
Temporal and spatial zonation of the demersal trawl fauna of the central Great Barrier Reef
Upload
departmentconservationnewzealandCategory
view
4download
0
Polar Biol
DOI 10.1007/s00300-006-0192-8ORIGINAL PAPER
Origin, age, sex and breeding status of wandering albatrosses (Diomedea exulans), northern (Macronectes halli) and southern giant petrels (Macronectes giganteus) attending demersal longliners in Falkland Islands and Scotia Ridge waters, 2001–2005
Helen Otley · Tim Reid · Richard Phillips · Andy Wood · Ben Phalan · Isaac Forster
Received: 10 April 2006 / Revised: 17 July 2006 / Accepted: 17 July 2006© Springer-Verlag 2006
Abstract A total of 547 sightings of 291 banded wan-dering albatrosses Diomedea exulans and 21 sightingsof 14 banded giant petrels Macronectes spp. were madefrom toothWsh longliners operating on the southernPatagonian Shelf during 2001–2005. This included 25%of the wandering albatrosses with Darvic bands thatbred at Bird Island (South Georgia) during this period.Thirteen of the northern Macronectes halli and south-ern giant petrels Macronectesgiganteus had beenbanded at South Georgia, and there was one sightingof a southern giant petrel from Argentina. Male andfemale wandering albatrosses of all age classes exceptyoung birds (<15 years old) were equally likely toattend longline vessels. Most sightings of all age classeswere made during the incubation period and fewestduring the brood period. Eighty-six percent of birdssighted had bred at least once before, with half cur-rently breeding and half on sabbatical (i.e. betweenbreeding attempts). Almost half of the wandering alba-trosses were sighted on more than one occasion. Thedata conWrms that the southern Patagonian shelf is an
important foraging area for wandering albatrosses andnorthern and southern giant petrels, and that someindividuals show consistent associations in multipleyears with longline vessels Wshing in the region.
Introduction
South Georgia is home to a variety of albatross, petreland prion species that have been the subject of detailedlong-term studies at Bird Island, oV its north-westerlypoint, since the late 1950s (Tickell 1968; Croxall et al.1998). Several of these are threatened; the populationsof wandering Diomedea exulans, black-browed Tha-lassarche melanophrys and grey-head albatrosses T.chrysostoma at South Georgia have all declined by30% since 1984 (Poncet et al. 2006). Systematic band-ing of albatrosses at Bird Island was instituted in theearly 1970s, with colour banding with numeric oralphanumeric plastic Darvic bands of all wanderingalbatross chicks and adults since 1979 and 1980, respec-tively, in a study area (Wanderer Ridge) that holdsapproximately 100 pairs per year (Croxall et al. 1999).In addition, intermittently from summer 1978/1979onwards, northern and southern giant petrel chicks andadults have been banded with British Trust for Orni-thology (BTO) metal bands, and breeding adults alsowith Darvic bands. Starting in the 2002/2003 summer,all adult birds in a study area that holds approximately150 and 250 pairs of southern and northern giantpetrels, respectively, have been Wtted with BTO andDarvic bands.
Since there is little evidence for reduced breedingsuccess at colonies, it appears that reduced survival of
H. Otley (&)Falkland Islands Fisheries Department, Stanley, FIQQ 1ZZ, Falkland Islands, UKe-mail: [email protected]
T. ReidFalklands Conservation, PO Box 26, Stanley, FIQQ 1ZZ, Falkland Islands, UK
R. Phillips · A. Wood · B. Phalan · I. ForsterBritish Antarctic Survey, Natural Environment Research Council, High Cross, Madingley Road, Cambridge, CB3 0ET, UK
123

Polar Biol
immatures and adults when foraging at sea, particu-larly incidental mortality associated with longline andtrawl Wsheries, has caused the recent populationdeclines (Croxall and Gales 1998; Croxall et al. 1998).Foraging areas of wandering albatross and northernand southern giant petrels have been determined bysightings of banded birds at sea and tracking using sat-ellite transmitters (Prince et al. 1998; Croxall et al.1999; González-Solis et al. 2000; Xavier et al. 2004).Satellite tracking, predominantly of breeding birdsindicates that Wandering albatrosses use the Patago-nian shelf-edge area year round, whereas northern andsouthern giant petrels mainly visit the region duringincubation and the post-breeding period (Croxall andWood 2002). Much less is known about the foragingareas of pre-breeders and non-breeding birds of bothspecies.
In Falkland Island waters, demersal longliners Wsh forPatagonian toothWsh Dissostichus eleginoides on theedge of the Patagonian shelf and consequently, with onboard scientiWc observers, these vessels provide aunique opportunity to observe seabirds at sea, particu-larly banded individuals (Croxall et al. 1999). We reporton sightings of wandering albatross, northern and south-ern giant petrels banded on Bird Island and also of asouthern giant petrel banded in southern Argentina, attoothWsh longliners during 2001–2005 in the FalklandIslands (51°S 57°W) and at a seamount on the ScotiaRidge at 54°S, 52°W (hereafter referred to ‘ScotiaRidge’), located half way between the Falkland Islandsand South Georgia. This study extends the previousanalysis of at-sea sightings in the area (Croxall et al.1999), and complements the studies of behaviouralinteractions of wandering albatross and also royal alba-tross Diomedea epomophora with Wsheries in the south-western Atlantic and eastern New Zealand waters,respectively (Xavier et al. 2004; Waugh et al. 2005).
Methods
Wandering albatross
Wandering albatrosses recruit into the breeding popu-lation at a modal age of 9–10 years, and thereafter, ifsuccessful, breeding is usually biennial, with attemptsmade in consecutive years only when eggs or chicks arelost prior to June (Tickell 2000). All breeding attemptson the island by Darvic banded birds are recorded aspart of the annual monitoring programme and in addi-tion, from early January to mid-March, a check is madefor non-breeding birds with Darvics in the WandererRidge study area (approximately 100 breeding pairs) at
least once a week, and less frequently in breeding areaselsewhere on the island (Croxall et al. 1999). A total of1,009 birds with Darvics were recorded making breed-ing attempts at Bird Island during the 2002–2005breeding seasons.
Giant petrels
At South Georgia, northern giant petrels lay in lateSeptember to early October, and southern giant petrelsin early to mid-November (Hunter 1984). Chicks hatch60 days later, and Xedge after a further 105–132 days.Given the diVerence in laying dates, northern giantpetrel chicks Xedge in mid to late March, whereassouthern giant petrel chicks Xedge in late April to mid-May. All breeding attempts by banded birds arerecorded as part of the annual monitoring programme,but the presence of non-breeding banded birds is notrecorded.
Banding of southern giant petrels also occurs inArgentina, with Centro Nacional Patagónico, ConicetArgentina and Aves Argentinas implementing a band-ing program of Xedglings at two adjacent sites in theChubut province of Argentina in 2004. In March, 263females and 233 males were banded with BTO bandsand yellow Darvic bands at Islas Arce (45°00�S,65°50�W) and Gran Robredo (45°08�S, 66°03�W).
Longlining for toothWsh
Exploratory longline Wshing commenced in the Falk-land Islands in 1992 and became commercial in 1994with the granting of two licences to a locally registeredcompany. In the Falkland Islands, two longliners arelicenced to Wsh in waters deeper than 600 m and one tothree other longliners also registered to the FalklandIslands obtain a licence in some years to Wsh on thehigh seas, often concentrating eVorts at Scotia Ridge.Scientists from the Falkland Islands Fisheries Depart-ment (FIFD) and Falklands Conservation (FC)observe Wshing activities aboard longliners in Falklandwaters and FIFD observers may also join FalklandIslands registered longliners working on the high seas.
During 2001–2004, licensed longliners caught any-where between 100 and 12,000 kg of whole toothWshper day (Laptikhovsky and Brickle 2005). For every100 kg of whole toothWsh processed, 37 kg of heads,tails and other oVal is discarded. There is also substan-tial bycatch, with a mean § SD of 185 § 184 kg of Mac-rourus spp., 36 § 40 kg of Antimora rostrata and153 § 180 kg of large rays (Raja and Bathyraja spp.),discarded whole per day (Otley 2005). Based on dis-card ingestion rates, the soft-Xeshed A. rostrata is the
123

Polar Biol
species preferred by seabirds (HO and TR, personalobservation). Bait that returns on hooks during haulingis also discarded but the amount is not estimated.
Field observations
During November 2001–February 2005 (51 months),12 observers from FIFD and FC made 58 one- to ten-week trips during all months aboard longliners Wshingfor Patagonian toothWsh. Only two observers were everaboard longline vessels at any one time. All trips weretaken in Falkland Island waters, except for two 3-weektrips in January and February 2004 to Scotia Ridge.
On a number of occasions during line hauling, sea-bird species abundance was recorded in a 500 £ 500 m2
area from the hauling bay on the bow to 500 m asternand 250 m out of the port and starboard sides (Reidand Sullivan 2004). Where the total number of any spe-cies was less than 20–30, birds were counted individu-ally. If more abundant, estimates were to §5individuals (30–100 individuals present) and to §10individuals (>100 individuals present). Northern andsouthern giant petrels were not diVerentiated.
Detailed observations of attendant birds were madeusing binoculars during hauling activities, often fromabove the discard chute. Not all Darvic bands could beread because of problems associated with rough seastate, poor line of sight from vantage points on somevessels and bird movements. A reading was obtainedfor the majority of wandering albatrosses but onlyapproximately half of the giant petrels with Darvicsbecause of the smaller lettering used for the smallerspecies. Thus, the number of birds identiWed on eachcruise is not a reliable indicator of the relative numbersof wandering albatrosses and giant petrels or of bandedand unbanded birds.
Six of 12 observers made sightings of banded sea-birds, with considerable variation among observers:four individuals made 7–16 records, one (HO) made241 and another (TR) made 263 records. Records atsea were matched with information on sex, age andprevious and current breeding status from British Ant-arctic Survey (BAS) databases.
Data analysis
Sightings were stratiWed into three regions: (1) ScotiaRidge, (2) Falklands Zone and (3) Burdwood BankZone (Fig. 1). Wandering albatross sightings were alsostratiWed temporally according to the four diVerentstages of the breeding cycle: (1) pre-laying (1 Novem-ber–15 December), (2) incubation (16 December–20March), (3) brooding (21 March–20 April) and (4)
post-brooding (21 April–31 October) (Tickell 1968 andBAS, unpublished data).
Prior to statistical comparisons, tests for normalityand homogeneity of variance were conducted, and datatransformed where possible in order to use parametrictests.
Results
Seabird abundance
Direct counts showed signiWcant diVerences betweenbreeding periods in the abundance of wandering alba-trosses at longliners (F3,532 = 32.65, P < 0.001, usingmaximum number present per haul). This was a conse-quence of the relatively low abundance during post-brooding (mean of 4.8 birds) compared to other peri-ods (means of 9.7–13.6 birds; Table 1). Giant petrelnumbers did not vary signiWcantly between breedingperiods (F3,780 = 2.09, P < 0.101, using maximum num-ber present per haul), with means of 86–114 birds pres-ent throughout the study (Table 1).
Wandering albatross sex, age and status
A total of 547 records of 291 colour-banded wanderingalbatrosses were made. This included 25% of the 1,009birds with Darvics that bred at Bird Island in the 2002–2005 breeding seasons. There were a further 11 sight-ings where the recorded alphanumeric Darvic codecould not be matched to Bird Island records.
Banded birds of all ages were seen (Table 2). Theyoungest individual sighted was 2 years of age and theeldest was 45 years, with the majority (75%) aged 10–29 years, and few older than 30 years.
One hundred and forty two individuals sighted atlongline vessels were female, 126 were males and 23were of unknown sex (Table 2). The overall sex ratiowas not signiWcantly diVerent from 1:1 (normal approx-imation to the binomial test, Z = 0.98, P = 0.36). Con-sidering only birds of known age, the proportion offemales was signiWcantly greater amongst birds aged14 years or less, than in older age classes (69.6% of 56birds, cf. 47.8% of 186 birds; �2 (corrected forcontinuity) = 7.4, df = 1, P < 0.01).
Two hundred and Wfty-one birds sighted (86%) hadbred at least once before (Table 2). These birds were aslikely to be breeding (226 observations) as on sabbati-cal (229 observations) at the time of sighting (Table 3).Twenty-seven birds seen at sea (9% of the total) hadbeen sighted at Bird Island but never recorded asbreeding, and 13 birds (4%) had not been sighted since
123

Polar Biol
ringing (Table 2). Three of these birds were ringed atan unknown age as adults, eight birds were less than10 years of age (9 years being the modal age of Wrstbreeding) and hence may not have started breeding,and two birds were aged 20–29 at the time of sighting.
Six birds aged 5–8 years and 44 birds aged 9–45 years (17%) had not been sighted at Bird Island formore than 4 (maximum 28) years. Thirty-six of thesebirds (72%) were only sighted once at sea. Seventeenbirds had previously bred at Bird Island, 20 birds hadbeen sighted but not bred there and 13 birds, aged 5–27 years of age, had not been sighted at Bird Islandsince Xedging.
Timing and location of sightings
The number of sightings/day varied signiWcantlybetween the breeding periods (�2 = 415.09, df = 3,P < 0.001). Most sightings and the highest daily sightingrate of Darvic banded wandering albatrosses took placeduring the incubation period (December 16–March 20),with the least number of sightings made during brood-ing (Table 3). There was no signiWcant diVerence in theproportion of sabbatical and current breeders sightedduring each period (�2 = 5.63, df = 3, P > 0.05).
Birds were sighted throughout the shelf-slope, butmostly in regions with water depths of 1,000–1,500 m(Fig. 1). A total of 313, 101 and 133 sightings were madein Burdwood Bank, Falklands and Scotia Ridge Zones,respectively (Table 3; Fig. 1). Despite the high numberof sightings (many of them of the same birds on severaloccasions) in Falkland Island waters, only ten birdswere sighted in both Burdwood Bank and FalklandZones. Fifteen of the 45 birds seen at Scotia Ridge werealso seen in Falklands and/or Burdwood Bank Zones.
Association with Wshing vessel
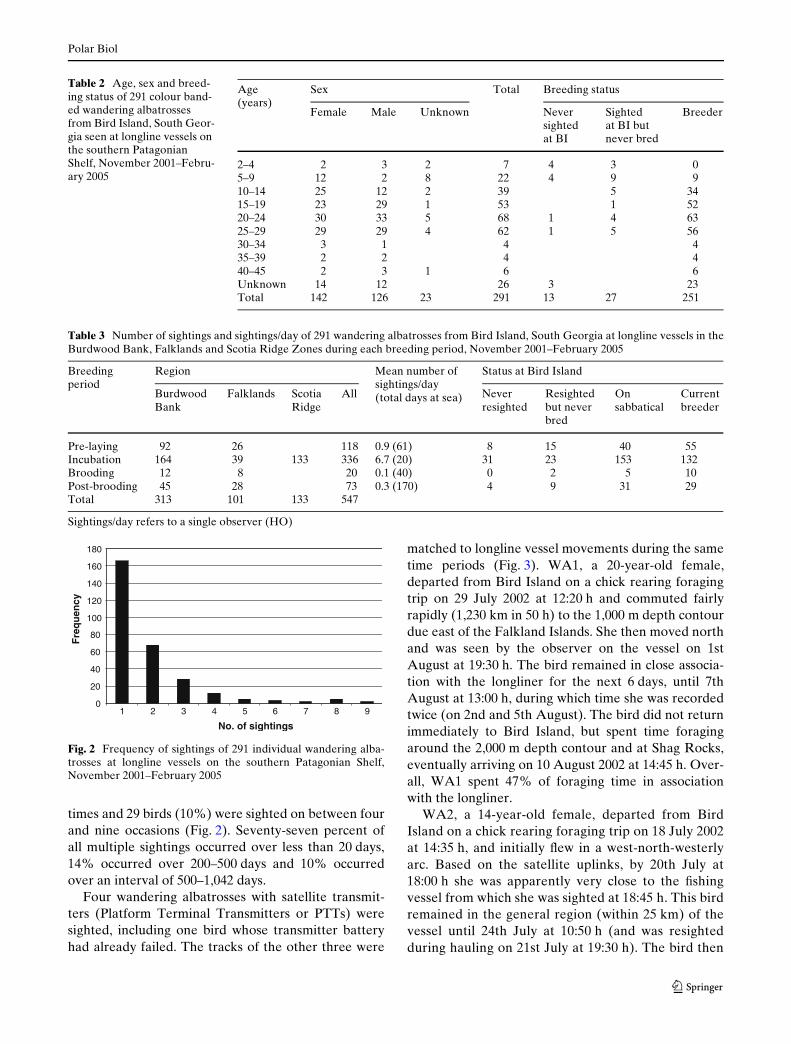
One hundred and sixty-six individuals (57%) weresighted once, 90 birds (33%) were sighted two or three
Fig. 1 Location of the 547 sightings of wandering albatrosses atlongline vessels and areas of the Falklands, Scotia Ridges andBurdwood Bank Zones on the southern Patagonian Shelf,November 2001–February 2005. Shaded area around the Falk-
lands Islands is the extent of the Falklands Inner/Outer Conser-vation Zone and the shaded area around South Georgia is theSouth Georgia Maritime Zone
Table 1 Mean number and standard deviation of wanderingalbatrosses and giant petrels sighted at longline vessels on thesouthern Patagonian Shelf during each wandering albatrossbreeding period, November 2001–February 2005
n = number of surveys
Wandering albatrossbreeding period
Wandering albatross Giant petrel
Mean SD n Mean SD n
Pre-laying 9.7 11.2 84 86 93 135Incubation 13.6 13.7 37 114 126 69Brooding 11.2 9.5 40 91 64 32Post-brooding 4.8 6.1 372 112 123 545
123
Polar Biol
times and 29 birds (10%) were sighted on between fourand nine occasions (Fig. 2). Seventy-seven percent ofall multiple sightings occurred over less than 20 days,14% occurred over 200–500 days and 10% occurredover an interval of 500–1,042 days.
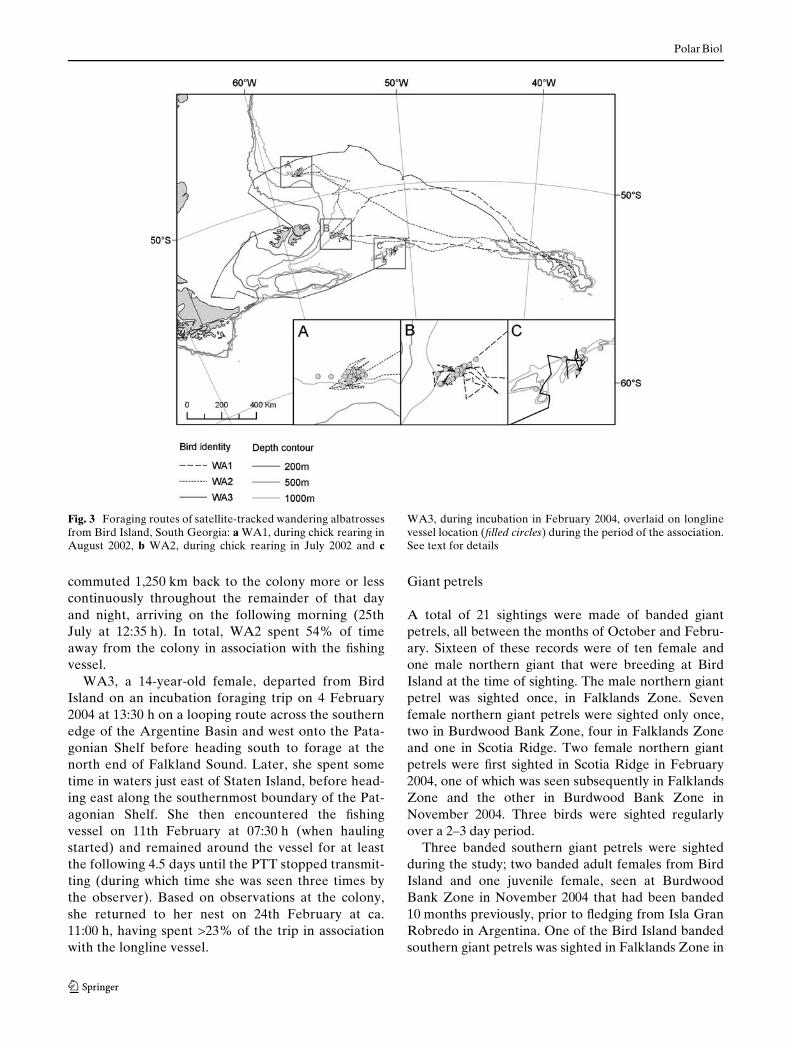
Four wandering albatrosses with satellite transmit-ters (Platform Terminal Transmitters or PTTs) weresighted, including one bird whose transmitter batteryhad already failed. The tracks of the other three were
matched to longline vessel movements during the sametime periods (Fig. 3). WA1, a 20-year-old female,departed from Bird Island on a chick rearing foragingtrip on 29 July 2002 at 12:20 h and commuted fairlyrapidly (1,230 km in 50 h) to the 1,000 m depth contourdue east of the Falkland Islands. She then moved northand was seen by the observer on the vessel on 1stAugust at 19:30 h. The bird remained in close associa-tion with the longliner for the next 6 days, until 7thAugust at 13:00 h, during which time she was recordedtwice (on 2nd and 5th August). The bird did not returnimmediately to Bird Island, but spent time foragingaround the 2,000 m depth contour and at Shag Rocks,eventually arriving on 10 August 2002 at 14:45 h. Over-all, WA1 spent 47% of foraging time in associationwith the longliner.
WA2, a 14-year-old female, departed from BirdIsland on a chick rearing foraging trip on 18 July 2002at 14:35 h, and initially Xew in a west-north-westerlyarc. Based on the satellite uplinks, by 20th July at18:00 h she was apparently very close to the Wshingvessel from which she was sighted at 18:45 h. This birdremained in the general region (within 25 km) of thevessel until 24th July at 10:50 h (and was resightedduring hauling on 21st July at 19:30 h). The bird then
Table 2 Age, sex and breed-ing status of 291 colour band-ed wandering albatrosses from Bird Island, South Geor-gia seen at longline vessels on the southern Patagonian Shelf, November 2001–Febru-ary 2005
Age (years)
Sex Total Breeding status
Female Male Unknown Never sighted at BI
Sighted at BI but never bred
Breeder
2–4 2 3 2 7 4 3 05–9 12 2 8 22 4 9 910–14 25 12 2 39 5 3415–19 23 29 1 53 1 5220–24 30 33 5 68 1 4 6325–29 29 29 4 62 1 5 5630–34 3 1 4 435–39 2 2 4 440–45 2 3 1 6 6Unknown 14 12 26 3 23Total 142 126 23 291 13 27 251
Table 3 Number of sightings and sightings/day of 291 wandering albatrosses from Bird Island, South Georgia at longline vessels in theBurdwood Bank, Falklands and Scotia Ridge Zones during each breeding period, November 2001–February 2005
Sightings/day refers to a single observer (HO)
Breeding period
Region Mean number of sightings/day (total days at sea)
Status at Bird Island
Burdwood Bank
Falklands Scotia Ridge
All Never resighted
Resighted but never bred
On sabbatical
Current breeder
Pre-laying 92 26 118 0.9 (61) 8 15 40 55Incubation 164 39 133 336 6.7 (20) 31 23 153 132Brooding 12 8 20 0.1 (40) 0 2 5 10Post-brooding 45 28 73 0.3 (170) 4 9 31 29Total 313 101 133 547
Fig. 2 Frequency of sightings of 291 individual wandering alba-trosses at longline vessels on the southern Patagonian Shelf,November 2001–February 2005
0
20
40
60
80
100
120
140
160
180
54321 8 976
Fre
qu
ency
No. of sightings
123
Polar Biol
commuted 1,250 km back to the colony more or lesscontinuously throughout the remainder of that dayand night, arriving on the following morning (25thJuly at 12:35 h). In total, WA2 spent 54% of timeaway from the colony in association with the Wshingvessel.
WA3, a 14-year-old female, departed from BirdIsland on an incubation foraging trip on 4 February2004 at 13:30 h on a looping route across the southernedge of the Argentine Basin and west onto the Pata-gonian Shelf before heading south to forage at thenorth end of Falkland Sound. Later, she spent sometime in waters just east of Staten Island, before head-ing east along the southernmost boundary of the Pat-agonian Shelf. She then encountered the Wshingvessel on 11th February at 07:30 h (when haulingstarted) and remained around the vessel for at leastthe following 4.5 days until the PTT stopped transmit-ting (during which time she was seen three times bythe observer). Based on observations at the colony,she returned to her nest on 24th February at ca.11:00 h, having spent >23% of the trip in associationwith the longline vessel.
Giant petrels
A total of 21 sightings were made of banded giantpetrels, all between the months of October and Febru-ary. Sixteen of these records were of ten female andone male northern giant that were breeding at BirdIsland at the time of sighting. The male northern giantpetrel was sighted once, in Falklands Zone. Sevenfemale northern giant petrels were sighted only once,two in Burdwood Bank Zone, four in Falklands Zoneand one in Scotia Ridge. Two female northern giantpetrels were Wrst sighted in Scotia Ridge in February2004, one of which was seen subsequently in FalklandsZone and the other in Burdwood Bank Zone inNovember 2004. Three birds were sighted regularlyover a 2–3 day period.
Three banded southern giant petrels were sightedduring the study; two banded adult females from BirdIsland and one juvenile female, seen at BurdwoodBank Zone in November 2004 that had been banded10 months previously, prior to Xedging from Isla GranRobredo in Argentina. One of the Bird Island bandedsouthern giant petrels was sighted in Falklands Zone in
Fig. 3 Foraging routes of satellite-tracked wandering albatrossesfrom Bird Island, South Georgia: a WA1, during chick rearing inAugust 2002, b WA2, during chick rearing in July 2002 and c
WA3, during incubation in February 2004, overlaid on longlinevessel location (Wlled circles) during the period of the association.See text for details
123

Polar Biol
November 2003 and the other was sighted in Burd-wood Bank Zone in January 2002 and then on twooccasions over 3 days in October 2002. Neither birdwas breeding at Bird Island at the time of sighting.Banded southern giant petrels were not sighted at Sco-tia Ridge.
Discussion
This at-sea study recorded 547 sightings of 291 wander-ing albatross and 21 sightings of 14 northern and south-ern giant petrels from toothWsh longliners Wshing in thesouthern Patagonian Shelf, extending the spatial cover-age beyond that of the earlier study by Croxall et al.(1999) which was based primarily on sightings at theBurdwood Bank.
Wandering albatross sex, age and status
Large numbers of males and females of all age classesof wandering albatross banded at Bird Island attendedlongline vessels in this region. This agrees with resultsof satellite tracking of breeding birds from Bird Islandindicating that both sexes use the southern PatagonianShelf and shelf-slope to considerable extents (Princeet al. 1998; Croxall and Wood 2002; Xavier et al. 2003a,2004). The signiWcantly higher proportion of females inthe youngest age classes sighted (<15 years) may reXectthe longer time spent at the colony by males comparedwith females in the years from Wrst return to Wrstbreeding attempt (Pickering 1989). As females willreadily compete for oVal and discards even when thelarger males are present, both sexes are vulnerable toincidental mortality. In contrast, in most other alba-trosses and petrels, longline mortality in toothWsh Wsh-eries is biased towards males (Gales et al. 1998; Ryanand Boix-Hinzen 1999).
Croxall et al. (1999) suggested, and this study con-Wrms over a wider spatial and temporal scale, that ahigh proportion of birds of all known age classes,except young birds (<10 years of age), feed on thesouthern Patagonian Shelf. The vast majority of adultbirds were established breeders, with half of thesebreeding at the time of sighting and half on sabbaticalyears. The ratio of current to sabbatical breeders wasmore or less constant across all sampling periods.Although breeding birds from South Georgia havebeen tracked extensively, information on the move-ments of adults on sabbatical is restricted primarily tothat gleaned from ringing recoveries (Prince et al.1998). The importance of the southern PatagonianShelf to sabbatical breeders throughout the year has
not been shown previously, and is of considerable con-servation interest given the high Wshing eVort by a vari-ety of methods in the region.
A surprisingly high percentage of sightings in thisstudy (17%) were of individuals not seen at Bird Islandfor 5 years or more (also see Croxall et al. 1999). Thus,as Croxall et al. (1999) suggests, factors other thanproximity to the breeding colony are involved in thedecision of non-breeders to remain in, or return toSouth Atlantic waters. Some wandering albatrossessighted in our study had not been seen since Xedging(up to 28 years previously) on Bird Island. A propor-tion of these records are likely to be misread bands;43% of birds were only sighted once and of thosesighted on multiple occasions, only four were seen overintervals of more than 5 days. However, this is unlikelyto account for all these unexpected records: care wastaken to ensure that combinations were valid and notjust single character diVerences (particularly betweenletters and digits of similar shape) from Darvic codes ofbirds sighted at Bird Island in recent years.
Wandering albatross colonies elsewhere are subjectto immigration and emigration (de la Mare and Kerry1994; Cooper and Weimerskirch 2003). Fourteen birdsbanded at Bird Island, including ten that were breed-ing, were sighted at one of the other 30 colonies duringa South Georgia island-wide wandering albatross cen-sus in 2003/2004 (S. Poncet, personal communication).One non-breeding banded Wandering albatross seen atAlbatross Island, 60 km southeast of Bird Island, inJanuary 2004 was sighted from a longliner at ScotiaRidge 1 month subsequent; the bird had not beensighted since being banded as an adult on Bird Islandin 1991.
Alternatively, birds with or without breeding experi-ence that are not resighted at the colony for manyyears may be chronic non-breeders (which may nevervisit the island, or do so only for short periods). Theseindividuals may be of lower intrinsic quality, unable toacquire suYcient body condition to attempt breeding,or have found it diYcult to secure a nest site or replacea dead partner (Chastel et al. 1995; Catry et al. 1998).
Timing and location of sightings
Although Croxall et al. (1999) reported no sightings ofcurrent breeders in the southern Patagonian Shelfduring the brooding period, we observed ten individu-als whose chicks had hatched recently. This concurswith recent satellite-tracking data (Xavier et al. 2003b;BAS unpublished data) which indicate that althoughthe majority of trips during brooding last 4 days orless and are conWned to the local South Georgia shelf
123

Polar Biol
and shelf-slope, a small proportion of breeders travelas far as Falkland waters (cf. Prince et al. 1998). How-ever, the great majority of birds that we recorded dur-ing this period would have been immatures or non-breeders (many on sabbatical).
Most wandering albatrosses were sighted over waterdepths of 1,000–1,500 m, the preferred depth at whichthe longliners Wshed for toothWsh. This concurs withsatellite-tracking data indicating that wandering alba-trosses from South Georgia tend to feed in oceanicwaters or on the shelf slope, with only a small propor-tion of time spent in shallow waters (<200 m) usuallyonly in the southern portion of the Patagonian Shelf(Nicholls et al. 2002).
It is noteworthy that most birds recorded on multi-ple occasions were only seen in one zone and thatdespite an interval of up to 10 years between the sight-ings of Croxall et al. (1999) and this study, records of11 of the 17 birds sighted in both studies were from thesame zone. This suggests that some wandering alba-trosses show Wdelity between years to particular areason the Patagonian Shelf. Serial recaptures of the samewandering albatrosses oV New South Wales, and arecent tracking study of migrating black-browed alba-trosses, similarly indicate consistent preferences forparticular foraging sites or regions by individual non-breeders (Nicholls et al. 1995; Phillips et al. 2005).
Association with Wshing vessel
Contrary to the 1 year survey of Croxall et al. (1999) inwhich few birds were resighted and a high turnoverrate of individuals attending vessels was suggested, ourstudy proposes that many wandering albatrosses showstrong and long-lasting associations with longliners onthe southern Patagonian Shelf. Almost half of the wan-dering albatrosses sighted in this study were seen morethan once and 17 of the 86 birds seen by Croxall et al.(1999) during 1996/1997 were sighted again during2001–2005. In addition, Wve individuals seen duringboth studies were sighted on multiple occasions. Simi-larly, analysis of the satellite tracks overlaid on vesselmovements (Fig. 3) conWrms that a particular wander-ing albatross may associate with a longliner for severaldays. Indeed, the two birds tracked over their entirefeeding trip spent around half of the time away fromthe colony in the vicinity of a single vessel. This is com-paratively higher than satellite tracked Westlandpetrels (Procellaria westlandica), which spent an aver-age of one third of their foraging trip within 5 km ofWnWsh trawlers (Freeman et al. 2001). This appeared tobe a facultative association because two birds alsospent considerable time foraging in other areas, possi-
bly on natural prey, during the trip, as was alsoreported for Westland petrels (Freeman et al. 2001).The strong association of South Georgia wanderingalbatross with longliners agrees well with recent dietstudies, where toothWsh remains represented 43–63%of the estimated wet mass of stomach contents (Xavieret al. 2004). “FinWsh” trawlers are estimated to provide4% of the total annual energy requirements of black-browed albatrosses breeding in the Falkland Islands(Thompson and Riddy 1995) and it would appear thattoothWsh longliners Wshing in southern PatagonianShelf and South Georgia waters may well provide ahigher proportion of the diet of wandering albatrossesfrom South Georgia at certain times of year.
Giant petrels
Although male northern and southern giant petrelstend to depend primarily on seabird and seal carcassesat South Georgia, in some years their diet and that offemales includes a substantial proportion of krill(Hunter 1983). Recent research also suggests that giantpetrels elsewhere feed to a considerable extent on Wsh,squid and crustaceans (Forero et al. 2004). Our studyindicates that at least a proportion of the diet of Wshand squid identiWed by Forero et al. (2004) could bescavenged—as discarded oVal, bycatch species andsquid bait—from longliners.
More sightings were made of females than males,and of banded northern giant petrels than southerngiant petrels from Bird Island, and sightings of north-ern giant petrels were generally in more northerly loca-tions. This conWrms what is known of foraging areasfrom satellite tracking of incubating birds of both spe-cies (González-Solis et al. 2000) and at-sea surveys inFalkland Island waters (White et al. 2002) and suggeststhat the general separation between females that for-age far from the colony in pelagic waters or on distantshelves, and males that tend to remain around theSouth Georgia coasts, also occurs outside of the incu-bation period.
Five of the 14 banded northern and southern giantpetrels were sighted at vessels over 2–3 days and threeindividuals were also resighted at intervals of10 months, Wrst on Scotia Ridge and then in FalklandIslands waters. This suggests that, like wandering alba-trosses, some northern and southern giant petrels showconsistent associations with longliners on the southernPatagonian Shelf.
Southern giant petrels breeding on the Argentinecoastline forage across the Patagonian Shelf fromcoastal areas to the shelf break (Quintana andDell’Arciprete 2002). Of the total of 45 banded south-
123

Polar Biol
ern giant petrel Xedglings from the Chubut province ofArgentina that have been sighted at sea (includingfrom the Argentine coastline), 84% were of individualsassociating with Wshing vessels on the Patagonian, Bra-zilian and Uruguayan shelves (Copello et al. 2005).
Fisheries have a variety of impacts on marine preda-tors such as albatrosses and petrels. They can aVectpopulation numbers directly, through direct mortalityor provision of discards, or indirectly, through theireVects on the marine ecosystem and seabird prey (Reidet al. 2004; Votier et al. 2004; Sullivan et al. 2006).Improving our knowledge of where and how alba-trosses interact with Wsheries is essential if we are tofully understand the impacts of Wsheries, and to informconservation eVorts.
Acknowledgements Special thanks to FIFD observers AndyBlack, Ross James, Matthew Pearce and Oli Yates for their sight-ings, J. Pompert who supervised the FIFD observer program andJanet Silk for the Wgures. Flavio Quintana (Centro Nacional Pa-tagónico, Conicet Argentina) and Fabian Rabufetti (Aves Ar-gentinas) kindly provided information about their southern giantpetrel banding program. We also express thanks to Sally Poncetfor providing records of Bird Island banded albatrosses seen dur-ing the 2003/2004 South Georgia albatross census and advice onan earlier draft of this paper. A. Arkhipkin and two anonymousreferees also helped to improve the manuscript. The support ofthe Fisheries Patrol OYcers and the crew of the Fishery PatrolVessels Dorada and Sigma for the transfer of observers were crit-ical. H.O. wishes to thank Consolidated Fisheries Limited whoprovided funding for the collation of the data and to the Govern-ment of South Georgia and South Sandwich Islands for generousassistance during manuscript preparation. We thank all thosewho have banded wandering albatrosses and giant petrels at BirdIsland since 1958.
References
Catry P, Phillips RA, Hamer KC, RatcliVe N, Furness RW(1998) The incidence of non-breeding by adult great skuasand parasitic jaegers from Foula, Shetland. Condor100:448–455
Chastel O, Weimerskirch H, Jouventin P (1995) InXuence ofbody condition on reproductive decision and reproductivesuccess in the blue petrel. Auk 112:964–972
Cooper J, Weimerskirch H (2003) Exchange of the Wanderingalbatross Diomedea exulans between the Prince Edward andCrozet Islands: implications for conservation. Afr J Mar Sci25:519–523
Copello S, Quintana F, Rabufetti (2005) Dispersión de juvenilesde petrel gigante del sur, Macronectes giganteus, anillados encolonias del norte de Patagonia. XI Reuníón Argentina deOrnotología. 7–10 September 2005, Buenos Aires
Croxall JP, Gales R (1998) An assessment of the conservationstatus of albatrosses. In: Robertson G, Gales R (eds) Alba-tross biology and conservation. Surrey Beatty and Sons,Chipping Norton, pp 46–65
Croxall JP, Wood AG (2002) The importance of the PatagonianShelf for top predator species breeding at South Georgia.Aquat Conserv Mar Freshw Ecosyst 12:101–118
Croxall JP, Prince PA, Rothery P, Wood AG (1998) Populationchanges in albatrosses at South Georgia. In: Robertson G,Gales R (eds) Albatross biology and conservation. SurreyBeatty and Sons, Chipping Norton, pp 69–83
Croxall JP, Black AD, Wood AG (1999) Age, sex and status ofwandering albatrosses Diomedea exulans L. in Falkland Is-lands waters. Antarct Sci 11:150–156
de la Mare WK, Kerry KR (1994) Population dynamics of thewandering albatross (Diomedea exulans) on Macquarie Is-land and the eVects of mortality from longline Wshing. PolarBiol 14:231–241
Forero MG, Bortolloti, Hobson KA, Donazar JA, Bertelloti M,Blanco G (2004) High trophic overlap within the seabirdcommunity of Argentinean Patagonia: a multiscale ap-proach. J Anim Ecol 73:789–801
Freeman AND, Wilson KJ, Nicholls DG (2001) Westland petrelsand the hoki Wshery: determining co-occurrence using satel-lite telemetry. Emu 101:47–56
Gales R, Brothers N, Reid T (1998) Seabird mortality in the Jap-anese tuna longline Wshery around Australia. Biol Conserv86:37–56
González-Solis J, Croxall JP, Wood AG (2000) Foraging parti-tioning between giant petrels Macronectes spp. and its rela-tionship with breeding population changes at Bird Island,South Georgia. Mar Ecol Prog Ser 204:279–288
Hunter S (1983) The food and feeding ecology of the giant petrelsMacronectes halli and M. giganteus at South Georgia. J ZoolLond 200:521–538
Hunter S (1984) Breeding biology and population dynamics of gi-ant petrels Macronectes at South Georgia (Aves: Procellarii-formes). J Zool Lond 203:441–460
Laptikhovsky V, Brickle P (2005) The Patagonian toothWsh Wsh-ery in Falkland Islands’ waters. Fish Res 74:11–23
Nicholls DG, Murray D, Battam H, Robertson G, Moors P,Butcher E, Hildebrandt M (1995) Satellite tracking of thewandering albatross Diomedea exulans around Australiaand in the Indian Ocean. Emu 95:223–230
Nicholls DG, Robertson CJR, Prince PA, Murray MD, WalkerKJ, Elliott GP (2002) Foraging niches of three Diomedeaalbatrosses. Mar Ecol Prog Ser 231:269–277
Otley H (2005) Seabird attendance at Patagonian toothWsh long-liners in the Falkland Islands over three years. Falkland Is-lands Fisheries Department, Stanley, Falkland Islands
Phillips RA, Silk JRD, Croxall JP, Afanasyev V, Bennett VJ(2005) Summer distribution and migration of nonbreedingalbatrosses: individual consistencies and implications forconservation. Ecology 86:2386–2396
Pickering SPC (1989) Attendance patterns and behaviour in rela-tion to experience and pair-bond formation in the wanderingalbatross Diomedea exulans at South Georgia. Ibis 131:183–195
Poncet S, Robertson G, Phillips RA, Lawton K, Phalan B, Tra-than PN, Croxall JP (2006) Status and distribution of wan-dering, black-browed and grey-headed albatrosses breedingat South Georgia. Polar Biol (online Wrst)
Prince PA, Croxall JP, Trathan PN, Wood AG (1998) The pelagicdistribution of South Georgia albatrosses and their relation-ships with Wsheries. In: Robertson G, Gales R (eds) Alba-tross biology and conservation. Surrey Beatty and Sons,Chipping Norton, pp 137–167
Quintana F, Dell’Arciprete OP (2002) Foraging grounds ofsouthern giant petrels (Macronectes giganteus) on the Pata-gonian shelf. Polar Biol 25:159–161
Reid TA, Sullivan BJ (2004) Longliners, black-browed albatrossmortality and bait scavenging in Falkland Island waters:what is the relationship? Polar Biol 27:131–139
123

Polar Biol
Reid TA, Sullivan BJ, Pompert J, Enticott JW, Black AD (2004)Seabird mortality associated with Patagonian toothWsh(Dissostichus eleginoides). Emu 104:317–325
Ryan PG, Boix-Hinzen C (1999) Consistent male-biased seabirdmortality in the Patagonian toothWsh longline Wshery. Auk116:851–854
Sullivan BJ, Reid TA, Bugoni L (2006) Seabird mortality on fac-tory trawlers in the Falkland Islands and beyond. Biol Con-serv 131:495–504
Thompson KR, Riddy MD (1995) Utilization of oVal and discardsfrom “WnWsh” trawlers around the Falkland Islands by theBlack-browed albatross Diomedea melanophris. Ibis137:198–206
Tickell WLN (1968) The biology of the great albatrosses, Diome-dea exulans and Diomedea epomophora. In: Austin OL Jr(ed) Antarctic bird studies. American Geophysical Union,Washington, pp 1–55
Tickell WLN (2000) Albatrosses. Pica Press, East SussexVotier SC, Furness RW, Bearhop S, Crane JE, Caldow RWG,
Catry P, Ensor K, Hamer KC, Hudson AV, Kalmbach E,Klomp NI, PfeiVer P, Phillips RA, Prieto I, Thompson DR
(2004) Changes in Wsheries discard rates and seabird commu-nities. Nature 427:727–730
Waugh S, Filippi D, Fukuda A, Suzuki M, Higuchi H, SetiawanA, Davis L (2005) Foraging of royal albatrosses, Diomedeaepomophora, from the Otago Peninsula and its relationshipsto Wsheries. Can J Fish Aquat Sci 62:1410–1421
White RW, Gillon KW, Black AD, Reid JB (2002) The distribu-tion of seabirds and marine mammals in Falkland Islands wa-ters. JNCC and Falklands Conservation, Peterborough
Xavier JC, Croxall JP, Trathan PN, Wood AG (2003a) Feedingstrategies and diets of breeding grey-headed and wanderingalbatrosses at South Georgia. Mar Biol 143:221–232
Xavier JC, Croxall JP, Trathan PN, Wood AG (2003b) Interan-nual variation in the diets of two albatross species breedingat South Georgia: implications for breeding performance.Ibis 145:593–610
Xavier JC, Trathan PN, Croxall JP, Wood AG, Podesta G, Rod-house PG (2004) Foraging ecology and interactions with Wsh-eries of wandering albatrosses (Diomedea exulans) breedingat South Georgia. Fish Oceanogr 13:324–344
123
Copyright © 2022 FDOKUMEN




















