
Organization-dependent effects of toxic bivalent ions
9
Eur. J. Biochem. 267, 4731–4739 (2000) q FEBS 2000 Organization-dependent effects of toxic bivalent ions Microtubule assembly and glycolysis Ka ´ roly Liliom 1 , Ga ´ bor Wa ´ gner 1 , Anita Pa ´ cz 1 , Marta Cascante 2 , Ja ´ nos Kova ´ cs 3 and Judit Ova ´ di 1 1 Institute of Enzymology, Biological Research Center of the Hungarian Academy of Sciences, Budapest, Hungary; 2 Department of Biochemistry and Molecular Biology, Faculty of Chemistry, University of Barcelona, Catalonia, Spain; 3 Department of General Zoology, Faculty of Sciences, University of Eo ¨tvo ¨s Lora ´nd, Budapest, Hungary The effects of bivalent ions on tubulin dynamics and the upper phase of glycolysis were investigated at different organization levels in vitro. Cu 21 , Cd 21 , Hg 21 and CrO 2– 4 inhibit the tubulin polymerization at an IC 50 of 14–24 mm with high cooperativity and also induce microtubule disassembly. The apparent binding constants of the ions to tubulin, estimated by fluorescence quenching, vary between 6 and 28 mm. BIAcore measurements for tubulin–tubulin interaction suggest that the presence of Cu 21 affects neither k off nor k on , but the amount of the bound tubulin. While the inhibitory effect of Cu 21 on tubulin polymerization is partially abolished by cross- linking of microtubules with substoichiometric amounts of phosphofructokinase or decoration of tubules with cytosolic proteins, in the presence of kinase but not with cytosolic proteins the tubules are resistant to CrO 4 2– . No inhibitory effect of Cu 21 or CrO 2– 4 on microtubule assembly was detected in the MAP-containing cytosolic fraction. Electron microscopy revealed that tubules assembled in the presence of Cu 21 or CrO 2– 4 ions contain aggregates of thread-like oligomers that are less conspicuous in the presence of cytosolic proteins. Cu 21 , Cd 21 , and Hg 21 inhibit the glycolytic flux in the cytosolic fraction characterized at equilibrium by an IC 50 of 10–14 mm with high cooperativity. Tubulin diminishes the inhibitory effect of the cations. These data indicate that the responses elicited by the bivalent ions are highly dependent on the supramolecular organization of the systems. Keywords: glycolysis; microtubule; organization; toxic ions. Metals are essential for many cell functions. The key roles of the major cations, Na 1 ,K 1 , Mg 21 and Ca 21 , have been extensively characterized. There are trace elements (e.g. Cu 21 ) that are essential for functions of specific proteins: metalloen- zymes, stress proteins, oxygen transporters, and redox activities [1]. As their concentration exceeds the physiological level, however, their effects on cells become deleterious. Eventually they become toxic and can cause pathological effects. The nonessential metals, such as Cd 21 and Hg 21 , are potentially very toxic if their internal bioavailable concentration exceeds the capacity of physiological detoxification processes. The principal target for many toxic metals are cells of the nervous system [2]; many diseases are related to metal poisoning against excess cations. Microtubules (MTs) organize axonal vesicular traffic and are thus important and abundant sub- cellular components of neurocytes. Hexavalent chromium (e.g. CrO 2– 4 ) has the ability to penetrate all tissues in the body and produces oxidative stress and oxidative DNA damage by generating active oxygen radicals and reactive Cr(Vc) species [3]. The assembly/disassembly process of MTs built up of tubulin dimers has been observed in intact cells, cell-free preparations, and reconstituted systems. The assembly and function of MTs are regulated by MAPs and a variety of other effectors and environmental conditions. Sulfhydryl reagents block tubulin polymerization in vitro [4] by reacting with two of the eight SH-groups per tubulin dimer [5] that are essential for MT assembly [6]. An early study indicated the interaction of sulfhydryl groups of tubulin with Cu 21 , Cd 21 , and Hg 21 [7]. Copper also affects cells’ energy production by decreasing the glycolytic rate in the brain cytosolic fraction [8] and also in the muscle cytosolic fraction [9]. Recently, we provided evidence for the combined enhance- ment of MT-assembly and glucose metabolism, which resulted in decreased sensitivity to copper toxicity [10]. The mutual enhancement effect appeared to be specific for neuronal systems as it was observed in microtubular protein-free cytosolic fractions (MPFCFs) from whole brain and from cultured neuroblastoma cells but not from muscle cytosolic fraction. Here we have extended our studies to other bivalent ions, Cu 21 , Cd 21 , CrO 2– 4 , Hg 21 , and Pb 21 , as well as the monovalent anion NO – 2 . We characterized the binding of the ions to target proteins as well as its structural and functional consequences. These studies at different organization levels rendered it possible to establish a role for macromolecular associations in response to different stress effects. MATERIALS AND METHODS Chemicals Cadmium acetate, potassium chromate, copper chloride, lead acetate, mercury nitrate and sodium nitrite were obtained from Correspondence to J. Ova ´di, Institute of Enzymology, Biological Research Center of the Hungarian Academy of Sciences, H-1518 Budapest, P.O. Box 7, Hungary. E-mail: [email protected] Abbreviations: BSA, bovine serum albumin; CCF, crude cytosolic fraction; G-6-PDH, glucose-6-phosphate dehydrogenase; IC 50 , half inhibitory concentration; MAP, microtubule-associated protein; Mes, 2-[N-morpholino]ethanesulfonic acid; MPFCF, microtubular protein-free cytosolic fraction; MT, microtubule; PFK, phosphofructokinase. Note: the first two authors contributed equally to this work. (Received 6 March 2000, revised 30 May 2000, accepted 30 May 2000)
Transcript of Organization-dependent effects of toxic bivalent ions

Eur. J. Biochem. 267, 4731±4739 (2000) q FEBS 2000
Organization-dependent effects of toxic bivalent ionsMicrotubule assembly and glycolysis
Ka roly Liliom1, Ga bor Wa gner1, Anita Pa cz1, Marta Cascante2, Ja nos Kova cs3 and Judit Ova di1
1Institute of Enzymology, Biological Research Center of the Hungarian Academy of Sciences, Budapest, Hungary; 2Department of Biochemistry
and Molecular Biology, Faculty of Chemistry, University of Barcelona, Catalonia, Spain; 3Department of General Zoology, Faculty of Sciences,
University of EoÈtvoÈs LoraÂnd, Budapest, Hungary
The effects of bivalent ions on tubulin dynamics and the upper phase of glycolysis were investigated at
different organization levels in vitro. Cu21, Cd21, Hg21 and CrO2±4 inhibit the tubulin polymerization at an IC50
of 14±24 mm with high cooperativity and also induce microtubule disassembly. The apparent binding constants
of the ions to tubulin, estimated by fluorescence quenching, vary between 6 and 28 mm. BIAcore measurements
for tubulin±tubulin interaction suggest that the presence of Cu21 affects neither koff nor kon, but the amount of the
bound tubulin. While the inhibitory effect of Cu21 on tubulin polymerization is partially abolished by cross-
linking of microtubules with substoichiometric amounts of phosphofructokinase or decoration of tubules with
cytosolic proteins, in the presence of kinase but not with cytosolic proteins the tubules are resistant to CrO42±. No
inhibitory effect of Cu21 or CrO2±4 on microtubule assembly was detected in the MAP-containing cytosolic
fraction. Electron microscopy revealed that tubules assembled in the presence of Cu21 or CrO2±4 ions contain
aggregates of thread-like oligomers that are less conspicuous in the presence of cytosolic proteins. Cu21, Cd21,
and Hg21 inhibit the glycolytic flux in the cytosolic fraction characterized at equilibrium by an IC50 of 10±14 mm
with high cooperativity. Tubulin diminishes the inhibitory effect of the cations. These data indicate that the
responses elicited by the bivalent ions are highly dependent on the supramolecular organization of the systems.
Keywords: glycolysis; microtubule; organization; toxic ions.
Metals are essential for many cell functions. The key roles ofthe major cations, Na1, K1, Mg21 and Ca21, have beenextensively characterized. There are trace elements (e.g. Cu21)that are essential for functions of specific proteins: metalloen-zymes, stress proteins, oxygen transporters, and redox activities[1]. As their concentration exceeds the physiological level,however, their effects on cells become deleterious. Eventuallythey become toxic and can cause pathological effects. Thenonessential metals, such as Cd21 and Hg21, are potentiallyvery toxic if their internal bioavailable concentration exceedsthe capacity of physiological detoxification processes. Theprincipal target for many toxic metals are cells of the nervoussystem [2]; many diseases are related to metal poisoningagainst excess cations. Microtubules (MTs) organize axonalvesicular traffic and are thus important and abundant sub-cellular components of neurocytes. Hexavalent chromium (e.g.CrO2±
4 ) has the ability to penetrate all tissues in the body andproduces oxidative stress and oxidative DNA damage bygenerating active oxygen radicals and reactive Cr(Vc) species[3].
The assembly/disassembly process of MTs built up of tubulindimers has been observed in intact cells, cell-free preparations,and reconstituted systems. The assembly and function of MTsare regulated by MAPs and a variety of other effectors andenvironmental conditions. Sulfhydryl reagents block tubulinpolymerization in vitro [4] by reacting with two of the eightSH-groups per tubulin dimer [5] that are essential for MTassembly [6]. An early study indicated the interaction ofsulfhydryl groups of tubulin with Cu21, Cd21, and Hg21 [7].Copper also affects cells' energy production by decreasing theglycolytic rate in the brain cytosolic fraction [8] and also in themuscle cytosolic fraction [9].
Recently, we provided evidence for the combined enhance-ment of MT-assembly and glucose metabolism, which resultedin decreased sensitivity to copper toxicity [10]. The mutualenhancement effect appeared to be specific for neuronalsystems as it was observed in microtubular protein-freecytosolic fractions (MPFCFs) from whole brain and fromcultured neuroblastoma cells but not from muscle cytosolicfraction. Here we have extended our studies to other bivalentions, Cu21, Cd21, CrO2±
4 , Hg21, and Pb21, as well as themonovalent anion NO±
2. We characterized the binding of theions to target proteins as well as its structural and functionalconsequences. These studies at different organization levelsrendered it possible to establish a role for macromolecularassociations in response to different stress effects.
M A T E R I A L S A N D M E T H O D S
Chemicals
Cadmium acetate, potassium chromate, copper chloride, leadacetate, mercury nitrate and sodium nitrite were obtained from
Correspondence to J. OvaÂdi, Institute of Enzymology, Biological Research
Center of the Hungarian Academy of Sciences, H-1518 Budapest,
P.O. Box 7, Hungary. E-mail: [email protected]
Abbreviations: BSA, bovine serum albumin; CCF, crude cytosolic
fraction; G-6-PDH, glucose-6-phosphate dehydrogenase; IC50, half
inhibitory concentration; MAP, microtubule-associated protein;
Mes, 2-[N-morpholino]ethanesulfonic acid; MPFCF, microtubular
protein-free cytosolic fraction; MT, microtubule; PFK,
phosphofructokinase.
Note: the first two authors contributed equally to this work.
(Received 6 March 2000, revised 30 May 2000, accepted 30 May 2000)

Merck (Merck KGaA, Darmstadt, Germany). Bovine serumalbumin (BSA), glucose-6-phosphate dehydrogenase (G-6-PDH) from yeast, phosphofructokinase (PFK) from rabbitmuscle, taxol, Mes, EGTA, and GTP were purchased fromSigma Chemical Co. (St Louis, MO). All other chemicals werereagent grade commercial preparations. High purity water of aMilli-Q system was used for preparing the solutions (Millipore,Bedford, MA).
Routine measurements of protein concentration were doneby the Bradford method [11], using the Bio-Rad Protein AssayKit (Bio-Rad Laboratories GmbH, MuÈnchen, Germany).
Preparation of tubulin
MAP-free tubulin was purified from calf brain as described byNa and Timasheff [12]. For assembly experiments, the storedtubulin was dialyzed against 50 mm Mes buffer, pH 6.8 at 4 8Cas described previously [10].
Preparation of cytosolic fractions
Microtubular protein-free cytosolic fraction (MPFCF) frombovine brain was prepared using the method of Vallee [13] withslight modifications as described recently [10]. Briefly, braintissue was homogenized in 100 mm piperazine-N,N 0-bis[2-ethanesulfonic acid] buffer, pH 6.6, with 1 mm EGTA and1 mm MgCl2, also containing 2 mm dithioerythritol, 0.6 mmphenylmethylsulfonylfluoride, and 1 mm benzamidine. Thehomogenate was centrifuged first at 30 000 g at 4 8C for20 min, then the supernatant was further centrifuged at100 000 g at 4 8C for 30 min. The supernatant of this lattercentrifugation comprised the soluble proteins of cells, includingtubulin, MAPs, glycolytic enzymes, as well as other cytosolicproteins, and small molecules. This preparation was used as thecrude cytosolic fraction (CCF). To remove tubulin and MAPs,1 mm GTP and 20 mm taxol were added to the supernatant,warmed up to 37 8C and kept warm for 40 min. Then thesolution was centrifuged at 100 000 g at 37 8C for 30 min andthe supernatant was gel-filtered through a Sephadex G-25column to remove small molecules and in some casesconcentrated with a Miniplus device (Millipore) if necessary.This preparation (MPFCF) was free from microtubular proteinsas shown by gel electrophoresis.
Turbidity measurements
The assembly of tubulin dimers into microtubules was followedby turbidimetry [14] by monitoring the change in absorbance at350 nm with a Jasco V-550 UV/VIS spectrophotometer (JascoCorporation, Tokyo, Japan). The temperature of the sample wascontrolled by a Grant model Y6 thermostatically regulatedliquid circulator (Grant Instruments Ltd, Royston, England).Tubulin was diluted to 10 mm calculated as dimers in 50 mmMes buffer, pH 6.8, containing 100 mm KCl, 1 mm EGTA,1 mm MgCl2. After the temperature of the sample reached37 8C in the cuvette holder, microtubule assembly was initiatedby adding taxol to a final concentration of 20 mm. Tubulin wasincubated with the compounds before the initiation ofpolymerization at 4 8C for 20 min. From the time-dependenceof turbidity, the duration of the initial lag-phase, the maximalslope of the initial rise and the net increase of turbidity weredetermined.
Flourescence quenching
The binding of ligands to tubulin dimers was determined byfluorescence quenching titrations with a Jasco FP-777 (Jasco
Corporation, Tokyo, Japan) spectrofluorometer. The excitationwavelength was 290 nm and the emitted light was measured bysweeping the range between 320 and 350 nm. Both theexcitation and the emission light bandwidths were 1.5 nm.Fluorescence measurements were carried out in 20 mmphosphate buffer, pH 6.9, containing 1 mm EGTA and0.5 mm MgCl2. From the fluorescence data, the maximalfractional quenching and the apparent dissociation constantwere determined. When the inner filter effect was significant,correction was done by implementing the graphical correctionprocedure of Mertens and KaÈgi [15] (see details below).
Surface plasmon resonance
Real-time monitoring of the binding kinetics of tubulin±tubulininteraction was done using a BIAcore X instrument (BIAcoreAB, Uppsala, Sweden). The analysis for the dissociation andassociation rate constants was done by the BIAevaluation 3.0software supplied by the manufacturer. Tubulin was biotiny-lated with a sulfosuccinimidyl ester derivative of biotin havingan extended spacer arm (B-6352, Molecular Probes) in Mesbuffer (50 mm, pH 7.0, 2 mm dithioerythritol, 1 mm MgCl2,1 mm EGTA) at 4 8C for 90 min, followed by stopping thereaction by 120 mm K-glutamate and separating the biotiny-lated protein in a Sephadex G-25 column. The biotinylatedtubulin was then immobilized on a streptavidin-coated sensorchip (Sensor Chip SA, product code BR-1000±32, BIAcore) ata concentration of 50 mg´mL21, at a flow rate of 5 mL´min21
for 5 min, yielding a net increase in the sensorgram of 1800RU. All BIAcore experiments were done at 20 8C. The chipwas equilibrated with the running buffer until a stable baselinewas obtained (50 mm Mes buffer pH 7.0, 2 mm dithioerythritol,1 mm MgCl2, 1 mm EGTA) and samples were injected over thesurface at 5 mL´min21 for 3 min.
Measurement of the glycolytic flux
The steady-state flux of the upper phase of glycolysis catalyzedby hexokinase, glucosephosphate isomerase, PFK and aldolasewas measured in the MPFCF by coupling the reaction withexcess triosephosphate isomerase and glycerophosphate dehy-drogenase activities. The glycolytic flux was proportional to therate of decrease in NADH concentration followed at 340 or384 nm. The experiments were carried out in N-[2-hydroxy-ethyl]piperazine-N 0-[2-ethansulfonic acid] buffer (50 mm,pH 7.4, containing 100 mm KCl, 10 mm MgCl2, 10 mmNa2HPO4), 11 U´mL21 triosephosphate isomerase,1.1 U´mL21 glycerophosphate dehydrogenase, 0.16 or 0.8 mmNADH detected at 340 or 384 nm, respectively, and 2 mm Mg-ATP, starting the reactions with the addition of 10 mm glucose.The NADH-consumption was monitored by a Jasco V550 or aCary 50 spectrophotometer at 25 8C.
Statistical evaluations of data
The dose-dependent inhibition of microtubule assembly causedby the toxic ligands were quantified using the followingequation:
E � 100 � �1 2 M � Ln/�ICn50 1 Ln�� �1�
where E is the relative effect in percentages normalized to thecontrol, obtained at a given concentration of the ligand (L); M isfor the maximal inhibition at infinite ligand concentration; IC50
is the concentration of the ligand that elicits half of the maximaleffect; and n is the pseudo-Hill coefficient.
Apparent dissociation constants were determined from thefluorescent quenching data. The inner filter effect was corrected
4732 K. Liliom et al. (Eur. J. Biochem. 267) q FEBS 2000

according to Mertens and KaÈgi [15]. The observed residualfluorescence, F, can be given by the equation:
F � F0 � �1 2 Q max � L/�K appd 1 L�� � exp �2IFE � L� �2�
where F0 is the fluorescence intensity measured in the absenceof the absorber; Q max is the maximal fractional quenching; IFEis a constant characterizing the inner filter effect caused by theligand; and K app
d is the apparent dissociation constant of theinteraction between tubulin and the ligand (L) assuming 1 : 1stoichiometry. The first term accounts for the quenching whilethe second term accounts for the inner filter effect of the ligand.The value of IFE depends on the light path through the solution.For the evaluation of the binding data, the natural logarithm ofequation 2 was fitted to the data points by nonlinear regression:
ln F � ln F0 1 ln �1 2 Q max � L/�K appd 1 L��2 IFE � L �3�
A Student's t-test at 95% probability level was carried out in allcases when comparison was made between mean values.
Transmission electron microscopy
MT-containing samples polymerized in the absence andpresence of cytosolic fractions with and without Cu21 orCrO4
2± were pelleted by centrifugation at 100 000 g at 37 8Cfor 20 min. The pellets were prefixed (2% glutaraldehyde (v/v),0.2% tannic acid (w/v), 0.1 m sodium cacodylate pH 7.4, atambient temperature for 60 min). The pellets were washed(0.1 m sodium cacodylate pH 7.4), postfixed (0.5% OsO4 (w/v)in 0.1 m sodium cacodylate pH 7.4, at an ambient temperaturefor 60 min), washed again and stained in one batch (1% uranylacetate (w/v), 30 min). Stained samples were dehydrated ingraded ethanol and embedded in Durcupan (Fluka). Thinsections were contrasted with uranyl acetate and lead citrate.The samples were examined and photographed in a JEOL CX100 electron microscope operated at an accelerating voltage of60 kV. Magnification was calibrated with a diffraction gratingreplica (2160 line per mm, Balzers).
R E S U L T S
Effects of ions in separated systems
Tubulin polymerization. Four cations, Cu21, Cd21, Hg21, Pb21,and two anions, CrO2±
4 and NO22 , were tested for their
ability to influence the taxol-assisted polymerization ofpurified MAP-free tubulin followed by turbidimetry. Normal-ized dose±response curves obtained from the maximalslopes of the turbidity enhancement for the four most effectiveions are shown in Fig. 1. The polymerization in three out of thefour ions in Fig. 1 levels off at 30±40% of the control value.From the dose±response curves the IC50 values and the pseudo-Hill coefficients (n) were estimated by fitting equation 1 to theexperimental points (Table 1). The IC50 values and the pseudo-Hill coefficients obtained from the net increase in turbidity,which is directly proportional to the total amount of polymerformed [14], were similar to those presented in Table 1 (datanot shown). Pb2± and NO2
2 appear to be weak inhibitors of MTassembly (Table 1).
When Cu21, Cd21, CrO2±4 , and Hg21 were added to the
preassembled MT, the turbidity levels were reduced by 30±40%and reached turbidity levels similar to those obtained when theions were added before polymerization (Table 1).
Fluorescence quenching. The binding of the ions to tubulin wasquantified by fluorescence quenching measurements. Thefluorescence emission of tubulin dimers was significantlyquenched by the four ions, Cu21, Cd21, CrO2±
4 , and Hg21. Asshown in Fig. 2, in the case of Hg21, the decrease influorescence intensity reaches a well-defined limit value,while in the case of Cu21 the curve is biphasic due to theinner filter effect caused by the free Cu21 ions. These titrationswere carried out at a relatively low tubulin concentration(1 mm) under conditions unfavourable for tubulin oligomeriza-tion. Thus the dissociation constants estimated by fittingequation 3 to the titration data (Table 1) likely refer to theequilibrium binding of the ions to the tubulin dimer. The
Fig. 1. Inhibition of tubulin polymerization
by (A) Cu21, (B) Cd21, (C) Hg21 and (D)
CrO2±4 . The dose±response curves were
generated by fitting the general logistic function
(equation 1) to the data by nonlinear regression.
The estimated IC50 values for the pseudo-Hill
coefficient, n, are given in Table 1.
Normalized data points are the mean values of
3±5 independent determinations with standard
errors of 6±7%.
q FEBS 2000 Organization-dependent responses to stimuli (Eur. J. Biochem. 267) 4733
titrations with Cd21 and CrO2±4 yielded similar results, although
the quenching efficiencies of Hg21 and Cu21 are higher thanthose of Cd21 and CrO2±
4 . The apparent dissociation constantsobtained for the binding of different ions to tubulin arecomparable and are in the range where they inhibit tubulinpolymerization.
Tubulin±tubulin interaction. The effect of Cu21 on tubulin±tubulin interaction as the initial step of MT assembly wasinvestigated by a commercial surface plasmon resonancebiosensor, BIAcore. Biotinylated tubulin was immobilized ona streptavidin matrix covalently attached to the sensor chip'ssurface and tubulin was injected over this layer. As shown inFig. 3, the binding of tubulin led to an increase in the amount ofprotein bound to the surface. As there is a linear relationshipbetween the mass of protein bound to the surface and therefractive index change monitored by the instrument, theassociation/dissociation rate constants of tubulin in solution to
the immobilized tubulin could be quantified in the absence andpresence of Cu21. Tubulin (0.1 mm) was injected over thetubulin-coated surface for 3 min. The values of the kineticconstants do not differ significantly in the absence and presenceof Cu21: kon, 1.47 and 1.34 � 104 m21´s21, respectively; koff,2.07 and 2.09 � 1023´s21, respectively. The dissociationconstants 1.41 and 1.56 � 1027 m, in the absence and presenceof Cu21, respectively, are not dependent on the presence ofCu21, although the amount of tubulin bound to the tubulin layerdecreased by 30% due to the presence of Cu21.
Flux of glycolysis. The steady-state flux of conversion ofglucose into triosephosphates was measured in 0.8 mg´mL21
brain cell-free extract at pH 7.4 at 25 8C. The consecutivereactions were coupled with excess triosephosphate isomerase
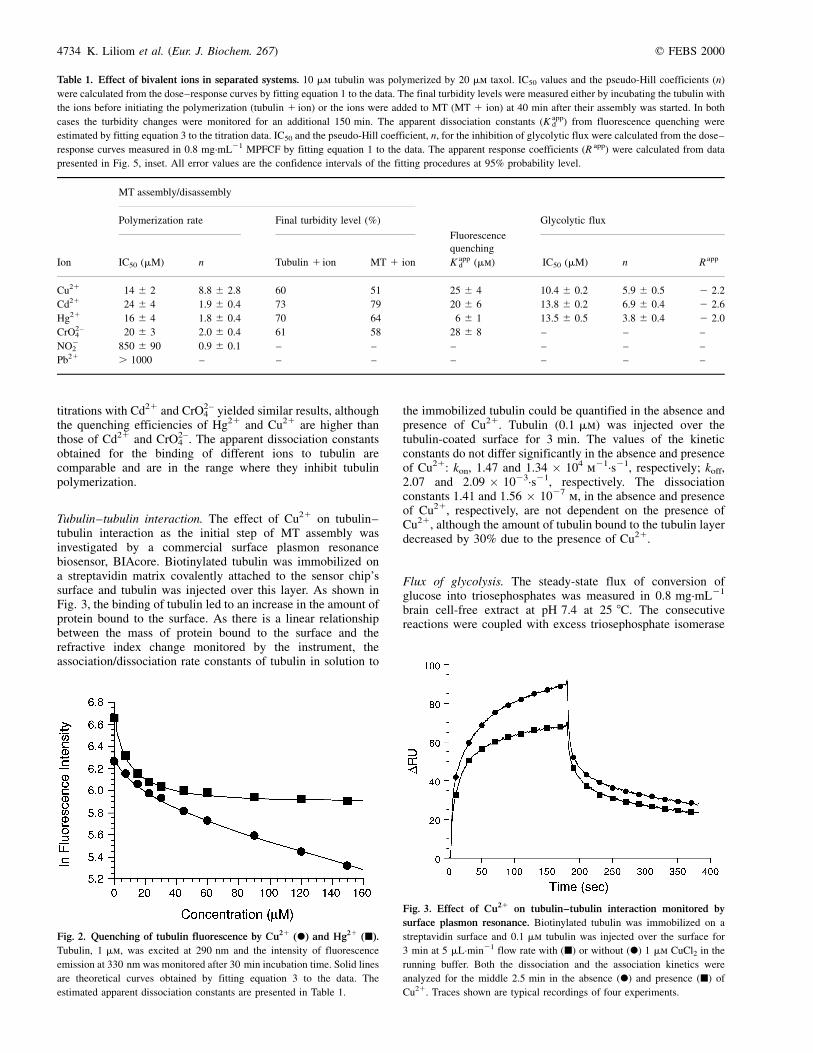
Table 1. Effect of bivalent ions in separated systems. 10 mm tubulin was polymerized by 20 mm taxol. IC50 values and the pseudo-Hill coefficients (n)
were calculated from the dose±response curves by fitting equation 1 to the data. The final turbidity levels were measured either by incubating the tubulin with
the ions before initiating the polymerization (tubulin 1 ion) or the ions were added to MT (MT 1 ion) at 40 min after their assembly was started. In both
cases the turbidity changes were monitored for an additional 150 min. The apparent dissociation constants (K appd ) from fluorescence quenching were
estimated by fitting equation 3 to the titration data. IC50 and the pseudo-Hill coefficient, n, for the inhibition of glycolytic flux were calculated from the dose±
response curves measured in 0.8 mg´mL21 MPFCF by fitting equation 1 to the data. The apparent response coefficients (R app) were calculated from data
presented in Fig. 5, inset. All error values are the confidence intervals of the fitting procedures at 95% probability level.
MT assembly/disassembly
Polymerization rate Final turbidity level (%)
Fluorescence
Glycolytic flux
Ion IC50 (mM) n Tubulin 1 ion MT 1 ion
quenching
K appd (mm) IC50 (mM) n R app
Cu21 14 �^ 2 8.8 �^ 2.8 60 51 25 �^ 4 10.4 �^ 0.2 5.9 �^ 0.5 2 2�.2
Cd21 24 �^ 4 1.9 �^ 0.4 73 79 20 �^ 6 13.8 �^ 0.2 6.9 �^ 0.4 2 2�.6
Hg21 16 �^ 4 1.8 �^ 0.4 70 64 6 �^ 1 13.5 �^ 0.5 3.8 �^ 0.4 2 2�.0
CrO2±4 20 �^ 3 2.0 �^ 0.4 61 58 28 �^ 8 ± ± ±
NO22 850 �^ 90 0.9 �^ 0.1 ± ± ± ± ± ±
Pb21 . 1000 ± ± ± ± ± ± ±
Fig. 2. Quenching of tubulin fluorescence by Cu21 (X) and Hg21 (B).
Tubulin, 1 mm, was excited at 290 nm and the intensity of fluorescence
emission at 330 nm was monitored after 30 min incubation time. Solid lines
are theoretical curves obtained by fitting equation 3 to the data. The
estimated apparent dissociation constants are presented in Table 1.
Fig. 3. Effect of Cu21 on tubulin±tubulin interaction monitored by
surface plasmon resonance. Biotinylated tubulin was immobilized on a
streptavidin surface and 0.1 mm tubulin was injected over the surface for
3 min at 5 mL´min21 flow rate with (B) or without (X) 1 mm CuCl2 in the
running buffer. Both the dissociation and the association kinetics were
analyzed for the middle 2.5 min in the absence (X) and presence (B) of
Cu21. Traces shown are typical recordings of four experiments.
4734 K. Liliom et al. (Eur. J. Biochem. 267) q FEBS 2000

and glycerophosphate dehydrogenase. The flux calculated fromthe linear segment of the absorbance versus time curve was7.8 ^ 0.3 U´g21 protein (n � 10). We have reported recentlythat 50 mm Cu21 decreases the flux of the upper phase ofglycolysis in the cytosolic fractions prepared from brain andmuscle [9,10]. Now we found that the addition of Cu21, Cd21,and Hg21 inhibited the flux similarly, and the effects of thethree cations were time and concentration dependent (data notshown). Pb21, CrO2±
4 , or NO±2 were ineffective up to a
concentration of 50 mm even after prolonged incubation time.The time-dependent and concentration-dependent inhibition
of glycolytic flux caused by Cu21 is shown in Fig. 4A. Thesolid lines are computed curves fitted to the dose±response datausing Eqn (1). IC50 values are decreased by increasing theincubation time from 25 to 10 mm. After 3 h of incubation withthe toxic ions the position and shape of the dose±responsecurve did not change significantly, which was indicative of
approaching the equilibrium state. The dose±response curvesfor Cd21 and Hg21 were also measured at 3 h incubation time.The IC50 values and cooperativity factors (pseudo-Hillcoefficient, n) estimated by curve fitting are presented inTable 1.
The sensitivity of a pathway towards perturbing effectors canbe quantitatively characterized by the response coefficient, R[16]. The experimental points of dose±response curves do notlie on straight lines in the log±log plot as supposed, but theslopes (R) change with the concentrations of metal ions(Fig. 4B). At low concentrations R is practically zero as theflux remains nearly maximal. Threshold values are reached byincreasing the concentrations of the ions (around 8±10 mmdepending on the ions), which are accompanied by dramaticchanges from low to high R-values in a narrow range of ionconcentrations. At these ion concentrations an abrupt decreaseof the glycolytic flux is observed; consequently R-values alsochange abruptly to values around 2 or higher. Thus, in this zonea little change in ion concentration results in a large change inR-values, and thus determinations of R-values are inaccurate.Above this region of metal concentration the flux decreasessteeply, and a quasi-linear dependence of the log-log plot isobserved at which flux becomes practically negligible and
Fig. 4. Dose±response curves for the inhibition of glycolytic flux in the
cytosolic fraction (MPFCF). (A) Time depencence of the effect of Cu21 at
incubation times: B, 5 min; X, 30 min; V, 60 min; P, 120 min; O,
180 min. Glycolysis was started by addition of glucose and measured for
15 min. The solid lines are theoretical curves obtained by fitting equation 1
to the experimental data. (B) Log±log plots of dose±response curves
constructed from the flux data measured after 3 h incubation with the
cations (O, Cu21 B, Cd21 X, Hg21). Equation 1 was fitted to the data by
nonlinear regression. Data points represent the mean of 3±4 independent
determinations with a standard error of 7%.
Fig. 5. Effect of 50 mm Cu21 and 50 mm CrO2±4 on MT assembly at
different organization levels. MT was assembled from purified MAP-free
tubulin in the presence of (A) 4 mm PFK, or (B) 2 mg´mL21 MPFCF; (C) or
the endogenous tubulin of the CCF was polymerized. Each panel also
contains the polymerization curves measured in the absence of additives.
(A) Curves a, b, and c, without PFK; curves d, e, f, with PFK. (B) Curves a,
b, and c, without MPFCF; curves d, e, f, with MPFCF. (C) Curves a, b, and
c, CCF at 3.4 mg´mL21 total protein concentration; curves d, e, f, CCF at
9.0 mg´mL21 total protein concentration. In (A±C): a and d, no ions; b and
e, with Cu21 c and f, with CrO2±4 ions.
q FEBS 2000 Organization-dependent responses to stimuli (Eur. J. Biochem. 267) 4735

R-values remain between 2 and 4. At an ion concentration of< 30 mm the flux becomes negligible.
To characterize quantitatively the effect of toxic ions on theglycolytic flux we suggest the IC50 value as an operationalpoint for evaluation of an apparent response coefficient (R app).The values of R app obtained graphically from the log-log plotsfor Cu21, Cd21, and Hg21 are presented in Table 1. As theslopes of the curves at IC50 values are very steep, the error inthe determinations is relatively large. However, if R app isevaluated from the direct dose±response curves by taking intoaccount the whole set of experimental points, the error ofdetermination can be reduced.
The dose±response curves can be generally described by thelogistic function:
J � Jo �1 2 X n/�IC 50n 1 Xn��
where J and Jo are the fluxes in the presence and absence of ion(X), respectively; and n is a pseudo-Hill coefficient. By derivingR J
X � (2J/J)/(2X/X) with the logistic function, and substitutingJ � Jo/2 and X � IC50, a special formula can be obtained:
Rappcalc � 2n/2
that is, sensitivity factors can be determined directly from thepseudo-Hill coefficients evaluated from the dose±responsecurves. If these values are compared with the R app valueobtained by graphical derivation (Table 1), it is clear that thetwo values are in good agreement in the case of Hg21, where nis relatively small, but the difference between the values is
obvious in the case of Cu21 and Cd21, where the values of n arehigh.
Effect of ions in combined systems
Tubulin polymerization at different organization levels. Theinhibitory effect of 50 mm Cu21 or CrO2±
4 on tubulinpolymerization was investigated in the absence and presenceof cytosolic proteins of different compositions. A typical set oftubulin polymerization curves without and with 4 mm(0.3 mg´mL21) PFK is presented in Fig. 5A. The bundlingactivity of PFK counteracts the inhibitory effect of both ions atsubstochiomeric concentrations; although Cu21 decreased theturbidity with respect to that measured with PFK but was stillsignificantly higher than the control (no PFK, no ions), thePFK-containing system appeared to be resistant to the CrO2±
4
evoked inhibition. When 0.3 or 0.6 mg´mL21 BSA or0.4 mg´mL21 yeast G-6-PDH were added to tubulin insteadof PFK, neither the control nor Cu21 inhibited polymerizationwere perturbed (data not shown).
As we demonstrated recently, the cytosolic fraction MPFCFabove 2 mg´mL21 concentration caused a substantial increasein the initial rate of turbidity and decreased the duration of thelag-phase without significantly influencing the final netincrease of turbidity [10] (Fig. 5B, curve d). This effect isentirely different from the effect caused by muscle PFK ontubulin polymerization (Fig. 5A, curve d). The addition ofMPFCF does not affect the inhibition by 50 mm CrO2±
4 , while
Fig. 6. Electron microscopic pictures of MT in
the absence and presence of Cu21 and CrO2±4
with or without cytosolic fraction. Microtubules
assembled in the absence (A±C) or presence
(D±F) of MPFCF without (A,D) or with addition
of Cu21 (B,E) or CrO2±4 (C,F) ions. Note the
decoration on MTs in (D±F) and the S-shaped
sheets (arrow) and oligomeric structures in
(B,C,E). Protofilaments curling away from the
end of MTs (arrowhead) is seen in (F).
Bar: 150 nm.
4736 K. Liliom et al. (Eur. J. Biochem. 267) q FEBS 2000

the inhibitory effect of 50 mm Cu21 is partially abolished(Fig. 5B).
In the next set of experiments the effects of the two bivalentions were investigated under more complex conditions, when acrude cytosolic fraction (CCF) was used. This preparation is amore physiological mixture of the cytosolic proteins as tubulinand MAP were not isolated from the extract, it containsMPFCF, tubulin and MAPs. In CCF the endogenous tubulinaccounts for about 15% of the total protein content as estimatedfrom SDS/PAGE patterns (data not shown). The effect of50 mm Cu21 or CrO2±
4 was investigated on the polymerizationof endogenous tubulin initiated by the addition of 20 mm taxolat two different total protein concentrations. At 3.4 mg´mL21
total protein concentration both ions reduced the polymeriza-tion of tubulin, although at 9 mg´mL21 total protein concentra-tion the addition of the ions was virtually ineffective (Fig. 5C).
Morphology of microtubules. To visualize the effects exerted by50 mm Cu21 or CrO2±
4 and/or cytosolic protein fractions at2 mg´mL21 total protein concentration on the structure of MT,an electron microscopy studies was performed. Randomlyarranged smooth-surfaced intact MT with a minimal amount ofoligomeric threads in the background were present in samplesassembled from purified MAP-free tubulin (Fig. 6A). Theaddition of either Cu21 or CrO2±
4 to tubulin resulted in theformation of large aggregates consisting of thread-likeoligomers (Fig. 6B,C, respectively). In addition, numerous`C'-shaped and `S'-shaped profiles, representing cross-sec-tioned microtubular sheets, cumulated in both samples(Fig. 6B,C). MTs assembled in the presence of MPFCF aredecorated by tiny filaments and globules representing MT-attached cytosolic proteins (Fig. 6D). When Cu21 or CrO2±
4
was added to samples also containing the cytosolic fraction, adifferent picture emerged. In the sample where Cu21 was addedtogether with MPFCF, decorated tubules can be visualizedtogether with oligomeric aggregates (Fig. 6E), while CrO2±
4
significantly damaged the tubules even in the presence of thiscytosolic fraction (Fig. 6F). Occasionally, filaments peelingaway from the tip of MTs were found, a phenomenon mainlyobserved in samples assembled in the presence of CrO2±
4 ions(Fig. 6F), and the amount of proteins attached to the surface ofMTs also seems to be reduced.
Flux of glycolysis. As discussed above, cytosolic proteins caninfluence the inhibition of tubulin polymerization caused byions in a rather complex manner and the effect depends on thenature of the ions. Recently, we presented evidence for theenhancement of the glycolytic flux in neuronal cell-free
extracts by MTs [10]. This effect counteracts the Cu21-inducedinhibition and seems to be specific as no stimulation ofglycolysis could be detected in muscle extracts under similarconditions. Tubulin (1 mg´mL21), which does not significantlyinfluence the upper phase of the glycolytic rate in MPFCF, isable to suspend the inhibition of the flux caused by 30 mmCu21, Cd21, or Hg21, which otherwise caused a nearlycomplete inhibition of the flux in the absence of tubulin(Table 2). To see whether the stimulating effect of tubulin onthe glycolytic flux can be preserved in the presence of the ions,the flux was measured in the presence of 30 mm ion plus3 mg´mL21 tubulin. This concentration of Cu21 was not able toreduce the tubulin-stimulated rate of glycolysis (Table 2).
D I S C U S S I O N
Some transition metal ions are essential for many metabolicprocesses and their homeostasis is crucial for life. Aberrationsin the cellular concentration of metal ions may lead to celldeath and severe diseases. Cells regulate the traffic of transitionmetal ions (such as copper), maintaining the amount necessaryfor biological function while avoiding excess levels, which aretoxic [17]. There is considerable interest in factors that controlthe concentration of transition metal ions inside the cell. Forexample, intracellular Cu21 is limited to less than one freecopper ion per cell [18]. A chaperone has been identified forcopper in humans and mice, which delivers copper to theantioxidant enzyme copper-zinc superoxide dismutase [17].
Many trace elements enter the cell by passive or activetransport mechanisms as well as by endocytosis. For example,Cd21 has an ion radius similar to that of Ca21, thus a part of itsuptake is mediated by voltage-dependent Ca21 channels [19].In another case, Cu21 can be taken up by a combination ofpassive and facilitated diffusion in intestinal cells, mediated bya specific copper carrier system [20]. Some trace elements arebound intracellularly to certain proteins, forming stable metal±protein complexes that involve sulfur atoms, amines or thecarboxyl groups of proteins. As the metals get into the cell,their chemical forms or availability are usually changed, thustheir intracellular level can be relatively high even if theirconcentration in the environment is low; some metals can beconcentrated by a factor of 1032105.
The principal target for many toxic metals are cells of thenervous system. Their filamentous structures can bind metalsdue to their high content of cysteine and methionine [2]. MTsare important and abundant subcellular components of neuronalcells as they organize the axonal vesicular traffic. Compoundsformed by metal ions reacting with two of the eight SH-groupsper tubulin [5], which are essential for MT assembly [6], blocktubulin polymerization in vitro [4]. We demonstrated that Cu21,Cd21, Hg21 and CrO2±
4 inhibit the tubulin polymerization at anIC50 of 14±24 mm with high cooperativity (Fig. 2) and alsoinduce microtubule disassembly. A plausible explanationfor these findings is that these active bivalent cations modifythe SH groups of tubulin involved in MT assembly [5,6]. TheCrO2±
4 anion could cause a similar effect acting indirectly byincreasing the oxygen reactive species. NO2
2 can producecomplete inhibition of tubulin polymerization; however, thiseffect occurs at a very high concentration (< 10 mm), and it isprobably exerted via nonspecific interactions between tubulinand this anion. Pb21, in contrast to the other bivalent cations,causes only a marginal inhibition of the polymerization up to aconcentration of 1 mm. According to previous studies [21],organic lead is a potent inhibitor of tubulin polymerization
Table 2. Preventative effect of tubulin on the inhibition of glycolytic
flux caused by bivalent metal ions. The rate of glucose conversion in
0.8 mg´mL21 microtubular protein-free cytosolic fraction (MPFCF) was
measured in the absence and presence of tubulin without and with 30 mm
bivalent ions after 2 h incubation time as described in Materials and
methods. The numbers are mean values of 3±5 determinations.
Normalized flux (%)
No ions 1 Cu21 1 Cd21 1 Hg21
MPFCF 100 1�.3 1�.0 1�.0
1 tubulin, 1 mg´mL21 112 111 104 98
1 tubulin, 3 mg´mL21 137 130 136 135
q FEBS 2000 Organization-dependent responses to stimuli (Eur. J. Biochem. 267) 4737

(300 mm trimethyl lead completely blocks the formation ofMT) while inorganic lead has only a minor effect.
We have determined the apparent dissociation constantsfor tubulin±metal ion complexes obtained by fluorescencequenching measurements (Fig. 2), which are in the micromolarconcentration range. Our data suggest, on the one hand, that theaffinities of the active ions to tubulin are similar, and on theother hand, that the apparent dissociation constants are inthe range where these ions significantly inhibit tubulinpolymerization. The BIAcore experiment (Fig. 3) suggestedthat tubulin in complex with Cu21 does not interact with theimmobilized tubulin. The mechanism responsible for theinhibitory effects could, therefore, be directly related tothe binding of the ion to tubulin.
It has been demonstrated that MAPs and cytosolic proteinssuch as glycolytic enzymes copolymerize with tubulin and theMTs formed are extensively decorated by macromolecules [22].The bundling activity of PFK is unique as the tetrameric PFKmolecules periodically cross-link the tubules, as visualized byelectron microscopy [23]. Comparison of the data presented inFig. 5 illustrates that the sensitivity of tubules for ions in thecombined system is different from that observed in theseparated system (purified tubulin alone), where they causedextensive inhibition. This organization-dependent effect oftoxic ions is similar to that observed previously withantimicrotubular agents [24], which suggests that the proteinshave a protective role on MT assembly. Obviously, the toxiceffect of ions can be reduced by increasing the concentration ofproteins, although the protecting effects obtained with differentcompositions of the proteins disprove this hypothesis. Forexample, 4 mm (0.3 mg´mL21) PFK, but not BSA or G-6-PDHin comparable concentrations, influenced the inhibitory effectof 50 mm Cu21. In the case of 50 mm Cu21, 2 mg´mL21 brainextract, which, in addition to neuronal cells, containspredominantly glial cells, exhibited a similar protecting effectto 0.25 mg´mL21 neuroblastoma extract [10]. These dataindicate that the toxic effect does depend on the superstructureof MTs created by the proteins present in the polymerizationassay. The electron microscopic images strengthened the viewthat both Cu21 and CrO2±
4 ions significantly reduced theformation of MT; mostly aberrant oligomers of differentmorphology are formed, although cytosolic protein couldabolish, at least partly, the toxic effect of ions by associatingwith the tubules.
Copper also affects the cell energy production by decreasingthe glycolytic rate in both brain and muscle cytosolic fraction[1,9,25]. We have recently reported that 50 mm Cu21 decreasesthe flux of the upper phase of glycolysis in the cytosolicfractions prepared from brain and muscle [9,10]. Now we findthat the addition of Cu21, Cd21, and Hg21, but not Pb21,CrO2±
4 , or NO22, to the brain cytosolic fraction completely
inhibited the flux of glucose conversion to triosephosphates byIC50 of 10±14 mm with high cooperativity (Table 1). Tubulindiminishes the inhibitory effect of the cations. This effect seemsto be specific as bovine serum albumin at similar conditionswas much less effective (data not shown). As under similarconditions the tubulin polymerization was also protected bycytosolic proteins against copper inhibition (Fig. 5), a possibleexplanation for these findings is that the binding sites of theions causing inactivation of the glycolytic enzymes are blockedby tubulin in the combined system.
Reliable estimates of the free concentration of transitionmetal ions other than copper in cells are not available yet.Uncontrolled transition metal ion concentrations in cells havebeen associated with several human diseases including
neurological disorders, Parkinson's disease and Wilson'sdisease [25,26]. Current studies into the mechanisms ofcommon neurodegenerative diseases suggest that copper-uptakeinto neuronal cells may be mediated by amyloid precursorprotein [27] and prion protein [28], which are directly linked toAlzheimer's and Creutzfeldt±Jacob's diseases, respectively.
Copper-dependent enzymes require accessory factors tocompete with chelators and processes that sequester essentiallyall intracellular free copper. On the other hand, elements ofprotein structures or metabolic pathways could be damaged bycomplex formation or oxidation processes if crucial metalbinding sites were available for the free or donor-deliveredmetals. It has been suggested that copper chaperones protectcells against copper toxicity [18]. Here we suggest that apossible protection mechanism could be the formation oforganized structures. We did observe protective effects on MTassembly and glycolytic flux against bivalent ions of themicrotubular superstructure formed in complex protein sys-tems. The protecting effect appeared to be specific, dependingon the particular proteins in the system, the complexity of thesystem as well as on the chemical nature of the ions. Some ionswere toxic only in the separated system, while others were toxicin both the separated and the combined systems. The protectingeffect of structural origin might be due to reduced availabilityof binding sites on tubulin and on glycolytic enzymes inorganized states; the reactive ions could thus be sequestered byother sites. In this respect, the function of the organizedstructure is just the opposite to that suggested for chaperones,which direct the metal to the target enzyme. This issue might beimportant in understanding how brain cells function even athigh total concentrations of copper (80±100 mm [29]), or in thecases of other ions that are `toxic' in test tubes, wherecomponents are separated. It might also help in understandingthe pathological features of diseases caused by the uncontrolledor elevated concentrations of pollutants in cells, specifically inneuronal cells.
A C K N O W L E D G E M E N T S
The excellent technical contribution of Emma Hlavanda and Mimi
NuridsaÂny is gratefully acknowledged. This work was supported by grants
from the Hungarian National Science Foundation OTKA (T-025291 and
T-031892 to J. O. and T-2227 to J. K.), the Spanish Government Grant,
BIO98±0365 from DGICYT to M. C., and from the European Community
(Grant ERBIC15CT960307 to J. O. and M. C.). K. L. receives the `Bolyai'
fellowship of the Hungarian Academy of Sciences.
R E F E R E N C E S
1. Camakaris, J., Voskoboinik, I. & Mercer, J.F. (1999) Molecular
mechanism of copper homeostasis. Biochem. Biophys. Res. Commun.
261, 225±232.
2. Clarkson, T.W. (1987) Metal toxicity in the central nervous system.
Environ. Health. Persp. 75, 59±64.
3. Costa, M. (1997) Toxicity and carcinogenicity of Cr (VI) in animal
models and humans. Crit. Rev. Toxicol. 27, 431±442.
4. Kuriyama, R. & Sakai, H. (1974) Role of tubulin-SH groups in
polymerization to microtubules. Functional SH groups in tubulin for
polymerization. J. Biochem. 76, 651±654.
5. Wallin, M., Larsson, H. & EdstroÈm, A. (1977) Tubulin sulfhydryl
groups and polymerization in vitro. Effects of di- and trivalent
cations. Exp. Cell Res. 107, 219±225.
6. Arai, T., Ihara, Y., Arai, K.I. & Kaziro, Y. (1975) Purification of
tubulin from bovine brain and its interaction with guanine
nucleotides. J. Biochem. 77, 647±658.
4738 K. Liliom et al. (Eur. J. Biochem. 267) q FEBS 2000

7. Larsson, H., Wallin, M. & EdstroÈm, A. (1976) Induction of a sheet
polymer of tubulin by Zn21. Exp. Cell Res. 100, 104±110.
8. Lai, J.C.K. & Blass, J.P. (1984) Neurotoxic effects of copper:
Inhibition of glycolysis and glycolytic enzymes. Neurochem. Res.
9, 1699±1710.
9. Jannaschik, D., Burgos, M., Centelles, J.J., OvaÂdi, J. & Cascante, M.
(1999) Application of metabolic control analysis to the study of toxic
effects of copper in muscle extracts. FEBS Lett. 445, 144±148.
10. Liliom, K., WaÂgner, G., KovaÂcs, J., Comin, B., Cascante, M., Orosz, F.
& OvaÂdi, J. (1999) Combined enhancement of microtubule assembly
and glucose metabolism in neuronal systems in vitro. Decreased
sensitivity to copper toxicity. Biochem. Biophys. Res. Commun. 264,
605±610.
11. Bradford, M.M. (1976) A rapid and sensitive method for the
quantitation of micogram quantities of protein utilizing the principle
of protein-dye binding. Anal. Biochem. 72, 248±254.
12. Na, C.N. & Timasheff, S.N. (1986) Interaction of vinblastine with calf
brain tubulin: multiple equilibria. Biochemistry 25, 6214±6222.
13. Vallee, R.B. (1986) Reversible assembly purification of microtubules
without assembly-promoting agents and further purification of
tubulin, microtubule-associated proteins and MAP fragments.
Methods Enzymol. 134, 89±127.
14. Gaskin, F., Cantor, C.R. & Shelanski, M.L. (1974) Turbidimetric
studies of the in vitro assembly and disassembly of procine
neurotubules. J. Mol. Biol. 89, 737±758.
15. Mertens, M.C. & KaÈgi, J.H. (1979) A graphical correction procedure
for inner filter effect in fluorescence quenching titrations. Anal.
Biochem. 96, 448±455.
16. Fell, D. (1997) Understanding the control of metabolism. In Frontiers
in Metabolism, Vol. 2 (Snell, K., ed.) Portland Press, London, UK.
17. Lippard, S.J. (1999) Free copper ions in the cell? Science 284,
748±749.
18. Rae, T.D., Schmidt, P.J., Pufahl, R.A., Culotta, V.C. & O'Halloran, T.V.
(1999) Undetectable intracellular free copper: the requirement of a
copper chaperone for superoxide dimutase. Science 284, 805±808.
19. Hinkle, P.M., Shanshala, I.E.D. & Nelson, E.J. (1992) Measurement of
intracellular cadmium with fluorescent dyes. J. Biol. Chem. 267,
25553±25559.
20. Wang, Z., Ward, W.E., Lonnerdal, B. & Atkinson, S.A. (1993) Copper
uptake and its interaction with zinc and iron in intestinal brush-border
membrane-vesicles (Bbmv) in the developing piglet. FASEB J. 7,
A299.
21. Roderer, G. & Doenges, K.H. (1983) Influence of trimethyl lead and
inorganic lead on the in vitro assembly of microtubules from
mammalian brain. Neurotoxicology 4, 171±180.
22. Orosz, F., WaÂgner, G., Liliom, K., KovaÂcs, J., BaroÂti, K., HoraÂnyi, M.,
Farkas, T., HollaÂn, S. & OvaÂdi, J. (2000) Enhanced association of
mutant triosephosphate isomerase to red cell membranes and to brain
microtubules. Proc. Natl Acad. Sci. USA 97, 1026±1031.
23. Lehotzky, A., PaÂlfia, Z., KovaÂcs, J., MolnaÂr, A. & OvaÂdi, J. (1994)
Ligand-modulated cross-bridging of microtubules by phos-
phofructokinase. Biochem. Biophys. Res. Comm. 204, 585±591.
24. Orosz, F., KovaÂcs, J., LoÃw, P., VeÂrtessy, B.G., UrbaÂnyi, Z., AÂ cs, T.,
Keve, T. & OvaÂdi, J. (1997) Interaction of a new bis-indol derivative,
KAR-2 with tubulin and its antimitotic activity. Br. J. Pharmacol. 21,
947±954.
25. Nelson, N. (1999) Metal ion transporters and homeostasis. EMBO J.
18, 4361±4371.
26. Radisky, D. & Kaplan, J. (1999) Regulation of transition metal
transport across the yeast plasma membrane. J. Biol. Chem. 274,
4481.
27. Multhaup, G., Schlicksupp, A., Hesse, L., Beher, D., Ruppert, T.,
Masters, C.L. & Beyreuther, K. (1996) The amyloid precursor protein
of Alzheimer's disease in the reduction of copper (II) to copper.
Science 271, 1406±1409.
28. Prince, R.C. & Gunson, D.E. (1998) Prions are copper-binding
proteins. Trends Biochem. Sci. 23, 197±198.
29. Soto, M. & MarigoÂmez, I. (1995) Cell Biology in Environmental
Toxicology. (Cajaraville, M.P., ed.). University of the Basque
Country, Bilbao, Spain.
q FEBS 2000 Organization-dependent responses to stimuli (Eur. J. Biochem. 267) 4739




















