
Ostreopsis cf. ovata bloom in the northern Adriatic Sea during summer 2009: Ecology, molecular...
8
Ostreopsis cf. ovata bloom in the northern Adriatic Sea during summer 2009: Ecology, molecular characterization and toxin profile Stefano Accoroni a , Tiziana Romagnoli a , Federica Colombo a , Chiara Pennesi a , Cristina Gioia Di Camillo a , Mauro Marini b , Cecilia Battocchi c , Patrizia Ciminiello d , Carmela Dell’Aversano d , Emma Dello Iacovo d , Ernesto Fattorusso d , Luciana Tartaglione d , Antonella Penna c , Cecilia Totti a,⇑ a Dipartimento di Scienze del Mare, Università Politecnica delle Marche, Via Brecce Bianche, 60131 Ancona, Italy b Istituto di Scienze Marine (CNR), Largo Fiera della Pesca, 60125 Ancona, Italy c Dipartimento di Scienze Biomolecolari, Università di Urbino, Viale Trieste 296, 61100 Pesaro, Italy d Dipartimento di Chimica delle Sostanze Naturali, Università degli Studi di Napoli ‘‘Federico II’’, Via D. Montesano 49, 80131 Napoli, Italy article info Keywords: Adriatic Sea Ostreopsis cf. ovata Benthic dinoflagellates HABs Palytoxins Nutrients abstract Intense blooms of the benthic dinoflagellate Ostreopsis cf. ovata have occurred in the northern Adriatic Sea since 2006. These blooms are associated with noxious effects on human health and with the mortality of benthic organisms because of the production of palytoxin-like compounds. The O. cf. ovata bloom and its relationships with nutrient concentrations at two stations on the Conero Riviera (northern Adriatic Sea) were investigated in the summer of 2009. O. cf. ovata developed from August to November, with the high- est abundances in September (1.3 Â 10 6 cells g 1 fw corresponding to 63.8 Â 10 3 cells cm 2 ). The pres- ence of the single O. cf. ovata genotype was confirmed by a PCR assay. Bloom developed when the seawater temperature was decreasing. Nutrient concentrations did not seem to affect bloom dynamics. Toxin analysis performed by high resolution liquid chromatography–mass spectrometry revealed a high total toxin content (up to 75 pg cell 1 ), including putative palytoxin and all the ovatoxins known so far. Ó 2011 Elsevier Ltd. All rights reserved. 1. Introduction Over the past decades, reports of harmful algal blooms have in- creased concerning both the geographical distribution of the spe- cies involved and the frequency of phenomena in both the northern and the southern hemispheres. This has been attributed either to global climate changes (Gómez and Claustre, 2003; Halle- graeff, 2010; Lejeusne et al., 2010) or to anthropogenic impacts, such as eutrophication and ballast water transfer (Anderson, 2009; Hallegraeff, 2010; Heisler et al., 2008), or it has been inter- preted as an apparent phenomenon, due to the worldwide increase in monitoring programs (Van Dolah, 2000). Benthic dinoflagellates belonging to the family Ostreopsidaceae are common members of benthic microalgal communities in both tropical and temperate areas (Faust et al., 1996; Rhodes, 2011). Two Ostreopsis species, O. cf. ovata and O. cf. siamensis, have been found with increasing frequency in several Mediterranean coastal areas (Aligizaki and Nikolaidis, 2006; Mangialajo et al., 2008; Penna et al., 2005; Totti et al., 2010; Turki et al., 2006; Vila et al., 2001). The correct identification of the two Mediterranean Ostreopsis species is important, since they have distinctive toxin profiles with different effects on human health and marine organisms. Neverthe- less, the identification of the Ostreopsis morphotypes at the species level continues to be problematic, at least in routine microscopy analysis, because of the species’ high morphometric variability, especially in terms of antero-posterior and dorso-ventral diameter ratios. Therefore, alternative diagnostic methods based on rapid, accurate and less time-consuming molecular methodologies can be highly advantageous. Recently, molecular analyses based on the PCR assay, which was developed and validated using species- specific Ostreopsis primers, were performed on environmental samples collected in different areas of the Mediterranean Sea. The results demonstrated that the O. cf. ovata genotype is more widespread than that of O. cf. siamensis in both the western and eastern Mediterranean (Battocchi et al., 2010; Penna et al., 2010). Ostreopsis blooms have often been associated with noxious ef- fects on human health (Gallitelli et al., 2005) and with the mortal- ity of benthic marine organisms (Simoni et al., 2003; Shears and Ross, 2009, 2010), because of the production of palytoxin-like com- pounds. Palytoxin (PLTX) is one of the most potent non-protein marine toxins (Deeds and Schwartz, 2010) known so far. It exhibits high toxicity in mammals with LD50 in the range of 25– 450 ng kg 1 , depending on the animal model and the administra- tion route. Recently, liquid chromatography–mass spectrometry 0025-326X/$ - see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.marpolbul.2011.08.003 ⇑ Corresponding author. Tel.: +39 071 2204647; fax: +39 071 2204650. E-mail address: [email protected] (C. Totti). Marine Pollution Bulletin 62 (2011) 2512–2519 Contents lists available at SciVerse ScienceDirect Marine Pollution Bulletin journal homepage: www.elsevier.com/locate/marpolbul
Transcript of Ostreopsis cf. ovata bloom in the northern Adriatic Sea during summer 2009: Ecology, molecular...

Marine Pollution Bulletin 62 (2011) 2512–2519
Contents lists available at SciVerse ScienceDirect
Marine Pollution Bulletin
journal homepage: www.elsevier .com/locate /marpolbul
Ostreopsis cf. ovata bloom in the northern Adriatic Sea during summer 2009:Ecology, molecular characterization and toxin profile
Stefano Accoroni a, Tiziana Romagnoli a, Federica Colombo a, Chiara Pennesi a, Cristina Gioia Di Camillo a,Mauro Marini b, Cecilia Battocchi c, Patrizia Ciminiello d, Carmela Dell’Aversano d, Emma Dello Iacovo d,Ernesto Fattorusso d, Luciana Tartaglione d, Antonella Penna c, Cecilia Totti a,⇑a Dipartimento di Scienze del Mare, Università Politecnica delle Marche, Via Brecce Bianche, 60131 Ancona, Italyb Istituto di Scienze Marine (CNR), Largo Fiera della Pesca, 60125 Ancona, Italyc Dipartimento di Scienze Biomolecolari, Università di Urbino, Viale Trieste 296, 61100 Pesaro, Italyd Dipartimento di Chimica delle Sostanze Naturali, Università degli Studi di Napoli ‘‘Federico II’’, Via D. Montesano 49, 80131 Napoli, Italy
a r t i c l e i n f o
Keywords:Adriatic SeaOstreopsis cf. ovataBenthic dinoflagellatesHABsPalytoxinsNutrients
0025-326X/$ - see front matter � 2011 Elsevier Ltd.doi:10.1016/j.marpolbul.2011.08.003
⇑ Corresponding author. Tel.: +39 071 2204647; faxE-mail address: [email protected] (C. Totti).
a b s t r a c t
Intense blooms of the benthic dinoflagellate Ostreopsis cf. ovata have occurred in the northern Adriatic Seasince 2006. These blooms are associated with noxious effects on human health and with the mortality ofbenthic organisms because of the production of palytoxin-like compounds. The O. cf. ovata bloom and itsrelationships with nutrient concentrations at two stations on the Conero Riviera (northern Adriatic Sea)were investigated in the summer of 2009. O. cf. ovata developed from August to November, with the high-est abundances in September (1.3 � 106 cells g�1 fw corresponding to 63.8 � 103 cells cm�2). The pres-ence of the single O. cf. ovata genotype was confirmed by a PCR assay. Bloom developed when theseawater temperature was decreasing. Nutrient concentrations did not seem to affect bloom dynamics.Toxin analysis performed by high resolution liquid chromatography–mass spectrometry revealed a hightotal toxin content (up to 75 pg cell�1), including putative palytoxin and all the ovatoxins known so far.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Over the past decades, reports of harmful algal blooms have in-creased concerning both the geographical distribution of the spe-cies involved and the frequency of phenomena in both thenorthern and the southern hemispheres. This has been attributedeither to global climate changes (Gómez and Claustre, 2003; Halle-graeff, 2010; Lejeusne et al., 2010) or to anthropogenic impacts,such as eutrophication and ballast water transfer (Anderson,2009; Hallegraeff, 2010; Heisler et al., 2008), or it has been inter-preted as an apparent phenomenon, due to the worldwide increasein monitoring programs (Van Dolah, 2000).
Benthic dinoflagellates belonging to the family Ostreopsidaceaeare common members of benthic microalgal communities in bothtropical and temperate areas (Faust et al., 1996; Rhodes, 2011).Two Ostreopsis species, O. cf. ovata and O. cf. siamensis, have beenfound with increasing frequency in several Mediterranean coastalareas (Aligizaki and Nikolaidis, 2006; Mangialajo et al., 2008;Penna et al., 2005; Totti et al., 2010; Turki et al., 2006; Vila et al.,2001).
All rights reserved.
: +39 071 2204650.
The correct identification of the two Mediterranean Ostreopsisspecies is important, since they have distinctive toxin profiles withdifferent effects on human health and marine organisms. Neverthe-less, the identification of the Ostreopsis morphotypes at the specieslevel continues to be problematic, at least in routine microscopyanalysis, because of the species’ high morphometric variability,especially in terms of antero-posterior and dorso-ventral diameterratios. Therefore, alternative diagnostic methods based on rapid,accurate and less time-consuming molecular methodologies canbe highly advantageous. Recently, molecular analyses based onthe PCR assay, which was developed and validated using species-specific Ostreopsis primers, were performed on environmentalsamples collected in different areas of the Mediterranean Sea. Theresults demonstrated that the O. cf. ovata genotype is morewidespread than that of O. cf. siamensis in both the western andeastern Mediterranean (Battocchi et al., 2010; Penna et al., 2010).
Ostreopsis blooms have often been associated with noxious ef-fects on human health (Gallitelli et al., 2005) and with the mortal-ity of benthic marine organisms (Simoni et al., 2003; Shears andRoss, 2009, 2010), because of the production of palytoxin-like com-pounds. Palytoxin (PLTX) is one of the most potent non-proteinmarine toxins (Deeds and Schwartz, 2010) known so far. It exhibitshigh toxicity in mammals with LD50 in the range of 25–450 ng kg�1, depending on the animal model and the administra-tion route. Recently, liquid chromatography–mass spectrometry

S. Accoroni et al. / Marine Pollution Bulletin 62 (2011) 2512–2519 2513
(LC–MS) disclosed the presence of putative PLTX and five newpalytoxin analogues, named ovatoxin-a, ovatoxin-b, ovatoxin-c,ovatoxin-d and ovatoxin-e, in field and in cultured samples of O.cf. ovata that were collected along the Italian coasts (Ciminielloet al., 2006, 2008, 2010, 2011; Guerrini et al., 2010). High resolu-tion (HR) LC–MS measurements, coupled with MS/MS experi-ments, indicated that ovatoxins present molecular formulaesimilar to those of palytoxins, with differences in their elementalcompositions residing either in the A or the B moiety of the paly-toxin molecule (Fig. 1). Palytoxin-like compounds can accumulatein shellfish, especially in filter-feeding molluscs, as discovered dur-ing monitoring programs of molluscs in Greece (Aligizaki andNikolaidis, 2006) and Campania (Italy) (ARPAC, 2008), where theyexert cytotoxic effects (Louzao et al., 2010). Regarding the toxic ef-fect of palytoxin in humans, it is known that the presence of paly-toxin in seafood can lead to clupeotoxism, i.e., an illness linked tohuman mortality (Onuma et al., 1999).
The dynamics of Ostreopsis blooms observed along the Mediter-ranean coasts are still not well understood because of the lack oftemporal and spatial series of data sets that analyze the microalgalabundances and their relationships with environmental factors.The effects of environmental factors on Ostreopsis blooms aresomewhat unclear. It has been suggested that temperature is akey factor for bloom development, attributing the recent Ostreopsisexpansion in the Mediterranean Sea to global warming (Granéliet al., 2011). However, it has also been recognized that the roleof temperature is not the same in all Mediterranean areas (Mang-ialajo et al., 2011). Hydrodynamics have been shown to play animportant role in regulating benthic/planktonic dinoflagellateabundances, especially in shallow coastal areas, enhancing theresuspension of benthic cells in the water column (Totti et al.,2010), although the fate of planktonic Ostreopsis cells is still poorlyknown. The relationship between benthic dinoflagellate blooms
Fig. 1. Structure of palytoxin. Molecular formulae (M) of ovatoxins and elemental comexperiments. (From Ciminiello et al. (2008, 2010).)
and nutrient concentration has been only sporadically investi-gated, although Armi et al. (2010) found a positive correlation be-tween nutrients and abundances for Coolia monotis.
In this study, we investigated the O. cf. ovata bloom and its rela-tionships with nutrient concentrations along the Conero Riviera(northern Adriatic Sea) during the summer of 2009. Furthermore,a molecular PCR-based analysis was applied to environmentalsamples to confirm the identity of Ostreopsis spp. genotypesthroughout the study period. The toxin profile of epiphytic O. cf.ovata samples was evaluated both qualitatively and quantitativelywith HR LC–MS analyses.
2. Materials and methods
2.1. Study area
The study was conducted at two stations along the Conero Rivi-era (Ancona, NW Adriatic Sea): st. 1 (Passetto) and st. 2 (Portonovo)(Fig. 2). Both stations are characterized by rocky bottoms and shal-low depths (<1 m) and are affected moderately by human impactduring the summer season. At st. 1, two sites were sampled, char-acterized by different hydrodynamic conditions: st. 1/L (sheltered)and st. 1/H (exposed). A third sheltered site (st. 1/LP) was sampledin order to collect samples for the toxin analysis. At st. 2, only onesite was sampled, which was exposed and therefore was character-ized by high hydrodynamic conditions.
2.2. Sampling
Sampling was conducted fortnightly from March 25 (st. 2) andMay 7 (st. 1) to October 28, 2009, until the appearance of the firstOstreopsis cells; the sampling frequency was then intensified (oncea week) until the bloom declined.
position of their relevant A and B moieties, as deduced by HR LC–MS and MS2
Fig. 2. Map of the study area, showing the location of the sampling stations alongthe Conero Riviera. 1: Passetto; 2: Portonovo.
2514 S. Accoroni et al. / Marine Pollution Bulletin 62 (2011) 2512–2519
The surface temperature was measured with a CTD, Model 30Handheld Salinity, YSI (Yellow Spring, OH, USA). Meteomarine con-ditions were annotated for each sampling site according to theDouglas scale, integrating those records with data from the web-site www.ilmeteo.it, which reports the wave height along the Ital-ian coast every 3 h.
Water samples for the nutrient analysis were collected in poly-ethylene bottles (50 ml) near the sampled substrata (see below),avoiding resuspension phenomena, immediately filtered throughGF/F Whatman filters (25 mm, nominal pore size 0.7 lm) andstored in triplicate in 4 ml polyethylene bottles at �22 �C untilthe analysis.
Benthic dinoflagellate abundances and composition were ana-lyzed both in benthic substrata (macrophytes and pebbles) andin the water column. Macrophytes and pebbles were collected inthree replicates at a depth of 0.5 m. Two seaweed species, Ulva rig-ida (Ulvophyceae) and Dictyota dichotoma (Phaeophyceae), werechosen among the most common macroalgal species in the studyarea. At the end of the summer, during the bloom peak, death ofmacroalgae was observed in the sampling area (see below), caus-ing the disappearance of D. dichotoma thalli. Therefore, Dictyopterispolypodioides (Phaeophyceae) and Hypnea musciformis (Rhodophy-ceae) were also sampled. To avoid a loss of epiphytic cells, all of thesubstrata were collected underwater in plastic bags and were pro-tected from light until they were transferred to the laboratory(within 2 h). At each station, the surface seawater (1 replicate)was also sampled to analyze the abundance of benthic dinoflagel-lates in the water column.
The toxin content was investigated in natural O. cf. ovata epi-phytic assemblages. Seaweed samples were collected on Septem-ber 18 at st. 1/L and st. 1/LP, September 19 at st. 2 and October21 at st. 1/L. The samples were treated as described below to allowthe detachment of the epiphytic cells.
2.3. Nutrient analysis
The analysis of nitrates, nitrites, ammonium, phosphates andsilicates was conducted following the colorimetric method byStrickland and Parsons (1968), using an Autoanalyzer QuAAtro
Axflow. Detectability limits were 0.02 lmol l�1 for nitrates, nitrites,ammonia and silica and 0.03 lmol l�1 for orthophosphates.
2.4. Sample treatment
The sample treatment was performed following the proceduredescribed in Totti et al. (2010). Seaweed samples were vigorouslyshaken for a few minutes to allow the dislodgement of epiphyticcells and then rinsed at least three times with FSW until the epi-phytic microalgae was completely removed.
Macrophyte thalli were then weighed to determine the fresh (gfw) and dry (g dw) weight. The area of seaweeds was calculatedwith image analysis software to obtain the surface area (cm2) ofdigitalized images obtained with a scanner. The fresh weight/dryweight/area ratio was then calculated for each macroalgal specieson a significant number of individuals to obtain a conversion fac-tor, allowing the thallus area to be estimated only by measuringwet or dry weight. Pebbles were accurately scraped with a bladeand rinsed with FSW. The area of the scraped surface was thenmeasured using a tape measure. The samples were preserved with0.8% neutralized formaldehyde and stored in the dark at 4 �C untilthe microscopy and molecular analyses.
2.5. Molecular analysis
A total of 22 macrophyte (Ulva rigida) and 21 seawater sampleswere analyzed. The volume of formalin-fixed samples was 40 mlfor macrophytes and 50 ml for seawater samples. Macrophytesamples were collected by centrifugation at 4000 rpm for 10 minat room temperature. The supernatant was discarded, 30 ml of fil-tered seawater was added, and the samples were again centrifugedat 4000 rpm for 10 min at room temperature. Surface seawatersamples were filtered through a 25 mm diameter Isopore mem-brane of pore size 3 lm (Millipore, USA). The pellets and filterscontaining Ostreopsis epiphytic cells obtained from the above pro-cedures were processed for total genomic DNA extraction, as de-scribed in Battocchi et al. (2010). The PCR amplification assaywas carried out using genus- (Ostreopsis) and species-specificprimers (O. cf. ovata and O. cf. siamensis), as described in Pennaet al. (2007) and Battocchi et al. (2010).
2.6. Microscopy counting
Benthic dinoflagellates were identified and counted using an in-verted microscope (Zeiss Axiovert 135) equipped with phase con-trast, at 200� magnification. Sub-samples (5–25 ml, dependingon observed cell abundances) were settled in counting chambersafter homogenization, according to the Utermöhl’s sedimentationmethod (Hasle, 1978). The identification of O. cf. ovata is basedon the layout and thecal plate morphology and dimensions (Stei-dinger and Tangen, 1997); thus, the samples were observed in epi-fluorescence after staining with fluorochrome (Calcofluor white).Counting was performed on a known area (10–30 random fields,1–2 transects, or the whole sedimentation chamber) in order tocount a representative cell number. The final data were expressedas cells g�1 fw or dw and cells cm�2 for macrophytes, cells cm�2 forhard substrata, and cells l�1 for the water column.
2.7. Evaluation of the toxin profile
All organic solvents were of the distilled-in-glass grade (CarloErba, Milan, Italy). Water was distilled and passed through a MilliQwater purification system (Millipore Ltd., Bedford, MA, USA). Aceticacid (Laboratory grade) was purchased from Carlo Erba. The ana-lytical standard of palytoxin was purchased from Wako ChemicalsGmbH (Neuss, Germany).

Table 1Retention times and most abundant ion peaks of the [M+2H�H2O]2+ and [M+2H+K]3+
ion clusters of palytoxin and ovatoxins used in quantitative analysis.
Toxin Rt (min) [M+2H�H2O]2+ [M+2H+K]3+
Palytoxin 10.78 1331.7436 906.8167Ovatoxin-a 11.45 1315.7498 896.1572Ovatoxin-b 11.28 1337.7623 910.8318Ovatoxin-c 10.90 1345.7584 916.1628Ovatoxin-d and -e 11.07 1323.7456 901.4884
S. Accoroni et al. / Marine Pollution Bulletin 62 (2011) 2512–2519 2515
The epiphytic samples (1A–4A) were treated as described aboveto detach epiphytic Ostreopsis cells. A 50 ml aliquot of each samplewas preserved with 0.8% neutralized formaldehyde for cell identi-fication and counting, while the rest was preserved for the chemi-cal analysis.
All the samples (except sample 3A, which was processed with-out separation as the filtering apparatus was damaged) were fil-tered under low pressure (<5 mbar) through nylon net filters(11 lm diameter, Millipore type NY 11) to separate cell pellets(1A = 1,340,456 cells; 2A = 1,028,503 cells; 4A = 37,314 cells) fromseawater (1A = 1700 ml; 2A = 1600 ml; 4A = 2000 ml). Pellet andseawater samples were separately processed for extraction. Eachpellet sample was added with 50 ml of a methanol/water (1:1, v/v) solution and sonicated for 30 min in pulse mode while coolingin an ice bath. The mixture was centrifuged at 3000 � g for30 min, the supernatant was decanted, and the pellet was washedtwice with 50 ml of methanol/water (1:1, v/v). The extracts werecombined, and the volume was adjusted to 160 ml with extractingsolvent. The obtained mixture was analyzed directly by HR LC–MS(5 ll injected). Each seawater sample was extracted five times withan equal volume of butanol. The butanol layer was evaporated todryness, dissolved in 45 ml of methanol/water (1:1, v/v) and ana-lyzed directly by LC–MS (5 ll injected).
The sample collected at st. 2 on September 19 (3A), containing12.5 � 106 cells in 1670 ml, was sonicated and centrifuged at3000 � g for 30 min. The supernatant was extracted five times withan equal volume of butanol. The butanol layer was evaporated todryness, dissolved in 250 ml of methanol/water (1:1, v/v) and ana-lyzed directly by LC–MS (5 ll injected).
The efficiency of the above extraction procedures was estimatedto be 98% for the pellet extracts (1A, 2A, and 4A) and 75% for sea-water extracts (1A–4A). These recovery percentages were used tocorrect the toxin content measured by HR LC–MS.
High resolution liquid chromatography–mass spectrometry (HRLC–MS) experiments were conducted on an Agilent 1100 LC binarysystem (Palo Alto, CA, USA) coupled to a hybrid linear ion trap LTQOrbitrap XL™ Fourier Transform MS (FTMS) that was equippedwith an ESI ION MAX™ source (Thermo-Fisher, San Josè, CA,USA). Chromatographic separation was accomplished using a3 lm gemini C18 (150 � 2.00 mm) column (Phenomenex, Tor-rance, CA, USA) maintained at room temperature and eluted at0.2 ml/min with water (eluent A) and 95% acetonitrile/water (elu-ent B), both containing 30 mM acetic acid. A slow gradient elutionwas used: 20–50% B over 20 min, 50–80% B over 10 min, 80–100%B in 1 min, and hold 5 min (Ciminiello et al., 2008). HR full MSexperiments (positive ions) were acquired at a resolving powerof 15,000 in the range m/z 800–1400, where each palytoxin-likecompound has bi-charged ions due to [M+H+K]2+, [M+H+Na]2+,and [M+2H]2+; tri-charged ions due to [M+2H+K]3+ and[M+2H+Na]3+; and a number of ions due to multiple water lossesfrom the [M+2H]2+ and [M+3H]3+ ions. The following source set-tings were used in all the HR LC–MS experiments: a spray voltageof 4 kV, a capillary temperature of 290 �C, a capillary voltage of22 V, a sheath gas and auxiliary gas flow of 35 and 1 (arbitraryunits). The tube lens voltage was set at 110 V.
The quantitative determination of putative palytoxin and theindividual ovatoxins in the extracts was accomplished using thecalibration curve of the palytoxin standard (triplicate injection)at four concentration levels of (25, 12.5, 6.25, and 3.13 ng ml�1).Because of the lack of standards for ovatoxins, ovatoxins were as-sumed to have the same molar response as palytoxin (Ciminielloet al., 2010). Extracted ion chromatograms (XIC) were obtainedby selecting the most abundant ion peaks of both [M+2H�H2O]2+
and [M+2H+K]3+ ion clusters contained in the HR full MS spectraof palytoxin and ovatoxins (Table 1). A mass tolerance of 5 ppmwas used. The measured detection limit for palytoxin for the sys-
tem was 1.9 ng ml�1. For samples 1A, 2A and 4A, the toxin contentsof the relevant pellet and seawater extracts were added up to ex-press the detailed and total toxin content of each sample.
2.8. Statistical analysis
Differences in the abundances of benthic dinoflagellates be-tween stations, substrata, and areas affected by different hydrody-namic regimen were assessed through a one-way analysis ofvariance (ANOVA). O. cf. ovata abundances were also tested for sig-nificant correlations (Pearson) with nutrient concentrations (phos-phate, silicate, nitrate, nitrite, ammonium and dissolved inorganicnitrogen (DIN)). When significant differences for the main effectwere observed (p < 0.05), a Tukey’s pairwise comparison test wasalso performed. The statistical analyses were conducted using Stat-istica (Statsoft) software.
3. Results
3.1. PCR-based analysis
PCR-based assays were performed on the natural microphyto-benthic and phytoplankton formalin-fixed samples. The PCRamplification reactions revealed only the presence of the genotypeOstreopsis cf. ovata in all examined samples by giving the amplifiedproducts of expected sizes of Ostreopsis sp. (92 bp) and O. cf. ovata(210 bp).
3.2. Spatial and temporal variability
O. cf. ovata abundances showed a comparable trend at both sta-tions. At st. 1/L, the peak was observed approximately 15 days ear-lier and presented higher abundances than at st. 2. The firstoccurrence of O. cf. ovata was recorded on July 29 at st. 1 and on July31 at st. 2 with low abundances (62 cells cm�2). The temporal trendin O. cf. ovata abundances on macroalgae and on rocks is shown inFig. 3c and d for st. 1/L and st. 2, respectively. The maximum abun-dances on macroalgae were recorded at st. 1/L on September 9(1.3 � 106 cells g�1 fw corresponding to 1.6 � 107 cells g�1 dw and63.8 � 103 cells cm�2). A second peak was observed on September29 (7.8 � 105 cells g�1 fw corresponding to 6.9 � 106 cells g�1 dwand 20.6 � 103 cells cm�2). Maximum abundances on pebbles wererecorded on September 15 (28.1 � 103 cells cm�2). At st. 2, only onepeak occurred between the end of September and the first week ofOctober (max. 26.1 � 103 cells cm�2, recorded on pebbles on Sep-tember 23). Abundance values were persistently high until the endof September at both stations while from the second week of Octo-ber they started to decrease until the bloom declined during the lastdays of the month, in correspondence to a strong meteorologicalperturbation.
O. cf. ovata cells were also observed in the water column at bothstations with quite variable abundances. Cells in the water columnappeared a few days after their appearance on the benthic sub-strata, showing a comparable temporal trend. However, we notedthat the abundances in the water column increased after moderate
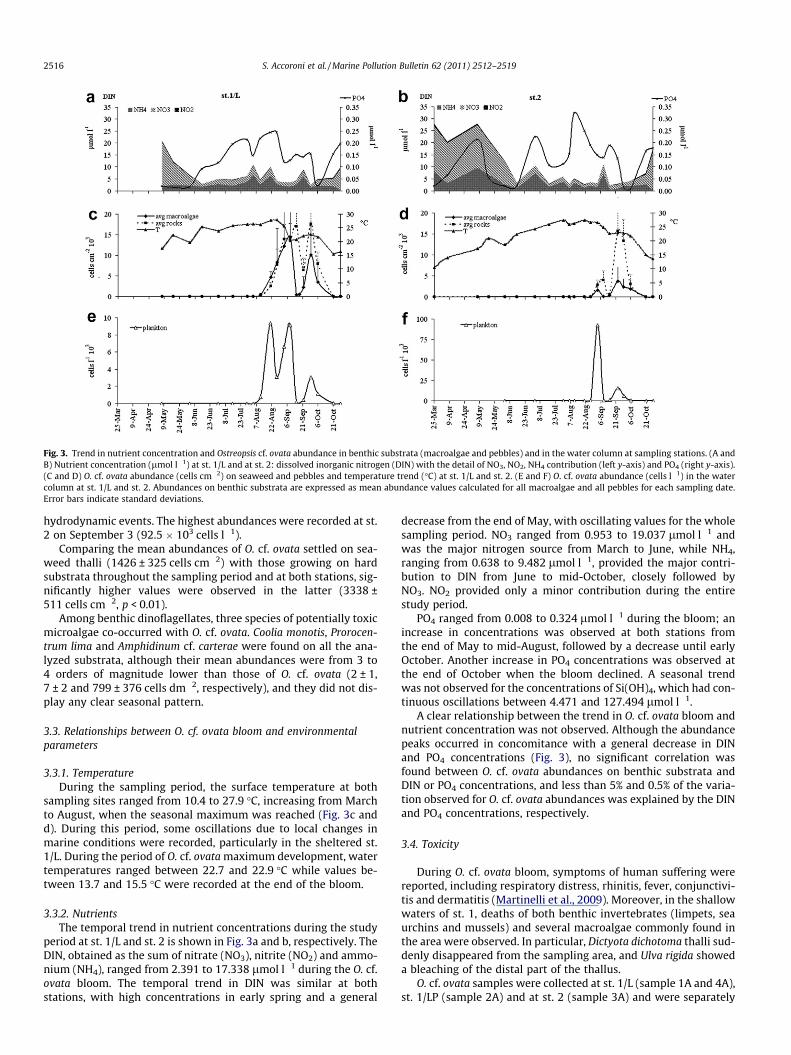
Fig. 3. Trend in nutrient concentration and Ostreopsis cf. ovata abundance in benthic substrata (macroalgae and pebbles) and in the water column at sampling stations. (A andB) Nutrient concentration (lmol l�1) at st. 1/L and at st. 2: dissolved inorganic nitrogen (DIN) with the detail of NO3, NO2, NH4 contribution (left y-axis) and PO4 (right y-axis).(C and D) O. cf. ovata abundance (cells cm�2) on seaweed and pebbles and temperature trend (�C) at st. 1/L and st. 2. (E and F) O. cf. ovata abundance (cells l�1) in the watercolumn at st. 1/L and st. 2. Abundances on benthic substrata are expressed as mean abundance values calculated for all macroalgae and all pebbles for each sampling date.Error bars indicate standard deviations.
2516 S. Accoroni et al. / Marine Pollution Bulletin 62 (2011) 2512–2519
hydrodynamic events. The highest abundances were recorded at st.2 on September 3 (92.5 � 103 cells l�1).
Comparing the mean abundances of O. cf. ovata settled on sea-weed thalli (1426 ± 325 cells cm�2) with those growing on hardsubstrata throughout the sampling period and at both stations, sig-nificantly higher values were observed in the latter (3338 ±511 cells cm�2, p < 0.01).
Among benthic dinoflagellates, three species of potentially toxicmicroalgae co-occurred with O. cf. ovata. Coolia monotis, Prorocen-trum lima and Amphidinum cf. carterae were found on all the ana-lyzed substrata, although their mean abundances were from 3 to4 orders of magnitude lower than those of O. cf. ovata (2 ± 1,7 ± 2 and 799 ± 376 cells dm�2, respectively), and they did not dis-play any clear seasonal pattern.
3.3. Relationships between O. cf. ovata bloom and environmentalparameters
3.3.1. TemperatureDuring the sampling period, the surface temperature at both
sampling sites ranged from 10.4 to 27.9 �C, increasing from Marchto August, when the seasonal maximum was reached (Fig. 3c andd). During this period, some oscillations due to local changes inmarine conditions were recorded, particularly in the sheltered st.1/L. During the period of O. cf. ovata maximum development, watertemperatures ranged between 22.7 and 22.9 �C while values be-tween 13.7 and 15.5 �C were recorded at the end of the bloom.
3.3.2. NutrientsThe temporal trend in nutrient concentrations during the study
period at st. 1/L and st. 2 is shown in Fig. 3a and b, respectively. TheDIN, obtained as the sum of nitrate (NO3), nitrite (NO2) and ammo-nium (NH4), ranged from 2.391 to 17.338 lmol l�1 during the O. cf.ovata bloom. The temporal trend in DIN was similar at bothstations, with high concentrations in early spring and a general
decrease from the end of May, with oscillating values for the wholesampling period. NO3 ranged from 0.953 to 19.037 lmol l�1 andwas the major nitrogen source from March to June, while NH4,ranging from 0.638 to 9.482 lmol l�1, provided the major contri-bution to DIN from June to mid-October, closely followed byNO3. NO2 provided only a minor contribution during the entirestudy period.
PO4 ranged from 0.008 to 0.324 lmol l�1 during the bloom; anincrease in concentrations was observed at both stations fromthe end of May to mid-August, followed by a decrease until earlyOctober. Another increase in PO4 concentrations was observed atthe end of October when the bloom declined. A seasonal trendwas not observed for the concentrations of Si(OH)4, which had con-tinuous oscillations between 4.471 and 127.494 lmol l�1.
A clear relationship between the trend in O. cf. ovata bloom andnutrient concentration was not observed. Although the abundancepeaks occurred in concomitance with a general decrease in DINand PO4 concentrations (Fig. 3), no significant correlation wasfound between O. cf. ovata abundances on benthic substrata andDIN or PO4 concentrations, and less than 5% and 0.5% of the varia-tion observed for O. cf. ovata abundances was explained by the DINand PO4 concentrations, respectively.
3.4. Toxicity
During O. cf. ovata bloom, symptoms of human suffering werereported, including respiratory distress, rhinitis, fever, conjunctivi-tis and dermatitis (Martinelli et al., 2009). Moreover, in the shallowwaters of st. 1, deaths of both benthic invertebrates (limpets, seaurchins and mussels) and several macroalgae commonly found inthe area were observed. In particular, Dictyota dichotoma thalli sud-denly disappeared from the sampling area, and Ulva rigida showeda bleaching of the distal part of the thallus.
O. cf. ovata samples were collected at st. 1/L (sample 1A and 4A),st. 1/LP (sample 2A) and at st. 2 (sample 3A) and were separately

Table 2Toxin content (pg cell�1) determined by HR LC–MS of epiphytic Ostreopsis cf. ovata samples collected along the northern Adriatic coast.
Sample # Station Date Palytoxin Ovatoxin-a Ovatoxin-b Ovatoxin-c Ovatoxin-d + e Total
1A St. 1/L 18th September 2 40 13 4 13 722A St. 1/LP 18th September 1 21 6 1 6 353A St. 2 19th September 0.3 8 2 0.4 3 144A St. 1/L 21st October nd 49 15 nd 11 75
nd: not detectable.
S. Accoroni et al. / Marine Pollution Bulletin 62 (2011) 2512–2519 2517
extracted as reported in the experimental procedure. High resolu-tion LC–MS experiments were conducted on the crude extracts toevaluate the toxin profile by using a slow gradient system, whichallowed chromatographic separation of the major components ofthe toxin profile. The presence of putative palytoxin and all theovatoxins (ovatoxin-a, ovatoxin-b, ovatoxin-c, ovatoxin-d, andovatoxin-e) recently identified in O. cf. ovata was demonstratedin all the analyzed samples.
Table 2 reports the total toxin content of the analyzed samples,expressed as pg cell�1, and the detailed toxin profile. On a per-cellbasis, samples 1A and 4A showed a very high total toxin content(72–75 pg cell�1), followed by samples 2A (35 pg cell�1) and 3A(14 pg cell�1). Ovatoxin-a was the major component of the toxinprofile for all the analyzed samples. It was present at levels of21–49 pg cell�1 in samples 1A, 2A and 4A, and at levels of8 pg cell�1 in sample 3A, followed by ovatoxin-b (2–15 pg cell�1)and ovatoxin-d + e (3–13 pg cell�1). Ovatoxin-c and putative paly-toxin were, indeed, present at very low levels (0.3–2 and 0.4–4 pg cell�1, respectively) in samples 1A, 2A and 3A. They wereundetectable in sample 4A.
4. Discussion
Ostreopsis cf. ovata blooms occurring along the Conero Rivieraseem to be among the most intense of the entire Mediterranean ba-sin (Mangialajo et al., 2011), with maximum abundances reachingan order of magnitude of 106 cells g�1 fw (corresponding to107 cells g�1 dw and 104 cells cm�2) in late summer, as observedduring the 2007 bloom. Significantly higher O. cf. ovata abundanceswere reached in the sheltered site than in the exposed one, as al-ready observed in several other studies, highlighting the importanceof hydrodynamism as a major factor affecting the dynamics andintensity of benthic dinoflagellate blooms (Barone, 2007; Shearsand Ross, 2009; Totti et al., 2010). Comparing the mean abundancesof O. cf. ovata on benthic substrata, significantly higher values wereobserved on pebbles, confirming that living substrata support lowerconcentrations of epibionts (Totti et al., 2010), probably due to theproduction of some allelopathic compounds (Jin and Dong, 2003).
4.1. Relationships with environmental parameters
In this study, the highest abundances were recorded with tem-peratures between 22.7 and 22.9 �C, i.e., after the maximum sum-mer value (27.9 �C). Some authors have suggested that globalwarming may be considered the cause of Ostreopsis expansion inthe Mediterranean (Granéli et al., 2011; Hallegraeff, 2010; Lejeusneet al., 2010), and several studies have hypothesized high sea watertemperature to be the major factor affecting the bloom (Aligizakiand Nikolaidis, 2006; Mangialajo et al., 2008; Tognetto et al.,1995). However, in the N Adriatic, such a positive correlation be-tween Ostreopsis proliferation and water temperatures has notbeen clearly shown, as blooms develop and reach maximum abun-dances at temperature values well below the summer maximum(Monti et al., 2007; Totti et al., 2010). This was also confirmed inexperimental conditions, as a strain isolated from the Conero areareached higher cell yields at 20 �C than at 25 and 30 �C (Pezzolesi
et al., in press). The Ostreopsis proliferation was also not associatedwith high temperature values in the Catalan Sea (Vila et al., 2001)and in Russian waters, where maximum abundances(10.9 � 103 cells g�1 dw) were found at a water temperature of9 �C (Selina and Orlova, 2010). This finding suggests that O. cf. ovat-a from different areas may exhibit different temperature affinities.Further studies are needed to clarify whether high temperaturesmay affect algal development at any stage of the life cycle, suchas cyst germination. Moreover, temperature may also have aninfluence on the toxin content, as it has been shown that the levelof toxicity per cell increases at temperature values that are sub-optimal for cell growth (Granéli et al., 2011).
Recent studies have provided increasing evidence of a link be-tween the nutrient enrichment of coastal waters (anthropogeniceutrophication) and harmful algal events (Glibert and Burkholder,2006; Glibert et al., 2010). However, such a relationship has notbeen shown for O. cf. ovata. Along the Conero Riviera, nutrient con-centrations were quite high throughout the sampling period, as ex-pected (Marini et al., 2002, 2008), since samples were collected nearthe sea bottom in high resuspension areas, and they did not appearto have decreased to limiting values. In our study, we did not ob-serve any clear relationship between the bloom trend and nutrientconcentrations. The observed decrease of DIN and PO4 in correspon-dence to the abundance peak may be explained by nutrient uptakeby algal cells during the period of bloom development. Our resultsare in agreement with literature data: Vila et al. (2001) and Shearsand Ross (2009) did not find any relation between epiphytic O. cf.ovata and nutrients, and Parsons and Preskitt (2007) found a corre-lation with nutrients for only a few epiphytic microalgal species,but not for O. cf. ovata. However, the scarcity of field measurementsof nutrient concentrations during O. cf. ovata blooms does not allowthe relationship between cell abundances and nutrient levels to beevaluated. Along the Italian coast, O. cf. ovata blooms have been ob-served at very different nutrient levels, (A.A.V.V., 2010), and world-wide Ostreopsis spp. appear to proliferate both in eutrophicated(Hallegraeff, 2010; Heisler et al., 2008) and in oligotrophic areas(Tindall and Morton, 1998; Zeldis et al., 2004). The trophic behaviorof Ostreopsis spp. is still not clear, and further studies are needed toclarify this aspect. Under experimental conditions, O. cf. ovatagrowth decreased as N and P decreased, and such effects seemedmore evident under N limited conditions (Pistocchi, personalcommunication). Barone (2007) hypothesized a mixotrophicbehavior, describing a particular defensive strategy performed byOstreopsis, where small organisms remain trapped in a mucilagi-nous web or mat. Moreover, we should consider that many HABgenera, including dinoflagellates, cyanobacteria, raphidophytesand pelagophytes, can use organic (dissolved or particulate) formsof nutrients for some or all of their nitrogen, phosphorus and/orcarbon demands (Cucchiari et al., 2008; Heisler et al., 2008).
4.2. Toxicity
This is the first study to provide the detailed toxin profile of nat-ural O. cf. ovata samples from the northern Adriatic Sea. Toxin pro-files of samples collected from different sites on the Conero Rivieraappeared to be dominated by ovatoxin-a, but the presence of newovatoxins recently detected in Adriatic O. cf. ovata cultures

2518 S. Accoroni et al. / Marine Pollution Bulletin 62 (2011) 2512–2519
(Ciminiello et al., 2010), as well as putative palytoxins, was alsohighlighted. Relative abundances of individual toxins were similarin different samples: ovatoxin-a was by far the major componentof the toxin profile, representing 55–60% of the total toxin content,followed by ovatoxin-b (14–24%), ovatoxin-d and -e (15–21%), ova-toxin-c (3–5%) and putative palytoxin (2–3%). The observed rela-tive abundance of individual compounds is in agreement withpreviously reported data. Ovatoxin-a also dominated the toxin pro-files of field and cultured O. cf. ovata samples from the Liguriancoasts (Ciminiello et al., 2008), as well as Tyrrhenian and AdriaticO. cf. ovata cultures (Ciminiello et al., 2010; Guerrini et al., 2010).
However, some variability in total toxin content, on a per-cellbasis, was observed among samples collected at different sites (Ta-ble 2). As an example, the major component of the toxin profile,ovatoxin-a, was contained at levels of 40 and 49 pg cell�1 in sam-ples 1A and 4A, respectively, whereas much lower levels weremeasured in samples collected at st. 1/LP (21 pg cell�1) and at st.2 (8 pg cell�1). It should be noted that sample 4A, despite the highovatoxin-a content, did not contain a detectable amount of eitherputative palytoxin or ovatoxin-c. Ovatoxin-a content in samplescollected at st. 1/L from the Conero Riviera was comparable withthat of Adriatic O. cf. ovata cultures reported by Guerrini et al.(2010) (42 pg cell�1) and Ciminiello et al. (2010) (18 pg cell�1)proving that the toxin profiles of natural samples and strainsgrown in the laboratory are quite similar. Moreover, the observedovatoxin-a levels appear higher than those previously found infield (1–3 pg cell�1) and cultured (3.8 pg cell�1) samples from theLigurian Sea. This finding confirms previous observations, i.e., thatnorthern Adriatic samples present a higher toxin content thanthose from the Ligurian (Bellocci et al., 2008) and Tyrrhenian Seas(Guerrini et al., 2010; Rossi et al., 2010). Thus, regarding toxin con-tent, Adriatic O. cf. ovata blooms appear to be more toxic thanthose occurring in other Italian areas with a higher potential risk(Guerrini et al., 2010). This toxicity is reflected in the intoxicationcases for humans (Martinelli et al., 2009) and in the death of ben-thic marine organisms, such as shellfish (Patella coerulea, Mytilusgalloprovincialis) and sea urchins (Paracentrotus lividus), in the Con-ero coast, typically observed every year only during periods of O. cf.ovata blooms.
Toxins were also found in mussel tissues collected from naturalbanks in the bloom area at a concentration of 91 lg kg�1 soft tissueof OVTX-a (Poletti, personal communication). Rhodes et al. (2006)reported concentrations of 1.2–14.5 lg kg�1 of total palytoxin inbivalve molluscs and sea urchins after feeding with Ostreopsissiamensis in the laboratory, and high levels of putative palytoxinwere reported in shellfish collected from Greek natural banks(33.3–96.8 lg kg�1, Aligizaki et al., 2008), reaching values compa-rable with our results. Although, to date, the effects of palytoxinsin invertebrates have not been clarified, evidence of the death ofbenthic marine organisms observed in several areas and variouscases of noxious effects on human health associated with blooms,indicate that further studies are needed on toxin production andtheir effects to optimize the management of coastal monitoring.
Acknowledgments
This research was funded by ISPRA-Italian Ministry of the Envi-ronment and MURST (PRIN 2007).
References
A.A.V.V., 2010. Relazione finale del Programma di Ricerca Ostreopsis ovata eOstreopsis spp.: nuovi rischi di tossicità microalgale nei mari italiani’’. ISPRA –MATTM, luglio 2010, p. 492.
Aligizaki, K., Nikolaidis, G., 2006. The presence of the potentially toxic generaOstreopsis and Coolia (Dinophyceae) in the north Aegean Sea, Greece. HarmfulAlgae 5, 717–730.
Aligizaki, K., Katikou, P., Nikolaidis, G., Panou, A., 2008. First episode of shellfishcontamination by palytoxin-like compounds from Ostreopsis species (AegeanSea, Greece). Toxicon 51, 418–427.
Anderson, D.M., 2009. Approaches to monitoring, control and management ofharmful algal blooms (HABs). Ocean Coast. Manage. 52, 342–347.
Armi, Z., Turki, S., Trabelsi, E., Ben Maiz, N., 2010. First recorded proliferation ofCoolia monotis (Meunier, 1919) in the North Lake of Tunis (Tunisia)correlation with environmental factors. Environ. Monit. Assess. 164, 423–433.
ARPAC, 2008. Il monitoraggio dell’ Ostreopsis ovata lungo il litorale della Campania(giugno–agosto 2007). Available from: <http://www.arpacampania.it/pubblicazioni.asp/>.
Barone, R., 2007. Behavioural trait of Ostreopsis ovata (Dinophyceae) inMediterranean rock pools: the spider’s strategy. Harmful Algae News, 1–3.
Battocchi, C., Totti, C., Vila, M., Masó, M., Capellacci, S., Accoroni, S., Reñé, A.,Scardi, M., Penna, A., 2010. Monitoring toxic microalgae Ostreopsis(dinoflagellate) species in coastal waters of the Mediterranean Sea usingmolecular PCR-based assay combined with light microscopy. Mar. Pollut. Bull.60, 1074–1084.
Bellocci, M., Ronzitti, G., Milandri, A., Melchiorre, N., Grillo, C., Poletti, R., Yasumoto,T., Rossini, G.P., 2008. A cytolytic assay for the measurement of palytoxin basedon a cultured monolayer cell line. Anal. Biochem. 374, 48–55.
Ciminiello, P., Dell’Aversano, C., Fattorusso, E., Forino, M., Magno, G.S., Tartaglione,L., Grillo, C., Melchiorre, N., 2006. The Genoa 2005 outbreak. Determination ofputative palytoxin in Mediterranean Ostreopsis ovata by a new liquidchromatography–tandem mass spectrometry method. Anal. Chem. 78, 6153–6159.
Ciminiello, P., Dell’Aversano, C., Fattorusso, E., Forino, M., Tartaglione, L., Grillo, C.,Melchiorre, N., 2008. Putative palytoxin and its new analogue, ovatoxin-a, inOstreopsis ovata collected along the Ligurian coasts during the 2006 toxicoutbreak. J. Am. Soc. Mass Spectrom. 19, 111–120.
Ciminiello, P., Dell’Aversano, C., Dello Iacovo, E., Fattorusso, E., Forino, M., Grauso, L.,Tartaglione, L., Guerrini, F., Pistocchi, R., 2010. Complex palytoxin-like profile ofOstreopsis ovata. Identification of four new ovatoxins by high-resolution liquidchromatography/mass spectrometry. Rapid Commun. Mass Spectrom. 24,2735–2744.
Ciminiello, P., Dell’Aversano, C., Dello Iacovo, E., Fattorusso, E., Forino, M.,Tartaglione, L., 2011. LC–MS of palytoxin and its analogues: state of the artand future perspectives. Toxicon 57, 376–389.
Cucchiari, E., Guerrini, F., Penna, A., Totti, C., Pistocchi, R., 2008. Effect of salinity,temperature, organic and inorganic nutrients on growth of cultured Fibrocapsajaponica (Raphidophyceae) from the northern Adriatic Sea. Harmful Algae 7,405–414.
Deeds, J.R., Schwartz, M.D., 2010. Human risk associated with palytoxin exposure.Toxicon 56, 150–162.
Faust, M.A., Morton, S.L., Quod, J.P., 1996. Further SEM study of marinedinoflagellates: the genus Ostreopsis (Dinophyceae). J. Phycol. 32, 1053–1065.
Gallitelli, M., Ungaro, N., Addante, L.M., Silver, N.G., Sabba, C., 2005. Respiratoryillness as a reaction to tropical algal blooms occurring in a temperate climate. J.Am. Med. Assoc. 293, 2599–2600.
Glibert, P.M., Burkholder, J.M., 2006. The complex relationships between increasesin fertilization of the earth, coastal eutrophication and proliferation of harmfulalgal blooms. In: Graneli, E., Turner, J.T. (Eds.), Ecology of Harmful Algae.Springer, Berlin, Heidelberg, pp. 341–354.
Glibert, P.M., Allen, J.I., Bouwman, A.F., Brown, C.W., Flynn, K.J., Lewitus, A.J.,Madden, C.J., 2010. Modeling of HABs and eutrophication status, advances,challenges. J. Mar. Syst. 83, 262–275.
Gómez, F., Claustre, H., 2003. The genus Asterodinium (Dinophyceae) as a possiblebiological indicator of warming in the western Mediterranean Sea. J. Mar. Biol.Assoc. UK 83, 173–174.
Granéli, E., Vidyarathna, N.K., Funari, E., Cumaranatunga, P.R.T., Scenati, R., 2011.Can increases in temperature stimulate blooms of the toxic benthicdinoflagellate Ostreopsis ovata? Harmful Algae 10, 165–172.
Guerrini, F., Pezzolesi, L., Feller, A., Riccardi, M., Ciminiello, P., Dell’Aversano, C.,Tartaglione, L., Dello Iacovo, E., Fattorusso, E., Forino, M., Pistocchi, R., 2010.Comparative growth and toxin profile of cultured Ostreopsis ovata from theTyrrhenian and Adriatic Seas. Toxicon 55, 211–220.
Hallegraeff, G.M., 2010. Ocean climate change, phytoplankton communityresponses, and harmful algal blooms: a formidable predictive challenge. J.Phycol. 46, 220–235.
Hasle, G.R., 1978. The inverted microscope method. In: Sournia, A. (Ed.),Phytoplankton Manual. UNESCO, Paris, pp. 88–96.
Heisler, J., Glibert, P.M., Burkholder, J.M., Anderson, D.M., Cochlan, W., Dennison,W.C., Dortch, Q., Gobler, C.J., Heil, C.A., Humphries, E., Lewitus, A., Magnien, R.,Marshall, H.G., Sellner, K., Stockwell, D.A., Stoecker, D.K., Suddleson, M., 2008.Eutrophication and harmful algal blooms: a scientific consensus. Harmful Algae8, 3–13.
Jin, Q., Dong, S., 2003. Comparative studies on the allelopathic effects of twodifferent strains of Ulva pertusa on Heterosigma akashiwo and Alexandriumtamarense. J. Exp. Mar. Biol. Ecol. 293, 41–55.
Lejeusne, C., Chevaldonne, P., Pergent-Martini, C., Boudouresque, C.F., Perez, T.,2010. Climate change effects on a miniature ocean: the highly diverse, highlyimpacted Mediterranean Sea. Trends Ecol. Evol. 25, 250–260.
Louzao, M.C., Espiña, B., Cagide, E., Ares, I.R., Alfonso, A., Vieytes, M.R., Botana, L.M.,2010. Cytotoxic effect of palytoxin on mussel. Toxicon 56, 842–847.

S. Accoroni et al. / Marine Pollution Bulletin 62 (2011) 2512–2519 2519
Mangialajo, L., Bertolotto, R., Cattaneo-Vietti, R., Chiantore, M., Grillo, C., Lemee, R.,Melchiorre, N., Moretto, P., Povero, P., Ruggieri, N., 2008. The toxic benthicdinoflagellate Ostreopsis ovata: quantification of proliferation along thecoastline of Genoa, Italy. Mar. Pollut. Bull. 56, 1209–1214.
Mangialajo, L., Ganzin, N., Accoroni, S., Asnaghi, V., Blanfuné, A., Cabrini, M.,Cattaneo-Vietti, R., Chavanon, F., Chiantore, M., Cohu, S., Costa, E., Fornasaro, D.,Grossel, H., Marco-Miralles, F., Masó, M., Reñé, A., Rossi, A.M., Sala, M.M.,Thibaut, T., Totti, C., Vila, M., Lemée, R., 2011. Trends in Ostreopsis proliferationalong the Northern Mediterranean coasts. Toxicon 57, 408–420.
Marini, M., Fornasiero, P., Artegiani, A., 2002. Variations of hydrochemical featuresin the coastal waters of Monte Conero: 1982–1990, P.S.Z.N.. Mar. Ecol. 23(Suppl. 1), 258–271.
Marini, M., Jones, B.H., Campanelli, A., Grilli, F., Lee, C.M., 2008. Seasonal variabilityand Po River plume influence on biochemical properties along western Adriaticcoast. J. Geophys. Res. 113, C05S90. doi:10.1029/2007JC004370.
Martinelli, E., Morbidoni, M., Farroni, L., Mengarelli, C., Mariottini, M., Verna, M.,2009. Inchiesta epidemiologica per sospetta epidemia da aerosol marino.L’episodio del 23 agosto 2009 ad Ancona, ARS 123, 36–39. Available from:<www.arpa.marche.it/doc/Pdf/epidem/epid_ostreopsis.pdf/>.
Monti, M., Minocci, M., Beran, A., Iveša, L., 2007. First record of Ostreopsis cfr. ovataon macroalgae in the Northern Adriatic Sea. Mar. Pollut. Bull. 54, 598–601.
Onuma, Y., Satake, M., Ukena, T., Roux, J., Chanteau, S., Rasolofonirina, N.,Ratsimaloto, M., Naoki, H., Yasumoto, T., 1999. Identification of putativepalytoxin as the cause of clupeotoxism. Toxicon 37, 55–65.
Parsons, M.L., Preskitt, L.B., 2007. A survey of epiphytic dinoflagellates from thecoastal waters of the island of Hawai’i. Harmful Algae 6, 658–669.
Penna, A., Vila, M., Fraga, S., Giacobbe, M.G., Andreoni, F., Riobó, P., Vernesi, C., 2005.Characterization of Ostreopsis and Coolia (Dinophyceae) isolates in the westernMediterranean Sea based on morphology, toxicity and internal transcribedspacer 5.8s rDNA sequences. J. Phycol. 41, 212–225.
Penna, A., Bertozzini, E., Battocchi, C., Galluzzi, L., Giacobbe, M.G., Vila, M., Garces, E.,Lugliè, A., Magnani, M., 2007. Monitoring of HAB species in the MediterraneanSea through molecular methods. J. Plankton Res. 29, 19–38.
Penna, A., Fraga, S., Battocchi, C., Casabianca, S., Giacobbe, M.G., Riobó, P., Vernesi, C.,2010. A phylogeographical study of the toxic benthic dinoflagellate genusOstreopsis Schmidt. J. Biogeogr. 37, 830–841.
Pezzolesi, L., Guerrini, F., Ciminiello, P., Dell’Aversano, C., Dello Iacovo, E., Fattorusso,E., Forino, M., Tartaglione, L., Pistocchi, R., in press. Influence of temperature andsalinity on Ostreopsis cf. ovata growth and evaluation of toxin content throughHR LC–MS and biological assays. Water Res.
Rhodes, L., 2011. World-wide occurrence of the toxic dinoflagellate genus OstreopsisSchmidt. Toxicon 57, 400–407.
Rhodes, L.L., Munday, R., Briggs, L.R., Holland, P.T., Miles, C.O., Loader, J., Jensen, D.,Cooney, J., 2006. Ostreopsis siamensis and palytoxin-related compounds in New
Zealand: a risk to human health? In: Twelfth International Conference onHarmful Algae, Copenhagen, Denmark, p. 41.
Rossi, R., Castellano, V., Scalco, E., Serpe, L., Zingone, A., Soprano, V., 2010. Newpalytoxin-like molecules in Mediterranean Ostreopsis cf. ovata (dinoflagellates)and in Palythoa tuberculosa detected by liquid chromatography–electrosprayionization time-of-flight mass spectrometry. Toxicon 56, 1381–1387.
Selina, M.S., Orlova, T.Y., 2010. First occurrence of the genus Ostreopsis(Dinophyceae) in the Sea of Japan. Bot. Mar. 53, 243–249.
Shears, N.T., Ross, P.M., 2009. Blooms of benthic dinoflagellates of the genusOstreopsis: an increasing and ecologically important phenomenon on temperatereefs in New Zealand and worldwide. Harmful Algae 8, 916–925.
Shears, N.T., Ross, P.M., 2010. Toxic cascades: multiple anthropogenic stressors havecomplex and unanticipated interactive effects on temperate reefs. Ecol. Lett. 13,1149–1159.
Simoni, F., Gaddi, A., Di Paolo, C., Lepri, L., 2003. Harmful epiphytic dinoflagellate onTyrrhenian Sea reefs. Harmful Algae News 24, 13–14.
Steidinger, K.A., Tangen, K., 1997. Dinoflagellates. In: Thomas, C.R. (Ed.), IdentifyingMarine Phytoplankton. Academic Press, San Diego, pp. 387–584.
Strickland, J.D.H., Parsons, T.R., 1968. A practical handbook of seawater analysis. J.Fish. Res. Board Can. 167, 310.
Tindall, D.R., Morton, S.L., 1998. Community dynamics and physiology of epiphytic/benthic dinoflagellates associated with ciguatera. In: Anderson, D.M., Cembella,A.D., Hallegraeff, G.M. (Eds.), Physiological Ecology of Harmful Algal Blooms,NATO ASI Series, Series G, Ecological Sciences, vol. 41. Springer-Verlag, Berlin,pp. 291–313.
Tognetto, L., Bellato, S., Moro, I., Andreoli, C., 1995. Occurrence of Ostreopsis ovata(Dinophyceae) in the Tyrrhenian Sea during summer 1994. Bot. Mar. 38, 291–295.
Totti, C., Accoroni, S., Cerino, F., Cucchiari, E., Romagnoli, T., 2010. Ostreopsisovata bloom along the Conero Riviera (northern Adriatic Sea): relation-ships with environmental conditions and substrata. Harmful Algae 9, 233–239.
Turki, S., Harzallah, A., Sammari, C., 2006. Occurrence of harmful dinoflagellates intwo different Tunisian ecosystems: the lake of Bizerte and the gulf of Gabes.Cah. Biol. Mar. 47, 253–259.
Van Dolah, F.M., 2000. Marine algal toxins: origins, health effects, and theirincreased occurrence. Environ. Health Perspect. 108, 133–141.
Vila, M., Garcés, E., Masó, M., 2001. Potentially toxic epiphytic dinoflagellateassemblages on macroalgae in the NW Mediterranean. Aquat. Microb. Ecol. 26,51–60.
Zeldis, J.R., Walters, R.A., Greig, M.J.N., Image, K., 2004. Circulation over thenortheastern New Zealand continental slope, shelf and adjacent Hauraki Gulf,during spring and summer. Cont. Shelf Res. 24, 543–561.




![9Z[RS SR_ Z_ T]Rddc``^ gR]ZU cf]Vd](https://static.fdokumen.com/doc/165x107/63326f1e3108fad7760e9f01/9zrs-sr-z-trddc-grzu-cfvd.jpg)

![AcVk Cf]V Z_ >RYRcRdYecR aRceZVd cV]ZVgVU](https://static.fdokumen.com/doc/165x107/6328451de491bcb36c0baf97/acvk-cfv-z-ryrcrdyecr-arcezvd-cvzvgvu.jpg)


![t] x| f} + of] hgf -cf=j](https://static.fdokumen.com/doc/165x107/631922cab41f9c8c6e099861/t-x-f-of-hgf-cfj.jpg)










