
Distribution of recent benthic foraminifera in shelf carbonate environments of the Western...
13
MarineGeology 134(1996) 113-125 ELSEVIER Distribution of recent benthic foraminifera in a subarctic fjord-delta: Tana, Norway Geoffrey D. Corner *, Per Ivar Steinsund, Roald Aspeli Department of Geology, IBG, University of Tromsg, N-9037, Tromse, Norway Received 3 November 1995; accepted 3 May 1996 Abstract Analysis of benthic foraminifera (living + dead fauna) in surface sediments from a deltaic and fjord environment at Tana, northern Norway, shows a marked contrast in foraminiferal assemblages between delta platform and delta slope-fjord basin provinces, and a correspondence between test abundance, percentage living of total foraminifera and inferred rate of sedimentation. The relatively shallow (<25 m), high-energy, sandy delta platform province is characterized by an absence or low abundance of foraminifera and a dominance of calcareous forms. Delta plain and tidal strait environments of this province contain a sparse Elphidium albiumbilicatum assemblage and a resedimented Cibicides iobatulus assemblage, respectively. The deeper (25-125 m), lower energy, silty-sandy to sandy-muddy delta slope-fjord basin province is characterized by a relatively high abundance of foraminifera and a dominance of agglutinated forms comprising an Adercotryma glomerata-Spiroplectammina biformis assemblage. Foraminiferal abundance in the delta slope-fjord basin province tends to increase with increasing depth and distance from the river mouth, suggesting a correlation with sedimentation rate. A generally high percentage of living foraminifera is thought to mainly reflect high sedimentation rates in the deltaic environment. Local variations in foraminiferal abundance, percentage living foraminifera, and fauna1 composition are ascribed mainly to local. variations in sedimentation rates and processes, and postmortem changes. Key words: Benthic foraminifera; Fjord; Delta; Norway 1. Introduction The Tana delta is a sandy, steep-faced, fjord- head delta prograding into Tanafjord, arctic Norway (70”31’N, 28”24’E), (Fig. 1). The distribu- tion of living and dead benthic foraminifera in surface sediments of the delta and adjacent fjord area was analysed to study the relationship * Corresponding author. Fax: +47 77 64 56 00. E-mail: [email protected] 0025-3227/96/$15.00 0 1996 Elsevier Science B.V. All rights reserved PZZ SOO25-3227(96)00039-4 between fauna1 assemblages and depositional envi- ronment, to estimate relative rates of sedimenta- tion, and to provide a comparative basis for future stratigraphic studies. Deltas do not form a distinct environment for foraminiferal fauna, but rather an assemblage of environments such as occur elsewhere (Murray, 1973). At Tana, this assemblage includes intertidal delta plain, steep delta slope, and deep fjord basin depositional environments. Previous foraminiferal studies of such environments, and of steep-faced
-
Upload
uni-hamburg -
Category
Documents
-
view
0 -
download
0
Transcript of Distribution of recent benthic foraminifera in shelf carbonate environments of the Western...

MarineGeology 134(1996) 113-125 ELSEVIER
Distribution of recent benthic foraminifera in a subarctic fjord-delta: Tana, Norway
Geoffrey D. Corner *, Per Ivar Steinsund, Roald Aspeli
Department of Geology, IBG, University of Tromsg, N-9037, Tromse, Norway
Received 3 November 1995; accepted 3 May 1996
Abstract
Analysis of benthic foraminifera (living + dead fauna) in surface sediments from a deltaic and fjord environment at Tana, northern Norway, shows a marked contrast in foraminiferal assemblages between delta platform and delta slope-fjord basin provinces, and a correspondence between test abundance, percentage living of total foraminifera and inferred rate of sedimentation.
The relatively shallow (<25 m), high-energy, sandy delta platform province is characterized by an absence or low abundance of foraminifera and a dominance of calcareous forms. Delta plain and tidal strait environments of this province contain a sparse Elphidium albiumbilicatum assemblage and a resedimented Cibicides iobatulus assemblage, respectively. The deeper (25-125 m), lower energy, silty-sandy to sandy-muddy delta slope-fjord basin province is characterized by a relatively high abundance of foraminifera and a dominance of agglutinated forms comprising an Adercotryma glomerata-Spiroplectammina biformis assemblage.
Foraminiferal abundance in the delta slope-fjord basin province tends to increase with increasing depth and distance from the river mouth, suggesting a correlation with sedimentation rate. A generally high percentage of living foraminifera is thought to mainly reflect high sedimentation rates in the deltaic environment. Local variations in foraminiferal abundance, percentage living foraminifera, and fauna1 composition are ascribed mainly to local. variations in sedimentation rates and processes, and postmortem changes.
Key words: Benthic foraminifera; Fjord; Delta; Norway
1. Introduction
The Tana delta is a sandy, steep-faced, fjord-
head delta prograding into Tanafjord, arctic
Norway (70”31’N, 28”24’E), (Fig. 1). The distribu-
tion of living and dead benthic foraminifera in surface sediments of the delta and adjacent fjord area was analysed to study the relationship
* Corresponding author. Fax: +47 77 64 56 00. E-mail: [email protected]
0025-3227/96/$15.00 0 1996 Elsevier Science B.V. All rights reserved PZZ SOO25-3227(96)00039-4
between fauna1 assemblages and depositional envi-
ronment, to estimate relative rates of sedimenta-
tion, and to provide a comparative basis for future
stratigraphic studies.
Deltas do not form a distinct environment for
foraminiferal fauna, but rather an assemblage of environments such as occur elsewhere (Murray, 1973). At Tana, this assemblage includes intertidal delta plain, steep delta slope, and deep fjord basin depositional environments. Previous foraminiferal studies of such environments, and of steep-faced

114 G.D. Corner et al.lMarine Geology 134 (1996) 113-125
Fig. 1. Location map of the Tana delta and adjacent fjord showing depositional environments, foraminiferal sampling sites, dated core sites, and hydrographic station in Leirpollen. The delta and its feeder system are stippled. Full sample numbers are given in Table 1. Isobaths (constructed from sea chart no. 110) and spot heights are in metres.
deltas in particular, are limited. They include work 1990; Alve, 1990,1991,1995) and western Norway on the Frazer River delta of western Canada (Austin and Sejrup, 1994). In north Norwegian (Williams, 1989; see references in Lutenauer et al., fjord areas, foraminiferal studies have focussed 1994) and fjords of North America (Lagoe et al., almost exclusively on Late Quaternary faunas, and 1994), Greenland (Jennings and Helgadottir, recent benthic foraminifera are described only in 1994), southern Norway (Alve and Nagy, 1986, some early (pre-1945) studies (see references in

G.D. Corner et al./Marine Geology 134 (1996) 113-125 115
Hald, 1986) and in a more recent, unpublished study from Altafjord (Strand, 1979).
2. Depositional environment
The delta lies at the mouth of the Tana River and is currently prograding into Tanafjord along its northeastern flank. A previously formed part of the delta extends eastwards into the semi- enclosed Leirpollen fjord basin (Fig. 1).
Four main depositional environments in two sub-areas (Tanafjord and Leirpollen) are recog- nized on the basis of morphology: (1) a wave- influenced, predominantly intertidal ( 1.5-2.5 m tidal range) delta plain, (2) an 8-15 m deep tidal strait connecting Leirpollen with Tanafjord, (3) steep (20-2”) delta slopes descending from the delta rim at 5 m below sea level to the fjord floor at a depth of 30-40 m in Leirpollen and about 175 m in Tanafjord, and (4) fjord basin comprising the sides and floor of the fjord beyond the delta slope in Tanafjord and Leirpollen. The delta plain and tidal strait are relatively shallow, high-energy environments comprising what is here termed the delta platform province to distinguish them from deeper, lower energy environments of the delta slope-fjord basin province.
Sediment is transported to the delta and fjord basin predominantly during spring snowmelt river flood, and is deposited on and beyond the delta slope mainly from suspension fallout. Reworking by waves and tidal currents prevails on the delta platform, and redeposition by gravity mass trans- port processes is probably important locally on the upper and middle delta slope. In general, sedimentation rates decrease away from the river mouth, as indicated by basinward-thinning bed- ding geometry seen on seismic profiles (Corner et al., 1994; Mauring et al., 1995), and by distal fining of sediment which ranges from sand on the delta plain and in the tidal strait, through silty sand and muddy sand on the delta slope, to sandy mud in the fjord basin. Acoustic profiling shows that the boundary between the delta slope and adjacent fjord basin is generally distinct, reflecting probably downslope creep.
Radiocarbon dated sediment cores from two
sites just beyond the delta slope in Tanafjord and Leirpollen (sites a and b, Fig. 1) indicate average sedimentation rates of 0.5-2.5 mm/yr during the last 1500-4500 years. Present sedimentation rates in Leirpollen are probably lower than both the obtained Late Holocene average as well as present rates generally in Tanafjord, since the delta front in Leirpollen is no longer active. Sedimentation rates on the upper delta slope are probably at least an order of magnitude higher than in the fjord basin, judging from a comparison with other north Norwegian deltas (Corner et al., 1990; Comer and Fjalstad, 1993).
The area has, comparable to other inner fjord areas of northern Norway (NordgBrd et al., 1982), an estuarine hydrography with a fresh to brackish surface layer (2-5 m thick) in summer, overlying brackish to marine water reaching year round salinity values of 33-34%0 below a depth of about 20 m (Fig. 2). Bottom water in Leirpollen is relatively well oxygenated (c. 5 ml/l, Fig. 2) sug- gesting annual replenishment. The temperature of
E 20 c E. 0” t P t/i
30
-u- Temperature
4 5 6 7 8 9 “C 25 27 29 31 33 35 %0 4.5 4.7 4.9 5.1 5.3 5.5 ml/l
Fig. 2. Vertical variation in salinity, temperature, and dissolved
oxygen in Leirpollen (site b, Fig. 1) measured during
foraminiferal sampling, 7 October 1992.

116 G.D. Corner et aLlMarine Geology 134 (1996) 113-125
the marine water is estimated to range seasonally between 2 and 8°C (cf. Nordgard et al., 1982).
3. Samples and methods
A total of 21 surface sediment samples were collected for foraminiferal analysis from various areas of the delta and adjacent fjord basin, at depths ranging from sea level to 124 m water depth (Fig. 1, Table 1). Delta plain samples (T93-1 to T93-7) were collected on 17-19 September 1993, subaqueous samples (T92-20 to T92-42) on 7-8 October 1992.
At each site, 2 dl samples were taken from a c. 150 cm2 area of the uppermost 1-2 cm sediment layer, either directly from wet in situ sediment (delta plain sites), or from undisturbed samples retrieved using a Van Veen grab (subaqueous sites). Samples were placed immediately in a 2 dl staining solution of rose Bengal (2 g/l) and alcohol (96%), preserved for three weeks, then dried at 40°C weighed, disaggregated in water, and washed through 0.1 and 1 mm mesh diameter sieves. The residue on the 0.1 mm sieve was dried and the foraminifera concentrated by flotation using sodium polytungstate diluted with water to a specific gravity of 1.8. A total of c. 250-290 tests (stained + unstained) was counted from each sample, except in the case of five low abundance samples in which all available specimens were counted (Table 2). Stained and unstained speci- mens are considered synonymous with living and dead fauna, respectively (Walton, 1952; Bernhard, 1988). Examining samples dry, rather than wet, should not have significantly affected the reliability of identifying stained samples, since species having thick-walled tests were uncommon. Drying samples will have caused some fragile agglutinated species to disintegrate, although this should have affected all samples equally. The results, after drying, should also be more comparable with results from standard stratigraphic analyses (Feyling-Hanssen, 1964; Meldgaard and Knudsen, 1979). Cluster analysis was performed using SYSTAT; distance measure 1 -Pearson correlation coefficient, complete linkage method (farthest neighbour).
Grain-size analysis of surface sediment samples was carried out using standard procedures (wet sieving and sedigraph). Total organic carbon was measured on separate samples from 2-5 cm below the sediment surface using a Leco induction oven.
4. Results
4.1. Foraminiferal abundance
Foraminiferal abundance, expressed as number of tests per gram dry sediment, varies considerably in different parts of the sampled area (Fig. 3, Table 1).
Lowest abundance was found on the intertidal delta plain, where 5 out of 7 samples were barren. Foraminifera were rare (~0.2 tests per gram) in the remaining two samples from the delta plain which derive, respectively, from a permanently inundated, ripple-bedded, fluviotidal channel on the lower delta plain (sample T93-l), and a rela- tively wave-sheltered, fine-sandy flat on the middle delta plain (sample T93-7). Sample T92-31 from the tidal strait (12 m depth) was barren and sample T92-32 from the tidal strait mouth (23 m depth) contained few (0.5 tests per gram) foraminifera.
Samples from the delta slope-fjord basin prov- ince, spanning depths of 26-124 m, contained mostly frequent to abundant foraminifera (2.6- 113 tests per gram in 10 out of 12 samples). Particularly high abundance (68- 113 tests per gram) was found at two sites in Leirpollen (T92-20, T92-21) and at a distal site in Tanafjord (T92-42). Sample T92-34, from the proximal delta slope in Tanafjord, con- tained few foraminifera (0.8 tests per gram), and sample T92-23, from the delta slope in Leirpollen, contained rare foraminifera (0.05 tests per gram). Abundance among delta slope-fjord basin samples correlates positively with increasing water depth, decreasing grain size, and increasing total organic carbon content (Fig. 4A).
4.2. Percentage living fauna
The percentage living (stained) of total foramini- feral fauna ranges from 7% to 88%, and averages 42%. Relatively low values (< 15%) are shown by
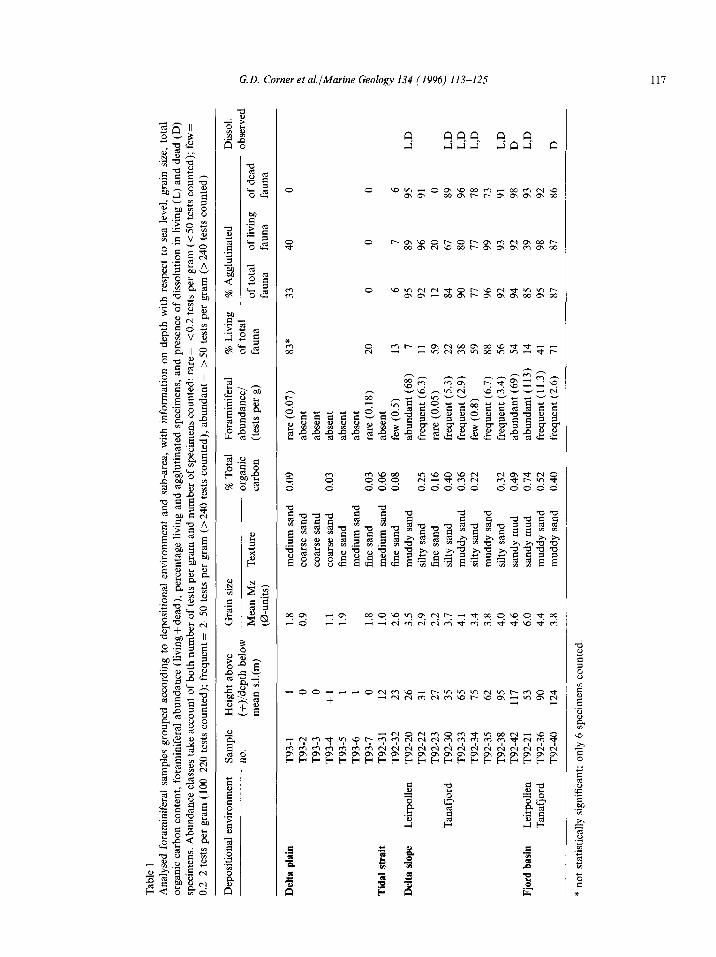
Tab
le
1
Ana
lyse
d fo
ram
inif
eral
sa
mpl
es
grou
ped
acco
rdin
g to
de
posi
tiona
l en
viro
nmen
t an
d su
b-ar
ea,
with
in
form
atio
n on
de
pth
with
re
spec
t to
se
a le
vel,
grai
n si
ze,
tota
l
orga
nic
carb
on
cont
ent,
fora
min
ifer
al
abun
danc
e (l
ivin
g +
dead
),
perc
enta
ge
livin
g an
d ag
glut
inat
ed
spec
imen
s,
and
pres
ence
of
di
ssol
utio
n in
liv
ing
(L)
and
dead
(D
)
spec
imen
s.
Abu
ndan
ce
clas
ses
take
ac
coun
t of
bot
h nu
mbe
r of
tes
ts
per
gram
an
d nu
mbe
r of
sp
ecim
ens
coun
ted:
ra
re
= ~0
.2
test
s pe
r gr
am
(i
50 t
ests
co
unte
d);
few
=
0.22
2 te
sts
per
gram
(l
oo-2
20
test
s co
unte
d);
freq
uent
=
2-50
te
sts
per
gram
(>
240
te
sts
coun
ted)
, ab
unda
nt
= >
50 t
ests
pe
r gr
am
(>
240
test
s co
unte
d)
Dep
ositi
onal
en
viro
nmen
t Sa
mpl
e H
eigh
t ab
ove
Gra
in
size
%
Tot
al
Fora
min
ifer
al
% L
ivin
g %
Agg
lutin
ated
D
isso
l.
no.
(+)/
dept
h be
low
or
gani
c ab
unda
nce/
of
to
tal
obse
rved
mea
n s.
l.(m
) M
ean
Mz
Tex
ture
ca
rbon
(t
ests
pe
r g)
fa
una
of
tota
l of
liv
ing
of
dead
(0-u
nits
) fa
una
faun
a fa
una
Del
ta
plai
n T
93-1
1
T93
-2
0
T93
-3
0
T93
-4
+1
T93
-5
1
T93
-6
1
T93
-7
0
Tid
al
stra
it T
92-3
1
12
T92
-32
23
Del
ta
slop
e L
eirp
olle
n T
92-2
0 26
T92
-22
31
T92
-23
27
Tan
afjo
rd
T92
-30
35
T92
-33
65
T92
-34
75
T92
-35
62
T92
-38
95
T92
-42
117
Fjor
d ba
sin
Lei
rpol
len
T92
-21
53
Tan
afjo
rd
T92
-36
90
T92
-40
124
1.8
0.9
1.1
1.9
1.8
1.0
2.6
3.5
2.9
2.2
3.1
4.1
3.4
3.8
4.0
4.6
6.0
4.4
3.8
med
ium
sa
nd
coar
se
sand
coar
se
sand
coar
se
sand
fine
sa
nd
med
ium
sa
nd
tine
sand
med
ium
sa
nd
fine
sa
nd
mud
dy
sand
silty
sa
nd
fine
sa
nd
silty
sa
nd
mud
dy
sand
silty
sa
nd
mud
dy
sand
silty
sa
nd
sand
y m
ud
sand
y m
ud
mud
dy
sand
mud
dy
sand
0.09
0.03
0.03
0.06
0.08
0.25
0.16
0.40
0.36
0.22
0.32
0.49
0.74
0.52
0.40
rare
(0
.07)
abse
nt
abse
nt
abse
nt
abse
nt
abse
nt
rare
(0
.18)
abse
nt
few
(0
.5)
abun
dant
(6
8)
freq
uent
(6
.3)
rare
(0
.05)
freq
uent
(5
.3)
freq
uent
(2
.9)
few
(0
.8)
freq
uent
(6
.7)
freq
uent
(3
.4)
abun
dant
(6
9)
abun
dant
(
113)
freq
uent
(
11.3
)
freq
uent
(2
.6)
83*
33
40
20
13 I 11
59
22
38
59
88
56
54
14
41
71
0 0
6 I
6
95
89
95
L,D
92
96
91
12
20
0
84
67
89
L,D
90
80
96
L
,D
71
71
78
L,D
96
99
13
92
93
91
L,D
94
92
98
D
85
39
93
L,D
95
98
92
87
87
86
D
0 0
* no
t st
atis
tical
ly
sign
ific
ant;
only
6
spec
imen
s co
unte
d.

N
G.D. Corner et aLlMarine Geology 134 (1996) 113-125

G.D. Corner et al/Marine Geology 134 (1996) 113-125 119
&LIVING/DEAC )
0 _- Living _- Dead
ABUNDANCE Tests per gram
sediment
@
loo
0 IO
91 0 0.1 +o
\
Fig. 3. Abundance (indicated by circle size) and percentage living and dead foraminifera (light and dark sectors) in surface sediments
at sampled localities (numbered) at Tana. Full sample numbers are given in Table 1.
most Leirpollen samples, and by a few relatively shallow water samples (T93-7, T92-30, T92-32), (Fig. 3). A particularly high value (88%) occurs in delta slope sample T92-35, situated directly beyond the river outlet. Percentage living foramini- fera values show a weak positive correlation with water depth (Fig. 4B).
4.3. Totalforaminiferal assemblages
Amongst a total of 42 benthic foraminiferal taxa identified to species or genus level, 34 comprise > 1% in any one sample (combined living and dead fauna) and 4 species comprise >25% in any one sample (Table 2). No planktic foraminifera were found.
Three different fauna1 assemblages are distinguished on the basis of the dominant
species: ( 1) Elphidium albiumbilicatum assemblage, (2) Cibicides lobatulus assemblage, ,and (3) Adercotryma glomerata-Spiroplectammina biformis assemblage. With a few exceptions, living and dead assemblages in each sample are similar. Fig. 5 illustrates the fauna1 affinity of samples from the different environments.
4.3.1. Elphidium albiumbilicatum assembluge Elphidium albiumbilicatum dominates (33-88%)
in foraminifera-bearing samples from the delta plain (T93-1, T93-7) and in a single sample from the delta slope in Leirpollen (T92-23). All three samples derive from relatively shallow water (< 27 m), consist of sand, and show low abundance. Agglutinated species are subordinate (O-33% of total specimens). Apart from E. albiumbilicatum, the fauna1 content of each sample differs. although

120 G. D. Corner et al./Marine Geology 134 (1996) X13-125
Leirpollen Tanafjord
Water depth (m): 0 0
Mean grain size Mz (0): n 0
TOC (%): + +
Delta plain Correlation
n - _ _ _ _ _
- - - _ _ _ _
0 1 2 3 4 5 60 0 1 2 3 4 5 60 I I, I I I. I. I. I I I * I I I. I I, 1
0 20 40 60 80 100 120 130m 0 20 40 60 60 100 120 130m I. I., . I- I, I a,, t I I . , . t. I I- I - I I,
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 % 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 %
Plot of foraminiferal abundance (A) and percentage living of total foraminifera (B) versus mean grain-size, water depth and total organic carbon (TOC), for samples from the delta plain and subaqueous environments (tidal strait, delta slope, fjord basin) in Tanafjord and Leirpollen. (A) foraminiferal abundance correlates with grain size, water depth and TOC at 99.956, 92.5% and 99.7% confidence levels, respectively. (B) percentage living fauna correlates with water depth at an 82.4% confidence level; no correlation is found with grain size or TOC.
the low number of tests in the samples reduces the statistical significance of this fact. E. albiumbilica- turn occurs, otherwise, only in small numbers in tidal strait sample T92-32. Several specimens of E. albiumbilicatum were enclosed in agglutinated cysts (Plate I, specimen 12).
4.3.2. Cibicides lobatulus assemblage Cibicides lobatulus dominates (71%) in a single,
low abundance (0.47 tests per gram), fine-sand sample (T92-32) from the mouth of the tidal strait at a depth of 23 m. Agglutinated species comprise 6% of the specimens in this sample. Tests of C. lobatulus were abraded, in contrast to unabraded tests of other species. C. lobatulus also occurs in very small numbers at some other sites, notably at two proximal sites on the delta slope in Tanafjord near the outlet of the tidal strait (samples T92-30, T92-33) and on the delta slope in Leirpollen (samples T92-20, T92-22).
4.3.3. Adercotryma glomerata-spiroplectammina biformis assemblage
The Adercotryma glomerata-Spiroplectammina biformis assemblage is characteristic of the delta slope and fjord basin, being found in all but one of the 12 samples from this province, between depths of 26 and 124 m. Agglutinated species dominate in this assemblage (77-96% of the speci- mens in each sample; Table 1). Both A. glomeruta and S. biformis form a substantial component (15-47% in each sample), except in sample T92-35 where S. biformis dominates strongly (84%). S. biformis tends to be more common than A. glo- meruta, particularly in shallower (< 75 m) and proximal samples, and lowest abundance of A. glomeruta is found in samples from proximal sites (T92-30, T92-34 and T92-35; Fig. 5). Cribrostomoides crassimargo, C. kosterensis, Bulimina elongata, Reophax jiisiformis and Recurvoides turbinatus occur as accessary species

G. D. Corner et al./Marine Geology 134 (1996) 113-125 121
A.glomerata - Sbiformis (Delta slope - fjord basin)
T92-40
T92-38
T92-38
T92-20
T92-33
T92-42
T92-22
T92-21
T92-34
T92-30
T92-35= I
T92-32.............................mw-et-w . . . . . . . . . . . . . . . ..*..*................ . . ..*................................. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ..**..*....*..*..... .*.........*.....*................... :::::::::::::::::::::::::::::::::::::::- . . . . . . . ..a.......... .~.~.~.~.~.~.'.~.~.'.~.~.'.~.~.'.~.~.'. .~.~.~.~.~.'.~.~.~.'.~.~.~.~.~.'.~.~.~. .'.~.~.~.~.'.'.~.~.'.~.~.~.~.~.'.~.~.~.
ok0 0125 ok0 0.>5 ii0
Pearson correlation coefficient I.&
Fig. 5. Cluster analysis dendrogram showing sample al3inity according to species composition. Oblique shading denotes an Adercotryma glomerata-Spiroplectammina biformis assemblage from the delta slope and fjord basin province; proximal samples, which contain a lower frequency of A. glomerata, are indicated by light shading. Dot-shading denotes assemblages predominantly from the delta platform province-a Cibicides lobatulus assemblage from the tidal strait (sample T92-32) and an Elphidium albiumbilicatum assemblage from the delta plain (samples T93-1 and T93-7) and delta slope (sample T92-23).
in most samples. Living and dead assemblages are similar, except in the case of Leirpollen samples, where living S. biformis is absent.
5. Discussion
5. I. Fauna1 assemblage and depositional province
A major distinction can be made, on the basis ing a deeper (25-125 m), low-energy, muddy- of foraminiferal assemblages, test abundance, and sandy to sandy-muddy, fully marine environment, physical criteria, between delta platform and is characterized by relatively high, though variable, delta slope-fjord basin fauna1 and depositional foraminiferal abundance, a dominance of provinces. agglutinated forms, and an A. glomerata-S.
5.1.1. Delta platform province The delta platform province, comprising rela-
tively shallow (~25 m), high-energy, sandy, partly brackish areas of the delta plain and tidal strait, is characterized by an absence or low abundance of foraminifera, a dominance of calcareous forms and, amongst the few samples from this province containing foraminifera, an E. albiumbilicatum assemblage on the delta plain and a C. lobatulus assemblage in a single tidal strait mouth sample (T92-32).
The low foraminiferal abundance in this prov- ince is most likely related to extremely harsh conditions for foraminifera in an environment which experiences rapid salinity changes and sedi- ment reworking by waves and tidal currents (cf. Williams, 1989). Sample T93-1, from the most dynamic (fluviotidal channel) of the two sites on the delta plain which yielded foraminifera, con- tained mostly living specimens (Table 1)) suggest- ing that dead specimens are quickly removed by winnowing and attrition. Encystment, seen in sev- eral specimens of E. albiumbilicatum, may be a protective response to mechanical and chemical disturbances in the environment (Linke and Lutze, 1993).
E. albiumbilicatum in other regions typically occurs in shallow and brackish water (Austin and Sejrup, 1994; Alve, 1995). C. lobatulus lives attached to clasts or algae in high energy environ- ments (Nyholm, 1961; Williamson, 1985). Abraded specimens of C. lobatulus, which domi- nate in sandy tidal strait sample T92-32, were probably transported to the site from the adjacent sublittoral zone to the east, indicating a mixed assemblage. C. lobatulus specimens in other delta- proximal samples were probably also transported from nearby sublittoral areas, this being facilitated by their low weight relative to size (Kontrovitz et al., 1978).
5.1.2. Delta slope-fjord basin province The delta slope-fjord basin province, compris-

122 G.D. Corner et aLlMarine Geology 134 (1996) 113-125
PLATE I

G.D. Corner et al./Marine Geology 134 (1996) 113-125 123
bgormis assemblage. A relatively high frequency of S. biformis compared with A. glomerata is found in shallower (~75 m), more proximal samples, in accordance with findings from other fjord and shelf areas (Scott et al., 1980; Vilks, 1989; Alve, 1990,1995; Alve and Nagy, 1990). Apart from this slight fauna1 depth zonation, and an absence of living specimens of S. bzjiormis in Len-pollen (related perhaps to ecological conditions at the time of sampling), the total assemblage within the delta slope-fjord basin province is fairly uniform, and no appreciable difference is apparent between the constituent environments of delta slope and adjacent fjord basin, nor between the open Tanafjord and the semi-enclosed Leirpollen, sug- gesting broadly similar ecological conditions in these areas.
The dominant species of the A. glomerata- S. biformis assemblage are common components in many estuarine, shelf and fjord associations of northern regions (see references in Murray, 1991; Williamson, 1985; Schafer and Cole, 1988; Vilks, 1989; Schroder-Adams et al., 1990; Jennings and Helgadottir, 1994). These species show a wide environmental tolerance range, particularly S. biformis which tolerates low salinity (20-31.2%0) and low dissolved oxygen levels (~2 ml/l), (Williamson et al., 1984; Alve, 1990,1991; Alve and Nagy, 1990). Spiroplectammina sp. and A. glomerata form characteristic species of a “fjord biofacies” at Yakutat, Alaska (Lagoe et al., 1994) and are relatively common in the fjords of Svalbard where a number of assemblages, includ- ing an A. glomerata assemblage, occur (M. Hald and S. Korsun, pers. comm., 1995). In Altafjord, northern Norway, agglutinated species, particu- larly A. gfomerata and, to a lesser extent, S. biformis are relatively common at sites close to the delta, shallower than 200 m, but not at deeper, more distal localities (Strand, 1979). Farther north, on the southern Barents Sea shelf, S. biformis and A. glomerata are relatively uncommon
(Hald and Steinsund, 1992). Thus, in the context of the northern Norway-Barents Sea area, the A. glomerata-S. biformis assemblage at Tana appears to represent an inner fjord biofacies.
5.2. Population variability within the delta slope-fjord basin province
Foraminiferal abundance within the delta slo- pe-fjord basin province is variable but tends to increase with increasing water depth and decreas- ing grain size (Fig. 4A). This trend corresponds to increasing distal position and an inferred decrease in sedimentation rate away from the river mouth. For example, highest abundance is found at distal sites T92-20, T92-21 and T92-42 (Fig. 3). High abundance samples also contain a higher content of total organic carbon (Fig. 4A) implying improved nutrient conditions at distal sites and perhaps enhancement, by biological factors, of the effect of differences in sedimentation rate.
High percentages of living foraminifera in most samples probably reflect generally high sedimenta- tion rates in the deltaic environment, since rapid burial of dead foraminifera will cause a higher frequency of living specimens. The weak positive correlation of percentage living of total foramini- fera values with water depth (Fig. 4B) is thought not to be significant. Relatively low values (< 15%) of living foraminifera combined with high test abundance in most samples from Leirpollen sug- gest that sedimentation rates there are generally lower than beyond the active delta front in Tanafjord, in accordance with Leirpollen’s distal position.
The variability in foraminiferal abundance and percentage living of total fauna in relation to depth and proximal-distal position, amongst delta slope-fjord basin samples, may be caused by a number of factors:
( 1) Postmortem disaggregation of fragile agglu- tinated tests and dissolution of calcareous tests
Plate I
1, 2. Reophax jiisiformis (Williamson). 3-5. Spiroplectammina biformis (Parker and Jones). 6. Adercotryma glomerata (Brady).
7. Cribrostomoides kosterensis (Hbglund). 8. Cribrostomoides crassimargo (Norman). 9, 10. Bulimina elongata (d’orbigny).
11, 12. Elphidium albiumbilicatum (Weiss). 13. Elphidium excavatum (Terquem). 14. Cibicides lobatulus (Walker and Jacob). Note
traces of dissolution in specimen 9, abrasion in specimens 11 and 13, and partial encystment of specimen 12.

124 G.D. Corner et aL/Marine Geology 134 (1996) 113-125
may reduce populations at rates which vary spa- tially. Stratigraphic studies from Altafjord indicate that disaggregation occurs slowly (Strand, 1979). Its effect on recent populations at Tana is therefore thought to be small. Calcium carbonate dissolution was observed (cf. Plate I) on both stained and unstained tests in several samples and appears to have reduced the number of calcareous tests to some extent, since the proportion of calcareous tests relative to agglutinated tests is generally lower among the dead than the living fauna (Table 1). However, the effect of dissolution on total abun- dance patterns has probably been minor since calcareous species are subordinate (< 33%) to agglutinated species among the living population (Table 1 ), and dissolution occurs among both high and low abundance samples, at various depths, and in all subenviroments except on the delta platform.
(2) Local variations in sedimentation rates can be expected in the delta slope-fjord basin province, particularly at more proximal sites, and this factor may have influenced abundance patterns. One sample in particular (T92-23), from a proximal delta slope site in Leirpollen, contains an unusually low abundance of foraminifera for a sample from this depth (27 m), as well as an anomalous E. albiumbilicatum fauna more characteristic of the delta plain. Resedimentation by downslope mass transport or possibly by tidal currents seems likely in this case.
(3) The effect of foraminiferal reproduction rates and the influence of scavengers on abundance patterns is difficult to assess. Sample T92-35, from the delta slope at a depth of 65 m directly beyond the river mouth, contains an unusually high abun- dance of foraminifera for a sample so close to the sediment source. Its fauna is also dominated by living specimens of a single species (94% S. biformis), reflecting perhaps population patchiness or a bloom at the time of sampling (cf. Murray, 1991).
6. Conclusions
Foraminiferal assemblages at Tanafjord- Leirpollen show a marked contrast between the
stressed environment of the delta platform prov- ince and the deeper environment of the delta slope-fjord basin province. Within the latter prov- ince, no contrast in foraminiferal assemblage is seen between the morphologically defined delta slope and adjacent fjord basin environments, nor between the open Tanafjord and the semi-enclosed Leirpollen basin. Fauna1 composition changes slightly with depth. A relatively high abundance of foraminifera at distal sites and a low proportion of living fauna in Leirpollen probably reflect lower sedimentation rates in these areas. Local variations in sedimentation rates and processes, postmortem dissolution and dissagregation of tests, and biologi- cal factors have probably contributed to the vari- ability in abundance and proportion of living of total fauna observed in the samples.
Acknowledgment
The work was carried out as part of a University of Tromso study of the Tana delta, sponsored by Norsk Hydro. The marine sediment samples were collected during a cruise with UiTo’s R/V Johan Ruud. Marit Berntsen and Mary Raste carried out the laboratory analyses, Froydis Strand and Audun Igesund made the drawings, Reidar Myklebust assisted with the SEM pictures, and Gunvor Granaas made the photographic repro- ductions. Radiocarbon dating was carried out by the Radiological Dating Laboratory, Trondheim. Critical comments on earlier drafts of the manu- script by M. Hald, M.B. Lagoe, D.B. Scott, C.J. Schriider-Adams and S. Snyder are much appreciated.
References
Alve, E., 1990. Variations in estuarine foraminiferal biofacies with diminishing oxygen conditions in Drammensfjord, southeast Norway. In: C. Hemleben et al. (Editors), Palaeo- ecology, Biostratigraphy, Palaeoceanography, and Taxon- omy of Agglutinated Foraminifera. Kluwer, Dortrecht, pp. 661-694.
Alve, E., 1991. Foraminifera, climate change, and pollution: a study of late Holocene sediments in Drammensfjord, southeast Norway. Holocene, 1: 243-261.

G.D. Corner et al/Marine Geology 134 (1996) 113-125 125
Alve, E., 1995. Benthic foraminiferal distribution and recolonization of formerly anoxic environments in Dram- mensfjord, southern Norway. Mar. Micropaleontol., 25: 1699186.
Alve, E. and Nagy, J., 1986. Estuarine foraminiferal distribution in Sandebukta, a branch of the Oslo Fjord. J. Foraminif. Res., 16: 261-284.
Alve, E. and Nagy, J., 1990. Main features of foraminiferal distribution reflecting estuarine hydrography in Oslo Fjord. Mar. Micropaleontol., 16: 181-206.
Austin, W.E.N. and Sejrup, H.P., 1994. Recent shallow water benthic foraminifera from western Norway: ecology and palaeoecological significance. Cushman Found. Spec. Publ., 32: 103-125.
Bernhard, J.M., 1988. Postmortem vital staining in benthic fora- minifera: duration and importance in population and distri- butional studies. J. Foraminif. Res., 18: 143-146.
Corner, G.D. and Fjalstad, A., 1993. Spreite trace fossils (Teichichnus) in a raised Holocene fjord-delta, Breidvikeidet, Norway. Ichnos, 2: 1555164.
Corner, G.D., Nordahl, E., Munch-Ellingsen, K. and Robert- sen, K.R., 1990. Morphology and sedimentology of an emer- gent fjord-head Gilbert-type delta: Alta delta, Norway. In: A. Colella and D.B. Prior (Editors), Coarse-Grained Deltas. Spec. Publ. Int. Assoc. Sedimentol., 10: 155-168.
Corner, G.D., Andreassen, K., Running, J.S. and Mauring, E., 1994. Morphology, sedimentology and structure of the Tana delta, northern Norway. In: 15th Reg. Int. Assoc. Sedi- mental. Meet., Ischia, 13-15 April 1994 (Abstr.).
Feyling-Hanssen, R.W., 1964. Foraminifera in Late Quaternary deposits in the Oslofjord area. Nor. Geol. Unders., 225, 383 pp.
Hald, M., 1986. Late Quaternary stratigraphy and paleo- environments on the continental shelf off Troms, Northern Norway, with special emphasis on the use of foraminifera. Thesis. Univ. Tromss.
Hald, M. and Steinsund, P.I., 1992. Distribution of surface sediment benthic foraminifera in the southwestern Barents Sea. J. Foraminif. Res., 22: 347-362.
Jennings, A.E. and Helgadottir, G., 1994. Foraminiferal assem- blages from the fjords and shelf of eastern Greenland. J. Foraminif. Res., 24: 123-144.
Kontrovitz, M., Snyder, S.W. and Brown, R.J., 1978. A flume study of the movement of foraminifera tests. Palaeogeogr. Palaeoclimatol. Palaeoecol. 23: 141-150.
Lagoe, M.B., Eyles, C.H. and Eyles, N., 1994. Foraminiferal biofacies and paleoenvironments in a Pliocene megachannel of the glaciomarine Yakataga Formation, Gulf of Alaska. Cushman Found. Spec. Publ., 32: 127-139.
Linke, P. and Lutze, G.F., 1993. Microhabitat preferences of benthic foraminifera-a static concept or a dynamic adapta- tion to optimize food aquisition? Mar. Micropaleontol., 20: 2155234.
Lutenauer, J.L., Barrie, J.V., Christian, H.A., Clague, J.J., Evoy, R. W., Hart, B.S., Hunter, J.A., Killeen, P.G., Kos-
taschuk, R.A., Mathewes, R.W., Monahan, P.A., Moslow, T.F., Mwenifumbo, C.J., Olynyk, H.W., Patterson, R.T., Pullan, S.E., Roberts, M.C., Robertson, P.K., Tarbotton, M.R., and Woeller, D.J., 1994. Fraser River delta: geology, geohazards and human impact. In: J.W.H. Monger (Editor), Geology and Geological Hazards of the Vancouver Region, Southwestern British Columbia. Geol. Surv. Can. Bull., 481: 197-220.
Mauring, E., Running, J.S. and Danielsen, E., 1995 Georadar og refleksjonsseismiske malinger i Tanadalen, Tana kom- mune, Finnmark. NGU Rapp., 95.007, 11 pp.
Meldgaard, S. and Knudsen, K.L., 1979. Metoder til indsamling og opparbejding af prover til foraminifer-analyser. Dan. Natur, Dan. Skole Arskr., 1979: 48857.
Murray, J.W., 1973. Distribution and Ecology of Living Benthic Foraminiferids. Heinemann, London, 274 pp.
Murray, J.W., 1991. Ecology and Palaeoecology of Benthic Foraminifera. Longman, 397 pp.
Nordgard, O., Normann, U., Pettersen, F. and Skarberg, N.E., 1982. Havmiljodata fra nordnorske fjorder 1981. Tromura, Naturvitensk., 38, Tromss Mus., 168 pp.
Nyholm, K.G., 1961. Morphogenesis and biology of the fora- minifer Cibicides lobatulus. Zool. Bidr. Upps., 33: 157-197.
Schafer, CT. and Cole, F.E., 1988. Environmental associations of Baffin Island fjord agglutinated foraminifera. Abh. Geol. Bundesanst., 41: 3077323.
Schroder-Adams, C.J., Cole, F.E., Medioli, F.S., Mudie, P.J., Scott, D.B. and Dobbin, L., 1990. Recent arctic shelf fora- minifera: seasonally ice covered vs. perenially ice covered areas. J. Foraminif. Res., 20: 8836.
Scott, D.B., Schafer, CT. and Medioli, F.S., 1980. Eastern Canadian estuarine foraminifera: a framework for compari- son. J. Foraminif. Res., 10: 2055234.
Strand, J.E., 1979. Paleoklimatisk og stratigrafisk undersekelse av senkvartrere marine sedimenter fra Altafjorden og Troms- oflaket, Nord-Norge ved hjelp av bentoniske foraminiferer. Thesis. Univ. Oslo, 107 pp.
Vilks, G., 1989. Ecology of recent foraminifera on the Canadian continental shelf of the Arctic Ocean. In: Y. Herman (Editor), The Arctic Seas. Climatology, Oceanography, Geology, and Biology. Van Nostrand Reinhold, New York, pp. 497-569.
Walton, W.R., 1952. Techniques for recognition of living fora- minifera. Contrib. Cushman Found. Foraminif. Res., 3: 56660.
Williams, H.F.L., 1989. Foraminiferal zonations on the Frazer River delta and their application to paleoenvironmental interpretations. Palaeogeogr. Palaeoclimatol. Palaeoecol.. 73: 39950.
Williamson, M.A., 1985. Recent foraminiferal diversity on the continental margin off Nova Scotia, Canada. J. Foraminif. Res., 15: 43-51.
Williamson, M.A., Keen, C.E. and Mudie, P.J., 1984. Foramini- feral distribution on the continental margin off Nova Scotia. Mar. Micropaleontol., 9: 219-239.




















