
PAH-Domain-Specific Interactions of the Arabidopsis Transcription Coregulator SIN3LIKE1 (SNL1) with...
13
PAH-Domain-Specific Interactions of the Arabidopsis Transcription Coregulator SIN3-LIKE1 (SNL1) with Telomere-Binding Protein 1 and ALWAYS EARLY2 Myb-DNA Binding Factors Adam J. Bowen 1 , Deyarina Gonzalez 1 , Jonathan G. L. Mullins 1 , Anuj M. Bhatt 2 , Alberto Martinez 3 and R. Steven Conlan 1 ⁎ 1 Institute of Life Science, School of Medicine, Swansea University, Singleton Park, Swansea SA2 8PP, UK 2 Department of Plant Sciences, University of Oxford, South Parks Road, Oxford OX1 3RB, UK 3 Syngenta Ltd, Jealott's Hill International Research Centre, Bracknell, Berkshire RG42 6EY, UK Received 21 May 2009; received in revised form 25 September 2009; accepted 29 November 2009 Available online 4 December 2009 The eukaryotic SIN3 protein is the central component of the evolutionarily conserved multisubunit SIN3 complex that has roles in regulating gene expression and genome stability. Here we characterise the structure of the SIN3 protein in higher plants through the analysis of SNL1 (SIN3-LIKE1), SNL2, SNL3, SNL4, SNL5 and SNL6, a family of six SIN3 homologues in Arabidopsis thaliana. In an Arabidopsis-protoplast β-glucuronidase reporter gene assay, as well as in a heterologous yeast repression assay, full-length SNL1 was shown to repress transcription in a histone-deacetylase-dependent manner, demonstrating the conserved nature of SIN3 function. Yeast two-hybrid screening identified a number of DNA binding proteins each containing a single Myb domain that included the Arabidopsis ALWAYS EARLY proteins AtALY2 and AtALY3, and two telomere binding proteins AtTBP1 and AtTRP2/TRFL1 as SNL1 partners, suggesting potential functions for SNL1 in development and telomere maintenance. The interaction with telomere-binding protein 1 was found to be mediated through the well- defined paired amphipathic helix domain PAH2. In contrast, the AtALY2 interaction was mediated through the PAH3 domain of SNL1, which is structurally distinct from PAH1 and PAH2, suggesting that evolution of this domain to a more novel structural motif has occurred. These findings support a diverse role of SNL1 in the regulation of transcription and genome stability. © 2009 Elsevier Ltd. All rights reserved. Edited by J. O. Thomas Keywords: SIN3; SNL; PAH; Arabidopsis; coregulator Introduction The packaging of DNA into chromatin within the nucleus prevents access of the transcriptional machinery to the regulatory regions of genes, thereby providing an important mechanism for the regulation of gene expression. Chromatin is a dynamic structure and is constantly being ‘opened’ and ‘closed’ by transcription factors to regulate the accessibility of genes. One mechanism by which a cell is able to regulate the dynamics of chromatin is by altering the acetylation state of histones, the core protein components of nuclesomes. It is widely accepted that the acetylation of histone proteins correlates with gene expression, while deacetylation is associated with gene repression. The general transcription coregulator SIN3 is a component of a multisubunit protein complex typi- *Corresponding author. E-mail address: [email protected]. Present addresses: A. J. Bowen, Laboratory of Chromatin & Gene Expression, The Babraham Institute, Babraham Research Campus, Cambridge CB22 3AT, UK; A. Martinez, Syntaxin, Barton Lane, The Quadrant, Abingdon OX14 3YS, UK. Abbreviations used: HDAC, histone deacetylase; Rb, retinoblastoma; PAH, paired amphipathic helix; HID, HDAC interaction domain; TBP1, telomere-binding protein 1; qRT-PCR, quantitative real-time PCR; TSA, trichostatin A; SID, selected interacting domain; SNL, SIN3-LIKE; ONPG, o-nitrophenol-β-d-galactosidase-D- galactosidase; SDM, site-directed mutagenesis. doi:10.1016/j.jmb.2009.11.065 J. Mol. Biol. (2010) 395, 937–949 Available online at www.sciencedirect.com 0022-2836/$ - see front matter © 2009 Elsevier Ltd. All rights reserved.
Transcript of PAH-Domain-Specific Interactions of the Arabidopsis Transcription Coregulator SIN3LIKE1 (SNL1) with...

doi:10.1016/j.jmb.2009.11.065 J. Mol. Biol. (2010) 395, 937–949
Available online at www.sciencedirect.com
PAH-Domain-Specific Interactions of the ArabidopsisTranscription Coregulator SIN3-LIKE1 (SNL1) withTelomere-Binding Protein 1 and ALWAYS EARLY2Myb-DNA Binding Factors
Adam J. Bowen1, Deyarina Gonzalez1, Jonathan G. L. Mullins1,Anuj M. Bhatt2, Alberto Martinez3 and R. Steven Conlan1⁎
1Institute of Life Science, School ofMedicine, Swansea University,Singleton Park, Swansea SA28PP, UK2Department of Plant Sciences,University of Oxford, South ParksRoad, Oxford OX1 3RB, UK3Syngenta Ltd, Jealott's HillInternational Research Centre,Bracknell, Berkshire RG42 6EY,UKReceived 21 May 2009;received in revised form25 September 2009;accepted 29 November 2009Available online4 December 2009
*Corresponding author. E-mail [email protected] addresses: A. J. Bowen, La
Chromatin & Gene Expression, TheBabraham Research Campus, CambA. Martinez, Syntaxin, Barton Lane,Abingdon OX14 3YS, UK.Abbreviations used: HDAC, histo
retinoblastoma; PAH, paired amphiHDAC interaction domain; TBP1, teprotein 1; qRT-PCR, quantitative reatrichostatin A; SID, selected interactSIN3-LIKE; ONPG, o-nitrophenol-β-galactosidase; SDM, site-directed m
0022-2836/$ - see front matter © 2009 E
The eukaryotic SIN3 protein is the central component of the evolutionarilyconserved multisubunit SIN3 complex that has roles in regulating geneexpression and genome stability. Here we characterise the structure of theSIN3 protein in higher plants through the analysis of SNL1 (SIN3-LIKE1),SNL2, SNL3, SNL4, SNL5 and SNL6, a family of six SIN3 homologues inArabidopsis thaliana. In an Arabidopsis-protoplast β-glucuronidase reportergene assay, as well as in a heterologous yeast repression assay, full-lengthSNL1was shown to repress transcription in a histone-deacetylase-dependentmanner, demonstrating the conserved nature of SIN3 function. Yeasttwo-hybrid screening identified a number of DNA binding proteins eachcontaining a single Myb domain that included the Arabidopsis ALWAYSEARLY proteins AtALY2 and AtALY3, and two telomere binding proteinsAtTBP1 andAtTRP2/TRFL1 as SNL1partners, suggesting potential functionsfor SNL1 in development and telomere maintenance. The interaction withtelomere-binding protein 1 was found to be mediated through the well-defined paired amphipathic helix domain PAH2. In contrast, the AtALY2interaction was mediated through the PAH3 domain of SNL1, which isstructurally distinct from PAH1 and PAH2, suggesting that evolution of thisdomain to amore novel structuralmotif has occurred. These findings supporta diverse role of SNL1 in the regulation of transcription and genome stability.
© 2009 Elsevier Ltd. All rights reserved.
Edited by J. O. Thomas
Keywords: SIN3; SNL; PAH; Arabidopsis; coregulatoress:
boratory ofBabraham Institute,ridge CB22 3AT, UK;The Quadrant,
ne deacetylase; Rb,pathic helix; HID,lomere-bindingl-time PCR; TSA,ing domain; SNL,d-galactosidase-D-utagenesis.
lsevier Ltd. All rights reserve
Introduction
The packaging of DNA into chromatin within thenucleus prevents access of the transcriptionalmachinery to the regulatory regions of genes,thereby providing an important mechanism for theregulation of gene expression. Chromatin is adynamic structure and is constantly being ‘opened’and ‘closed’ by transcription factors to regulate theaccessibility of genes. One mechanism by which acell is able to regulate the dynamics of chromatin isby altering the acetylation state of histones, the coreprotein components of nuclesomes. It is widelyaccepted that the acetylation of histone proteinscorrelates with gene expression, while deacetylationis associated with gene repression.The general transcription coregulator SIN3 is a
component of a multisubunit protein complex typi-
d.

938 Arabidopsis SIN3-LIKE (SNL) Coregulators
cally associated with the histone deacetylase(HDAC) RPD3 and has been shown to be importantin the modulation of gene expression in a number ofeukaryotes including Saccharomyces cerevisiae (bud-ding yeast), Drosophila melanogaster and mammals.1
SIN3 functions as a scaffold protein in the core SIN3complex, which also includes the SIN3-associatedproteins SAP-18, -25, -30, -45, -130 and -180, and the
Fig. 1. Conservation of SIN3 peptide sequences. Fifty-onenumber of genomic databases and aligned using ClustalX 2.0.1nodes are bootstrap values, which are based on 1000 replicates
retinoblastoma (Rb)-associated proteins RbAp46/48.1 SIN3 is thought to mediate multiple protein–protein interactions between core and other moreperipheral proteins via paired amphipathic helix(PAH) domains and an HDAC interaction domain(HID).1 SIN3 contains no demonstrated intrinsicDNA binding activity, but is recruited to target DNAthrough interactions with site-specific DNA binding
unique SIN3 peptide sequences were identified from a0. The tree was constructed using NJplot. Numbers at the. The scale bar represents 0.1 peptide substitutions per site.

Fig. 2. SNL-predicted structure and functional domains. Pfam analysis identified paired amphipathic helices (PAH,green), HDAC interaction domains (HID, red) and other nonannotated domains (shaded regions). Numbers indicatepredicted peptide amino acid length.
939Arabidopsis SIN3-LIKE (SNL) Coregulators
factors to achieve its gene-specific regulatory activ-ity. Through regulation of interactions with DNAbinding proteins, the activity of SIN3 can be tightlymodulated.In Schizosaccharomyces pombe (fission yeast), SIN3
(PST1 and PST2) has been shown to be important inthe regulation of centromeres, playing a critical rolein the equal distribution of chromosomes to daugh-ter cells during both mitosis and meiosis.2 SIN3 hasbeen shown to associate temporally with centro-meres, where it appears to be involved in the initialdeacetylation necessary for subsequent steps inheterochromatin formation, and therefore has arole in maintaining the genomic stability.2 Genomeintegrity is also maintained by telomere stabilitywhere they prevent degradation and fusion ofchromosome ends. Loss of SIN3 has been associatedwith shortened telomere length in genetic screens inS. cerevisiae.3
SIN3 is essential inDrosophila, as sin3 null mutantsdie during the larval stage of development due to adelay in the G2 phase of the cell cycle.
4 The deletionof mSIN3A in mouse results in lethality at thepostimplantation stage of development.5 Further-more, in mouse primary and established cell lines,the deletion of mSIN3A results in defects at allphases of the cell cycle, including the DNA synthesisphase, as well as enhanced apoptotic responses,altered expression of many genes linked to cellregulation, DNA replication/repair, mitochondrialmetabolism and histone modification.5,6
In plants, homologous components of the coreSIN3 complex, including SNL3 (see Results forArabidopsis SIN3 nomenclature),7 AtSAP18,8
AtRPD39 and the Rb-associated protein RbAp46/48 homologues including AtMSI1/2,10 have beenidentified. These components are involved in severalbiological pathways, including stress response7 andseed development.10
In this study, we have identified six SIN3orthologues that are expressed in Arabidopsis. Weshow that SNL1 is able to repress transcriptionthrough a highly conserved mechanism. A yeasttwo hybrid screen identified a number of Myb-domain-containing DNA binding proteins, includ-ing AtALY2 and telomere-binding protein 1 (TBP1)that interacted with SNL1 domains PAH2 and
PAH3, respectively, demonstrating how the SNLproteins can be differentially recruited to targetgenes. Our data show that specific PAH domainsare likely to be involved in the targeted recruitmentof the coregulator by DNA binding factors involvedin development and genome stability.
Results
Identification and characterisation of theArabidopsis SIN3-like gene family
Interrogation of the Arabidopsis genome databasereturned six predicted peptide sequences homolo-gous to the yeast SIN3 corepressor. These werenamed SIN3-LIKE (SNL) 1 to 6 in accordance withTAIR guidelines, which we propose as the nomen-clature for SIN3 inArabidopsis. These sequences werealigned using ClustalX (Fig. 1 and SupplementaryFig. 1). The resultant phylogenetic tree placed theSNL peptide sequences into three distinct groupscontaining SNL1 (AT3G01320) and SNL2(AT5G15020), SNL3 (AT1G24190) and SNL4(AT1G70060), SNL5 (AT1G59890) and SNL6(AT1G10450). To examine the relationship betweenSNLs and SIN3s encoded for by other eukaryoticgenomes, an exhaustive phylogenetic analysis ofpeptide sequences was performed. Using a numberof genomic databases, we identified and alignedapproximately 50 unique SIN3 peptide sequences(Fig. 1). In this extensive analysis, SIN3 homologuesfell into a number of expected clades. Yeasts andfungi fell largely into a distinct clade, suggestingconsiderable divergence from higher eukaryotes.Metazoa SIN3 sequences were placed in the sameclade,with all SIN3A and SIN3B sequences groupingseparately. Mammalian SIN3A sequences groupedtogether with Xenopus (frog) and Tetraodon nigrovir-idis (puffer fish) sequences, whereas on the adjacentbranch of this clade with mammalian SIN3B wereDanio rario (zebra fish) and Strongylocentrotus pur-puratus (sea urchin) sequences. Insect sequencesformed a separate group that diverged from theanimal clade at an early stage. Plant sequencesformed a distinct group with representative se-

Fig. 3. Structural modelling of the PAH2 domains of SNL1, SNL2 and SNL5. Homology modelling of the PAH2domains of SNL1 (a), SNL2 (b) and SNL5 (c) showing the locations of basic residues in blue and acidic residues in red. Thestructures of SNL1 and SNL2 are very similar in terms of distribution and identity of charged residues. R172 and R201(SNL1), R166 and R195 (SNL2) project into the interhelical binding groove. While the tertiary structure is generallysimilar, there is more variation in the location of charged residues in SNL5, where R159 adopts a different orientation andthe other projecting basic residue is a histidine (H188). In SNL5, there is an additional basic residue (R194) in the loopregion immediately following helix 4, whereas there is no basic residue in the equivalent regions of PAH2 in SNL1 andSNL2. All models were visualized using the molecular graphics program Chimera (http://www.cgl.ucsf.edu/chimera/).
940 Arabidopsis SIN3-LIKE (SNL) Coregulators
quences from Oryza sativa (rice), Zea mays (maize)andMarchantia polymorpha (a liverwort). Interesting-ly, only two copies of SIN3 were identified in maize,which corresponded to the SNL1/2 group and theSNL3/4 group in Arabidopsis.Analysis of the six SNL peptide sequences using
Clustal-X and the Pfam database† revealed that allcontain PAH domains and HIDs; however, SNL 5and 6 contained only PAH domains 1 and 2 (Fig. 2and Supplementary Fig. 1). Both PAH and HIDdomains have been shown to be critical for SIN3function in all other organisms in which SIN3 hasbeen functionally characterised. In a previous studyof yeast SIN3 and two mouse SIN3 homologues(mSIN3A/B), it was found that the regions ofhighest similarity were centred around the PAHdomains.11
Structural modelling of the PAH2 domains ofSNL1, SNL2 and SNL5 was carried out based onsequence identity of approximately 46% with theresolved crystal structures of PAH domains depos-ited in the Protein Data Bank (PDB) (2F05_A,1PD7_A, 1S5R_B, 1G1E_B, 1E91_A, 1S5Q_B). Theconservation of amino acid residues on the innersurface of PAH2 domains suggests that the functionof this domain is highly conserved and that thishydrophobic pocket created by the α-helix bundlestructure is likely to act as the primary interactionsurface for protein–protein interactions.12 In gener-al, there is a particularly high level of conservationon the inner surface of the longer α1 and α2 helices.These two helices form a deep hydrophobic pocketthat has been found to be the primary interactionsurface between the PAH2 domain and MAD1.12 Inthis work, discrete differences are seen betweenhelix 4 of the PAH2 domains of SNL1 and SNL2when compared with, for example, SNL5 (Fig. 3,
†http://www.sanger.ac.uk/Software/Pfam/
basic residues, blue; acidic residues, red). The PAH2structures of SNL1 and SNL2 are very similar interms of distribution and identity of chargedresidues. R201 (SNL1) and R195 (SNL2) projectinto the interhelical binding groove. While thetertiary structure is generally similar, there is morevariation in the location of charged residues inSNL5, where the projecting basic residue is ahistidine (H188). In SNL5, there is an additionalbasic residue (R194) in the loop region immediatelyfollowing helix 4, whereas there is no basic residuein the equivalent regions of PAH2 in SNL1 andSNL2. Such differences are likely to play animportant role in interactions between individualSNL family members and DNA binding proteins.
Expression analysis of SNL genes and SNL1protein
The expression of each SNL gene was determinedusing quantitative real-time PCR (qRT-PCR) ofmRNA extracted from rosette leaves and wassupported by EST and genome-wide microarrayexpression data‡ demonstrating that none of the sixSNL family members were pseudogenes (Fig. 4a).Comparative analysis of qRT-PCR data with ATH1microarray data revealed similar expression levelsfor each of the SNL genes and suggests that they areall likely to encode functional proteins; however,specific SNL protein antibodies are not yet availableto evaluate protein expression.A full-length cDNA clone was obtained for SNL1
and verified by DNA sequencing before being usedin subsequent experiments. The expression patternand subcellular localisation of full-length SNL1 wasdetermined using an SNL1-CFP translational fusionprotein. Tobacco leaf cells were infiltrated with
‡http://Arabidopsis.org/index.jsp
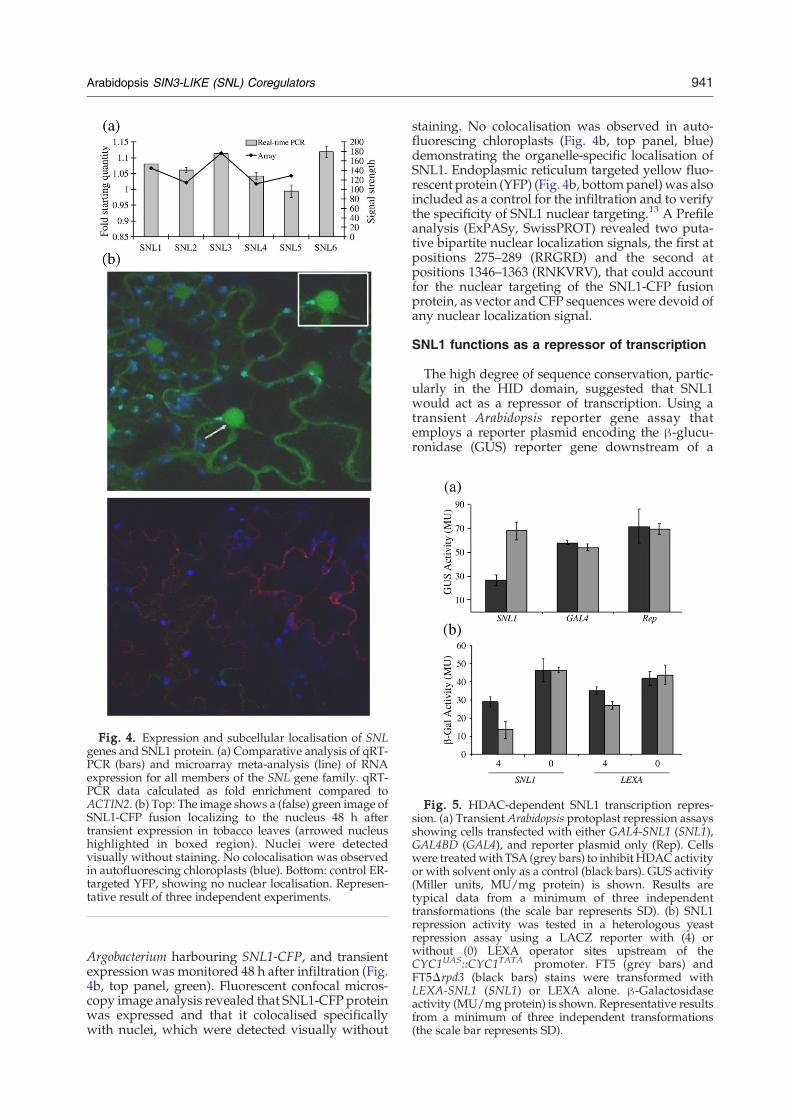
Fig. 4. Expression and subcellular localisation of SNLgenes and SNL1 protein. (a) Comparative analysis of qRT-PCR (bars) and microarray meta-analysis (line) of RNAexpression for all members of the SNL gene family. qRT-PCR data calculated as fold enrichment compared toACTIN2. (b) Top: The image shows a (false) green image ofSNL1-CFP fusion localizing to the nucleus 48 h aftertransient expression in tobacco leaves (arrowed nucleushighlighted in boxed region). Nuclei were detectedvisually without staining. No colocalisation was observedin autofluorescing chloroplasts (blue). Bottom: control ER-targeted YFP, showing no nuclear localisation. Represen-tative result of three independent experiments.
Fig. 5. HDAC-dependent SNL1 transcription repres-sion. (a) Transient Arabidopsis protoplast repression assaysshowing cells transfected with either GAL4-SNL1 (SNL1),GAL4BD (GAL4), and reporter plasmid only (Rep). Cellswere treatedwith TSA (grey bars) to inhibit HDACactivityor with solvent only as a control (black bars). GUS activity(Miller units, MU/mg protein) is shown. Results aretypical data from a minimum of three independenttransformations (the scale bar represents SD). (b) SNL1repression activity was tested in a heterologous yeastrepression assay using a LACZ reporter with (4) orwithout (0) LEXA operator sites upstream of theCYC1UAS::CYC1TATA promoter. FT5 (grey bars) andFT5Δrpd3 (black bars) stains were transformed withLEXA-SNL1 (SNL1) or LEXA alone. β-Galactosidaseactivity (MU/mg protein) is shown. Representative resultsfrom a minimum of three independent transformations(the scale bar represents SD).
941Arabidopsis SIN3-LIKE (SNL) Coregulators
Argobacterium harbouring SNL1-CFP, and transientexpression was monitored 48 h after infiltration (Fig.4b, top panel, green). Fluorescent confocal micros-copy image analysis revealed that SNL1-CFP proteinwas expressed and that it colocalised specificallywith nuclei, which were detected visually without
staining. No colocalisation was observed in auto-fluorescing chloroplasts (Fig. 4b, top panel, blue)demonstrating the organelle-specific localisation ofSNL1. Endoplasmic reticulum targeted yellow fluo-rescent protein (YFP) (Fig. 4b, bottompanel)was alsoincluded as a control for the infiltration and to verifythe specificity of SNL1 nuclear targeting.13 A Prefileanalysis (ExPASy, SwissPROT) revealed two puta-tive bipartite nuclear localization signals, the first atpositions 275–289 (RRGRD) and the second atpositions 1346–1363 (RNKVRV), that could accountfor the nuclear targeting of the SNL1-CFP fusionprotein, as vector and CFP sequences were devoid ofany nuclear localization signal.
SNL1 functions as a repressor of transcription
The high degree of sequence conservation, partic-ularly in the HID domain, suggested that SNL1would act as a repressor of transcription. Using atransient Arabidopsis reporter gene assay thatemploys a reporter plasmid encoding the β-glucu-ronidase (GUS) reporter gene downstream of a

942 Arabidopsis SIN3-LIKE (SNL) Coregulators
minimal tCUP promoter and 4 GAL4 bindingsites,14,15 we tested the repression function of aconstitutively expressed full-length SNL1 fused tothe DNA binding domain of the GAL4 protein(GAL4-SNL1). Reporter and effecter plasmids werecotransfected into Arabidopsis leaf protoplasts. Whenrecruited to the reporter plasmid, the GAL4-SNL1fusion reduced GUS reporter gene expression by63% (Fig. 5a), demonstrating the ability of SNL1 todirectly repress transcription when artificially teth-ered to a test promoter. The presence of theHID in allSNLpeptide sequences suggested that the repressionfunction of SNL1 was likely to occur through anHDAC-dependent mechanism. Therefore, repres-sion assays were repeated in the presence of theHDAC inhibitor trichostatin A (TSA). The effect ofTSA was to completely abolish the repressionactivity of SNL1 (Fig. 5a), demonstrating therequirement for HDAC activity in transcriptionrepression by SNL1. Similarly, in a heterologousyeast repression assay system, SNL1 demonstrated arequirement for the HDAC RPD3, the specificHDAC associated with SIN3 function in S. cerevisiae.Full-length SNL1 was fused to the LEXA DNAbinding domain to create a constitutively expressedLEXA-SNL1 fusion analogous to the plant GAL4-SNL1 fusion clone. LEXA-SNL1 was co-transformedinto a wild-type yeast strain (FT5) together with β-galactosidase (LacZ) reporter plasmids with(pJK1621) or without (pLG312ΔS) four LEXA ope-rator sites located upstream of the CYC1UAS::CYC1TATA promoter.16 When SNL1 was directlyrecruited to the test promoter via LEXA (LEXA-SNL1), LacZ reporter gene expression was reducedby 72% compared to the LEXA-only control (Fig. 5b),demonstrating that SNL1 has repressor activity inthis heterologous system and supporting the notionthat it functions through a highly conserved repres-sionmechanism. In the absence ofRPD3 (FT5Δrpd3),SNL1 was unable to fully repress transcription whenrecruited directly to a test promoter. LacZ reportergene expression was reduced by only 37% comparedto that of the control (Fig. 5b), indicating that SNL1was able to utilise a heterologous HDAC protein toeffect repression function. It is possible that otherHDACsmay have partially compensated for the lossof RPD3, or that SNL1 may be able to represstranscription through other mechanism(s).
Table 1. SNL1 interacting DNA binding proteins identified i
Locus tag Gene name Length (residues)
AT3G05380 AtALY2 975AT3G21430 AtALY3 1133AT5G56860 GNC 399AT5G13820 TBP1 640AT3G46590 AtTRP2/TRFL1 548AT2G36960 TKI1 723AT5G47660 Similar to GTL1 398AT5G04390 C2H2-like 363AT2G30470 HSI2 791AT4G32620 1514
SID indicates the consensus interaction region for each prey protein r
Identification of SNL1 interacting proteins
Sin3 is a known component of multiproteincomplexes that is recruited to the promoters oftarget genes by DNA binding proteins in order toexert function; however, no such protein partnershave yet been reported in plants. To identify SNL1partners, a yeast two-hybrid assay testing a total of88 million interactions was undertaken using full-length SNL1 as bait. A total of 262 interactingpartners encoding for 43 individual proteins (Sup-plementary Table 1) were identified and given aconfidence score,17 which has been shown topositively correlate with biologically significantinteractions.18 Overlapping prey fragments derivedfrom the same gene were clustered, and thisconserved sequence was defined as the selectedinteracting domain (SID)17 (Table 1 and Supple-mentary Table 1). Interacting partners with knownor predicted nonnuclear subcellular localisationswere considered as false interactions, as SNL1 wasshown to be localised to the nucleus in transienttransfection assays. Analysis of the remaininginteracting partners revealed 10 known or putativeDNA binding proteins, including AtALY2,AtALY3,19 HSI2,20 TKI1,21 and TBP1 and AtTRP2/TRFL122,23 that function in transcription repression/gene silencing processes, suggesting that like itshomologues in other eukaryotic species, SNL1 isrecruited to target genes through site-specifictranscription factors (Table 1). AtALY2 andAtALY319 are orthologues of the ALWAYS EARLYDrosophila (ALY) protein in Drosophila, LIN9 inCaenorhabditis elegans, hLIN9 in humans and theDrosophila Myb interacting protein 130 (MIP13)/Twilight (TWIT) family of proteins.19 Unlike otherALY proteins, which do not contain any knownDNA binding motif or reported DNA bindingactivity, AtALY2 and AtALY3 each contain a singleMyb domain that has been shown to bind DNA (Fig.6a), suggesting that the plant ALY orthologues haveevolved intrinsic DNA-binding function, and couldserve to recruit SNL1 directly to target genes.Further analysis of the aligned AtALY2 andAtALY3 SNL1 interacting regions revealed a regionof strong homology with a predicted helical struc-ture (AFDALQALAELS, AtALY2) as a potentialSNL1 interaction domain (Fig. 6b). AtTBP1 and
n a large-scale yeast two-hybrid screen
SID Interprot domains
244–427 Myb, homeodomain, DIRP, Tudor365–453 Myb, homeodomain, DIRP268–399 Myb18–296 Myb, homeodomain, ubiquitin10–193 Myb, homeodomain, SANT332–538 Myb, homeodomain, SANT47–221 Myb157–248 zinc finger (C2H2 type)188–393 B3 DNA binding domain, CW-type zinc finger211–406 Tudor
elative to the start codon.

Fig. 6. Structure and alignment of SNL1 interacting proteins. Schematic representation of (a) AtALY3 and AtALY2,and (c) TBP1 and AtTRP2/TRFL1 showing location ofMyb domains, other prominent structural domains (e.g., DRIP) andthe SNL1 interacting domain (SID). Secondary-structure domains were determined within the SID using Jpred3 (H, α-Helix; HeH, extended α-helix). (b) ClustalX alignments of the consensus SID between AtALY2 and AtALY3 with theconserved alpha-helix region underlined.
943Arabidopsis SIN3-LIKE (SNL) Coregulators
AtTRP2/TRFL1 encode for two of the telomericDNA binding proteins found in Arabidopsis, andboth peptide sequences for these proteins againcontain a single Myb DNA binding domain22,23
(Fig. 6c). Further analysis of the aligned AtTBP1 andAtTRP2/TRFL1 SNL1 interacting regions revealedregions of strong homology, although no conservedhelical domain was identified in the aligned SIDs(not shown). The identification of telomere-specificprotein partners of SNL1 is consistent with the roleplayed by SIN3 in maintaining genome stability inother organisms and, very excitingly, shows for thefirst time a direct interaction between a telomere-binding protein and a SIN3 protein.A second group of SNL1 interacting proteins
identified in the yeast two-hybrid screen haveknown or predicted functions in protein stability.ASK2 (Arabidopsis SKP1-LIKE 2), a ubiquitin-proteinligase expressed in metaphase, telophase and duringcytokinesis in male meiocytes24 that is required forearly development inArabidopsis,25 ARABIDILLO-1,which contains an F-Box and may therefore functionin protein degradation,26 and the E3 ubiquitin-protein ligase AtPUB14.27 Previously, the functionof the SIN3 complex in regulating silencing at HMR,telomeric and rDNA loci in yeast has been shown tobe dependent on the ubiquitin-conjugating enzyme
Rad6;28 therefore, the association of SNL1 with theseproteins is not unexpected.
Interaction of SNL1 with AtALY2 and TBP1involves distinct PAH domains
Having identified a group of single Myb-domain-containing DNA binding proteins in the yeast two-hybrid screen, we selected two members of thisgroup of proteins to determine whether they de-monstrated any interaction specificity with SNL1.Full-length LexA-tagged SNL1 and sequential C-terminal truncations of LexA-SNL1 terminating afterPAH1, PAH2, PAH3, or HIDwere synthesised usinga cell-free transcription-translation system (Fig. 7a).These proteins were then incubated with [35S]methionine-labelled AtALY2 or TBP1 producedusing the same cell-free transcription–translationsystem before samples were immunoprecipitatedusing an anti-LexA antibody, separated by PAGE,and analysed by phosphoimaging. This analysisrevealed that distinct PAH domains were requiredfor interactionwith each of theMyb-domain proteinstested. Deletion of PAH2 from the C-terminal-truncated SNL1 protein resulted in loss of interactionwith TBP1, indicating that, like many other reportedSIN3–DNA binding factor interactions,1 PAH2

Fig. 7. Specific PAH domains of the SNL1 protein are required for interaction with AtALY2 and TBP1. (a) LexA fusionproteins for SNL1 (1–1520 aa) and C-terminal truncated SNL1 derivatives terminating after PAH1 (1–137 aa), PAH2 (1–276 aa), PAH3 (1–455 aa) and HID (1–654 aa) were incubated with full-length [35S]methionine-labelled TBP1, AtALY2 orluciferase (LUC) control protein. (b, d, and f) Samples were then incubated with an anti-LexA antibody toimmunoprecipitate SNL1 (or SNL1-derivative) interacting proteins. Immunoprecipitated samples were separated bySDS-PAGE and any immunoprecipitated 35S-labelled proteins were visualised using a phosphorimager. (c, e, and g) Thepresence of unlabelled SNL1 and C-terminal truncated derivatives in immunoprecipitated samples was confirmed byprotein blotting using an anti-LexA antibody. Input= 10% volume of input protein used in interaction assays. (h)Structural model of SNL1 PAH3 domain showing the first two shortened helices and the different positions of equivalentbasic side chains of the key arginine residues (Arg222 and Arg249) relative to the binding pocket and other arginine(Arg198) and lysine (Lys194) residues surrounding the pocket.
944 Arabidopsis SIN3-LIKE (SNL) Coregulators
supports the interaction between SNL1 and TBP1(Fig. 7b and c). However, the region of TBP1 that wasidentified in our yeast two-hybrid screen (aminoacids 18–296) does not contain the proposed PAH2interaction consensus sequence identified in amurine system (A[A/V]X[V/A][L/V]),29 suggestingthat a plant-specific motif supports this interaction.
In contrast, deletion of PAH3 from the C-terminalof SNL1 resulted in loss of interaction with AtALY2(Fig. 7d and e), indicating that PAH3 is required forthe interaction between SNL1 and AtALY2. Only alimited number of other proteins have been shown tointeract with PAH3, and most of these experimentsalso included regions flanking the PAH3 domain.1

945Arabidopsis SIN3-LIKE (SNL) Coregulators
Structuralmodelling of the SNL1 PAH3domain (Fig.7h), as with PAH1 and PAH2 domains, revealed afour-helix structure, although the first two helices aremarkedly shorter and the positions of the equivalentbasic side chains of the key arginine residues (Arg222and Arg249) are different relative to the bindingpocket, suggesting that there may be bindinginteractions involving other arginine (Arg198) andlysine (Lys194) residues surrounding the pocket. Thenotably different distribution of charged residuescompared to PAH1 and PAH2 domains supports thedistinct binding capacity observed for AtALY2.
Discussion
We have identified a family of Arabidopsis SIN3coregulator encoding genes that have been placedinto three distinct groups following phylogeneticanalysis. We propose systematically naming thesegenes SNL1, SNL2, SNL3, SNL4, SNL5 and SNL6.Each member of this coregulator family was shownto be expressed in Arabidopsis plants, suggesting thatthey each play a functional role. Analysis of SNLpeptide sequences identified the presence of con-served PAHandHIDdomains that have been shownto be essential for SIN3 function in other eukaryotes.Using homology modelling, we have shown thatwhilst the PAH2 domain of SNL proteins show ahigh degree of conservation with the NMR structureof mSIN3A, discrete structural differences betweenSNL family members on the inner DNA bindingprotein interaction surface of this domain are likelyto result in target gene specificity. Furthermore, themodelled structure of PAH3 indicates that there maybe more profound structural differences whencompared to PAH1 and PAH2 domains, leading toa generally wider specificity.Through Arabidopsis protoplast-based artificial re-
cruitment repression assays, it was found that SNL1possesses an inherent transcription repression capa-bility that was dependent on functional HDACactivity in plants. Heterologous yeast assays demon-strated that the repression activity of SNL1 wasgreatly reduced in the absence of the HDAC RPD3,highlighting the degree of conservation in themolecular mechanism(s) utilised by SIN3 to represstranscription. SNL1 repression activity was notcompletely abolished in the absence of RPD3, whichis consistent with ySIN3 studies where ySIN3repression function was similarly reduced in anrpd3Δ yeast strain.30 This is not surprising, as fiveclass I (RPD3-like) HDACs are encoded by theS. cerevisiae genome, suggesting that a degree offunctional redundancy occurs. Consistently, mSIN3was found to interact with both HDAC1 andHDAC2, two highly related homologues of theyeast HDAC RPD3. In Arabidopsis, four class 1HDACs have been identified. HDA19 interacts withthe transcription corepressor LUG,31 whilst HDA6has been implicated in rDNA gene silencing.32
However, these two HDACs have a redundant rolein the repression of embryonic properties through the
repression of embryo-specific gene function.33 It willtherefore be interesting to define the specificity ofHDAC utilisation by SNL1 and the other members ofthe SNL coregulator protein family.A large-scale yeast two-hybrid screen identified a
number of significant interacting partners that wereconsistent with the nuclear localisation and functionof SNL1. One highly represented group contained anumber of known or predicted DNA bindingtranscription factors. The identification of thesetranscription factor partners offers a route to identi-fying the functional role of SNL1 in Arabidopsis.These predominantly Myb-domain-containing pro-teins included the Arabidopsis ALY homologuesAtALY2 and AtALY3. When tested for any PAHdomain specificity, AtALY2 was found to interactwith the PAH3 domain of SNL1. The retinoblastoma(Rb) tumour suppressor protein complex has beenshown to associate with both Drosophila and humanALY/LIN-9 proteins, which are involved in therepression of E2F-regulated genes. The Rb repressioncomplex associates with the SIN3 complex, with Rbfunctioning as a bridging molecule between E2Ftranscription factors and SAP18, a component of theSIN3 core complex.34 The interaction betweenAtALY2/3 and SNL1 raises the possibility thatthese proteins may have a similar function inArabidopsis. It is feasible that, like ALY/LIN-9proteins, both AtALY2 and AtALY3 may alsointeract directly with Rb to form an AtRb-SNL1-AtALY repression complex as they also contain aDIRP motif. A second group of proteins identified inour two-hybrid screen were the Arabidopsis telomere-binding proteins AtTBP1 and AtTRP2/TRFL1. Anal-ysis revealed that PAH2 was required to support theinteraction between SNL1 and AtTBP1. AtTBP1 andAtTRP2/TRFL1 have both been shown to specificallybind to the plant telomeric repeat sequence 5′-TTTAGGG-3′.22,23 In eukaryotes, the telomere isessential for the preservation of chromosome integ-rity and for protection from exonuclease degrada-tion. Both AtTBP1 and AtTRP2/TRFL1 are membersof the double-stranded telomere-repeat DNA-bind-ing protein family35 and encode a characteristic C-terminal Myb domain required for telomere binding.AtTBP1 and AtTRP2/TRFL1 are homologues of thehuman Myb-like telomere-binding proteins TRF1and TRF2 and the yeast homologue yRAP1. yRAP1is a multifunctional protein that has a specific role inthe maintenance of telomeres. Mutation of RAP1results in altered telomere length and also the loss oftelomeric silencing, which is suppressed throughmutations in either SIN3 or RPD3.36 Furthermore,loss of SIN3 has also been associated with shortenedtelomere length in genetic screens in S. cerevisiae.3
Given the conserved function of SIN3 and theidentification of the two telomere-binding proteinsAtTBP1 and AtTRP2/TRFL1, it seems plausible tosuggest that SNL1 may have a role in telomerestability in Arabidopsis.Through this and other studies, it is becoming clear
that members of the Arabidopsis SNL family andputative components of a core SNL complex play

946 Arabidopsis SIN3-LIKE (SNL) Coregulators
important biological roles. Initial evidence pointstoward the diverse role of SNL in regulatingtranscription and maintaining genome stabilitybeing conserved in Arabidopsis. Biochemical purifica-tion, genome-wide localisation, and further analysisin vivo using small interfering RNA knockdown areall now required to understand the importance of thisfamily of transcription coregulators in higher plants.
Methods
Bioinformatics and structural modelling
The ySIN3peptide sequence (M36822)was obtained fromthe SaccharomycesGenomeDatabase§ and used to search forArabidopsis orthologues using BLASTP at the MunichInformation Centre for Protein Sequences (MIPS)A. thalianaDatabase (MAtDB)‖. To identify other orthologous SIN3peptide sequences, a BLASTP screen of both the NationalCenter for Biotechnology Information (NCBI) and MIPSdatabases was performed. Returned sequences were man-ually screened to remove duplication, resulting in 51 uniquesequences. Peptide sequences were aligned using ClustalX2.0.10. Within the Protein Gap Parameters, Residue-specificand hydrophilic Penalties were ‘on’, with HydrophilicResidues being GPSNDQEKR. The Gap Separation Dis-tance [0–100] was 4 with End Gap Separation ‘off’. WithintheAlignment Parameters theMultiple Parameters were setto the following: Gap Opening [0–100] 10, Gap Extension[0–100] 0.2, Delay Divergent Sequences (%) 30, DNATransition Weight [0–1] 0.5 and Use Negative Matrix ‘off’.The Gonnet Series Protein Weight Matrix was used. Thetree and bootstrap values were then plotted using NJplot.Homology models were generated for the PAH2
domains of SNL1, SNL2 and SNL5 based on the resolvedcrystal structures of PAH domains deposited in the PDB(2F05_A, 1PD7_A, 1S5R_B, 1G1E_B, 1E91_A, 1S5Q_B).37
Structural modelling was carried out using a homologymodelling pipeline built with the Biskit structural bioin-formatics platform.38 Our pipeline workflow incorporatesthe NCBI tools platform,39 including the BLASTprogram40 for similarity searching of sequence databases.In this case, protein sequences corresponding to the PDBof protein structures were searched for homology with theSNL proteins in to identify putative structural homo-logues. T-COFFEE41 was used for alignment of the testsequence with the template, followed by 10 iterations ofthe MODELLER program42 for homology modelling, andthe DSSP algorithm43 for secondary-structure validation.The PAH2 domains, residues 131–210 of SNL1, 125–204 ofSNL2 and 118–197 of SNL5 were modelled by 46%identity with the PAH crystal structure homologues, andthe PAH3 domain of SNL1, residues 331–400, wasmodelled by 27.5% identity. The resultant homologymodels were validated by cross-reference to the second-ary-structure predictions. Homology models were gener-ated with 10 iterations of the MODELLER program.
Plant culture
All plants were in Columbia-0 background and weregrown in Aratrays (Betatech, Belgium) under constant
§http://www.yeastgenome.org/‖http://mips.gsf.de/proj/plant/jsf/athal/index.jsp
conditions (20 °C, constant 2400-lx globe lighting).Rosette leaves were removed (stage 6.9) for DNA andRNA analysis.
Real-time PCR
Total RNA was isolated (RNeasy Plant Mini system,with on-column DNase I digestion step; Qiagen) from100 mg of rosette leaves. Random decamer primers wereused to reverse-transcribe 1 μg of total RNA (RETRO-script, Ambion). The cDNA was used as a template foranalysing target gene expression (Absolute QPCR SYBRgreen, ABgene) using gene-specific primer pairs (BeaconDesigner, Premier BioSoft) (see Supplementary Data forprimer sequences).ACTIN2was used as an internal reference, and genomic
DNA and RNA were used as positive and negativecontrols, respectively. Relative quantification of geneexpression data was carried out using the thresholdcycle (Ct) values for each sample. Serial dilutions ofcDNA were used to trace a calibration curve, which wasthen used to quantify gene expression levels in each groupof samples by plotting their respective Ct value in thecurve. Expression levels were then calculated as foldenrichment compared to that of ACTIN2 and plotted.
Arabidopsis protoplast repression assay
The SNL1 Arabidopsis (Columbia) root cDNA cloneAV547444 in pBluescriptII SK− was obtained fromKazuza DNA Research Institute, Japan,44 and cloned intopJG31 following site-directed mutagenesis (SDM) to createan SmaI restriction site upstream of the SNL1 translationstart (QuickChange XL SDMKit, Stratagene). Primers usedwere forward, 5′-GTGGACAATCCCCGGGCCACGG-CAGTGG-3′, and reverse, 5′-CCACTGCCGTGGCCCG-GGGATTGTCCAC-3′. To verify SDM, the recoveredplasmid was sequenced using M13 forward and reverseprimers. An SmaI–SnaBI fragment containing SNL1 wasexcised from the AV547444 pBluescriptII SK-constructcontaining the engineered SmaI site and ligated into theSmaI and SnaBI restriction sites of pJG.31 pGAL4-SNL1wasrecovered and sequenced (using the primer 5′-TCATCG-GAAGAGAGTAG-3′) to ascertain the orientation of theSNL1 insert.Protoplast isolation, transfection, and GUS repression
assays were performed as previously described.31 Briefly,protoplasts were prepared from Arabidopsis rosette leavesand transfected with 10 μg of each reporter–effecterplasmid combination using polyethylene glycol. Follow-ing overnight incubation, β-glucuronidase was extractedand activity was determined fluorometrically using 4-methylumbelliferyl glucuronide (MUG) substrate. Sam-ples were normalised for total protein content and forbackground fluorescence using untransfected protoplasts.Fluorescence activity was measured at 365 nm (excita-tion)/460 nm (emission) using a PolarStar Galaxy fluo-rimeter (BMG Labtech, UK). For HDAC inhibition assaysprotoplasts were treated with 20 μm TSA (Sigma-Aldrich)and incubated for an additional 6 h before GUS activitywas measured.
Heterologous yeast repression assay
Wild-type FT5 and FT5Δrpd3 yeast strains45 were usedin heterologous yeast repression assays. Two reporterplasmids were used that encode the β-galactosidase

947Arabidopsis SIN3-LIKE (SNL) Coregulators
(LACZ) gene, with (pJK1621) or without (pLG312ΔS) fourLEXA operator (binding) sites upstream of the CYC1UAS::CYC1TATA promoter sequence.45 The effecter plasmid(pAS64F2) encodes for the DNA binding protein LEXAin a low copy number yeast centromeric plasmid. SNL1was excised from pBluescriptII SK− using the restrictionenzymes AatII and SacII. pGEM7 (Promega) was digestedusing EcoRI. The 5′ overhang created by the EcoRIdigestion was filled in using Klenow (NEB). The Kle-now-treated pGEM7 was then digested with AatII. SNL1was then ligated into pGEM7. Plasmid DNA was purifiedand sequenced to confirm the presence of SNL1. Full-length SNL1 was digested with AatII, then blunt-endedusing mung bean nuclease (Promega) and subsequentlydigested with KpnI. This DNA fragment was cloned intopAS64F2 using SmaI and KpnI to give a LEXA-SNL1translational fusion under the control of the high-levelconstitutive ADH1 promoter.Yeast were grown overnight in liquid media and
transformed with the selected plasmids using a polyeth-ylene glycol–lithium acetate method. The transformedyeast were then plated onto appropriate selective media.Individual transformants were grown overnight in liquidmedia to early log phase and assayed for β-galactosidaseactivity. Briefly β-galactosidase was extracted from yeastcells and incubated with o-nitrophenol-β-D-galactosidase(ONPG). Hydrolysis of ONPG to a yellow-colouredderivative, o-nitrophenol, was measured at 420 nmusing a Beckman DU530 spectrophotometer. Valueswere normalised to OD600.
SNL1-CFP fusion construct
Full-length cDNA for SNL1was amplified by PCR fromCol-0 inflorescence-derived cDNA using the primers 5′-ATGAAGCGGATAAGAGATGAT-3′ and 5′-AGCTGA-GAAACCTATGGAACC-3′ with Pyrobest Taq polymer-ase (TaKaRa). attB1 and attB2 sites were introduced intothe amplified SNL1 open reading frame (lacking its stopcodon) by 20 cycles of nested PCR using chimericoligonucleotides containing attB1 plus 5′ SNL1 and attB2plus 3′ SNL1 sequences (Invitrogen), respectively, usingPyrobest Taq polymerase. Purified PCR product wascloned into pDONR Zeo (Invitrogen) using BP clonase,the SNL1 insert was sequenced, confirmed to lack anysequence errors and subsequently used to make a C-terminal CFP fusion in pEARLEYGATE 102 using LRclonase (Invitrogen). The SNL1-CFP junction was se-quenced to confirm it was in frame, and the SNL1-CFPplasmid was introduced into Agrobacterium tumefaciensGV3101 by electroporation. ER-YFP has been previouslydescribed.13
Transient expression of SNL1 in tobacco leaves
Expanded leaves (fourth to sixth) of Nicotiana tabacumSR1 plants were infiltrated with an overnight culture of A.tumefaciens GV3101 containing different constructs.46
Bacterial cells were washed twice in infiltration buffercontaining 10 μM acetosyringone (Aldrich) diluted to anOD600 of 0.03 or 0.1 in the same buffer, and infiltrated onthe abaxial surface of leaves. Infiltrated plants werereturned to the growth chamber at 20 °C and were usedfor microscopic analysis after 48 h. Sections (0.5 cm2) werecut from leaves, distal to the primary infiltration site, andsections were mounted in water and analysed for SNL1-CFP (or ER-YFP) expression by confocal microscopy on aZeiss CLSM 510.
Yeast two-hybrid analysis
The full-length coding sequence for SNL1 was PCR-amplified and cloned into pB66 as a C-terminal fusion toGal4 DNA-binding domain (N-Gal4-SNL1-C). The con-struct was checked by sequencing the entire insert andused as a bait to screen a random-primed A. thalianaseedlings cDNA library cloned into pP6. pB66 and pP6 arederived from pAS2ΔΔ47 and pGADGH plasmids, respec-tively. Eighty-eight million clones (8.8-fold the complexityof the library) were screened using a mating approachwith Y187 (matα) and CG1945 (mata) yeast strains aspreviously described.47 A total of 262 His+ colonies wereselected on a medium lacking tryptophan, leucine andhistidine. The prey fragments of the positive clones wereamplified by PCR and sequenced at their 5′ and 3′junctions. The resulting sequences were used to identifythe corresponding interacting proteins in the GenBankdatabase (NCBI). A confidence score (predicted biologicalscore) was attributed to each interaction as previouslydescribed.17
Protein–protein interaction assays
Full-length SNL1 was isolated from wild-type yeast(FT5) transformed with pAS-SNL1 as a LexA-hybridprotein as previously described.31 LexA-SNL1 deletionderivatives were produced from PCR templates using aPromega TNT T7 Quick for PCR in vitro transcription–translation system using template DNA amplified frompAS-SNL1 (see Supplementary Data online for primersequences). Total protein isolated from yeast or TNTreactions was incubated overnight at 4 °C with 15 μl ofanti-LexA mouse monoclonal antibody [LexA (2-12),Santa Cruz Biotechnology] in immunoprecipitation buffer[50 mM Tris–HCl (pH 8.0), 150 mM NaCl, 5 mMethylenediaminetetraacetic acid (EDTA), 1% Triton X-100, 1 mM DTT and complete protease inhibitors cocktailII (Calbiochem)]. Following incubation, 50 μl of activatedprotein G magnetic beads (Qiagen) was added to theextracts and incubated on ice for 2 h with vortexing at 15-min intervals before immobilised proteins were washedwith immunoprecipitation buffer. Plasmids containingAtALY219 and TBP148 were used to direct coupledtranscription–translation using T7 polymerase (TNT Sys-tem, Promega). [35S]Methionine-labelled protein (10 μl)was incubated with 100 μl of bead-immobilised LexA-hybrid protein in 1 ml binding buffer [20 mM Tris–HCl(pH 8), 50 mM NaCl, 50 mM KCl, 5 mM MgCl2, 0.2%Triton X-100, 0.2% bovine serum albumin, 0.2 mM EDTA,1 mM DTT and complete protease inhibitors cocktail II(Calbiochem)] overnight at 4 °C. Following incubation thebeads were washed extensively in binding buffer, elutedin SDS-PAGE gel loading buffer, and analysed by SDS-PAGE for detection of 35S-labeled proteins by phosphor-imaging (Pharos FX Plus, Bio-Rad), and LexA-hybridproteins by immunoblot with the antibody anti-LexA(dilution 1:200) and chemiluminescence detection (GEHealthcare).
Acknowledgements
We gratefully acknowledge M. G. Hwang andM. H. Cho for the gift of an AtTBP1 containing

948 Arabidopsis SIN3-LIKE (SNL) Coregulators
plasmid. We would like to thank Zhongchi Liu forcomments and discussions. Yeast two-hybrid screen-ing was performed by Hybrigenics, S.A., Paris,France (http://www.hybrigenics.com). This workwas funded by a grant from the School of BiologicalSciences, Swansea University, UK, and SyngentaLtd, Jealots Hill, UK, and a research project grantfrom The Royal Society, UK, to RSC.
Supplementary Data
Supplementary data associated with this articlecan be found, in the online version, at doi:10.1016/j.jmb.2009.11.065
References
1. Silverstein, R. A. & Ekwall, K. (2005). Sin3: a flexibleregulator of global gene expression and genomestability. Curr. Genet. 47, 1–17.
2. Silverstein, R. A., Richardson, W., Levin, H., Allshire,R. & Ekwall, K. (2003). A new role for the transcrip-tional corepressor SIN3; regulation of centromeres.Curr. Biol. 13, 68–72.
3. Askree, S. H., Yehuda, T., Smolikov, S., Gurevich, R.,Hawk, J., Coker, C. et al. (2004). A genome-widescreen for Saccharomyces cerevisiae deletion mutantsthat affect telomere length. Proc. Natl Acad. Sci. USA,101, 8658–8663.
4. Pennetta, G. & Pauli, D. (1998). The Drosophila Sin3gene encodes a widely distributed transcription factoressential for embryonic viability. Dev. Genes Evol. 208,531–536.
5. Dannenberg, J. H., David, G., Zhong, S., van der Torre,J., Wong, W. H. & Depinho, R. A. (2005). mSin3Acorepressor regulates diverse transcriptional networksgoverning normal and neoplastic growth and survival.Genes Dev. 19, 1581–1595.
6. Cowley, S. M., Iritani, B. M., Mendrysa, S. M., Xu, T.,Cheng, P. F., Yada, J. et al. (2005). The mSin3Achromatin-modifying complex is essential for embryo-genesis and T-cell development. Mol. Cell. Biol. 25,6990–7004.
7. Song, C. P., Agarwal, M., Ohta, M., Guo, Y., Halfter,U., Wang, P. & Zhu, J. K. (2005). Role of an ArabidopsisAP2/EREBP-type transcriptional repressor in abscisicacid and drought stress responses. Plant Cell, 17,2384–2396.
8. Song, C. P. & Galbraith, D. W. (2006). AtSAP18, anorthologue of human SAP18, is involved in theregulation of salt stress and mediates transcriptionalrepression in Arabidopsis. Plant Mol. Biol. 60, 241–257.
9. Wu, K., Malik, K., Tian, L., Brown, D. & Miki, B.(2000). Functional analysis of a RPD3 histone deace-tylase homologue in Arabidopsis thaliana. Plant Mol.Biol. 44, 167–176.
10. Hennig, L., Taranto, P., Walser, M., Schonrock, N. &Gruissem, W. (2003). Arabidopsis MSI1 is required forepigenetic maintenance of reproductive development.Development, 130, 2555–2565.
11. Halleck, M. S., Pownall, S., Harder, K. W., Duncan, A.M., Jirik, F. R. & Schlegel, R. A. (1995). A widelydistributed putative mammalian transcriptional regu-lator containing multiple paired amphipathic helices,with similarity to yeast SIN3. Genomics, 26, 403–406.
12. Brubaker, K., Cowley, S. M., Huang, K., Loo, L.,Yochum, G. S., Ayer, D. E. et al. (2000). Solutionstructure of the interacting domains of the Mad-Sin3complex: Implications for recruitment of a chromatin-modifying complex. Cell, 103, 655–665.
13. Bhatt, A. M., Etchells, J. P., Canales, C., Lagodienko,A. & Dickinson, H. (2004). VAAMANA-a BEL1-likehomeodomain protein, interacts with KNOX proteinsBP and STM and regulates inflorescence stem growthin Arabidopsis. Gene, 328, 103–111.
14. Sridhar, V. V., Surendrarao, A., Gonzalez, D., Conlan,R. S. & Liu, Z. (2004). Transcriptional repression oftarget genes by LEUNIG and SEUSS, two interactingregulatory proteins for Arabidopsis flower develop-ment. Proc. Natl Acad. Sci. USA, 101, 11494–11499.
15. Foster, E., Hattori, J., Labbe, H., Ouellet, T., Fobert,P. R., James, L. E. et al. (1999). A tobacco crypticconstitutive promoter, tCUP, revealed by T-DNAtagging. Plant. Mol. Biol. 41, 45–55.
16. Tzamarias, D. & Struhl, K. (1994). Functional dissec-tion of the yeast Cyc8-Tup1 transcriptional co-repressor complex. Nature, 369, 758–761.
17. Formstecher, E., Aresta, S., Collura, V., Hamburger,A., Meil, A., Trehin, A. et al. (2005). Protein interactionmapping: a Drosophila case study. Genome Res. 15,376–384.
18. Wojcik, J., Boneca, I. G. & Legrain, P. (2002).Prediction, assessment and validation of proteininteraction maps in bacteria. J. Mol. Biol. 323, 763–770.
19. Bhatt, A. M., Zhang, Q., Harris, S. A., White-Cooper,H. & Dickinson, H. (2004). Gene structure andmolecular analysis of Arabidopsis thaliana ALWAYSEARLY homologs. Gene, 336, 219–229.
20. Tsukagoshi, H., Saijo, T., Shibata, D., Morikami, A. &Nakamura, K. (2005). Analysis of a sugar responsemutant of Arabidopsis identified a novel B3 domainprotein that functions as an active transcriptionalrepressor. Plant Physiol. 138, 675–685.
21. Ehsan, H., Reichheld, J. P., Durfee, T. & Roe, J. L.(2004). TOUSLED kinase activity oscillates during thecell cycle and interacts with chromatin regulators.Plant Physiol. 134, 1488–1499.
22. Hwang, M. G., Chung, I. K., Kang, B. G. & Cho, M. H.(2001). Sequence-specific binding property of Arabi-dopsis thaliana telomeric DNA binding protein 1(AtTBP1). FEBS Lett. 503, 35–40.
23. Hwang, M. G., Kim, K., Lee, W. K. & Cho, M. H.(2005). AtTBP2 and AtTRP2 in Arabidopsis encodeproteins that bind plant telomeric DNA and induceDNA bending in vitro. Mol. Genet. Genom. 273, 66–75.
24. Wang, Y. & Yang, M. (2006). The ARABIDOPSISSKP1-LIKE1 (ASK1) protein acts predominately fromleptotene to pachytene and represses homologousrecombination in male meiosis. Planta, 223, 613–617.
25. Liu, F., Ni, W., Griffith, M. E., Huang, Z., Chang, C.,Peng, W. et al. (2004). The ASK1 and ASK2 genes areessential for Arabidopsis early development. Plant Cell,16, 5–20.
26. Xiao, W. & Jang, J. (2000). F-box proteins inArabidopsis. Trends Plant. Sci. 5, 454–457.
27. Andersen, P., Kragelund, B. B., Olsen, A. N., Larsen,F. H., Chua, N. H., Poulsen, F. M. & Skriver, K.(2004). Structure and biochemical function of aprototypical Arabidopsis U-box domain. J. Biol. Chem.279, 40053–40061.
28. Sun, Z. W. & Hampsey, M. (1999). A generalrequirement for the Sin3-Rpd3 histone deacetylasecomplex in regulating silencing in Saccharomycescerevisiae. Genetics, 152, 921–932.

949Arabidopsis SIN3-LIKE (SNL) Coregulators
29. Le Guezennec, X., Vermeulen, M. & Stunnenberg,H. G. (2006). Molecular characterization of Sin3PAH-domain interactor specificity and identificationof PAH partners. Nucleic Acids Res. 34, 3929–3937.
30. Kadosh, D. & Struhl, K. (1998). Histone deacetylaseactivity of Rpd3 is important for transcriptionalrepression in vivo. Genes Dev. 12, 797–805.
31. Gonzalez, D., Bowen, A. J., Carroll, T. S. & Conlan, R.S. (2007). The transcription corepressor LEUNIGinteracts with the histone deacetylase HDA19 andmediator components MED14 (SWP) and CDK8(HEN3) to repress transcription. Mol. Cell. Biol. 27,5306–5315.
32. Earley, K., Lawrence, R. J., Pontes, O., Reuther, R.,Enciso, A. J., Silva, M. et al. (2006). Erasure of histoneacetylation by Arabidopsis HDA6 mediates large-scalegene silencing in nucleolar dominance. Genes Dev. 20,1283–1293.
33. Tanaka, M., Kikuchi, A. & Kamada, H. (2008). TheArabidopsis histone deacetylases HDA6 and HDA19contribute to the repression of embryonic propertiesafter germination. Plant Physiol. 146, 149–161.
34. Binda, O., Roy, J. S. & Branton, P. E. (2006). RBP1family proteins exhibit SUMOylation-dependent tran-scriptional repression and induce cell growth inhibi-tion reminiscent of senescence. Mol. Cell. Biol. 26,1917–1931.
35. Kuchar, M. & Fajkus, J. (2004). Interactions of putativetelomere-binding proteins in Arabidopsis thaliana:identification of functional TRF2 homolog in plants.FEBS Lett. 578, 311–315.
36. Vannier, D., Damay, P. & Shore, D. (2001). A role forSds3p, a component of the Rpd3p/Sin3p deacetylasecomplex, in maintaining cellular integrity in Saccharo-myces cerevisiae. Mol. Genet. Genomics, 265, 560–568.
37. Berman, H. M., Westbrook, J., Feng, Z., Gilliland, G.,Bhat, T. N., Weissig, H. et al. (2000). The Protein DataBank. Nucleic Acids Res. 28, 235–242.
38. Grunberg, R., Nilges, M. & Leckner, J. (2007). Biskit-asoftware platform for structural bioinformatics.Bioinformatics, 23, 769–770.
39. Wheeler, D. L., Barrett, T., Benson, D. A., Bryant, S. H.,Canese, K., Chetvernin, V. et al. (2008). Databaseresources of the National Center for BiotechnologyInformation. Nucleic Acids Res. 36, D13–21.
40. Altschul, S. F., Gish, W., Miller, W., Myers, E. W. &Lipman, D. J. (1990). Basic local alignment search tool.J. Mol. Biol. 215, 403–410.
41. Notredame, C., Higgins, D. G. & Heringa, J. (2000). T-Coffee: a novel method for fast and accurate multiplesequence alignment. J. Mol. Biol. 302, 205–217.
42. Eswar, N., John, B., Mirkovic, N., Fiser, A., Ilyin, V. A.,Pieper, U. et al. (2003). Tools for comparative proteinstructure modeling and analysis. Nucleic Acids Res. 31,3375–3380.
43. Kabsch, W. & Sander, C. (1983). Dictionary of proteinsecondary structure: pattern recognition of hydrogen-bonded and geometrical features. Biopolymers, 22,2577–2637.
44. Asamizu, E., Nakamura, Y., Sato, S. & Tabata, S.(2000). A large scale analysis of cDNA in Arabidopsisthaliana: generation of 12,028 non-redundantexpressed sequence tags from normalized and size-selected cDNA libraries. DNA Res. 7, 175–180.
45. Kadosh, D. & Struhl, K. (1997). Repression by Ume6involves recruitment of a complex containing Sin3corepressor and Rpd3 histone deacetylase to targetpromoters. Cell, 89, 365–371.
46. Batoko, H., Zheng, H. Q., Hawes, C. & Moore, I.(2000). A rab1 GTPase is required for transportbetween the endoplasmic reticulum and Golgi appa-ratus and for normal Golgi movement in plants. PlantCell, 12, 2201–2218.
47. Fromont-Racine, M., Rain, J. C. & Legrain, P. (1997).Toward a functional analysis of the yeast genomethrough exhaustive two-hybrid screens.Nat. Genet. 16,277–282.
48. Hwang, M. G. & Cho, M. H. (2007). Arabidopsisthaliana telomeric DNA-binding protein 1 is requiredfor telomere length homeostasis and its Myb-exten-sion domain stabilizes plant telomeric DNA binding.Nucleic Acids Res. 35, 1333–1342.




















