
Boron isotopes and B/Ca in benthic foraminifera: Proxies for the deep ocean carbonate system
11
Boron isotopes and B/Ca in benthic foraminifera: Proxies for the deep ocean carbonate system James W.B. Rae a,b, ⁎, Gavin L. Foster a,b,1 , Daniela N. Schmidt b , Tim Elliott a,b a Bristol Isotope Group, United Kingdom b Department of Earth Sciences, University of Bristol, Wills Memorial Building, Queen's Rd, Bristol, BS8 1RJ, United Kingdom abstract article info Article history: Received 16 July 2010 Received in revised form 14 December 2010 Accepted 15 December 2010 Available online 15 January 2011 Editor: P. DeMenocal Keywords: boron isotopes B/Ca foraminifera pH proxy pore water Accurate records of the state of the ocean carbonate system are critical for understanding past changes in pCO 2 , ocean acidification and climate. The chemical principles underlying the proxy of oceanic pH provided by the boron isotope ratio of foraminiferal carbonate are relatively well understood, but the proxy's reliability has been questioned. We present 76 new Multi-Collector Inductively-Coupled Plasma Mass Spectrometry (MC-ICPMS) δ 11 B measurements on a range of benthic foraminifera from 23 late-Holocene samples from the Atlantic that reaffirm the utility of the δ 11 B-pH proxy. Our boron isotope measurements on ~ 10 benthic foraminifera tests typically yield a precision of ~ ± 0.25‰ at 2 s.d. (equivalent to ~ ± 0.03 pH units). δ 11 B values of epifaunal species are within analytical uncertainty of those predicted from a simple model assuming sole incorporation of B(OH) 4 − from seawater and no vital effects, using the independently determined fractionation factor of 1.0272 between 11 B/ 10 B of aqueous boron species. Infaunal foraminifera are consistent with this model, but record the combined effects of lower pore-water δ 11 B and pH. No influence of partial dissolution or shell size on δ 11 B is observed. We have also measured the B/Ca ratios of the same samples. For individual Cibicidoides species, B/Ca shows a good correlation with Δ[CO 3 2− ], but the B/Ca of different co- occurring species morphotypes varies considerably. These effects are not seen in δ 11 B, which may therefore provide a more robust proxy of the ocean carbonate system. Whilst in theory δ 11 B and B/Ca can be combined to provide a quantitative reconstruction of alkalinity and dissolved inorganic carbonate (DIC), in practice this is precluded by propagated uncertainties. δ 11 B data give significant constraints on foraminifera calcification mechanisms, and seem most simply explained by incorporation of B(OH) 4 − into a HCO 3 − pool, which is then completely incorporated in foraminiferal CaCO 3 . Our demonstration of the predictable variation of δ 11 B with pH, across a wide range of species and locations, provides confidence in the application of MC-ICPMS measurements of foraminiferal δ 11 B to reconstruct past changes in the ocean carbonate system. © 2010 Elsevier B.V. All rights reserved. 1. Introduction The boron isotope pH proxy is a potentially powerful tool for reconstructing changes in the ocean's carbon cycle. The basis of the proxy is firmly grounded in inorganic chemistry, but its application has attracted some controversy due to differences between measured boron isotope values and those expected from theory (Pagani et al., 2005). 1.1. The boron isotope pH proxy Boron exists mainly in two molecular species in the ocean: boric acid (B(OH) 3 , trigonal) and borate ion (B(OH) 4 − , tetrahedral), related by the acid–base equilibrium: B OH ð Þ 3 +2H 2 O ⇌ B OH ð Þ − 4 +H 3 O þ : ð1Þ The stoichiometric equilibrium constant of reaction 1, K B ⁎ , is 10 −8.6 (or pK B ⁎ = 8.6) in seawater at 25 °C, 35 salinity and atmospheric pressure (Dickson, 1990). As pK B ⁎ is close to typical ocean pH the proportions of B(OH) 3 and B(OH) 4 − vary significantly with changes in ocean pH (Fig. 1a). Earth and Planetary Science Letters 302 (2011) 403–413 ⁎ Corresponding author. Department of Earth Sciences, University of Bristol, Wills Memorial Building, Queen's Rd, Bristol, BS8 1RJ, United Kingdom. E-mail address: [email protected] (J.W.B. Rae). 1 Present address: School of Ocean and Earth Sciences, National Oceanography Centre, Southampton, University of Southampton, European Way, Southampton, SO14 3ZH, United Kingdom. 0012-821X/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.epsl.2010.12.034 Contents lists available at ScienceDirect Earth and Planetary Science Letters journal homepage: www.elsevier.com/locate/epsl
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Boron isotopes and B/Ca in benthic foraminifera: Proxies for the deep ocean carbonate system

Earth and Planetary Science Letters 302 (2011) 403–413
Contents lists available at ScienceDirect
Earth and Planetary Science Letters
j ourna l homepage: www.e lsev ie r.com/ locate /eps l
Boron isotopes and B/Ca in benthic foraminifera: Proxies for the deep oceancarbonate system
James W.B. Rae a,b,⁎, Gavin L. Foster a,b,1, Daniela N. Schmidt b, Tim Elliott a,b
a Bristol Isotope Group, United Kingdomb Department of Earth Sciences, University of Bristol, Wills Memorial Building, Queen's Rd, Bristol, BS8 1RJ, United Kingdom
⁎ Corresponding author. Department of Earth SciencMemorial Building, Queen's Rd, Bristol, BS8 1RJ, United
E-mail address: [email protected] (J.W.B. Rae)1 Present address: School of Ocean and Earth Scie
Centre, Southampton, University of Southampton, Europ3ZH, United Kingdom.
0012-821X/$ – see front matter © 2010 Elsevier B.V. Adoi:10.1016/j.epsl.2010.12.034
a b s t r a c t
a r t i c l e i n f oArticle history:Received 16 July 2010Received in revised form 14 December 2010Accepted 15 December 2010Available online 15 January 2011
Editor: P. DeMenocal
Keywords:boron isotopesB/CaforaminiferapHproxypore water
Accurate records of the state of the ocean carbonate system are critical for understanding past changes inpCO2, ocean acidification and climate. The chemical principles underlying the proxy of oceanic pH provided bythe boron isotope ratio of foraminiferal carbonate are relatively well understood, but the proxy's reliabilityhas been questioned. We present 76 new Multi-Collector Inductively-Coupled Plasma Mass Spectrometry(MC-ICPMS) δ11B measurements on a range of benthic foraminifera from 23 late-Holocene samples from theAtlantic that reaffirm the utility of the δ11B-pH proxy. Our boron isotope measurements on ~10 benthicforaminifera tests typically yield a precision of ~±0.25‰ at 2 s.d. (equivalent to ~±0.03 pH units). δ11Bvalues of epifaunal species are within analytical uncertainty of those predicted from a simple model assumingsole incorporation of B(OH)4− from seawater and no vital effects, using the independently determinedfractionation factor of 1.0272 between 11B/10B of aqueous boron species. Infaunal foraminifera are consistentwith this model, but record the combined effects of lower pore-water δ11B and pH. No influence of partialdissolution or shell size on δ11B is observed. We have also measured the B/Ca ratios of the same samples. Forindividual Cibicidoides species, B/Ca shows a good correlation with Δ[CO3
2−], but the B/Ca of different co-occurring species morphotypes varies considerably. These effects are not seen in δ11B, which may thereforeprovide a more robust proxy of the ocean carbonate system. Whilst in theory δ11B and B/Ca can be combinedto provide a quantitative reconstruction of alkalinity and dissolved inorganic carbonate (DIC), in practice thisis precluded by propagated uncertainties. δ11B data give significant constraints on foraminifera calcificationmechanisms, and seem most simply explained by incorporation of B(OH)4
− into a HCO3− pool, which is then
completely incorporated in foraminiferal CaCO3. Our demonstration of the predictable variation of δ11B withpH, across a wide range of species and locations, provides confidence in the application of MC-ICPMSmeasurements of foraminiferal δ11B to reconstruct past changes in the ocean carbonate system.
es, University of Bristol, WillsKingdom..nces, National Oceanographyean Way, Southampton, SO14
ll rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
The boron isotope pH proxy is a potentially powerful tool forreconstructing changes in the ocean's carbon cycle. The basis of theproxy is firmly grounded in inorganic chemistry, but its applicationhas attracted some controversy due to differences between measuredboron isotope values and those expected from theory (Pagani et al.,2005).
1.1. The boron isotope pH proxy
Boron exists mainly in two molecular species in the ocean: boricacid (B(OH)3, trigonal) and borate ion (B(OH)4−, tetrahedral), relatedby the acid–base equilibrium:
B OHð Þ3+2H2O⇌ B OHð Þ−4 +H3Oþ: ð1Þ
The stoichiometric equilibrium constant of reaction 1, KB⁎, is 10−8.6
(or pKB⁎=8.6) in seawater at 25 °C, 35 salinity and atmospheric
pressure (Dickson, 1990). As pKB⁎ is close to typical ocean pH the
proportions of B(OH)3 and B(OH)4− vary significantly with changes inocean pH (Fig. 1a).
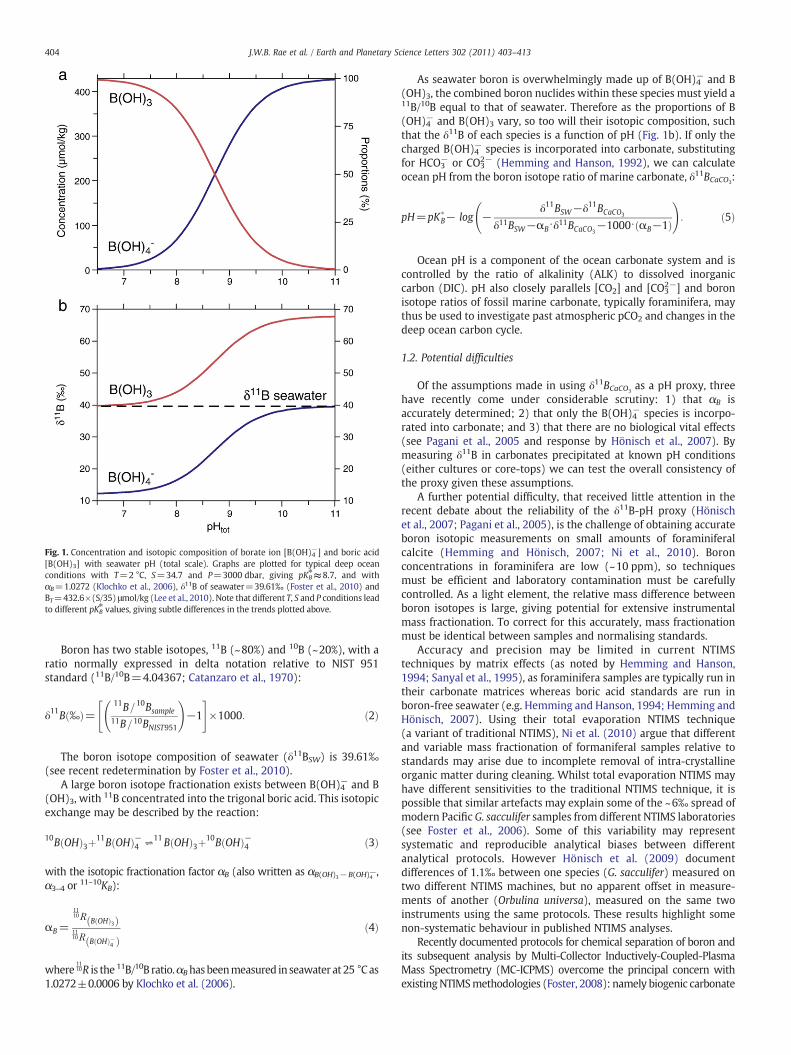
Fig. 1. Concentration and isotopic composition of borate ion [B(OH)4−] and boric acid[B(OH)3] with seawater pH (total scale). Graphs are plotted for typical deep oceanconditions with T=2 °C, S=34.7 and P=3000 dbar, giving pKB
⁎≈8.7, and withαB=1.0272 (Klochko et al., 2006), δ11B of seawater=39.61‰ (Foster et al., 2010) andBT=432.6×(S/35) μmol/kg (Lee et al., 2010). Note that different T, S and P conditions leadto different pKB
⁎ values, giving subtle differences in the trends plotted above.
404 J.W.B. Rae et al. / Earth and Planetary Science Letters 302 (2011) 403–413
Boron has two stable isotopes, 11B (~80%) and 10B (~20%), with aratio normally expressed in delta notation relative to NIST 951standard (11B/10B=4.04367; Catanzaro et al., 1970):
δ11B ‰ð Þ=11B=10Bsample11B=10BNIST951
!−1
" #×1000: ð2Þ
The boron isotope composition of seawater (δ11BSW) is 39.61‰(see recent redetermination by Foster et al., 2010).
A large boron isotope fractionation exists between B(OH)4− and B(OH)3, with 11B concentrated into the trigonal boric acid. This isotopicexchange may be described by the reaction:
10B OHð Þ3þ11B OHð Þ−4 ⇌11B OHð Þ3þ10B OHð Þ−4 ð3Þ
with the isotopic fractionation factor αB (also written as αB(OH)3−B(OH)4−,
α3–4 or 11–10KB):
αB=
1110R B OHð Þ3ð Þ1110R B OHð Þ−4ð Þ
ð4Þ
where1110R is the 11B/10B ratio.αBhasbeenmeasured in seawater at 25 °Cas
1.0272±0.0006 by Klochko et al. (2006).
As seawater boron is overwhelmingly made up of B(OH)4− and B(OH)3, the combined boron nuclides within these species must yield a11B/10B equal to that of seawater. Therefore as the proportions of B(OH)4− and B(OH)3 vary, so too will their isotopic composition, suchthat the δ11B of each species is a function of pH (Fig. 1b). If only thecharged B(OH)4− species is incorporated into carbonate, substitutingfor HCO3
− or CO32− (Hemming and Hanson, 1992), we can calculate
ocean pH from the boron isotope ratio of marine carbonate, δ11BCaCO3:
pH=pK�B− log −
δ11BSW−δ11BCaCO3
δ11BSW−αB⋅δ11BCaCO3−1000⋅ αB−1ð Þ
!: ð5Þ
Ocean pH is a component of the ocean carbonate system and iscontrolled by the ratio of alkalinity (ALK) to dissolved inorganiccarbon (DIC). pH also closely parallels [CO2] and [CO3
2−] and boronisotope ratios of fossil marine carbonate, typically foraminifera, maythus be used to investigate past atmospheric pCO2 and changes in thedeep ocean carbon cycle.
1.2. Potential difficulties
Of the assumptions made in using δ11BCaCO3as a pH proxy, three
have recently come under considerable scrutiny: 1) that αB isaccurately determined; 2) that only the B(OH)4− species is incorpo-rated into carbonate; and 3) that there are no biological vital effects(see Pagani et al., 2005 and response by Hönisch et al., 2007). Bymeasuring δ11B in carbonates precipitated at known pH conditions(either cultures or core-tops) we can test the overall consistency ofthe proxy given these assumptions.
A further potential difficulty, that received little attention in therecent debate about the reliability of the δ11B-pH proxy (Hönischet al., 2007; Pagani et al., 2005), is the challenge of obtaining accurateboron isotopic measurements on small amounts of foraminiferalcalcite (Hemming and Hönisch, 2007; Ni et al., 2010). Boronconcentrations in foraminifera are low (~10 ppm), so techniquesmust be efficient and laboratory contamination must be carefullycontrolled. As a light element, the relative mass difference betweenboron isotopes is large, giving potential for extensive instrumentalmass fractionation. To correct for this accurately, mass fractionationmust be identical between samples and normalising standards.
Accuracy and precision may be limited in current NTIMStechniques by matrix effects (as noted by Hemming and Hanson,1994; Sanyal et al., 1995), as foraminifera samples are typically run intheir carbonate matrices whereas boric acid standards are run inboron-free seawater (e.g. Hemming and Hanson, 1994; Hemming andHönisch, 2007). Using their total evaporation NTIMS technique(a variant of traditional NTIMS), Ni et al. (2010) argue that differentand variable mass fractionation of formaniferal samples relative tostandards may arise due to incomplete removal of intra-crystallineorganic matter during cleaning. Whilst total evaporation NTIMS mayhave different sensitivities to the traditional NTIMS technique, it ispossible that similar artefacts may explain some of the ~6‰ spread ofmodern Pacific G. sacculifer samples from different NTIMS laboratories(see Foster et al., 2006). Some of this variability may representsystematic and reproducible analytical biases between differentanalytical protocols. However Hönisch et al. (2009) documentdifferences of 1.1‰ between one species (G. sacculifer) measured ontwo different NTIMS machines, but no apparent offset in measure-ments of another (Orbulina universa), measured on the same twoinstruments using the same protocols. These results highlight somenon-systematic behaviour in published NTIMS analyses.
Recently documented protocols for chemical separation of boron andits subsequent analysis by Multi-Collector Inductively-Coupled-PlasmaMass Spectrometry (MC-ICPMS) overcome the principal concern withexistingNTIMSmethodologies (Foster, 2008): namely biogenic carbonate

405J.W.B. Rae et al. / Earth and Planetary Science Letters 302 (2011) 403–413
matrix is removed from samples by ion chromatography, so samples andstandards are run under very similar conditions. Although machine-induced mass fractionation is significant (~15% per amu), sample-standard bracketing is rapid, accurately captures machine variability,and thus corrects for this fractionation. The analytical reliability of ourMC-ICPMS technique is supported by standard addition experiments(Ni et al., 2010), and our reproducibility has been substantiated byextensive repeat analyses of boric acids and total procedural replicates ofin-house carbonate standards (see Supplementary Fig. 1).
Given the previously documented accuracy of our MC-ICPMStechnique, we can re-examine the boron isotope-pH proxy bycomparison of δ11B measurements in core-top benthic foraminiferato carbonate system data from nearby hydrographic sites. Theepifaunal benthic foraminifera analysed have a generally stablehabitat and lack symbionts, allowing proxy testing with fewerbiological and ecological complications than those associated withplanktic species (Hönisch and Hemming, 2004; Hönisch et al., 2003).If our understanding of the δ11B-pH proxy is correct, measured δ11B inmodern epifaunal foraminifera should closely match the calculatedδ11BB(OH)4− in seawater.
2. Materials and methods
2.1. Samples
Late Holocene samples have been taken from 23 cores spanning376–4427 m and 58 °N–48 °S in the Atlantic Ocean (Fig. 2), focussingon three depth transects, at the Canary Islands, the Namibian Coastand Walvis Ridge. Exact core locations and the cruises on which theywere collected are listed in the Supplementary information. Corematerial was washed and sieved N63 μm with deionised water.Foraminifera were picked in discrete size-fractions in the CanaryIsland samples (200–250, 250–300, 300–350, 350–425, N425 μm totest for any shell size effect) and N200 μm elsewhere. Sample sizesrange from 5 to 75 foraminifera (0.35–3 mg CaCO3), with an averageof ~15 foraminifera giving ~1 mg CaCO3 and ~10 ng boron. A largerange of common hylaine benthic foraminifera species have beenexamined. Epifaunal species include Cibicidoides wuellerstorfi, Cibici-doides mundulus and Planulina ariminensis, which have been shown tolive in epifaunal or elevated epifaunal environments (Lutze and Thiel,1989) and are commonly used to reconstruct bottomwater chemistry(e.g. Hillaire-Marcel and De Vernal, 2007). A range of otherCibicidoides have also been studied, and several infaunal species,
Fig. 2. Locations of core samples on an Atlantic section of calculated δ11B of B(OH)4−, withoceanographic conditions in the Atlantic, spanning pH 7.87–8.07 and 2–13 °C, with three dcores from the high North Atlantic and Southern Ocean.
including Oridorsalis umbonatus, Uvigerina peregrina, Gyroidina solda-nii, Lenticulina vortex, Ammonia beccarii and Melonis zaandamae. Thearagonitic benthic foraminifer, Hoeglundina elegans, has also beenanalysed. In addition four intervals were picked from the top 35 cm ofcore PS2495 to compare relative offsets between different co-occurring Cibicidoides species morphotypes at the same site.
Pore waters were sampled at cm intervals from three multicoresfrom the Namibian Coast using Rhizons, thus filtering the samples tob0.1 μm (Seeberg-Elverfeldt et al., 2005). pH was analysed on thesesamples using pH electrodes and “NBS” buffers, and alkalinitydetermined by titration (Zabel, unpublished data).
2.2. Hydrographic data
Hydrographic data used in this study is taken from nearby GLODAPsites (Key et al., 2004) located using Ocean Data View (Schlitzer,2010). The nearest two profiles to the Canary Island samples havebeen combined to improve depth resolution. Carbonate systemparameters are calculated from alkalinity and total dissolvedinorganic carbon (DIC) using a modified version of CO2SYS-MATLAB(van Heuven et al., 2009). DIC is corrected for anthropogenic CO2
(Sabine et al., 2004) at the Canary Island and planktonic sites of Foster(2008); this data was not available for the Walvis or Namibianprofiles, but comparison of these sites to the nearest locations inwhich anthropogenic influence has been assessed suggests minimalperturbation at the depths of interest. Carbonate calculations includethe effects of temperature, salinity, pressure and phosphate andsilicate alkalinity. Formulations are those recommended by Dickson etal. (2007) (including the Lueker et al., 2000 refit of the K1 and K2carbonic acid dissociation constants of Mehrbach et al., 1973), exceptthe total boron concentration of seawater, for which we use therecently redetermined value of Lee et al. (2010) (BT=432.6×(S/35)μmol/kg-sw). Temperature and pressure independent concentrationunits of mol/kg-sw are used throughout (Zeebe and Wolf-Gladow,2001).
δ11B of B(OH)4− is calculated using
δ11BB OHð Þ−4 =δ11BSW⋅BT−1000⋅ αB−1ð Þ⋅ B OHð Þ3
� �B OHð Þ−4� �
+αB⋅ B OHð Þ3� � ð6Þ
(see Zeebe and Wolf-Gladow, 2001) with [B(OH)3] and [B(OH)4−]determined as in CO2SYS, using KB
⁎ of Dickson (1990) with thepressure correction of Culberson and Pytkowicz (1968) (as refit by
pH contours, calculated from GLODAP data. Samples are taken from a large range ofepth transects at the Canary Islands, Namibian Coast and Walvis Ridge, and individual

406 J.W.B. Rae et al. / Earth and Planetary Science Letters 302 (2011) 403–413
Millero, 1979 and neglecting the salinity term). The constants forcarbonate system pressure corrections have often been misquotedand are given in full in the Supplementary information. Uncertaintyon the calculated δ11BB(OH)4− was assessed using the statistics featureof Ocean Data View (Schlitzer, 2010), and is given a representative2 s.d. value of 0.25‰, equivalent to pH uncertainty of 0.032 for benthicconditions and 0.020 for planktic.
2.3. Analytical techniques
Analytical techniques used follow Foster (2008), with oxidativecleaning developed from Barker et al. (2003). Foraminifera samplesare crushed between clean glass slides and transferred to clean Tefloncentrifuge tubes. Clay is then removed with repeat rinses of MilliQN18.2 MΩ/cm water and ultrasonication. Organic matter is oxidisedwith 1% hydrogen peroxide (buffered with 0.1 M ammoniumhydroxide) at 80 °C. Samples are then treated with 0.0005 M HNO3
to remove any adsorbed contaminants, and dissolved in 330–1000 μlof 0.075 M HNO3 depending on sample size. Samples are centrifugedimmediately after dissolution and carefully transferred to screw topTeflon vials for storage, leaving a residual ~10 μl, which helps excludeany remaining undissolved contaminants. All HNO3 is Teflon-distilledwith manitol, a boron-free MilliQ package is used to generate cleanwater, and work is carried out in a dedicated flow hood within anover-pressured clean lab that uses boron-free HEPA filters.
A small aliquot (b5%) of the dissolved sample is analysed for traceelements (Ca, Mg, B, Al, Mn, Fe, Ba, Sr, Li, Na, Cd, U, Nd) on a ThermoFinnigan Element 2 ICP-MS, matching sample and standard Ca concen-tration (typically 1 mmol/l) and using ammonia gas to improve boronwashout (Al-Ammar et al., 2000). B/Ca ratios obtained in this manner areaccurate and precise to ±5% at 2 s.d., as determined by isotope dilutionanalyses and evidenced by the reproducibility of a range of in-houseconsistency standards (Ni et al., 2007).
Prior to boron isotope analysis, boron is separated from the samples'carbonate matrix using the boron-specific anionic exchange resinAmberlite IRA 743 (Kiss, 1988) in a 20 μl micro-column. Samples areloaded in a 2 M sodium acetate–0.5 M acetic acid buffer, rinsed withMilliQ water, and collected in 450 μl of 0.5 M HNO3. Column yields,checked by isotope dilution, are N95% (Foster, 2008) and the elution tailof every sample is checked with an extra acid rinse to ensure nosignificant amount of sample boron remains. In all cases this tailrepresents b1% of the boron loaded. Boron concentrations and possiblecontamination from the Na-rich buffer are checked with a 20 μl aliquotprior to analysis. b5% of samples are influenced by the latter process,which likely results from imperfect removal of buffer from the columnprior to boron collection. As reported by Foster (2008, Supplementaryinformation), these samples show anomalous mass fractionationbehaviour and are rejected. Depending on foraminiferal sample sizeand boron concentration, boron isotope samples range from 2 to 20 ng,yielding solutions for analysis of ~5–50 ppb (ng/g).
Boron isotope analyses are performed on a Thermo Finnigan NeptuneMC-ICPMS. Before sample analysis, the instrument is tuned andoptimisedfor maximum 11B/10B stability (following Foster, 2008). Solutions areintroduced using a Teflon barrel spray chamber, and 2–3 ml/min ofammonia gas is added to improvewashout (Al-Ammar et al., 2000; Foster,2008). Exposure to the laboratory atmosphere of samples in open vials inthe autosampler can cause significant blank contamination (10 pg/h). Tomonitor and correct for this accumulatingblank, vials containing the samevolumeof0.5 MHNO3as thesamplesareanalysedevery secondsample.Atotal procedural blank (TPB) is measured with every column batch,averaging ~100 pg (~1% of sample size) with a δ11B of 5‰. A TPBcorrection is made using this average isotopic composition and themeasured amount of blank B from each batch of chemistry, giving a TPBcorrectionon δ11Bb0.2‰. Samples arebracketedwith a50 ppb solutionofNIST SRM 951 boric acid standard, which is used to correct for machine-inducedmass-fractionation, and converts 11B/10B ratios to delta notation.
We find no need to match the standard to the variable sampleconcentrations within the measured range of ~5–50 ppb (Foster, 2008).Each sample is measured twice and the mean δ11B value is used.
Reproducibility has been assessed by repeat analysis of in-house boricacid and coral standards at a range of concentrations (see SupplementaryFig. 1). These data give a relationship betweenmeasured sample intensity([11B] in volts) and external reproducibility (at 2 standarddeviations) thatis well fitted by a two-term exponential relationship
2 s:d:=1:7⋅ exp−29½11B�+0:31⋅ exp−0:75½11B� ð7Þ
which we use to assess sample δ11B uncertainty (2 s.d. is typically0.23‰ at 50 ppb and 0.33‰ at 10 ppb). An in-house coral standard isalso run with each batch of column chemistry as a further check onreproducibility.
Pore water samples were diluted with 0.5 M HNO3 to check Namatrix concentrations, then analysed for trace element concentra-tions (B, Mg, Ca, Sr), matching Na concentrations between samplesand bracketing standards. Column chemistry was carried out in anidentical fashion to carbonate samples, using 18 μl of pore water toobtain samples of ~40 ng B, which were then diluted andmeasured at50 ppb.
3. Results
3.1. Foraminiferal δ11B
Boron isotope ratios were measured on 79 benthic foraminiferasamples. Of these 3 were discarded due to high Na content aftercolumn chemistry, indicating buffer contamination. Measured δ11Bvalues in the epifaunal species C. wuellerstorfi, C. mundulus and P.ariminensis range from 14.5 to 16.8‰ and show a close match to thecalculated δ11B of their local seawater B(OH)4− (Figs. 3 and 4). Thereare no systematic δ11B offsets between different epifaunal species andno significant changes in δ11B with foraminiferal test size (seeSupplementary Fig. 2). There is also no apparent offset of samplesshowing visual signs of partial dissolution from the rest of the dataset.Infaunal foraminifera typically have lower δ11B than epifaunal speciesfrom the same core, though the magnitudes of these offsets are highlyvariable, ranging from ~0.2 to 2.5‰ (Fig. 3). The aragonitic foraminiferH. elegans has low and relatively invariant δ11B for each transect, withone exception at ~3000 m at Walvis Ridge. Similar offsets betweenepifaunal foraminifera and infaunal foraminifera and H. elegans werealso noted by Hönisch et al. (2008).
The fit of our δ11B data to expected δ11BB(OH)4− values, weighted for
uncertainties, can be quantified by the mean squared weighteddeviation (MSWD). MSWD of our epifaunal benthic data is 3.0, withindividual values of 1.9 for C. wuellerstorfi and 3.8 for C. mundulus,suggesting C. wuellerstorfi is the more reliable recorder of bottomwater conditions (in keeping with results from various other proxies,e.g. Mackensen et al., 1993). These MSWD values are slightly highrelative to the 95% confidence limit of 1.8 for sample sets the size ofour C. wuellerstorfi and C. mundulus data. Such minor scatter isperhaps not surprising, given the potential for variable conditions atthe sample location compared to bottomwater at the nearest GLODAPsite.
3.2. Foraminiferal trace elements
Epifaunal foraminifera typically have B/Ca ~130–230 μmol/kg(Fig. 5). B/Ca ratios in C. wuellerstorfi and C. mundulus show largevariations, correlated with the degree of carbonate ion saturation(Δ[CO3
2−]), in agreement with Yu and Elderfield (2007), where:
Δ½CO2−3 �=½CO2−
3 �in situ−½CO2−3 �sat ð8Þ

Canary Islands Namibian Coast Walvis Ridge
Dep
th (
m)
δ11B
C. wuel. C. mund. Lenticulina H. elegansGyroidinaO. umb. UvigerinaPlanulina C. mund. var.
δ11BB(OH)4
-
0
500
1000
1500
2000
2500
3000
3500
4000
4500
0
500
1000
1500
2000
2500
3000
3500
13 14 15 16 17 18 13 14 15 16 17 1819 13 14 15 16 17 18 190
500
1000
1500
2000
2500
3000
3500
4000
11 12
δ11B δ11B
Fig. 3.Depth profiles showingmeasured δ11B of common infaunal and epifaunal foraminifera and calculated δ11B of B(OH)4− at nearby GLODAP sites. Grey bands indicate typical 2 s.d.uncertainty on δ11BB(OH)4− of 0.25‰. Single measurements exist on several other species — see Fig. 7 — but are not shown for clarity. Note that scales vary between profiles.
407J.W.B. Rae et al. / Earth and Planetary Science Letters 302 (2011) 403–413
½CO2−3 �sat= ½CO2−
3 �in situ
Ωcalciteð9Þ
and
Ωcalcite=½Ca2+ �⋅½CO2−
3 �Ksp
ð10Þ
where Ksp is the solubility constant of calcite (Zeebe and Wolf-Gladow, 2001). For consistency with Yu and Elderfield (2007) we usethe partial molar volume (ΔV) of Ingle (1975) for the pressuredependence of Ksp. Infaunal species' B/Ca values are lower(~10–80 μmol/kg) and less variable (Fig. 5). B/Ca in H. elegans isalso low and quite scattered, and has no significant correlation withΔ[CO3
2−]. In contrast with δ11B, absolute values and trends in B/Ca arestrongly species specific. These vital effects are significant even forvery subtle changes in species morphology from the same sample(Fig. 6).
All samples are screened for potential clay contamination usingmeasured Al/Ca ratios. These average 52 μmol/kg. 13 samples haveAl/Ca higher than 100 μmol/kg, but no correlation is seen betweenhigh Al/Ca and B/Ca or δ11B, or offsets of these values from those
15.8 16.0 16.2
δ11B fo
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
14.5 15.5
PlanulinaC. wuellerstorfiC. mundulus
G. ruberG. sacculifer
Δ δ11
B fo
ram
- s
w B
(OH
) 4-
Fig. 4. Offsets of measured δ11B in epifaunal benthic (this study) and shallow-dwelling plplotted against measured foraminiferal δ11B (note changing x axis scale). C. mundulus var. is inby the dashed lines. Planktic foraminifera data are from Foster (2008) and are offset above
predicted from core-top calibration (Yu and Elderfield, 2007) or thepH of local seawater (R2b0.1). Data on other trace elements (Li, Mg, Sretc.) is being prepared for a separate publication.
3.3. Pore-water δ11B and pH
Measured bottom water δ11B from the Namibian multicores of39.5–39.9‰ (depth zero in Fig. 7) is similar to open seawater (39.61‰Foster et al., 2010). Pore water values show a relatively steadydecrease to 38.65±0.15‰ at 5 cm depth (Fig. 7).
pH and alkalinity measurements were also made on the Namibiancores' pore waters at centimetre intervals (Zabel unpublished data),but may be compromised by changes in pressure between the oceanfloor and measurement at the surface. Although changes in closedsystem carbonate chemistry due to differences in temperature andpressure can be accounted for, the effects of CO2 outgassing andcarbonate precipitation (see Archer et al., 1989) cannot easily becorrected. Bottom water pH data from our multicores is offset fromnearby GLODAPmeasurements by up to ~−0.2 units (~−0.4 withoutclosed system temperature and pressure corrections), presumably asa result of carbonate precipitation, and it is likely that pore waters aresimilarly affected. The offset between GLODAP and “bottomwater” pHhas thus been applied to the pore-water pH data (Fig. 7c) but even
19.5 20.5-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
16.4 16.6
ram
anktic foraminifera (Foster, 2008) from calculated δ11BB(OH)4− at nearby GLODAP sites,dicated by a light green circle, as in Fig. 3. A 0.25‰ uncertainty in δ11BB(OH)4− is indicatedδ11BB(OH)4− by ~0.7‰, likely due to symbiont photosynthesis.

Fig. 5. B/Ca in core-top foraminifera plotted againstΔ[CO32−] from nearby GLODAP sites.
The core-top dataset of Yu and Elderfield (2007) is also shown in open symbols, alongwith Yu and Elderfield's (2007) trend lines for C. wuellerstorfi and C. mundulus. Of thethree C. wuellerstorfi samples lying outside the scatter of Yu and Elderfield's (2007)data, two show some variability from perfect C. wuellerstorfi morphotype and one hashigh Al/Ca (366 μmol/kg).
408 J.W.B. Rae et al. / Earth and Planetary Science Letters 302 (2011) 403–413
these values should be treated with caution, as further evidenced bythe unrealistic variability in some pH profiles.
4. Discussion
4.1. Comparison to NTIMS
Our results differ significantly from previously published (NTIMS)δ11B data from benthic foraminifera (Fig. 9). The mixed benthic
Fig. 6. a) δ11B from samples with two or more Cibicidoides species. Different Cibicidoides specmorphotypes from 4 depths in the same core. Arrow indicates ~40 μmol/mol difference beΔ[CO3
2−]. c) Examples of the Cibicidoidesmorphotypes and species plotted in b. C. wuellerstorslightly less curved chambers, a more rounded outline and a less pronounced final chamberesembling the shallow water species C. ungerianus.
measurements of Sanyal et al. (1995) lie above the calculated δ11BB
(OH)4− by ~5‰, while Hönisch et al.'s (2008) data from C. wuellerstorfi atWalvis Ridge are offset above our own by ~1.2‰.
The large discrepancy between the C. wuellerstorfi NTIMS data(Hönisch et al., 2008) and the mixed benthics of Sanyal et al. (1995)has been discussed by Hönisch et al. (2008). Sanyal et al. (1995)acknowledge that their data are unlikely to be accurate due todifferences in run conditions between samples and standards.Intriguingly, using the same techniques, G. sacculifer measured bySanyal et al. (1995) is only 1‰ heavier than that measured by Hönischand Hemming (2005), whereas the offset in δ11B in benthics betweenthe same respective authors is much larger. As noted by Hönisch et al.(2008), the use of a mixed benthic sample by Sanyal et al. (1995)should if anything lower the δ11B relative to pure C. wuellerstorfi data,and thus reduce the difference between these measurements. Therethus appear to be some unpredictable analytical biases betweendifferent NTIMS studies.
It is encouraging, however, to note that although the data ofHönisch et al. (2008) are offset from our own, this analytical offset isrelatively small and invariant over the limited δ11B range analysed(Fig. 9). Thus both techniques have the potential to record similarchanges in pH. Furthermore, Hönisch et al. (2008) find similar δ11Bdifferences between C. wuellerstorfi and infaunal foraminifera and H.elegans to those observed in this study.
4.2. δ11B-pH proxy systematics
The good agreement between our epifaunal foraminiferal δ11B andcalculated δ11B of seawater borate suggests we may accuratelyreconstruct ocean pH from δ11B (Fig. 8), and allows us to putconstraints on previously controversial components of the δ11B proxy.
4.2.1. Fractionation factorThe isotopic fractionation factor (αB)betweenB(OH)3 andB(OH)4−has
been determined experimentally by Klochko et al. (2006) as 1.0272±0.0006. This is the only direct experimental measurement of αB inseawater, and is used in our calculations of δ11BB(OH)4− and δ11B-calculatedpH.Given theexcellent correspondencebetweenpredictedandmeasured
ies and morphotypes are plotted against C. wuellerstorfi. b) B/Ca in different Cibicidoidestween C. wuellerstorfi sensu stricto and C. wuellerstorfi var., equivalent to ~30 μmol/kgfi var. or C. wuellerstorfi spp. has a very similar appearance to C. wuellerstorfi s.s., but withr, and C. mundulus var. appears intermediate between C. wuellerstorfi and C. mundulus,

-4 -2 0 2
Dep
th in
MU
C (
cm)
Δδ11B foram - SW B(OH)4-
7.4 7.8
pHtot Δδ11B PW - SW B(OH)4-
a b c d0
1
2
3
4
5
-2.0 -1.5 -1.0 -0.5 0.00
1
2
3
4
5
38.5 39.5δ11B PW
12801
12808
12811
Cibicidoides wuellerstorfiCibicidoides mundulus
Planulina ariminensis
Cibicidoides lobatusCibicidoides ungerianus
Cibicidoides robertsonianusOridorsalis umbonatus
Gyroidina soldanii
Lenticulina vortexAmmonia beccariiMelonis zaandamaeUvigerina peregrina
Hoeglundina elegans
Epifaunal
Infaunal
Aragonite
(‰) (‰) (‰)
Fig. 7. a) Offsets between measured foraminiferal δ11B and δ11B of seawater B(OH)4− for all studied species. b) Measured δ11B of pore-water. c) Pore water pH (total scale), correctedso that depth 0 matches nearby GLODAP values. d) Offset of pore-water B(OH)4− δ11B, calculated from pore water pH and measured pore-water δ11B, relative to δ11B of B(OH)4− atdepth 0. Note that the depths of pore-water samples in b–d only approximately apply to the depths of foraminifera habitat in the left hand column, and that the aragonitic H. elegansis epifaunal.
409J.W.B. Rae et al. / Earth and Planetary Science Letters 302 (2011) 403–413
δ11B of our epifaunal benthic foraminifera, using this independentlyconstrained fractionation factor, it seems appropriate to use this value.Moreover, using αB of Klochko et al. (2006) yields good correspondencebetween predicted and measured δ11B of planktic foraminifera, withmeasured values offset above δ11B of B(OH)4− by 0.7‰. This offset isconsistentwith the sense, if not the absolutemagnitude, of the increase inlocal pH due to symbiont photosynthesis (Jørgensen et al., 1985; Rinket al., 1998; Zeebe et al., 2003). Consistencywith the Klochko et al. (2006)
Fig. 8. Comparison of epifaunal benthic foraminifera δ11B data from MC-ICPMS (thisstudy) to NTIMS data for mixed benthic foraminifera (Sanyal et al., 1995) and C.wuellerstorfi (Hönisch et al., 2008). Note the difference between full procedural 2 s.d.uncertainties on the MC-ICPMS data, and published 2 s.e. uncertainties on NTIMS data,calculated on three or more replicates of each sample.
alpha, given physiological offsets in planktic foraminifera, has also beensuggested by Kasemann et al. (2009).
Earlier authors typically used a value of 1.0194, as reported byKakihana et al. (1977) from theoretical calculations based onvibrational frequency data (Kakihana and Kotaka, 1977; Kotaka andKakihana, 1977). However Rustad and Bylaska (2007) show that amajor vibrational mode was improperly assigned in making theseestimates, yielding spuriously low values. More recent theoreticalwork (Liu and Tossell, 2005; Oi, 2000; Rustad et al., 2010; Zeebe,2005) and adsorption and precipitation experiments (Oi et al., 1991;Sonoda et al., 2000) give significantly larger values (~1.025–1.035),much closer to that of Klochko et al. (2006).
It is likely that αB exhibits some temperature dependence, but themagnitude of this effect is currently unquantified. Zeebe (2005)discusses a potential temperature influence on theoretical calcula-tions of αB, but states that
Given the range of outcome for αB3–B4 at 300 K calculated in thecurrent paper, no recommendation will be made regarding α'stemperature dependence, which equally depends on the frequen-cies/methods chosen.
Klochko et al. (2006) conducted experiments to determine αB at 25and 40 °C, but the uncertainties in the high temperature experiments arelarge and thedata arewithin error. Hönisch et al. (2008) nonetheless usedthe calculations of Zeebe (2005) as a basis for a temperature dependenceonαB of 1.0194 and 1.0361, and interpolated these values forαBof 1.0272.Applying this temperature effect onαBof 1.0272 toour calculations resultsin significant and variable offsets between measured foraminiferal δ11Band the calculated δ11BB(OH)4− (Supplementary Fig. 3). Furthermore, theoffsets of epifaunal foraminiferal δ11B from δ11BB(OH)4−, using αB=1.0272with no temperature dependence, show no correlationwith temperature(Supplementary Fig. 4). Thus although a temperature dependence on thefractionation factor is likely to exist, our data suggests that it is notsignificant over the temperature range relevant forour samples (~2–13 °Cin benthics).
Given the excellent fit of ourmeasured boron isotope data to aqueousborate values predicted using the fractionation factor of 1.0272 (Klochkoet al., 2006) with no temperature effect, we use these assumptions in ourparameterisation of the natural δ11BCaCO3
relationship. Nevertheless we
410 J.W.B. Rae et al. / Earth and Planetary Science Letters 302 (2011) 403–413
realise there is potential for further isotopic fractionations duringincorporation of boron into foraminiferal calcite, and recognise that thecloseness of our fit to simple “inorganic theory” may be fortuitous.However until further work is done to improve our understanding of themechanisms of boron isotope incorporation into benthic foraminifera, wechose to follow this straightforward approach.
4.2.2. Boron incorporationIn their initial δ11B measurements on marine carbonates, Hemming
andHanson (1992) suggested incorporation of borate ion to explain theirlight values relative to seawater. They hypothesised that this chargedspecies was able to interact with growing crystal surfaces, and observedchanges in the growth and geometry of calcite crystals when grown inhigh boron concentration (Hemming et al., 1995). The boron isotope-pHproxy relies on the assumption of exclusive B(OH)4− incorporation.
Recent NMR data (Klochko et al., 2009) showed that trigonal (BO3)and tetrahedral (BO4) coordinations of boron exist in biogenic calcitein roughly equal proportions. However this test of boron coordinationin the crystal lattice is not a test of which molecule — B(OH)3 or B(OH)4− — is incorporated from solution. As Klochko et al. (2009) pointout (their Fig. 9), boron may switch between coordination with 3 or 4oxygens during adsorption and incorporation into borocarbonate,independent of the species initially adsorbed.
Isotope composition is a powerful indirect test of boron speciesincorporation, due to the large isotopic offset between B(OH)3 and B(OH)4− (Fig. 1). Although seawater boron is dominated by B(OH)3(N90%) at the environmental conditions in our core-top dataset, ourδ11B data provide evidence for exclusive B(OH)4− incorporation inforaminiferal calcite: just 2% B(OH)3 would clearly offset our datafrom the close match to the calculated δ11B of seawater B(OH)4−.
4.2.3. Vital effectsVital effects have previously been invoked to explain a number of
offsets of δ11B from theoretical δ11BB(OH)4− values in planktic forami-nifera (Hönisch et al., 2007; Ni et al., 2007). Our benthic data show noevidence of such an effect: epifaunal species closely match δ11BB(OH)4−,and offset values in infaunal speciesmay be explained bymicrohabitat(Section 4.3). The only exception to this is the aragonitic H. elegans,
7.7
7.8
7.9
8.0
8.1
8.2
8.3
7.7 7.8 7.9 8.0 8.2 8.3
PlanulinaC. wuellerstorfiC. mundulusG. ruberG. sacculifer
pHtot seawater
pHto
t for
am
8.1
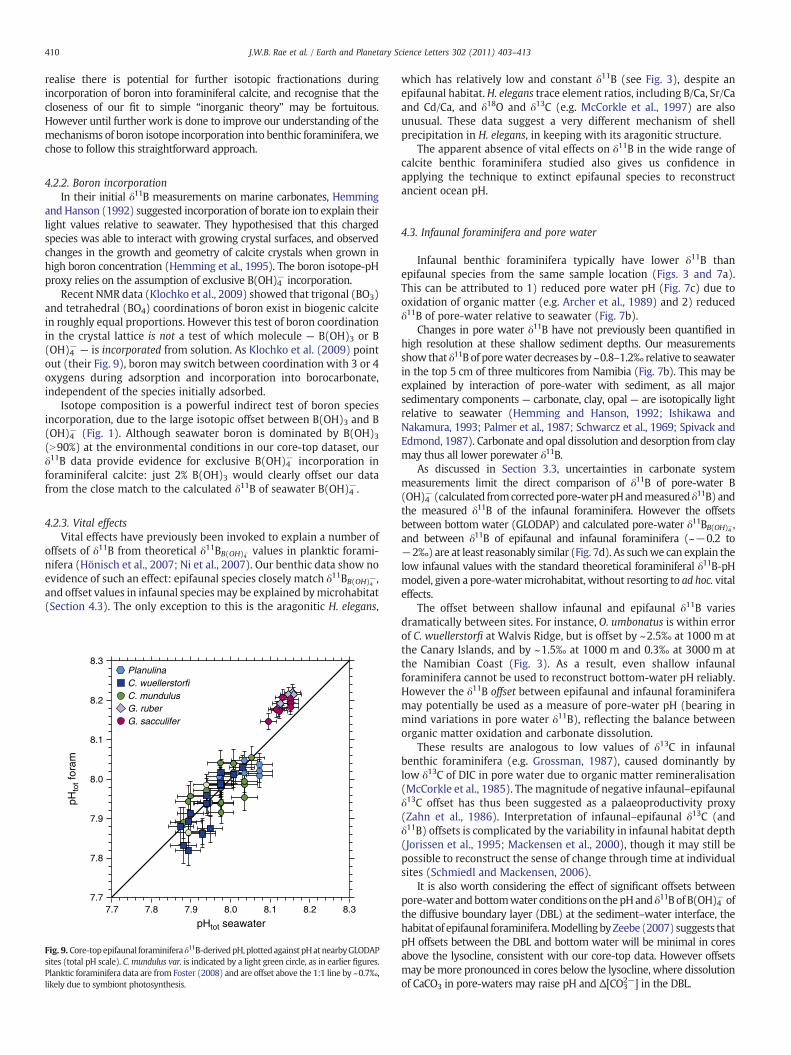
Fig. 9. Core-topepifaunal foraminifera δ11B-derivedpH,plottedagainst pHatnearbyGLODAPsites (total pH scale). C. mundulus var. is indicated by a light green circle, as in earlier figures.Planktic foraminifera data are from Foster (2008) and are offset above the 1:1 line by ~0.7‰,likely due to symbiont photosynthesis.
which has relatively low and constant δ11B (see Fig. 3), despite anepifaunal habitat.H. elegans trace element ratios, including B/Ca, Sr/Caand Cd/Ca, and δ18O and δ13C (e.g. McCorkle et al., 1997) are alsounusual. These data suggest a very different mechanism of shellprecipitation in H. elegans, in keeping with its aragonitic structure.
The apparent absence of vital effects on δ11B in the wide range ofcalcite benthic foraminifera studied also gives us confidence inapplying the technique to extinct epifaunal species to reconstructancient ocean pH.
4.3. Infaunal foraminifera and pore water
Infaunal benthic foraminifera typically have lower δ11B thanepifaunal species from the same sample location (Figs. 3 and 7a).This can be attributed to 1) reduced pore water pH (Fig. 7c) due tooxidation of organic matter (e.g. Archer et al., 1989) and 2) reducedδ11B of pore-water relative to seawater (Fig. 7b).
Changes in pore water δ11B have not previously been quantified inhigh resolution at these shallow sediment depths. Our measurementsshow that δ11B of porewater decreases by ~0.8–1.2‰ relative to seawaterin the top 5 cm of three multicores from Namibia (Fig. 7b). This may beexplained by interaction of pore-water with sediment, as all majorsedimentary components — carbonate, clay, opal — are isotopically lightrelative to seawater (Hemming and Hanson, 1992; Ishikawa andNakamura, 1993; Palmer et al., 1987; Schwarcz et al., 1969; Spivack andEdmond, 1987). Carbonate and opal dissolution and desorption from claymay thus all lower porewater δ11B.
As discussed in Section 3.3, uncertainties in carbonate systemmeasurements limit the direct comparison of δ11B of pore-water B(OH)4− (calculated fromcorrectedpore-waterpHandmeasured δ11B) andthe measured δ11B of the infaunal foraminifera. However the offsetsbetween bottom water (GLODAP) and calculated pore-water δ11BB(OH)4−,and between δ11B of epifaunal and infaunal foraminifera (~−0.2 to−2‰) are at least reasonably similar (Fig. 7d). As suchwe can explain thelow infaunal values with the standard theoretical foraminiferal δ11B-pHmodel, given a pore-watermicrohabitat, without resorting to ad hoc. vitaleffects.
The offset between shallow infaunal and epifaunal δ11B variesdramatically between sites. For instance, O. umbonatus is within errorof C. wuellerstorfi at Walvis Ridge, but is offset by ~2.5‰ at 1000 m atthe Canary Islands, and by ~1.5‰ at 1000 m and 0.3‰ at 3000 m atthe Namibian Coast (Fig. 3). As a result, even shallow infaunalforaminifera cannot be used to reconstruct bottom-water pH reliably.However the δ11B offset between epifaunal and infaunal foraminiferamay potentially be used as a measure of pore-water pH (bearing inmind variations in pore water δ11B), reflecting the balance betweenorganic matter oxidation and carbonate dissolution.
These results are analogous to low values of δ13C in infaunalbenthic foraminifera (e.g. Grossman, 1987), caused dominantly bylow δ13C of DIC in pore water due to organic matter remineralisation(McCorkle et al., 1985). Themagnitude of negative infaunal–epifaunalδ13C offset has thus been suggested as a palaeoproductivity proxy(Zahn et al., 1986). Interpretation of infaunal–epifaunal δ13C (andδ11B) offsets is complicated by the variability in infaunal habitat depth(Jorissen et al., 1995; Mackensen et al., 2000), though it may still bepossible to reconstruct the sense of change through time at individualsites (Schmiedl and Mackensen, 2006).
It is also worth considering the effect of significant offsets betweenpore-water andbottomwater conditions on thepHand δ11B of B(OH)4−ofthe diffusive boundary layer (DBL) at the sediment–water interface, thehabitat of epifaunal foraminifera.ModellingbyZeebe (2007) suggests thatpH offsets between the DBL and bottom water will be minimal in coresabove the lysocline, consistent with our core-top data. However offsetsmay bemore pronounced in cores below the lysocline, where dissolutionof CaCO3 in pore-waters may raise pH and Δ[CO3
2−] in the DBL.

411J.W.B. Rae et al. / Earth and Planetary Science Letters 302 (2011) 403–413
4.4. B/Ca
There is good reason to believe that B/Ca in marine carbonate shouldhave a relationship with the carbonate system, given exclusive incorpo-ration of boron as the charged B(OH)4− ion (Hemming and Hanson, 1992,Section4.2.2) and thedependenceof [B(OH)4−] onpH.However no simplerelationship is foundbetweenB/Ca and [B(OH)4−], pH, or anyhypotheticalpartition coefficient of boron into calcite (Foster, 2008; Tripati et al., 2009;Yu and Elderfield, 2007; Yu et al., 2007). Comparison of similar benthicforaminifera species from the same sample habitat reveals large vitaleffects on B/Ca (Yu and Elderfield, 2007, and Fig. 6), suggesting theremaybe a strong physiological control on the B/Ca of foraminiferal calcite.
Yu and Elderfield (2007) demonstrated a good correlation betweenbenthic B/Ca in C. wuellerstorfi and C. mundulus and carbonate ionsaturation state (Δ[CO3
2−]). Our C. wuellerstorfi and C. mundulus B/Ca datafit this empirical relationship (Fig. 5), but even subtle variants inmorphotype show significant deviations from this trend (Fig. 6). Suchspecimensareoftencombined forδ18Oandδ13Canalyses, but clearlymorestringent selection criteria are required for B/Ca ratios, which are up to~40 μmol/mol lower in C. wuellerstorfi var. than C. wuellerstorfi sensustricto, equivalent to ~30 μmol/kg Δ[CO3
2−] (the whole deep Atlanticglacial–interglacial change; Yu and Elderfield, 2007). These vital effectsalso preclude the use of B/Ca as a quantitative proxy ofΔ[CO3
2−] in extinctspecies. In contrast, δ11B of these different species from the same coresample lie within error (Fig. 6a), allowing the possibility of combiningthese species for δ11B analysis when sample sizes are small.
4.5. Carbonate system determination
Theoceanic carbonate systemhas6 componentswith4 independentequations between them; if we can quantify 2 of these components, wecan determine the state of the whole system. δ11B and B/Ca record pHand Δ[CO3
2−] (and thereby [CO32−]) respectively; in theory this should
allow reconstruction of the “master variables,” dissolved inorganiccarbon (DIC) and alkalinity (ALK), from these proxies. However evenwhen foraminifera-derived pH and [CO3
2−] data are very close toseawater values, our core-top data demonstrate that such quantitativereconstruction is impractical (Fig. 10a). This is due to the very closecorrelation of pH and [CO3
2−] in DIC-ALK space (Fig. 10b), meaning thatany errors and uncertainties are greatly magnified in attempting todetermine their intersection (also noted by Yu et al., 2010). Fullpropagation of δ11B and B/Ca measurement uncertainties gives averageabsolute 2 s.d. uncertainty on calculated DIC at Walvis Ridge of
ALK
(m
mol
/kg)
0
1000
2000
3000
4000
1500 2000 2500 3000
DIC (μmol/kg)
Dep
th (
m)
a) b)
Fig. 10. a) Calculated DIC from δ11B-derived pH and B/Ca-derived [CO32−]. The grey bar indica
of DIC and ALK. Close correlation of pH and [CO32−] leads to large errors on reconstructed D
~300 μmol/kg sw (~13% relative uncertainty), and ALK of ~230 μmol/kg sw (~10% relative uncertainty). As a result our DIC estimates span agreater range than the entire modern ocean.
Nevertheless, both proxies should record similar changes in thecarbonate system, due to the similar trends of pH, CO2 and CO3
2− inDIC-ALK space, and may thus be used in a complementary fashion (Yuet al., 2010). Since B/Ca measurements are more analytically straightfor-ward and can be made on smaller amounts of sample, they provide thepotential to generate large datasets more easily than with δ11B (Rickabyet al., 2010; Yu et al., 2010). However, the caveat of B/Ca sensitivity tospeciesmorphotype, whichmay change systematically down-core, needsto be carefully assessed in such applications.
4.6. Boron incorporation and foraminiferal calcification
The fact that bulk foraminiferal δ11B matches δ11BB(OH)4− at
seawater pH is a powerful constraint on mechanisms of foraminiferalcalcification. Assuming that foraminifera calcify from an internalreservoir or vacuole of modified seawater (de Nooijer et al., 2009a;Elderfield et al., 1996; Erez, 2003), we speculate on two scenarios thatare compatible with our observations:
1. incorporation of a small amount of B(OH)4− from a seawater-likecalcifying medium at seawater pH;
2. removal of a small amount of B(OH)4− to a HCO3− pool, which may
then vary its pH as long as all the boron in this pool is incorporated.
In either scenario the quantity of B(OH)4− removed from the vacuolemust be small, as if we remove significant amounts of B(OH)4− from afinite reservoir, Rayleigh fractionation will occur, shifting the compo-sition of the borate (and the foraminiferal calcite) towards seawaterδ11B. Given thevery small partition coefficientof boron into foraminifera(foraminiferal B/Ca being ~200 times lower than seawater) extensiveboron removal from the vacuole seems unlikely.
A relatively direct mechanism of foraminiferal calcification fromseawater, as in scenario 1, is suggested by the extensive and rapidstreaming of seawater vacuoles to the site of chamber formation, asobserved by Bentov et al. (2009). However calcification in or directly fromthese vacuoles is not consistent with our boron isotope data, given thesignificant increases in vacuole pH (~+0.5) observed during chamberformation (Bentov et al., 2009; de Nooijer et al., 2009b). Such elevated pHshould increase vacuole δ11BB(OH)4−, and thus δ11B of foraminiferal CaCO3,by ~5‰ above δ11BB(OH)4− in the surrounding seawater.
2.25
2.30
2.35
2.40
2.45
1.95 2.00 2.05 2.10 2.15
DIC (mmol/kg)
pH300
200
200
100
8.2
8.2
8.1
8.1
8.0
7.9
7.9
7.8
[CO3
2- ]
250
250
150
DIC/A
LK
tes the full range of modern ocean DIC (Key et al., 2004). b) pH and [CO32−] as a function
IC or ALK, shown figuratively by the grey shaded area.

412 J.W.B. Rae et al. / Earth and Planetary Science Letters 302 (2011) 403–413
Culture experiments have suggested that foraminifera concentrateand store Ca2+ andDIC internally (Anderson and Faber, 1984; Erez, 2003;ter Kuile and Erez, 1987, 1988; ter Kuile et al., 1989), possibly by pumpingCa2+ and HCO3
− into a secondary reservoir (de Nooijer et al., 2009a;Toyofuku et al., 2008). It has been suggested that a Ca2+ pump may be“leaky”with respect to other cations, largely discriminatingly in favour ofCa but letting in trace quantities of other cations of similar size and charge(e.g. Mg2+; de Nooijer et al., 2009a; Erez, 2003; Raitzsch et al., 2009). In asimilar way, a HCO3
− transporter might be leaky to small quantities of thecharged B(OH)4− ion, but not B(OH)3. Once B(OH)4− is taken into thisreservoir, pH could vary significantly without affecting foraminiferal δ11B,as long as all the boron in this pool is incorporated.
Themajor challenge to this scenario is that whilst B(OH)4− is removedfrom the original seawater vacuole, the pH of this vacuole must remainequal to seawater. This may be difficult to achieve given removal ofsignificant quantities of Ca2+ and HCO3
−, and would require tightbiological controls on the ALK/DIC ratio. However it is not impossiblethat such controls exist to maintain charge balance during pumping, forinstance by exchange with Na+ or Cl− (Flatman, 1984; Günzel et al.,1997).
Pumping of B(OH)4− also allows different foraminifera species tohave the same δ11B as B(OH)4− but significantly different B/Ca ratios.Given the ~200 fold decrease in B/Ca ratio relative to seawater, onlysmall changes to the efficiency of this process are required to causesignificant differences in foraminiferal B/Ca.
5. Conclusions
The good agreement of the δ11B of our core-top epifaunal benthicforaminifera to calculated δ11B of B(OH)4− in ambient seawater stronglysupports the concept of the δ11B proxy of seawater pH (Hemming andHanson, 1992). This is based on simple, inorganic fractionation of boronisotopes between B(OH)3 and B(OH)4− in seawater, assuming soleincorporation of B(OH)4− into foraminiferal carbonate and no vital effects,and using the independently determined αB of 1.0272 (Klochko et al.,2006). Infaunal foraminifera also fit with this inorganic model but recordδ11B of B(OH)4− reflecting the δ11B and pH of pore water, which are offsetfrom bottom water. Our B/Ca data correlate with seawater Δ[CO3
2−], inagreement with the work of Yu and Elderfield (2007), but marked B/Cavariations are found between very similar Cibicidoides species, whichshow no significant change in δ11B. While δ11B and B/Ca proxy twodifferent parameters of the oceanic carbonate system, reconstruction ofalkalinity and DIC is precluded by error propagation. The δ11B and B/Cadata are consistent with a calcification mechanism where B(OH)4− isincorporated from a seawater vacuole to a HCO3
− pool, which is then fullyincorporated into foraminiferal carbonate.
Supplementarymaterials related to this article can be found onlineat doi:10.1016/j.epsl.2010.12.034.
Acknowledgements
We thank: Barbara Donner and colleagues at the Marum, Universityof Bremen, for samples from the Canary Islands and Walvis Ridge;Matthias Zabel and colleagues at theUniversity of Bremen, formulticoreand pore-water samples from the Namibian Coast; AndreasMackensenand Hannes Grobe at the AWI Bremerhaven for samples from PS2495;Andreas Mackensen, Gerhard Schmiedl (University of Hamburg) andEllen Thomas (Yale University) for advice on Cibicidoidesmorphotypes;Chris Coath for crucial assistance in the laboratory; participants at theboron mini-conference at Lamont-Doherty Earth Observatory for openand stimulating discussions; four anonymous reviewers for carefulcomments that improved thismanuscript; and Peter DeMenocal for defteditorial handling. This researchwas funded by a NERC studentship to J.R., an advanced NERC fellowship to G.F. and a Royal Society URF to D.S.,and benefitted from analytical developments made as part of NER/A/S/2001/10213 and NE/C510983/1 to T.E.
References
Al-Ammar, A., Gupta, R., Barnes, R., 2000. Elimination of boron memory effect ininductively coupled plasma-mass spectrometry by ammonia gas injection into thespray chamber during analysis. Spectrochim. Acta B 55 (6), 629–635.
Anderson, O., Faber, W., 1984. An estimation of calcium carbonate deposition rate in aplanktonic foraminifer Globigerinoides sacculifer using 45Ca as a tracer: arecommended procedure for improved accuracy. J. Foraminiferal Res. 14, 303–308.
Archer, D., Emerson, S., Reimers, C., 1989. Dissolution of calcite in deep-sea sediments:pH and O2 microelectrode results. Geochim. Cosmochim. Acta 53 (11), 2831–2845.
Barker, S., Greaves, M., Elderfield, H., 2003. A study of cleaning procedures used forforaminiferal Mg/Ca paleothermometry. Geochem. Geophys. Geosyst. 4 (9), 8407.
Bentov, S., Brownlee, C., Erez, J., 2009. The role of seawater endocytosis in thebiomineralization process in calcareous foraminifera. Proc. Natl. Acad. Sci. 106 (51),21,500–21,504.
Catanzaro, E., Champion, C., Garner, E., Marinenko, G., Sappenfield, K., Shields, W., 1970.Boric acid: isotopic and assay standard reference materials. NBS (US) Spec. Publ.,260, pp. 1–70.
Culberson, C., Pytkowicz, R., 1968. Effect of pressure on carbonic acid, boric acid, and thepH in seawater. Limnol. Oceanogr. 13 (3), 403–417.
de Nooijer, L., Langer, G., Nehrke, G., Bijma, J., 2009a. Physiological controls on seawateruptake and calcification in the benthic foraminifer Ammonia tepida. Biochem.Biophys. Res. Commun. 6 (4), 7083–7102.
de Nooijer, L., Toyofuku, T., Kitazato, H., 2009b. Foraminifera promote calcification byelevating their intracellular pH. Proc. Natl. Acad. Sci. 106 (36), 15,374–15,379.
Dickson, A.G., 1990. Thermodynamics of the dissociation of boric acid in syntheticseawater from 273.15 to 318.15 K. Deep Sea Res. A 37 (5), 755–766.
Dickson, A., Sabine, C., Christian, J. (Eds.), 2007. Guide to Best Practices for Ocean CO2
Measurements. North Pacific Marine Science Organization, Sidney, BritishColumbia.
Elderfield, H., Bertram, C., Erez, J., 1996. A biomineralizationmodel for the incorporationof trace elements into foraminiferal calcium carbonate. Earth Planet. Sci. Lett. 142,409–423.
Erez, J., 2003. The source of ions for biomineralization in foraminifera and theirimplications for paleoceanographic proxies. Rev. Mineral. Geochem. 54 (1), 115–149.
Flatman, P., 1984. Magnesium transport across cell membranes. J. Membr. Biol. 80 (1),1–14.
Foster, G., 2008. Seawater pH, pCO2 and [CO32−] variations in the Caribbean Sea over the
last 130 kyr: a boron isotope and B/Ca study of planktic foraminifera. Earth Planet.Sci. Lett. 271 (1–4), 254–266.
Foster, G., Ni, Y., Haley, B., Elliott, T., 2006. Accurate and precise isotopic measurementof sub-nanogram sized samples of foraminiferal hosted boron by total evaporationNTIMS. Chem. Geol. 230 (1–2), 161–174.
Foster, G.L., Pogge von Strandmann, P.A.E., Rae, J.W.B., 2010. Boron and magnesiumisotopic composition of seawater. Geochem. Geophys. Geosyst. 11 (8).
Grossman, E., 1987. Stable isotopes in modern benthic foraminifera; a study of vitaleffect. J. Foraminiferal Res. 17 (1), 48–61.
Günzel, D., Durry, S., Schlue, W., 1997. Intracellular alkalinization causes Mg2+ releasefrom intracellular binding sites in leech Retzius neurones. Pflügers Arch. Eur. J.Physiol. 435 (1), 65–73.
Hemming, N.G., Hanson, G.N., 1992. Boron isotopic composition and concentration inmodern marine carbonates. Geochim. Cosmochim. Acta 56, 537–543.
Hemming, N., Hanson, G., 1994. A procedure for the isotopic analysis of boron bynegative thermal ionization mass spectrometry. Chem. Geol. 114 (1–2), 147–156.
Hemming, N., Hönisch, B., 2007. Boron isotopes in marine carbonate sediments and theph of the ocean. In: Hillaire-Marcel, C., De Vernal, A. (Eds.), Proxies in Late CenozoicPaleoceanography, Vol. 1 (17). Elsevier Science Ltd, pp. 717–734.
Hemming, N., Reeder, R., Hanson, G., 1995. Mineral-fluid partitioning and isotopicfractionation of boron in synthetic calcium carbonate. Geochim. Cosmochim. Acta59 (2), 371–379.
Hillaire-Marcel, C., De Vernal, A. (Eds.), 2007. Proxies in Late Cenozoic Paleoceanography.Elsevier Science.
Hönisch, B., Hemming, G.N., 2004. Ground-truthing the boron isotope-paleo-pH proxyin planktonic foraminifera shells: partial dissolution and shell size effects.Paleoceanography 19, PA4010.
Hönisch, B., Hemming, N.G., 2005. Surface ocean pH response to variations in pCO2
through two full glacial cycles. Earth Planet. Sci. Lett. 236 (1–2), 305–314.Hönisch, B., Bijma, J., Russell, A.D., Spero, H.J., Palmer, M.R., Zeebe, R.E., Eisenhauer, A.,
2003. The influence of symbiont photosynthesis on the boron isotopic compositionof foraminifera shells. Mar. Micropaleontol. 49 (1–2), 87–96.
Hönisch, B., Hemming, N., Loose, B., 2007. Comment on “A critical evaluation of theboron isotope-pH proxy: The accuracy of ancient ocean pH estimates” byM. Pagani,D. Lemarchand, A. Spivack and J. Gaillardet. Geochim. Cosmochim. Acta 71 (6),1636–1641.
Hönisch, B., Bickert, T., Hemming, N., 2008. Modern and Pleistocene boron isotopecomposition of the benthic foraminifer Cibicidoides wuellerstorfi. Earth Planet. Sci.Lett. 272 (1–2), 309–318.
Hönisch, B., Hemming, N.G., Archer, D., Siddall, M., McManus, J.F., 2009. Atmosphericcarbon dioxide concentration across the mid-Pleistocene transition. Science 324(5934), 1551–1554.
Ingle, S., 1975. Solubility of calcite in the ocean. Mar. Chem. 3 (4), 301–319.Ishikawa, T., Nakamura, E., 1993. Boron isotope systematics of marine sediments. Earth
Planet. Sci. Lett. 117 (3–4), 567–580.Jørgensen, B., Erez, J., Revsbach, N., Cohen, Y., 1985. Symbiotic photosynthesis in a planktonic
foraminifera, Globigerinoides sacculifer (brady), studied with microelectrodes. Limnol.Oceanogr. 30, 1253–1267.

413J.W.B. Rae et al. / Earth and Planetary Science Letters 302 (2011) 403–413
Jorissen, F., de Stigter, H., Widmark, J., 1995. A conceptual model explaining benthicforaminiferal microhabitats. Mar. Micropaleontol. 26, 3–15.
Kakihana, H., Kotaka, M., 1977. Equilibrium constants for boron isotope-exchangereactions. Bull. Res. Lab. Nucl. React. 2, 1–12.
Kakihana, H., Kotaka, M., Satoh, S., Nomura, M., Okamoto, M., 1977. Fundamentalstudies on the ion-exchange separation of boron isotopes. Bull. Chem. Soc. Jpn. 50,158–163.
Kasemann, S.A., Schmidt, D.N., Bijma, J., Foster, G.L., 2009. In situ boron isotope analysisin marine carbonates and its application for foraminifera and palaeo-pH. Chem.Geol. 260 (1–2), 138–147.
Key, R.M., Kozyr, A., Sabine, C.L., Lee, K., Wanninkhof, R., Bullister, J.L., Feely, R.A., Millero, F.J.,Mordy, C., Peng, T.-H., 2004. A global ocean carbon climatology: results from global dataanalysis project (GLODAP). Global Biogeochem. Cycles 18, GB4031.
Kiss, E., 1988. Ion-exchange separation and spectrophotometric determination of boronin geological materials. Anal. Chim. Acta 211, 243–256.
Klochko, K., Kaufman, A.J., Yao, W., Byrne, R.H., Tossell, J.A., 2006. Experimentalmeasurement of boron isotope fractionation in seawater. Earth Planet. Sci. Lett. 248(1–2), 276–285.
Klochko, K., Cody, G.D., Tossell, J.A., Dera, P., Kaufman, A.J., 2009. Re-evaluating boronspeciation in biogenic calcite and aragonite using 11B MAS NMR. Geochim.Cosmochim. Acta 73 (7), 1890–1900.
Kotaka, M., Kakihana, H., 1977. Thermodynamic isotope effect of trigonal planar andtetrahedral species. Bull. Res. Lab. Nucl. React. 2, 13–29.
Lee, K., Kim, T.-W., Byrne, R.H., Millero, F.J., Feely, R.A., Liu, Y.-M., 2010. The universalratio of boron to chlorinity for the North Pacific and North Atlantic oceans.Geochim. Cosmochim. Acta 74 (6), 1801–1811.
Liu, Y., Tossell, J.A., 2005. Ab initio molecular orbital calculations for boron isotopefractionations on boric acids and borates. Geochim. Cosmochim. Acta 69 (16),3995–4006.
Lueker, T.J., Dickson, A.G., Keeling, C.D., 2000. Ocean pCO2 calculated from dissolvedinorganic carbon, alkalinity, and equations for K1 and K2: validation based onlaboratory measurements of CO2 in gas and seawater at equilibrium. Mar. Chem. 70(1–3), 105–119.
Lutze, G., Thiel, H., 1989. Epibenthic foraminifera from elevated microhabitats:Cibicidodes wuellerstorfi and Planulina arimensis. J. Foraminiferal Res. 19 (2),153–158.
Mackensen, A., Hubberten, H.W., Bickert, T., Fischer, G., Fütterer, D.K., 1993. The δ13C inbenthic foraminiferal tests of Fontbotia wuellerstorfi (Schwager) relative to the δ13Cof dissolved inorganic carbon in southern ocean deepwater: implications for glacialocean circulation models. Paleoceanography 8 (5), 587–610.
Mackensen, A., Schumacher, S., Radke, J., Schmidt, D.N., 2000. Microhabitat preferencesand stable carbon isotopes of endobenthic foraminifera: clue to quantitativereconstruction of oceanic new production? Mar. Micropaleontol. 40 (3), 233–258.
McCorkle, D., Emerson, S., Quay, P., 1985. Stable carbon isotopes in marine porewaters.Earth Planet. Sci. Lett. 74 (1), 13–26.
McCorkle, D., Corliss, B., Farnham, C., 1997. Vertical distributions and stable isotopiccompositions of live (stained) benthic foraminifera from the North Carolina andCalifornia continental margins. Deep Sea Res. I 44 (6), 983–1024.
Mehrbach, C., Culberson, C., Hawley, J., Pytkowicz, R., 1973. Measurement of theapparent dissociation constants of carbonic acid in seawater at atmosphericpressure. Limnol. Oceanogr. 18 (6), 897–907.
Millero, F., 1979. The thermodynamics of the carbonate system in seawater. Geochim.Cosmochim. Acta 43, 1651–1661.
Ni, Y., Foster, G.L., Bailey, T., Elliott, T., Schmidt, D.N., Pearson, P., Haley, B., Coath, C.,2007. A core top assessment of proxies for the ocean carbonate system in surfacedwelling foraminifers. Paleoceanography 22 (3), PA3212.
Ni, Y., Foster, G.L., Elliott, T., 2010. The accuracy of δ11B measurements of foraminifers.Chem. Geol. 274 (3–4), 187–195.
Oi, T., 2000. Calculations of reduced partition function ratios of monomeric and dimericboric acids and borates by the ab initio molecular orbital theory. J. Nucl. Sci.Technol. 37 (2), 166–172.
Oi, T., Kato, J., Ossaka, T., Kakihana, H., 1991. Boron isotope fractionation accompanyingboron mineral formation from aqueous boric acid–sodium hydroxide solutions at25 °C. Geochem. J. 25, 377–385.
Pagani, M., Lemarchand, D., Spivack, A., Gaillardet, J., 2005. A critical evaluation of theboron isotope-pH proxy: the accuracy of ancient ocean pH estimates. Geochim.Cosmochim. Acta 69 (4), 953–961.
Palmer, M., Spivack, A., Edmond, J., 1987. Temperature and pH controls over isotopicfractionation during adsorption of boron on marine clay. Geochim. Cosmochim.Acta 51 (9), 2319–2323.
Raitzsch, M., Dueñas-Bohórquez, A., Reichart, G., de Nooijer, L., Bickert, T., 2009. Theimpact of seawater calcite saturation state by modifying Ca ion concentrations onMg and Sr incorporation in cultured benthic foraminifera. Biochem. Biophys. Res.Commun. 6, 11,347–11,375.
Rickaby, R.E.M., Elderfield, H., Roberts, N., Hillenbrand, C.D., Mackensen, A., 2010.Evidence for elevated alkalinity in the glacial southern ocean. Paleoceanography 25(1), PA1209.
Rink, S., Kühl, M., Bijma, J., Spero, H.J., 1998. Microsensor studies of photosynthesis andrespiration in the symbiotic foraminifer Orbulina universa. Mar. Biol. 131 (4),583–595.
Rustad, J.R., Bylaska, E.J., 2007. Ab initio calculation of isotopic fractionation in B(OH)3(aq) and B(OH)4− (aq). J. Am. Chem. Soc. 129 (8), 2222–2223.
Rustad, J., Bylaska, E., Jackson, V., Dixon, D., 2010. Calculation of boron-isotopefractionation between B(OH)3 (aq) and B(OH)4− (aq). Geochim. Cosmochim. Acta74 (10), 2843–2850.
Sabine, C.L., Feely, R.A., Gruber, N., Key, R.M., Lee, K., Bullister, J.L., Wanninkhof, R.,Wong, C.S., Wallace, D.W.R., Tilbrook, B., Millero, F.J., Peng, T.-H., Kozyr, A., Ono, T.,Rios, A.F., 2004. The oceanic sink for anthropogenic CO2. Science 305 (5682),367–371.
Sanyal, A., Hemming, N., Hanson, G., Broecker, W., 1995. Evidence for a higher pH in theglacial ocean from boron isotopes in foraminifera. Nature 373, 234–236.
Schlitzer, R., 2010. Ocean Data View. http://www.odv.awi.de2010.Schmiedl, G., Mackensen, A., 2006. Multispecies stable isotopes of benthic foraminifers
reveal past changes of organic matter decomposition and deepwater oxygenationin the arabian sea. Paleoceanography 21 (4), PA4213.
Schwarcz, H., Agyei, E., McMullen, C., 1969. Boron isotopic fractionation during clayadsorption from seawater. Earth Planet. Sci. Lett. 6, 1–5.
Seeberg-Elverfeldt, J., Schlüter, M., Feseker, T., Kölling, M., 2005. Rhizon sampling ofpore waters near the sediment/water interface of aquatic systems. Limnol.Oceanogr. Methods 3, 361–371.
Sonoda, A., Makita, Y., Ooi, K., Takagi, N., Hirotsu, T., 2000. pH-dependence of thefractionation of boron isotopes with N-Methyl-D-Glucamine resin in aqueoussolution systems. Bull. Chem. Soc. Jpn. 73 (5), 1131–1133.
Spivack, A., Edmond, J., 1987. Boron isotope exchange between seawater and theoceanic crust. Geochim. Cosmochim. Acta 51 (5), 1033–1043.
ter Kuile, B., Erez, J., 1987. Uptake of inorganic carbon and internal carbon cycling insymbiont-bearing benthonic foraminifera. Mar. Biol. 94 (4), 499–509.
ter Kuile, B., Erez, J., 1988. The size and function of the internal inorganic carbon pool ofthe foraminifer Amphistegina lobifera. Mar. Biol. 99 (4), 481–487.
ter Kuile, B., Erez, J., Padan, E., 1989. Mechanisms for the uptake of inorganic carbon bytwo species of symbiont-bearing foraminifera. Mar. Biol. 103 (2), 241–251.
Toyofuku, T., de Nooijer, L., Yamamoto, H., Kitazato, H., 2008. Real-time visualization ofcalcium ion activity in shallow benthic foraminiferal cells using the fluorescentindicator Fluo-3 AM. Geochem. Geophys. Geosyst. 9 (5).
Tripati, A.K., Roberts, C.D., Eagle, R.A., 2009. Coupling of CO2 and ice sheet stability overmajor climate transitions of the last 20 million years. Science 326, 1394–1397.
van Heuven, S., Pierrot, D., Lewis, E., Wallace, D.W.R., 2009. MATLAB ProgramDeveloped for CO2 System Calculations. Vol. ORNL/CDIAC-105b. Carbon DioxideInformation Analysis Center, Oak Ridge National Laboratory, U.S. Department ofEnergy, Oak Ridge, Tennessee.
Yu, J., Elderfield, H., 2007. Benthic foraminiferal B/Ca ratios reflect deepwater carbonatesaturation state. Earth Planet. Sci. Lett. 258 (1–2), 73–86.
Yu, J., Elderfield, H., Hönisch, B., 2007. B/Ca in planktonic foraminifera as a proxy forsurface seawater pH. Paleoceanography 22 (3), PA2202.
Yu, J., Foster, G.L., Elderfield, H., Broecker, W.S., Clark, E., 2010. An evaluation of benthicforaminiferal B/Ca and δ11B for deep ocean carbonate ion and pH reconstructions.Earth Planet. Sci. Lett. 293 (1–2), 114–120.
Zahn, R., Winn, K., Sarnthein, M., 1986. Benthic foraminiferal δ13C and accumulationrates of organic carbon: Uvigerina peregrina group and Cibicidoides wuellerstorfi.Paleoceanography 1 (1), 27–42.
Zeebe, R.E., 2005. Stable boron isotope fractionation between dissolved B(OH)3 and B(OH)4−. Geochim. Cosmochim. Acta 69 (11), 2753–2766.
Zeebe, R., 2007. Modeling CO2 chemistry, δ13C, and oxidation of organic carbon andmethane in sediment porewater: implications for paleo-proxies in benthicforaminifera. Geochim. Cosmochim. Acta 71 (13), 3238–3256.
Zeebe, R.E., Wolf-Gladow, D., 2001. CO2 in Seawater: Equilibrium, Kinetic, Isotopes.Elsevier, Amsterdam.
Zeebe, R.E., Wolf-Gladrow, D.A., Bijma, J., Hönisch, B., 2003. Vital effects in foraminiferado not compromise the use of d11b as a paleo-pH indicator: evidence frommodeling. Paleoceanography 18 (2).




















