. Giussani DA, Riquelme RA, Hanson MA,lanos AJ. Is the rapid and intense peripheralasoconstriction occurring during acute hy-oxaemia in the llama fetus an arterial chemore-ex? Adv Exp Med Biol 1994;360:341-4.4. Gardner DS, Fletcher AJ, Bloomfield MR,owden AL, Giussani DA. Effects of prevail-
ng hypoxaemia, acidaemia or hypoglycaemiapon the cardiovascular, endocrine and meta-olic responses to acute hypoxaemia in thevine fetus. J Physiol 2002;540:351-66.5. Hawkins P, Steyn C, McGarrigle HH, et al.ardiovascular and hypothalamic-pituitary-ad-
enal axis development in late gestation fetalheep and young lambs following modest ma-ernal nutrient restriction in early gestation. Re-rod Fertil Dev 2000;12:443-56.6. Pardal R, Lopez-Barneo J. Carotid bodyhin slices: responses of glomus cells to hypoxiand K(�)-channel blockers. Respir Physioleurobiol 2002;132:69-79.7. Zhang M, Buttigieg J, Nurse CA. Neuro-ransmitter mechanisms mediating low-glucoseignalling in cocultures and fresh tissue slices ofat carotid body. J Physiol 2007;578:735-50.8. Burrage D, Green LR, Moss TJ, et al. Thearotid bodies influence growth responses tooderate maternal undernutrition in late-gesta-
ion fetal sheep. BJOG 2008;115:261-8.9. Burrage DM, Braddick L, Cleal JK, et al. The
ate gestation fetal cardiovascular response toypoglycaemia is modified by prior peri-implan-ation undernutrition in sheep. J Physiol 2009;87:611-24.0. Roberts CT, Sohlstrom A, Kind KL, et al.aternal food restriction reduces the exchange
urface area and increases the barrier thicknessf the placenta in the guinea-pig. Placenta
001;22:177-85. EMONTH 2010 Ameri
1. Arnett DK, Evans GW, Riley WA. Arterialtiffness: a new cardiovascular risk factor? Am Jpidemiol 1994;140:669-82.2. Cruickshank K, Riste L, Anderson SG,right JS, Dunn G, Gosling RG. Aortic pulse-ave velocity and its relationship to mortality iniabetes and glucose intolerance: an integrated
ndex of vascular function? Circulation 2002;06:2085-90.3. Akira M, Yoshiyuki S. Placental circulation,
etal growth, and stiffness of the abdominalorta in newborn infants. J Pediatr 2006;48:49-53.4. Murotsuki J, Challis JR, Han VK, Fraher LJ,agnon R. Chronic fetal placental embolizationnd hypoxemia cause hypertension and myo-ardial hypertrophy in fetal sheep. Am J Physiol997;272:R201-7.5. Kitanaka T, Alonso JG, Gilbert RD, Siu BL,lemons GK, Longo LD. Fetal responses to
ong-term hypoxemia in sheep. Am J Physiol989;256:R1348-54.6. Anderson PAW, Kleinman CS, Lister G, Tal-er NS. Cardiovascular function during normaletal and neonatal development and with hy-oxic stress. In: Polin RA, Fox WW, eds. Fetalnd neonatal physiology. Philadelphia: W.B.aunders; 1998.7. Kempf H, Linares C, Corvol P, Gasc JM.harmacological inactivation of the endothelin
ype A receptor in the early chick embryo: aodel of mispatterning of the branchial archerivatives. Development 1998;125:4931-41.8. Byrne AM, Bouchier-Hayes DJ, Harmey JH.ngiogenic and cell survival functions of vascu-
ar endothelial growth factor (VEGF). J Cell Moled 2005;9:777-94.9. Kourembanas S, Morita T, Liu Y, Christou. Mechanisms by which oxygen regulatesene expression and cell-cell interaction in theasculature. Kidney Int 1997;51:438-43.0. Kim YH, Veille JC, Cho MK, et al. Chronicypoxia alters vasoconstrictive responses ofemoral artery in the fetal sheep. J Korean Medci 2005;20:13-9.1. Ruijtenbeek K, Kessels CG, Janssen BJ, etl. Chronic moderate hypoxia during in ovo de-elopment alters arterial reactivity in chickens.flugers Arch 2003;447:158-67.2. Garcia FC, Stiffel VM, Gilbert RD. Effects of
ong-term high-altitude hypoxia on isolated fetalvine coronary arteries. J Soc Gynecol Investig000;7:211-7.3. Thompson LP, Aguan K, Zhou H. Chronicypoxia inhibits contraction of fetal arteries by
ncreased endothelium-derived nitric oxide androstaglandin synthesis. J Soc Gynecol Investig004;11:511-20.4. Cheema KK, Dent MR, Saini HK, Arou-iounova N, Tappia PS. Prenatal exposure toaternal undernutrition induces adult cardiacysfunction. Br J Nutr 2005;93:471-7.5. Brower GL, Gardner JD, Forman MF, et al.he relationship between myocardial extracel-
ular matrix remodeling and ventricular function.
ur J Cardiothorac Surg 2006;30:604-10.can Journal of Obstetrics & Gynecology 1.e11
Copyright © 2022 FDOKUMEN




















