
Paleontology pnd?Stratigra:phy - USGS Publications Repository
Upload
bryanskhistorfacultyCategory
view
2download
0
114
УДК903.4/569*56.012.2
ПАЛЕОНТОЛОГИЯ, АРХЕОЗООЛОГИЯ И ТАФОНОМИЯ
ВЕРХНЕПАЛЕОЛИТИЧЕСКОГО ПОСЕЛЕНИЯ ХОТЫЛЕВО 2
(по материалам раскопок 1994-2003 гг.)
А. А. Чубур
Брянский государственный университет им. академика И.Г. Петровского
241036 г.Брянск, ул. Бежицкая, 14 [email protected]
Остеологический материал из раскопок 1994-2003 гг. насчитывает свыше 1700 костей млекопитающих. Определен видовой и анатомический состав, изучены природные механические повреждения, химические изменения под влиянием почвообразования, погрызы, следы работы человека (расчленение туш, питание, добывание костного мозга, использование костей в конструкциях). Основной вывод: раскопами вскрыты следы охотничьего лагеря теплого сезона.
Ключевые слова: зооархеология, палеолит, Хотылево 2, мамонт, мамонтовая фауна
В последнее время возрос интерес к проблемам археозоологии и тафономии памятников палеолита, связанный с процессом смены парадигм в отечественном палеолитоведении. Ископаемые костные остатки, наряду с каменной индустрией, – основная часть коллекций, собираемых в процессе раскопок. На базе их анализа формируются представления, как о фауне прошлого, так и о характере взаимодействия животного мира и первобытного человека. Видовой состав позволяет судить об окружающем ландшафте, показывает промысловых животных, биостратиграфическое положение поселения. Изучение костей млекопитающих даёт сведения о палеоэкологии человека (характер и сезонность пребывания на стоянке, пищевая стратегия, приемы охоты и разделки добычи, взаимоотношения с хищниками и др.). Тафономический анализ позволяет узнать о процессе седиментации и последующих трансформациях культурного слоя.
Объект исследования – верхнепалеолитическое поселение Хотылево 2, открытое в 1968 г. Ф.М. Заверняевым. Он производил раскопки 10 сезонов, вскрыв 500 м
2 В 1990 г. памятник был
обследован автором, а с 1993 г. раскопки продолжил К.Н. Гаврилов, они ведутся им уже свыше 10 сезонов. Культурный слой залегает в слабо гумусированном горизонте, находящемся в основании пачки поздневалдайских лессовидных супесей. Его радиоуглеродная датировка находится в пределах 22-24 тыс. лет. Каменный и костяной инвентарь позволяют относить поселение к кругу памятников восточного граветта с узкой пластинчатой заготовкой, сближая с Центрально-Европейским павловьеном с одной стороны и с восточноевропейским поселением Гагарино с другой (Гаврилов, 1997; Амирханов, 1998; Чубур, 2004).
Материал и методика Остеологический материал из культурного слоя Хотылево 2 (раскопки 1994-2003 гг.)
насчитывает свыше 1700 костей млекопитающих крупного и среднего размера (табл.1) (кости грызунов не учитывались), а также обломок 1 лучевой кости небольшой птицы. Материал был разделен по вертикали (нивелировочные отметки и микростратиграфия) и по горизонтали (квадраты раскопа). После видового определения для каждого вида выявлено число определимых костных остатков и минимальное число особей. С учетом степени стирания зубов и степени срастания швов черепа и эпифизов с диафизами определен индивидуальный возраст животных. Выполнена стандартная морфометрия костных остатков. Костные остатки могли попадать в культурные слои разным путем: в результате охотничьей и собирательской деятельности человека, естественной гибели животных, пищевой активности хищников, заноса водными потоками. Поэтому важно проследить тафономию: степень выветренности костного материала (это показывает, как долго находился он на дневной поверхности), его фрагментарность и характер повреждений на поверхности костей – погрызы хищников и следы деятельности человека (следы огня, порезы, обработка). Степень выветренности поверхности костей определялась по схеме, ранее использованной в Юдиново (Бурова, 2002). Нами изучены пространственное распределение и набор скелетных элементов крупных млекопитающих, основные типы их повреждения, образовавшиеся в результате хозяйственно-бытовой деятельности древнего человека и способы использования им костей. По ряду признаков мы попытались установить сезонность охоты обитателей вскрытого участка Хотылевской стоянки.
Систематика и морфология Видовой состав фауны раскопок 1994-2003 гг. несколько беднее, чем в раскопах Ф.М.
Заверняева (Величко, Грибченко, Маркова, Ударцев, 1977; Soffer, 1985; Чубур, 1995, 2003; Чубур,

115
Тимошкин, Головачева, 1997). Так, нет здесь донского зайца, лисички-корсака, бурого медведя, шерстистого носорога. Впрочем, они весьма немногочисленны и в раскопах 1968-1981-х годов, в процессе которых были исследованы более насыщенные участки культурного слоя. Далее мы приводим описание костных остатков млекопитающих в соответствии с принятой систематикой.
Отряд ХИЩНИКИ (Carnivora), семейство СОБАЧЬИ (Canidae)
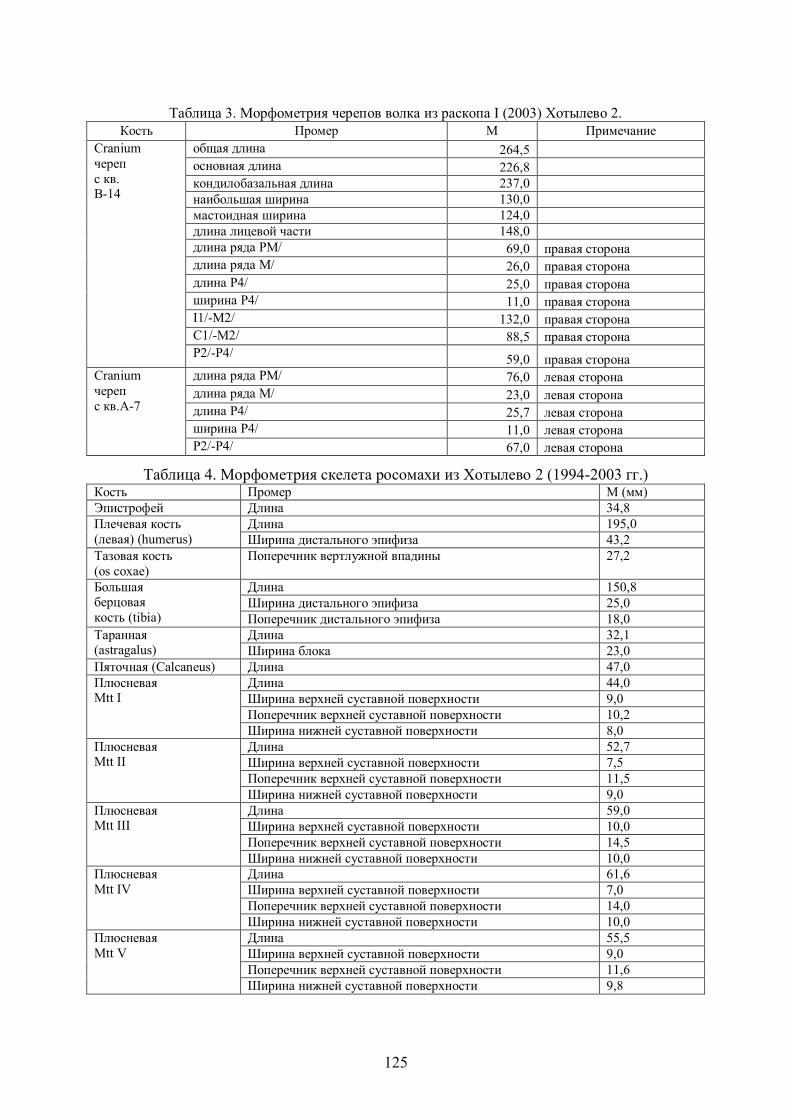
ВОЛК (Canis lupus L. 1788). Материал: 80 костей минимум от 4 особей – череп и 36 фрагментов другого черепа, зубы (Р4/, по два Р2/, Р1/ и С1/), позвонок, 4 фрагмента ребер, 6 плечевых, 5 локтевых, 3 лучевых, бедренная, 2 больших берцовых, малая берцовая, обломок метаподия, пястные кости (Mtc2: длина 80,5 мм, ширина проксимальная 9 мм; Mtc3: длина 90,5 мм, ширина проксимальная 11,9 мм; Mtc4: длина 89,5 мм, ширина проксимальная 10,5 мм; Mtc5: длина 77,7 мм, ширина проксимальная 16 мм) в анатомической связи с 5 костями запястья и 3 фалангами I. Основные промеры даны в табл.2 и 3. Сравнение: Хотылевский волк, с учетом материалов Ф.М. Заверняева, занимает по показателям зубной системы и морфологии посткраниального скелета промежуточное положение между костенковскими волками из верхней гумусированной толщи (32-25 тыс.л.н.) и лессовидных суглинков Костенок (23-19 тыс.л.н.), опубликованными И.Е. Кузьминой и М.В. Саблиным (1994). Волк Хотылево 2 крупнее рецентного из зон тундры и тайги. Морфологических признаков доместикации у него не наблюдается.
ПЕСЕЦ (Alopex lagopus rossicus Sablin et Kuzmina, 1992) Материал: Из раскопа 12-14 (Р-14) происходят 12 костей песца, принадлежащих 1 особи – правая альвеоляра нижней челюсти, 4 позвонка, две лучевых, плечевая, большая берцовая, пяточная, таранная и метаподий. Из раскопа I (далее – Р-I) происходят 9 костей от 1 особи – 2 зуба, правая альвеоляра нижней челюсти, 2 позвонка грудного отдела, локтевая, лучевая, метаподий и ребро. Морфометрические показатели удалось снять с локтевой кости (поперечник в полулунной ямке 7,2 мм, высота 11,8 мм) и с зубного ряда нижней челюсти (С/1-М/3 - 55 мм, Р/1-М/2 - 50,5 мм М/1 - 13,2 х 5 мм). Сравнение: Зубная система песца из Хотылево 2 близка к таковой из Юдиново, Костенок, Быков (Кузьмина, Саблин, 1993; Саблин, 1994; Чубур, 2001) и отличается от рецентной популяции меньшей относительной шириной хищнических зубов и большей относительной длиной зубного ряда. Малый объем материала не позволяет делать далеко идущих сопоставлений.
Семейство КУНЬИ РОСОМАХА (Gulo gulo L., 1788)
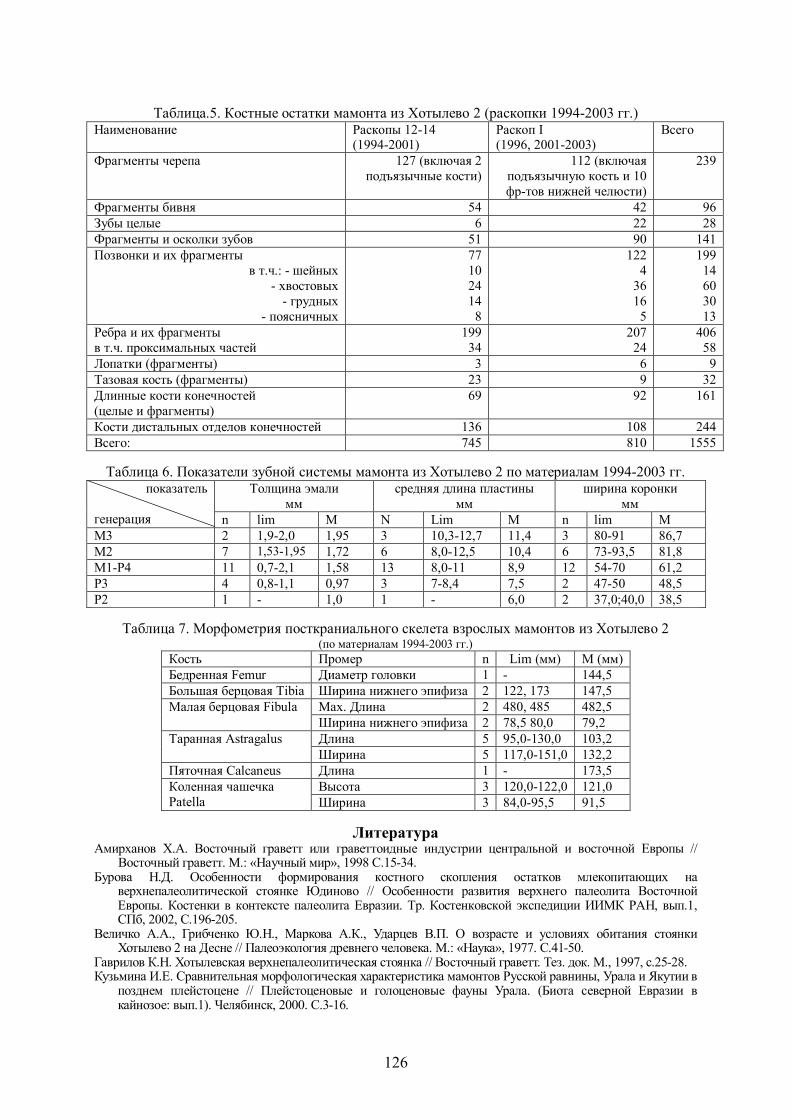
Материал и сравнение: 11 фрагментов черепа, 13 позвонков (включая атлант, эпистрофей, 1 шейный, 1 хвостовой, 4 грудных и 5 поясничных), 3 фрагмента ребер, 4 фрагмента тазовой кости, 2 плечевых, 2 бедренных кости (одна сильно разрушена), большая берцовая, малая берцовая, астрагал, кость предплюсны, 3 метаподия, 12 фаланг и 25 костей в анатомической группе дистальной части задней конечности (5 плюсневых, 9 фаланг, пяточная, астрагал, кости предплюсны, сессамовидные, большая и малая берцовые) – всего 77 костей от 1 особи. Морфометрия костей приведена в табл.4. Сравнительный материал по ископаемой росомахе Русской равнины не опубликован. Росомаха из Хотылево 2, с учетом данных по коллекции из раскопок Ф.М. Заверняева (Чубур, Головачева, Тимошкин, 1997) на 13-19% крупнее рецентной.
Отряд ARTIODACTYLA - ПАРНОКОПЫТНЫЕ Семейство BOVIDAE - ПОЛОРОГИЕ
Бизон плейстоценовый (Bison priscus Boj, ) Материал: бедренная кость (Р-12), а также несколько ребер и позвонков из Р-13 и найденных на границе с Р-8 и 11 Ф.М.Заверняева, в которых также отмечались малочисленные остатки бизона. Сравнение: Объекты непригодны для морфометрии, однако, учитывая данные, полученные нами коллекции из раскопок Ф.М. Заверняева (Чубур, Головачева, Тимошкин, 1997), можно полагать, что остатки принадлежат мелкой адаптации позднеплейстоценового бизона.
Семейство CERVIDAE - ОЛЕНЬИ Северный олень (Rangifer tarandus L. 1758)
Материал: В Р-13 – единственный обломок рога с нарезкой, в Р-I вид представлен 9 костями (3 фрагмента рога, 4 фрагмента черепа, обломок ребра и большая берцовая кость без эпифизов) от одной молодой особи.
116
Отряд PROBOSCIDEA - ХОБОТНЫЕ Семейство ELEPHANTIDAE - CЛОНЫ
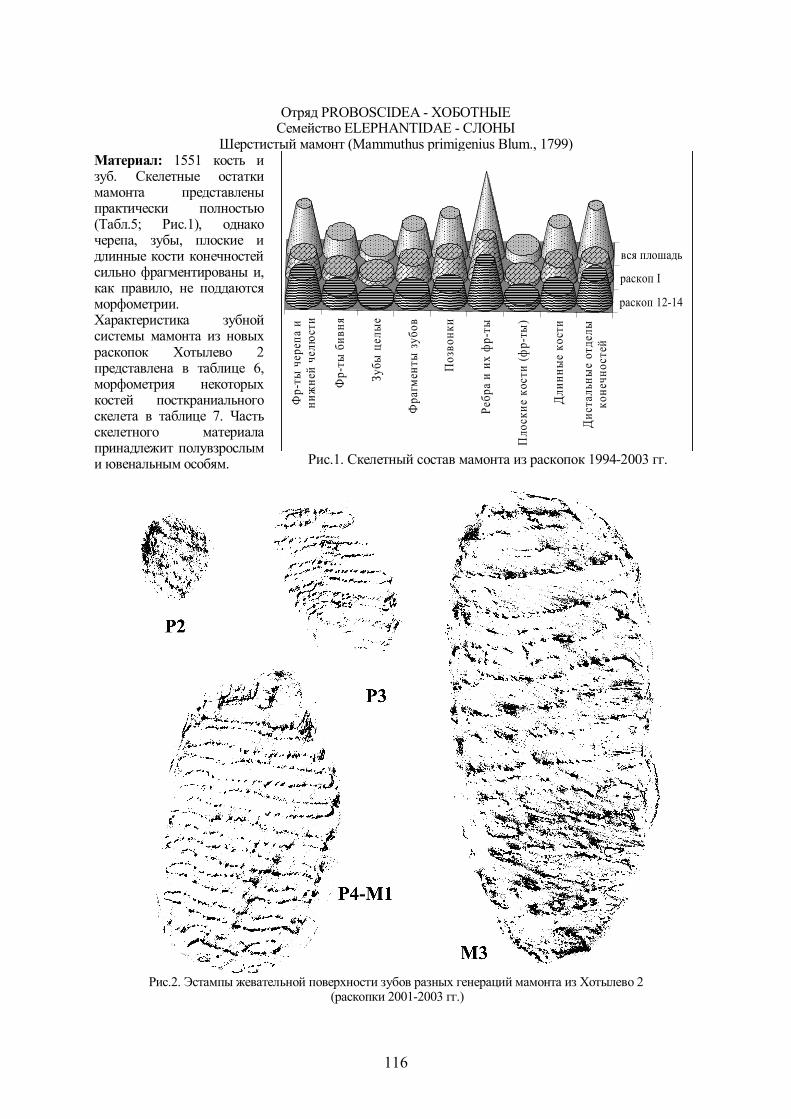
Шерстистый мамонт (Mammuthus primigenius Blum., 1799) Материал: 1551 кость и зуб. Скелетные остатки мамонта представлены практически полностью (Табл.5; Рис.1), однако черепа, зубы, плоские и длинные кости конечностей сильно фрагментированы и, как правило, не поддаются морфометрии. Характеристика зубной системы мамонта из новых раскопок Хотылево 2 представлена в таблице 6, морфометрия некоторых костей посткраниального скелета в таблице 7. Часть скелетного материала принадлежит полувзрослым и ювенальным особям.
Фр
-ты
чер
епа
ин
иж
ней
чел
юст
и
Фр
-ты
би
вн
я
Зу
бы
цел
ые
Фр
агм
енты
зу
бо
в
По
зво
нк
и
Реб
ра
и и
х ф
р-т
ы
Пл
оск
ие
ко
сти
(ф
р-т
ы)
Дл
ин
ны
е к
ост
и
Ди
стал
ьны
е о
тдел
ык
он
ечн
ост
ей
раскоп 12-14
раскоп I
вся плошадь
Рис.1. Скелетный состав мамонта из раскопок 1994-2003 гг.
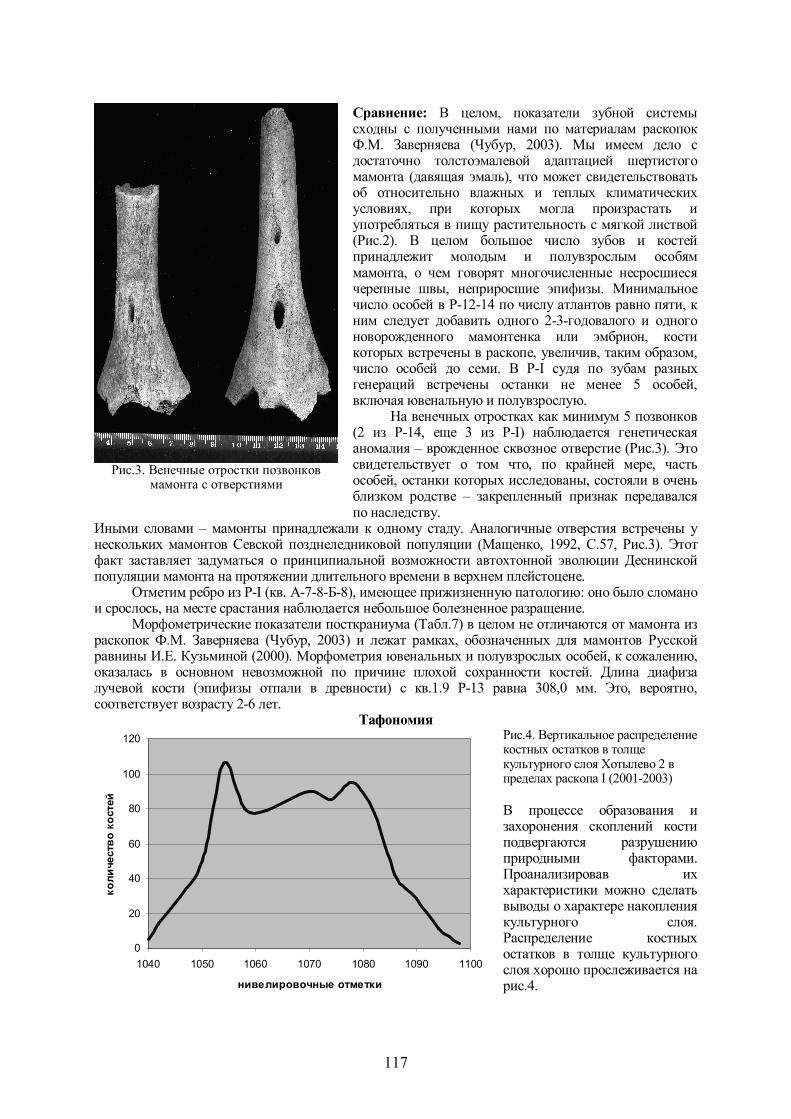
Рис.2. Эстампы жевательной поверхности зубов разных генераций мамонта из Хотылево 2
(раскопки 2001-2003 гг.)
117
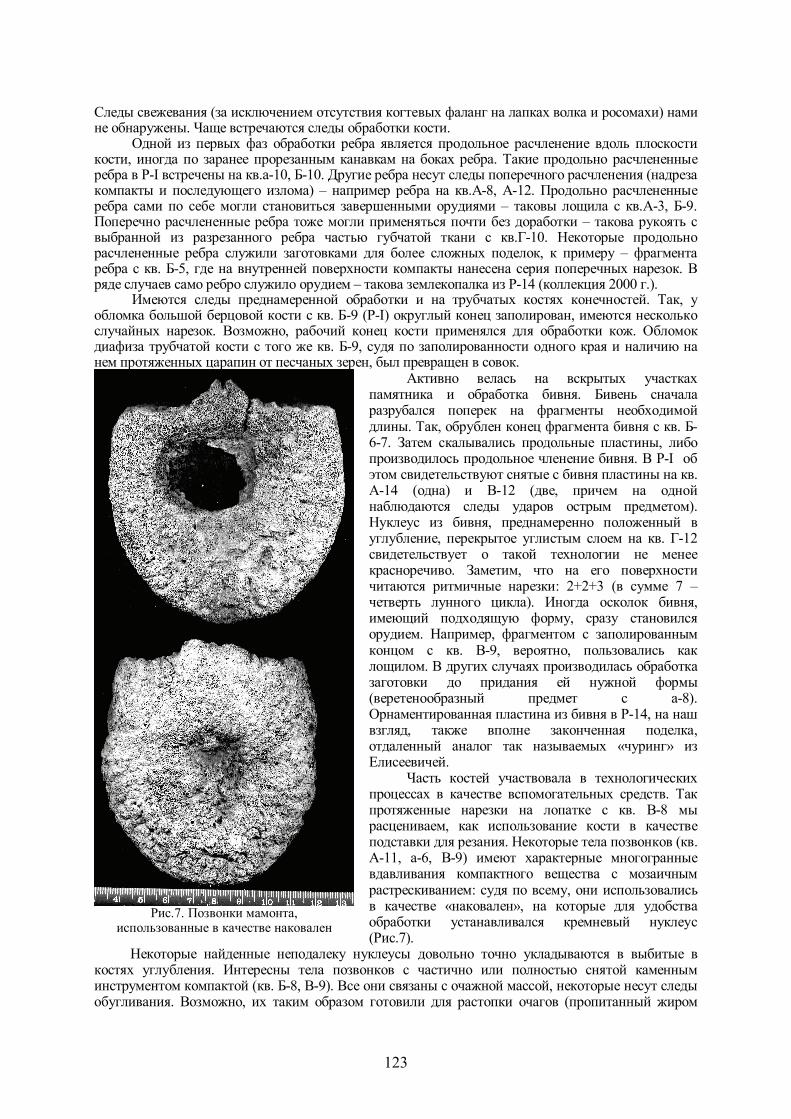
Рис.3. Венечные отростки позвонков
мамонта с отверстиями
Сравнение: В целом, показатели зубной системы сходны с полученными нами по материалам раскопок Ф.М. Заверняева (Чубур, 2003). Мы имеем дело с достаточно толстоэмалевой адаптацией шертистого мамонта (давящая эмаль), что может свидетельствовать об относительно влажных и теплых климатических условиях, при которых могла произрастать и употребляться в пищу растительность с мягкой листвой (Рис.2). В целом большое число зубов и костей принадлежит молодым и полувзрослым особям мамонта, о чем говорят многочисленные несросшиеся черепные швы, неприросшие эпифизы. Минимальное число особей в Р-12-14 по числу атлантов равно пяти, к ним следует добавить одного 2-3-годовалого и одного новорожденного мамонтенка или эмбрион, кости которых встречены в раскопе, увеличив, таким образом, число особей до семи. В Р-I судя по зубам разных генераций встречены останки не менее 5 особей, включая ювенальную и полувзрослую.
На венечных отростках как минимум 5 позвонков (2 из Р-14, еще 3 из Р-I) наблюдается генетическая аномалия – врожденное сквозное отверстие (Рис.3). Это свидетельствует о том что, по крайней мере, часть особей, останки которых исследованы, состояли в очень близком родстве – закрепленный признак передавался по наследству.
Иными словами – мамонты принадлежали к одному стаду. Аналогичные отверстия встречены у нескольких мамонтов Севской позднеледниковой популяции (Мащенко, 1992, С.57, Рис.3). Этот факт заставляет задуматься о принципиальной возможности автохтонной эволюции Деснинской популяции мамонта на протяжении длительного времени в верхнем плейстоцене.
Отметим ребро из Р-I (кв. А-7-8-Б-8), имеющее прижизненную патологию: оно было сломано и срослось, на месте срастания наблюдается небольшое болезненное разращение.
Морфометрические показатели посткраниума (Табл.7) в целом не отличаются от мамонта из раскопок Ф.М. Заверняева (Чубур, 2003) и лежат рамках, обозначенных для мамонтов Русской равнины И.Е. Кузьминой (2000). Морфометрия ювенальных и полувзрослых особей, к сожалению, оказалась в основном невозможной по причине плохой сохранности костей. Длина диафиза лучевой кости (эпифизы отпали в древности) с кв.1.9 Р-13 равна 308,0 мм. Это, вероятно, соответствует возрасту 2-6 лет.
Тафономия
0
20
40
60
80
100
120
1040 1050 1060 1070 1080 1090 1100
нивелировочные отметки
ко
ли
че
ств
о к
остей
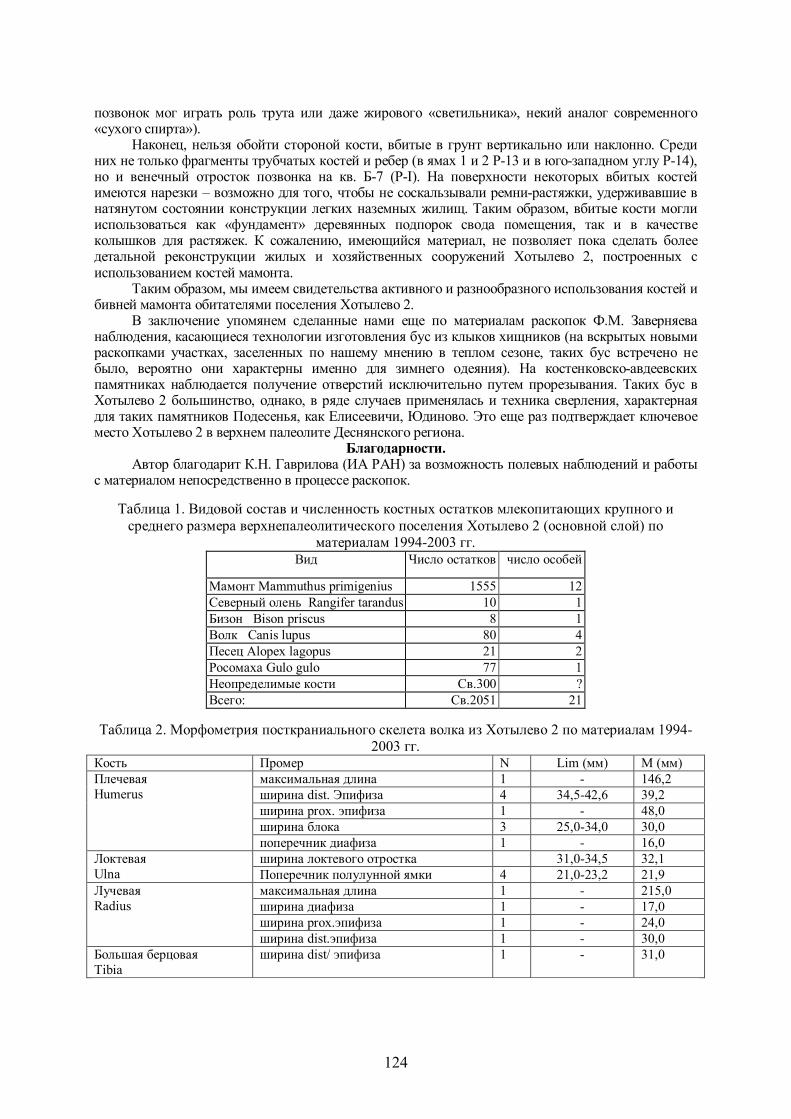
Рис.4. Вертикальное распределение костных остатков в толще культурного слоя Хотылево 2 в пределах раскопа I (2001-2003)
В процессе образования и захоронения скоплений кости подвергаются разрушению природными факторами. Проанализировав их характеристики можно сделать выводы о характере накопления культурного слоя. Распределение костных остатков в толще культурного слоя хорошо прослеживается на рис.4.

118
Cудя по построенной нами кривой, скорость накопления костных остатков мало менялась в процессе формирования культурного слоя, усиление процесса связано с горизонтами, лежащими у отметок -1055 и -1080. По-видимому, они связаны с активизацией жизни на данном участке поселения на определенных этапах его существования – в начале существования и ближе к его концу. Ярче эти уровни прослеживаются при анализе вертикального распределения осколков трубчатых костей и костей дистальных отделов конечностей мамонта.
Цвет. Кости в основном светло-бежевого оттенка. Вместе с тем, независимо от видовой принадлежности, имеются немногочисленные костные остатки темно-серого и коричневого цветов, а также единичные элементы с локально расположенными на поверхности черными пятнами замарганцованности и рыжими пятнами и разводами ожелезнения. Это свидетельствует о накоплении костных остатков в субаэральных условиях, в ряде случаев на них отразились процессы почвообразования и переувлажнения грунта стоячими водами.
Выветривание. Одной из важнейших характеристик сохранности является интенсивность выветривания остеологического материала. Н.Д. Буровой (2002) при работе с материалами Юдиново были выделены стадии климатического разрушения костных остатков, которые применимы и для Хотылево 2: 0 – нет изменений; 0/1-1 – наблюдается слабое растрескивание поверхности (низкая степень выветривания); 1 – появление продольных трещин, 1/2 – незначительное расслаивание верхнего слоя костей. По нашим наблюдениям, наиболее высокой степенью выветренности (1–1/2) отличаются кости, лежащие на поверхности культурного слоя (причем выветривание наблюдается только обращенной кверху стороны кости). Это не более 3% остеологического материала (преимущественно неопределимые кости – мелкие фрагменты, ребра и некоторые фрагменты бивня). Основная часть материала не несет на себе изменений такого рода. Из этого следует, что пребывание костных скоплений на субаэральной поверхности было непродолжительным, в результате сравнительно быстрого осадконакопления в ходе активных склоновых процессов.
Повреждения костей. Среди поверхностных природных повреждений костей имеют место признаки механического воздействия, химические видоизменения и биоповреждения. В ископаемом материале доминируют кости с неповрежденной поверхностью.
У ископаемых костей из Хотылево 2 природные механические повреждения редки. Наиболее часто они связаны с криогенным фактором. Отдельные кости разорваны и смещены трещинами мелкополигональной сетки криодеформаций, что говорит об усилении криоаридных условий после завершения седиментации культурного слоя. В качестве примера приведем разорванные мерзлотными трещинами ребра на кв.6.2, скуловую дугу молодого мамонта на кв.3.2 Р-13. Плечевая кость росомахи не только разорвана, но и разнесена на соседние кв. 1.3 и 2.3.
Химические видоизменения связаны в основном с действием корней растений, росших на культурном слое после его седиментации. Они встречены на нижних поверхностях некоторых костей, лежавших у верхнего контакта культурного слоя (например, на ребрах мамонта с кв.Г-8, а-9, А-9 в Р-I и др.). Кости с отпечатками корневой системы растений, часто имеющие темно-серый цвет, первоначально были погребены в почвенном слое с повышенным воздействием гуминовых кислот. К химическим изменениям можно отнести и образование на некоторых костях известковых наростов, причиной чему такжеявляются процессы древнего почвообразования.
Биоповреждения, не связанные с деятельностью человека – это погрызы. Их следы очень редки, такие кости составляют менее 0,25% общей массы. Погрызы преимущественно нанесены мышевидными грызунами (фаланги и метаподии мамонта), лишь один случай можно трактовать, как деятельность хищника – следы зубов на головке ребра молодого мамонта (Р-I кв.А-3). Хищники, видимо, редко рисковали проникать на территорию поселения.
Фрагментированность. Помимо климатического разрушения, как до захоронения, так и во время осадконакопления, ископаемый материал подвергается механическому разрушению. Оказалось, что наиболее сильно раздроблены черепа, кости скелета поясов и свободных конечностей. Позвонки часто имеют естественный излом, отделяющий тело позвонка от дуги и дугу от венечного отростка. Значительную часть раздробленных костных остатков составляют фрагменты диафизов трубчатых костей. Нам представляется, что большая часть фрагментации костей имеет в своей основе антропогенное воздействие. Иное дело – в каком случае мы наблюдаем преднамеренное, а в каком – случайное раскалывание (например, растаптывание)? Костные остатки не переносились водными потоками, солифлюкцией и склоновыми процессами, они попали в отложения стоянки лишь за счет хозяйственной деятельности человека. При этом нельзя

119
исключить, что какая-то часть костей мамонта изъята из располагавшегося неподалеку природного скопления – «мамонтового кладбища», сходного с объектом исследованным в Севске (Мащенко, 2000). В таких «кладбищах» шло накопление туш мамонта, сносившихся половодьями с поймы и скелетировавшихся уже на месте отложения.
Археозоология Изучение остеологического материала показало, что в отложениях основного
верхнепалеолитического культурного слоя поселения Хотылево 2 доминируют костные остатки мамонта, который являлся одним из главных объектов как охоты, так и собирательства. Второе место по количеству особей занимают хищники, на третьем - северный олень и бизон.
Планиграфический анализ показал, что костные остатки распределены по площади культурного слоя неравномерно. В Р-12-14 костный материал имеет два центра сосредоточения (Рис.5). Один из них – в восточной части исследованной площадки – связан с окраиной скоплений, вскрытых Р-8 и 11 Ф.М. Заверняева. Второй расположен в районе округлого зольного скопления (возможно следов легкого наземного жилища). В Р-I центром сосредоточения костного материала стала зона очага и участок, лежащий к северу от него (вероятно также следы легкого наземного жилища) (Рис.6). Однако распределение отдельных категорий костей не всегда повторяет эту картину.
Рис.5. Распределение костных остатков в Хотылево 2 на площади раскопов 12-14 (1994-2001 гг.)
40-60
20-40
0-20
Рис.6. Распределение костных остатков в Хотылево 2 на площади раскопа I (2001-2003)
Череп мамонта. Все черепа сильно фрагментированы, установить их число на исследованной территории трудно, но можно учесть, что слуховые капсулы и затылочные мыщелки принадлежат как минимум 3 особям, подъязычные кости – 2 особями (включая одного мамонтенка). По крайней мере у одной из них не все черепные швы срослись к моменту гибели, у другой срослись неполно. Видимо два черепа были разрушены в районе восточной стенки Р-13 (раздробленный материал тянется из раскопов Ф.М. Заверняева), два компактных скопления наблюдается на южной периферии зольного скопления и к западу от него (один череп здесь принадлежал полувзрослой особи), но еще одно, несомненно, уходит под юго-западный угол Р-14. Распределение фрагментов черепов мамонта явственно связано с расположением скоплений зольной массы (вероятно, остатков наземных легких сезонных жилищ). Вполне возможно, что черепа использовались в конструкциях, но не сохранились в целости, поскольку не были защищены грунтом. Таким образом, в юго-западном углу Р-14 можно предполагать край нового такого объекта. Об этом косвенно свидетельствуют также понижение микрорельефа и наличие двух вбитых в материк ребер. В Р-I концентрация фрагментов черепа имеется в районе очага и на участке к северу от него. Еще два черепа, вероятно, распались в северной и юго-восточной частях раскопа. Черепа здесь принадлежали как минимум трем разновозрастным особям.
Зубы мамонта. Целых зубов в Р-12-14 мало. Практически все они расположены в пределах Р-14, где более быстрое захоронение, вероятно, не дало им разрушиться. Что же касается фрагментов зубов – основная их часть перемешана с обломками черепа в средней части раскопа у восточной стенки. Это, на наш взгляд, еще одно свидетельство того, что мы в этом случае имеем дело с черепом, разрушившимся в результате продолжительного нахождения на дневной поверхности. В Р-I зубы и их фрагменты (включая крошку) сосредоточены также в местах

120
предполагаемого разрушения черепов (у очага и в северной и юго-восточной частях). В целом сохранность и плотность расположения целых зубов здесь выше, чем в Р-12-14.
Бивни мамонта. Все бивни фрагментированы, как правило, размер фрагментов не более 10 см, хотя длина некоторых превышает 25 см. Большая часть группируется на востоке вскрытой площади, маркируя периферию комплексов, вскрытых раскопами Ф.М. Заверняева. Многочисленность бивневой крошки и продольно расчлененные фрагменты бивней заставляют полагать, что мы имеем дело с местом первичной обработки бивневого сырья. Два компактных, но слабо насыщенных скопления отмечены в восточной и западной частях Р-14. Кстати, продольное расщепление некоторых бивней можно расценивать как косвенное свидетельство того, что для обработки использовался в числе прочего бивень, некоторое время пролежавший на открытом воздухе. В Р-I место обработки бивня связано, скорее всего, с юго-восточной его частью, под стенку которой, вероятно, уходит неисследованный пока, но многообещающий комплекс. Небольшая концентрация фрагментов бивня наблюдается и к северу от очага.
Осевой скелет мамонта. В Р-13-14 позвонки группируются в основном к западу от зольного скопления (в районе очага), небольшая концентрация наблюдается и на границе с Р-8 Ф.М. Заверняева (также у зольного скопления). Зона же с наибольшей плотностью ребер и их фрагментов практически соответствует зоне максимальной концентрации костных остатков. Большое количество ребер в районе округлого золистого скопления (возможных остатков легкого наземного жилища) позволяет предполагать, что их использовали не только в качестве вбитых в грунт клиньев в ямах 1 и 2, но, возможно и в качестве некоего каркаса легкой наземной конструкции. В Р-I позвонки также явно группируются в районе очага на квадратах а-В-8-10. Часть из них обожжена (иногда только со стороны, обращенной к углистому слою), с некоторых снята компакта. Не остается сомнений, эти кости, богатые жировой тканью, обитателями стоянки использовались в основном в качестве топлива, как мы полагали и ранее (Чубур, 2003). Ребра же в Р-I, напротив, весьма компактно группируются на северных краях из зольных скоплений, причем залегают преимущественно в верхней части культурного слоя (отметка -1060 от условного репера и выше). Нельзя исключать, что они не просто складированы, а использовались для сооружения каркаса легкой конструкции типа шалаша или чума у очага. Одно из ребер (кв. В-13) вбито в материк, что может свидетельствовать о наличии новых объектов к югу от вскрытой площади.
Плоские кости мамонта все сохранились исключительно в обломках, как правило, не более 15 см в поперечнике. Концентрация таких фрагментов в Р-14 наблюдается, как и в случае с позвонками, в районе очага, к востоку от округлого золистого скопления. По всей вероятности – это остатки одной разбитой тазовой кости. Такое дробление могло производиться, скорее всего, для использования кости в качестве топлива. В Р-I расположение осколков плоских костей хаотично, 6 из 10 фрагментов тазовой кости связано с нижним горизонтом культурного слоя.
Длинные кости конечностей мамонта, а точнее их осколки, в Р-12-14 напротив, концентрируются преимущественно в пределах округлого золистого скопления и в ближайших его окрестностях. Большая часть из них, вероятно, следы трапез. Об этом говорит характерный спиралевидный излом, характерный для искусственного раскалывания трубчатых костей с целью добывания костного мозга. Часть обломков, в первую очередь с сохранившимися суставными поверхностями, была использована и в качестве клиньев, вбитых здесь же в землю (ямы 1 и 2) и являющих собой, по нашему мнению, деталь конструкции легкого наземного жилища. В Р-I осколки диафизов трубчатых костей сосредоточены в районе очага и к северу от него, также являя собой следы трапез. Осколки эпифизов распределены более хаотично. Целые берцовые кости на кв.А-7, Б-7, А-4 и Б-4 могут быть деталями сооружения. Так или иначе, преднамеренность их расположения не оставляет сомнений.
Кости дистальных отделов конечностей мамонта (предплюсна, запястье, метаподии, фаланги, сессамовидные кости) концентрируются в центре и на востоке раскопа. Создается впечатление, что территория, занятая округлым золистым скоплением (предполагаемым жилищем), преднамеренно расчищена от этих костей. В Р-I зона концентрации таких костей перекрывает собой очаг и прилегающие к нему территории.
Интересно, что и в Р-13-14 и в Р-I кости мамонтенка группируются в окрестностях скоплений очажной массы – эти детеныши, несомненно, были съедены обитателями поселения. С другой стороны, доля ювенальных особей в охотничьей добыче весьма мала, то есть забой детенышей, несмотря на то, что он более прост, явление в целом нехарактерное для охотничьей стратегии населения Хотылево 2.
Кости росомахи – разрозненные фрагменты черепа и скелета разбросаны почти по площади Р-12-14 и принадлежат 1 особи. Лишь в 2 случаях кости сохранили анатомический порядок: задняя

121
лапка в кв.2.3 (берцовые кости и дистальный отдел, за исключением когтевых фаланг), а также четыре поясничных позвонка с вонзенным в них кремневым наконечником, послужившим причиной гибели животного. Метаподии и фаланги (включая когтевые) были обнаружены и на кв.9.4. Нам представляется, что они отделены от тушки вместе со шкурой. Тушка убитой росомахи была освежевана, выброшена, а затем, вероятно, растерзана песцами. Из этого следует предположение о сезонном обитании людей в этой части поселения, тем паче трудно представить себе разлагающийся освежеванный труп росомахи непосредственно у обитаемого жилья.
Кости песца – немногочисленны, в Р-12-14 все они, кроме анатомической группы из таранной и пяточной кости из кв.1.5, беспорядочно, но компактно разбросаны на кв.-1-4 – 11-15 к западу от предполагаемого жилища. Еще более хаотично расположены кости песца в Р-I. Нельзя исключать, что песца, как в Юдиново, эпизодически употребляли в пищу, однако фактов, достоверно свидетельствующих в пользу такой гипотезы, не наблюдается.
Кости волка в Р-12-14 очень малочисленны и расположены хаотично. Что касается размещения костей волка в Р-I, можно заметить, что кости, включая анатомические группы костей конечностей и осколки черепа, сосредоточены к югу от очага и в его пределах (но не обожжены, из чего следует, что во время их отложения очаг уже не использовался). Скорее всего, тушка волка растерзана песцами, безнаказанно себя чувствовавшими во время необитаемости данного участка поселения. Осколки еще одного черепа лежат среди скопления ребер мамонта к северу от очага – не исключено, что этот череп венчал конструкцию предполагаемого легкого жилища и разрушился при падении. Третий череп и скопление костей волка лежали в основании культурного слоя в юго-западном углу у края еще одного скопления зольной массы. Выше уже не раз отмечалось, что там может скрываться окраина неисследованного нового объекта.
Кости бизона и оленя в Р-12-14 немногочисленны, организованности, упорядоченности в их расположении заметить не удалось. Кости и фрагменты рога северного оленя сосредоточены в северной части Р-I с менее сохранившимся культурным слоем, поврежденным современным оврагом.
Неопределимые кости. К сожалению, достаточно трудно судить о распределении неопределимых костей на большей части Р-12-14, поскольку не все они поначалу сохранялись в коллекции. В Р-I их распределение в целом повторяет картину распределения совокупности остеологического материала, наибольшее их число связано с очагами и скоплениями зольной массы. По всей видимости, большинство неопределимых осколков костей – не что иное, как преднамеренно раздробленный для использования в качестве топлива костный материал.
Палеоарт. Отдельно следует остановиться на находке, отнесенной нами к категории палеоарта – использованным для хозяйственных нужд и украшения палеонтологическим объектам. В кв.1.8 в предполагаемых пределах остатков жилища, встречена раковина моллюска Gryphaea dilatata, возможно использовавшаяся, как емкость для минеральной краски. Такие же раковины известны из кв.31 Р-11, кв.28 Р-4, ямы на кв.31 Р-6, ямы 6 Р-3 Ф.М.Заверняева. Происходят они из серых и черных глин келловейского яруса юрской системы, выходящих на поверхность в 15-20 км к северо-востоку от Хотылево в долине р.Болва. Раковины могли добываться одновременно с сидеритом, использовавшимся для получения минеральной краски.
Итак, участки, вскрытые Р-12-14 и I, существенно отличает от ранее исследованных Ф.М. Заверняевым отсутствие групп костей мамонта в анатомической связи (исключая отдельные фрагменты хвостовой части позвоночного столба), тогда как в Р-1-8 количество таких групп достаточно велико. Практически отсутствуют здесь целые и слабо фрагментированные трубчатые кости мамонта, целые его черепа, лопатки, тазовые кости, обычные для большей части площади, исследованной Ф.М.Заверняевым. Все это, на наш взгляд, косвенно свидетельствует о различном происхождении этих костных остатков (результат собирательства – кости некоторых анатомических групп и крупные целые кости с одной стороны, и разрозненные и раздробленные костные кухонные отбросы с другой). Имеются и общие для всей территории стоянки черты – использование позвонков мамонта в качестве топлива, размещение черепов мамонта у скоплений зольной массы, наличие анатомических групп костей хищников. Возрастной состав мамонта, определенный по сменам зубов несколько отличается от данных, полученных по материалам Ф.М. Заверняева. Если в Р-1-11 доля полувзрослых и молодых животных составляет около 37% (Чубур, 2003, табл.4), то в новых раскопах не менее 57% животных. Вероятно, К.Н.Гавриловым исследован функционально иной участок поселения – кости здесь лежат преимущественно не в конструкциях (для которых могли быть получены путем собирательства), а в кухонных отбросах. Это, как нам представляется, следы собственно охотничьего лагеря теплого сезона. Именно в это время, на наш взгляд, происходило накопление костей мамонта, бизона, а возможно и северного оленя. Не

122
противоречат весенне-летнему периоду обитания и встреченные в Р-13 костные остатки сформировавшегося эмбриона мамонта или новорожденного мамонтенка.
По набору скелетных элементов и по костям, сохранившимся в анатомическом сочленении, можно сделать вывод, что добытые хищники, часто приносились на стоянку целиком (возможно, когда смерть настигала их в непосредственной близости от поселения). Шкурки песцов, волков, росомах снимались с фалангами, а порой и с метаподиями. Период добычи хищников, скорее всего, не совпадал со временем обитания на вскрытой площадке: мех, ради которого их добывали, имеет оптимальное качество в зимнее время, да и разлагающиеся части тушек в районе жилья представить нелегко. Сомнительно и то, чтобы мелкие хищники терзали эти тушки в непосредственной близости от людей. Таким образом, останки хищников, по нашему мнению, отражают зимний период жизни поселения, когда данный участок был необитаем. Достоверных следов употребления мяса хищников в пищу в Хотылево 2 нами пока не отмечено.
Копытные и другие крупные животные, судя по всему, разделывались сразу после завершения охоты, поскольку остывшая, а тем паче подмороженная туша требует многократного увеличения трудозатрат (Haynes, 1982). Малое количество и выборочность элементов скелета копытных представленных на стоянке, можно объяснить как раз тем, что свежевание и расчленение туш проводилось непосредственно на месте забоя (Binford, 1981). Там малоценные части оставлялись, а на поселение приносились, главным образом, шкуры и наиболее мясистые части туш, реже трубчатые кости конечностей, богатые костным мозгом, изредка черепа, рога. Нижние части конечностей бизона могли оставаться со снимаемой с туши шкурой и соответственно транспортироваться анатомически сочлененными – на это указывает находка такого фрагмента из Р-7 Ф.М.Заверняева (Чубур, Головачева, Тимошкин, 1997). Кроме того, малое число костей копытных может быть связано с малочисленностью данных животных в окрестностях поселения в теплом сезоне, к которому мы относим обитание человека на вскрытом участке. Разделка туш (или их частей) мамонта велась и непосредственно на территории стоянки, о чем свидетельствуют обнаруженные на костях порезы, нанесенные каменными орудиями. Таким образом, мы переходим к рассмотрению следов деятельности человека в остеологической коллекции 1994-2003 гг.
Повреждение костей при добыче костного мозга. Многие трубчатые кости мамонта преднамеренно расколоты. Относительная длина фрагментов и спиральная форма края слома аналогичны повреждениям костей, образующихся и в результате дробления челюстями хищников, и в результате растаптывания костей ногами копытных и других крупных животных, и при раскалывании человеком для добычи костного мозга, что убедительно показано Г. Хейнесом (Haynes, 1983). Таким образом, доказать антропогенный фактор раскалывания может только совокупность фактов – распределение расколотых костей, наличие иных следов употребления мяса и т.д. В данном случае, учитывая наличие локальных концентраций таких осколков, а также следов расчленения туш и срезания мяса с костей, раскалывание костей мамонта человеком можно считать убедительно доказанным. Сказать того же о плечевой кости волка из Р-I мы не можем – она, будучи более хрупкой, могла быть расколота вследствие случайного механического воздействия и не может быть свидетельством употребления мяса и костного мозга хищников в пищу.
Порезы. При анализе порезов главными критериями для их выявления было наличие следующих признаков: короткие параллельные штрихи, нанесенные серией ударов по одному месту, в профиль имеющие рельефно выраженные края; сравнительно широкое поперечное сечение; совпадение цвета кости и пореза (Binford, 1981). Перечисленные признаки и положение повреждений на элементах скелета, повторяемость, форма и направление исключают их случайное образование. Повреждения, нанесенные орудиями, делятся на 4 основных типа (снятие шкуры, расчленение, обработка костей, неопределимые порезы). В Р-I порезы зафиксированы примерно на 2% костного материала, в Р-12-14 – на еще меньшем количестве костей.
В основном это следы расчленения туш – глубокие короткие насечки и нарезки на эпифизах и суставных поверхностях – следы разрубания сухожилий (например, нижние эпифизы бедренных костей с кв. А-8 и В-6, коленная чашечка с кв. В-8, таранная с кв.В-4). Интересен атлант молодого мамонта из Р-14, на суставных поверхностях которого наблюдаются глубокие зарубки – по-видимому так первый шейный позвонок отделялся от затылочных мыщелков черепа. Как разновидность нарезок, связанных с расчленением, назовем следы срезания мяса с костей: например, на ребре взрослого мамонта с кв. В-8 имеется множество косых субпараллельных нарезок (следы снятия мяса) и скобления кости. Еще одно свидетельство древних трапез – 9 субпараллельных нарезок на обломке бедренной кости из кв.1.8. Сходна с ними и часть лопатки с кв. Б-8, на которой имеются субпараллельные короткие нарезки длиной около сантиметра каждая.
123
Следы свежевания (за исключением отсутствия когтевых фаланг на лапках волка и росомахи) нами не обнаружены. Чаще встречаются следы обработки кости.
Одной из первых фаз обработки ребра является продольное расчленение вдоль плоскости кости, иногда по заранее прорезанным канавкам на боках ребра. Такие продольно расчлененные ребра в Р-I встречены на кв.а-10, Б-10. Другие ребра несут следы поперечного расчленения (надреза компакты и последующего излома) – например ребра на кв.А-8, А-12. Продольно расчлененные ребра сами по себе могли становиться завершенными орудиями – таковы лощила с кв.А-3, Б-9. Поперечно расчлененные ребра тоже могли применяться почти без доработки – такова рукоять с выбранной из разрезанного ребра частью губчатой ткани с кв.Г-10. Некоторые продольно расчлененные ребра служили заготовками для более сложных поделок, к примеру – фрагмента ребра с кв. Б-5, где на внутренней поверхности компакты нанесена серия поперечных нарезок. В ряде случаев само ребро служило орудием – такова землекопалка из Р-14 (коллекция 2000 г.).
Имеются следы преднамеренной обработки и на трубчатых костях конечностей. Так, у обломка большой берцовой кости с кв. Б-9 (Р-I) округлый конец заполирован, имеются несколько случайных нарезок. Возможно, рабочий конец кости применялся для обработки кож. Обломок диафиза трубчатой кости с того же кв. Б-9, судя по заполированности одного края и наличию на нем протяженных царапин от песчаных зерен, был превращен в совок.
Рис.7. Позвонки мамонта,
использованные в качестве наковален
Активно велась на вскрытых участках памятника и обработка бивня. Бивень сначала разрубался поперек на фрагменты необходимой длины. Так, обрублен конец фрагмента бивня с кв. Б-6-7. Затем скалывались продольные пластины, либо производилось продольное членение бивня. В Р-I об этом свидетельствуют снятые с бивня пластины на кв. А-14 (одна) и В-12 (две, причем на одной наблюдаются следы ударов острым предметом). Нуклеус из бивня, преднамеренно положенный в углубление, перекрытое углистым слоем на кв. Г-12 свидетельствует о такой технологии не менее красноречиво. Заметим, что на его поверхности читаются ритмичные нарезки: 2+2+3 (в сумме 7 – четверть лунного цикла). Иногда осколок бивня, имеющий подходящую форму, сразу становился орудием. Например, фрагментом с заполированным концом с кв. В-9, вероятно, пользовались как лощилом. В других случаях производилась обработка заготовки до придания ей нужной формы (веретенообразный предмет с а-8). Орнаментированная пластина из бивня в Р-14, на наш взгляд, также вполне законченная поделка, отдаленный аналог так называемых «чуринг» из Елисеевичей.
Часть костей участвовала в технологических процессах в качестве вспомогательных средств. Так протяженные нарезки на лопатке с кв. В-8 мы расцениваем, как использование кости в качестве подставки для резания. Некоторые тела позвонков (кв. А-11, а-6, В-9) имеют характерные многогранные вдавливания компактного вещества с мозаичным растрескиванием: судя по всему, они использовались в качестве «наковален», на которые для удобства обработки устанавливался кремневый нуклеус (Рис.7).
Некоторые найденные неподалеку нуклеусы довольно точно укладываются в выбитые в костях углубления. Интересны тела позвонков с частично или полностью снятой каменным инструментом компактой (кв. Б-8, В-9). Все они связаны с очажной массой, некоторые несут следы обугливания. Возможно, их таким образом готовили для растопки очагов (пропитанный жиром
124
позвонок мог играть роль трута или даже жирового «светильника», некий аналог современного «сухого спирта»).
Наконец, нельзя обойти стороной кости, вбитые в грунт вертикально или наклонно. Среди них не только фрагменты трубчатых костей и ребер (в ямах 1 и 2 Р-13 и в юго-западном углу Р-14), но и венечный отросток позвонка на кв. Б-7 (Р-I). На поверхности некоторых вбитых костей имеются нарезки – возможно для того, чтобы не соскальзывали ремни-растяжки, удерживавшие в натянутом состоянии конструкции легких наземных жилищ. Таким образом, вбитые кости могли использоваться как «фундамент» деревянных подпорок свода помещения, так и в качестве колышков для растяжек. К сожалению, имеющийся материал, не позволяет пока сделать более детальной реконструкции жилых и хозяйственных сооружений Хотылево 2, построенных с использованием костей мамонта.
Таким образом, мы имеем свидетельства активного и разнообразного использования костей и бивней мамонта обитателями поселения Хотылево 2.
В заключение упомянем сделанные нами еще по материалам раскопок Ф.М. Заверняева наблюдения, касающиеся технологии изготовления бус из клыков хищников (на вскрытых новыми раскопками участках, заселенных по нашему мнению в теплом сезоне, таких бус встречено не было, вероятно они характерны именно для зимнего одеяния). На костенковско-авдеевских памятниках наблюдается получение отверстий исключительно путем прорезывания. Таких бус в Хотылево 2 большинство, однако, в ряде случаев применялась и техника сверления, характерная для таких памятников Подесенья, как Елисеевичи, Юдиново. Это еще раз подтверждает ключевое место Хотылево 2 в верхнем палеолите Деснянского региона.
Благодарности. Автор благодарит К.Н. Гаврилова (ИА РАН) за возможность полевых наблюдений и работы
с материалом непосредственно в процессе раскопок.
Таблица 1. Видовой состав и численность костных остатков млекопитающих крупного и
среднего размера верхнепалеолитического поселения Хотылево 2 (основной слой) по материалам 1994-2003 гг.
Вид Число остатков число особей
Мамонт Mammuthus primigenius 1555 12
Северный олень Rangifer tarandus 10 1
Бизон Bison priscus 8 1
Волк Canis lupus 80 4
Песец Alopex lagopus 21 2
Росомаха Gulo gulo 77 1
Неопределимые кости Св.300 ?
Всего: Св.2051 21
Таблица 2. Морфометрия посткраниального скелета волка из Хотылево 2 по материалам 1994-2003 гг.
Кость Промер N Lim (мм) M (мм)
максимальная длина 1 - 146,2
ширина dist. Эпифиза 4 34,5-42,6 39,2
ширина prox. эпифиза 1 - 48,0
ширина блока 3 25,0-34,0 30,0
Плечевая
Humerus
поперечник диафиза 1 - 16,0
ширина локтевого отростка 31,0-34,5 32,1 Локтевая
Ulna Поперечник полулунной ямки 4 21,0-23,2 21,9
максимальная длина 1 - 215,0
ширина диафиза 1 - 17,0
ширина prox.эпифиза 1 - 24,0
Лучевая
Radius
ширина dist.эпифиза 1 - 30,0
Большая берцовая
Tibia
ширина dist/ эпифиза 1 - 31,0
125
Таблица 3. Морфометрия черепов волка из раскопа I (2003) Хотылево 2.
Кость Промер М Примечание
общая длина 264,5
основная длина 226,8
кондилобазальная длина 237,0
наибольшая ширина 130,0
мастоидная ширина 124,0
длина лицевой части 148,0
длина ряда РМ/ 69,0 правая сторона
длина ряда М/ 26,0 правая сторона
длина Р4/ 25,0 правая сторона
ширина Р4/ 11,0 правая сторона
I1/-М2/ 132,0 правая сторона
С1/-М2/ 88,5 правая сторона
Cranium
череп
с кв. В-14
Р2/-Р4/ 59,0 правая сторона
длина ряда РМ/ 76,0 левая сторона
длина ряда М/ 23,0 левая сторона
длина Р4/ 25,7 левая сторона
ширина Р4/ 11,0 левая сторона
Cranium
череп
с кв.А-7
Р2/-Р4/ 67,0 левая сторона
Таблица 4. Морфометрия скелета росомахи из Хотылево 2 (1994-2003 гг.) Кость Промер M (мм)
Эпистрофей Длина 34,8
Длина 195,0 Плечевая кость
(левая) (humerus) Ширина дистального эпифиза 43,2
Тазовая кость
(os coxae)
Поперечник вертлужной впадины 27,2
Длина 150,8
Ширина дистального эпифиза 25,0
Большая берцовая
кость (tibia) Поперечник дистального эпифиза 18,0
Длина 32,1 Таранная (astragalus) Ширина блока 23,0
Пяточная (Calcaneus) Длина 47,0
Длина 44,0
Ширина верхней суставной поверхности 9,0
Поперечник верхней суставной поверхности 10,2
Плюсневая Mtt I
Ширина нижней суставной поверхности 8,0
Длина 52,7
Ширина верхней суставной поверхности 7,5
Поперечник верхней суставной поверхности 11,5
Плюсневая Mtt II
Ширина нижней суставной поверхности 9,0
Длина 59,0
Ширина верхней суставной поверхности 10,0
Поперечник верхней суставной поверхности 14,5
Плюсневая Mtt III
Ширина нижней суставной поверхности 10,0
Длина 61,6
Ширина верхней суставной поверхности 7,0
Поперечник верхней суставной поверхности 14,0
Плюсневая
Mtt IV
Ширина нижней суставной поверхности 10,0
Длина 55,5
Ширина верхней суставной поверхности 9,0
Поперечник верхней суставной поверхности 11,6
Плюсневая
Mtt V
Ширина нижней суставной поверхности 9,8
126
Таблица.5. Костные остатки мамонта из Хотылево 2 (раскопки 1994-2003 гг.) Наименование Раскопы 12-14
(1994-2001) Раскоп I (1996, 2001-2003)
Всего
Фрагменты черепа 127 (включая 2
подъязычные кости)
112 (включая
подъязычную кость и 10
фр-тов нижней челюсти)
239
Фрагменты бивня 54 42 96
Зубы целые 6 22 28
Фрагменты и осколки зубов 51 90 141
Позвонки и их фрагменты
в т.ч.: - шейных
- хвостовых
- грудных
- поясничных
77
10
24
14
8
122
4
36
16
5
199
14
60
30
13
Ребра и их фрагменты
в т.ч. проксимальных частей
199
34
207
24
406
58
Лопатки (фрагменты) 3 6 9
Тазовая кость (фрагменты) 23 9 32
Длинные кости конечностей
(целые и фрагменты)
69 92 161
Кости дистальных отделов конечностей 136 108 244
Всего: 745 810 1555
Таблица 6. Показатели зубной системы мамонта из Хотылево 2 по материалам 1994-2003 гг. Толщина эмали
мм
средняя длина пластины
мм
ширина коронки
мм
показатель
генерация n lim M N Lim M n lim M
М3 2 1,9-2,0 1,95 3 10,3-12,7 11,4 3 80-91 86,7
М2 7 1,53-1,95 1,72 6 8,0-12,5 10,4 6 73-93,5 81,8
М1-Р4 11 0,7-2,1 1,58 13 8,0-11 8,9 12 54-70 61,2
Р3 4 0,8-1,1 0,97 3 7-8,4 7,5 2 47-50 48,5
Р2 1 - 1,0 1 - 6,0 2 37,0;40,0 38,5
Таблица 7. Морфометрия посткраниального скелета взрослых мамонтов из Хотылево 2 (по материалам 1994-2003 гг.)
Кость Промер n Lim (мм) M (мм)
Бедренная Femur Диаметр головки 1 - 144,5
Большая берцовая Tibia Ширина нижнего эпифиза 2 122, 173 147,5
Max. Длина 2 480, 485 482,5 Малая берцовая Fibula
Ширина нижнего эпифиза 2 78,5 80,0 79,2
Длина 5 95,0-130,0 103,2 Таранная Astragalus
Ширина 5 117,0-151,0 132,2
Пяточная Calcaneus Длина 1 - 173,5
Высота 3 120,0-122,0 121,0 Коленная чашечка
Patella Ширина 3 84,0-95,5 91,5
Литература Амирханов Х.А. Восточный граветт или граветтоидные индустрии центральной и восточной Европы //
Восточный граветт. М.: «Научный мир», 1998 С.15-34. Бурова Н.Д. Особенности формирования костного скопления остатков млекопитающих на
верхнепалеолитической стоянке Юдиново // Особенности развития верхнего палеолита Восточной Европы. Костенки в контексте палеолита Евразии. Тр. Костенковской экспедиции ИИМК РАН, вып.1, СПб, 2002, С.196-205.
Величко А.А., Грибченко Ю.Н., Маркова А.К., Ударцев В.П. О возрасте и условиях обитания стоянки Хотылево 2 на Десне // Палеоэкология древнего человека. М.: «Наука», 1977. С.41-50.
Гаврилов К.Н. Хотылевская верхнепалеолитическая стоянка // Восточный граветт. Тез. док. М., 1997, с.25-28. Кузьмина И.Е. Сравнительная морфологическая характеристика мамонтов Русской равнины, Урала и Якутии в
позднем плейстоцене // Плейстоценовые и голоценовые фауны Урала. (Биота северной Евразии в кайнозое: вып.1). Челябинск, 2000. С.3-16.

127
Кузьмина И.Е., Саблин М.В. Песцы позднего плейстоцена верховьев Десны // Материалы по мезозойской и кайнозойской истории наземных позвоночных. Труды Зоологического института РАН, т.249,СПб, 1993, С.93-104
Кузьмина И.Е., Саблин М.В. Волк из позднепалеолитических стоянок Костенки на Дону // Четвертичная фауна северной Евразии. Труды Зоологического института РАН, т.256, СПб, 1994, С.59-68,
Мащенко Е.Н. Структура стада мамонтов из Севского позднеплейстоценового местонахождения (Брянская обл.) // История крупных млекопитающих и птиц северной Евразии. Труды Зоологического института РАН, т.246, СПб, 1992.С.41-59.
Мащенко Е.Н. Последние мамонты Русской равнины // Химия и жизнь, 2000 - №8, С.32-37. Саблин М.В. Позднеплейстоценовый песец из Костенок Воронежской области // Четвертичная фауна северной
Евразии. Труды Зоологического института РАН, т.256, СПб, 1994, С.59-68. Чубур А.А. О мамонтах с верхнепалеолитической стоянки Хотылево 2 (по материалам раскопок Ф.М.
Заверняева) // Деснинские древности, Вып.1, Материалы 1-й Межгос. науч. конф. памяти Ф.М. Заверняева. Брянск, 1995, С.28-31.
Чубур А.А. Быки. Новый палеолитический микрорегион и его место в верхнем палеолите Русской равнины. Брянск: «Брянск-Сегодня», 2001.
Чубур А.А. Мамонт Mammuthus primigenius Blumenbach (1799) c верхнепалеолитической стоянки Хотылево 2 // Фондовая работа в естественнонаучном музее (Труды Государственного Дарвиновского музея. Вып.6), М., 2003. С.226-237.
Чубур А.А. Связи населения бассейнов Дона и Десны в верхнем палеолите // Верхнее Подонье. Природа, археология, история, т.1, Тула, 2004. С.75-93.
Чубур А.А., Головачева И.В., Тимошкин В.М. Хищники, жвачные и непарнопалые с верхнепалеолитической стоянки Хотылево-2 // Песоченский историко-археологический сборник, вып.3, ч.1. Киров, 1977 С.14-20.
Binford L.R. Bones: Ancient men and modern muths. Orlando: Academic press, 1984. Haynes G. Utilization and skeletal disturbances of North American prey carcasses // Arctic, vol.35, No 2, 1982, p.266-
281. Haynes G. Frequencies of spiral and green-bone fractures on ungulate limb bones in modern surface assemblages //
American antiquity, vol.45 No1, 1983. p.102-114. Soffer O. The Upper Paleolithic of the Central Russian plain. Orlando: Academic press, 1985.
PALEONTOLOGY, ARHEOZOOLOGY AND TAFONOMY OF THE UPPER PALEOLITHIC
SETTLEMENT CHOTYLEVO 2 (BASED ON EXCAVATIONS OF 1994-2003).
A.A. Chubur Osteological material from the excavations of 1994-2003. employs more than 1700 mammal bones. The species and anatomical structure, are the natural mechanical damage, chemical changes under the influence of soil formation, nibble, traces of man (dissection of carcasses, food, getting bone marrow, the use of bones in the design). The main conclusion: Excavation revealed traces of a hunting camp of the warm season.
Keywords: zooarheologiya, Paleolithic, Khotylevo 2, mammoth, mammoth fauna
УДК 370.176
ИСПОЛЬЗОВАНИЕ ИНТЕРНЕТ-ТЕХНОЛОГИЙ ДЛЯ ОРГАНИЗАЦИИ НАУЧНО-ИССЛЕДОВАТЕЛЬСКОЙ РАБОТЫ В ВУЗЕ
Т.А. Николаева
Брянский государственный университет имени академика И.Г. Петровского
Данная статья представляет основные направления и задачи действующей с целью эффективной организации научно-исследовательской работы в БГУ лаборатории «Интернет образование и образовательные ресурсы». Материал статьи демонстрирует актуальность использования образовательных ресурсов сети Интернет в системе обучения, открывающих широкие возможности для всех участников образовательного процесса. Ключевые слова: научно-исследовательская лаборатория, исследовательская деятельность, образовательные ресурсы сети Интернет.
Основой подготовки специалистов мирового уровня в университете является интеграция в образовательный процесс научных исследований. Для реализации объявленного принципа ведется разработка междисциплинарных подходов к учебному процессу, создание комплексных научных центров и лабораторий для проведения различных исследований с учетом развития непрерывного образования. С этой целью оздаются условия для привлечения в университет молодых кадров,
Copyright © 2022 FDOKUMEN




















![Emersonian : [Emerson College yearbook]](https://static.fdokumen.com/doc/165x107/6321be5e117b4414ec0b98a5/emersonian-emerson-college-yearbook.jpg)