
On the signals underlying conscious awareness of action
9
On the signals underlying conscious awareness of action Sukhvinder S. Obhi a,b, * , Peggy J. Planetta a,b , Jordan Scantlebury c a Cognition in Action Laboratory, Centre for Cognitive Neuroscience, Wilfrid Laurier University, Waterloo, Ontario, Canada b Department of Psychology, Wilfrid Laurier University, Waterloo, Ontario, Canada c Department of Psychology, Dalhousie University, Halifax, Nova Scotia, Canada article info Article history: Received 8 February 2008 Revised 12 October 2008 Accepted 13 October 2008 Available online xxxx Keywords: Conscious awareness of Action Efference copy Reafference Motor timing Movement timing Perception of action Agency Awareness Consciousness abstract To investigate whether conscious judgments of movement onset are based solely on pre- movement signals (i.e., premotor or efference copy signals) or whether sensory feedback (i.e., reafferent) signals also play a role, participants judged the onset of finger and toe movements that were either active (i.e., self initiated) or passive (i.e., initiated by the experimenter). Conscious judgments were made by reporting the position of a rotating clock hand presented on a computer screen and were then compared to the actual mea- sured time of movement onset. In line with previous studies, judgment errors were found to be anticipatory for both finger and toe movements. There was a significant difference between judgment errors for active and passive movements, with judgments of active movements being more anticipatory than judgments of passive movements. This is consis- tent with a pre-movement (from here on referred to as an ‘‘efferent”) account of action awareness because premotor and efference copy signals are only present in active move- ments, whereas the main source of movement information in passive movements is sen- sory feedback which is subject to time delays of conduction (and hence predicts later judgment times for passive movements). However, judgments of active toe movement onset time were less anticipatory than judgments of active finger movement onset time. This pattern of results is not consistent with a pure efferent account of conscious aware- ness of action onset – as this account predicts more anticipatory judgments for toe move- ments compared to finger movements. Instead, the data support the idea that conscious judgments of movement onset are based on efferent (i.e., premotor, efference copy) and reafferent (i.e., feedback from the movement) components. Ó 2008 Elsevier B.V. All rights reserved. 1. Introduction The study of consciousness has taken centre stage in contemporary experimental psychology and cognitive neu- roscience (e.g., Arshavsky, 2006; Baars, 2005; Dehaene & Naccache, 2001; Gazzaniga, 1998; Haggard, 2005; Maia & Cleeremans, 2005; Mandler, 2005; Zeman, 2001, 2006). One important general question relates to the contents of consciousness, and many studies have been aimed at addressing this issue (see Rees, 2007). In the domain of hu- man action, the contents of consciousness are strongly tied to the subjective experience of acting. What aspects of our actions are we aware of and what is the relationship of these aspects to the workings of the central nervous sys- tem? The classic study of Libet, Gleason, Wright, and Pearl (1983) sparked numerous investigations, most of which have attempted to further understand the relationship be- tween the temporal sequence of neural events and the associated temporal sequence of conscious experience of action (e.g., Franck, Psada, Pichon & Haggard, 2005; Hag- gard, Clark, & Kalogeras, 2002; Haggard & Eimer, 1999; Haggard, Newman, & Magno, 1999; Libet, 1985, 1996). This 0010-0277/$ - see front matter Ó 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.cognition.2008.10.009 * Corresponding author. Address: Room N2023, Department of Psy- chology, Wilfrid Laurier University, 75 University Avenue West, Waterloo, Ontario, Canada N2L 3C5. Tel.: +1 519 884 0710x2423; fax: +1 519 746 7605. E-mail address: [email protected] (S.S. Obhi). Cognition xxx (2008) xxx–xxx Contents lists available at ScienceDirect Cognition journal homepage: www.elsevier.com/locate/COGNIT ARTICLE IN PRESS Please cite this article in press as: Obhi, S. S., et al. On the signals underlying conscious awareness of action. Cognition (2008), doi:10.1016/j.cognition.2008.10.009
Transcript of On the signals underlying conscious awareness of action

Cognition xxx (2008) xxx–xxx
ARTICLE IN PRESS
Contents lists available at ScienceDirect
Cognition
journal homepage: www.elsevier .com/locate /COGNIT
On the signals underlying conscious awareness of action
Sukhvinder S. Obhi a,b,*, Peggy J. Planetta a,b, Jordan Scantlebury c
a Cognition in Action Laboratory, Centre for Cognitive Neuroscience, Wilfrid Laurier University, Waterloo, Ontario, Canadab Department of Psychology, Wilfrid Laurier University, Waterloo, Ontario, Canadac Department of Psychology, Dalhousie University, Halifax, Nova Scotia, Canada
a r t i c l e i n f o
Article history:Received 8 February 2008Revised 12 October 2008Accepted 13 October 2008Available online xxxx
Keywords:Conscious awareness of ActionEfference copyReafferenceMotor timingMovement timingPerception of actionAgencyAwarenessConsciousness
0010-0277/$ - see front matter � 2008 Elsevier B.Vdoi:10.1016/j.cognition.2008.10.009
* Corresponding author. Address: Room N2023,chology, Wilfrid Laurier University, 75 University AvOntario, Canada N2L 3C5. Tel.: +1 519 884 0710x247605.
E-mail address: [email protected] (S.S. Obhi).
Please cite this article in press as: Obhi(2008), doi:10.1016/j.cognition.2008.10.0
a b s t r a c t
To investigate whether conscious judgments of movement onset are based solely on pre-movement signals (i.e., premotor or efference copy signals) or whether sensory feedback(i.e., reafferent) signals also play a role, participants judged the onset of finger and toemovements that were either active (i.e., self initiated) or passive (i.e., initiated by theexperimenter). Conscious judgments were made by reporting the position of a rotatingclock hand presented on a computer screen and were then compared to the actual mea-sured time of movement onset. In line with previous studies, judgment errors were foundto be anticipatory for both finger and toe movements. There was a significant differencebetween judgment errors for active and passive movements, with judgments of activemovements being more anticipatory than judgments of passive movements. This is consis-tent with a pre-movement (from here on referred to as an ‘‘efferent”) account of actionawareness because premotor and efference copy signals are only present in active move-ments, whereas the main source of movement information in passive movements is sen-sory feedback which is subject to time delays of conduction (and hence predicts laterjudgment times for passive movements). However, judgments of active toe movementonset time were less anticipatory than judgments of active finger movement onset time.This pattern of results is not consistent with a pure efferent account of conscious aware-ness of action onset – as this account predicts more anticipatory judgments for toe move-ments compared to finger movements. Instead, the data support the idea that consciousjudgments of movement onset are based on efferent (i.e., premotor, efference copy) andreafferent (i.e., feedback from the movement) components.
� 2008 Elsevier B.V. All rights reserved.
1. Introduction
The study of consciousness has taken centre stage incontemporary experimental psychology and cognitive neu-roscience (e.g., Arshavsky, 2006; Baars, 2005; Dehaene &Naccache, 2001; Gazzaniga, 1998; Haggard, 2005; Maia &Cleeremans, 2005; Mandler, 2005; Zeman, 2001, 2006).One important general question relates to the contents of
. All rights reserved.
Department of Psy-enue West, Waterloo,
23; fax: +1 519 746
, S. S., et al. On the sig09
consciousness, and many studies have been aimed ataddressing this issue (see Rees, 2007). In the domain of hu-man action, the contents of consciousness are strongly tiedto the subjective experience of acting. What aspects of ouractions are we aware of and what is the relationship ofthese aspects to the workings of the central nervous sys-tem? The classic study of Libet, Gleason, Wright, and Pearl(1983) sparked numerous investigations, most of whichhave attempted to further understand the relationship be-tween the temporal sequence of neural events and theassociated temporal sequence of conscious experience ofaction (e.g., Franck, Psada, Pichon & Haggard, 2005; Hag-gard, Clark, & Kalogeras, 2002; Haggard & Eimer, 1999;Haggard, Newman, & Magno, 1999; Libet,1985, 1996). This
nals underlying conscious awareness of action. Cognition

2 S.S. Obhi et al. / Cognition xxx (2008) xxx–xxx
ARTICLE IN PRESS
experiential sequence includes ‘early’ high level processessuch as intention as well as ‘later’ processes such as theperception of the consequences of action. Libet et al.’s(1983) study revealed the controversial finding that con-scious experience of the intention to act occurred afterthe onset of neural events associated with response prepa-ration, but early enough for the action to be inhibited ifnecessary. This finding provoked a great deal of interestfrom psychologists and philosophers alike because itthrows into question the Cartesian notion of a mind-bodydirection of causation. Another, perhaps less controversialfinding from the study was that the conscious experienceof actually moving occurred before movement onset asmeasured using electromyography (EMG). In the originalstudy participants were approximately 90 ms early in theirjudgments of when they moved (i.e., they judged them-selves as having moved approximately 90 ms before theyreally did).
It must be noted that the methodology used and conclu-sions drawn in Libet style experiments have been the sub-ject of much scrutiny. Indeed, several published criticismshave been leveled at them (e.g., Breitmeyer, 2002; Pockett,2002a,b). In the context of Libet style experiments on mo-tor awareness most of these criticisms have been dealtwith previously and the interested reader is referred to thiswork (e.g., Haggard & Libet, 2001; Haggard et al., 1999;Obhi, 2007). Suffice to say, whereas the validity of absolutemeasured times in Libet experiments is indeed open toquestion, patterns of differences between timing measuresobtained in conditions that are all equally subject to poten-tial methodological biases can usefully inform theories ofaction awareness (e.g., Haggard et al., 1999).
The anticipatory awareness of action results have beenreplicated several times and has been generally explainedwith reference to pre-movement processes (Blakemore,Wolpert, & Frith, 2002; Frith, Blakemore, & Wolpert,2000; Haggard & Eimer, 1999; Haggard et al., 1999). Forexample, Blakemore et al. (2002) have argued that thefunctioning of an internal forward model which uses effer-ence copy to predict the sensory consequences of an actionprior to its execution can account for the finding. By thisview, what we are aware of when we move is not the ac-tion itself but a prediction of the sensory consequencesof the action (Wolpert & Ghahramani, 2000; Wolpert &Miall, 1996). This explanation and other forms of premotorexplanations will be referred to as the ‘‘efferent account” ofaction awareness for the remainder of this paper.
Despite the appeal of an efferent account of actionawareness, to date, there has been very little empiricalsupport to substantiate this assertion. Indeed, (to ourknowledge) the only experiment to provide empirical sup-port for any type of premotor account of action awarenessin Libet type paradigms was performed by Haggard et al.(1999). These authors found that judgments of action onsetwere affected by the length of a typed sequence such thatjudgments became more anticipatory as the length of thesequence increased. They interpreted their results in thecontext of Sternberg’s model of action generation (1978)and asserted that awareness of action arises from somepremotor process that occurs prior to the dispatch of motorcommands for the response (i.e., these authors differenti-
Please cite this article in press as: Obhi, S. S., et al. On the sig(2008), doi:10.1016/j.cognition.2008.10.009
ate between an efference copy hypothesis and a premotorhypothesis on the basis that efference copy is only avail-able after the selection of motor commands). In spite ofthis elegant result, which does support a role for premotorprocesses in the generation of action awareness, experi-ments which directly investigate the possible role of sen-sory feedback in conscious judgments of action onset inLibet tasks are lacking. Instead, an assumption that sensoryfeedback is not involved seems to have been made. Thus,the theoretical assertion that judgments of action onsetare based solely on conscious access to preparatory motorstates has gained popular support, without empiricalvalidation.
However, in a recent study Obhi (2007) reported find-ings that seemed to call into question this proposed mech-anism. In the critical conditions of the study, participantsmade key press actions that were either active (i.e., gener-ated by the participant) or passive (i.e., generated by theexperimenter pushing the participants finger down ontoa key at an unpredictable time). These manipulations werecarried out in a Libet style experiment in which partici-pants judged the onset times of the action by referring tothe position of a clock hand rotating on a screen in frontof them (see Libet et al., 1983 and Haggard et al., 1999for more details on this methodology). Firstly, the resultsconfirmed the previously reported anticipatory awarenessof action - and this was the case for active and passivemovements. However, there was absolutely no differencebetween the judgments for active versus passive move-ments. Because passive movements are not preceded bypremotor processing (and they do not generate efferencecopy), and active and passive movements produced similarjudgments, it seems plausible that the judgments were bebased on a signal that is common to both scenarios, namelya sensory feedback signal. In view of these findings it wassuggested that anticipatory awareness of action cannot so-lely be linked to pre-movement processes such as the pre-diction of the sensory consequences of a to-be-executedaction.
Thus, movement related feedback may play more of arole in our conscious experience of action than previouslyacknowledged. It might be that the anticipatory judgmentsare the result of a reconstructive process that uses patternsof movement related feedback to estimate when the move-ment occurred. Generalizations of this idea should bemade with caution though, since there are several poten-tial signals that might underlie conscious judgments, andthe combination of signals used may depend on the pre-vailing task-operator characteristics. For example, in a re-cent study it was found that deafferented patientsexhibited anticipatory timing of action in a synchroniza-tion task in which finger taps must be made in time withisochronous beeps (Stenneken, Prinz, Cole, Paillard, &Aschersleben, 2006). Hence, for these individuals, a pre-movement predictive signal may have been the only viablesource of information about when a movement started.
Despite the apparent support for the role of feedback inconscious judgments of action onset in the study of Obhi(2007), there was one problem with their paradigm whichcould have produced the observed pattern of results for ac-tive and passive movements, thus rendering them some-
nals underlying conscious awareness of action. Cognition

S.S. Obhi et al. / Cognition xxx (2008) xxx–xxx 3
ARTICLE IN PRESS
what inconclusive. Specifically, in that experiment, theexperimenter only ever touched the participant’s fingerin the passive movement conditions. Participants sat withtheir finger resting lightly on a key and at an unpredictabletime the experimenter made contact with their finger andpushed it down to initiate a key press. This was not thecase in the active movement condition, where the experi-menter never touched the participant’s finger. Hence, thereis the very real possibility that a premotor (anticipatory)awareness effect in the active movement condition wassimply replaced with a temporally similar effect based onthe tactile sensation of the experimenter’s touch in thepassive condition. That is, participants may have used theonset of experimenter touch to judge the time of the move-ment in the passive condition. In reality, there is no way toexclude this possibility without matching the potential ef-fects of experimenter touch across the active and passiveconditions.
The present study was designed to further elucidate thesignals that contribute to the conscious awareness of ac-tion. Specifically, we sought to determine whether con-scious awareness of action does indeed depend solely onpremotor, efferent signals, or whether feedback signalsalso play a role, at least in Libetian paradigms. To thisend, we conducted a careful experiment similar to that ofObhi (2007) in which participants made active and passivemovements and judged their onsets using a Libet set-up.However, in contrast to our previous study, we manipu-lated conduction length by requiring subjects to make fin-ger movements in one condition and toe movements inanother. Crucially, we also made sure that experimentertouch was equalized across the experimental conditionsby ensuring that the experimenter maintained light con-tact with the participant’s finger/toe in both active andpassive conditions. Using such a set-up, some specific pre-dictions are possible based on whether the signals used tomake the judgment are mostly efferent or mostly reaffer-ent/feedback related in nature. It has previously been dem-onstrated that in simple reaction time tasks, responses ofthe finger are faster than responses of the foot (Stenneken,Aschersleben, Cole, & Prinz, 2002; Bard, Paillard, Teasdale,Fleury, & Lajoie, 1991; Billon, Bard, Fleury, Blouin & Tease-dale, 1996). This simple reaction time difference is thoughtto be due to the difference in motor cortex-finger and mo-tor cortex-foot conduction length, and hence is explainedby an efferent signal conduction account. Of relevance isthat estimates of conduction velocity of the motor nervesinnervating the hand and foot have been found to be inthe 40–60 m/s range with lower conduction velocitiesfor the nerves innervating foot muscles (e.g., Thomas,Sears, and Gilliatt, 1959). Estimates of muscle afferentconduction velocity have been found to be of the order of58 m/s (Shefner & Logigian, 1994).
In the present study this ‘efferent’ account can only ap-ply to the active movement conditions and predicts thatsince motor preparation for a toe movement made at a par-ticular time begins earlier than preparation for a fingermovement, the judgment for the toe movement might bemore anticipatory than the judgment for the finger move-ment. On the other hand, it has also previously been shownthat, when subjects attempt to produce simultaneous fin-
Please cite this article in press as: Obhi, S. S., et al. On the sig(2008), doi:10.1016/j.cognition.2008.10.009
ger and foot movements in a self initiated manner, footmovements must precede finger movements in order fortemporal simultaneity to be perceived (Bard et al., 1991;Billon, Bard, Fleury, & Blouin, 1996; Stenneken, Aschersle-ben, Cole, & Prinz, 2002). This finding is thought to be dueto the fact that the judged timing of self produced move-ments is based on somatosensory feedback representedat the central level, and such feedback takes longer to reachthe brain from the foot than from the finger. In the presentstudy, such a ‘feedback’ account might apply to the passivemovement conditions (and possibly also the active move-ment conditions) and predicts that the toe movement con-dition might yield later judgments than the fingermovement condition. These predictions are outlined sche-matically in Fig. 1. Of course, in active movement condi-tions, there is a possibility that efferent signals andfeedback signals are both used for awareness of onset judg-ments. Such combining of feedback signals with efferentsignals would also be expected to affect patterns of judg-ment error. Specifically, the addition of a feedback compo-nent to the signal used to form a conscious percept ofmovement onset might in general delay the percept, witha greater delay for toe movements than for finger move-ments (due to the increased time required for feedbackto reach the brain from the toe compared to the finger).
2. Methods
2.1. Participants
Twenty four naïve volunteers (9 males, 15 females;mean age, 21 years) participated in exchange for coursecredit or payment. All participants provided written in-formed consent, and reported being right-handed and freeof bone, joint, and muscle problems. The procedures in thisexperiment were approved by the Research Ethics Board atWilfrid Laurier University.
2.2. Apparatus
The experiment was programmed using DirectRT soft-ware (Empirisoft Corp., NY) and run on a Dell laptop com-puter. Responses were made on the DirectIN Response BoxSystem (Empirisoft Corp., NY), which interfaced with thecomputer via the USB port. The temporal precision of themeasured response times was 1 ms.
To measure the force with which responses were made,a force sensitive resistor (FSR) was adhered to the top sur-face of the response button, and was connected to a Bagn-oli-8 EMG System (Delsys Inc., Boston, MA). The EMGsystem interfaced with a second Dell laptop computer viaa National Instruments data acquisition card. EMGworks3.1 signal acquisition software (Delsys Inc., Boston, MA)sampled the data at a frequency of 1000 Hz.
2.3. Design and procedure
The experiment consisted of four conditions (active-fin-ger, passive finger, active toe, passive-toe), each presentedin blocks of 30 trials. The blocks were presented in a pseu-
nals underlying conscious awareness of action. Cognition
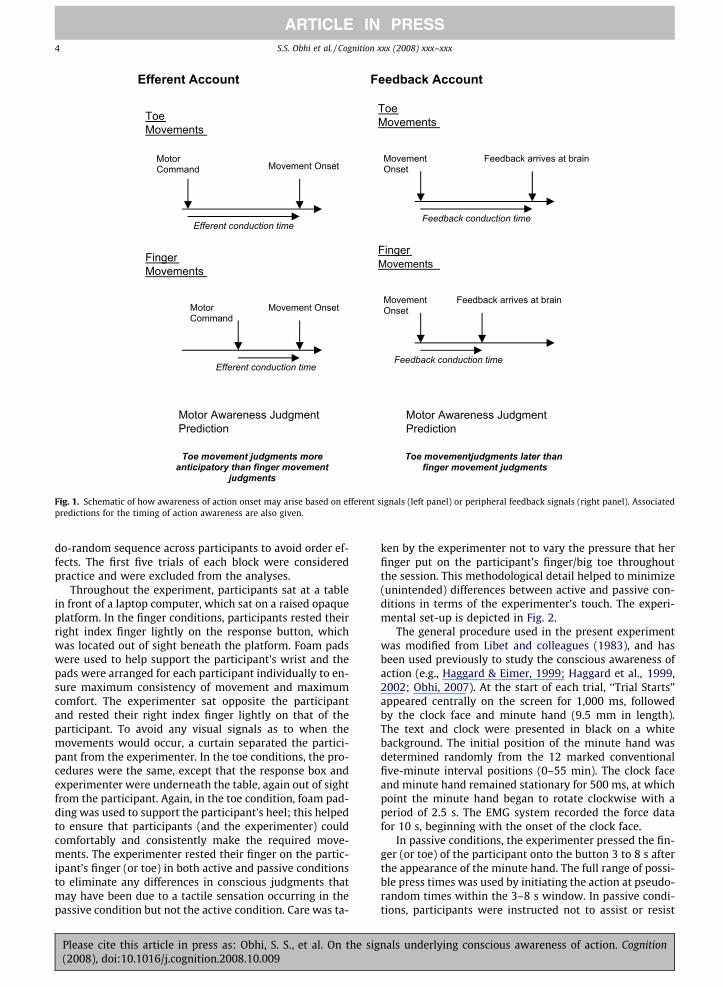
Efferent Account Feedback Account
Feedback arrives at brain Movement Onset
Toe Movements
Feedback arrives at brain
Finger Movements
Feedback conduction time
Feedback conduction time
Movement Onset
Toe movementjudgments later than finger movement judgments
Motor Awareness Judgment Prediction
Motor Command Movement Onset
Toe Movements
Movement Onset
Finger Movements
Efferent conduction time
Efferent conduction time
Motor Command
Toe movement judgments more anticipatory than finger movement
judgments
Motor Awareness Judgment Prediction
Fig. 1. Schematic of how awareness of action onset may arise based on efferent signals (left panel) or peripheral feedback signals (right panel). Associatedpredictions for the timing of action awareness are also given.
4 S.S. Obhi et al. / Cognition xxx (2008) xxx–xxx
ARTICLE IN PRESS
do-random sequence across participants to avoid order ef-fects. The first five trials of each block were consideredpractice and were excluded from the analyses.
Throughout the experiment, participants sat at a tablein front of a laptop computer, which sat on a raised opaqueplatform. In the finger conditions, participants rested theirright index finger lightly on the response button, whichwas located out of sight beneath the platform. Foam padswere used to help support the participant’s wrist and thepads were arranged for each participant individually to en-sure maximum consistency of movement and maximumcomfort. The experimenter sat opposite the participantand rested their right index finger lightly on that of theparticipant. To avoid any visual signals as to when themovements would occur, a curtain separated the partici-pant from the experimenter. In the toe conditions, the pro-cedures were the same, except that the response box andexperimenter were underneath the table, again out of sightfrom the participant. Again, in the toe condition, foam pad-ding was used to support the participant’s heel; this helpedto ensure that participants (and the experimenter) couldcomfortably and consistently make the required move-ments. The experimenter rested their finger on the partic-ipant’s finger (or toe) in both active and passive conditionsto eliminate any differences in conscious judgments thatmay have been due to a tactile sensation occurring in thepassive condition but not the active condition. Care was ta-
Please cite this article in press as: Obhi, S. S., et al. On the sig(2008), doi:10.1016/j.cognition.2008.10.009
ken by the experimenter not to vary the pressure that herfinger put on the participant’s finger/big toe throughoutthe session. This methodological detail helped to minimize(unintended) differences between active and passive con-ditions in terms of the experimenter’s touch. The experi-mental set-up is depicted in Fig. 2.
The general procedure used in the present experimentwas modified from Libet and colleagues (1983), and hasbeen used previously to study the conscious awareness ofaction (e.g., Haggard & Eimer, 1999; Haggard et al., 1999,2002; Obhi, 2007). At the start of each trial, ‘‘Trial Starts”appeared centrally on the screen for 1,000 ms, followedby the clock face and minute hand (9.5 mm in length).The text and clock were presented in black on a whitebackground. The initial position of the minute hand wasdetermined randomly from the 12 marked conventionalfive-minute interval positions (0–55 min). The clock faceand minute hand remained stationary for 500 ms, at whichpoint the minute hand began to rotate clockwise with aperiod of 2.5 s. The EMG system recorded the force datafor 10 s, beginning with the onset of the clock face.
In passive conditions, the experimenter pressed the fin-ger (or toe) of the participant onto the button 3 to 8 s afterthe appearance of the minute hand. The full range of possi-ble press times was used by initiating the action at pseudo-random times within the 3–8 s window. In passive condi-tions, participants were instructed not to assist or resist
nals underlying conscious awareness of action. Cognition

Fig. 2. The Experimental set-up. Note that this schematic depicts thepassive finger movement condition in which the experimenter initiatesthe participant’s finger movement by pushing their finger to depress abutton. The participant cannot see the experimenter’s face because thereis a curtain placed between them and they cannot see the experimenter’sfinger because all movements are made under an opaque platform whichoccludes participants’ vision of the set-up. In passive toe movements, theexperimenter was under the table instead of seated on a chair andinitiated passive movements by pushing the participant’s toe down todepress a button. Again, the participant was unable to see the experi-menter’s face or finger in this condition. Active movements used the sameset-up, with the experimenter gently resting their finger on the partic-ipant’s finger or toe, and taking care not to assist the participant’s selfinitiated movements.
S.S. Obhi et al. / Cognition xxx (2008) xxx–xxx 5
ARTICLE IN PRESS
the presses initiated by the experimenter. In active condi-tions, the participants pressed the button themselves usingtheir finger or toe, again at a random time (this time of theirown choosing) within the 3–8 s window after the appear-ance of the minute hand. Participants were instructed toavoid initiating button presses at predetermined or stereo-typed times and to avoid intentionally timing buttonpresses to coincide with particular positions of the clockhand. Additionally, participants were instructed to pressthe button by flexing only their finger or toe and wereexplicitly requested to avoid flexing their entire hand orfoot. When necessary, if the participant flexed more thantheir finger or toe (i.e., exhibited whole hand or foot flex-ion), the experimenter issued a reminder at the end of thetrial that they should attempt to move only their finger ortoe. Following each trial, participants verbally reportedthe position of the clock hand at the instant they startedmoving. The subsequent trial was initiated by a key presswhen the experimenter and participant were ready.
To familiarize themselves with the task, prior to theexperiment proper, the participant initiated several buttonpresses with their right index finger and right big toe. Theyalso practiced obtaining similar levels of force and durationin finger and toe movements. During this procedure theexperimenter also initiated several passive button presses
Please cite this article in press as: Obhi, S. S., et al. On the sig(2008), doi:10.1016/j.cognition.2008.10.009
to practice obtaining similar force and duration data asthe participant. This pre-experiment session was impor-tant as it helped to minimize any differences in the physi-cal characteristics of the finger and toe responses andbetween the active and passive conditions. Throughoutthe experiment, care was taken to equalize the nature offinger and toe movements in active and passive conditions,especially with regard to the degree of whole hand or footflexion that occurred. On the whole participants and theexperimenter were successful at minimizing differencesbetween active and passive conditions in the physicalmovements.
2.4. Data analysis
2.4.1. Errors and outliersTrials in which the button was not pressed or pressed
more than once were excluded from the analyses, as weretrials in which the judgment error exceeded 300 ms (or 7‘‘minutes” on the clock).
2.4.2. Judgment error dataJudgment error was calculated for each trial as the dif-
ference between the perceived onset of movement and theactual onset of movement. As such, negative values repre-sent anticipatory judgments of movement and positive val-ues represent delayed judgments of movement. In eachcondition, the mean judgment error (in ms) was calculatedfor each participant.
2.4.3. Force dataEMGworks 3.1 software (Delsys, Inc., Boston, MA) was
used to process the FSR data. For each condition, for eachtrial the peak to peak amplitude of the FSR signal wasdetermined as a quantification of the force of the buttonpress in volts (V). In addition to the peak amplitude data,we also determined the duration of each key press by cal-culating the difference between the onset and offset timeof the FSR trace. Force data were not converted from Voltsto Newtons as the force measurements in this experimentwere not the main dependent measures of interest, andserved only as a means to compare potential confoundingeffects of force differences across conditions.
3. Results
Data from one subject were excluded due to the factthat this participant failed to follow the experimenter’sinstructions. The application of the outlier exclusion crite-rion resulted in the removal of 3.5% of the data.
3.1. Judgment error data
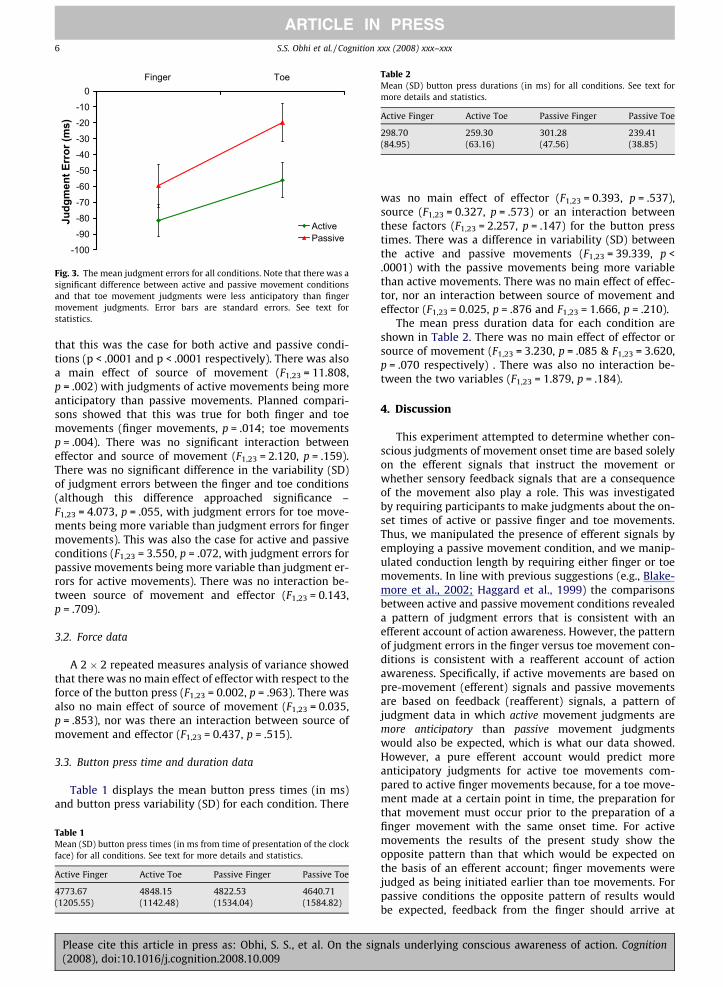
Mean judgment errors (in ms) for each condition areshown in Fig. 3. The judgment error data were subjectedto a 2 (effector: finger, toe) � 2 (source: active, passive) re-peated measures analysis of variance which revealed amain effect of effector, F1,23 = 52.634, p < .0001, with thejudgments of movement initiation being more anticipatoryfor the finger than the toe. Planned comparisons confirmed
nals underlying conscious awareness of action. Cognition
-100
-90
-80
-70
-60
-50
-40
-30
-20
-10
0Finger Toe
Judg
men
t Err
or (m
s)
ActivePassive
Fig. 3. The mean judgment errors for all conditions. Note that there was asignificant difference between active and passive movement conditionsand that toe movement judgments were less anticipatory than fingermovement judgments. Error bars are standard errors. See text forstatistics.
Table 2Mean (SD) button press durations (in ms) for all conditions. See text formore details and statistics.
Active Finger Active Toe Passive Finger Passive Toe
298.70 259.30 301.28 239.41(84.95) (63.16) (47.56) (38.85)
6 S.S. Obhi et al. / Cognition xxx (2008) xxx–xxx
ARTICLE IN PRESS
that this was the case for both active and passive condi-tions (p < .0001 and p < .0001 respectively). There was alsoa main effect of source of movement (F1,23 = 11.808,p = .002) with judgments of active movements being moreanticipatory than passive movements. Planned compari-sons showed that this was true for both finger and toemovements (finger movements, p = .014; toe movementsp = .004). There was no significant interaction betweeneffector and source of movement (F1,23 = 2.120, p = .159).There was no significant difference in the variability (SD)of judgment errors between the finger and toe conditions(although this difference approached significance –F1,23 = 4.073, p = .055, with judgment errors for toe move-ments being more variable than judgment errors for fingermovements). This was also the case for active and passiveconditions (F1,23 = 3.550, p = .072, with judgment errors forpassive movements being more variable than judgment er-rors for active movements). There was no interaction be-tween source of movement and effector (F1,23 = 0.143,p = .709).
3.2. Force data
A 2 � 2 repeated measures analysis of variance showedthat there was no main effect of effector with respect to theforce of the button press (F1,23 = 0.002, p = .963). There wasalso no main effect of source of movement (F1,23 = 0.035,p = .853), nor was there an interaction between source ofmovement and effector (F1,23 = 0.437, p = .515).
3.3. Button press time and duration data
Table 1 displays the mean button press times (in ms)and button press variability (SD) for each condition. There
Table 1Mean (SD) button press times (in ms from time of presentation of the clockface) for all conditions. See text for more details and statistics.
Active Finger Active Toe Passive Finger Passive Toe
4773.67 4848.15 4822.53 4640.71(1205.55) (1142.48) (1534.04) (1584.82)
Please cite this article in press as: Obhi, S. S., et al. On the sig(2008), doi:10.1016/j.cognition.2008.10.009
was no main effect of effector (F1,23 = 0.393, p = .537),source (F1,23 = 0.327, p = .573) or an interaction betweenthese factors (F1,23 = 2.257, p = .147) for the button presstimes. There was a difference in variability (SD) betweenthe active and passive movements (F1,23 = 39.339, p <.0001) with the passive movements being more variablethan active movements. There was no main effect of effec-tor, nor an interaction between source of movement andeffector (F1,23 = 0.025, p = .876 and F1,23 = 1.666, p = .210).
The mean press duration data for each condition areshown in Table 2. There was no main effect of effector orsource of movement (F1,23 = 3.230, p = .085 & F1,23 = 3.620,p = .070 respectively) . There was also no interaction be-tween the two variables (F1,23 = 1.879, p = .184).
4. Discussion
This experiment attempted to determine whether con-scious judgments of movement onset time are based solelyon the efferent signals that instruct the movement orwhether sensory feedback signals that are a consequenceof the movement also play a role. This was investigatedby requiring participants to make judgments about the on-set times of active or passive finger and toe movements.Thus, we manipulated the presence of efferent signals byemploying a passive movement condition, and we manip-ulated conduction length by requiring either finger or toemovements. In line with previous suggestions (e.g., Blake-more et al., 2002; Haggard et al., 1999) the comparisonsbetween active and passive movement conditions revealeda pattern of judgment errors that is consistent with anefferent account of action awareness. However, the patternof judgment errors in the finger versus toe movement con-ditions is consistent with a reafferent account of actionawareness. Specifically, if active movements are based onpre-movement (efferent) signals and passive movementsare based on feedback (reafferent) signals, a pattern ofjudgment data in which active movement judgments aremore anticipatory than passive movement judgmentswould also be expected, which is what our data showed.However, a pure efferent account would predict moreanticipatory judgments for active toe movements com-pared to active finger movements because, for a toe move-ment made at a certain point in time, the preparation forthat movement must occur prior to the preparation of afinger movement with the same onset time. For activemovements the results of the present study show theopposite pattern than that which would be expected onthe basis of an efferent account; finger movements werejudged as being initiated earlier than toe movements. Forpassive conditions the opposite pattern of results wouldbe expected, feedback from the finger should arrive at
nals underlying conscious awareness of action. Cognition

S.S. Obhi et al. / Cognition xxx (2008) xxx–xxx 7
ARTICLE IN PRESS
the brain before feedback from the toe predicting a moreanticipatory judgment for finger movements compared totoe movements. Indeed, the pattern of results from thepassive movement conditions is consistent with such afeedback account; toe movement judgments were lessanticipatory than finger movement judgments. This is thecase even though all judgments were seemingly anticipa-tory in absolute time as measured with the Libet method.However, as previously discussed, this does not imply thatuseful comparisons cannot be made between the differentexperimental conditions. Taken together, results from thecurrent study provide the first direct evidence of the cru-cial role for both premotor signals and feedback signals indetermining conscious awareness of action onset.
Our effort to reduce the physical differences betweenmovement conditions by inclusion of a practice sessionwas successful in that there were no differences in forceor duration between active and passive movements, or fin-ger and toe movements (although there was a trend to-wards toe movements being shorter and less variablethan the finger movements). Thus, we can be quite surethat differences in the physical nature of the movementsdid not contribute to the observed pattern of judgment er-rors. In addition, our a priori decision to exclude trials inwhich judgments were greater than 300 ms early or late(compared to the actual time of the key press) could becriticized for its potential to bias our results. To be clear,our reason for choosing this cut-off is based on our previ-ous experience with this paradigm which suggests thatparticipants are virtually never 300 ms early or late in theirjudgments (corresponding to 7 min on the clock), unlessthey are not paying attention to the task. Furthermore,these exclusion criteria produce results that are completelyin line with previous studies including the original study ofLibet et al. (1983) and other replication studies (e.g., Hag-gard et al., 1999). Lastly, application of the exclusion crite-ria resulted in removal of a negligible percentage of trials.Hence, we are confident that our choice of exclusion crite-ria did not induce bias into our results.
One aspect of the data that is noteworthy is that alljudgments (including passive conditions) were seemingly(i.e., measured as being) anticipatory. It could be arguedthat the reason for this pattern of anticipatory judgmentsfor active and passive movements might be due to a kindof movement ‘expectancy’ on the part of subjects whichmay have triggered some form of premotor activation inpassive movement conditions. However, it has previouslybeen shown that passive movements do not yield electro-myographic (EMG) responses for foot flexion–extensionmovements or finger flexion–extension movements (Mimaet al., 1999; Sahyoun, Floyer-Lea, Johansen-Berg, & Mat-thews, 2004). Further, in an earlier pilot study in our labwe also confirmed that pushing a subject’s index fingeronto a key at unpredictable times in a fashion similar tothe manipulation used here produced no measurableEMG activity in the first dorsal interosseous muscle. Thisevidence coupled with neuroimaging results showing alack of motor cortical activation in passive movementssuggests that there was no premotor activity (includingefference copy) in the passive movement condition in thepresent study (e.g., Mima et al., 1999). However, as previ-
Please cite this article in press as: Obhi, S. S., et al. On the sig(2008), doi:10.1016/j.cognition.2008.10.009
ously mentioned, it is wise not to place too much faith inthe absolute numerical values of judgments in Libet styleparadigms as there are several potential problems associ-ated with the methodology (for wide-ranging critiques ofLibet’s experiments and interpretations including method-ological issues see Dennett & Kinsbourne, 1992; Gomes,1998; Gomes, 2002; Pockett, 2002a; Pockett, 2002b;Ramakrishna, 2002; Breitmeyer, 2002; Haggard & Libet,2001; Lau, Rogers, & Passingham, 2006). In addition, theabsolute values of judgment errors that we see may notsimply be biased due to problems with the Libet method;there also exists the possibility that the brain actuallyreconstructs the time at which the movement would haveoccurred (in passive conditions), so that the judgment ofonset appears anticipatory.
The fact that conscious judgments for active move-ments were more anticipatory than for passive movementslends some support to the idea that pre-movement signalsinvolving the generation and use of efference copy may bethe dominant signals underlying conscious awareness ofaction onset. However, a previous study has suggested thatfeedback from the moving limb may also be an importantsignal used to base conscious awareness judgments on(Obhi, 2007). Briefly, in that experiment, active and passivemovements were found to yield statistically identical con-scious judgments of movement onset. This pattern of re-sults was suggested to fit with a feedback basedexplanation for conscious judgments of action onsets. Atface value, the fact that the present study found differencesin judgment errors between active and passive conditionsand the previous study by Obhi (2007) did not, may seemcause for concern. However, we place more faith in thepresent results compared to the previous results basedon three lines of reasoning: First, as highlighted in theintroduction, the present experiment eliminated an impor-tant confound in the previous study; namely the possibilityof participant’s mistaking the experimenter’s touch as theonset of the movement in the passive condition. This wasachieved by having the experimenter maintain light fingercontact on the participant’s finger throughout each block oftrials (i.e., in both active and passive conditions). In con-trast, in the previous experiment the experimenter didnot touch the participant’s finger at all in active conditionsbut did touch the participant’s finger at unpredictabletimes to push it onto the key in passive conditions. Thisprovides the very real possibility that a premotor (efferent)effect in the active condition was replaced by a ‘‘perceptionof the experimenters touch” effect in the passive condition.This would yield a similar estimation of movement onsettime for active and passive conditions (which is what wereported in that paper). Indeed, unless the two conditionsare matched for the potentially confounding effect ofexperimenter touch there is no way to be sure about thecause of the equivalent judgments found in the previousstudy. This matching was ensured in the current studyand hence the present results are more convincing thanthe former. Second, the current study used double thenumber of participants than the previous study, which fur-ther increases confidence in the present data. Third, andperhaps most importantly, the current active-passive re-sults are in completely line with those reported in studies
nals underlying conscious awareness of action. Cognition

8 S.S. Obhi et al. / Cognition xxx (2008) xxx–xxx
ARTICLE IN PRESS
by other authors showing that active voluntary (i.e., active)movements are judged as occurring earlier than involun-tary (i.e., passive) movements (e.g., Haggard et al., 2002;Haggard & Clark, 2003). Thus, the discrepancy betweenthe present results and those of Obhi (2007) is most likelydue to a significant improvement of experimental proce-dures in the current study.
How then, can such a result be explained? There are atleast three possibilities: First, the signal used to judge theonset of an action may not be a pure pre-movement signalat all, but may instead be a composite signal that incorpo-rates feedback arising from the moving limb. Such incorpo-ration of feedback information would delay the consciousjudgment more for toe than finger movements (due to dif-ferences in conduction length), and could produce the pat-tern of data that we observe (i.e., more anticipatoryjudgments for finger movements – or, put differently, moredelayed judgments of toe movements). Note that a ‘‘delay”in this discussion is synonymous with a less anticipatoryjudgment. A related possibility that is in keeping with thefact that all active movement judgments in the presentstudy were anticipatory is that the two signals consist ofthe generation of efference copy, and the prediction of thesensory consequences of the commanded movement (asopposed to the real sensory feedback). The observed pat-tern of judgment errors for active movements could be ob-tained if predictions are slower for toe movements than forfinger movements. This possibility would require the inter-nal forward model (Wolpert & Miall, 1996) that predictsthe sensory consequences of the efferent commands to re-tain the relative conduction time differences between fin-ger and toe movements in its simulation. This possibleexplanation, which is a predictive variant of the first possi-bility (combined efferent and actual reafference explana-tion) would fit with the data and would avoid having toinvoke a potential reconstructive account of action aware-ness to explain the fact that all judgments were anticipa-tory. However, the explanation does not account for theanticipatory judgments observed for passive movementswhich are not preceded by a predictive process performedby an internal forward model. Thus, it is unlikely that thisexplanation is correct. The third possible explanation isthat the time course over which efference copy signalsactivate their target areas in the brain for finger and toemovements might differ, perhaps on the basis that fingermovements are much more common than toe movements.Hence, areas in the brain that are responsible for process-ing an efference copy signal, might be better ‘‘primed” toprocess an index finger efference copy signal comparedto the much more rarely produced toe pre-movement sig-nal (based on the frequency of past finger movementscompared to toe movements). This suggestion is akin tothe mechanisms believed to underlie perceptual priming,where detectors for more frequent stimuli are betterprimed than those for less frequent stimuli and have ahigher baseline level of activity, thus requiring weaker in-puts to push their activation level up to the ‘‘threshold”(e.g., Howes & Solomon, 1951; Jacoby & Dallas, 1981);the only difference is that in this case, the stimulus is a pat-tern of efference copy, and not a pattern of sensory stimu-lation. Such an explanation would also yield a later percept
Please cite this article in press as: Obhi, S. S., et al. On the sig(2008), doi:10.1016/j.cognition.2008.10.009
of movement onset for toe movements (i.e., a less anticipa-tory conscious judgment) compared to finger movements,which is what we find. This suggestion in itself raises aninteresting question: What role does movement typicality(i.e., the frequency with which a particular movement oc-curs) play in the processing of the efference copy signalaccompanying self produced movement? Are the brainareas responsible for processing premotor or other signalssubject to priming effects whereby more typical move-ments activate these areas more easily (and quickly) thanmore atypical movements? Despite the potential for thisexplanation to account for the current results, it remainsspeculation, and the more plausible (and indeed simpler)idea is that actual feedback signals are indeed importantsources of information underlying conscious judgmentsof movement onset.
The present data provide new (and direct) empiricalsupport for a reconceptualisation of the mechanisms medi-ating conscious awareness of action in Libet tasks. The vastmajority of previous work has explained the anticipatoryawareness of action finding with sole reference to the pro-cessing of efference copy by an internal model or someother premotor process (Haggard & Eimer, 1999; Haggardet al., 1999; Blakemore et al., 2002; Obhi & Haggard,2004; Frith et al., 2000). To be clear, no evidence for thisclaim has (to our knowledge) ever been cited. Rather thisclaim is more of an assumption based on the idea that apredictive signal is available prior to movement onsetwhereas feedback signals are necessarily delayed andhence cannot contribute to the (anticipatory) judgmentof action onset. The present experiment provides the firstreal direct test of this assumption by incorporating thekey manipulation of conduction length in conjunction withactive and passive movement conditions. The data showdirectly that a premotor account alone is not adequate,and that feedback signals from the limb also play a contrib-utory role.
In summary, the data from the current experiment pro-vide the first clear support for the idea that efferent signals(i.e., premotor signals) and movement related feedback(i.e., reafference) are used as a basis for conscious judg-ments of movement onset in Libet paradigms. This dataprompts reevaluation of a sole premotor mechanismunderlying action awareness judgments. Over the comingyears, several more experiments will be needed, perhapsusing other paradigms in addition to the Libet paradigmto fully determine the factors that affect the weighting gi-ven to efferent and reafferent signals in conscious judg-ments of action onset. In constructing such a theory ofhow action awareness arises, it will also be important toverify whether judgments of action onset really are (inabsolute terms) anticipatory. In particular, this will helpevaluate any potential reconstructive accounts of how ac-tion awareness is generated.
Acknowledgements
This work was supported by a Discovery Grant awardedto SSO from the Natural Sciences and Engineering ResearchCouncil of Canada and infrastructure support from theCanada Foundation for Innovation. The authors would like
nals underlying conscious awareness of action. Cognition

S.S. Obhi et al. / Cognition xxx (2008) xxx–xxx 9
ARTICLE IN PRESS
to thank Kara Mendlowitz for assistance with data collec-tion. We are also grateful for the comments of two anony-mous reviewers and Bruno H. Repp for helpful commentson an earlier draft of this paper.
References
Arshavsky, Y. I. (2006). ‘‘Scientific roots” of dualism in neuroscience.Progress in Neurobiology, 79(4), 190–204.
Baars, B. J. (2005). Global workspace theory of consciousness: Toward acognitive neuroscience of human experience. Progress in BrainResearch, 150, 45–53.
Bard, C., Paillard, J., Teasdale, N., Fleury, M., & Lajoie, Y. (1991). Self-induced versus reactive triggering of synchronous hand and heelmovement in young and old subjects. New York, NY, USA: KluwerAcademic/Plenum Publishers.
Billon, M., Bard, C., Fleury, M., & Blouin, J. (1996). Simultaneity of twoeffectors in synchronization with a periodic external signal. HumanMovement Science, 15(1), 25–38.
Billon, M., Bard, C., Fleury, M., Blouin, J., & Teasdale, N. (1996).Simultaneity of two effectors in synchronization with a periodicexternal signal. Human Movement Science, 15(1), 25–38.
Blakemore, S., Wolpert, D. M., & Frith, C. D. (2002). Abnormalities in theawareness of action. Trends in Cognitive Sciences, 6(6), 237–242.
Breitmeyer, B. G. (2002). In support of pockett’s critique of LIbet’s studiesof the time course of consciousness. Consciousness and Cognition: AnInternational Journal. Special Issue: Timing Relations between Brain andWorld, 11(2), 280–283.
Dehaene, S., & Naccache, L. (2001). Towards a cognitive neuroscience ofconsciousness: Basic evidence and a workspace framework. Cognition.Special Issue: The Cognitive Neuroscience of Consciousness, 79(1–2),1–37.
Dennett, D. C., & Kinsbourne, M. (1992). Time and the observer: Thewhere and when of consciousness in the brain. Behavioral and BrainSciences, 15(2), 183–247.
Franck, N., Posada, A., Pichon, S., & Haggard, P. (2005). Altered subjectivetime of events in schizophrenia. The Journal of Nervous and MentalDisease, 193(5), 350–353.
Frith, C. D., Blakemore, S. J., & Wolpert, D. M. (2000). Abnormalities in theawareness and control of action. Philosophical Transactions of the RoyalSociety of London. Series B, Biological Sciences, 355(1404), 1771–1788.
Gazzaniga, M. S. (1998). Brain and conscious experience. Advances inNeurology, 77, 181–192. discussion 192–3.
Gomes, G. (1998). The timing of conscious experience. A critical reviewand reinterpretation of libet’s research. Consciousness and Cognition:An International Journal, 7(4), 559–595.
Gomes, G. (2002). The interpretation of libet’s results on the timing ofconscious events: A commentary. Consciousness and Cognition, 11(2),221–230. discussion 308–13, 314–25.
Haggard, P. (2005). Conscious intention and motor cognition. Trends inCognitive Sciences, 9(6), 290–295.
Haggard, P., & Clark, S. (2003). Intentional action: Conscious experienceand neural prediction. Consciousness and Cognition, 12(4), 695–707.
Haggard, P., Clark, S., & Kalogeras, J. (2002). Voluntary action andconscious awareness. Nature Neuroscience, 5(4), 382–385.
Haggard, P., & Eimer, M. (1999). On the relation between brain potentialsand the awareness of voluntary movements. Experimental BrainResearch. Experimentelle Hirnforschung. Experimentation Cerebrale,126(1), 128–133.
Haggard, P., & Libet, B. (2001). Conscious intention and brain activity.Journal of Consciousness Studies, 8(11), 47–63.
Haggard, P., Newman, C., & Magno, E. (1999). On the perceived time ofvoluntary actions. British Journal of Psychology (London, England:1953), 90(Pt 2), 291–303.
Howes, D., & Solomon, R. (1951). Visual duration thresholds as a functionof word probability. Journal of Experimental Psychology, 41, 401–410.
Jacoby, L. L., & Dallas, M. (1981). On the relationship betweenautobiographical memory and perceptual learning. Journal ofExperimental Psychology: General, 3, 306–340.
Please cite this article in press as: Obhi, S. S., et al. On the sig(2008), doi:10.1016/j.cognition.2008.10.009
Lau, H. C., Rogers, R. D., & Passingham, R. E. (2006). On measuring theperceived onsets of spontaneous actions. The Journal of Neuroscience:The Official Journal of the Society for Neuroscience, 26(27), 7265–7271.
Libet, B. (1985). Subjective antedating of a sensory experience and mind-brain theories: Reply to Honderich (1984). Journal of TheoreticalBiology, 114(4), 563–570.
Libet, B. (1996). Neural processes in the production of conscious experience.Florence, KY, USA: Taylor & Frances/Routledge.
Libet, B., Gleason, C. A., Wright, E. W., & Pearl, D. K. (1983). Time ofconscious intention to act in relation to onset of cerebral activity(readiness-potential). The unconscious initiation of a freely voluntaryact. Brain: A Journal of Neurology, 106(Pt 3), 623–642.
Maia, T. V., & Cleeremans, A. (2005). Consciousness: Converging insightsfrom connectionist modeling and neuroscience. Trends in CognitiveSciences, 9(8), 397–404.
Mandler, G. (2005). The consciousness continuum: From ‘‘qualia” to ‘‘freewill”. Psychological Research/Psychologische Forschung, 69(5–6),330–337.
Mima, T., Sadato, N., Yazawa, S., Hanakawa, T., Fukuyama, H., Yonekura, Y.,et al (1999). Brain structures related to active and passive fingermovements in man. Brain: A Journal of Neurology, 122(10), 1989–1997.
Obhi, S. S. (2007). Evidence for feedback dependent conscious awarenessof action. Brain Research, 1161, 88–94.
Obhi, S. S., & Haggard, P. (2004). Free will and free won’t. AmericanScientist, 92(4), 358–365.
Pockett, S. (2002a). On subjective back-referral and how long it takes tobecome conscious of a stimulus: A reinterpretation of libet’s data.Consciousness and Cognition: An International Journal. Special Issue:Timing Relations between Brain and World, 11(2), 144–161.
Pockett, S. (2002b). Backward referral, flash-lags, and quantum free will:A response to commentaries on articles by pockett, klein, gomes, andtrevena and miller. Consciousness and Cognition: An InternationalJournal. Special Issue: Timing Relations between Brain and World, 11(2),314–325.
Ramakrishna, C. (2002). Real latencies and facilitation. Consciousness andCognition: An International Journal. Special Issue: Timing Relationsbetween Brain and World, 11(2), 300–303.
Rees, G. (2007). Neural correlates of the contents of visual awareness inhumans. Philosophical Transactions of The Royal Society of London SeriesB-Biological Sciences, 362(1481), 877–886.
Sahyoun, C., Floyer-Lea, A., Johansen-Berg, H., & Matthews, P. M. (2004).Towards an understanding of gait control: Brain activation during theanticipation, preparation and execution of foot movements.NeuroImage, 21(2), 568–575.
Shefner, J. M., & Logigian, E. L. (1994). Conduction velocity in motor,cutaneous afferent, and muscle afferent fibers within the same mixednerve. Muscle and Nerve, 17(7), 785–792.
Stenneken, P., Aschersleben, G., Cole, J., & Prinz, W. (2002). Self-inducedversus reactive triggering of synchronous movements in adeafferented patient and control subjects. Psychological Research/Psychologische Forschung, 66(1), 40–49.
Stenneken, P., Prinz, W., Cole, J., Paillard, J., & Aschersleben, G. (2006). Theeffect of sensory feedback on the timing of movements: Evidencefrom deafferented patients. Brain Research, 1084(1), 123–131.
Sternberg, S., Monsell, S., Knoll, R. L., & Wright, C. E. (1978). The latencyand duration of rapid movement sequences: Comparisons of speechand typewriting. In G. E. Stelmach (Ed.), Information processing inmotor control and learning (pp. 117–152). Amsterdam: North-Holland.
Thomas, P. K., Sears, T. A., & Gilliatt, R. W. (1959). The range of conductionvelocity in normal motor nerve fibres to the small muscles of thehand and foot. Journal of Neurology, Neurosurgery, and Psychiatry, 22,175–181.
Wolpert, D. M., & Ghahramani, Z. (2000). Computational principles ofmovement neuroscience. Nature Neuroscience, 3(Suppl.), 1212–1217.
Wolpert, D. M., & Miall, R. C. (1996). Forward models for physiologicalmotor control. Neural Networks: The Official Journal of the InternationalNeural Network Society, 9(8), 1265–1279.
Zeman, A. (2001). Consciousness. Brain: A Journal of Neurology, 124(7),1263–1289.
Zeman, A. (2006). Consciousness. Philadelphia, PA, USA: LippincottWilliams and Wilkins Publishers.
nals underlying conscious awareness of action. Cognition




















