
Paleoceanographic evolution of the northeastern Indian Ocean during the Miocene: Evidence from...
10
Indian Journal of Marine Sciences Vol. 36(4), December 2007, pp. 332-341 Paleoceanographic evolution of the northeastern Indian Ocean during the Miocene: Evidence from deep-sea benthic foraminifera (DSDP Hole 216A) Ajoy K. Bhaumik, Anil K. Gupta*, M. Sundar Raj, K. Mohan, Soma De & Sudipta Sarkar Department of Geology and Geophysics, Indian Institute of Technology, Kharagpur-721 302, India *[E-mail: [email protected] ] Statistical analyses (factor and cluster) were performed on 30 highest ranked deep-sea benthic foraminifer species from >149 μm size fraction from Deep Sea Drilling Project Hole 216A to understand Miocene (~20.5 to ~7 Ma) paleoceanographic evolution of the northeastern Indian Ocean. Factor and cluster analyses enabled us to identify five biofacies defining five clusters. Known ecological preferences of benthic foraminifera were used for environmental interpretations. The faunal data documents a shift in deep-sea ventilation and productivity at 15-14 Ma, coinciding with the abrupt cooling in the middle Miocene. This coincides with the beginning of permanent ice sheets in Antarctica during the middle Miocene. [Key words: Benthic foraminifera, Indian Ocean, paleoceanography, Miocene] Introduction Benthic foraminifera have good potential for paleoenvironmental reconstruction owing to their diverse morphological variations, greater abundance, wide distribution and well preserved fossil record. The vertical distribution of benthic foraminifera is mainly controlled by the quality and quantity of food and deep-sea oxygenation 1-8 . The eutrophic regions are characterized by the dominance of low-oxygen species due to decreasing oxygen content along the sediment water interface. The oligotrophic environments are favorable for epifaunal, well- oxygenated species 9, 10 . Some workers have found a relation between benthic foraminifera and deep-sea water masses. For instance, Cibicides wuellerstorfi is widely used as an indicator of well ventilated North Atlantic Deep Water (NADW) whereas Nuttallides umbonifera is widely used as an indicator of carbonate undersaturated Antarctic Bottom Water (AABW) 4, 11-13 . Some attempts have been made to reconstruct paleoclimate variability including the Indian monsoon during the Quaternary and older intervals using this group 7, 10, 14, 15 . In the present study we attempt to understand paleoceanographic changes in the northeastern Indian Ocean during the Miocene using multivariate data of deep-sea benthic foraminifera combined with published stable oxygen and carbon isotopic values of Oridorsalis umbonatus 16 from Deep Sea Drilling Project Hole 216A, Leg 22 (Fig. 1). Known ecological preferences of benthic foraminifera from different ocean basins have been used to understand paleoceanographic changes in the northeastern Indian Ocean. Materials and Methods Deep Sea Drilling Project (DSDP) Hole 216A (lat 01° 27.73΄ N; long 90° 12.48΄ E; water depth 2262 m) is located on the northern side of the Ninetyeast Ridge below the equatorial divergence in the northeastern Indian Ocean basin (Fig. 1). The deep and intermediate currents in the eastern Indian Ocean are generated by the mixing of water masses from the Atlantic and Southern oceans 17 as shown in Fig. 1. The top 1000 m water column in the equatorial Indian Ocean is characterized by high productivity, low salinity and oxygen-poor deep watermass 18 . Depths between 1000 and 4000 m are bathed by the southward flowing North Indian Deep Water (NIDW), which is a mixture of NADW, AABW and deep water mass of northern Indian Ocean 19 . The well-oxygenated, cold AABW (having a potential temperature 0.9°C to 1.2°C) lies below 4000 m, which originates from the Weddell Sea and the Ross Sea 20, 21 . The benthic foraminiferal lysocline 14 lies near 3600 m. The present day deep- _________ *Corresponding author: Ph: +91-3222-283368 Fax: +91-3222-282700
Transcript of Paleoceanographic evolution of the northeastern Indian Ocean during the Miocene: Evidence from...

Indian Journal of Marine Sciences Vol. 36(4), December 2007, pp. 332-341
Paleoceanographic evolution of the northeastern Indian Ocean during the Miocene: Evidence from deep-sea benthic foraminifera (DSDP Hole 216A)
Ajoy K. Bhaumik, Anil K. Gupta*, M. Sundar Raj, K. Mohan, Soma De & Sudipta Sarkar Department of Geology and Geophysics, Indian Institute of Technology, Kharagpur-721 302, India
*[E-mail: [email protected] ]
Statistical analyses (factor and cluster) were performed on 30 highest ranked deep-sea benthic foraminifer species from >149 µm size fraction from Deep Sea Drilling Project Hole 216A to understand Miocene (~20.5 to ~7 Ma) paleoceanographic evolution of the northeastern Indian Ocean. Factor and cluster analyses enabled us to identify five biofacies defining five clusters. Known ecological preferences of benthic foraminifera were used for environmental interpretations. The faunal data documents a shift in deep-sea ventilation and productivity at 15-14 Ma, coinciding with the abrupt cooling in the middle Miocene. This coincides with the beginning of permanent ice sheets in Antarctica during the middle Miocene.
[Key words: Benthic foraminifera, Indian Ocean, paleoceanography, Miocene]
Introduction Benthic foraminifera have good potential for paleoenvironmental reconstruction owing to their diverse morphological variations, greater abundance, wide distribution and well preserved fossil record. The vertical distribution of benthic foraminifera is mainly controlled by the quality and quantity of food and deep-sea oxygenation1-8. The eutrophic regions are characterized by the dominance of low-oxygen species due to decreasing oxygen content along the sediment water interface. The oligotrophic environments are favorable for epifaunal, well-oxygenated species9, 10. Some workers have found a relation between benthic foraminifera and deep-sea water masses. For instance, Cibicides wuellerstorfi is widely used as an indicator of well ventilated North Atlantic Deep Water (NADW) whereas Nuttallides umbonifera is widely used as an indicator of carbonate undersaturated Antarctic Bottom Water (AABW)4, 11-13. Some attempts have been made to reconstruct paleoclimate variability including the Indian monsoon during the Quaternary and older intervals using this group7, 10, 14, 15. In the present study we attempt to understand paleoceanographic changes in the northeastern Indian Ocean during the Miocene using multivariate data of
deep-sea benthic foraminifera combined with published stable oxygen and carbon isotopic values of Oridorsalis umbonatus16 from Deep Sea Drilling Project Hole 216A, Leg 22 (Fig. 1). Known ecological preferences of benthic foraminifera from different ocean basins have been used to understand paleoceanographic changes in the northeastern Indian Ocean. Materials and Methods Deep Sea Drilling Project (DSDP) Hole 216A (lat 01° 27.73΄ N; long 90° 12.48΄ E; water depth 2262 m) is located on the northern side of the Ninetyeast Ridge below the equatorial divergence in the northeastern Indian Ocean basin (Fig. 1). The deep and intermediate currents in the eastern Indian Ocean are generated by the mixing of water masses from the Atlantic and Southern oceans17 as shown in Fig. 1. The top 1000 m water column in the equatorial Indian Ocean is characterized by high productivity, low salinity and oxygen-poor deep watermass18. Depths between 1000 and 4000 m are bathed by the southward flowing North Indian Deep Water (NIDW), which is a mixture of NADW, AABW and deep water mass of northern Indian Ocean19. The well-oxygenated, cold AABW (having a potential temperature 0.9°C to 1.2°C) lies below 4000 m, which originates from the Weddell Sea and the Ross Sea20, 21. The benthic foraminiferal lysocline14 lies near 3600 m. The present day deep-
_________ *Corresponding author: Ph: +91-3222-283368 Fax: +91-3222-282700

BHAUMIK et al.: FORAMINIFERA OF DSDP HOLE 216A
333
Fig. 1—Location map of DSDP Hole 216A in the northeastern Indian Ocean. Also shown are main deep-ocean currents (modified from Kawagata et al.17).
water temperature, salinity and oxygen concentration at DSDP Hole 216A are 2.36°C, 34.75 and 135 µM/kg, respectively22.
DSDP Hole 216A has good sediment preservation and was further south of its present position, during the Miocene15, 23. In the earliest Miocene (22 Ma), Hole 216A was situated ~11° S from its present position within the equatorial high productivity belt24. This Site has also been above the Calcite Compensation Depth (CCD) since the Miocene. The paleodepth of Hole 216A was 1869 m in the early Miocene (22 Ma) and 2182 m in the late Miocene24 at 8 Ma. This study is based on the data from seventy samples from the Miocene sequence of Hole 216A (Srinivasan & Gupta25). Samples were processed as described in Srinivasan & Gupta25. Each sample of 10 cc volume was soaked in water with 4-5 drops of H2O2 for 8-12 hours and washed over 63 µm size sieve. Samples were dried at ~50-60°C and then transferred into glass vials. For microscopic observations, samples were dry-sieved over a 149 µm sieve and all specimens of benthic foraminifera from the entire sample were picked and counted. Relative abundances of all benthic foraminiferal species were calculated. Ages are based on planktic foraminiferal faunal datums25 and updated to the age model of Berggren et al.26.
Table 1—List of species with relative abundance >5% and present at least in five samples, used in R-mode factor and Q-mode cluster analyses. Anomalina globulosa Chapman and parr, 1937Astrononion umbilicatulum Uchio, 1952Bulimina alazanensis Cushman, 1927Buliminella carteri Bhatia, 1955Chrysalogonium equisetiformis Schwager, 1866Cibicides bradyi (Trauth, 1918)Cibicides kullenbergi Parker, 1953Cibicides wuellerstorfi Schwager, 1866Eggerella bradyi (Cushman, 1911)Ehrenbergina hystrix Brady, 1884Epistominella exigua (Brady, 1884)Favocassidulina favus Brady, 1884Favocassidulina indica Gupta and Srinivasan, 1990Globocassudulina pacifica Cushman, 1925Globocassudulina subglobosa (Brady, 1884)Gyroidinoides cibaoensis (Bermudez, 1949)Gyroidinoides nitidula (Schwager, 1866)Laticarinina pauperata Parker and Jones, 1865Martinotiella scabra Cushman, 1936Osangularia culter (Parker and Jones, 1865)Planulina marialina gigus Keijzer, 1845Pleurostomella alternans Schwager, 1866Pullenia bulloides (d'Orbigny, 1846)Pullenia osloensis Feyling-Hanssen, 1954Pullenia quinqueloba (Reuss, 1851)Sphaeroidina bulloides d'Orbigny, 1826Stilostomella lepidula (Schwager, 1866)Uvigerina proboscidea Schwager, 1866Vagunilina elegans d'Orbigny, 1933Vulvulina nicobarica Schwager, 1866

INDIAN J. MAR. SCI., VOL. 36, NO. 4, DECEMBER 2007
334
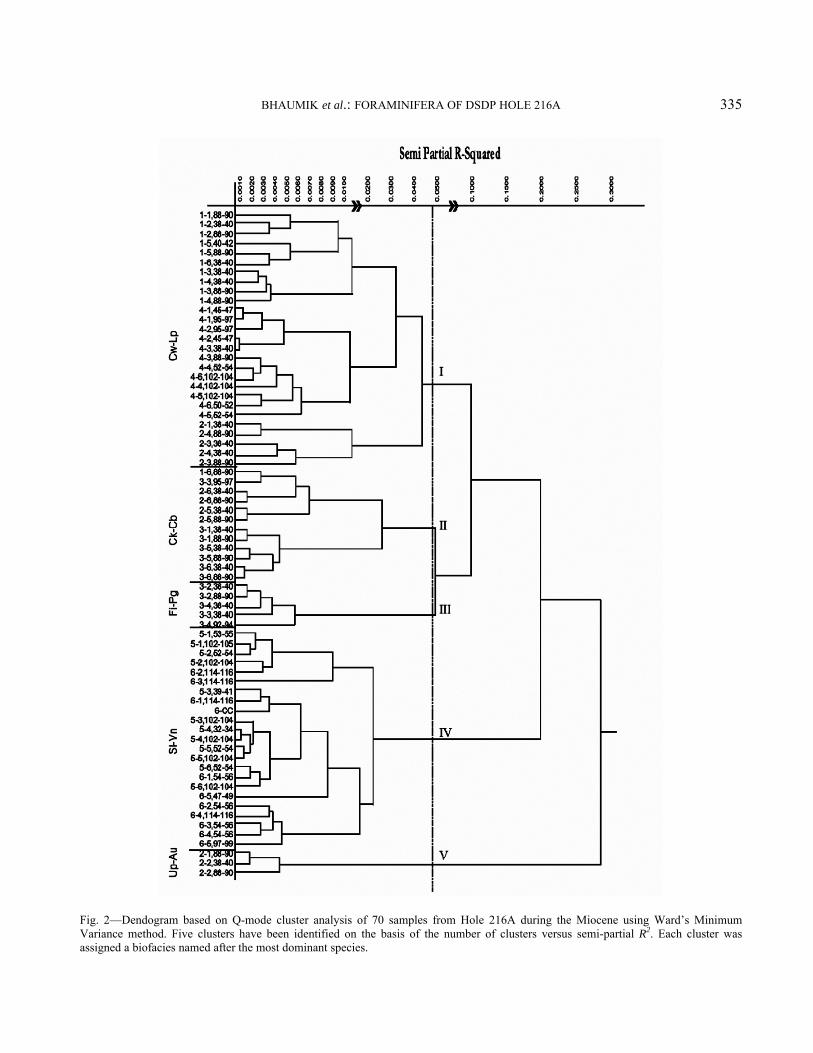
To remove post-depositional noise from the dataset for better paleoceanographic interpretations, factor and cluster analyses were performed on census counts of 30 highest ranked benthic species using SAS/STAT package27. Species were selected on the basis of their relative abundance of 5% or more in any one sample and present in at least 5 samples (Table 1). R-mode Principal Component Analysis (PCA) was performed on the correlation matrix. A scree (x-y) plot of eigen values versus the number of factors and screening of factor scores allowed us to retain 5 factors, accounting for 60.14% of the total variance. The missing values were coded to obtain a better result, because the presence of large number of zeros can bias the result28. Q-mode cluster analysis was performed using Ward’s Minimum Variance method to identify sample groups. To standardize the dataset, a PCA was performed on the covariance matrix of 30 highest ranked benthic foraminifer species prior to cluster analysis. Based on the plot of semi-partial R-squared values versus the number of clusters, 5 clusters were identified (Fig. 2). Principal Components (PCs) that show significant species associations were considered to define biofacies. Factors that do not show significant species associations were not used to define benthic faunal biofacies. In this way,
5 bio-facies were identified and their paleo-environmental preferences were inferred based on the recent distribution of benthic foraminifera (Table 2). Stable oxygen and carbon isotope data of benthic foraminifer Oridorsalis umbonatus (Fig. 3) is from Vincent et al.16.
Table 2—Interpretation of characteristic species comprising different biofacies at Hole 216A
Species Interpretation Astrononion umbilicatulum Oligotrophic, well ventilation10, 29, 30, High salinity29, 31, High organic carbon7, 32, 33
Bulimina alazanensis Infaunal, low oxygen, high food34, High continuous food supply7, NADW and warm benthos fauna4
Buliminella carteri Infaunal34, Low oxygen, intermediate organic flux10
Cibicides bradyi Well oxygen7, 10, 35, 36, Adaptive to low oxygen37
Cibicides kullenbergi Intermediate to low organic flux13, 38, warm deep water39, 40, NADW41, 42
Cibicides wuellerstorfi Epibenthic, prefers to live on elevated substrate, suspension feeder, high energy12, 43, 44, Oligotrophic44, Seasonal food supply45, NADW4, 12, 13, AABW46, 47
Eggerella bradyi Moderate, degraded organic matter, well oxygenated condition31-33, High organic flux, low seasonality33
Epistominella exigua Epibenthic, cosmopolitan, abyssal, opportunistic, phytodetritus feeders48, 49, Low organic flux, well oxygenation32
Favocassidulina favus Suboxic50
Globocassidulina pacifica Cosmopolitan51, 52, Low oxygen, intermediate food7
Gyroidinoides cibaoensis Food limited or pulsed food12, 53, Oligotrophic10
Gyroidinoides nitidula Intermediate organic flux, high seasonality33, 54
Laticarinina pauperata Strongly pulsed, low to intermediate organic flux54
Pleurostomella alternans Deep infaunal, low oxygen, high organic carbon55, 56
Pullenia osloensis High food supply7
Sphaeroidina bulloides High productivity13, 57, Low oxygen species58, 59
Stilostomella lepidula Cosmopolitan38, 61-63, Low oxygen and High organic carbon55, 64, 65
Uvigerina proboscidea High organic carbon, independent of oxygenation37, 38, High productivity7, 10, 14, 31, 66
Results Following biofacies were identified that define benthic assemblages at Hole 216A: Biofacies Sl-Vn This biofacies is defined by the species having high negative scores on Factor 1, ranging from 20.4 to 14.5 Ma. The characteristic species of this biofacies are Stilostomella lepidula, Vulvulina nicobarica, Buliminella carteri, Sphaeroidina bulloides, Pleurostomella alternans, Gyroidinoides nitidula and Globocassidulina pacifica indicating oxygen-depleted, organic carbon rich deep-sea environment (Tables 2 and 3). Biofacies Cw-Lp Cibicides wuellerstorfi, Laticarinina pauperata, Vaginulina elegans, Bulimina alazanensis, Gyroidinoides cibaoensis, Cibicides kullenbergi, Favocassidulina favus and Epistominella exigua are
BHAUMIK et al.: FORAMINIFERA OF DSDP HOLE 216A
335
Fig. 2—Dendogram based on Q-mode cluster analysis of 70 samples from Hole 216A during the Miocene using Ward’s Minimum Variance method. Five clusters have been identified on the basis of the number of clusters versus semi-partial R2. Each cluster was assigned a biofacies named after the most dominant species.
INDIAN J. MAR. SCI., VOL. 36, NO. 4, DECEMBER 2007
336
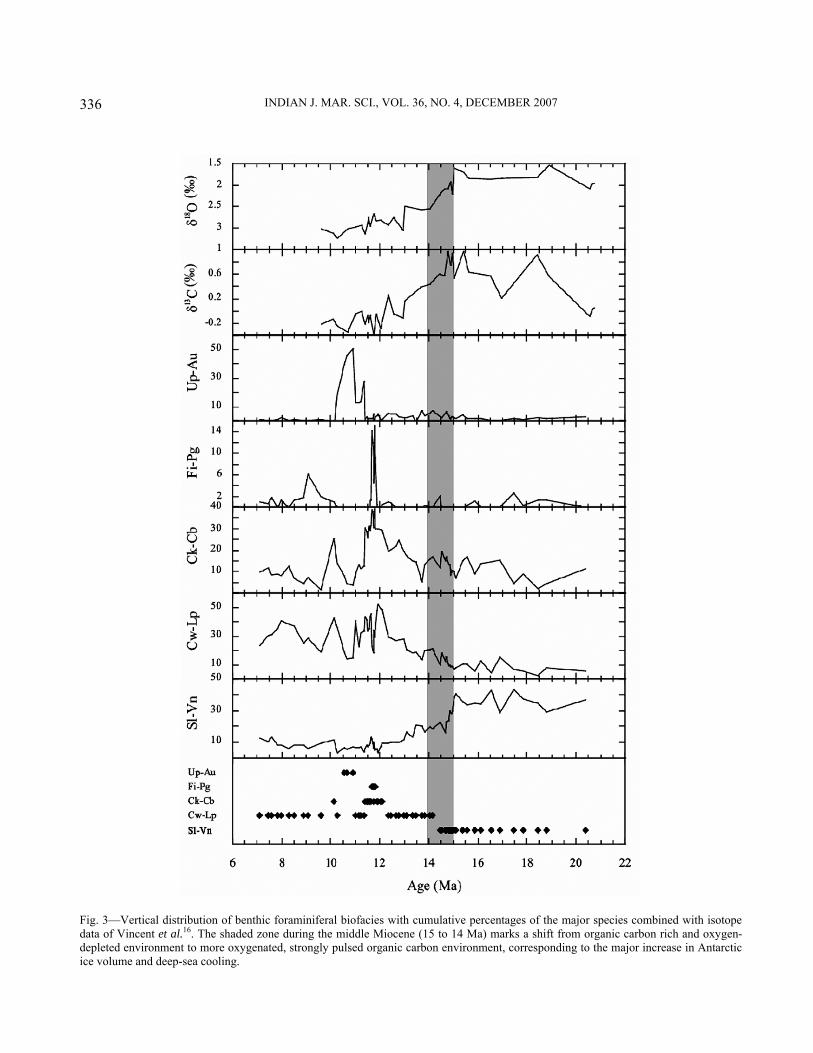
Fig. 3—Vertical distribution of benthic foraminiferal biofacies with cumulative percentages of the major species combined with isotope data of Vincent et al.16. The shaded zone during the middle Miocene (15 to 14 Ma) marks a shift from organic carbon rich and oxygen-depleted environment to more oxygenated, strongly pulsed organic carbon environment, corresponding to the major increase in Antarctic ice volume and deep-sea cooling.

BHAUMIK et al.: FORAMINIFERA OF DSDP HOLE 216A
337
the characteristic species of this biofacies having high positive scores on Factor 1. This biofacies ranges from 14.5 to 7.1 Ma and is distributed over 27 samples. Species association of this biofacies indicates pulsed, low flux of organic matter, well-oxygenated environment and strong ocean current (Tables 2 and 3). Biofacies Ck-Cb Dominant species of this biofacies are Cibicides kullenbergi, Cibicides bradyi, Pullenia osloensis, Favocassidulina indica and Eggerella bradyi with high negative scores on Factor 2, spreading over 12.1 to 11.4 Ma in 12 samples. This biofacies also has a short-lived presence at 10.1 Ma. Species assemblage of this biofacies indicates presence of NADW-like watermass characterized by high oxygen and low organic carbon (Tables 2 and 3).
Biofacies Fi-Pg
Table 3—Benthic foraminiferal biofacies and their interpreted environments at Hole 216A, northeastern Indian Ocean based on Table 2. Stable isotope analysis was done on benthic foraminifer Oridorsalis umbonatus (adapted from Vincent et al.16).
Biofacies Factor scores Environment Sl-Vn (Factor 1 negative) Stilostomella lepidula Vulvulina nicobarica Buliminella carteri Sphaeroidina bulloides Pleurostomella alternans Gyroidinoides nitidula Globocassudulina pacifica
-0.76 -0.75 -0.65 -0.63 -0.56 -0.54 -0.52
Warm interval with oxygen depleted, organic carbon rich environment
Cw-Lp (Factor 1 positive) Cibicides wuellerstorfi Laticarinina pauperata Vagunilina elegans Bulimina alazanensis Gyroidinoides cibaoensis Cibicides kullenbergi Favocassidulina favus Epistominella exigua
0.74 0.68 0.57 0.53 0.51 0.50 0.45 0.36
Pulsed, low flux organic matter (oligotrophic), well oxygenated environment with strong ocean current, high seasonality
Ck-Cb (Factor 2 negative) Cibicides kullenbergi Cibicides bradyi Pullenia osloensis Favocassidulina indica Eggerella bradyi
-0.52 -0.51 -0.51 -0.47 -0.46
NADW like water mass, high oxygen, low organic carbon environment
Fi-Pg (Factor 6 positive) Favocassidulina indica Planulina marialina gigus
0.48 0.39
Well-oxygenated, low-organic carbon environment
Up-Au (Factor 4 negative) Uvigerina proboscidea Astrononion umbilicatulum Martinottiella scabra
-0.65 -0.46 -0.33
High, sustained flux of organic matter, low seasonality
This biofacies consists of two species, Favocassidulina indica and Planulina marialina gigus, bearing high positive scores on Factor 6. This biofacies has a short-lived presence during 11.8 to 10.7 Ma (5 samples). These species have rare and sporadic occurrence in the Indian Ocean and not much is known about their environmental preferences51. Their association with C. wuellerstorfi as reported in few studies from the Indian Ocean suggests their preference to low-organic flux and well-oxygenated environment51. Biofacies Up-Au Biofacies Up-Au comprises Uvigerina probos-cidea, Astrononion umbilicatulum and Martinottiella scabra, showing high negative scores on Factor 4. This biofacies occurs in only 3 samples (10.9 to 10.6

INDIAN J. MAR. SCI., VOL. 36, NO. 4, DECEMBER 2007
338
Ma) and indicates the presence of a short period of high, sustained flux organic matter. This was also a time of major change in the Indian monsoon system14. The distribution of biofacies shows a major change at ~14.5 Ma (Fig. 3) coinciding with the major increase in Antarctic ice volume during the Miocene16,67. From 20.5 to 14.5 Ma the benthic fauna are dominated by biofacies Sl-Vn, which is replaced by biofacies Cw-Lp, Ck-Cb, Fi-Pg and Up-Au during 14.5 to 7 Ma (Fig. 3, Table 3). The interval 14.5-10 Ma is marked by three biofacies Cw-Lp, Ck-Cb, Fi-Pg and Up-Au, suggesting widespread changes in the northeastern Indian Ocean.
Discussion The Miocene represents a link between the warm interval of the Paleogene and the cold sphere of the Neogene67, 68. The Miocene was a critical time in the evolution of Earth’s climate marked by a major increase in Antarctic ice volume67, 68 and widespread deep-sea hiatuses69. The Miocene ice volume brought significant changes in the deep sea. To understand these changes, numerous studies were taken up under Cenozoic Paleoceanography Project (CENOP) using faunal and geochemical proxies68, 70. The middle Miocene marks a major shift towards cold climates and deep waters that may have influenced the deep-sea fauna. During this time, a major increase in δ18O values has been observed throughout the Indian, Pacific and Atlantic oceans representing a major and permanent accumulation of East Antarctic ice sheet, and cooling of deep waters16, 68, 71. Benthic foraminiferal faunal and stable isotope record shows a major transition across the early-middle Miocene at Hole 216A. The deep-sea conditions were warm, oxygen-depleted and organic carbon rich during 20.4 to 14.5 Ma (biofacies Sl-Vn). The δ18O values were lighter but δ13C values were heavier during this time. The high δ13C values are at odds to the dominance of high productivity deep-sea benthic foraminifera during this time (Fig. 3). This contradiction can be explained as the early Miocene was an interval of climate warmth during which time tropical forests were widespread and the continental organic biosphere would have been more extensive, which might have contributed to the positive δ13C values in the early Miocene72. The warm deep-sea temperatures may have also contributed to the heavier values of δ13C in the early Miocene69. Since the middle Miocene (~14.5 Ma), biofacies Cw-Lp, Ck-Cb and Fi-Pg became dominant at Hole
216A indicating low organic flux (oligotrophic) during a weaker Indian monsoon with high seasonality, and stronger deep oceanic currents (Fig. 3). The δ18O values also became heavier since the middle Miocene indicating prolong deep-sea cooling due to the formation of permanent ice sheets in East Antarctica10, 67, 72-74. The interval 12-10 Ma is characterized by four biofacies indicating widespread changes in the northeastern Indian Ocean (Fig. 3). A short-lived peak of biofacies Up-Au during 10.9 to 10.6 Ma indicates intense upwelling and high surface productivity probably driven by the intense Indian monsoon14, 75. The δ13C values became lighter since the middle Miocene indicating increased productivity in the ocean basins. The dominance of low productivity benthic fauna at Hole 216A since the middle Miocene may be attributed to increased availability of suspended food particles and well oxygenated, cold deep waters. Conclusion The biofacies distribution pattern at DSDP Hole 216A divides the Miocene interval into two climatic realms across the early-middle Miocene boundary. During the late early Miocene (20 to 14.5 Ma) warm interval, the deep-sea was rich in organic food and depleted in oxygen. The onset of permanent ice sheets on Antarctica since the middle Miocene brought a significant change in deep-sea circulation as well as deep-sea fauna, increasing the vigor of deep-sea circulation. The middle Miocene climate transition is marked by a shift in δ13C towards lighter values and δ18O towards heavier values, which is also visible in other ocean basins. Thus, late Miocene was similar to the present day world in many aspects. Acknowledgement Authors acknowledge Deep Sea Drilling Project for providing the core samples. We are thankful to Rajiv Nigam for inviting us to contribute this article. References: 1 Corliss B H & Emerson S, Distribution of Rose Bengal
stained deep-sea benthic foraminifera from the Nova Scotian continental margin and Gulf of Maine, Deep-Sea Res, 37 (1990) 381-400.
2 Sen Gupta B K & Machain-Castillo M L, Benthic foraminifera in oxygen-poor habitats, Mar Micropaleontol, 20 (1993) 183-201.
3 Jorissen F J, De Stigter H C & Widmark J, A conceptual model explaining benthic foraminiferal microhabitats, Mar Micropaleontol, 26 (1995) 3-15.

BHAUMIK et al.: FORAMINIFERA OF DSDP HOLE 216A
339
4 Schmiedl G & Mackensen A, Late Quaternary paleoproductivity and deep water circulation in the eastern South Atlantic Ocean: Evidence from benthic foraminifera, Palaeogeogr Palaeoclimatol Palaeoecol, 130 (1997) 43-80.
5 Van der Zwaan G J, Duijnstee I A P, den Dulk M, Ernst S R, Jannink N T & Kowenhoven T J, Benthic foraminifers: proxies or problems? A review of paleoecological concepts, Earth-Sci. Rev., 46 (1999) 213-236.
6 Gupta A K, Latest Pliocene through Holocene paleoceanography of the eastern Indian Ocean: benthic foraminiferal evidence, Mar Geol, 161 (1999) 63-73.
7 Gupta A K & Thomas E, Latest Miocene-Pleistocene productivity and deep-sea ventilation in the northwestern Indian Ocean (DSDP Site 219), Paleoceanography, 14 (1999) 62-73.
8 den Dulk M, Reichart G J, van Heyst S, Zachariasse W J & Van der Zwaan G J, Benthic foraminifera as proxies of organic matter flux and bottom water oxygenation? A case history from the northern Arabian Sea, Palaeogeogr Palaeoclimatol Palaeoecol, 161 (2000) 337-359.
9 Gooday A J, Deep-sea benthic foraminiferal species which exploit phytodetritus: Characteristic features and controls on distribution, Mar Micropaleontol, 22 (1993) 197-205.
10 Singh R K & Gupta A K, Late Oligocene–Miocene paleoceanographic evolution of the southeastern Indian Ocean: evidence from deep-sea benthic foraminifera (ODP Site 757), Mar Micropaleontol, 51 (2004) 153-170.
11 Bremer M L & Lohmann G P, Evidence for primary control of the distribution of certain Atlantic Ocean benthonic foraminifera by degree of carbonate saturation, Deep-Sea Res, 29 (1982) 987-998.
12 Mackensen A, Schmiedl G, Harloff J & Giese M, Deep-sea foraminifera in the South Atlantic Ocean: Ecology and assemblage generation, Micropaleontology, 41 (1995) 342-358.
13 Gooday A J, Benthic Foraminifera (Protista) as Tools in Deep-Water Paleoceanography: Environmental influences on Faunal Characteristics, Adv. Mar. Biol., 46 (2003) 1-90.
14 Gupta A K & Srinivasan M S, Uvigerina proboscidea abundances and paleoceanography of the Northern Indian Ocean DSDP Site 214 during the late Neogene, Mar Micropaleontol, 19 (1992) 355-367.
15 Gupta A K, Singh R K, Joseph S & Thomas E, Indian Ocean high-productivity event (10-8 Ma): linked to global cooling or to the initiation of the Indian monsoons? Geology, 32 (2004) 753-756.
16 Vincent E, Killingley J S & Berger W H, Miocene oxygen and carbon isotope stratigraphy of the tropical Indian Ocean, In: The Miocene ocean, edited by Kennett J P, Geol Soc Am Mem, 163 (1985) pp. 103-130.
17 Kawagata S, Hayward B W & Gupta A K, Benthic foraminiferal extinctions linked to late Pliocene-Pleistocene deep-sea circulation changes in the northern Indian Ocean (ODP Sites 722 and 758), Mar Micropaleontol, 58 (2006) 219-242.
18 Tchernia P, Descriptive regional oceanography, (Pergamon Press, New York), 1980, pp. 253.
19 Vincent E, Cenozoic planktonic biostratigraphy and paleoceanography of the tropical western Indian Ocean, Initial Rep Deep Sea Drill Proj, 24 (1974) 1111-1150.
20 Kolla V, Sullivan L, Streeter S S & Langseth M G, Spreading of Antarctic bottom-water and its effects on the floor of the Indian Ocean inferred from bottom-water potential temperature, turbidity, and sea-floor photography, Mar Geol, 21 (1976) 171-189.
21 Wyrtki K, Oceanographic atlas of the International Indian Ocean Expedition (A A Balkemen, Rotterdam, Netherlands), 1988.
22 GEOSECS, India Ocean Expedition, Hydrographic data, 1977-1978, Vol. 5 (United States Government Printing Office, Washington, DC), 1983, pp. 1-48.
23 Peirce J, Weissel J, Taylor E, Dehn J, Driscoll N, Farrell J, Fourtanier E, Frey F A, Gamson P, Gee J, Gibson I L, Janecek T, Klootwijk C, Lawrence J R, Litke R, Newman J S, Nomura R, Owen R M, Pospichal J J, Rea D K, Resiwati P, Saunders A D, Smit J, Smith G M, Tamaki K, Weis D & Wilkinson C, Proc. ODP, Sci. Program, Initial Reports, 121 (1989) 359-453.
24 Sclater J G, Meinke L, Bennett A & Murphy C, The depth of the ocean through the Neogene. In: The Miocene ocean, edited by Kennett, J P, Geol Soc Am Mem, 163 (1985) 1-19.
25 Srinivasan M S & Gupta A K, Miocene benthic foraminifera and paleoceanography of DSDP Sites along Ninetyeast Ridge, northern Indian Ocean, J. Palaeontol. Soc. India, 35 (1990) 61-72.
26 Berggren W A, Kent D V, Swisher III C C & Aubry M–P, A revised Cenozoic geochronology and chronostratigraphy, In: Geochronology timescale and global stratigraphic correlation, edited by Berggren W A, Kent D V, Aubry M-P & Hardenbol J, SEPM (Society for Sedimentary Geology) Special Publication, 54 (AAPG, USA), 1995, pp. 129-212.
27 SAS Institute Inc., SAS/STAT users’ guide, Release 6.03 edition, Cary N C, SAS Institute Inc. 1988, pp.1-1003.
28 Parker W C & Arnold A J, Quantitative methods of data analysis in foraminiferal ecology, In: Modern foraminifera, edited by Sen Gupta B K, (Kluwer Academic Publishers, Great Britan), 1999, pp. 71-89.
29 Burke S C, Berger W H, Coulbourn W T & Vincent E, Benthic foraminifera in Box Core ERDC 112, Ontong Java Plateau, J Foraminiferal Res, 23 (1993) 19-39.
30 Almogi-Labin A, Schmiedl G, Hemleben C, Siman-tov R, Segl M & Meischner D, The influence of the NE winter monsoon on productivity changes in the Gulf of Aden, NW Arabian Sea, during the last 530 ka as recorded by foraminifera, Mar Micropaleontol, 40 (2000) 295-319.
31 Gupta A K, Paleoceanographic and paleoclimatic history of the Somali Basin during the Pliocene-Pleistocene: Multivariate analyses of benthic foraminifera from DSDP Site 241 (leg 25), J Foraminiferal Res, 27 (1997) 196-208.
32 Gupta A K, Sarkar S & Mukherjee B, Paleoceanographic changes during the past 1.9 Myr at DSDP Site 238, Central Indian Ocean Basin: Benthic foraminiferal proxies, Mar Micropaleontol, 60 (2006) 157-166.
33 Gupta A K, Das M & Bhaskar K, South Equatorial Current (SEC) driven changes at DSDP Site 237, Central Indian Ocean, during the Plio-Pleistocene: Evidence from benthic foraminifera and stable isotopes, J Asian Earth Sci, 28 (2006) 276-290.

INDIAN J. MAR. SCI., VOL. 36, NO. 4, DECEMBER 2007
340
34 Corliss B H & Chen C, Morphotype patterns of Norwegian Sea deep-sea benthic foraminifera and ecological implications, Geology, 16 (1988) 716-719.
35 Danne R A & Sen Gupta B K, Association of bathyal foraminifera with water masses in the northwestern Gulf of Mexico, Mar Micropaleontol, 17 (1991) 173-193.
36 Barmawidjaja D M, Jorissen F J, Puskaric S & van der Zwaan G J, Microhabitat selection by benthic foraminifera in the northern Adriatic Sea, J Foraminiferal Res, 22 (1992) 297-317.
37 Rathburn A E & Corliss B H, The ecology of living (stained) benthic foraminifera from the Sulu Sea, Paleoceanography, 9 (1994) 87-150.
38 Lutze G F & Coulbourn W T, Recent benthic foraminifera from the continental margin of Northwest Africa: community structure and distribution, Mar Micropaleontol, 8 (1984) 361-401.
39 Woodruff F, Changes in Miocene deep-sea benthic foraminiferal distribution in the Pacific Ocean: relationship to paleoceanography, In: The Miocene ocean, edited by Kennett J P, Geol Soc Am Mem, 163 (1985) 131-176.
40 Gupta A K & Srinivasan M S, Response of Northern Indian Ocean deep-sea benthic foraminifera to global climate change during Pliocene-Pleistocene, Mar Micropaleontol, 16 (1990) 77-91.
41 Lohmann G P, Abyssal benthonic foraminifera as hydrographic indicators in the western South Atlantic Ocean, J Foraminiferal Res, 8 (1978) 6-34.
42 Schnitker D, Quaternary deep-sea benthic foraminifers and water masses, Ann. Rev. Earth Planet. Sci., 8 (1980) 343-370.
43 Lutze G F & Thiel H, Epibenthic foraminifera from elevated microhabitats: Cibicidoides wuellerstorfi and Planulina ariminensis, J Foraminiferal Res, 19 (1989) 153-158.
44 Linke P & Lutze G F, Microhabitat preferences of benthic foraminifera-a static concept or a dynamic adaptation to optimise food acquisition? Mar Micropaleontol, 20 (1993) 215-234.
45 Loubere P & Fariduddin M, Quantitative estimates of global patterns of surface ocean biological productivity and its seasonal variation on time scales from centuries to millennia, Glob. Biogeochem. Cyc., 13 (1999) 115-133.
46 Corliss B H, Recent deep-sea benthonic foraminiferal distributions in the southeast Indian Ocean: Inferred bottom water routes and ecological implications, Mar Geol, 31 (1979) 115-138.
47 Corliss B H, Distribution of Holocene deep-sea benthonic foraminifera in the southwest Indian Ocean, Deep-Sea Res, 30 (1983) 95-117.
48 Gooday A J, A response by benthic foraminifera to the deposition of phytodetritus in the deep sea, Nature, 332 (1988) 70-73.
49 Gooday A J & Turley C M, Response by benthic organisms to inputs of organic material to the Ocean floor: a review, Phil. Trans. Roy. Soc. London, A 331 (1990) 119-138.
50 Kaiho K, Effect of organic carbon flux and dissolved oxygen on the benthic foraminiferal oxygen index (BFOI), Mar Micropaleontol, 37 (1999) 67-76.
51 Gupta A K, Taxonomy and bathymetric distribution of Holocene deep-sea benthic foraminifera in the Indian Ocean and the Red Sea, Micropaleontology, 40 (1994) 351-367.
52 Fariduddin M & Loubere P, The surface ocean productivity response of deep water benthic foraminifera in the Atlantic Ocean, Mar Micropaleontol, 32 (1997) 289-310.
53 De Rijk S, Troelstra S R & Rohling E J, Benthic foraminiferal distribution in the Mediterranean Sea, J Foraminiferal Res, 29 (1999) 93-103.
54 Gupta A K & Thomas E, Initiation of Northern Hemisphere glaciation and strengthening of the northeast Indian monsoon: Ocean Drilling Program Site 758, eastern equatorial Indian Ocean, Geology, 31 (2003) 47-50.
55 Kahio K, Eocene to Quaternary benthic foraminifers and paleobathymentry of the Izu-Bonin Arc, Legs 125 and 126. In: Proce Ocean Drill Prog, Sci Res, 126 (1992), 285-310.
56 Hess S & Kuhnt W, Neogene and Quaternary paleoceanographic changes in the southern South China Sea (Site 1143): the benthic foraminiferal record, Mar Micropaleontol, 54 (2005) 63-87.
57 Loubere P, The surface ocean productivity and bottom water oxygen signals in deep water benthic foraminiferal assemblages, Mar Micropaleontol, 28 (1996) 247-261.
58 Burke S C, Recent benthic foraminifera of the Ontong Java Plateau, J Foraminiferal Res, 11 (1981) 1-19.
59 Hermelin J O R & Shimmield G B, The importance of the oxygen minimum zone and sediment geochemistry in the distribution of recent benthic foraminifera in the northwest Indian Ocean, Mar Geol, 91 (1990) 1-29.
60 Mullineaux L S & Lohmann G P, Late Quaternary stagnations of the eastern Mediterranean: changes in the deep water recorded by fossil benthic foraminifera, J. Foraminiferal Res, 11 (1981) 20-39.
61 Gupta A K, Neogene deep water benthic foraminifera from the northern Indian Ocean DSDP Sites (Leg 22), Ph. D. thesis, Banaras Hindu University, India, 1987.
62 Weinholz P & Lutze G F, The Stilostomella extinction, In: Proceedings of the Ocean Drilling Program, Scientific Results edited by Ruddiman W, Sarnthein M, et al., (Ocean Drilling Program, College Station, TX), 108 (1989) 113-117.
63 Gupta A K, Biostratigraphic vs. paleoceanographic importance of Stilostomella lepidula (Schwager) in the Indian Ocean, Micropaleontology, 39 (1993) 47-51.
64 Thomas D J, Bralower T J & Zachos J C, New evidence for subtropical warming during the late Paleocene thermal maximum: stable isotopes from Deep Sea Drilling Project 527, Walvis Ridge, Paleoceanography, 14 (1999) 561-570.
65 Hayward B W, Late Pliocene to middle Pleistocene extinctions of deep-sea benthic foraminifera (“Stilostomella extinction”) in the southwest Pacific, J. Foraminiferal Res, 32 (2002) 274-307.
66 Gupta A K, Dhingra H, Melice J L & Anderson D M, Earth’s eccentricity cycles and Indian summer monsoon variability over the past 2 million years: evidence from deep-sea benthic foraminifer, Geophys. Res. Letts., 28 (2001) 4131-4134.
67 Kennett J P & Barker P F, Latest Cretaceous to Cenozoic climate and oceanographic developments in the Weddell Sea,

BHAUMIK et al.: FORAMINIFERA OF DSDP HOLE 216A
341
Antarctica: an Ocean-Drilling Perspective, Proc Ocean Drill Prog, Sci Results, 113, (1990) 937-960.
68 Savin S M, Douglas R G et al., Miocene benthic foraminiferal isotope records: a synthesis, Mar Micropaleontol, 6 (1981).
69 Keller G & Barron J A, Paleoceanographic implications of Miocene deep-sea hiatuses, Geol Soc Am Bull, 94 (1983) 590-613.
70 Kennett J P, The Miocene Ocean: Paleoceanography and Biogeography, Geol Soc Am Mem, 163 (1985) 1-337.
71 Shackleton N J & Backman J, Oxygen isotope calibration of ice rafting in DSDP Site 552 A: History of glaciation in the North Atlantic region, Nature, 307 (1984) 620-623.
72 Kennett J P & von der borch C, Southwest Pacific Cenozoic Paleoceanography, Initial Rep DSDP, 90 (1986), 1493-1517.
73 Kennett J P, Cenozoic evolution of Antarctic glaciation, the circum-Antarctic Ocean, and their impact on global paleoceanography, J Geophys Res, 82 (1977) 3843-3860.
74 Zachos J, Pagani M, Sloan L, Thomas E, & Billups K, Trends, Rhythms, and Aberrations in Global Climate 65 Ma to Present, Science, 292 (2001) 686-693.
75 Kroon D, Steens T N F & Troelstra S R, Onset of monsoonal related upwelling in the western Arabian sea as revealed by planktonic foraminifer, Proc Ocean Drill Prog Sci Res, 117 (1991) 257-263.




















