
Fingerprinting electronic structure of heme iron by ab initio ...
Upload
jamiahamdard-inCategory
view
3download
0
B
SSa
b
c
d
a
ARRAA
KBPPCM
1
lscpimpcmpsa[i
ami
0d
International Journal of Biological Macromolecules 47 (2010) 109–115
Contents lists available at ScienceDirect
International Journal of Biological Macromolecules
journa l homepage: www.e lsev ier .com/ locate / i jb iomac
oron stabilizes peroxide mediated changes in the structure of heme proteins
hakir Ali a,∗, Humaira Farooqia,b, Ram Prasada, Mohammad Naimea, Indusmita Routraya,avita Yadavc, Faizan Ahmadd
Department of Biochemistry, Faculty of Science, Jamia Hamdard (Hamdard University), Hamdard Nagar, New Delhi 110062, IndiaDepartment of Biotechnology, Faculty of Science, Jamia Hamdard, Hamdard Nagar, New Delhi 110062, IndiaDepartment of Biophysics, All India Institute of Medical Sciences (AIIMS), New Delhi 110029, IndiaCenter for Interdisciplinary Research in Basic Sciences, Jamia Millia Islamia, New Delhi 110025, India
r t i c l e i n f o
rticle history:eceived 15 April 2010eceived in revised form 13 May 2010ccepted 17 May 2010vailable online 31 May 2010
eywords:
a b s t r a c t
Boron is reported in this study to stabilize the structure of heme proteins exposed to peroxides. Theoxidized heme protein (15 �M) was treated with H2O2 (10 mM) in 1 M glycine–NaOH buffer (pH 9.2) at25 ◦C in absence/presence of boron, and characterized by visible absorption spectroscopy, gel exclusionchromatography, native PAGE, HPLC and DLS. Spectral analysis of exposed heme proteins revealed adecrease in absorbance in the Soret region, which was stabilized by boron. The native PAGE analysisof exposed heme proteins showed high molecular weight products; the band intensity was lesser in
oronrotein stabilizationeroxideytochrome cetmyoglobin
presence of boron. Further, elution profile of the exposed heme proteins on Sephadex G-200 column andHPLC revealed more than one peak (aggregate formation) when compared to the respective untreatedproteins. DLS, which measures the hydrodynamic radius (RH), was used to ascertain whether the peakscorrespond to monomer, dimer or aggregate forms. The RH of boron pretreated heme proteins was closeto RH of the respective heme protein. Non-heme protein RNase did not show any change when exposed
er, reites o
to peroxide. Taken togethmight be due to specific s
. Introduction
Folded structure of a protein, which is largely stabilized by cova-ent and non-covalent interactions, is thermodynamically moretable and active than its unfolded form. However, under stressonditions such as the oxidative stress, the 3-D folded structure iserturbed, and leads to the loss of protein function. Oxidative stress
s produced by the excessive production of partially reduced oxygenoieties such as the superoxide anion radical (O2
•−) and hydrogeneroxide, and has been implicated in large number of pathologicalonditions such as Alzheimer’s disease and various chronic inflam-atory disorders [1–3]. The partially reduced chemical species (ex:
eroxides) are produced from both the exogenous and endogenousources as inescapable by-product of normal aerobic metabolism,nd may inflict damage to nucleic acids, lipids, sugars and proteins4–8]. Reactions of H2O2 with heme proteins have been extensivelynvestigated and reported [9–19].
In the cell, alterations in protein structure are usually counter-cted by specific chemical chaperones capable of correcting theisfolded conformations, thus preventing the loss of protein stabil-
ty and biological activity. In this study, we have investigated the
∗ Corresponding author. Tel.: +91 11 26059688.E-mail addresses: [email protected], [email protected] (S. Ali).
141-8130/$ – see front matter © 2010 Elsevier B.V. All rights reserved.oi:10.1016/j.ijbiomac.2010.05.013
sults conclude that boron stabilizes the structure of heme proteins, whichn heme proteins that can bind to borate ions.
© 2010 Elsevier B.V. All rights reserved.
effect of boron on peroxide mediated alterations in the structureof heme proteins. Boron is a dynamic trace element with multi-ple effects. It is important in mineral metabolism, brain functionand performance and prevention of osteoporosis and osteoarthritis[20]. Boron strengthens the body’s immune system [21], and pre-vents the progression of tissue injury [22]. We hypothesize theseeffects to be due to the ability of boron to preserve the three dimen-sional structure of protein, and in the current study, demonstratedthat boron in the form of borax stabilizes the structure of oxidizedheme proteins, cytochrome c (Cyt c) and metmyoglobin (met-Mb)exposed to peroxides in vitro. Considering the widespread role ofiron-containing proteins, this study has both clinical and industrialimplications.
2. Materials and methods
2.1. Materials
Horse heart cytochrome c and metmyoglobin were purchased
from Sigma and Sephadex G-200 was purchased from Pharmacia,Sweden. Other reagents used in this study were of high purity pur-chased from the standard commercial sources in India. Hydrogenperoxide (30%) and methanol was purchased from E. Merck, India,and borax was purchased from CDH.
110 S. Ali et al. / International Journal of Biological Macromolecules 47 (2010) 109–115
F M) ac withS oron.i
2
if
2s
a
ig. 1. (A) Spectroscopic measurements of Cyt c (15 �M) in presence of H2O2 (10 mhange in absorbance with time. (B) Spectra of Cyt c monitored after 1 h incubationoret absorbance peak in H2O2 treated samples both in presence and absence of bons. The spectra were taken as function of time for 1 h.
.2. Preparation of protein stock solutions
To ensure that the proteins used in the experiments were alln the ferric form, each sample was oxidized in 0.1% potassiumerricyanide. Excess of this salt was removed by dialysis.
.3. Preparation of H2O2-activated metMb and oxidized Cyt cystem
The commercial solution of H2O2 (10 M) was diluted to 10 mMnd mixed with the oxidized Cyt c and metMb to initiate the
nd absence (a) or presence (b) of borax (40 mM). Arrows indicate the direction ofhydrogen peroxide in absence or presence of borax. Note a 2 nm shift to the left of(C) Time-scan of change at A409 for cytochrome c in absence or presence of borate
oxidation in vitro. These systems were used as positive control.Absorption spectra of the respective proteins were also taken,which served as control.
2.4. Preparation of borax stock solution (133 mM)
The stock solution of borax was prepared by dissolving 1.268 gborax in 20 ml water or glycine–NaOH buffer (1 M, pH 9.2). pH ofthe mixture was adjusted to 9.2, and the final volume was made upto 25 ml in distilled water or the desired buffer. Molarity of borax inthe final reaction mixture containing the protein was 40 mM, which
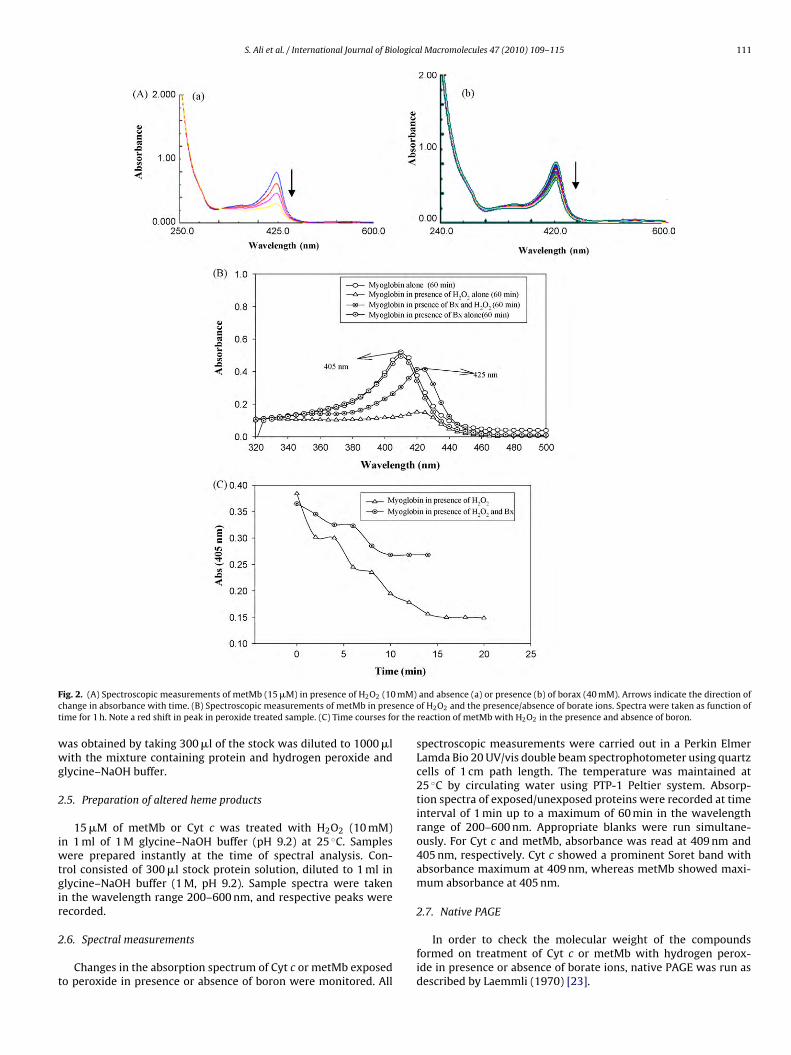
S. Ali et al. / International Journal of Biological Macromolecules 47 (2010) 109–115 111
F mM)c encet r the
wwg
2
iwtgir
2
t
ig. 2. (A) Spectroscopic measurements of metMb (15 �M) in presence of H2O2 (10hange in absorbance with time. (B) Spectroscopic measurements of metMb in presime for 1 h. Note a red shift in peak in peroxide treated sample. (C) Time courses fo
as obtained by taking 300 �l of the stock was diluted to 1000 �lith the mixture containing protein and hydrogen peroxide and
lycine–NaOH buffer.
.5. Preparation of altered heme products
15 �M of metMb or Cyt c was treated with H2O2 (10 mM)n 1 ml of 1 M glycine–NaOH buffer (pH 9.2) at 25 ◦C. Samples
ere prepared instantly at the time of spectral analysis. Con-rol consisted of 300 �l stock protein solution, diluted to 1 ml inlycine–NaOH buffer (1 M, pH 9.2). Sample spectra were takenn the wavelength range 200–600 nm, and respective peaks wereecorded.
.6. Spectral measurements
Changes in the absorption spectrum of Cyt c or metMb exposedo peroxide in presence or absence of boron were monitored. All
and absence (a) or presence (b) of borax (40 mM). Arrows indicate the direction ofof H2O2 and the presence/absence of borate ions. Spectra were taken as function ofreaction of metMb with H2O2 in the presence and absence of boron.
spectroscopic measurements were carried out in a Perkin ElmerLamda Bio 20 UV/vis double beam spectrophotometer using quartzcells of 1 cm path length. The temperature was maintained at25 ◦C by circulating water using PTP-1 Peltier system. Absorp-tion spectra of exposed/unexposed proteins were recorded at timeinterval of 1 min up to a maximum of 60 min in the wavelengthrange of 200–600 nm. Appropriate blanks were run simultane-ously. For Cyt c and metMb, absorbance was read at 409 nm and405 nm, respectively. Cyt c showed a prominent Soret band withabsorbance maximum at 409 nm, whereas metMb showed maxi-mum absorbance at 405 nm.
2.7. Native PAGE
In order to check the molecular weight of the compoundsformed on treatment of Cyt c or metMb with hydrogen perox-ide in presence or absence of borate ions, native PAGE was run asdescribed by Laemmli (1970) [23].

1 ological Macromolecules 47 (2010) 109–115
2
t(
2
mTT1i
2p
wamDiCfoktbDf
3
3a
9i1(tcdss
Fig. 3. (A) Spectroscopic analysis of RNase in presence of H2O2 and borate ions.
Fi
12 S. Ali et al. / International Journal of Bi
.8. Gel filtration
To check further the nature of product(s) formed, gel filtra-ion chromatography was performed on Sephadex G-200 column2.2 cm × 45 cm).
.9. High performance liquid chromatography (HPLC)
The HPLC of treated/untreated samples was performed in “Schi-adzu SCL-10AVP” fitted with a C-18 (5 micron) analytical column.
he solvent system consisted of acetonitrile (0.1% TFA)/water (0.1%FA); TFA: triflouroacetic acid. The flow rate was maintained atml/min, and the time gradient was 1%/min (5–70% actetonitrile
n 5–70 min).
.10. Dynamic light scattering (DLS) analysis of the altered hemeroduct
DLS measurements of the exposed/unexposed heme proteinsere done in a Laser-Spectroscatter 201(RiNA) at 25 ◦C. Data was
nalysed using PMgrv 3.01p17 software supplied with the instru-ent. All chemicals were filtered through 0.1 �m mesh size filters.iode laser of wavelength 660 nm was used as a source. The result-
ng autocorrelation functions were analysed with the programONTIN to obtain hydrodynamic radii distribution. Refractive index
or the globular molecules was 1.330. Light scattered at a fixed anglef 90◦ was collected by a filter/monochromator or interferometer,ept in front of the photomultiplier tube. Concentration of the pro-ein was 10 mg/ml, and the concentrations of H2O2 (10 mM) andorax (40 mM) were kept the same as in the spectroscopic studies.iffusion coefficient was calculated into the hydrodynamic radius
or a sphere through the Stokes–Einstein equation.
. Results
.1. Absorption spectra of proteins exposed to peroxide inbsence or presence of boron
The absorption spectra of Cyt c alone in glycine–NaOH buffer (pH.2) in the wavelength range 200–600 nm showed a single peak
n Soret (�) region at 409 nm (Fig. 1A). Treatment of Cyt c with0 mM H2O2 caused a decrease in the absorbance in Soret region409–410 nm) and a blue shift was observed that led to a shift in
he peak from 409 nm to 407 nm (Fig. 1B). Addition of peroxide toytochrome c pretreated with borax reported a considerably lessecrease in absorbance at 409 nm as compared to control (Fig. 1C),uggesting the protective effect of borate ions on the three dimen-ional structural stability of protein. Treatment of metmyoglobinig. 4. Native PAGE analysis of Cyt c (A), metMb (B) and RNase-A(C) in glycine–NaOH bun presence of H2O2 and in presence of H2O2 and borate ions.
Spectra were taken as function of time. (B) Time courses for the reaction of RNAseA with hydrogen peroxide in presence or absence of boron showed no change inabsorbance with time.
with peroxide also showed similar changes. There was a decreasein absorbance in the Soret region (408–410 nm), and a red shiftwas observed with a shift in peak from 406 nm to 420 nm in pres-ence of hydrogen peroxide (Fig. 2A–C). Metmyoglobin pretreated
with boron was found to be considerably more stable. On the otherhand, no change in absorbance at 280 nm was recorded when thenon-heme protein RNAse A was treated with hydrogen peroxide inpresence or absence of borax (Fig. 3A and B).ffer at pH 9.2. Lanes 1, 2 and 3 represent proteins, respectively, in absence of H2O2,

S. Ali et al. / International Journal of Biological Macromolecules 47 (2010) 109–115 113
F4a
3
eFbePiPIai(o(i(
3p
2piTc
ig. 5. (A) Elution profile of Cyt c treated with H2O2 in presence/absence of boron at10 nm. (B) Elution profile of Cyt c treated with H2O2 in presence/absence of boront 280 nm.
.2. Native PAGE analysis
The native PAGE revealed the formation of specific products onxposure of proteins in presence or absence of boron. Lane 1 inig. 4A shows the native PAGE of Cyt c; the protein appears as aand corresponding to molecular mass of 12 kDa. However, in pres-nce of H2O2, an aggregate of 38–46 kDa (Lane 2) was found. TheAGE analysis of Cyt c pretreated with boron and exposed to perox-de showed a 25-kDa band (Lane 3). On the other hand, the nativeAGE analysis of metmyoglobin showed a 17-kDa band (Lane 1).ntensity of the band decreased in presence of hydrogen peroxide;streak was observed in the region corresponding to 30–38 kDa,
ndicating the formation of higher molecular mass compoundsLane 2). Native PAGE of metMb exposed to peroxide in presencef boron showed an increase in band (22 kDa) intensity (Lane 3)Fig. 4B). The native PAGE of non-heme protein exposed to perox-de did not show any change either in presence or absence of boronFig. 4C).
.3. Gel filtration analysis of heme/non-heme proteins exposed toeroxide in presence or absence of boron
Elution profile of Cyt c (treated/untreated) on a sephadex G-
00 column is shown in Fig. 5. Fig. 5A and B represents the elutionrofile of Cyt c alone, and Cyt c treated with hydrogen peroxiden presence or absence of borax at 409 and 280 nm, respectively.reatment with peroxide affected the elution profile of cytochrome. Similar results were found with metMb (Fig. 6A and B). No change
Fig. 6. (A) Elution profile of metMb treated with H2O2 in presence/absence of boronat 410 nm. (B) Elution profile of metMb treated with H2O2 in presence/absence ofboron at 280 nm.
in the elution profile of non-heme proteins was recorded (data notshown).
3.4. HPLC
HPLC of Cyt c, Cyt c exposed to H2O2 and Cyt c exposed to per-oxides in presence/absence of boron was performed (Fig. 7A–C).Untreated Cyt c appeared as monomer. However, presence of sto-ichiometric amount of peroxide resulted in more than one peak,suggesting an aggregate. In presence of boron, HPLC profile of Cytc exposed to peroxides revealed two peaks. The HPLC profile ofmetMb alone, metMb in presence of H2O2, and metMb in pres-ence of borax and H2O2 is shown in Fig. 7D–F. Presence of a singlepeak suggests monomer. Treatment of metMb with hydrogen per-oxide revealed more than one peak, suggesting the formation ofan aggregate. HPLC profile of metmyoglobin in presence of hydro-gen peroxide and borax revealed two peaks (dimer). However, itis difficult to ascertain whether the peaks correspond to a dimer!Subsequent study on DLS proved the existence of dimer.
3.5. DLS
DLS measurements clearly distinguish between monomer,dimer or aggregate forms (Table 1). The hydrodynamic radius (RH)of Cyt c alone was 1.38 nm, which suggests a monomer. The valueof RH for the protein exposed to peroxide was 5.88 nm, indicat-ing an 18-mer product. However, in presence of boron, RH was2.68 nm, which suggests a dimer. The RH value of untreated metMbin glycine–NaOH buffer at pH 9.2 was 1.80 nm, suggesting the exis-
tence of monomer at pH 9.2. However, in presence of H2O2 inglycine–NaOH buffer at pH 9.2, RH was 47.65 nm, suggesting theformation of a high molecular weight aggregate at pH 9.2. In pres-ence of peroxide and borax in glycine–NaOH buffer at pH 9.2, RHwas 2.50 nm, suggesting the existence of metMb as a dimer.
114 S. Ali et al. / International Journal of Biological Macromolecules 47 (2010) 109–115
F LC prH D) HPLm 2 at pw ent sy
4
aaavspTg
TTli
NaacH
ig. 7. (A) HPLC profile of untreated Cyt c at pH 9.2 in glycine–NaOH buffer. (B) HP2O2 at pH 9.2 for 30 min. (C) HPLC profile of Cyt c in presence of borax and H2O2. (etMb in presence of H2O2. Reaction mixture consisted of metMb treated with H2Oere injected onto an HPLC column (Schimadzu SCL-10AVP) equilibrated with solv
. Discussion
Iron-containing proteins have been found susceptible to dam-ge by peroxides, which are usually produced in living systems andct by various mechanism such as by causing the cross-linking of anctive site tyrosine residue to the heme [12]. In this study, we pro-ide experimental evidence to show that boron in the form of borax
tabilizes the three dimensional structure of oxidized Fe-containingroteins, Cyt c and metMb, exposed to hydrogen peroxide in vitro.he protein was characterized by visible absorption spectroscopy,el exclusion chromatography, native PAGE, HPLC and dynamicable 1he hydrodynamic radii (RH) of various heme proteins obtained through dynamicight scattering (DLS) of cytochrome c and myoglobin treated with hydrogen perox-de in presence or absence of borate ions.
Proteins Hydrodynamic radii
Untreated H2O2 Borate ion + H2O2
Cytochrome c 1.38 nm 5.89 nm 2.68 nmMyoglobin 1.80 nm 47.65 nm 2.50 nm
ote the formation of dimer in case of iron-containing proteins (both cytochrome cnd myoglobin) exposed to peroxide in presence of boron. Untreated cytochrome cnd myoglobin appeared as monomers, while in presence of peroxide, cytochromeresulted in the formation of an 18-mer and metmyoglobin in the formation of anMW: high molecular weight.
ofile of Cyt c in presence of H2O2. Reaction mixture consisted of Cyt c treated withC profile of untreated metMb at pH 9.2 in glycine–NaOH buffer. (E) HPLC profile of
H 9.2 for 30 min. (F) HPLC profile of metMb in presence of borax and H2O2. Samplesstem acetonitrile (0.1%TFA)/water (0.1%TFA) at a flow rate of 1 ml/min.
light scattering. Oxidized forms of these proteins were treated withborax followed by exposure to hydrogen peroxide. Spectroscopicmeasurements on Cyt c and metMb clearly show that peroxidedestabilizes the structure of native Cyt c and metMb (Figs. 1,2).However presence of borate ions considerably minimized the dam-age. In presence of boron, the exposed heme proteins showed Soretspectra near the control value (Fig. 1A). The Soret spectra of theheme proteins were recorded at regular time interval, and the per-oxide mediated damage was found to be time dependent (Fig. 1C).Boron could counteract the effect of peroxide. A possible explana-tion might be the ability of boron to prevent free radical mediateddamage in vivo, which has been demonstrated earlier [22,24].
Size exclusion chromatography of iron-containing proteinsexposed to peroxides indicates a change in the elution volume (Ve)of Cyt c (Fig. 5A and B). Ve of untreated Cyt c was 145 ml, whilein presence of H2O2, it was 122 ml. This led us to hypothesize theformation an aggregate or a high molecular weight (HMW) prod-uct. These findings are in agreement with earlier reports indicatingthat the exposure of heme proteins to H2O2 causes aggregation orcross-linking reactions [12]. This study further reveals that Ve ofCyt c in presence of borate ions followed by the peroxide treatment
was 133 ml, indicating the formation of an intermediate molecularweight product and the prevention of aggregation to a significantextent. For untreated metMb, the elution volume (136 ml) changedto 115 ml in presence of H2O2, and in presence of borate ions, itwas found to be 123 ml (Fig. 6A and B). No change in elution profile
logica
wNubnt
tuo((IwattwmtHaa9ho
sriorlotwtrharwCopscstodobwo
b‘ifTwost
[
[[
[[
[[[[
[[[
[[[[25] K.N. King, M.E. Winfield, Aust. J. Biol. Sci. 19 (1966) 211–217.
S. Ali et al. / International Journal of Bio
as seen when non-heme protein RNase was exposed to peroxide.ative PAGE analysis provided further information on the molec-lar weights of various products formed in presence or absence oforon when the heme proteins were exposed to peroxides (Fig. 4);on-iron protein did not show any sign of damage when exposedo peroxide (Fig. 4C).
HPLC and DLS experiments provided convincing evidence forhe role of borate ions as a structural stabilizer of heme proteinsnder oxidative stress conditions. When exposed to peroxide, thexidized heme protein resulted in the formation of an aggregateTable 1). HPLC of Cyt c treated with H2O2 alone revealed four peaksFig. 7B, peaks I–IV) suggesting the formation of an aggregate. PeaksII and IV have been previously described [12,16]. Similar results
ere obtained with metMb exposed to peroxides in presence orbsence of boron (Fig 7E and F). In presence of boron, peroxidereatment resulted in two peaks. However, it was difficult to ascer-ain whether the peak corresponds to a dimer! DLS measurementsere carried out to ascertain this difficulty. Results of DLS measure-ents showed the presence of an 18-mer in case of Cyt c exposed
o H2O2 and a dimer (2.68 nm), when the protein was exposed to2O2 in presence of borate ions. Similarly, in case of metMb, a HMWggregate (47.5 nm) was formed in presence of peroxide alone, anddimer (2.50 nm) was found in presence of borate and H2O2 at pH.2. The hydrodynamic radii from the DLS measurements of variouseme proteins suggest the formation of monomer, dimer, 18-merr a higher-mer aggregate under different conditions (Table 1).
The oxidant attack on proteins has been reported to causeite-specific amino acid modifications, aggregation of cross-linkedeaction products, altered electrical charge, increased susceptibil-ty to proteolysis, and fragmentation of the peptide chain. metMb,ne of the best characterized heme protein, for example has beeneported to react with H2O2 and lead to intermolecular cross-inking of myoglobin to form oligomers, or between myoglobin andther molecules as well as to the cross-linking of heme to the pro-ein at tyrosine residue 103 [25]. Initial steps in the reaction of H2O2ith metMb have been extensively investigated [13,26], and appear
o involve the formation of a Fe14OH heme species and hydroxyladical equivalent, which could be localized on the protein or theeme. When the radical is localized on the protein, an aromaticmino acid radical is formed, which most likely involve tyrosineesidues. The protein radical can lead to cross-linking reactionsith other proteins [26] and organic molecules [25], or with heme.ross-linking of heme to the protein moiety has been reported toccur by the attack of a tyrosine radical (residue 103) to the heme,ossibly at meso position. Boron can probably act by binding specificites on the heme proteins and counteracting the attack. The exacthemistry behind the reaction, however, remains elusive. In earliertudies [27], borate ions have been reported to stabilize the struc-ure of �-galactosidase, a non-heme glycoprotein from Aspergillusryzae under stress conditions. Borate ions bind to the carbohy-rate moiety of �-galactosidase, which changes the conformationf protein without inactivating the enzyme. Borate ions might sta-ilize the molecular structure of some polyhydroxy compounds,hich appear to compete favourably with the carbohydrate moiety
f glycoprotein.The molecular dynamics simulations have revealed distinct
inding sites on heme proteins such as cytochrome c. These areloose binding sites’ and ‘tight binding sites’. Loose binding sitesnclude (Lys25/Lys27-I site and Lys55/Lys73-II site) [28]. Borate ionorms two to three hydrogen bonds to the nearest lysine residue.
he binding is transient. The ion binds to a single lysine residueith three hydroxyl groups pointing towards the amine nitrogenf lysine, forming three hydrogen bonds. This demonstrates the fea-ibility of fast transfer of ion from one such site to another (Lys55
o Lys73) or fast exchange of such loosely bound ions with free ions
[[[[
l Macromolecules 47 (2010) 109–115 115
in solution, and suggests that this ion is nonspecific, and, therefore,can attach to Lys25, Lys27, Lys55, or Lys73. It has been reported fur-ther that the third site for binding of boron is near Lys13 and Lys86
of the native cytochrome c [28]. The site was postulated as the tightbinding site based on its relative proximity to the heme allowing. Ithas been reported that this surface site, which binds to borate iontightly is also the known binding site of Pi [29]. We propose thatbinding of borate ions to heme proteins probably changes the con-formation of protein to a more stable form that can help counteractthe damaging effect of peroxides. Since binding is transient at twosites (Lys25/Lys27-I site and Lys55/Lys73-II), a complete preventionof oxidative damage could not be seen.
5. Conclusions
This study reports that boron stabilizes the structure of hemeproteins under oxidative stress conditions in vitro. The study hasboth industrial and clinical implications as boron can be used tostabilize the peroxide-sensitive heme proteins; earlier studies fromour lab have reported boron to prevent tissue injury under oxida-tive stress conditions in animal model. The exact chemistry of thestabilizing effect, however, cannot be deduced at this stage. Hemeproteins are suggested to possess sites that can bind to borate ions.
Acknowledgments
The research work presented in this paper is a part of the PhDthesis of HF. HF is currently working as Assistant Professor in theDepartment of Biotechnology, Jamia Hamdard, and acknowledgesthe support of Prof. MZ Abdin, Head of the Department of Biotech-nology. UGC and DST (FIST) are acknowledged for providing fundsfor infrastructure development to the Department of Biochemistry.
References
[1] P.I. Moreira, Akihiko Nunomura, Xiongwei Zhu, Hyoung-Gon Lee, GjumrakchAliev, A. Mark, Smith, George Perry, in: S Vearsy (Ed.), Oxidative Neural Injury,Humana Press, 2009, pp.109–120.
[2] R.S. Sohal, Free Radical Biol. Med. 33 (2002) 37–44.[3] J.A. Jesberger, J.S. Richardson, Int. J. Neurosci. 57 (1991) 1–17.[4] S. Ali, S.K. Jain, M. Abdulla, M. Athar, Biochem. Mol. Biol. Int. 39 (1996) 63.[5] J.M. Gutteridge, B. Halliwell, Trends Biochem. Sci. 15 (1990) 129–135.[6] P. Cerutti, Lancet 334 (1994) 862–863.[7] B. Demple, A. Johnson, D. Fung, Proc. Natl. Acad. Sci. (1986) 7331–7735.[8] R.I. Hall, G. Dawber, A. McConkey, M.A. MacDonald, G.C. King, Phys. Rev. Lett.
68 (1992) 2751–2754.[9] D.J. Kelman, J.A. DeGray, R.P. Mason, J. Biol. Chem. 269 (1994) 7458–7463.10] N.K. King, F.D. Looney, M.E. Winfield, Biochim. Biophys. Acta 133 (1967) 65–
82.11] H.H. Ruf, U. Raab-Brill, C. Blau, Biochem. Soc. Trans. 21 (1993) 739–744.12] C.E. Catalano, Y.S. Choe, P.R. Ortiz de Montellano, J. Biol. Chem. 264 (1989)
10534–10541.13] D. Tew, P.R. Ortiz de Montellano, J. Biol. Chem. 263 (1988) 17880–17886.14] W.L. Smith, T.E. Eling, R.J. Kulmacz, L.J. Marnett, A. Tsai, Biochemistry 31 (1992)
3–7.15] M.J. Davies, Free Radical Res. Commun. 10 (1990) 361–370.16] Y. Osawa, K. Korzekwa, Proc. Natl. Acad. Sci. U.S.A. 88 (1991) 7081–7085.17] L. Cantoni, A.H. Gibbs, F. De Matteis, Int. J. Biochem. 13 (7) (1981) 823–830.18] S.E.G. Fligiel, E.C. Lee, J.P. McCoy, K.J. Johnson, J. Varani, Am. J. Pathol. 115 (1984)
418–425.19] K.J.A. Davies, S.W. Lin, R.E. Pacifici, J. Biol. Chem. 262 (1987) 9914–9920.20] S. Gregory, N.D. Kelly, Alt. Med. Rev. 2 (1) (1997) 48–56.21] W. Jerry Spears, Todd Armstrong, in: F Nu et al (Ed.), Advances in Plant and
Animal Boron nutrition, Springer, Netherlands, 2007, pp. 269–276.22] S. Ali, Trace Elem. Med. 7 (2) (2006) 57–62.23] U.K. Laemmli, Nature 227 (1970) 680–685.24] R.E. Newnham, Int. Clin. Nutr. Rev. 11 (1991) 68–70.
26] P. George, D.H. Irvine, Biochem. J. 52 (1952) 511–517.27] S. Shubhada, P.V. Sundaram, Enzyme Microb. Technol. 15 (10) (1993) 881–886.28] G. Taler, G. Navon, M. Becker, Biophys. J. 75 (1998) 2461–2468.29] G.W. Pettigrew, G.R. Moore, Cytochrome c: Biological Aspects, Springer-Verlag,
Berlin/Heidelberg/New York, 1987.
Copyright © 2022 FDOKUMEN




















