
Oligosaccharide-Induced Whey-Specific CD25+ Regulatory T-Cells Are Involved in the Suppression of...
218
Transcript of Oligosaccharide-Induced Whey-Specific CD25+ Regulatory T-Cells Are Involved in the Suppression of...


Cow’s milk allergy
immune modulation by
dietary intervention
Bastiaan Schouten

ISBN/EAN: 978-90-393-5210-6
Cover photo, design and thesis lay-out: Bastiaan Schouten
Printed by: GVO | Ponsen & Looijen B.V., Ede, The Netherlands
The printing of this thesis was financially supported by:Danone Research – Centre for Specialised Nutrition, J.E. Jurriaanse Stichting, Utrecht Institute for Pharmaceutical Sciences, TNO / Utrecht Center for Food Allergy, Infection & Immunity Center Utrecht, BD Biosciences, FrieslandCampina Domo
© 2009 Bastiaan SchoutenAll rights reserved. No part of this thesis may be reproduced or transmitted in any form, by any means, electronic or mechanical, without prior written permission of the author.

Cow’s milk allergyimmune modulation by dietary intervention
Koemelkallergieimmuunmodulatie door dieetinterventie
(met een samenvatting in het Nederlands)
Proefschrift
ter verkrijging van de graad van doctor aan de Universiteit Utrecht op gezag van de rector magnificus, prof.dr. J.C. Stoof, ingevolge het besluit van het college voor promoties,
in het openbaar te verdedigen op woensdag 25 november 2009 des middags te 2.30 uur
door
Bastiaan Schouten
geboren op 27 februari 1981 te Utrecht

Promotor: Prof.dr. J. Garssen
Co-promotoren: Dr. L.E.M. Willemsen
Dr. L.M.J. Knippels



Contents
Chapter 1 General introduction 9
Chapter 2 Nutrition and food allergy 25
Chapter 3 Acute allergic skin reactions and intestinal contractility 55 changes in mice orally sensitized against casein or whey
Chapter 4 Contribution of IgE and immunoglobulin free light chain in 77 the allergic reaction to cow’s milk proteins
Chapter 5 Non-digestible oligosaccharides reduce immunoglobulin 95 free light-chain levels in infants at risk for allergies
Chapter 6 Cow’s milk allergy symptoms are reduced in mice fed 103 dietary synbiotics during oral sensitization with whey
Chapter 7 Oligosaccharide induced whey-specific CD25+ regulatory 119 T-cells are crucial in the suppression of cow’s milk allergy in mice
Chapter 8 Dietary non-digestible carbohydrates inhibit the allergic 141 effector response via induction of casein-specific CD25+ regulatory T-cells in orally sensitized mice
Chapter 9 iNKT cells contribute to the allergic response in cow’s milk 163 protein sensitized mice
Chapter 10 General discussion 181 Miscellaneous 197 Summary 198 Nederlandse samenvatting 202 Dankwoord 207 Curriculum Vitae 212 List of publications 213 Abbreviations 217


1General introduction

10
1 General Introduction
The adaptive immune response and regulatory T-cells
The human body is continuously under attack by all kinds of microbes like bacteria, fungi, parasites and viruses and has at least three different parts to overcome these invaders. The first is a physicochemical barrier, the epithelial lining of the skin, respiratory and intestinal mucosa forming intrinsic tight junctions which prevent paracellular passage of antigenic particles. In the intestinal and respiratory mucosa the mucus layer forms an extrinsic barrier, preventing direct contact of the microbes with the epithelial lining. The second line of defense is the innate immune system, which comprises the complement system, natural killer cells, basophils, neutrophils and eosinophils, they are rigid and attack pathogens non-specific. The adaptive immune system forms the third immune defense and can be divided in a humoral and a cellular part which are able to recognize and remember specific antigens. In allergic individuals an adaptive immune response is generated towards a harmless antigen. In food allergic patients immunity instead of oral tolerance is raised towards food proteins. This results in a T
H2 type effector immune response and, more chronically, involves an allergen specific TH1 response [1]. Other effector cells like TH17, TH3 and others have not been implicated in the pathology of cow’s milk allergy. Tolerance or immunity to food antigens is generated in the gut associated lymphoid tissue (GALT) (Figure 1). The Peyer’s patches (PP) and mesenteric lymph nodes (MLN) are the inductive sites of the GALT, here immunity or oral tolerance is induced. Food proteins can induce clonal deletion or anergy (high dose tolerance), rendering cells non-responsive for the antigen, or active suppression by regulator T-cells (Treg cells) (low dose tolerance) [2]. Antigens are taken up by dendritic cells (DC) in the PP or lamina propria and travel to the MLN [3]. In the MLN and PP, DC present antigens to naïve T-cells and induce tolerance (anergy/deletion or suppression by Treg cells) or an effector response against the antigen. Antigen-responsive T-cells (CD4+) leave the MLN, travel subsequently through the bloodstream and home back into the intestinal mucosa (lamina propria) and transfer to the peripheral immune system (e.g. spleen) [2, 3]. Different Treg cell subsets are able to actively suppress effector T-cell responses and regulate immune homeostasis of the GALT. Antigen specific T-cells or Treg cells from the MLN in this respect can relocate local induced immune responses to the periphery. Cytokines produced by allergic T
H2 effector cells instruct isotype switching and maturation of B cells resulting in immunoglobulin (Ig)E production in human and in mice both IgE and IgG1 antibodies were produced against this allergen. These antibodies bind to Fc receptors on mast cells and after a second allergen exposure, receptor crosslinking results in mast cell activation and degranulation. Mast cell mediator release such as histamine and leukotriens cause the acute allergic symptoms like eczema, diarrhea, asthma and in some cases anaphylaxis.
The humoral response of the murine adaptive immune system is complex and covers both

11
1Chapter 1
TH1/Treg (IgG2a and IgA), TH2 (IgE, IgG1) and Ig free light chain (Ig-fLC) molecules. Ig-fLC are present in serum and their production is augmented under inflammatory conditions including allergic as well as some autoimmune diseases [4-6]. For example, in patients suffering from multiple sclerosis free kappa light chains correlate with disability prognosis [7] and in rheumatoid arthritis there is a significant correlation between kappa and lambda fLC and the disease activity score [8]. There is another subset of T-cells that regulates T
H1 and TH2, these are the Treg cells. The Treg cells are a heterogeneous group of cells that act via cytokine secretion or via direct cell-cell contact. In food allergy the delicate balance between oral tolerance and hypersensitivity is regulated by these cells [2, 3]. Cells with regulatory properties include TGF-β-producing TH3 cells, IL-10-producing Tr1 cells, adaptive and natural Foxp3 regulatory T (Treg)-cells and natural killer (NK) T cells [9-11]. In addition, the TGF-β and IL-10 producing naturally occurring CD4+CD25+Foxp3+ Treg cells were described to be important for maintaining peripheral tolerance and for suppression of CD4+ and CD8+ T cell proliferation [12, 13]. Treg represent 5-10% of the CD4+ T lymphocyte population and express the transcription factor forkhead box p3 (Foxp3), which is confirmed as a key gene for generation and maintenance of Treg cells. Adaptive Treg cells are educated in the GALT in particular and cells have similar surface markers; however their inhibitory capacities act via cell-cell contact instead of cytokines [14]. Treg cells are believed to alter mast cell degranulation via IL-10 or TGF-β production or cell-cell contact [15, 16]. In addition, Treg cells have been shown to suppress Fc-receptor expression on mast cells which would reduce the capacity of these cells to degranulate and induce an acute skin response [12]. NKT cells express a T cell receptor, but recognize glycolipids presented by CD1d molecules [17, 18]. In allergic asthma α-galactosylceramide induced NKT cells have been shown to suppress the airway hyper responsiveness, hence also NKT cells can modulate the allergic response [19].
Cow’s milk allergy
Food allergy is a growing problem in Western Europe and the USA. Clinical symptoms may involve the skin, respiratory tract and gastrointestinal tract, which can even lead to a systemic anaphylactic reaction [20, 21]. Cow’s milk allergy (CMA) is one of the leading causes of food allergy in adults [22]. In developed countries approximately 2 to 3% of infants exhibit CMA. Although most infants outgrow CMA before their fifth year, IgE-mediated CMA predisposes the development of other (food) allergies and even asthma later in life [23, 24]. Cow’s milk contains two main protein classes, which are the caseins (30 g/L) and whey proteins (5 g/L). The caseins consist mainly of αS1-, αS2-, κ- and β-casein, whereas whey proteins comprise of β-lactoglobulin, α-lactalbumin, bovine serum albumin, serum immunoglobulins and lactoferrin [25]. Large population studies with cow’s milk allergic infants have shown that the major allergens are β-lactoglobulin and αS1-casein [26, 27].

12
1 General Introduction

13
1Chapter 1
Figu
re 2
. W
orki
ng h
ypot
hesi
s of
thi
s th
esis
. a
Die
tary
cow
’s m
ilk p
rote
ins
such
as
case
in a
nd w
hey
can
caus
e se
nsiti
zatio
n by
bre
akin
g to
lera
nce
in t
he in
testi
nal m
ucos
a; h
ydro
lysa
tes
and
amin
o ac
id fo
rmul
as a
re d
evel
oped
to
redu
ce t
he r
isk
of s
ensi
tizati
on. b
If t
hese
food
co
mpo
nent
s (a
nd o
ther
sub
stan
ces)
are
tak
en u
p by
M-c
ells
, an
tigen
pre
senti
ng d
endr
itic
cells
(D
C) i
n th
e Pe
yer’
s pa
tche
s (P
P) p
rese
nt
thes
e pr
otei
ns t
o na
ïve
T ce
lls (
T H0)
, w
hich
diff
eren
tiate
, de
pend
ent
on t
he c
ytok
ine
mili
eu,
into
TH1
or T
H2
effec
tor
cells
(im
mun
ity)
or
into
reg
ulat
ory
T (T
reg)
-cel
ls w
hen
tole
ranc
e de
velo
ps.
In c
ase
of h
igh
dose
tol
eran
ce a
nerg
y or
clo
nic
dele
tion
occu
rs.
c D
C’s
can
also
tr
affic
from
the
PP
to t
he M
LN o
r d
sam
ple
antig
en f
rom
the
lum
en a
nd e
ffect
or s
ites
and
than
tra
ffic
to t
he M
LN w
ere
antig
ens
are
pres
ente
d to
nai
ve T
-cel
ls. e
Gen
erat
ed T
reg
cells
or
effec
tor
cells
ent
er t
he b
lood
stre
am a
nd h
ome
back
to t
he in
testi
nal m
ucos
a w
ere
they
w
ill b
ecom
e re
side
nt in
the
lam
ina
prop
ria. f
Tre
g ce
lls o
r eff
ecto
r T-
cells
gen
erat
ed in
the
MLN
can
als
o tr
affic
to t
he p
erip
hera
l im
mun
e sy
stem
and
tra
nsfe
r to
lera
nce
or i
mm
unity
. g/
h B-
cells
exp
anse
and
mat
ure
in s
plee
n an
d tr
affic
back
to
the
effec
tors
site
whe
re t
hey
prod
uce
spec
ific
IgE,
IgG
1, IgG
2a, I
g-fL
C or
IgA
. i T
hese
Ig’s
bin
d to
mas
t ce
lls, w
hich
res
ides
and
wai
t fo
r an
othe
r an
tigen
exp
osur
e. j
This
im
mun
e m
echa
nism
can
be
mod
ulat
ed b
y fo
od c
ompo
nent
s e.
g. p
rebi
otics
, pro
bioti
cs, t
he c
ombi
natio
n (s
ynbi
otics
) and
om
ega-
3 fa
tty
acid
s.
T H: T
hel
per c
ells
; Tre
g: R
egul
ator
y T
cells
; NKT
: Nat
ural
Kill
er T
cel
ls; D
C: d
entr
itic
cells
; pCH
: par
tial c
asei
n hy
drol
ysat
e; e
CH: e
xten
sive
cas
ein
hydr
olys
ate;
pW
H: p
artia
l whe
y hy
drol
ysat
e; e
WH
: ext
ensi
ve w
hey
hydr
olys
ate;
AA
: am
ino
acid
s

14
1 General Introduction
Development of a mouse model for orally induced CMA
Animal models of CMA provide a tool to unravel mechanisms involved in CMA and may explore new therapeutic and preventive approaches. However, in most existing food allergy models the animals are not sensitized via the oral route (e.g. intra peritoneal), while in reality most humans are sensitized orally [28-36]. To further assess mechanisms underlying cow’s milk allergy and/or test new concepts for prevention and/or treatment of CMA, the major goal of the present thesis was to introduce and develop new tools to address the pathological changes upon oral sensitization with casein and whey in mice. In this model humoral Ig and Ig-fLC responses were determined and the involvement of Treg cells in the allergic response was evaluated. Furthermore measurement of the acute allergic skin response, as a potential equivalent of the human skin prick test, was introduced as a new readout of systemic sensitization. In this thesis the preventive effects of dietary intervention with prebiotic non-digestible oligosaccharides and a candidate probiotic bacterial strain the Bifidobacterium Breve M-16V were studied.
Prebiotics
Non-digestible carbohydrates form a major constituent of breast milk. Human milk contains approximately 7-12 g/L oligosaccharides [37]. At least 130 different oligosaccharides have been isolated from human milk and the two main categories are neutral and acidic oligosaccharides [38]. These human milk oligosaccharides, but also specific dietary fibers, such as inulin derived from chicory, can selectively support growth of health promoting commensal bacteria in the gut and are therefore called prebiotics [39-42]. Prebiotics were defined as “food ingredients that beneficially affect the host by selectively stimulating the growth and/or activity of one or a limited number of bacterial species already resident in the colon, and thus attempt to improve host health” [39]. Recently this definition is adapted to: “A prebiotic is a selectively fermented ingredient that allows specific changes, both in the composition and/or activity in the gastrointestinal microflora that confers benefits upon host wellbeing and health” [43]. These bacterial species comprise among others the Gram-positive Lactobacillus genus and the Bifidobacterium genus. Besides their effects on the intestinal microbiota non-digestible carbohydrates may improve absorption of minerals and water holding capacity of the fecal bulk due to their physicochemical properties [44]. Short-chain fatty acids, released by for example lactic acid producing Bifidobacteria and Lactobacilli upon fermentation of prebiotics, are essential nutrients for intestinal epithelial cells and support gut function [45]. Besides the physicochemical properties of prebiotics, these non-digestible carbohydrates can enhance non-specific defense mechanisms [46]. There are

15
1Chapter 1
different types of dietary fibers e.g. (hemi)cellulose, lignin, β-glucans, pectins, gums, inulin and oligosaccharides [47]. Different non-digestible oligosaccharides with specific properties are obtained or manufactured from natural sources and used as dietary supplement. Dietary fibers are extracted from plants and/or produced enzymatically or chemically [48]. Short-chain galacto-oligosaccharides (scGOS), long-chain fructo-oligosaccarides (lcFOS) and pectin-derived acidic oligosaccharides (pAOS) are some examples of non-digestible oligosaccharides that mimic the functionality and molecular size distribution of human milk oligosaccharides and have been investigated in this thesis (Figure 2). Prebiotic scGOS are produced by enzymatic elongation of galactose (derived from lactose) by β-galactosidase, with a degree of polymerization (dp) of 3–8. Chicory inulin is a polymer from D-fructose residues linked by β(2-1) bonds with a terminal α(1-2) linked D-glucose (dp 2-60). lcFOS are produced from inulin and have a mean dp >23, whereas scFOS have mainly a dp <23. lcFOS are added to infant formulas. pAOS is a mixture of linear oligomers and small polymers of
Figure 2. Structures representing non-digestible carbohydrates used in this thesis. a Enzymatic elongation of galactose using β-galactosidase results in short-chain galacto-oligosaccharides (dp of 3-8). Structure represents two β(1-4) linked galactose molecules connected to a terminal glucose monomer. b Long-chain fructo-oligosaccharides are produced from chicory inulin (dp of 7-60 and mean dp > 23). Structure represents one glucose molecule and two fructose molecules connected with a β(2-1) linkage. c Basic chemical structure of pectin derived acidic oligosaccharides consisting of α(1-4) linked galacturonic acid molecules (dp of 1-20) (R=H or R=CH
3). Adapted from Vos et al. [49].
A
OHn<6
O
CH2OH
OH
O
CH2OH
OH
OH
O
O
CH2OH
OH
OH
OH
O
OH
B
O
CH2OH
OH
OH
OH
CH2OH
OCH2OH
OH
OH
OCH2OH
OH
OH
CH2
O
O
meann>21
O
COOR
OH
OH
O O
COOR
OH
OH
O O
COOR
OH
OH
OH
C n<18

16
1 General Introduction
galacturonic acid produced from from food grade pectin by pectin cleaving enzymes and have a dp of 1-20. Currently, it is not feasible to produce the precise human milk oligosaccharides. However, the used specific oligosaccharides have proven efficacy in clinical trials since they reduce the incidence of allergic disease and have been shown to enhance fecal Bifidobacteria counts [50-54]. Another prebiotic fructo-oligosaccharide, kestose, showed to be effective in reducing the incidence of atopic dermatitis in infants as well [55].
The intestinal microbiota and probiotics
The human gastrointestinal tract is composed of several compartments. The majority of macro- and micronutrients from food is absorbed in the duodenum and jejunum of the small intestine, while the ileum has residual absorptive capacities. After the ileum the chyme enters the large intestine which is mainly involved in water and electrolyte absorption. However, the colon is also the main reservoir for the intestinal microbiota which comprises a multitude of different bacterial strains. The human gastrointestinal tract contains ten times more bacteria than the total number of cells in the human body [56]. The colonization of the gastrointestinal tract by bacteria starts immediately after birth [57, 58]. Most of the bacteria in the gastrointestinal tract are commensals, bacteria that do not negatively affect human health. A well balanced intestinal microbiota consists of less than one percentage of pathogens, e.g. Klebsiella pneumoniae and Salmonella [59, 60]. A particular class of bacteria, i.e. probiotic bacteria, has been shown to be beneficial for the host. Probiotics can be defined as: “live microbial food ingredients that alter the enteric microflora and have a beneficial effect on health” [61]. Bifidobacteria together with Lactobacilli are well known for their health promoting functions. Bifidobacteria alone comprise of more than 60% in fecal samples of children [62]. Evidence is accumulating that both Bifidobacteria and Lactobacilli may account for as much as 90% of the total flora of breast fed infants [63]. Some of these are selected species and used as probiotics [20, 64, 65]. Probiotics stabilize the gut microflora hereby preventing pathogenic infection and pathogen induced diarrhea. Ongoing studies are being performed to unravel the immune modulatory effects of probiotics and their efficacy to reduce the incidence or relieve symptoms of various diseases caused by dysregulated immune responses [66].
Breast fed neonates have higher stool counts of Bifidobacteria compared with babies fed standard infant formula. When added to standard milk formula, the prebiotic mixture scGOS/lcFOS was found to enhance Bifidobacteria fecal counts in bottle fed infants to levels almost similar to those found in breast fed infants [67]. High Bifidobacteria counts were found to associate with a reduced risk of development of atopic disease in infants of westernized countries [20, 68]. Therefore probiotics have been investigated in multiple clinical studies for

17
1Chapter 1
the prevention and/or treatment of allergic diseases [65, 69-78]. However, there is still no consensus regarding the efficacy of these bacteria in having immune-modulatory effects and their clinical relevance [79]. In two Cochrane meta-analysis of randomized controlled trials the effectiveness of probiotics in the prevention or treatment of eczema is not confirmed [80, 81]. Several factors are suggested which influence the outcome of the studies, e.g. probiotic strain or mixture, environmental factors, intervention duration and timing and bacterial load [82, 83]. Most of the clinical studies are performed using Lactobacillus strains, however in breast fed infants in particular Bifidobacteria have been found in the feces [84]. In a bacterial strain comparison study with six different Bifidobacterium and Lactobacillus strains, the Bifidobacterium breve M-16V was identified as being the most potent anti-allergic strain in the treatment of ovalbumin-induced allergic asthma in mice [85]. Based on these results the Bifidobacterium breve M-16V was the strain of choice in the studies conducted in this thesis.
Synbiotics
To improve human health and reduce the development of allergies the combination of prebiotics and probiotics, called synbiotics, gain increased scientific attention. The hypothesis that pre- and probiotics together are more effective than the use of single preparations is very promising, although not many studies have been performed in relation to allergy [86-89]. The (preclinical) studies that have been carried out show potential for synbiotics to reduce the incidence and severity of allergic disease, since synbiotics were found to increase total serum (soluble) IgA antibodies and to reduce dermatitis-like skin lesions [86, 88].
Aim and scope of this thesis
This thesis provides preclinical data concerning prevention of orally induced cow’s milk allergy using dietary intervention with non-digestible carbohydrates and/or Bifidobacterium breve M-16V. Furthermore some of the underlying mechanisms of the pathophysiology of cow’s milk allergy are studied and in particular the function of Treg cells is addressed. Chapter 2 provides a literature study on food allergy and nutrition, its scope is wider than that of cow’s milk allergy solely, and multiple routes of intervention in the process of sensitization and allergy are described. Chapter 3 describes the establishment of a murine model of orally induced sensitization with casein or whey protein adapted from Li et al. [35] and Frossard et al. [36]. These groups have introduced an oral mouse model for whole cow’s milk protein or β-lactoglobulin, whereas in this thesis mice were sensitized with the complete whey or

18
1 General Introduction
casein protein mixture. In Europe most of the infant formulas are based on whey proteins, whereas in the USA the majority of infant formulas are based on casein proteins, however infant formulas with a mix of both casein and whey are on the market as well. We choose to develop a casein and a whey model. Surprisingly, the underlying mechanisms responsible for either casein- or whey-sensitization were found to be different. Chapter 4 describes the involvement of immunoglobulin free light chain (Ig-fLC) in the allergic effector response in casein sensitized mice using Ig-fLC blocker F991 in the active model and in passive transfer studies with spleen supernatants. The potential contribution of Ig-fLC in human allergic disease was evaluated as well. Ig-fLC (λ and κ) concentrations were determined in plasma samples from high risk children who developed atopic dermatitis (AD) and compared to children who did not develop AD. Chapter 5 describes the results of Ig-fLC measurement in sera of children at risk who were supplemented with the prebiotic mixture scGOS/lcFOS (Immunofortis®) for six months. The infants in this trial had fewer infections and fewer children suffered from atopic dermatitis [49-51]. In chapter 6 a dietary intervention study, with the same prebiotic mixture as tested in the clinical trial (scGOS/lcFOS), the candidate probiotic strain (Bifidobacterium breve M-16V) and the combination of both (synbiotics) was performed in whey sensitized mice in a preventive setup. The effectiveness of the diets in reducing the acute allergic skin response, anaphylaxis and multiple serological parameters was studied. In chapter 7 a third potential effective non-digestible carbohydrate, pAOS, was added to the already potent scGOS/lcFOS mixture and fed to the mice during oral sensitization with whey. In these mice the contribution of CD25+ regulatory T cells in the reduction of the allergic effector response caused by the diet was explored using adoptive transfer experiments. Chapter 8 describes in vivo depletion studies to explore CD25+ regulatory T cell function and in addition, an adoptive transfer experiment was performed to study the role of CD4+CD25+ Treg cells in more detail. Chapter 9 describes the contribution of another subset of regulatory T cells, the NKT cells, in allergic sensitization with whey using NKT cell agonist α-galactosylceramide. Chapter 10 provides an overall discussion of the results of this thesis putting together a small part of the cow’s milk allergy puzzle.
References
1. Romagnani, S., Immunologic influences on allergy and the TH1/TH2 balance. J Allergy Clin
Immunol, 2004. 113(3): p. 395-400.
2. Tsuji, N.M. and A. Kosaka, Oral tolerance: intestinal homeostasis and antigen-specific
regulatory T cells. Trends Immunol, 2008. 29(11): p. 532-40.
3. Mowat, A.M., Anatomical basis of tolerance and immunity to intestinal antigens. Nat Rev
Immunol, 2003. 3(4): p. 331-41.
4. Thio, M., et al., Free immunoglobulin light chains: a novel target in the therapy of

19
1Chapter 1
inflammatory diseases. Trends Pharmacol Sci, 2008. 29(4): p. 170-4.
5. Cooper, A. and R. Bluestone, Free immunoglobulin light chains in connective tissue diseases.
Ann Rheum Dis, 1968. 27(6): p. 537-43.
6. Kraneveld, A.D., et al., Elicitation of allergic asthma by immunoglobulin free light chains.
Proc Natl Acad Sci U S A, 2005. 102(5): p. 1578-83.
7. Rinker, J.R., 2nd, K. Trinkaus, and A.H. Cross, Elevated CSF free kappa light chains correlate
with disability prognosis in multiple sclerosis. Neurology, 2006. 67(7): p. 1288-90.
8. Gottenberg, J.E., et al., Serum immunoglobulin free light chain assessment in rheumatoid
arthritis and primary Sjogren’s syndrome. Ann Rheum Dis, 2007. 66(1): p. 23-7.
9. van Wijk, F., et al., CD4+CD25+ T cells regulate the intensity of hypersensitivity responses
to peanut, but are not decisive in the induction of oral sensitization. Clin Exp Allergy, 2007.
37(4): p. 572-81.
10. Akbari, O. and D.T. Umetsu, Role of regulatory dendritic cells in allergy and asthma. Curr
Opin Allergy Clin Immunol, 2004. 4(6): p. 533-8.
11. O’Garra, A. and P. Vieira, Regulatory T cells and mechanisms of immune system control. Nat
Med, 2004. 10(8): p. 801-5.
12. Kashyap, M., et al., Cutting edge: CD4 T cell-mast cell interactions alter IgE receptor
expression and signaling. J Immunol, 2008. 180(4): p. 2039-43.
13. Izcue, A. and F. Powrie, Special regulatory T-cell review: Regulatory T cells and the intestinal
tract--patrolling the frontier. Immunology, 2008. 123(1): p. 6-10.
14. Sun, C.M., et al., Small intestine lamina propria dendritic cells promote de novo generation
of Foxp3 T reg cells via retinoic acid. J Exp Med, 2007. 204(8): p. 1775-85.
15. Gri, G., et al., CD4+CD25+ regulatory T cells suppress mast cell degranulation and allergic
responses through OX40-OX40L interaction. Immunity, 2008. 29(5): p. 771-81.
16. Bundoc, V.G. and A. Keane-Myers, IL-10 confers protection from mast cell degranulation in a
mouse model of allergic conjunctivitis. Exp Eye Res, 2007. 85(4): p. 575-9.
17. Godfrey, D.I., et al., NKT cells: what’s in a name? Nat Rev Immunol, 2004. 4(3): p. 231-7.
18. Jukes, J.P., K.J. Wood, and N.D. Jones, Natural killer T cells: a bridge to tolerance or a pathway
to rejection? Transplantation, 2007. 84(6): p. 679-81.
19. Hachem, P., et al., Alpha-galactosylceramide-induced iNKT cells suppress experimental
allergic asthma in sensitized mice: role of IFN-gamma. Eur J Immunol, 2005. 35(10): p. 2793-
802.
20. Furrie, E., Probiotics and allergy. Proc Nutr Soc, 2005. 64(4): p. 465-9.
21. Kalach, N., et al., Intestinal permeability in children: variation with age and reliability in the
diagnosis of cow’s milk allergy. Acta Paediatr, 2001. 90(5): p. 499-504.
22. Schafer, T., et al., Epidemiology of food allergy/food intolerance in adults: associations with
other manifestations of atopy. Allergy, 2001. 56(12): p. 1172-9.
23. Rhodes, H.L., et al., Early life risk factors for adult asthma: a birth cohort study of subjects at
risk. J Allergy Clin Immunol, 2001. 108(5): p. 720-5.
24. Host, A., Frequency of cow’s milk allergy in childhood. Ann Allergy Asthma Immunol, 2002.

20
1 General Introduction
89(6 Suppl 1): p. 33-7.
25. Monaci, L., et al., Milk allergens, their characteristics and their detection in food: a review.
European Food Research and Technology, 2005(Volume 223, Number 2 / June, 2006): p.
149-179.
26. Bahna, S.L., Cow’s milk allergy versus cow milk intolerance. Ann Allergy Asthma Immunol,
2002. 89(6 Suppl 1): p. 56-60.
27. Ruiter, B., et al., Role of Human Leucocyte Antigen DQ in the Presentation of T Cell Epitopes
in the Major Cow’s Milk Allergen alphas1-Casein. Int Arch Allergy Immunol, 2007. 143(2): p.
119-126.
28. Knippels, L.M., et al., Oral sensitization to food proteins: a Brown Norway rat model. Clin Exp
Allergy, 1998. 28(3): p. 368-75.
29. Adel-Patient, K., et al., Oral administration of recombinant Lactococcus lactis expressing
bovine beta-lactoglobulin partially prevents mice from sensitization. Clin Exp Allergy, 2005.
35(4): p. 539-46.
30. Fritsche, R., et al., IgE-mediated rat mast cell triggering with tryptic and synthetic peptides
of bovine beta-lactoglobulin. Int Arch Allergy Immunol, 2005. 138(4): p. 291-7.
31. Bevilacqua, C., et al., Goats’ milk of defective alpha(s1)-casein genotype decreases intestinal
and systemic sensitization to beta-lactoglobulin in guinea pigs. J Dairy Res, 2001. 68(2): p.
217-27.
32. Buchanan, B.B. and O.L. Frick, The dog as a model for food allergy. Ann N Y Acad Sci, 2002.
964: p. 173-83.
33. Peng, H.J., et al., Effect of ingestion of cow’s milk hydrolysed formulas on whey protein-
specific Th2 immune responses in naive and sensitized mice. Clin Exp Allergy, 2004. 34(4): p.
663-70.
34. Peng, H.J., et al., Effect of ingestion of cow’s milk protein hydrolysate formulas on alpha-
casein-specific immunoglobulin E and G1 antibody responses in naive and sensitized mice. J
Pediatr Gastroenterol Nutr, 2005. 41(4): p. 438-44.
35. Li, X.M., et al., A murine model of IgE-mediated cow’s milk hypersensitivity. J Allergy Clin
Immunol, 1999. 103(2 Pt 1): p. 206-14.
36. Frossard, C.P., C. Hauser, and P.A. Eigenmann, Antigen-specific secretory IgA antibodies in
the gut are decreased in a mouse model of food allergy. J Allergy Clin Immunol, 2004. 114(2):
p. 377-82.
37. Boehm, G. and B. Stahl, Oligosaccharides from milk. J Nutr, 2007. 137(3 Suppl 2): p. 847S-
9S.
38. Nakhla, T., et al., Neutral oligosaccharide content of preterm human milk. Br J Nutr, 1999.
82(5): p. 361-7.
39. Gibson, G.R. and M.B. Roberfroid, Dietary modulation of the human colonic microbiota:
introducing the concept of prebiotics. J Nutr, 1995. 125(6): p. 1401-12.
40. Boehm, G., et al., Prebiotic concept for infant nutrition. Acta Paediatr Suppl, 2003. 92(441):
p. 64-7.

21
1Chapter 1
41. Boehm, G., et al., Prebiotic carbohydrates in human milk and formulas. Acta Paediatr Suppl,
2005. 94(449): p. 18-21.
42. Fanaro, S., et al., Galacto-oligosaccharides and long-chain fructo-oligosaccharides as
prebiotics in infant formulas: a review. Acta Paediatr Suppl, 2005. 94(449): p. 22-6.
43. Gibson, G.R., et al., Dietary modulation of the human colonic microbiota: updating the
concept of prebiotics. Nutr Res Rev, 2004. 17(2): p. 259-75.
44. Greger, J.L., Nondigestible carbohydrates and mineral bioavailability. J Nutr, 1999. 129(7
Suppl): p. 1434S-5S.
45. Schley, P.D. and C.J. Field, The immune-enhancing effects of dietary fibres and prebiotics. Br
J Nutr, 2002. 87 Suppl 2: p. S221-30.
46. Buddington, R.K., et al., Non-digestible oligosaccharides and defense functions: lessons
learned from animal models. Br J Nutr, 2002. 87 Suppl 2: p. S231-9.
47. Schneeman, B.O., Fiber, inulin and oligofructose: similarities and differences. J Nutr, 1999.
129(7 Suppl): p. 1424S-7S.
48. Meyer, P.D., Nondigestible oligosaccharides as dietary fiber. J AOAC Int, 2004. 87(3): p. 718-
26.
49 Vos, A.P., et al., Immune-modulatory effects and potential working mechanisms of orally
applied nondigestible carbohydrates. Crit Rev Immunol, 2007. 27(2): p. 97-140.
50. Moro, G., et al., A mixture of prebiotic oligosaccharides reduces the incidence of atopic
dermatitis during the first six months of age. Arch Dis Child, 2006. 91(10): p. 814-9.
51. Arslanoglu, S., et al., Early dietary intervention with a mixture of prebiotic oligosaccharides
reduces the incidence of allergic manifestations and infections during the first two years of
life. J Nutr, 2008. 138(6): p. 1091-5.
52. Arslanoglu, S., G.E. Moro, and G. Boehm, Early supplementation of prebiotic oligosaccharides
protects formula-fed infants against infections during the first 6 months of life. J Nutr, 2007.
137(11): p. 2420-4.
53. van Hoffen, E., et al., A specific mixture of short-chain galacto-oligosaccharides and long-
chain fructo-oligosaccharides induces a beneficial immunoglobulin profile in infants at high
risk for allergy. Allergy, 2009. 64(3): p. 484-7.
54. Boehm, G. and G. Moro, Structural and functional aspects of prebiotics used in infant
nutrition. J Nutr, 2008. 138(9): p. 1818S-1828S.
55. Shibata, R., et al., Clinical effects of kestose, a prebiotic oligosaccharide, on the treatment of
atopic dermatitis in infants. Clin Exp Allergy, 2009.
56. Guarner, F. and J.R. Malagelada, Gut flora in health and disease. Lancet, 2003. 361(9356): p.
512-9.
57. Blaser, M.J. and J.M. Musser, Bacterial polymorphisms and disease in humans. J Clin Invest,
2001. 107(4): p. 391-2.
58. Schiffrin, E.J. and S. Blum, Interactions between the microbiota and the intestinal mucosa.
Eur J Clin Nutr, 2002. 56 Suppl 3: p. S60-4.
59. Keynan, Y. and E. Rubinstein, The changing face of Klebsiella pneumoniae infections in the

22
1 General Introduction
community. Int J Antimicrob Agents, 2007. 30(5): p. 385-9.
60. Shah, N., H.L. DuPont, and D.J. Ramsey, Global etiology of travelers’ diarrhea: systematic
review from 1973 to the present. Am J Trop Med Hyg, 2009. 80(4): p. 609-14.
61. Shanahan, F., Probiotics and inflammatory bowel disease: is there a scientific rationale?
Inflamm Bowel Dis, 2000. 6(2): p. 107-15.
62. Harmsen, H.J., et al., Analysis of intestinal flora development in breast-fed and formula-fed
infants by using molecular identification and detection methods. J Pediatr Gastroenterol
Nutr, 2000. 30(1): p. 61-7.
63. Fanaro, S., et al., Galacto-oligosaccharides are bifidogenic and safe at weaning: a double-
blind randomized multicenter study. J Pediatr Gastroenterol Nutr, 2009. 48(1): p. 82-8.
64. Boehm, G., et al., Supplementation of a bovine milk formula with an oligosaccharide mixture
increases counts of faecal bifidobacteria in preterm infants. Arch Dis Child Fetal Neonatal Ed,
2002. 86(3): p. F178-81.
65. Kukkonen, K., et al., Probiotics and prebiotic galacto-oligosaccharides in the prevention
of allergic diseases: A randomized, double-blind, placebo-controlled trial. J Allergy Clin
Immunol, 2007. 119(1): p. 192-8.
66. Shida, K. and M. Nanno, Probiotics and immunology: separating the wheat from the chaff.
Trends Immunol, 2008.
67. Haarman, M. and J. Knol, Quantitative real-time PCR assays to identify and quantify fecal
Bifidobacterium species in infants receiving a prebiotic infant formula. Appl Environ
Microbiol, 2005. 71(5): p. 2318-24.
68. Strachan, D.P., Hay fever, hygiene, and household size. Bmj, 1989. 299(6710): p. 1259-60.
69. Moneret-Vautrin, D.A., et al., Probiotics may be unsafe in infants allergic to cow’s milk.
Allergy, 2006. 61(4): p. 507-8.
70. Sistek, D., et al., Is the effect of probiotics on atopic dermatitis confined to food sensitized
children? Clin Exp Allergy, 2006. 36(5): p. 629-33.
71. Kalliomaki, M., et al., Probiotics and prevention of atopic disease: 4-year follow-up of a
randomised placebo-controlled trial. Lancet, 2003. 361(9372): p. 1869-71.
72. Marschan, E., et al., Probiotics in infancy induce protective immune profiles that are
characteristic for chronic low-grade inflammation. Clin Exp Allergy, 2008. 38(4): p. 611-8.
73. Kalliomaki, M., et al., Probiotics in primary prevention of atopic disease: a randomised
placebo-controlled trial. Lancet, 2001. 357(9262): p. 1076-9.
74. Viljanen, M., et al., Probiotics in the treatment of atopic eczema/dermatitis syndrome in
infants: a double-blind placebo-controlled trial. Allergy, 2005. 60(4): p. 494-500.
75. Abrahamsson, T.R., et al., Probiotics in prevention of IgE-associated eczema: a double-blind,
randomized, placebo-controlled trial. J Allergy Clin Immunol, 2007. 119(5): p. 1174-80.
76. Brouwer, M.L., et al., No effects of probiotics on atopic dermatitis in infancy: a randomized
placebo-controlled trial. Clin Exp Allergy, 2006. 36(7): p. 899-906.
77. Kalliomaki, M., et al., Probiotics during the first 7 years of life: a cumulative risk reduction of
eczema in a randomized, placebo-controlled trial. J Allergy Clin Immunol, 2007. 119(4): p.

23
1Chapter 1
1019-21.
78. Soh, S.E., et al., Probiotic supplementation in the first 6 months of life in at risk Asian infants-
-effects on eczema and atopic sensitization at the age of 1 year. Clin Exp Allergy, 2009. 39(4):
p. 571-8.
79. del Giudice, M.M. and F.P. Brunese, Probiotics, prebiotics, and allergy in children: what’s
new in the last year? J Clin Gastroenterol, 2008. 42 Suppl 3 Pt 2: p. S205-8.
80. Boyle, R.J., et al., Probiotics for treating eczema. Cochrane Database Syst Rev, 2008(4): p.
CD006135.
81. Osborn, D.A. and J.K. Sinn, Probiotics in infants for prevention of allergic disease and food
hypersensitivity. Cochrane Database Syst Rev, 2007(4): p. CD006475.
82. Savilahti, E., M. Kuitunen, and O. Vaarala, Pre and probiotics in the prevention and treatment
of food allergy. Curr Opin Allergy Clin Immunol, 2008. 8(3): p. 243-8.
83. Prescott, S.L. and B. Bjorksten, Probiotics for the prevention or treatment of allergic diseases.
J Allergy Clin Immunol, 2007. 120(2): p. 255-62.
84. Gronlund, M.M., et al., Maternal breast-milk and intestinal bifidobacteria guide the
compositional development of the Bifidobacterium microbiota in infants at risk of allergic
disease. Clin Exp Allergy, 2007. 37(12): p. 1764-72.
85. Hougee, S., et al., Oral treatment with probiotics reduces allergic symptoms in ovalbumin-
sensitized mice: a bacterial strain comparison study. Int Arch Allergy Immunol, 2009.
86. Frece, J., et al., Synbiotic effect of Lactobacillus helveticus M92 and prebiotics on the
intestinal microflora and immune system of mice. J Dairy Res, 2009. 76(1): p. 98-104.
87. Fujita, S., et al., Regulatory dendritic cells protect against allergic airway inflammation in a
murine asthmatic model. J Allergy Clin Immunol, 2008. 121(1): p. 95-104 e7.
88. Ogawa, T., et al., A new synbiotic, Lactobacillus casei subsp. casei together with dextran,
reduces murine and human allergic reaction. FEMS Immunol Med Microbiol, 2006. 46(3): p.
400-9.
89. Schouten, B., et al., Cow milk allergy symptoms are reduced in mice fed dietary synbiotics
during oral sensitization with whey. J Nutr, 2009. 139(7): p. 1398-403.


2
Nutrition and food allergy
Bastiaan Schouten1
Betty C.A.M. van Esch1,2
Léon M.J. Knippels2
Linette E.M.Willemsen1
Johan Garssen1,2
1 Department of Pharmacology and Pathophysiology, Utrecht Institute for Pharmaceutical Sciences, Faculty of Science, Utrecht University, Utrecht, The Netherlands
2 Danone Research – Centre for Specialised Nutrition, Wageningen, The Netherlands
To be submitted for publication

26
2
Nutrition and food allergy
Content
Abstract 27 1. Introduction 28 2. Food Allergens 29 3 Diagnosis 31 3.1 Food Challenges 31 3.2 Skin Prick Test 31 3.3 Specific Immunoglobulins 31 4. Dietary intervention 32 4.1 Prevention 34 4.1.1 Breast Milk 34 4.1.2 Hypoallergenic Infant Formulas 34 4.1.3 Introduction of Solid Foods 35 4.1.4 Prebiotics 35 4.1.5 Probiotics 37 4.1.6 Synbiotics 38 4.1.7 Postbiotics 39 4.1.8 Polyunsaturated Fatty Acids (PUFA) 39 4.1.9 Imprinting During Pregnancy 40 4.2 Treatment 40 4.2.1 Epitope Exclusion 41 4.2.2 Hydrolysates 41 4.2.3 Pre-, Pro-, Synbiotics and PUFA 41 4.2.4 Traditional Chinese Herbal Medicine 42 4.2.5 Immune Therapy and Induction of Oral Tolerance 42 5. Conclusions 43 6. References 44

27
2
Chapter 2
Abstract
The increased incidence of food allergies gives rise to higher medical costs, but also warrants more intensive preclinical and clinical research in order to prevent and/or treat food allergy. In general, immunoglobulin (Ig)E and non-IgE mediated food allergies can be distinguished for which major food allergens have been identified. Standard operating procedures in diagnoses of food allergies have been developed and perfected; of which double-blind placebo controlled food challenges are the internationally accepted and recommended golden standard. Many dietary factors have been identified and tested for their potential to alter the outcome of food allergy in both pre-clinical as well as clinical studies. Among these components are prebiotics, probiotics, synbiotics, postbiotics, traditional Chinese herbal medicine, poly unsaturated fatty acids and peptides. There are several concepts that might inhibit the onset and severity of food allergic disorders. The timing of the intervention is a crucial aspect. Intervention during early life and even during pregnancy might interfere with immune development and the consequence in terms of allergy in the offspring. This can be recognized as the so-called immune imprinting hypothesis. This review aims to provide an overview regarding different dietary procedures for either prevention and/or treatment of food allergy.

28
2
Nutrition and food allergy
1. Introduction
There are five different types of immunological hypersensitivity reactions, type I-VI. Type I is immunoglobulin (Ig)E dependent and results in acute (<1 h) symptoms due to mast cell activation upon antibody (especially IgE) receptor crosslinking. Type II, III and IV hypersensitivity reactions result in delayed or intermediate delayed symptoms, 6-92 h, upon allergen challenge. Type II is IgG mediated and directed against cell-surface antigens. Type III is antibody mediated also and is directed against soluble antigens. Type IV hypersensitivity reactions are T-cell mediated and different mechanistic steps are involved in this complex type of hypersensitivity. An important feature is orchestrated by T
H1 cells and results in macrophage activation. Additionally, TH2 cells and eosinophils are involved as well in some subtypes. Finally, cytotoxic T-cells seem to play a role in this inflammatory type IV process as well, although this highly depends on the type of antigen or hapten the cells encounter. Type V uses a stimulating antibody, IgG antibodies react with tissue receptors like in Graves disease and also via antibody dependent cell mediated cytotoxicity. Target cells coated with antibodies are destroyed by specialized killer cells (e.g. NK cells and macrophages), which bear receptors for the Fc portion of the coated antibodies. Food allergy (FA) is the leading cause of anaphylaxis (type I hypersensitivity) treated in hospital emergency facilities in Western Europe and the USA and will be the major focus of this review.
Adverse food reactions include any abnormal reaction resulting from ingestion of a food, or food additive, and might be the result of food intolerances (e.g. lactase deficiency) or food hypersensitivity/allergy [1]. Food hypersensitivity, as stated by the European Academy of Allergy and Clinical Immunology nomenclature task force [2], can be divided in non-allergic food hypersensitivity, in which an immunological mechanism is excluded, and FA, in which an immunological mechanism is defined or strongly suspected. The latter can be divided into IgE-mediated FA (type I) and non-IgE-mediated FA (Figure 1). However, it should be realized that a third category exists as well which is a mixed IgE- and non-IgE mediated group. In the first few years of life the prevalence of food hypersensitivities is highest, affecting about 6% of infants under the age of 3 years [3]. In westernized countries the prevalence of allergic disorders has increased the last decades, however the underlying ethiology is still unclear. Decline in breast-feeding, early introduction of solid foods, construction of air-tight highly-insulated homes and changes in diet (for example reduced intake of n-3 polyunsaturated fatty acids) are probably part of the explanation [4]. In addition, the ‘hygiene’ or ‘microbial deprivation’ hypothesis is suggested, which states that a lower exposure to infectious agents during early childhood might cause a rapid rise in atopic disorders due to defective development of the adaptive immune system [5].

29
2
Chapter 2
2. Food Allergens
Among hundreds of different proteins in the human diet numerous are identified as allergens. Only a small part of them is responsible for the majority of hypersensitive adverse food reactions. The eight most common food allergies are cow’s milk, egg, peanut, tree nuts, fish, shellfish, soy and wheat allergy. The ‘big eight’ food allergens are well described and characterized (Table I).
Cow’s milk allergy has been regarded by pediatricians as a typical FA for several centuries [6]. It is the first allergy that children can develop in life, usually within their first year of life, and affects 2-3% of the children; 4 out of 5 children develop clinical tolerance by their fifth year of life. About 60% of these patients have cow’s milk specific IgE-mediated reactions [7, 8].
Chicken egg protein allergy is the second most common cause of FA in children, with an estimated prevalence of 2.5% [3, 9]. The majority of children outgrow their food allergy by school age [10].
Peanut allergy is regarded as the most severe, most dangerous type of food allergy. Most reported cases of anaphylaxis are caused by peanut, often induced by almost deniable doses. In contrast to cow’s milk allergy this allergy remains for life. The prevalence rates excess 1% [11, 12].
Nuts are known for their beneficial health effects since they may reduce the risk of cardiovascular disease and diabetes. Nuts have many favorable ingredients such as vitamins,
Figure 1. Nomenclature of food hypersensitivity (adapted from [2]).
Adverse food reactions
Intolerancee.g. lactose intolerance
Food hypersensitivity
Food allergy Non allergic food hypersensitivityimmune system not involved
IgE‐mediated food allergye.g. immediate type of food allergy
Non‐IgE‐mediated food allergye.g. delayed type of food allergy

30
2
Nutrition and food allergy
polyunsaturated fatty acids (PUFA) and monounsaturated fatty acids [13]. However, many tree nuts also can cause severe or less severe allergic reactions; the most common allergic nuts are hazelnut, walnut, cashew and almond. Other nuts like Pecan, Brazil, macadamia, pistachio, chestnut and coconut cause less severe allergic reactions [14, 15].
Fish and shellfish (seafood) allergy comprises 3 divisions of sea organisms, the Chordata, Mollusca and the Arthropoda, these include different kinds of fish (vertebrates), shrimps, crab (Crustacea) and mollusks [16, 17]. Europe, Spain, Portugal and obviously the Scandinavian countries have the highest fish consumption and prevalence of fish allergy. [18].
Soybean is a member of the legume family and an important nutrient, in particular in formula feeding as a substitute for cow’s milk [19, 20]. Soy allergy has been described primarily in young children with atopic dermatitis (AD) [21]. The immunological and the clinical basis is highly complex, which complicates the diagnosis of soy allergy and the advice in regard to risk management.
Table I. Major food allergens of the ‘big eight’ that have been identified.
Food allergy Major allergens (protein class) References
Cow’s milk allergy β-lactoglobulin, α-Lactalbumin, Bovine serum albumin (BSA), αS1-casein, αS2-casein, β-casein, κ-casein
[1, 25]
Chicken egg white allergy
Gal d 1 (ovomucoid), Gal d 2 (ovalbumin), Gal d 3 (ovotransferrin), Gal d 4 (lysozyme), Gal d 5 (α-Livetin)
[1, 26, 27]
Peanut allergy Ara h 1 (vicilin); Ara h 2, Arg h 6, Arg h 7 (conglutin); Ara h 3, Arg h 4 (glycinin); Ara h 5 (profilin); Ara h 8 (nonfusion protein); peanut oleosin
[28-33]
Tree nut allergy Cas s (chestnut), Cor a (hazelnut), Ber e (brazil nut), Jug n (black walnut), Jug r (English walnut), Ana o (cashew nut)
[1, 14, 15]
Seafood allergy (fish and shellfish allergy)
Met e 1 (shrimp), Hom a 1 (American lobster),Pan s 1 (spiny lobster), Cha f 1 (crab), Sal s 1 (Atlantic salmon), Gad c 1 (cod), Sco s 1 (mackerel)
[1, 16, 17, 34]
Soy allergy Gly m 1.0101 (hydrophobic protein), Gly m 2 (hull protein), Gly m 3 (profilin), Gly m 4, Gly m BD 30 k (vacuolar protein), glycinin, vicilin, β-conglycinin, Kunitz trypsin inhibitor
[1, 35, 36-38]
Wheat allergy Serpin, α-amylase inhibitor, γ-gliadin, low molecular weight glutenin
[39, 40]

31
2
Chapter 2
Wheat allergy and celiac disease are two different disorders. Celiac disease is caused by gluten intolerance which is believed to be in part non-immune mediated. However, gluten reactive T-cells have also been implicated in the pathophysiology of the disease [22]. In contrast, wheat allergy is an IgE mediated allergy. There are two forms of wheat allergy, bakers’ asthma (occupational asthma caused by flour inhalation) and food allergy to wheat proteins [23, 24].
3. Diagnosis
3.1 Food Challenges
The most effective approach to determine whether a person is allergic to food ingredients is by an oral challenge with the particular food. The risk of an anaphylactic reaction is always present and caution is needed when performing such a diagnostic food challenge. The oral food challenge can be performed open, single-blind placebo controlled or double-blind placebo controlled (DBPCFC) [41]. The latter is the golden standard for research and clinical purposes. Subjective symptoms are eliminated with the DBPCFC test since both the clinician and patient are unaware whether the placebo or food of interest is tested [42, 43]. Difficulties with the single-blind and the DBPCFC is to mask the food and flavor. Often this is performed by using another food or capsules.
3.2 Skin Prick Test
The skin prick puncture test (SPT) is commonly used to detect sensitization to certain foods. This SPT is performed to screen patients with suspected IgE-mediated FA. Food extracts are diluted and applied and the skin is punctured with a needle through the epidermis [44]. A positive (histamine) and negative (saline) control are applied as well. A patient is sensitized for a certain food when the wheel diameter is 3 mm or larger compared to the negative control. When compared to the DBPCFC the positive predictive accuracy of the SPT is less than 50% [45].
3.3 Specific Immunoglobulins
There are several analytical tools to determine allergen specific serum IgE levels. However

32
2
Nutrition and food allergy
the most recent, best validated, common one is the Pharmacia CAP-System FEIA. The detection limit of this system is 0.35 kU/L, therefore patients with lower levels are regarded as non-sensitized and patients with IgE levels above 0.35 kU/L are regarded as sensitized [46-49]. Although allergen specific IgE is typically involved in induction of the allergic symptoms other isotypes of allergen specific immunoglobulins (Ig) such as IgG have the properties to block the binding of IgE and therefore inhibit mast cell degranulation [50]. Ruiter et al. [51] demonstrated that the maintenance of oral tolerance to cow’s milk is characterized by high levels of cow’s milk specific IgG
4 in combination with low levels of cow’s milk specific IgE. Therefore, measurements of specific immunoglobulins will be more useful if all isotypes are determined and taken into account. Especially ratios between IgE and IgG isotypes might be predictive for sensitization linked to allergic symptoms and to tolerance.
A subgroup of patients do have the symptoms of immediate (mast cell mediated) type of hypersensitivity, however they lack the presence of specific IgE. There are indications that immunoglobulin free light chains are involved in some hypersensitivity disorders [52]. In particular preclinical data are available [53-55], however there are a few clinical studies in relation to immunoglobulin free light chains as well [56] (Schouten et al. submitted). Complete antibodies have light chains integrated in their structure, however immunoglobulin light chains are released as singular (free) molecules also. Immunoglobulin light chains are produced and secreted by B lymphocytes or plasma cells. Also Immunoglobulin light chains bind to mast cells and upon encounter with the specific allergen the mast cell degranulates and a type I hypersensitivity reaction is induced [57, 58]. More data regarding immunoglobulin free light chains are needed to address their utility as a predictive and/or diagnostic biomarker.
4. Dietary intervention
One of the major focus areas in this review is the possibility of dietary intervention to prevent or treat FA (Figure 2). Preventive intervention protocols preferably start early in life or importantly already during pregnancy since this might affect the immune system of the offspring. This phenomenon is called immune imprinting and as a consequence of the mother’s dietary restrictions or additives the offspring will develop less, or less severe, allergy. Currently, large cohort studies are being conducted to find evidence for this hypothesis.

33
2
Chapter 2
Figure 2. Schematic representation of the sensitization and effector phase occurring in food allergic patients. These are respectively targets of prevention (4.1) and treatment (4.2) using dietary intervention to control food allergy. Instead of oral tolerance, food allergic patients develop an IgE mediated allergic reaction towards harmless food antigens. Prevention of development of allergy should take place before or during the sensitization phase. Dietary intervention may target allergen presentation to naïve T-cells in the GALT (T
H1 instead of allergic TH2 polarization), enhance regulatory T-cell function (suppresses effector response) and/or reduce allergen specific IgE production by B-cells. Possible preventive approaches involve breast milk (4.1.1), hypoallergenic infant formulas (4.1.2), timing of introduction of solid foods (4.1.3), prebiotics (4.1.4), probiotics (4.1.5), synbiotics (4.1.6), postbiotics (4.1.7), polyunsaturated fatty acids (PUFA) (4.1.8) and imprinting during pregnancy (4.1.9). There might be overlap between the preventive and the treatment strategies. Treatment protocols focus on reduction of mast cell degranulation, which may occur direct or indirect via altered or reduced antibody production by B-cells or support of regulatory T-cell function. Ultimately oral tolerance is induced (desensitization). Possible treatment approaches include epitope exclusion (4.2.1), the use of hydrolysates (4.2.2), pre-, pro, synbiotics and PUFA (4.2.3) and traditional Chinese herbal medicine (4.2.4). PP: Peyer’s Patch; MLN: mesenteric lymph node; apc: antigen presenting cell; Treg: regulatory T-cell; T
H1: T helper 1 cell; TH2: T helper 2 cell.

34
2
Nutrition and food allergy
4.1 Prevention
Strategies to prevent or reduce sensitization for food allergens include dietary restriction (allergen avoidance) or dietary intervention to actively support the process of naturally occurring oral tolerance induction via improved immune development.
4.1.1 Breast Milk
The best nutritional intervention in early life is breastfeeding. It provides a unique combination of lipids, proteins, carbohydrates, vitamins, and minerals. Furthermore, there are numerous bioactive components in it with immunological properties like soluble IgA, antioxidants, oligosaccharides, bacterial fragments, Toll-like receptor ligands, cytokines, hormones, fatty acids and many more. Each component can individually, additionally or synergistically act on the immune system of the neonate. The breast milk content changes over time to ensure optimal passive and active protection and growth for the child [59-61]. However, the quality of the breast milk might vary between mothers and therefore also the effectiveness in the induction of oral tolerance. Little is known about the influence of dietary intervention on the quality of breast milk and more preclinical as well as clinical data are needed. Improvement of the quality of breast milk via a healthy balanced diet of the mother is a promising approach. It is known, however, that food antigens as well as IgE can be transferred to breast milk and therefore potentially can act as a risk factor for children genetically more susceptible for developing allergic disease [60]. Usually, the advantage of breast feeding is larger than the potential risk of sensitization.
4.1.2 Hypoallergenic Infant Formulas
Hypoallergenic infant formulas play a crucial role in the prevention of allergies in high risk children as well as in the treatment (4.2.2). Prevention might occur either by avoidance of the allergen early in life or by the induction of oral tolerance. It is believed that avoidance of the allergen might be a good approach for the prevention of allergy. However, for example cow’s milk allergy might predispose the development of other allergies later in life. Therefore active induction of immunological oral tolerance may be a preferable approach, since it may also positively affect the atopic constitution of the child. Hypoallergenic infant formulas or hydrolysates are categorized into partial and extensive hydrolysates, based upon the molecular weight and length of the remaining polypeptides which determines the degree of allergenicity. The allergenicity of proteins can be reduced using enzymatic hydrolysis,

35
2
Chapter 2
heat treatment and/or ultra filtration [62, 63]. Ultimately a third formula can be used consisting single amino acids only and cannot cause any hypersensitivity response, which is allergen avoidance. The main disadvantage of this formula is that it cannot induce any immunological oral tolerance [64]. A T cell epitope consists of 8-25 amino acids, depending on the MHC class. Single amino acids are therefore not capable of inducing any immune responsiveness and thus cannot induce immunological (oral) tolerance. Most of the (pre-)clinical studies that are performed to prevent allergic sensitization in infants at risk focus on the hydrolysates with low allergenic potential which may still poses tolerogenic capacities [65-67]. A difficulty in interpreting clinical results of different formulas is the heterogeneity in family history of allergies [68]. A formula, partial or extensive hydrolysate based, that shows no allergenic potential, but can still induce oral tolerance is the ultimate goal in this field of nutritional research.
4.1.3 Introduction of Solid Foods
Another factor that might influence the onset of allergies is the timing of introduction of solid foods. Initially much debate was going on regarding the introduction of solid foods, however there is consensus to avoid solid food before 17 weeks of age and to start no later than 26 weeks [69-71]. With the introduction of complementary feeding (e.g. nutrition other than derived from breast milk and infant formula) the risk for allergies is rising. Therefore caution is necessary, for children at risk for allergies, with the introduction of potential clinical significant allergens which can be found in peanuts, egg white, whole milk and (shell)fish. The late introduction of complementary foods cannot fully prevent the onset of allergies [71].
4.1.4 Prebiotics
Human breast milk contains approximately 7-12 g/L oligosaccharides [72]. At least 130 different oligosaccharides have been isolated from human milk and the two main categories are neutral and acidic oligosaccharides [73]. These oligosaccharides are non-digestible carbohydrates, or dietary fibers as they are frequently named, that have many different properties and are believed to act on the microbiotia in the gut [74-77]. Due to physicochemical properties of non-digestible carbohydrates the absorption of minerals and fecal consistency improves [78]. There are different types of dietary fibers e.g. (hemi)cellulose, lignin, β-glucans, pectins, gums, inulin and oligosaccharides [79]. In addition, different non-digestible oligosaccharides with specific properties are obtained or manufactured from

36
2
Nutrition and food allergy
natural sources [80]. Some of them have specific properties and can be used as prebiotics in a dietary supplement. Prebiotics are defined as “food ingredients that beneficially affect the host by selectively stimulating the growth and/or activity of one or a limited number of bacterial species already resident in the colon, and thus attempt to improve host health” [74]. Recently this definition is adapted to: “A prebiotic is a selectively fermented ingredient that allows specific changes, both in the composition and/or activity in the gastrointestinal microflora that confers benefits upon host wellbeing and health” [81]. Prebiotics enhance defense mechanisms of the host by stimulation of growth of Bifidobacteria and Lactobacilli [82] Short-chain fatty acids, released by these bacteria upon fermentation of prebiotics, are essential nutrients for intestinal epithelial cells and support gut function [83]. In vivo and in vitro studies have shown beneficial effects of prebiotics on the innate as well as the adaptive immune system [84].
Short-chain galacto-oligosaccharides (scGOS), long-chain fructo-oligosaccarides (lcFOS) and pectin-derived acidic oligosaccharides (pAOS) are some examples of non-digestible oligosaccharides that mimic the functionality and molecular size distribution of human milk oligosaccharides. Prebiotic scGOS are produced by enzymatic elongation of galactose (derived from lactose) by β-galactosidase, with a degree of polymerization (dp) of 3–8. FOS is derived from chicory inulin which is a polymer from D-fructose residues linked by β(2-1) bonds with a terminal α(1-2) linked D-glucose (dp 2-60). lcFOS are produced from inulin and have a mean dp >23, whereas scFOS have a mean dp <23. lcFOS are added to infant formula. pAOS is a mixture of linear oligomers and small polymers of galacturonic acid produced from from food grade pectin by pectin cleaving enzymes and have a dp of 1-20. Currently, it is not feasible to produce oligosaccharides identical to human milk oligosaccharides present in breast milk, however the used specific oligosaccharides (scGOS and lcFOS) have proven efficacy in clinical trials since they reduce the incidence of allergic disease and have been shown to enhance fecal Bifidobacteria counts [85-88].
The effects of scGOS and lcFOS (Immunofortis®) have been studied in a murine vaccination model [89, 90], in an allergic asthma model [91] and a cow’s milk allergy model [92, 93]. The response to vaccination of mice fed the scGOS/lcFOS diet was significant enhanced, as well as the fecal Bifidobacteria and Lactobacilli proportions in colon samples [90]. In addition, pAOS enhanced the murine vaccination response and the combination (scGOS/lcFOS/pAOS) was even more effective [89]. In infants ,pAOS is considered to be safe with regard to intestinal flora, stool characteristics, pH, growth, crying, vomiting and regurgitation patterns as compared to control infants [94]. Furthermore, systemic T
H1 dependent immune responses were enhanced using the prebiotics without inducing autoimmunity, as TH1 is low in newborn infants. On the other hand in an ovalbumin model for allergic asthma, which is TH2 dependent, the identical oligosaccharides reduced airway hyperresponsiveness and

37
2
Chapter 2
lowered influx of cells in the broncho-alveolar lavage fluid [91]. In addition, in a murine model for cow’s milk allergy dietary intervention with scGOS/lcFOS showed a significant decrease of the allergic response and increased specific IgG2a levels [93]. Another prebiotic fructo-oligosaccharide, kestose, showed to be effective in reducing the incidence of AD in infants as well [95].
Recently it was found that dietary intervention with scGOS/lcFOS/pAOS reduces the development of an acute allergic response upon antigen challenge, although specific immunoglobulins levels remain high. Both in vivo as well as ex vivo depletion of CD25+ Treg cell abrogated the diminished acute allergic response and imply crucial involvement of antigen specific CD25+ Treg cells in the suppression of the allergic effector response (Schouten et al., submitted).
Furthermore, clinical studies have been performed with the scGOS/lcFOS mixture in children at high risk for allergies. A reduction in the incidence of AD [85] and the incidence of allergic manifestations during the first 6 months of life [87] was observed, furthermore this reduction lasted at least until 2 years of age [86]. In another clinical study, using the scGOS/lcFOS mixture, fecal secretory IgA was increased in healthy infants [96]. There is cumulative evidence that prebiotic mixtures might beneficially affect the host in both T
H1 as well as TH2 prone settings as it might prevent food allergy (TH2) and enhances the vaccination response (TH1). Although it is believed that prebiotics exert their effect via stimulation of growth of selective bacterial species that beneficially improve host health, there is debate about this mechanism and there are potentially microbiota-independent mechanisms as well [84].
4.1.5 Probiotics
Probiotic means ‘for life’ and probiotics can be defined as: “live microbial food ingredients that alter the enteric microflora and have a beneficial effect on health” [97]. For the prevention of allergic disease probiotics are widely investigated in multiple clinical studies [98-108]. However, there is still no consensus regarding the efficacy of these bacteria in having immune-modulatory effects and their clinical relevance [109]. Several factors are able to influence the outcome of the studies, e.g. candidate probiotic strain or mixture, environmental factors, intervention duration, and timing and bacterial load [110, 111].
Among young infants 10% will develop AD, which can objectively be scored and approximately one-third of these children are affected with FA [112, 113]. When pregnant mothers with a family history of allergic disease and their babies were supplemented, for 6 mo, with Lactobacillus GG, the cumulative incidence of atopic disease was reduced after 2 y compared

38
2
Nutrition and food allergy
with placebo controls [103]. The supplementation was still effective after 4 y and even 7 y [101, 107]. In a similar study, Lactobacillus reuteri reduced the incidence of IgE-associated eczema and the occurrence of a positive skin prick test for almost 50% [105].
In germ-free mice the fundamental contribution of the gastrointestinal microbiota in the induction of oral tolerance was confirmed. Furthermore, the gastrointestinal microbiota composition may be a prerequisite for the development of allergy [114-116].
To gather more data regarding underlying mechanisms of immune modulation by probiotics animal models are widely used [93, 117-121]. The candidate probiotic strain B. breve M-16V effectively inhibited the development of the acute allergic skin response in mice orally sensitized with whey and increased serum levels of whey-specific IgG
2a, whereas no decrease in IgE and IgG1 levels was observed [93]. In addition, it was shown that supplementation of UV-killed bacteria of the same strain during the sensitization phase had similar effects (Schouten et al., unpublished data). In an ovalbumin induced mouse model oral supplementation of L. acidophilus, B. lactis, B. bifidum as well as L. casei, but not E. coli decreased ovalbumin-specific IgE levels [118, 119]. Furthermore, Feleszko et al. [121] showed that L. rhamnosus GG and B. lactis decreased the antigen specific recall proliferation of splenocytes as well as T
H2 type cytokine production of mesenteric lymph node cells in a mouse model of asthma.
These preclinical models can bridge the gap between in vitro selection of specific candidate probiotics and use in clinical studies. Furthermore, they can provide more insight in the underlying mechanisms of action.
4.1.6 Synbiotics
To improve human health and reduce the incidence of allergies in a natural way the combination of prebiotics and probiotics, called synbiotics, is being investigated as well. The hypothesis that pre- and probiotics together are more effective than the sum of single preparations is very promising, however not many studies have been performed in relation to allergy [93, 122-124]. Recently, in a mouse model of orally induced cow’s milk allergy indeed preventive dietary intervention with a synbiotic mixture of scGOS/lcFOS and B. breve MV-16 was found to be significantly more effective in reducing several allergic parameters when compared to intervention with pro- or prebiotics alone [93]. Hence, the (preclinical) studies that have been carried out show potential in reducing the incidence and severity of allergic disorders. Timing and dosage of both the prebiotics as well as the probiotics need further attention. In general, more research is needed to confirm the additional effects of

39
2
Chapter 2
combining prebiotics and probiotics.
4.1.7 Postbiotics
An emerging field in nutritional sciences is the supplementation of isolated bacterial products called postbiotics (ferments). These products include short-chain fatty acids, like butyrate, however other more unknown products (e.g. cytokines, toll like receptor ligands, carbohydrates and bacterial glycans) may also have important immunomodulatory effects [125]. So far, to our knowledge, there is no literature available regarding the use postbiotics in FA. However, in a stress-induced gut permeability rat model a probiotic strain in combination with its bacterial (or fermentation) products synergistically counteracted the stress-induced gut permeability and visceral sensitivity [126]. These data may open new avenues in the research concerning prevention or treatment of food allergies with postbiotics whether or not in combination with pre- pro- or synbiotics.
4.1.8 Polyunsaturated Fatty Acids (PUFA)
The increased prevalence of childhood allergic diseases coincides with altered dietary fatty acid intake in westernized countries. N-3 (alpha-linolenic acid) and n-6 PUFA (linoleic acid) are essential for humans and have to be taken up via the diet. PUFA are incorporated into the cellular membrane and are eicosanoid precursors hereby affecting the immune response. In this regard in particular long chain n-3 PUFA (EPA and DHA), which are obtained via diet or synthesized from alpha-linolenic acid, are regarded to be anti-inflammatory while n-6 PUFA are able to boost inflammatory responses. In westernized countries the n-3 fatty acid ingestion is no longer favored over omega-6 PUFA which may have implications for immune homeostasis. It has been stated that the original diet of the human race consists of a n-6 : n-3 ratio of approximately 1:1 and that this has changed over the past decades to a ratio of at least 15:1 [127]. This shift is suggested to be explanatory for the increased incidence of cardiovascular diseases, chronic inflammatory diseases and also allergic diseases. Therefore pre-clinical as well as clinical studies are performed in order to investigate this hypothesis. Several clinical studies have shown that enhanced n-3 PUFA intake reduces the incidence of allergic disorders, e.g. AD, less sensitization to egg (reviewed by [128]). Two studies by Dunstan et al. [129, 130] show reduced sensitization to egg and less AD when pregnant woman were supplemented with fish oil. Recently, Furuhjelm et al. showed that maternal n-3 PUFA intake decreased the risk of FA and IgE-associated eczema in children at risk for allergy [131]. In contrast, in a clinical study by Almqvist et al. [132] it is shown that supplementation

40
2
Nutrition and food allergy
of n-3 PUFA, starting at a maximum of six months of age, did not prevent children with a family history of asthma from developing atopy, eczema nor asthma at the age of 5 years. Hence, discrepancy on effects of n-3 PUFA in prevention of allergic disease exists. However, in a review by Prescott and Calder [133] it was put forward that such dietary factors are under-explored and that there is a need for novel synthetic PUFA which may be potent in the reduction of the allergic burden. One of the strategies that are proposed is the use of selected PUFA in the formula feeding of young children at high risk for allergies [134].
4.1.9 Imprinting During Pregnancy
In order to prevent an allergic outcome intervention during pregnancy has growing interest. The prenatal period is crucial in the priming of the immune system and therefore it may alter the susceptibility for development of allergies later in life. All of the above discussed dietary components, like pre-, pro-, synbiotics and PUFA have been studied for their immunomodulatory capacities [98, 128, 130, 135, 136, 137, 138]. Regarding pre- and probiotics the effects are variable. Kukkonen et al. [98] have shown that supplementation of pregnant women with a mixture of probiotics and supplementation of their offspring with prebiotics and the probiotics, decreased the incidence of atopic eczema. However a study by Boyle et al. [139] with one probiotic strain (Lactobacillus rhamnosus GG) supplemented only to the mothers (from 36 weeks gestation until delivery) did not result in any beneficial antigen-specific immune response of the neonate. Also enhanced dietary intake of n-3 PUFA during pregnancy may be able to alter the immune status of the neonate. PUFA can pass the placental barrier and are incorporated in the membranes of cells of the fetus [140]. Dietary intervention during pregnancy shows promising results, however these results are highly variable depending on the experimental setup. Therefore more studies with larger cohorts are necessary and if possible with dietary mixtures consisting of both synbiotics and PUFA.
4.2 Treatment
Besides prevention of the development of allergic disease treatment is essential as well. However, the immunological network is highly complex and although there are numerous pathways to intervene FA; e.g. anti-IgE therapy, corticosteroids, mast cell stabilizers, leukotriene inhibitors, TGF-β administration and anti-IL-5 therapy [141, 142], those interventions are still experimental. Here, the focus will be on the methods already used in the clinic, daily practice and dietary intervention studies with the objective to treat or reduce symptoms of allergic disease and in particular FA. For treatment both avoidance (epitope exclusion) as well as desensitization strategies are important.

41
2
Chapter 2
4.2.1 Epitope Exclusion
The single most effective strategy to prevent an adverse allergic reaction in food allergic individuals is by exclusion diets, which is avoidance of the allergen(s) [143]. If there is a suspicion of FA it is advised to eliminate several major food allergens from the diet. After several days there should be relief of symptoms and if there is relief one by one the different foods are introduced again until an allergic reaction occurs. Exclusion of possible allergens may be used for the management of FA. However, it should be ensured that during a multiple food exclusion diet the actual diet is still nutritionally adequate.
4.2.2 Hydrolysates
In general there are two different types of milk (infant formula) hydrolysates. In the USA, in particular casein protein based formulas are available, whereas in Europe most formulas are whey protein based. Partial hydrolysates (see also 4.1.2) contain peptides with a molecular weight ranging between 3.000 and 10.000 Da, while the extensive formula have generally peptide lengths under 3.000 Da [144]. Both partial as well as extensive hydrolysates can be whey or casein based. In some cases these small proteins are still able to trigger an allergic reaction [62, 63], Besides the two hydrolysate formula feedings, soy-based and amino acid-based formulas can be used [64]. The main advantage of soy- and amino acid-based formulas is that no cow’s milk allergenic potential is present, though oral tolerance to cow’s milk proteins cannot develop either, since there are no T-cell epitopes present.
4.2.3 Pre-, Pro-, Synbiotics and PUFA
To stimulate the immune system with the intention to prevent the onset of an allergic TH2 phenotype pre-, pro-, synbiotic and PUFA are under investigation. However, similar strategies are used for treatment with the intention to skew the immune response towards a more TH1/Treg polarized phenotype after the onset of allergy resulting in desensitization and ultimately oral tolerance induction. In parallel, modulation of the immune system using dietary components is highly interesting and might be helpful in order to induce a tolerogenic immune environment. In spite of the common belief that the intervention should take place as early in life as possible, several studies have shown successful treatment of AD in children above 2 years of age, suggesting that the immune system can still be influenced later in life [145-147]. In contrast several other studies show no positive effect of a probiotic intervention on AD [100, 106, 148]. Brouwer et al. [106] tested two different strains (Lactobacillus

42
2
Nutrition and food allergy
rhamnosus and Lactobacillus GG) and found no effect on AD in infants enrolled before the age of 5 years. Sistek et al. [100] tested the combination of Lactobacillus rhamnosus and Bifidobacteria lactis and found only a decrease in AD in a subgroup of children enrolled between age 1 and 10 years. Not much is known about the possible curative properties of PUFA in allergic disease however studies are being conducted to investigate this [135]. One study showed that female adults with high fish and DHA intake have a lower rate in allergic sensitization [149]. Use of pre-, pro-, synbiotics and PUFA to ameliorate or treat allergic disease may be advantageous, however the need for more and comprehensive (pre-)clinical data is essential.
4.2.4 Traditional Chinese Herbal Medicine
Complementary to regular medicine, alternative medicine have growing interest in the western world especially in the field of allergic asthma. Additionally scientific evidence for the treatment of FA is increasing [150, 151]. A few studies in preclinical models for FA using a traditional Chinese herbal formula have been published [152-154]. Two different formulas have been tested; the FA herbal formula (FAHF)-1 and FAHF-2, which are mixtures of 9-11 different herbs. Classically, these herbs were prescribed for gastrointestinal disorders, as diarrhea and vomiting and therefore thought to be effective in FA [151]. In the preclinical model these herbs were highly effective in protecting the animals from anaphylaxis after oral peanut challenge and reducing peanut-specific IgE levels [152, 153]. Currently, a phase I clinical trial is ongoing to test the safety of FAHF-2 in humans and phase II is planned to determine its efficacy in peanut allergic patients [141, 150].
4.2.5 Immune Therapy and Induction of Oral Tolerance
A more clinical approach in the treatment of FA is immune therapy. Several different procedures are under investigation. Subcutaneous injections of increasing doses of food allergen extracts have been shown to induce unacceptable side-effects, in contrast with other non-food allergens [155]. Other disadvantages are the introduction of sensitization to new allergens present in the used extracts and it is not effective in all patient groups [156]. To circumvent this major drawback, two alternative immunotherapies are being developed. In oral immunotherapy (OIT) also known as specific oral tolerance induction (SOTI) the allergen is being swallowed directly and in sublingual immunotherapy (SLIT) the allergen dose is held under the tongue for a few minutes before swallowing [142]. The dosages are increasing over time and the duration of these therapies is several months upto two years. Besides

43
2
Chapter 2
the effects on the clinical outcome of a provocation test, also some studies indicate that allergen specific IgE levels are altered, mostly decreased, which indicates desensitization [157-159]. In particular SOTI has been investigated with interesting results. There are two different mechanisms with regard to oral tolerance induction; low dose and high dose. Low dose tolerance is, in the rat, induced by orally applied multiple doses (1 mg) of the specific allergen, while high dose oral tolerance (20 mg) is induced after a single dosage [160]. Active suppression by means of regulatory T cells underlies the mechanism of action of low dose oral tolerance [161, 162]. Allergies develop as a consequence of a disparity in the T
H1/TH2 balance towards TH2 [163, 164]. Different Treg cell subsets (CD4+CD25+FOXP3+, TH3 and Tr1) actively regulate this balance locally in the gastro-intestinal tract via excretion of immunosuppressive cytokines IL-10 and TGF-β and via cell-cell contact. Antigens, or dendritic cells loaded with antigen, travel from the Peyer’s patches to the mesenteric lymph nodes (MLN) and are presented to naïve T-cells. Antigen-responsive T-cells (CD4+) travel subsequently through the bloodstream and home back into the intestinal mucosa or traffic to the peripheral immune system (e.g. spleen) [165, 166]. Antigen specific T-cells from the MLN in this respect can relocate systemic tolerance. There are several host factors contributing to the success of oral tolerance induction e.g. genetics, age and microbiotic flora [167]. For SOTI the allergen of interest is administered orally, from a very low dose which is daily increased. After a few months the amount of food that is tolerated is equivalent to a usual daily quantity. However a daily maintenance dose is required to remain tolerant [168]. The mechanism by which SOTI exerts its effect is not clear yet and it remains to be investigated whether specific immunological mechanisms underlie the increase in threshold dose. Hence, both oral and sublingual immunotherapy may be usefull for food allergic patients, however the protocols are time consuming and sometimes with transient efficacy. Although oral immunotherapy could be an interesting approach, more studies are needed to understand mechanisms of action.
5. Conclusions
Food allergies are of growing health concern in the Westernized world. Together with the rising incidence of food allergies the medical costs rise. If oral tolerance to a specific allergen is not induced, a subject becomes sensitized. After a recurrent encounter with the same allergen a clinical reaction is provoked, since mast cells degranulate and even anaphylaxis may occur. FA can be diagnosed, for example, by analyzing serum specific IgE [46-48]. Furthermore, a DBPCFC or a SPT may be performed to establish a reliable diagnose [41, 44]. In addition, Ig-fLC measurements may add to the diagnosis of food allergies [58], Schouten et al., submitted). Pre-clinical and clinical research on FA is expanding. Currently, no medicines to prevent or cure food allergies are available, however dietary interventions are safe and

44
2
Nutrition and food allergy
may prevent or ultimately cure food allergies as well. Therefore multiple approaches are studied, in particular for the prevention of allergies in children, using prebiotics, probiotics, synbiotics, postbiotics and PUFA. These food supplements are sometimes administered before birth, to the pregnant women, and are believed to induce beneficial immune imprinting in the neonate [98, 139].
Promising for the prevention of AD, which is related to food allergies, are the prebiotic mixtures of non-digestible carbohydrates and/or the use of specific probiotic strains. Future studies will have to reveal whether combined preparations of multiple dietary supplements, e.g. synbiotics and PUFA will further improve the efficacy.
FA is most probably caused by an imbalance between TH1, TH2 and Treg cell activity. The
mechanisms by which the different dietary interventions act are currently unclear, however enhanced CD25+ regulatory T cell function may be one of possibilities (Schouten et al., submitted). Research on food allergies should not only focus on testing more potential dietary components, but on the working mechanism also. If there is more insight in the mechanism, selection of food components might be easier. Multidisciplinary approaches of immunologists, nutritionists and allergists would help in a better understanding and approach to prevent and treat food allergy.
6. References
1. Sampson, H.A., Food allergy. Part 1: immunopathogenesis and clinical disorders. J Allergy
Clin Immunol, 1999. 103(5 Pt 1): p. 717-28.
2. Johansson, S.G., et al., A revised nomenclature for allergy. An EAACI position statement from
the EAACI nomenclature task force. Allergy, 2001. 56(9): p. 813-24.
3. Sampson, H.A., Update on food allergy. J Allergy Clin Immunol, 2004. 113(5): p. 805-19; quiz
820.
4. Chandra, R.K., Food allergy and nutrition in early life: implications for later health. Proc Nutr
Soc, 2000. 59(2): p. 273-7.
5. Bloomfield, S.F., et al., Too clean, or not too clean: the hygiene hypothesis and home hygiene.
Clin Exp Allergy, 2006. 36(4): p. 402-25.
6. Eigenmann, P.A., The spectrum of cow’s milk allergy. Pediatr Allergy Immunol, 2007. 18(3):
p. 265-71.
7. Rhodes, H.L., et al., Early life risk factors for adult asthma: a birth cohort study of subjects at
risk. J Allergy Clin Immunol, 2001. 108(5): p. 720-5.
8. Host, A., Frequency of cow’s milk allergy in childhood. Ann Allergy Asthma Immunol, 2002.
89(6 Suppl 1): p. 33-7.

45
2
Chapter 2
9. Heine, R.G., N. Laske, and D.J. Hill, The diagnosis and management of egg allergy. Curr Allergy
Asthma Rep, 2006. 6(2): p. 145-52.
10. Kemp, A.S., Egg allergy. Pediatr Allergy Immunol, 2007. 18(8): p. 696-702.
11. Sicherer, S.H. and H.A. Sampson, Peanut allergy: emerging concepts and approaches for an
apparent epidemic. J Allergy Clin Immunol, 2007. 120(3): p. 491-503; quiz 504-5.
12. Burks, A.W., Peanut allergy. Lancet, 2008. 371(9623): p. 1538-46.
13. Allen, L.H., Priority areas for research on the intake, composition, and health effects of tree
nuts and peanuts. J Nutr, 2008. 138(9): p. 1763S-1765S.
14. Roux, K.H., S.S. Teuber, and S.K. Sathe, Tree nut allergens. Int Arch Allergy Immunol, 2003.
131(4): p. 234-44.
15. Crespo, J.F., et al., Food allergy: nuts and tree nuts. Br J Nutr, 2006. 96 Suppl 2: p. S95-102.
16. Lehrer, S.B., R. Ayuso, and G. Reese, Seafood allergy and allergens: a review. Mar Biotechnol
(NY), 2003. 5(4): p. 339-48.
17. Sicherer, S.H., A. Munoz-Furlong, and H.A. Sampson, Prevalence of seafood allergy in the
United States determined by a random telephone survey. J Allergy Clin Immunol, 2004.
114(1): p. 159-65.
18. Pascual, C.Y., et al., Fish allergy in childhood. Pediatr Allergy Immunol, 2008. 19(7): p. 573-
9.
19. Turck, D., Soy protein for infant feeding: what do we know? Curr Opin Clin Nutr Metab Care,
2007. 10(3): p. 360-5.
20. Ballmer-Weber, B.K., et al., Clinical characteristics of soybean allergy in Europe: a double-
blind, placebo-controlled food challenge study. J Allergy Clin Immunol, 2007. 119(6): p.
1489-96.
21. Matricardi, P.M., et al., Primary versus secondary immunoglobulin E sensitization to soy and
wheat in the Multi-Centre Allergy Study cohort. Clin Exp Allergy, 2008. 38(3): p. 493-500.
22. Raki, M., et al., A unique dendritic cell subset accumulates in the celiac lesion and efficiently
activates gluten-reactive T cells. Gastroenterology, 2006. 131(2): p. 428-38.
23. Tatham, A.S. and P.R. Shewry, Allergy to wheat and related cereals. Clin Exp Allergy, 2008.
24. Jacobs, J.H., et al., Wheat allergen exposure and the prevalence of work-related sensitization
and allergy in bakery workers. Allergy, 2008. 63(12): p. 1597-604.
25. Monaci, L., et al., Milk allergens, their characteristics and their detection in food: a review.
European Food Research and Technology, 2005(Volume 223, Number 2 / June, 2006): p.
149-179.
26. Jacobsen, B., et al., The panel of egg allergens, Gal d 1-Gal d 5: Their improved purification
and characterization. Mol Nutr Food Res, 2008. 52 Suppl 2: p. S176-85.
27. Mine, Y. and M. Yang, Recent advances in the understanding of egg allergens: basic, industrial,
and clinical perspectives. J Agric Food Chem, 2008. 56(13): p. 4874-900.
28. Kleber-Janke, T., et al., Selective cloning of peanut allergens, including profilin and 2S
albumins, by phage display technology. Int Arch Allergy Immunol, 1999. 119(4): p. 265-74.
29. Mittag, D., et al., Ara h 8, a Bet v 1-homologous allergen from peanut, is a major allergen

46
2
Nutrition and food allergy
in patients with combined birch pollen and peanut allergy. J Allergy Clin Immunol, 2004.
114(6): p. 1410-7.
30. Pons, L., et al., The 18 kDa peanut oleosin is a candidate allergen for IgE-mediated reactions
to peanuts. Allergy, 2002. 57 Suppl 72: p. 88-93.
31. Rabjohn, P., et al., Molecular cloning and epitope analysis of the peanut allergen Ara h 3. J
Clin Invest, 1999. 103(4): p. 535-42.
32. Stanley, J.S., et al., Identification and mutational analysis of the immunodominant IgE binding
epitopes of the major peanut allergen Ara h 2. Arch Biochem Biophys, 1997. 342(2): p. 244-
53.
33. Burks, A.W., et al., Recombinant peanut allergen Ara h I expression and IgE binding in patients
with peanut hypersensitivity. J Clin Invest, 1995. 96(4): p. 1715-21.
34. Ree-Kim, L. and S.B. Lehrer, Seafood allergy. Curr Opin Allergy Clin Immunol, 2004. 4(3): p.
231-4.
35. Cordle, C.T., Soy protein allergy: incidence and relative severity. J Nutr, 2004. 134(5): p.
1213S-1219S.
36. Krishnan, H.B., et al., All three subunits of soybean beta-conglycinin are potential food
allergens. J Agric Food Chem, 2009. 57(3): p. 938-43.
37. Ballmer-Weber, B.K. and S. Vieths, Soy allergy in perspective. Curr Opin Allergy Clin Immunol,
2008. 8(3): p. 270-5.
38. Berkner, H., et al., Cross-reactivity of pollen and food allergens: soybean Gly m 4 is a member
of the Bet v 1 superfamily and closely resembles yellow lupine proteins. Biosci Rep, 2009.
29(3): p. 183-92.
39. Constantin, C., et al., Micro-arrayed wheat seed and grass pollen allergens for component-
resolved diagnosis. Allergy, 2009.
40. Akagawa, M., et al., Proteomic analysis of wheat flour allergens. J Agric Food Chem, 2007.
55(17): p. 6863-70.
41. Bock, S.A., Diagnostic evaluation. Pediatrics, 2003. 111(6 Pt 3): p. 1638-44.
42. Scibilia, J., et al., Wheat allergy: a double-blind, placebo-controlled study in adults. J Allergy
Clin Immunol, 2006. 117(2): p. 433-9.
43. Hill, D.J., C.S. Hosking, and L.V. Reyes-Benito, Reducing the need for food allergen challenges
in young children: a comparison of in vitro with in vivo tests. Clin Exp Allergy, 2001. 31(7): p.
1031-5.
44. Lied, G.A., Gastrointestinal food hypersensitivity: Symptoms, diagnosis and provocation
tests. Turk J Gastroenterol, 2007. 18(1): p. 5-13.
45. Sampson, H.A., Food allergy. Part 2: diagnosis and management. J Allergy Clin Immunol,
1999. 103(6): p. 981-9.
46. Celik-Bilgili, S., et al., The predictive value of specific immunoglobulin E levels in serum for
the outcome of oral food challenges. Clin Exp Allergy, 2005. 35(3): p. 268-73.
47. Mehl, A., et al., Utility of the ratio of food-specific IgE/total IgE in predicting symptomatic
food allergy in children. Allergy, 2005. 60(8): p. 1034-9.

47
2
Chapter 2
48. Garcia-Ara, C., et al., Specific IgE levels in the diagnosis of immediate hypersensitivity to
cows’ milk protein in the infant. J Allergy Clin Immunol, 2001. 107(1): p. 185-90.
49. Garcia-Ara, M.C., et al., Eliciting doses of positive challenge test in cow’s milk allergy are
related to cow’s milk specific IgE levels. Allergol Immunopathol (Madr), 2008. 36(6): p. 315-
9.
50. Ejrnaes, A.M., et al., Inhibition of rBet v 1-induced basophil histamine release with specific
immunotherapy -induced serum immunoglobulin G: no evidence that FcgammaRIIB
signalling is important. Clin Exp Allergy, 2006. 36(3): p. 273-82.
51. Ruiter, B., et al., Maintenance of tolerance to cow’s milk in atopic individuals is characterized
by high levels of specific immunoglobulin G4. Clin Exp Allergy, 2007. 37(7): p. 1103-10.
52. Groot Kormelink, T., et al., Atopic and non-atopic allergic disorders: current insights into the
possible involvement of free immunoglobulin light chains. Clin Exp Allergy, 2009. 39(1): p.
33-42.
53. Redegeld, F.A. and F.P. Nijkamp, Immunoglobulin free light chains and mast cells: pivotal role
in T-cell-mediated immune reactions? Trends Immunol, 2003. 24(4): p. 181-5.
54. Redegeld, F.A., et al., Immunoglobulin-free light chains elicit immediate hypersensitivity-like
responses. Nat Med, 2002. 8(7): p. 694-701.
55. Kraneveld, A.D., et al., Elicitation of allergic asthma by immunoglobulin free light chains.
Proc Natl Acad Sci U S A, 2005. 102(5): p. 1578-83.
56. Sediva, A., J. Kayserova, and E. Vernerova, Increase of serum immunoglobulin free light
chains in children with atopic dermatitis. Allergy, 2008. 63: p. 157.
57. Thio, M., et al., Free immunoglobulin light chains: a novel target in the therapy of
inflammatory diseases. Trends Pharmacol Sci, 2008. 29(4): p. 170-4.
58. Redegeld, F.A. and C.H. Wortel, IgE and immunoglobulin free light chains in allergic disease:
new therapeutic opportunities. Curr Opin Investig Drugs, 2008. 9(11): p. 1185-91.
59. Field, C.J., The immunological components of human milk and their effect on immune
development in infants. J Nutr, 2005. 135(1): p. 1-4.
60. Hoppu, U., et al., Breast milk--immunomodulatory signals against allergic diseases. Allergy,
2001. 56 Suppl 67: p. 23-6.
61. Lara-Villoslada, F., et al., Beneficial effects of probiotic bacteria isolated from breast milk. Br
J Nutr, 2007. 98 Suppl 1: p. S96-100.
62. Greer, F.R., S.H. Sicherer, and A.W. Burks, Effects of early nutritional interventions on the
development of atopic disease in infants and children: the role of maternal dietary restriction,
breastfeeding, timing of introduction of complementary foods, and hydrolyzed formulas.
Pediatrics, 2008. 121(1): p. 183-91.
63. Host, A., et al., Dietary products used in infants for treatment and prevention of food allergy.
Joint Statement of the European Society for Paediatric Allergology and Clinical Immunology
(ESPACI) Committee on Hypoallergenic Formulas and the European Society for Paediatric
Gastroenterology, Hepatology and Nutrition (ESPGHAN) Committee on Nutrition. Arch Dis
Child, 1999. 81(1): p. 80-4.

48
2
Nutrition and food allergy
64. Hill, D.J., et al., The efficacy of amino acid-based formulas in relieving the symptoms of cow’s
milk allergy: a systematic review. Clin Exp Allergy, 2007. 37(6): p. 808-22.
65. Halken, S., et al., Comparison of a partially hydrolyzed infant formula with two extensively
hydrolyzed formulas for allergy prevention: a prospective, randomized study. Pediatr Allergy
Immunol, 2000. 11(3): p. 149-61.
66. Niggemann, B. and K. Beyer, Diagnosis of food allergy in children: toward a standardization
of food challenge. J Pediatr Gastroenterol Nutr, 2007. 45(4): p. 399-404.
67. Niggemann, B., et al., Sensitizing capacity and residual allergenicity of hydrolyzed cow’s milk
formulae: results from a murine model. Int Arch Allergy Immunol, 2001. 125(4): p. 316-21.
68. von Berg, A., et al., The effect of hydrolyzed cow’s milk formula for allergy prevention in the
first year of life: the German Infant Nutritional Intervention Study, a randomized double-
blind trial. J Allergy Clin Immunol, 2003. 111(3): p. 533-40.
69. Calder, P.C., et al., Early nutrition and immunity - progress and perspectives. Br J Nutr, 2006.
96(4): p. 774-90.
70. Host, A. and S. Halken, Hypoallergenic formulas--when, to whom and how long: after more
than 15 years we know the right indication! Allergy, 2004. 59 Suppl 78: p. 45-52.
71. Agostoni, C., et al., Complementary feeding: a commentary by the ESPGHAN Committee on
Nutrition. J Pediatr Gastroenterol Nutr, 2008. 46(1): p. 99-110.
72. Boehm, G. and B. Stahl, Oligosaccharides from milk. J Nutr, 2007. 137(3 Suppl 2): p. 847S-
9S.
73. Nakhla, T., et al., Neutral oligosaccharide content of preterm human milk. Br J Nutr, 1999.
82(5): p. 361-7.
74. Gibson, G.R. and M.B. Roberfroid, Dietary modulation of the human colonic microbiota:
introducing the concept of prebiotics. J Nutr, 1995. 125(6): p. 1401-12.
75. Boehm, G., et al., Prebiotic concept for infant nutrition. Acta Paediatr Suppl, 2003. 92(441):
p. 64-7.
76. Boehm, G., et al., Prebiotic carbohydrates in human milk and formulas. Acta Paediatr Suppl,
2005. 94(449): p. 18-21.
77. Fanaro, S., et al., Galacto-oligosaccharides and long-chain fructo-oligosaccharides as
prebiotics in infant formulas: a review. Acta Paediatr Suppl, 2005. 94(449): p. 22-6.
78. Greger, J.L., Nondigestible carbohydrates and mineral bioavailability. J Nutr, 1999. 129(7
Suppl): p. 1434S-5S.
79. Schneeman, B.O., Fiber, inulin and oligofructose: similarities and differences. J Nutr, 1999.
129(7 Suppl): p. 1424S-7S.
80. Meyer, P.D., Nondigestible oligosaccharides as dietary fiber. J AOAC Int, 2004. 87(3): p. 718-
26.
81. Gibson, G.R., et al., Dietary modulation of the human colonic microbiota: updating the
concept of prebiotics. Nutr Res Rev, 2004. 17(2): p. 259-75.
82. Buddington, R.K., et al., Non-digestible oligosaccharides and defense functions: lessons
learned from animal models. Br J Nutr, 2002. 87 Suppl 2: p. S231-9.

49
2
Chapter 2
83. Schley, P.D. and C.J. Field, The immune-enhancing effects of dietary fibres and prebiotics. Br
J Nutr, 2002. 87 Suppl 2: p. S221-30.
84. Vos, A.P., et al., Immune-modulatory effects and potential working mechanisms of orally
applied nondigestible carbohydrates. Crit Rev Immunol, 2007. 27(2): p. 97-140.
85. Moro, G., et al., A mixture of prebiotic oligosaccharides reduces the incidence of atopic
dermatitis during the first six months of age. Arch Dis Child, 2006. 91(10): p. 814-9.
86. Arslanoglu, S., et al., Early dietary intervention with a mixture of prebiotic oligosaccharides
reduces the incidence of allergic manifestations and infections during the first two years of
life. J Nutr, 2008. 138(6): p. 1091-5.
87. Arslanoglu, S., G.E. Moro, and G. Boehm, Early supplementation of prebiotic oligosaccharides
protects formula-fed infants against infections during the first 6 months of life. J Nutr, 2007.
137(11): p. 2420-4.
88. van Hoffen, E., et al., A specific mixture of short-chain galacto-oligosaccharides and long-
chain fructo-oligosaccharides induces a beneficial immunoglobulin profile in infants at high
risk for allergy. Allergy, 2009. 64(3): p. 484-7.
89. Vos, A.P., et al., Dietary supplementation of neutral and acidic oligosaccharides enhances
Th1-dependent vaccination responses in mice. Pediatr Allergy Immunol, 2007. 18(4): p. 304-
12.
90. Vos, A.P., et al., A specific prebiotic oligosaccharide mixture stimulates delayed-type
hypersensitivity in a murine influenza vaccination model. Int Immunopharmacol, 2006. 6(8):
p. 1277-86.
91. Vos, A.P., et al., Dietary supplementation with specific oligosaccharide mixtures decreases
parameters of allergic asthma in mice. Int Immunopharmacol, 2007. 7(12): p. 1582-7.
92. Schouten, B., et al., Acute Allergic Skin Reactions and Intestinal Contractility Changes in Mice
Orally Sensitized against Casein or Whey. Int Arch Allergy Immunol, 2008. 147(2): p. 125-
134.
93. Schouten, B., et al., Cow milk allergy symptoms are reduced in mice fed dietary synbiotics
during oral sensitization with whey. J Nutr, 2009. 139(7): p. 1398-403.
94. Fanaro, S., et al., Acidic oligosaccharides from pectin hydrolysate as new component for infant
formulae: effect on intestinal flora, stool characteristics, and pH. J Pediatr Gastroenterol
Nutr, 2005. 41(2): p. 186-90.
95. Shibata, R., et al., Clinical effects of kestose, a prebiotic oligosaccharide, on the treatment of
atopic dermatitis in infants. Clin Exp Allergy, 2009.
96. Scholtens, P.A., et al., Fecal secretory immunoglobulin A is increased in healthy infants
who receive a formula with short-chain galacto-oligosaccharides and long-chain fructo-
oligosaccharides. J Nutr, 2008. 138(6): p. 1141-7.
97. Shanahan, F., Probiotics and inflammatory bowel disease: is there a scientific rationale?
Inflamm Bowel Dis, 2000. 6(2): p. 107-15.
98. Kukkonen, K., et al., Probiotics and prebiotic galacto-oligosaccharides in the prevention
of allergic diseases: A randomized, double-blind, placebo-controlled trial. J Allergy Clin

50
2
Nutrition and food allergy
Immunol, 2007. 119(1): p. 192-8.
99. Moneret-Vautrin, D.A., et al., Probiotics may be unsafe in infants allergic to cow’s milk.
Allergy, 2006. 61(4): p. 507-8.
100. Sistek, D., et al., Is the effect of probiotics on atopic dermatitis confined to food sensitized
children? Clin Exp Allergy, 2006. 36(5): p. 629-33.
101. Kalliomaki, M., et al., Probiotics and prevention of atopic disease: 4-year follow-up of a
randomised placebo-controlled trial. Lancet, 2003. 361(9372): p. 1869-71.
102. Marschan, E., et al., Probiotics in infancy induce protective immune profiles that are
characteristic for chronic low-grade inflammation. Clin Exp Allergy, 2008. 38(4): p. 611-8.
103. Kalliomaki, M., et al., Probiotics in primary prevention of atopic disease: a randomised
placebo-controlled trial. Lancet, 2001. 357(9262): p. 1076-9.
104. Viljanen, M., et al., Probiotics in the treatment of atopic eczema/dermatitis syndrome in
infants: a double-blind placebo-controlled trial. Allergy, 2005. 60(4): p. 494-500.
105. Abrahamsson, T.R., et al., Probiotics in prevention of IgE-associated eczema: a double-blind,
randomized, placebo-controlled trial. J Allergy Clin Immunol, 2007. 119(5): p. 1174-80.
106. Brouwer, M.L., et al., No effects of probiotics on atopic dermatitis in infancy: a randomized
placebo-controlled trial. Clin Exp Allergy, 2006. 36(7): p. 899-906.
107. Kalliomaki, M., et al., Probiotics during the first 7 years of life: a cumulative risk reduction of
eczema in a randomized, placebo-controlled trial. J Allergy Clin Immunol, 2007. 119(4): p.
1019-21.
108. Soh, S.E., et al., Probiotic supplementation in the first 6 months of life in at risk Asian infants-
-effects on eczema and atopic sensitization at the age of 1 year. Clin Exp Allergy, 2009. 39(4):
p. 571-8.
109. del Giudice, M.M. and F.P. Brunese, Probiotics, prebiotics, and allergy in children: what’s
new in the last year? J Clin Gastroenterol, 2008. 42 Suppl 3 Pt 2: p. S205-8.
110. Savilahti, E., M. Kuitunen, and O. Vaarala, Pre and probiotics in the prevention and treatment
of food allergy. Curr Opin Allergy Clin Immunol, 2008. 8(3): p. 243-8.
111. Prescott, S.L. and B. Bjorksten, Probiotics for the prevention or treatment of allergic diseases.
J Allergy Clin Immunol, 2007. 120(2): p. 255-62.
112. Hauk, P.J., The role of food allergy in atopic dermatitis. Curr Allergy Asthma Rep, 2008. 8(3):
p. 188-94.
113. Werfel, T. and K. Breuer, Role of food allergy in atopic dermatitis. Curr Opin Allergy Clin
Immunol, 2004. 4(5): p. 379-85.
114. Contractor, N.V., et al., Lymphoid hyperplasia, autoimmunity, and compromised intestinal
intraepithelial lymphocyte development in colitis-free gnotobiotic IL-2-deficient mice. J
Immunol, 1998. 160(1): p. 385-94.
115. Cebra, J.J., Influences of microbiota on intestinal immune system development. Am J Clin
Nutr, 1999. 69(5): p. 1046S-1051S.
116. Penders, J., et al., The role of the intestinal microbiota in the development of atopic disorders.
Allergy, 2007. 62(11): p. 1223-36.

51
2
Chapter 2
117. Adel-Patient, K., et al., Oral administration of recombinant Lactococcus lactis expressing
bovine beta-lactoglobulin partially prevents mice from sensitization. Clin Exp Allergy, 2005.
35(4): p. 539-46.
118. Kim, H., et al., Oral probiotic bacterial administration suppressed allergic responses in an
ovalbumin-induced allergy mouse model. FEMS Immunol Med Microbiol, 2005. 45(2): p.
259-67.
119. Kim, J.Y., Y.O. Choi, and G.E. Ji, Effect of Oral Probiotics (Bifidobacterium lactis AD011 and
Lactobacillus acidophilus AD031) Administration on Ovalbumin-Induced Food Allergy Mouse
Model. J Microbiol Biotechnol, 2008. 18(8): p. 1393-400.
120. Pessi, T., et al., Probiotics reinforce mucosal degradation of antigens in rats: implications for
therapeutic use of probiotics. J Nutr, 1998. 128(12): p. 2313-8.
121. Feleszko, W., et al., Probiotic-induced suppression of allergic sensitization and airway
inflammation is associated with an increase of T regulatory-dependent mechanisms in a
murine model of asthma. Clin Exp Allergy, 2007. 37(4): p. 498-505.
122. Frece, J., et al., Synbiotic effect of Lactobacillus helveticus M92 and prebiotics on the
intestinal microflora and immune system of mice. J Dairy Res, 2009. 76(1): p. 98-104.
123. Sato, T., et al., Isolation of lactate-utilizing butyrate-producing bacteria from human feces
and in vivo administration of Anaerostipes caccae strain L2 and galacto-oligosaccharides in a
rat model. FEMS Microbiol Ecol, 2008. 66(3): p. 528-36.
124. Ogawa, T., et al., A new synbiotic, Lactobacillus casei subsp. casei together with dextran,
reduces murine and human allergic reaction. FEMS Immunol Med Microbiol, 2006. 46(3): p.
400-9.
125. Neish, A.S., Microbes in gastrointestinal health and disease. Gastroenterology, 2009. 136(1):
p. 65-80.
126. Eutamene, H., et al., Synergy between Lactobacillus paracasei and its bacterial products
to counteract stress-induced gut permeability and sensitivity increase in rats. J Nutr, 2007.
137(8): p. 1901-7.
127. Simopoulos, A.P., The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular
disease and other chronic diseases. Exp Biol Med (Maywood), 2008. 233(6): p. 674-88.
128. Blumer, N. and H. Renz, Consumption of omega3-fatty acids during perinatal life: role in
immuno-modulation and allergy prevention. J Perinat Med, 2007. 35 Suppl 1: p. S12-8.
129. Dunstan, J.A., et al., The effect of supplementation with fish oil during pregnancy on breast
milk immunoglobulin A, soluble CD14, cytokine levels and fatty acid composition. Clin Exp
Allergy, 2004. 34(8): p. 1237-42.
130. Dunstan, J.A., et al., Fish oil supplementation in pregnancy modifies neonatal allergen-specific
immune responses and clinical outcomes in infants at high risk of atopy: a randomized,
controlled trial. J Allergy Clin Immunol, 2003. 112(6): p. 1178-84.
131. Furuhjelm, C., et al., Fish oil supplementation in pregnancy and lactation may decrease the
risk of infant allergy. Acta Paediatr, 2009.
132. Almqvist, C., et al., Omega-3 and omega-6 fatty acid exposure from early life does not affect

52
2
Nutrition and food allergy
atopy and asthma at age 5 years. J Allergy Clin Immunol, 2007. 119(6): p. 1438-44.
133. Prescott, S.L. and P.C. Calder, N-3 polyunsaturated fatty acids and allergic disease. Curr Opin
Clin Nutr Metab Care, 2004. 7(2): p. 123-9.
134. Vanderhoof, J.A., Hypoallergenicity and effects on growth and tolerance of a new amino
acid-based formula with DHA and ARA. J Pediatr Gastroenterol Nutr, 2008. 47 Suppl 2: p.
S60-1.
135. Calder, P.C., et al., Fatty acids and lymphocyte functions. Br J Nutr, 2002. 87 Suppl 1: p. S31-
48.
136. Sawada, J., et al., Ingestion of heat-treated Lactobacillus rhamnosus GG prevents
development of atopic dermatitis in NC/Nga mice. Clin Exp Allergy, 2007. 37(2): p. 296-303.
137. Vanderhoof, J.A., Probiotics in allergy management. J Pediatr Gastroenterol Nutr, 2008. 47
Suppl 2: p. S38-40.
138. Mihrshahi, S., et al., Effect of omega-3 fatty acid concentrations in plasma on symptoms of
asthma at 18 months of age. Pediatr Allergy Immunol, 2004. 15(6): p. 517-22.
139. Boyle, R.J., et al., Effects of Lactobacillus GG treatment during pregnancy on the development
of fetal antigen-specific immune responses. Clin Exp Allergy, 2008. 38(12): p. 1882-90.
140. Dutta-Roy, A.K., Cellular uptake of long-chain fatty acids: role of membrane-associated fatty-
acid-binding/transport proteins. Cell Mol Life Sci, 2000. 57(10): p. 1360-72.
141. Chehade, M., IgE and non-IgE-mediated food allergy: treatment in 2007. Curr Opin Allergy
Clin Immunol, 2007. 7(3): p. 264-8.
142. Skripak, J.M. and H.A. Sampson, Towards a cure for food allergy. Curr Opin Immunol, 2008.
20(6): p. 690-6.
143. Grimshaw, K.E., Dietary management of food allergy in children. Proc Nutr Soc, 2006. 65(4):
p. 412-7.
144. Hays, T. and R.A. Wood, A systematic review of the role of hydrolyzed infant formulas in
allergy prevention. Arch Pediatr Adolesc Med, 2005. 159(9): p. 810-6.
145. Isolauri, E., et al., Probiotics in the management of atopic eczema. Clin Exp Allergy, 2000.
30(11): p. 1604-10.
146. Rosenfeldt, V., et al., Effect of probiotic Lactobacillus strains in children with atopic dermatitis.
J Allergy Clin Immunol, 2003. 111(2): p. 389-95.
147. Passeron, T., et al., Prebiotics and synbiotics: two promising approaches for the treatment of
atopic dermatitis in children above 2 years. Allergy, 2006. 61(4): p. 431-7.
148. Betsi, G.I., E. Papadavid, and M.E. Falagas, Probiotics for the treatment or prevention of
atopic dermatitis: a review of the evidence from randomized controlled trials. Am J Clin
Dermatol, 2008. 9(2): p. 93-103.
149. Schnappinger, M., et al., Fish consumption, allergic sensitisation and allergic diseases in
adults. Ann Nutr Metab, 2009. 54(1): p. 67-74.
150. Li, X.M. and L. Brown, Efficacy and mechanisms of action of traditional Chinese medicines for
treating asthma and allergy. J Allergy Clin Immunol, 2009. 123(2): p. 297-306; quiz 307-8.
151. Li, X.M., Traditional Chinese herbal remedies for asthma and food allergy. J Allergy Clin

53
2
Chapter 2
Immunol, 2007. 120(1): p. 25-31.
152. Li, X.M., et al., Food Allergy Herbal Formula-1 (FAHF-1) blocks peanut-induced anaphylaxis in
a murine model. J Allergy Clin Immunol, 2001. 108(4): p. 639-46.
153. Srivastava, K.D., et al., The Chinese herbal medicine formula FAHF-2 completely blocks
anaphylactic reactions in a murine model of peanut allergy. J Allergy Clin Immunol, 2005.
115(1): p. 171-8.
154. Qu, C., et al., Induction of tolerance after establishment of peanut allergy by the food allergy
herbal formula-2 is associated with up-regulation of interferon-gamma. Clin Exp Allergy,
2007. 37(6): p. 846-55.
155. Staden, U., et al., Specific oral tolerance induction in food allergy in children: efficacy and
clinical patterns of reaction. Allergy, 2007. 62(11): p. 1261-9.
156. Niederberger, V., Allergen-specific immunotherapy. Immunol Lett, 2009. 122(2): p. 131-3.
157. de Boissieu, D. and C. Dupont, Sublingual immunotherapy for cow’s milk protein allergy: a
preliminary report. Allergy, 2006. 61(10): p. 1238-9.
158. Beyer, K. and U. Wahn, Oral immunotherapy for food allergy in children. Curr Opin Allergy
Clin Immunol, 2008. 8(6): p. 553-6.
159. Patriarca, G., et al., Oral specific desensitization in food-allergic children. Dig Dis Sci, 2007.
52(7): p. 1662-72.
160. Friedman, A. and H.L. Weiner, Induction of anergy or active suppression following oral
tolerance is determined by antigen dosage. Proc Natl Acad Sci U S A, 1994. 91(14): p. 6688-
92.
161. Burks, A.W., S. Laubach, and S.M. Jones, Oral tolerance, food allergy, and immunotherapy:
implications for future treatment. J Allergy Clin Immunol, 2008. 121(6): p. 1344-50.
162. van Wijk, F. and L. Knippels, Initiating mechanisms of food allergy: Oral tolerance versus
allergic sensitization. Biomed Pharmacother, 2007. 61(1): p. 8-20.
163. Romagnani, S., Immunologic influences on allergy and the TH1/TH2 balance. J Allergy Clin
Immunol, 2004. 113(3): p. 395-400.
164. Mayer, L. and L. Shao, Therapeutic potential of oral tolerance. Nat Rev Immunol, 2004. 4(6):
p. 407-19.
165. Mowat, A.M., Anatomical basis of tolerance and immunity to intestinal antigens. Nat Rev
Immunol, 2003. 3(4): p. 331-41.
166. Tsuji, N.M. and A. Kosaka, Oral tolerance: intestinal homeostasis and antigen-specific
regulatory T cells. Trends Immunol, 2008. 29(11): p. 532-40.
167. Ko, J. and L. Mayer, Oral tolerance: lessons on treatment of food allergy. Eur J Gastroenterol
Hepatol, 2005. 17(12): p. 1299-303.
168. Niggemann, B., et al., Specific oral tolerance induction in food allergy. Allergy, 2006. 61(7): p.
808-11.


3
Acute allergic skin reactions and intestinal contractility changes in mice orally sensitized
against casein or whey
Bastiaan Schouten1
Betty C.A.M. van Esch1,2
Gerard A. Hofman1
Lieke W.J. van den Elsen1 Linette E.M.Willemsen1
Johan Garssen1,2
1 Department of Pharmacology and Pathophysiology, Utrecht Institute for Pharmaceutical Sciences, Faculty of Science, Utrecht University, Utrecht, The Netherlands
2 Danone Research – Centre for Specialised Nutrition, Wageningen, The Netherlands
International Archives of Allergy and Immunology 2008;147:125-134S. Karger AG, Basel

56
3
Acute allergic skin reactions and intestinal contractility changes in mice orally sensitized against casein or whey
Abstract
Background: Cow’s milk allergy (CMA) is characterized by hypersensitivity against casein or whey, affecting 2.5% of young infants. The pathogenesis of CMA involves IgE as well as non-IgE-mediated reactions and clinical symptoms are found in the skin, lungs and gastrointestinal tract. In this study, local and systemic immunopathology was determined in whey- or casein-allergic mice.Methods: Mice were orally sensitized with casein or whey using cholera toxin as an adjuvant. Serum immunoglobulins and the acute allergic skin reaction (ear swelling 1 h after intradermal allergen challenge) were determined to reveal systemic hypersensitivity. Furthermore, pathophysiological changes were assessed within the intestine.Results: An acute allergic skin reaction was induced in both whey- and casein-sensitized mice. In these mice, whey-specific IgE, IgG
1, IgG2a and casein-specific IgG1 levels were found to be increased. In addition, the serum mouse mast cell protease-1 (mMCP-1) concentration was enhanced, reflecting mast cell degranulation. Indeed, the number of mMCP-1-positive mast cells within the colon was diminished in both whey- and casein-sensitized mice. Only in casein-sensitized mice isometric contraction of the colon was reduced, reflecting motility alterations.Conclusion: Mice, orally sensitized against casein or whey, revealed an allergen-specific acute allergic skin reaction. In casein-sensitized mice, hypocontractility of the colon reflected pathophysiological changes within the intestine. Allergen induced ear swelling and intestinal contractility changes are novel parameters in animal models of CMA which may add to the search for new therapeutic strategies to relieve symptoms of CMA.

57
3
Chapter 3
Introduction
Cow’s milk allergy (CMA) is one of the leading causes of food allergy in adults [1]. In developed countries, approximately 2–3% of infants exhibit CMA. Although most infants outgrow CMA before their fifth year, IgE mediated CMA predisposes the development of other (food) allergies and even asthma [2, 3]. Clinical features due to IgE-mediated reactions are expressed as immediate symptoms mostly. Clinical symptoms may involve the skin, respiratory tract and gastrointestinal tract, and can even lead to a systemic anaphylactic reaction [4, 5]. However, it should be realized that a particular group of patients, up to 40%, exhibit clinical features of CMA without detectable cow’s milk-specific serum IgE [6-8]. So far, the only therapeutic approach has been the elimination of cow’s milk proteins from the diet. Cow’s milk contains 2 main protein classes, caseins (30 g/l) and whey proteins (5 g/l). The caseins consist mainly of αS1-, αS2- and β-casein, whereas whey proteins comprise of β-lactoglobulin, α-lactalbumin, bovine serum albumin, serum immunoglobulins and lactoferrin. Large population studies with cow’s milk-allergic infants have shown that the major allergens are β-lactoglobulin, caseins and α-lactalbumin [9, 10]. The pathogenesis and development of CMA probably involves a partial dysfunction in immunological tolerance induction during early life combined with enhanced intestinal permeability [5]. There is growing evidence that intestinal allergic responses can initiate motility changes through attraction and activation of mast cells and production of T
H2 type of cytokines [11]. Motility changes of the intestine are a clinical problem in CMA patients, which manifests as either diarrhea or even constipation [12-14].
Animal models of CMA provide a tool to reveal mechanisms involved in CMA and may explore new therapeutic and preventive approaches in those models. However, in most existing food allergy models, the animals are not sensitized via the oral route (but, for example, intraperitoneally), while in reality humans are sensitized orally. Although numerous animal models for food allergy are available using an intraperitoneal sensitization protocol, only a few models use oral sensitization [15-23]. Li et al. [22] were the first to introduce a model in which mice were sensitized orally against complete cow’s milk, while Frossart et al. [23] showed oral sensitization to β-lactoglobulin. In these models, mice were sensitized by means of intragastric (i.g.) gavage using cholera toxin (CT) as an adjuvant. The primary read out parameters were IgA and IgE, and in particular cow’s milk-specific IgE. Furthermore, Li et al. [22] described systemic anaphylaxis upon i.g. challenge with cow’s milk. They described vascular leakage, mast cell degranulation and enhanced serum histamine levels in CMA animals. The systemic allergic responses to food allergens that are described in these models resemble features found in the clinic; however, the mechanisms behind the development of CMA are still only partially understood.

58
3
Acute allergic skin reactions and intestinal contractility changes in mice orally sensitized against casein or whey
To further assess mechanisms underlying CMA and/or test new concepts for prevention and/or treatment for CMA, in the present study new tools to address the pathological changes in casein and whey allergy in mice have been developed. Measurement of the acute allergic skin response, as an equivalent of the human skin prick test, was introduced to reflect a systemic sensitization. Intestinal contractility changes were measured to address local pathophysiological changes after oral challenge with the specific allergens.
Methods
Chemicals
Casein and whey were obtained from DMV International (Veghel, The Netherlands). Cholera toxin was purchased from Quadratech Diagnostics (Epsom, UK) and PBS from Cambrex Bio Science (Verviers, Belgium). Biotin-labeled rat anti-mouse IgE, IgG1 and IgG2a, and unlabeled rat anti mouse IgE were obtained from BD Biosciences (Alphen aan den Rijn, The Netherlands). All other chemicals were obtained from Sigma-Aldrich-Chemie (Zwijndrecht, The Netherlands).
Oral Sensitization and Challenge of Mice
Three- to five-week-old specific pathogen-free female C3H/HeOuJ mice (4–6 mice per group)
14 21 424035280 7
Sensitization by gavage with casein or whey + CT or CT alone (control)
Challenge i.d. 10 µg
‐ Serum parameters‐ Functional changes intestine‐ Histology
days
C3H/HeOuJFemale 3/5 wks Challenge i.g. 100 mg
Skin response 1, 4 and 24 hours post challenge
Figure 1. A schematic overview of the sensitization and challenge protocol and the parameters that are analyzed.

59
3
Chapter 3
were purchased from Charles River Laboratories (Maastricht, The Netherlands), maintained on cow’s milk protein-free mouse chow (Special Diets Services, Witham, UK) and housed in the animal facility at the Utrecht University. Animal care and use were performed in accordance with the guidelines of the Dutch Committee of Animal Experiments. Mice were sensitized i.g. with 0.5 ml homogenized casein or whey (40 mg/ml PBS) with CT (20 μg/ml PBS) as an adjuvant, using a blunt needle. Control mice received CT alone or PBS. Mice were boosted weekly for a period of 4–6 weeks, 1 week after the last sensitization mice were challenged i.g. with 100 mg casein or whey in 1 ml PBS (Figure 1). Blood samples were collected and centrifuged (15 min at 13,500 rpm). Sera were stored at –70° C. Mice were sacrificed by cervical dislocation 30 min after i.g. challenge.
Allergen-Specific Skin Response
The acute allergen-specific skin response was measured after injection of the specific protein in the ear pinnae intradermally (i.d.). Before i.g. challenge (t = 0), the control, casein- and whey-sensitized mice were injected i.d. in the left ear with 20 μl homogenized casein or whey (0.5 mg/ml in PBS), respectively. In the right ear, 20 μl PBS was injected as a vehicle control. Also, the CT and PBS sham-sensitized mice received a casein or whey ear challenge using PBS injections as control. Ear thickness was measured in duplicate using a digital micrometer (Mitutoyo, Veenendaal, The Netherlands), at t = 0 as well as 1, 4 and 24 h after challenge. The allergen-specific net ear swelling was calculated by subtracting the basal thickness (0 h) from the thickness measured at 1, 4 and 24 h after injection in the left ear. In addition, the control (right ear) swelling measured at the same time points was subtracted. The ear swelling is expressed as delta μm.
Measurement of Serum Immunoglobulin and Mouse Mast Cell Protease-1
Concentrations of total IgE and levels of casein- or whey-specific IgE, IgG1 and IgG2a were determined in serum of sacrificed mice by means of ELISA. Microlon plates (Greiner, Alphen aan den Rijn, The Netherlands) were coated with 100 μl whey or casein (20 μg/ml) in coating buffer or rat anti-mouse IgE (1 μg/ml; carbonate-bicarbonate buffer, 0.05 M , pH = 9.6) for 18 h at 4°C. Plates were washed and blocked for 1 h with 5% BSA. Serum samples were applied in several dilutions (4–100 times) and incubated for 2 h at room temperature. Plates were washed and incubated with biotin-labeled rat anti-mouse IgE, IgG
1 or IgG2a (1 μg/ml) for 90 min at room temperature and washed. The plates were incubated with streptavidin-horseradish peroxidase for 1 h, washed and developed with o-phenylendiamine. After 5

60
3
Acute allergic skin reactions and intestinal contractility changes in mice orally sensitized against casein or whey
min, the reaction was stopped with 4 M H2SO4 and absorbance was measured at 490 nm on a Benchmark microplate reader (Bio-Rad, Hercules, Calif., USA). Serum concentrations of mouse mast cell protease-1 (mMCP-1) were determined as described previously using a commercially available ELISA kit (Moredun Scientific Ltd., Penicuik, UK) [24].
Immunohistology of the Colon
For determination of histopathological alterations, Swiss rolls of 4 cm colon were prepared [25]. The colon was carefully dissected, opened longitudinally over the mesenteric line and luminal contents were removed by gently washing in saline. The colon was placed with the mucosal side down. Colons were rolled from the distal to the proximal end, fixed in ice-cold 10% formaldehyde in PBS (for 24–48 h) and embedded in paraffin (Leica EG1150c; Leica Microsystems, Rijswijk, The Netherlands). Sections of 5 μm were cut using a microtome (Leica RM2165; Leica Microsystems), stretched on water (Tissue Flotation Bath TFB45; Medite, Nunningen, Switzerland) and placed on poly-L-lysine-coated slides. Deparaffinized Swiss roll sections were stained with mMCP-1 to detect mucosal mast cells [26]. In short, after fixation in acetone, the sections were blocked with 10% normal goat serum, followed by incubation with rat anti-mouse mMCP-1 (kindly donated by Dr. H.R.P. Miller, Edinburgh, UK). The primary antibody was detected with a biotinylated polyclonal goat anti-rat antibody (Pharmingen, Aalst, Belgium), followed by incubation with streptavidin-horseradish peroxidase (Sanquin, Utrecht, The Netherlands). Color was developed with AEC chromogen staining kit (Sigma Chemical Co., St. Louis, Mo., USA) and sections were counterstained with hematoxylin. Mast cell counts were performed by counting 1 complete section of every animal.
Isometric Contraction of the Colon
The colon, caudal from the cecum, was dissected free of connective tissue and mesenterium. Colon parts of 1 cm length were mounted in an organ bath, using 2 small clamps, containing 10 ml tyrode buffer (in mM: NaCl 136.89, KCl 2.68, MgCl 2 1.05, CaCl 2 1.77, NaH2 PO4 0.42, NaHCO3 11.9 and glucose 5.55) [27-29]. The tissue was kept at 32°C, to prevent spontaneous contraction, and continuously gassed with a 5% CO2 and 95% O2 gas mixture. One clamp was attached to a fixed point in the organ bath and the other clamp was connected to an isometric transducer (Harvard Apparatus Ltd., Edenbridge, UK) with an analog recorder (BD40; Kipp & Zn., Delft, The Netherlands). Contractions were measured under a constant preload of 1.00 g. The preparations were equilibrated for 1 h in the organ bath before starting a log dose-response curve for carbachol (10–8 until 10–3 M). After every dosage, the organ

61
3
Chapter 3
bath was flushed twice and the tissue was allowed to recover for 10 min before addition of the next concentration. Also, basal activity and spontaneous contraction frequencies of the tissue were recorded.
Water Percentage in the Stool
Twenty-four hours after oral challenge, feces were collected from every animal. These samples were weighted, dried for several days and weighted again. The difference in weight is the evaporated water. The relative amount of water in the feces (relative to the total weight of the feces) was calculated and statistically evaluated.
Statistics
All data except for the isometric contractions were analyzed using one-way ANOVA and post hoc Dunnett’s test. Isometric contraction data were analyzed using repeated measures ANOVA and post hoc Dunnett’s test. Statistical analyses were conducted using GraphPad Prism software (version 4.0). For correlations, the Spearman rank order correlation coefficient test was used in SPSS 13.0 for Windows. Data are represented as means ± SEM.
Results
Acute Allergen-Specific Skin Reaction
In order to study allergic skin responses, mice were challenged i.d. in the ear pinnae with casein or whey 1 week after the last oral sensitization. One hour after dermal challenge, the allergen-specific acute ear swelling response was maximal when compared to PBS and CT sham-sensitized control mice. The delta ear swelling in the casein- and whey-sensitized animals was 71.2 ± 8.4 and 137.9 ± 21.7 μm, respectively, while this was neglectable in PBS and CT sham-treated mice (8.1 ± 9.4 and –4.6 ± 4.7 μm, respectively; Figure 2, p<0.01, n=4 in PBS group, n=6 in all other groups). All sensitized mice reacted positively to the allergen, indicating that there are no non-responders. Four hours after challenge, the ear swelling turned to basal levels in the casein group, while in the whey-sensitized group the swelling remained slightly enhanced up to 24 h after challenge (37.5 ± 12.6 μm in whey-sensitized vs. –3.9 ± 11.4 μm in CT-sensitized animals, p<0.05). In additional experiments (data not shown)

62
3
Acute allergic skin reactions and intestinal contractility changes in mice orally sensitized against casein or whey
it was found that the marginal 24 h response was not followed by a response at later time points. In sham-sensitized animals, no differences were found between casein and whey challenge at any time points. Furthermore, whey challenge in casein-sensitized mice or vice versa did not result in any significant swelling response, indicating the antigen specificity of the skin reaction (data not shown).
Increased Total IgE and Antigen-Specific Serum IgE, IgG1 and IgG2a Levels
Half an hour after oral challenge, serum was obtained from both sensitized and non-sensitized animals. Total IgE concentrations were increased in casein- and whey-sensitized mice (400 ± 32 and 364 ± 19 ng/ml, respectively) in comparison with CT and PBS sham-sensitized mice (243 ± 37 and 218 ± 31 ng/ml, respectively; data not shown, p<0.01 or p<0.05, n=4 in PBS group and n=6 in all other groups). Casein-specific IgE as well as casein-specific IgG
2a levels were not increased (Figure 3a and c). In contrast, casein-specific IgG1 levels were enhanced in the casein-sensitized mice (0.537 ± 0.135 OD A490) when compared to CT and PBS sham-sensitized animals (0.087 ± 0.017 and 0.062 ± 0.013, respectively; Figure 3b, p < 0.01). In
Figure 2. Induction of an acute ear swelling in casein- and whey-sensitized mice in comparison with sham-sensitized (CT and PBS) mice. Delta ear swelling (µm) is calculated by subtracting the specific ear swelling induced by the corresponding antigen with vehicle (PBS) induced swelling at 1, 4 and 24 hours. * p<0.05, ** p<0.01, n=4 in PBS group, n=6 in all other groups.
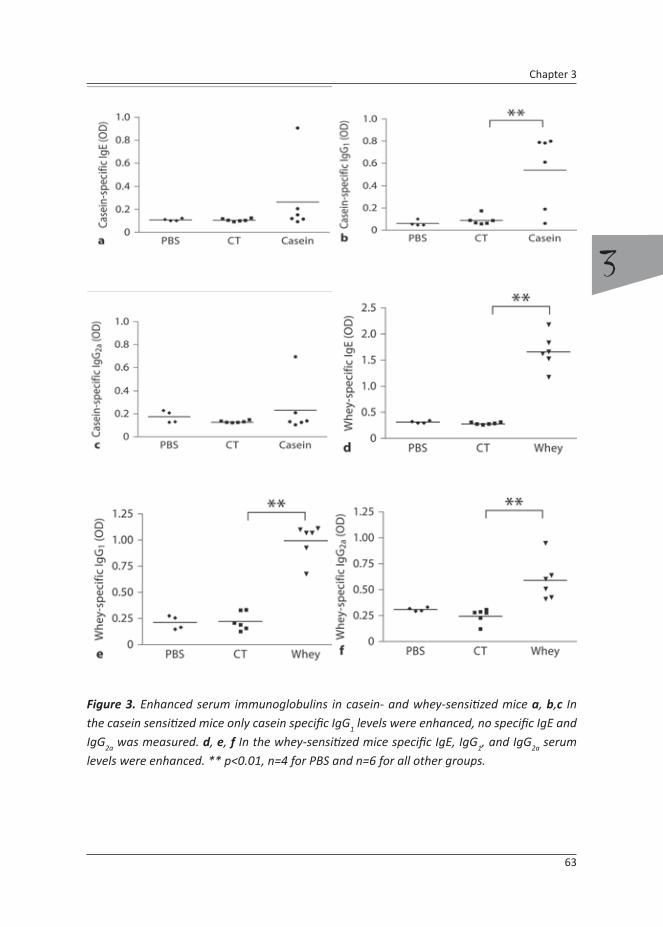
63
3
Chapter 3
Figure 3. Enhanced serum immunoglobulins in casein- and whey-sensitized mice a, b,c In the casein sensitized mice only casein specific IgG1 levels were enhanced, no specific IgE and IgG2a was measured. d, e, f In the whey-sensitized mice specific IgE, IgG1, and IgG2a serum levels were enhanced. ** p<0.01, n=4 for PBS and n=6 for all other groups.
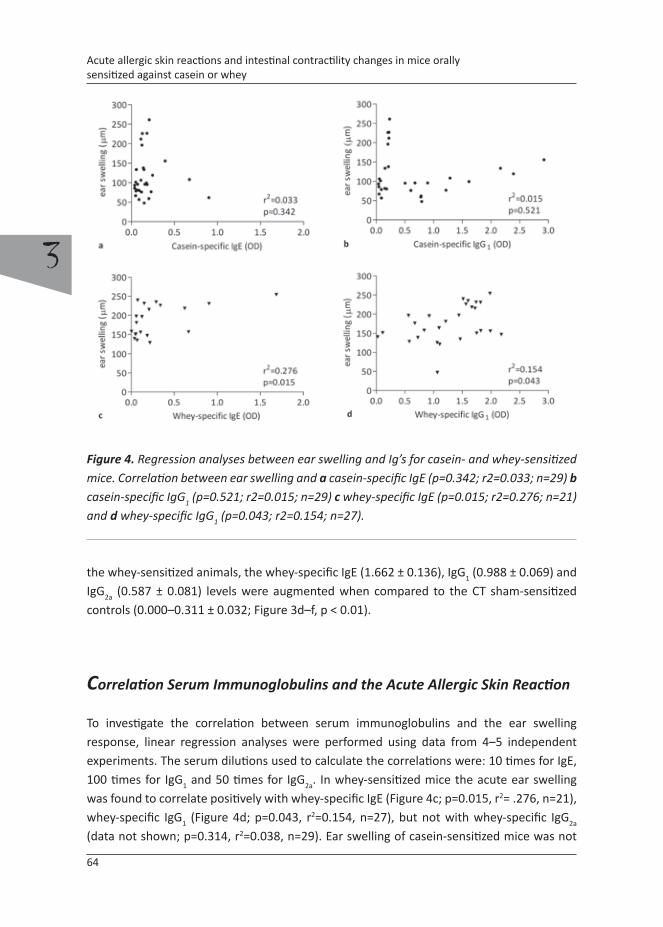
64
3
Acute allergic skin reactions and intestinal contractility changes in mice orally sensitized against casein or whey
Figure 4. Regression analyses between ear swelling and Ig’s for casein- and whey-sensitized mice. Correlation between ear swelling and a casein-specific IgE (p=0.342; r2=0.033; n=29) b casein-specific IgG1 (p=0.521; r2=0.015; n=29) c whey-specific IgE (p=0.015; r2=0.276; n=21) and d whey-specific IgG1 (p=0.043; r2=0.154; n=27).
the whey-sensitized animals, the whey-specific IgE (1.662 ± 0.136), IgG1 (0.988 ± 0.069) and IgG2a (0.587 ± 0.081) levels were augmented when compared to the CT sham-sensitized controls (0.000–0.311 ± 0.032; Figure 3d–f, p < 0.01).
Correlation Serum Immunoglobulins and the Acute Allergic Skin Reaction
To investigate the correlation between serum immunoglobulins and the ear swelling response, linear regression analyses were performed using data from 4–5 independent experiments. The serum dilutions used to calculate the correlations were: 10 times for IgE, 100 times for IgG1 and 50 times for IgG2a. In whey-sensitized mice the acute ear swelling was found to correlate positively with whey-specific IgE (Figure 4c; p=0.015, r2= .276, n=21), whey-specific IgG1 (Figure 4d; p=0.043, r2=0.154, n=27), but not with whey-specific IgG2a (data not shown; p=0.314, r2=0.038, n=29). Ear swelling of casein-sensitized mice was not
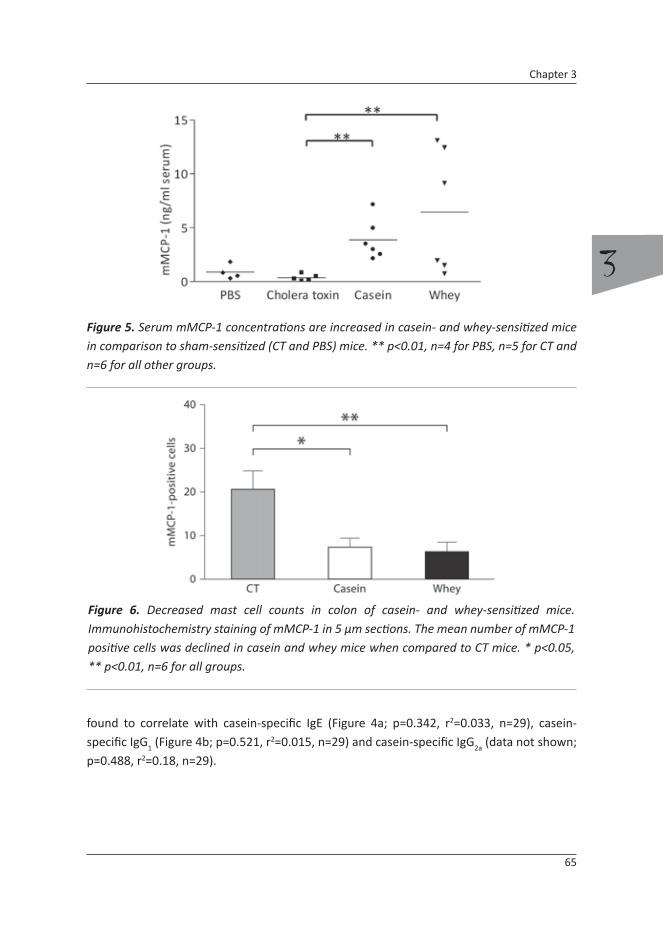
65
3
Chapter 3
found to correlate with casein-specific IgE (Figure 4a; p=0.342, r2=0.033, n=29), casein-specific IgG1 (Figure 4b; p=0.521, r2=0.015, n=29) and casein-specific IgG2a (data not shown; p=0.488, r2=0.18, n=29).
Figure 5. Serum mMCP-1 concentrations are increased in casein- and whey-sensitized mice in comparison to sham-sensitized (CT and PBS) mice. ** p<0.01, n=4 for PBS, n=5 for CT and n=6 for all other groups.
Figure 6. Decreased mast cell counts in colon of casein- and whey-sensitized mice. Immunohistochemistry staining of mMCP-1 in 5 µm sections. The mean number of mMCP-1 positive cells was declined in casein and whey mice when compared to CT mice. * p<0.05, ** p<0.01, n=6 for all groups.

66
3
Acute allergic skin reactions and intestinal contractility changes in mice orally sensitized against casein or whey
Increased mMCP-1 Levels in Serum
mMCP-1 is a protease specific for mouse mucosal mast cells and will appear in the bloodstream after mast cell degranulation. To assess mast cell degranulation, mMCP-1 levels were determined in the serum 30 min after oral challenge. mMCP-1 serum concentrations of casein-and whey-sensitized mice were enhanced when compared to CT and PBS controls (3.890 ± 0.769 and 6.472 ± 2.341 vs. 0.357 ± 0.139 and 0.878 ± 0.338 ng/ml, respectively; Figure 5; n=4–6, p<0.01).
Figure 7. Contractility changes in the colon of casein-sensitized mice after carbachol stimulation. Contractility of the colon of casein mice was decreased when compared to CT and whey treated mice. ** p<0.01, n=6 in all groups.
Figure 8. Water percentage water in the stool of casein- and whey-sensitized mice compared to CT controls. The stool water content was found to be increased in both casein and whey mice, 24 hours after oral challenge. * p<0.05, ** p<0.01, n=6 in all groups.

67
3
Chapter 3
Decreased Number of mMCP-1-Positive Mast Cells in the Colon
After oral challenge, mice were sacrificed and the colon was obtained for histological examination. mMCP-1-positive cells were counted after immunohistochemical staining. A decreased number of mucosal mast cells was counted in the colon of casein- (7.33 ± 5.13; p<0.05) and whey-sensitized animals (6.33 ± 5.32; p<0.01) when compared to non-sensitized control mice (20.67 ± 10.27; Figure 6; n=6).
Altered Isometric Contraction of the Colon
Isometric contraction of the colon was determined by assessing contractility upon exposure to muscarinic receptor agonist carbachol (noncumulative dose response, 10–8 – 10–3 M). Reduced contractility for all carbachol concentrations was found for the colon of casein-sensitized mice when compared to sham-treated mice (Figure 7; p<0.01, basal contractility did not differ between groups). This hyporesponsiveness of the colon cannot be characterized by a right movement of the EC50 concentration, but only by a reduction in maximal contraction force (Emax ) (4,975 ± 1,046, 2,242 ± 296 and 5,933 ± 1,071 mg in control, casein and whey groups). The hyporesponsiveness of the colon in the casein-sensitized mice was found to be consistently present in all experiments, while none of these effects were seen in the whey-sensitized animals.
Table 1. Overview of major parameters tested in both models. + : increased compared to control (CT) mice; - : decreased compared to control (CT) mice; ND : no difference observed regarding to control (CT) mice.
Parameter Casein Whey
Sensitization IgE ND +
IgG1 + +
ear swelling + +
Mast cell degranulation mMCP-1 + +
Physiology water percentage + +
motility - ND

68
3
Acute allergic skin reactions and intestinal contractility changes in mice orally sensitized against casein or whey
Water Percentage in the Stool
The relative amount of water in the feces was increased in the casein- (Figure 8; 61.97 ± 0.97%, p<0.05, n=6) and whey-sensitized animals (63.22 ± 1.78%, p<0.01, n=6) when compared to the control mice (56.68 ± 0.62%, n=6).
Discussion
One of the most common food allergies in childhood is CMA. CMA is diagnosed when symptomatic patients have enhanced serum levels of cow’s milk protein-specific IgE (radioallergosorbent test) and/or a positive skin prick test (SPT). In addition, a double-blind placebo-controlled oral challenge can be performed, which is the most reliable test for food allergy [9, 30, 31]. Currently, no immunotherapy is available for CMA patients, hence patients need to avoid cow’s milk allergens in the diet and use hydrolyzed milk formulas [9]. In the present study, oral sensitization against the cow’s milk proteins whey and casein is investigated in mice that were sensitized orally. Specific immunoglobulin levels were induced and local and systemic symptoms were evaluated by studying effects in the gastrointestinal tract and skin. The investigated parameters closely resemble diagnostic tools that are used in the clinic; they are summarized in table 1.
Besides screening for cow’s milk-specific serum IgE, the SPT is used for the diagnoses of CMA in humans [30]. In the current study, the allergen-induced ear swelling is introduced as a possible equivalent for the SPT. It is a new tool to determine systemic sensitization for casein and whey in mice. Oral sensitization with casein or whey consistently resulted in a positive acute allergic ear swelling response upon local allergen challenge. Furthermore, it can be concluded that sensitization via the oral route was able to induce systemic sensitization in mice treated either with casein or whey. In other models for skin hypersensitivity, comparative values for ear swelling responses have been found [32].
Although both whey- and casein-sensitized animals showed evidence of systemic allergy according to the ear swelling response, differences were found in serum immunoglobulin responses between the groups. Although total IgE concentrations and specific IgG
1 levels were enhanced in both the casein- and whey-sensitized mice, specific IgE and IgG2a were found to be increased only in the whey-sensitized mice. In the clinic, 40% of CMA patients have a negative radioallergosorbent test for cow’s milk proteins; however, they reveal to be positive in the SPT [6-8]. In a recent study, the SPT was found to correlate better with the double-blind food provocation test than serum immunoglobulins, although larger studies are necessary to confirm these findings [33]. Typically, in the casein-sensitized mice, no

69
3
Chapter 3
specific IgE was detected, while the mice indeed showed systemic allergy as seen in the acute allergen-induced ear swelling response. In contrast, the whey-sensitized mice showed the classical characteristics of type I IgE-mediated allergy, scoring positive for allergen-specific IgE and acute ear swelling, underlining the validity of this tool [34-36]. In whey-sensitized mice, the validity of the acute skin reaction test as a tool to determine systemic sensitization was supported by the finding that challenge-induced ear swelling correlated with whey-specific IgE and IgG
1. The acute allergic ear swelling in casein-sensitized mice was not associated with enhanced specific IgE levels, which may imply that there is no role for IgE in this model under the current protocol. In CMA patients, casein-specific IgE can be detected in the serum, but levels vary. It is known that patients with persistent CMA over the age of 9 years had elevated levels of casein-specific IgE compared to younger children with CMA. Therefore, low levels of casein-specific IgE early in life could indicate a non-persistent form of CMA [31]. Instead of IgE, the acute ear swelling in mice might have been triggered by IgG
1, which is generally known to play a crucial role in mast cell activation in rodents. It has been shown that immunoglobulins can significantly interfere, positively and negatively, with mast cell responses (reviewed by Bruhns et al. [37]). In addition, it is known from the literature [38] that even in FcεRI-deficient mice, anaphylactic reactions are possible, indicating that IgE is not always a prerequisite for anaphylaxis. Although, in casein-sensitized mice no significant correlation was found between ear swelling and levels of IgG
1, the scatter plot indeed shows a subgroup of mice in which the ear swelling is positively associated with the level of IgG1. Hence, although casein-allergic mice lack specific IgE, enhanced IgG1 levels in these mice may, at least partly, reflect allergic sensitization. Apart from IgE and IgG1, Redegeld et al. [39] have found that mast cell degranulation can occur with immunoglobulin light chain. Immunoglobulin light chain is produced in excess during the formation of immunoglobulins. Hence, casein-specific immunoglobulin light chain may have caused the ear swelling in casein mice with an acute allergic skin response that could not be explained by the presence of IgE nor IgG
1. However, this remains speculative and further research is necessary.
Whey-sensitized mice show a slight but significant ear swelling at 24 h. Together with the higher allergen-specific IgG2a levels in these animals, this might reflect a late-phase response after whey sensitization. Frossard et al. [40] also found enhanced serum levels of IgE/IgG1 and IgG2a, characteristic for a mouse TH2 and TH1 type of immune response, respectively, against β-lactoglobulin in a C3H/HeOuJ mice model for β-lactoglobulin. These levels of IgG2a are in accordance with enhanced levels of IgG2a that have been found in a similar mouse model for peanut allergy [22, 23, 41].
Clinical features of CMA are generally known to be elicited by mucosal mast cells [4, 22, 42]. Those mast cells are present in the intestinal mucosa and additionally drawn to the site of allergen challenge. In casein- and whey-sensitized mice, mast cell numbers of the

70
3
Acute allergic skin reactions and intestinal contractility changes in mice orally sensitized against casein or whey
colon were found to be declined in comparison to sham-sensitized mice. Mucosal mast cells were stained with mMCP-1, a β-chymase present in the mast cell granules, which end up in the bloodstream after degranulation [43]. In both whey- and casein-sensitized mice, serum mMCP-1 concentrations were increased, 30 min after i.g. challenge, reflecting mast cell degranulation. Upon oral allergen exposure, mucosal mast cells may have degranulated, resulting in enhanced mMCP-1 serum levels. After mast cell degranulation, the mast cell itself is not visible anymore with mMCP-1 staining, which might explain the drop in mast cell numbers in the colon of casein- and whey-sensitized mice [44].
One of the most prominent symptoms of food allergy is discomfort in the gastrointestinal tract which can be abdominal pain, diarrhea or sometimes constipation [4, 5]. The collected stool samples of casein- and whey-sensitized animals contained a higher water percentage than the controls, which is suggestive of diarrhea. Besides diarrhea, dysmotility of the intestine is a problem in CMA patients [12, 13]. The casein-sensitized mice showed contractility changes in organ bath studies, while whey-, CT and PBS-sensitized mice did not. Isometric contractions of colon segments of casein-sensitized animals were hyporeactive in comparison to CT controls. It is known that allergy has adverse effects on the gut health, causing dysmotility [45, 46]. In this study, smooth muscle contractility differences between control and casein-sensitized animals may reflect changes in motility, which is supported by findings of Kobayashi et al. [47] in a model for diarrhea. Motility measurements are often performed when studying inflammatory bowel diseases, in which similar intestinal symptoms can be found. In these studies, local intestinal inflammation is indicated as causative factor for motility changes reflecting alterations in smooth muscle contractions [48]. Hence, local intestinal inflammation may have induced hypomotility occurring in the casein mice. However, histological evaluation did not reveal any obvious signs of inflammatory cell infiltrates after oral challenge. Possibly during sensitization, casein might have caused local inflammation, hereby reducing sensitivity of cholinergic neurons for carbachol stimulations, known to alter colonic motility [49]. In addition, local mast cell degranulation may have induced motility changes [11]. Mule et al. [29, 50] have shown that activation of protease-activated receptors (PAR) can cause colon smooth muscle relaxation as well as contraction. PAR agonists such as mast cell-derived tryptase may have caused PAR activation in both casein- and whey-sensitized mice. It remains speculative why hypomotility of the colon was only observed in casein-sensitized mice; however, local levels of tryptase and PAR expression may differ between casein- and whey-sensitized mice and will be the focus of future investigation.
Additionally, casein and whey proteins differ with regard to physical, physiological and dietary properties and therefore may cause differential effects within the intestine. For example, it is known that there is a difference in digestive speed of casein and whey [51]. Casein protein is a slowly digested dietary protein, while whey protein is a fast one [52].

71
3
Chapter 3
In addition, both proteins have different effects on satiety and gastrointestinal hormone response [53]. Hence, these proteins differentially influence the intestinal physiology. Furthermore, it is known that casein by itself can cause DNA damage in the colon, which is associated with a thinner mucus barrier. In the same study, whey did not cause these effects [54]. Two studies have shown hypomotility of the intestine after ingestion of casein, which might relate to the presence of casomorphins that were found to reduce motility [55, 56]. Hence, casein may also have intrinsic properties that can cause hypomotility. At this stage, the exact mechanism behind the hypocontractility found in the casein-sensitized mice is not known and will be the topic of future studies.
Taken together, altered motility suggests subtle local changes in intestinal discomfort and provides a new tool to measure local intestinal alterations as a consequence of allergic sensitization in mice.
Both casein and whey sensitization consistently resulted in an acute allergic skin reaction after allergen challenge, which was associated with specific IgE and IgG
1 serum levels in whey-sensitized mice and specific IgG1 serum levels in casein-sensitized mice. Decreased numbers of mMCP-1-positive mucosal mast cells within the colon and enhanced mMCP-1 levels in the serum suggest a pathophysiological role for mucosal mast cells in this model. In addition, the stool water content of allergic mice was enhanced, reflecting occurrence of diarrhea 24 h after oral allergen challenge. In the casein-sensitized mice, reduced intestinal smooth muscle contraction was observed. This suggests dysmotility of the colon shortly after oral exposure to the allergen. Indeed, constipation and/or diarrhea are among the symptoms in patients affected with CMA. Overall, the tools described in this study might open new avenues to unravel underlying mechanisms of whey and casein allergy in mice.
Acknowledgement
We would like to thank Dr. Anneke Rijnierse for assistance with mMCP-1 immunohistochemistry and helpfull discussions.
References
1. Schafer, T., et al., Epidemiology of food allergy/food intolerance in adults: associations with
other manifestations of atopy. Allergy, 2001. 56(12): p. 1172-9.
2. Rhodes, H.L., et al., Early life risk factors for adult asthma: a birth cohort study of subjects at

72
3
Acute allergic skin reactions and intestinal contractility changes in mice orally sensitized against casein or whey
risk. J Allergy Clin Immunol, 2001. 108(5): p. 720-5.
3. Host, A., Frequency of cow’s milk allergy in childhood. Ann Allergy Asthma Immunol, 2002.
89(6 Suppl 1): p. 33-7.
4. Furrie, E., Probiotics and allergy. Proc Nutr Soc, 2005. 64(4): p. 465-9.
5. Kalach, N., et al., Intestinal permeability in children: variation with age and reliability in the
diagnosis of cow’s milk allergy. Acta Paediatr, 2001. 90(5): p. 499-504.
6. Vanto, T., et al., Prediction of the development of tolerance to milk in children with cow’s
milk hypersensitivity. J Pediatr, 2004. 144(2): p. 218-22.
7. Host, A. and S. Halken, A prospective study of cow milk allergy in Danish infants during
the first 3 years of life. Clinical course in relation to clinical and immunological type of
hypersensitivity reaction. Allergy, 1990. 45(8): p. 587-96.
8. Shek, L.P., et al., Humoral and cellular responses to cow milk proteins in patients with milk-
induced IgE-mediated and non-IgE-mediated disorders. Allergy, 2005. 60(7): p. 912-9.
9. Monaci, L., et al., Milk allergens, their characteristics and their detection in food: a review.
European Food Research and Technology, 2005(Volume 223, Number 2 / June, 2006): p.
149-179.
10. Bahna, S.L., Cow’s milk allergy versus cow milk intolerance. Ann Allergy Asthma Immunol,
2002. 89(6 Suppl 1): p. 56-60.
11. Saavedra, Y. and P. Vergara, Hypersensitivity to ovalbumin induces chronic intestinal
dysmotility and increases the number of intestinal mast cells. Neurogastroenterol Motil,
2005. 17(1): p. 112-22.
12. Murch, S., Allergy and intestinal dysmotility - evidence of genuine causal linkage? Curr Opin
Gastroenterol, 2006. 22(6): p. 664-8.
13. Magazzu, G. and R. Scoglio, Gastrointestinal manifestations of cow’s milk allergy. Ann Allergy
Asthma Immunol, 2002. 89(6 Suppl 1): p. 65-8.
14. Scaillon, M. and S. Cadranel, Food allergy and constipation in childhood: how functional is it?
Eur J Gastroenterol Hepatol, 2006. 18(2): p. 125-8.
15. Adel-Patient, K., et al., Oral administration of recombinant Lactococcus lactis expressing
bovine beta-lactoglobulin partially prevents mice from sensitization. Clin Exp Allergy, 2005.
35(4): p. 539-46.
16. Fritsche, R., et al., IgE-mediated rat mast cell triggering with tryptic and synthetic peptides
of bovine beta-lactoglobulin. Int Arch Allergy Immunol, 2005. 138(4): p. 291-7.
17. Knippels, L.M., et al., Comparison of antibody responses to hen’s egg and cow’s milk proteins
in orally sensitized rats and food-allergic patients. Allergy, 2000. 55(3): p. 251-8.
18. Bevilacqua, C., et al., Goats’ milk of defective alpha(s1)-casein genotype decreases intestinal
and systemic sensitization to beta-lactoglobulin in guinea pigs. J Dairy Res, 2001. 68(2): p.
217-27.
19. Buchanan, B.B. and O.L. Frick, The dog as a model for food allergy. Ann N Y Acad Sci, 2002.
964: p. 173-83.
20. Peng, H.J., et al., Effect of ingestion of cow’s milk hydrolysed formulas on whey protein-

73
3
Chapter 3
specific Th2 immune responses in naive and sensitized mice. Clin Exp Allergy, 2004. 34(4): p.
663-70.
21. Peng, H.J., et al., Effect of ingestion of cow’s milk protein hydrolysate formulas on alpha-
casein-specific immunoglobulin E and G1 antibody responses in naive and sensitized mice. J
Pediatr Gastroenterol Nutr, 2005. 41(4): p. 438-44.
22. Li, X.M., et al., A murine model of IgE-mediated cow’s milk hypersensitivity. J Allergy Clin
Immunol, 1999. 103(2 Pt 1): p. 206-14.
23. Frossard, C.P., C. Hauser, and P.A. Eigenmann, Antigen-specific secretory IgA antibodies in
the gut are decreased in a mouse model of food allergy. J Allergy Clin Immunol, 2004. 114(2):
p. 377-82.
24. Kraneveld, A.D., et al., Key role for mast cells in nonatopic asthma. J Immunol, 2002. 169(4):
p. 2044-53.
25. Moolenbeek, C. and E.J. Ruitenberg, The “Swiss roll”: a simple technique for histological
studies of the rodent intestine. Lab Anim, 1981. 15(1): p. 57-9.
26. Rijnierse, A., et al., Critical role for mast cells in the pathogenesis of 2,4-dinitrobenzene-
induced murine colonic hypersensitivity reaction. J Immunol, 2006. 176(7): p. 4375-84.
27. van Meeteren, M.E., et al., Intestinal permeability and contractility in murine colitis.
Mediators Inflamm, 1998. 7(3): p. 163-8.
28. Takaki, M., Gut pacemaker cells: the interstitial cells of Cajal (ICC). J Smooth Muscle Res,
2003. 39(5): p. 137-61.
29. Mule, F., M.C. Baffi, and M.C. Cerra, Dual effect mediated by protease-activated receptors on
the mechanical activity of rat colon. Br J Pharmacol, 2002. 136(3): p. 367-74.
30. Pourpak, Z., et al., The role of cow milk allergy in increasing the severity of atopic dermatitis.
Immunol Invest, 2004. 33(1): p. 69-79.
31. Sicherer, S.H. and H.A. Sampson, Cow’s milk protein-specific IgE concentrations in two age
groups of milk-allergic children and in children achieving clinical tolerance. Clin Exp Allergy,
1999. 29(4): p. 507-12.
32. van Houwelingen, A.H., et al., Topical application of F991, an immunoglobulin free light
chain antagonist, prevents development of contact sensitivity in mice. Clin Exp Allergy, 2007.
37(2): p. 270-5.
33. Ueno, H., K. Yoshioka, and T. Matsumoto, Usefulness of the skin index in predicting the
outcome of oral challenges in children. J Investig Allergol Clin Immunol, 2007. 17(4): p. 207-
10.
34. Tiemessen, M.M., et al., Cow’s milk-specific T-cell reactivity of children with and without
persistent cow’s milk allergy: key role for IL-10. J Allergy Clin Immunol, 2004. 113(5): p. 932-
9.
35. Busse, P.J., et al., Identification of sequential IgE-binding epitopes on bovine alpha(s2)-casein
in cow’s milk allergic patients. Int Arch Allergy Immunol, 2002. 129(1): p. 93-6.
36. Jarvinen, K.M., et al., IgE and IgG binding epitopes on alpha-lactalbumin and beta-
lactoglobulin in cow’s milk allergy. Int Arch Allergy Immunol, 2001. 126(2): p. 111-8.

74
3
Acute allergic skin reactions and intestinal contractility changes in mice orally sensitized against casein or whey
37. Bruhns, P., S. Fremont, and M. Daeron, Regulation of allergy by Fc receptors. Curr Opin
Immunol, 2005. 17(6): p. 662-9.
38. Dombrowicz, D., et al., Absence of Fc epsilonRI alpha chain results in upregulation of Fc
gammaRIII-dependent mast cell degranulation and anaphylaxis. Evidence of competition
between Fc epsilonRI and Fc gammaRIII for limiting amounts of FcR beta and gamma chains.
J Clin Invest, 1997. 99(5): p. 915-25.
39. Redegeld, F.A., et al., Immunoglobulin-free light chains elicit immediate hypersensitivity-like
responses. Nat Med, 2002. 8(7): p. 694-701.
40. Frossard, C.P., et al., Lymphocytes in Peyer patches regulate clinical tolerance in a murine
model of food allergy. J Allergy Clin Immunol, 2004. 113(5): p. 958-64.
41. Morafo, V., et al., Genetic susceptibility to food allergy is linked to differential TH2-TH1
responses in C3H/HeJ and BALB/c mice. J Allergy Clin Immunol, 2003. 111(5): p. 1122-8.
42. Fritsche, R., Animal models in food allergy: assessment of allergenicity and preventive
activity of infant formulas. Toxicol Lett, 2003. 140-141: p. 303-9.
43. Brown, J.K., et al., Constitutive secretion of the granule chymase mouse mast cell protease-1
and the chemokine, CCL2, by mucosal mast cell homologues. Clin Exp Allergy, 2003. 33(1): p.
132-46.
44. Pemberton, A.D., et al., Anaphylactic release of mucosal mast cell granule proteases: role of
serpins in the differential clearance of mouse mast cell proteases-1 and -2. J Immunol, 2006.
176(2): p. 899-904.
45. Cummings, J.H., et al., PASSCLAIM--gut health and immunity. Eur J Nutr, 2004. 43 Suppl 2: p.
II118-II173.
46. Crowe, S.E. and M.H. Perdue, Gastrointestinal food hypersensitivity: basic mechanisms of
pathophysiology. Gastroenterology, 1992. 103(3): p. 1075-95.
47. Kobayashi, M., et al., Regionally differential effects of sennoside A on spontaneous
contractions of colon in mice. Basic Clin Pharmacol Toxicol, 2007. 101(2): p. 121-6.
48. Cook, T.A., A.F. Brading, and N.J. Mortensen, Abnormal contractile properties of rectal
smooth muscle in chronic ulcerative colitis. Aliment Pharmacol Ther, 2000. 14(10): p. 1287-
94.
49. Mitolo-Chieppa, D., et al., Cholinergic stimulation and nonadrenergic, noncholinergic
relaxation of human colonic circular muscle in idiopathic chronic constipation. Dig Dis Sci,
1998. 43(12): p. 2719-26.
50. Mule, F., et al., Involvement of nitric oxide and tachykinins in the effects induced by protease-
activated receptors in rat colon longitudinal muscle. Br J Pharmacol, 2003. 139(3): p. 598-
604.
51. Calbet, J.A. and J.J. Holst, Gastric emptying, gastric secretion and enterogastrone response
after administration of milk proteins or their peptide hydrolysates in humans. Eur J Nutr,
2004. 43(3): p. 127-39.
52. Dangin, M., et al., Influence of the protein digestion rate on protein turnover in young and
elderly subjects. J Nutr, 2002. 132(10): p. 3228S-33S.

75
3
Chapter 3
53. Hall, W.L., et al., Casein and whey exert different effects on plasma amino acid profiles,
gastrointestinal hormone secretion and appetite. Br J Nutr, 2003. 89(2): p. 239-48.
54. Toden, S., et al., Differential effects of dietary whey, casein and soya on colonic DNA damage
and large bowel SCFA in rats fed diets low and high in resistant starch. Br J Nutr, 2007. 97(3):
p. 535-43.
55. Daniel, H., M. Vohwinkel, and G. Rehner, Effect of casein and beta-casomorphins on
gastrointestinal motility in rats. J Nutr, 1990. 120(3): p. 252-7.
56. Defilippi, C., et al., Inhibition of small intestinal motility by casein: a role of beta casomorphins?
Nutrition, 1995. 11(6): p. 751-4.


4
Contribution of IgE and Immunoglobulin free light chain in the allergic reaction to cow’s milk proteins
Bastiaan Schouten1
Betty C.A.M. van Esch1,2
Bart R.J. Blokhuis1 Tom Groot Kormelink1
Gerard A. Hofman1
Guido E. Moro3
Günther Boehm4,5
Sertac Arslanoglu3
Linette E.M.Willemsen1
Léon M.J. Knippels2
Frank A. Redegeld1
Johan Garssen1,2
1 Department of Pharmacology and Pathophysiology, Utrecht Institute for Pharmaceutical Sciences, Faculty of Science, Utrecht University, Utrecht, The Netherlands
2 Danone Research – Centre for Specialised Nutrition, Wageningen, The Netherlands3 Center for Infant Nutrition, Macedonio Melloni Maternity Hospital, Milan, Italy
4 Danone Research – Centre for Specialised Nutrition, Friedrichsdorf, Germany5 Sophia Children’s Hospital, Erasmus University, Rotterdam, The Netherlands
Submitted for publication

78
4
Contribution of IgE and Immunoglobulin free light chain in the allergic reaction to cow’s milk proteins
Abstract
Background: Cow’s milk allergy (CMA) is affecting 2-5% of young infants. A group of CMA patients has no detectable cow’s milk specific serum immunoglobulin (Ig)E. In previous studies it was observed that allergic sensitization of mice to the major cow’s milk allergens, casein and whey, led respectively to IgE-independent and IgE-dependent clinical responses.Objectives: In this study, the mechanism of the IgE-independent hypersensitivity response to casein was further explored.Methods: Mice were orally sham-, casein- or whey-sensitized. Acute allergen specific skin responses were determined, serum immunoglobulins and immunoglobulin free light chains (Ig-fLC) were measured. Ig-fLC dependency was investigated using a specific blocker and passive transfer studies of spleen supernatants.Results: After sensitization, no specific IgE was detectable in serum of casein-sensitized mice, while in whey-sensitized mice specific IgE was enhanced. Instead, Ig-fLC levels were increased in serum from casein-sensitized mice. Furthermore, blocking Ig-fLC strongly diminished allergic skin responses in casein-sensitized mice. In addition, allergic sensitization could be transferred using spleen cell culture supernatant from casein-sensitized mice and was found to be Ig-fLC dependent.Conclusions: This study indicates that sensitization with cow’s milk proteins can lead to both IgE-dependent and Ig free light chain-dependent allergic hypersensitivity responses and suggests that measurement of Ig-fLC may be important in the diagnosis of allergy. The possible clinical relevance of these results was further substantiated by the finding that kappa free light chains were significantly increased in mild atopic dermatitis patients.

79
4
Chapter 4
Introduction
Cow milk allergy (CMA) is a complex disorder, arising early in life and affecting 2-5% of the children of the western world. CMA is also associated with an increased risk to develop asthma later in life [1, 2]. The majority of CMA patients is allergic to the major milk proteins casein and/or whey [3, 4]. Usually, reactions to a skin prick test (SPT) are measured and/or titers of cow’s milk specific immunoglobulin (Ig)E are analyzed in the serum of patients. However, there are CMA patients which exhibit acute clinical features of CMA without detectable titers of specific IgE [3, 5, 6]. Therefore the ‘golden standard’ for diagnosis is still an orally induced double blind food challenge [7-10]. The main disadvantage of an oral food challenge is the time consuming character and the potential of inducing an anaphylactic shock. Although the incidence of anaphylaxis is low, its potential risk is quite stressful for the patient [7]. Therefore the search for another clinical tool to diagnose CMA is ongoing. So far, no (immuno)therapy is available and merely avoidance of cow’s milk and the use of hydrolyzed formulas are the only effective strategies to prevent symptoms of CMA [10].
Previously, two preclinical models for CMA have been introduced in which mice were sensitized orally for whey or casein [11]. In these models the acute allergic skin reaction was monitored as a possible equivalent of the SPT. In both models all mice exhibit an enhanced ear swelling upon intra dermal (i.d.) allergen challenge, which reflects systemic sensitization. The whey model resembles a typical type I allergy with high levels of whey-specific IgE and IgG
1. However, despite developing a pronounced acute allergic skin reaction upon local allergen challenge, the response to casein was not associated with detectable levels of casein-specific IgE. Although the casein-sensitized mice did have enhanced specific titers of IgG1 this was found not to correlate quantitatively with the skin reaction [11]. Therefore, an alternative explanation for the induction of the acute allergic skin reaction was explored. In previous studies, it was demonstrated that antigen-induced acute allergic responses could be elicited via immunoglobulin free light chains (Ig-fLC) [12-14]. For instance, transfer of trinitrophenol-specific Ig-fLC into naïve mice sensitized them to the respective antigen. Local challenge of the passively sensitized animals with the appropriate antigen resulted in the induction of mast cell degranulation, leading to a local inflammation. This Ig-fLC-elicited hypersensitivity response can be inhibited by local or systemic application of a specific antagonist, a 9-mer peptide F991. Acute allergic responses induced by IgG or IgE are not inhibited by F991 [12, 14]. Immunoglobulin free light chains (Ig-fLC) are present in serum and their production is augmented under inflammatory conditions including allergic as well as some autoimmune diseases [12, 15, 16]. For example, in patients suffering from multiple sclerosis, free kappa light chains correlate with disability prognosis [17] and in rheumatoid arthritis there is a significant correlation between kappa and lambda fLC and the disease activity score [18].

80
4
Contribution of IgE and Immunoglobulin free light chain in the allergic reaction to cow’s milk proteins
In this study it is demonstrated that circulatory Ig-fLC are increased after oral sensitization with casein. Moreover, acute allergic skin reactions to casein are inhibited by F991, which indicates that Ig-fLC plays a pivotal role in the allergic sensitization. Preliminary clinical data indicate that Ig-fLC concentrations are increased in children at risk for allergy with mild atopic dermatitis (AD).
Methods
Chemicals
Casein and whey were obtained from DMV international, Veghel, The Netherlands. Cholera toxin is purchased from Quadratech Diagnostics, Epsom, UK. PBS from Cambrex Bio Science, Verviers, Belgium. Biotin-labeled rat anti-mouse IgE and IgG1 from BD Biosciences, Alphen aan den Rijn, The Netherlands. The Ig-fLC antagonist F991 (AHWSGHCCL) was synthesized by Fmoc chemistry, Ansynth, Roosendaal, The Netherlands. All other chemicals were obtained from Sigma-Aldrich-Chemie, Zwijndrecht, The Netherlands.
Oral sensitization and challenge of mice
Three- to 5-week-old specific pathogen free female C3H/HeOuJ mice (n = 6 per group) were purchased from Charles River Laboratories (Maastricht, The Netherlands), maintained on cow’s milk protein free mouse chow (Special Diets Services, Witham, Essex, UK) and housed in the animal facility at the Utrecht University. Animal care and use were approved by and performed in accordance with the guidelines of the Animal Ethics Committee of the Utrecht University. Mice were sensitized i.g. with 0.5 mL homogenized casein or whey (40 mg/mL PBS) with cholera toxin (CT, 20 μg/mL PBS) as an adjuvant, using a blunt needle. Control mice received CT alone or PBS. Mice were boosted weekly for a period of 5 weeks. One week after last sensitization an i.d. ear challenge was performed. After 24 hours blood samples were collected and centrifuged (15 min at 16000 x g). Sera were stored at – 70oC. Mice were sacrificed by cervical dislocation. Cells of spleen and mesenteric lymph nodes of sensitized mice were collected and cultured in serum free RPMI (1 x 106 cells/mL) for 24 hours [14]. Supernatants were collected in order to measure Ig and/or Ig-fLC levels.

81
4
Chapter 4
Measurement of specific serum immunoglobulins
Levels of casein- or whey-specific IgE and IgG1 were determined in serum by means of ELISA. Microlon plates (Greiner, Alphen aan den Rijn, The Netherlands) were coated with 100 µL whey or casein (20 µg/mL) in coating buffer (carbonate-bicarbonate buffer, 0.05 M, pH=9.6) for 18 hours at 4oC. Plates were washed and blocked for 1 hour with 5% BSA. Serum samples were applied in several dilutions and incubated for 2 hours at room temperature. Plates were washed and incubated with biotin-labeled rat anti-mouse IgE or IgG1 (1 μg/mL) for one and a half hour at room temperature and washed. The plates were incubated with streptavidin-horseradish peroxidase (HRP) for one hour, washed and developed with o-phenylendiamine. After 5 min the reaction was stopped with 4M H2SO4 and absorbance was measured at 490 nm on a Benchmark microplate reader (Biorad, California, USA). Results were expressed as arbitrary units (AU), with pooled sera from casein and whey alum-i.p. immunized mice used as a positive reference serum to make the titration curve, as an internal standard.
Immunoblotting Ig-fLC
Prior to use, serum samples were precipitated to deplete high amounts of albumin using trichloroacetic acid/acetone, as previously described [19]. Samples were fractionated by SDS-PAGE under non-reducing conditions, electroblotted to PVDF membrane (Bio-Rad Laboratories, Veenendaal, The Netherlands) overnight and probed with anti-mouse kappa HRP Ab (Southern biotech Birmingham, Alabama USA). Immunoreactive bands were visualized using enhanced chemiluminescence. Optical density (OD) of the immunoreactive bands was quantified with a calibrated densitometer (BioRad, Veenendaal, The Netherlands).
Acute allergic skin reaction
The allergen-specific skin reaction was measured after injection of the specific protein in the ear pinnae. Non-sensitized mice were injected i.d. in the left ear with 20 μL casein (0.5 mg/mL in PBS) and in the right ear whey (0.5 mg/mL in PBS) to determine non-specific skin reactions. The casein- and whey-sensitized mice received in both ears either casein or whey, respectively. Ear thickness of mice was measured in duplicate using a digital micrometer (Mitutoyo, Veenendaal, The Netherlands), at t=0 and 1 hour after challenge. The ear swelling is expressed as μm.

82
4
Contribution of IgE and Immunoglobulin free light chain in the allergic reaction to cow’s milk proteins
Passive transfer with spleen supernatants
Spleen supernatants were concentrated using vivaspin 20 spin filters (Millipore). Concentrated fractions were 0.2 µm filtered and transferred i.v. (100 µL, approximately 400 µg protein/mL) in naïve recipient mice. Sham mice received PBS as a control. After half an hour skin reaction tests were performed as described above.
Antagonist studies
In the oral sensitized mice as well as in mice passively transferred with spleen supernatants of sensitized mice, F991, an Ig-fLC inhibitor [13, 20], or PBS as a control was administered i.v. (100 μL, 0.2 mg/mL F991) to determine whether the ear swelling was Ig-fLC dependent or not. Half an hour after F991 administration the ear swelling test was performed as described above.
Infants at high risk for allergy; serum Immunoglobulin kappa- and lambda-fLC levels
Term born infants with a parental history of atopic eczema, allergic rhinitis or asthma in either mother or father were eligible for a prebiotic intervention study, which was described before [21, 22]. The study protocol was approved by the Ethical Committee of the Macedonio Melloni Maternity Hospital in Italy. In a subpopulation plasma immunoglobulin levels were measured as described by Van Hoffen et al. (CMP-IgE, IgG
1, total- IgE, IgG1 and IgG2) [22]. In total, 25 infants had mild symptoms of AD (SCORAD 5.2 – 24.7) and 49 infants did not develop AD at the age of 6 months. For this study total levels of Ig-fLC kappa and lambda were determined in plasma as described previously [12, 23].
Statistical analysis
Ear swelling data and murine Ig-fLC data were analyzed using one way ANOVA and post hoc Dunnett’s test and all other serological data were analyzed using two-tailed Mann-Whitney tests. Statistical analyses were conducted using GraphPad Prism software (version 4.03). * means a p-value of <0.05, ** p<0.01 and *** p<0.001. Murine data are represented as mean ± SEM, whereas clinical data are represented as median with interquartile range (IQR).

83
4
Chapter 4
Results
Serum specific immunoglobulin levels
Serum levels of casein-specific IgE were not increased in mice orally sensitized with casein when compared to control mice (4.7 ± 1.6 vs 6.0 ± 3.5 AU, respectively) (Figure 1a). In contrast, casein-specific IgG1 levels were enhanced (404.8 ± 217.5 vs 0 ± 0 AU), although this was not significant due to two non-responders (Figure 1b). In whey-sensitized mice both specific IgE (239.4 ± 123.6 AU; p<0.05) and IgG1 (2671 ± 910 AU; p<0.001) were significantly increased compared to sham-sensitized mice (2.5 ± 1.1 and 14.0 ± 11.2 AU, respectively) (Figure 1c and d).
Ig-fLC levels in sensitized mice
Next, it was examined if sensitized mice had enhanced total levels of Ig-fLC. Sera of non-,
Figure 1. Specific serum immunoglobulin levels in casein- and whey-sensitized mice. In the casein-sensitized mice no increase in casein-specific IgE was found a, but IgG1 b was increased in 4 out of 6 animals. Whey-sensitization induced levels of specific IgE c and IgG1 d. * p<0.05, ** p<0.01 and n=6.

84
4
Contribution of IgE and Immunoglobulin free light chain in the allergic reaction to cow’s milk proteins
Figure 2. Densitometric analysis of kappa Ig-fLC in serum and supernatant from local mesenteric lymph nodes (MLN) revealed Ig-fLC levels to be increased. a Analysis of kappa Ig-fLC in serum from individual mice. In casein-sensitized mice, levels of Ig-fLC were significantly increased in comparison to control- or whey-sensitized mice. b Representative Western-blot of Ig-fLC levels in supernatants of MLN of control-, casein- and whey-sensitized mice. Igs are total immunoglobulins (IgA, IgD, IgE, IgG and IgM), (Lc)2 are Ig-fLC dimers and Ig-fLC are the monomers. * p<0.05 and n=6.
250150100
75
50
37
2520
15
kDaCasein
Whe
y
Control
Ig‐fLC
(Lc)2
Igs
b
Figure 3. Effect of F991 on the induction of an acute allergen-specific skin reaction in (a) casein- and (b) whey-sensitized mice. F991 decreased casein-induced ear swelling significantly but whey-induced ear swelling was unaffected. Ear swelling (µm) is calculated as the increase in ear thickness induced by the corresponding antigen at one hour after challenge. ** p<0.01, *** p<0.001 and n=6.

85
4
Chapter 4
Figure 4. Effect of F991 on the induction of an acute allergic skin reaction in naïve mice injected (i.v.) with spleen supernatants or PBS as a control. Passive transfer with spleen supernatants of a casein and b whey allergic mice resulted in acute allergic skin reaction upon i.d. allergen challenge. Only in casein recipients the acute allergic skin reaction was abrogated by F991. *** p<0.001 and n=6.
casein- and whey-sensitized mice were analyzed by immunoblotting for the presence of kappa Ig-fLC. The concentrations of kappa Ig-fLC in casein-sensitized mice were significantly increased compared to non- and whey-sensitized animals (p<0.05; non-sensitized: 66.6 ± 5.5; whey 64.2 ± 8.7 and casein 92.3 ± 9.5 OD\mm2, Figure 2a).
To analyze production of Ig-fLC in local lymph nodes, cells from mesenteric lymph nodes (MLN, n=6 pooled) were cultured for 24 hours and supernatants were analyzed for kappa Ig-fLC. Samples fractionated by SDS-PAGE under non-reducing conditions showed complete immunoglobulins (about 180-200 kDa), Ig-fLC dimers (about 45 kDa) and Ig-fLC monomers (20 kDa). The MLN of casein-sensitized mice produced higher levels of Ig-fLC compared to the control and whey-sensitized animals (Figure 2b).
Ig-fLC inhibitor F991 blocks the acute allergic skin reaction in casein-sensitized mice
To analyze whether the increase in Ig-fLC in casein-sensitized animals was functional relevant regarding the allergic reaction, the effect of the antagonist F991 was tested on the allergen-induced ear swelling. F991 greatly reduced the skin reaction in the casein-sensitized mice (147.4 ± 16.0 µm without and 83.3 ± 8.3 µm with F991, p<0.05, Figure 3a). As expected, the Ig-fLC antagonist did not affect the ear swelling of whey-sensitized mice (Figure 3b, 145.0 ± 9.66 and 148.5 ± 13.9 µm). Also non-specific ear swelling (irritant reaction) in control mice was not affected by the F991. Casein did not provoke an ear swelling in whey-allergic mice and vice versa (data not shown).

86
4
Contribution of IgE and Immunoglobulin free light chain in the allergic reaction to cow’s milk proteins
Table I. Immunoglobulin levels in infants at high risk for allergy, divided in healthy (SCORAD -) and AD (SCORAD +) groups.
Atopic dermatitis
SCORAD - SCORAD +
Number of infants n=49 n=25
Median (IQR) 0.0 (0) 6.9 (6.5)
Serum antibody Isotype Median (IQR) Median (IQR) p-value
Total IgE (kU/L) 4.50 (9) 6.00 (22) p=0.30
IgG1 (g/L) 2.69 (1.9) 2.95 (2.5) p=0.15
IgG2 (g/L) 0.78 (0.4) 0.91 (0.6) p=0.41
CMP-specific IgE (ng/mL) 1.95 (3.1) 2.90 (3.2) p=0.10
IgG1 (AU/mL) 1.50 (6.5) 3.25 (7.7) p=0.27
IQR, interquartile range
Figure 5. Infants at risk for allergy with mild symptoms of AD (SCORAD +, n=25) had enhanced levels of a kappa Ig-fLC compared to infants without AD (SCORAD -, p=0.0075, n=49). b Lambda Ig-fLC showed a similar tendency (p=0.14). Medians are presented.
Ig-fLC responsible for passive transfer of allergic sensitization to casein
In the previous experiments, it was shown that local lymphoid organs from casein-sensitized mice produced significantly more Ig-fLC compared to sham- and whey-sensitized animals. To investigate the involvement of Ig-fLC as a humoral factor in the induction of the acute allergic skin reaction, naïve recipient mice were injected intravenously (i.v.) with concentrated spleen supernatants (or PBS as a control), obtained from allergic mice, in presence or absence of F991 as previously described [14]. Subsequently, the cutaneous

87
4
Chapter 4
swelling was measured at one hour after intradermal (i.d.) allergen challenge. For recipient mice receiving spleen supernatants of casein-sensitized mice the allergen challenge induced a significant ear swelling (110.9 ± 10.2 µm). F991 inhibited the casein-induced ear swelling highly significantly (57.1 ± 5.7 µm, p<0.001, Figure 4a). The Ig-fLC antagonist did not affect the ear swelling of recipient mice receiving spleen supernatants of whey-sensitized mice (105.9 ± 6.6 and 105.0 ± 5.1 µm, Figure 4b), confirming that Ig-fLC did not play a significant role in the induction of the acute allergic skin reaction against whey protein. Casein did not provoke an ear swelling in whey spleen supernatant transferred mice and vice versa (data not shown).
Immunoglobulin levels and Ig-fLC levels in infants at high risk for allergy
In order to test the clinical relevance of the current findings, immunoglobulin levels and Ig-fLC levels were measured in plasma from children at risk of developing allergic disease from a double-blind, randomized, placebo controlled prebiotic intervention study [21, 22]. These infants were six months of age and developed AD (SCORAD between 5.2 and 24.7) or remained negative for AD (SCORAD -). No differences were found between the two groups for total levels of IgE, IgG
1, IgG2, cow’s milk-specific IgE and IgG1 (Table I).
In support to our findings, in the preclinical studies Ig-fLC kappa levels were found to be increased in the infants that developed AD (n=25) when compared to infants without AD (n=49) (median (IQR) 7.4 (3.0) vs 6.1 (4.0) µg/mL, p=0.0075, Figure 5a) Ig-fLC lambda levels showed the same tendency (7.7 (5.6) vs 6.1 (4.1) µg/mL, p=0.14, Figure 5b).
Discussion
In previous studies, it was shown that the allergic skin response in mice sensitized with casein or whey, both major allergens responsible for CMA, is substantially different [10, 11, 24]. Whey-sensitized mice show a classical type I allergic skin reaction combined with high levels of specific IgE. However, in casein-sensitized mice, acute allergic responses were elicited in the absence of casein-specific IgE. Here, new data provide evidence that casein-sensitization of mice results in the induction of an immunoglobulin free light chain (Ig-fLC) dependent hypersensitivity response.
Food allergy is thought to be mediated by IgE and non-IgE dependent mechanisms. IgE mediated food allergy is a classical type I allergy which manifests itself within a few hours.

88
4
Contribution of IgE and Immunoglobulin free light chain in the allergic reaction to cow’s milk proteins
Non-IgE mediated food allergy usually is defined as a delayed-type hypersensitivity with enhanced T-helper 1 activity and the symptoms are noticeable after one or more days while cellular mechanisms are involved [25-29]. However, of interest is a substantial group of patients which have symptoms of an immediate type hypersensitivity response, while lacking detectable levels of IgE [3, 5, 6]. In contrast to the murine model, in humans casein can elicit strong IgE responses [3]. The mice were sensitized for either casein or whey, while in infants hypersensitivity is raised against whey and casein derived from whole cow’s milk. Direct comparison between mice and human with regard to the sensitizing capacities for casein and whey are therefore difficult to perform. However, the concept of Ig-fLC being involved in the pathophysiology of human disease has been described for inflammatory as well as allergic disease [12, 14, 17, 18, 30].
Several lines of evidence suggest the critical involvement of Ig-fLC in the induction of casein hypersensitivity in mice. First, casein-sensitization results in increased levels of Ig-fLC in serum, while local lymph nodes of casein-sensitized mice show enhanced production of Ig-fLC ex vivo. Secondly, inhibition of Ig-fLC induced hypersensitivity using F991 prevented skin response in casein-sensitized mice. A minor part of the ear swelling in casein-sensitized mice was not affected by F991, which could suggest a residual role for IgG
1 or for IgE. In several studies it has been shown that F991 is specific for Ig-fLC and does not affect IgE nor IgG mediated skin responses. Furthermore, F991 prevents local inflammation in different hypersensitivity models [12, 14, 20, 31]. Thirdly, allergic sensitivity to casein could be transferred to naïve animals by i.v. injection of culture supernatant from spleens of casein-sensitized mice and was found to be dependent on Ig-fLC. Together, these data indicate a prominent role for Ig-fLC in the induction of casein allergy.
Other studies in murine models for cow’s milk allergy various results have been described. Oral sensitization with α-casein resulted in an increase of α-casein-specific IgE and IgG
1 titers in C3H mice [32]. On the other hand, Lara-Villoslada et al. [33] only detected whey and casein specific IgG1, while IgE was not detectable in a CMA model induced by simultaneous oral sensitization with casein and whey. The differences in route of sensitization, intraperitoneal vs oral, and variation in allergen fractions e.g. α-casein vs complete casein fraction or combined whey and casein, might explain the contrasting outcome in these and the present studies. The fundamental different mechanisms of allergy observed for casein and whey may be due to the differences in digestive speed, dietary, chemical, physical and physiological features of the proteins [34-37]. Therefore, these differences between the proteins influence not only the intestinal physiology, but likely the response of the immune system as well, which is a topic of further studies. Patients with IgE-mediated CMA have higher specific IgE concentrations to casein compared with whey proteins [3]. It would be interesting to analyze the presence of total or (e.g. casein-) specific Ig-fLC in those patients who suffer from cow´s milk allergy in absence of detectable increases in total or specific

89
4
Chapter 4
IgE.
In previous studies, it has been shown that Ig-fLC can mediate immediate hypersensitivity-like responses. Mast cells are the primary target cells for Ig-fLC and subsequent cross-linking of cell surface-bound Ig-fLC by cognate antigen results in mast cell activation and the induction of a local inflammatory response [14]. Polyclonal Ig-fLC levels are enhanced in chronic inflammatory diseases like inflammatory bowel disease [38], rheumatoid arthritis (16), Sjögren’s syndrome [18], multiple sclerosis [17, 39] and systemic lupus erythematosus [30]. Also in allergic diseases, increases in Ig-fLC have been described. For example, in individuals with allergic and non-allergic asthma, Ig-fLC levels are increased when compared to healthy controls [12]. It is estimated that in one third of the children suffering from AD the skin symptoms are triggered by sensitization to food [40, 41]. Notably, children with severe AD have higher levels of lambda and kappa light chain in serum [42]. Furthermore, in non-IgE mediated rhinitis patients both lambda and kappa Ig-fLC levels are found to be enhanced in nasal secretions (Powe, Groot Kormelink et al., JACI accepted). The present study confirms that increased Ig-fLC may be associated with development of allergic disease. In sera from infants at high risk for allergy neither enhancement of CMP-specific IgE and IgG
1 nor levels of total IgE, IgG1 and IgG2 were found. However, serum levels of kappa Ig-fLC were elevated significantly and lambda Ig-fLC showed a similar increment. This suggests that measurement of total and ultimately, specific levels of Ig-fLC may be an interesting parameter in the clinical diagnosis for AD.
In summary, sensitization with cow’s milk proteins can lead to both IgE-dependent and IgE-independent allergic hypersensitivity responses. In this study, evidence is provided indicating that the IgE-independent allergic response to casein is mediated, at least in part, by specific Ig-fLC. An increase in Ig-fLC levels may be of relevance in allergies without noticeable involvement of IgE. Further research is warranted to determine the clinical significance of assaying Ig-fLC for the diagnosis of allergic diseases.
References
1. Crittenden, R.G. and L.E. Bennett, Cow’s milk allergy: a complex disorder. J Am Coll Nutr,
2005. 24(6 Suppl): p. 582S-91S.
2. Ozol, D. and E. Mete, Asthma and food allergy. Curr Opin Pulm Med, 2008. 14(1): p. 9-12.
3. Shek, L.P., et al., Humoral and cellular responses to cow milk proteins in patients with milk-
induced IgE-mediated and non-IgE-mediated disorders. Allergy, 2005. 60(7): p. 912-9.
4. Fiocchi, A., et al., Clinical tolerance to lactose in children with cow’s milk allergy. Pediatrics,
2003. 112(2): p. 359-62.

90
4
Contribution of IgE and Immunoglobulin free light chain in the allergic reaction to cow’s milk proteins
5. Host, A. and S. Halken, A prospective study of cow milk allergy in Danish infants during
the first 3 years of life. Clinical course in relation to clinical and immunological type of
hypersensitivity reaction. Allergy, 1990. 45(8): p. 587-96.
6. Vanto, T., et al., Prediction of the development of tolerance to milk in children with cow’s
milk hypersensitivity. J Pediatr, 2004. 144(2): p. 218-22.
7. Gabriele, C., et al., Fractional exhaled nitric oxide in infants during cow’s milk food challenge.
Pediatr Allergy Immunol, 2008.
8. Sicherer, S.H. and H.A. Sampson, Cow’s milk protein-specific IgE concentrations in two age
groups of milk-allergic children and in children achieving clinical tolerance. Clin Exp Allergy,
1999. 29(4): p. 507-12.
9. Pourpak, Z., et al., The role of cow milk allergy in increasing the severity of atopic dermatitis.
Immunol Invest, 2004. 33(1): p. 69-79.
10. Monaci, L., et al., Milk allergens, their characteristics and their detection in food: a review.
European Food Research and Technology, 2005(Volume 223, Number 2 / June, 2006): p.
149-179.
11. Schouten, B., et al., Acute Allergic Skin Reactions and Intestinal Contractility Changes in Mice
Orally Sensitized against Casein or Whey. Int Arch Allergy Immunol, 2008. 147(2): p. 125-
134.
12. Kraneveld, A.D., et al., Elicitation of allergic asthma by immunoglobulin free light chains.
Proc Natl Acad Sci U S A, 2005. 102(5): p. 1578-83.
13. Redegeld, F.A. and F.P. Nijkamp, Immunoglobulin free light chains and mast cells: pivotal role
in T-cell-mediated immune reactions? Trends Immunol, 2003. 24(4): p. 181-5.
14. Redegeld, F.A., et al., Immunoglobulin-free light chains elicit immediate hypersensitivity-like
responses. Nat Med, 2002. 8(7): p. 694-701.
15. Thio, M., et al., Free immunoglobulin light chains: a novel target in the therapy of
inflammatory diseases. Trends Pharmacol Sci, 2008. 29(4): p. 170-4.
16. Cooper, A. and R. Bluestone, Free immunoglobulin light chains in connective tissue diseases.
Ann Rheum Dis, 1968. 27(6): p. 537-43.
17. Rinker, J.R., 2nd, K. Trinkaus, and A.H. Cross, Elevated CSF free kappa light chains correlate
with disability prognosis in multiple sclerosis. Neurology, 2006. 67(7): p. 1288-90.
18. Gottenberg, J.E., et al., Serum immunoglobulin free light chain assessment in rheumatoid
arthritis and primary Sjogren’s syndrome. Ann Rheum Dis, 2007. 66(1): p. 23-7.
19. Chen, Y.Y., et al., A modified protein precipitation procedure for efficient removal of albumin
from serum. Electrophoresis, 2005. 26(11): p. 2117-27.
20. van Houwelingen, A.H., et al., Topical application of F991, an immunoglobulin free light
chain antagonist, prevents development of contact sensitivity in mice. Clin Exp Allergy, 2007.
37(2): p. 270-5.
21. Moro, G., et al., A mixture of prebiotic oligosaccharides reduces the incidence of atopic
dermatitis during the first six months of age. Arch Dis Child, 2006. 91(10): p. 814-9.
22. van Hoffen, E., et al., A specific mixture of short-chain galacto-oligosaccharides and long-

91
4
Chapter 4
chain fructo-oligosaccharides induces a beneficial immunoglobulin profile in infants at high
risk for allergy. Allergy, 2009. 64(3): p. 484-7.
23. Abe, M., et al., Differences in kappa to lambda (kappa:lambda) ratios of serum and urinary
free light chains. Clin Exp Immunol, 1998. 111(2): p. 457-62.
24. Bahna, S.L., Cow’s milk allergy versus cow milk intolerance. Ann Allergy Asthma Immunol,
2002. 89(6 Suppl 1): p. 56-60.
25. Eigenmann, P.A., The spectrum of cow’s milk allergy. Pediatr Allergy Immunol, 2007. 18(3):
p. 265-71.
26. Sabra, A., et al., IgE and non-IgE food allergy. Ann Allergy Asthma Immunol, 2003. 90(6 Suppl
3): p. 71-6.
27. Lied, G.A., Gastrointestinal food hypersensitivity: Symptoms, diagnosis and provocation
tests. Turk J Gastroenterol, 2007. 18(1): p. 5-13.
28. Sletten, G.B., et al., Changes in humoral responses to beta-lactoglobulin in tolerant patients
suggest a particular role for IgG(4) in delayed, non-IgE-mediated cow’s milk allergy. Pediatr
Allergy Immunol, 2006. 17(6): p. 435-43.
29. Chehade, M., IgE and non-IgE-mediated food allergy: treatment in 2007. Curr Opin Allergy
Clin Immunol, 2007. 7(3): p. 264-8.
30. Hopper, J.E., et al., Clinical relapse in systemic lupus erythematosus: correlation with
antecedent elevation of urinary free light-chain immunoglobulin. J Clin Immunol, 1989. 9(4):
p. 338-50.
31. Huang, Z.Q. and P.W. Sanders, Localization of a single binding site for immunoglobulin light
chains on human Tamm-Horsfall glycoprotein. J Clin Invest, 1997. 99(4): p. 732-6.
32. Peng, H.J., et al., Effect of ingestion of cow’s milk protein hydrolysate formulas on alpha-
casein-specific immunoglobulin E and G1 antibody responses in naive and sensitized mice. J
Pediatr Gastroenterol Nutr, 2005. 41(4): p. 438-44.
33. Lara-Villoslada, F., M. Olivares, and J. Xaus, The balance between caseins and whey proteins
in cow’s milk determines its allergenicity. J Dairy Sci, 2005. 88(5): p. 1654-60.
34. Calbet, J.A. and J.J. Holst, Gastric emptying, gastric secretion and enterogastrone response
after administration of milk proteins or their peptide hydrolysates in humans. Eur J Nutr,
2004. 43(3): p. 127-39.
35. Dangin, M., et al., Influence of the protein digestion rate on protein turnover in young and
elderly subjects. J Nutr, 2002. 132(10): p. 3228S-33S.
36. Hall, W.L., et al., Casein and whey exert different effects on plasma amino acid profiles,
gastrointestinal hormone secretion and appetite. Br J Nutr, 2003. 89(2): p. 239-48.
37. Toden, S., et al., Differential effects of dietary whey, casein and soya on colonic DNA damage
and large bowel SCFA in rats fed diets low and high in resistant starch. Br J Nutr, 2007. 97(3):
p. 535-43.
38. Rijnierse, A., F.P. Nijkamp, and A.D. Kraneveld, Mast cells and nerves tickle in the tummy:
implications for inflammatory bowel disease and irritable bowel syndrome. Pharmacol Ther,
2007. 116(2): p. 207-35.

92
4
Contribution of IgE and Immunoglobulin free light chain in the allergic reaction to cow’s milk proteins
39. van der Heijden, M., A. Kraneveld, and F. Redegeld, Free immunoglobulin light chains as
target in the treatment of chronic inflammatory diseases. Eur J Pharmacol, 2006. 533(1-3):
p. 319-26.
40. Hauk, P.J., The role of food allergy in atopic dermatitis. Curr Allergy Asthma Rep, 2008. 8(3):
p. 188-94.
41. Werfel, T. and K. Breuer, Role of food allergy in atopic dermatitis. Curr Opin Allergy Clin
Immunol, 2004. 4(5): p. 379-85.
42. Sediva, A., J. Kayserova, and E. Vernerova, Increase of serum immunoglobulin free light
chains in children with atopic dermatitis. Allergy, 2008. 63: p. 157.

93
4
Chapter 4


5
Non-digestible oligosaccharides reduce immunoglobulin free light-chain levels in infants at
risk for allergies
Bastiaan Schouten1
Betty C.A.M. Van Esch1,2 Tom Groot Kormelink1
Guido E. Moro3
Sertac Arslanoglu3
Günther Boehm4,5
Léon M.J. Knippels2
Frank A. Redegeld1
Linette E.M. Willemsen1
Johan Garssen1,2
1 Department of Pharmacology and Pathophysiology, Utrecht Institute for Pharmaceutical Sciences, Faculty of Science, Utrecht University, Utrecht, The Netherlands
2 Danone Research – Centre for Specialised Nutrition, Wageningen, The Netherlands3 Center for Infant Nutrition, Macedonio Melloni Maternity Hospital, Milan, Italy
4 Danone Research – Centre for Specialised Nutrition, Friedrichsdorf, Germany5 Sophia Children’s Hospital, Erasmus University, Rotterdam, The Netherlands
Submitted for publication

96
5
Non-digestible oligosaccharides reduce immunoglobulin free light-chain levels in infants at risk for allergies
Abstract
Background: In previous studies it has been shown that a prebiotic mixture of short-chain galacto-oligosaccharides and long-chain fructo-oligosaccharides (Immunofortis®) reduces the incidence of atopic dermatitis. In this study the plasma immunoglobulin free light-chain (Ig-fLC) concentrations were measured in infants at risk for allergic disease.Methods: In a double-blind, randomized placebo-controlled trial infants received a hypoallergenic whey formula containing 8 g/L of the prebiotics or maltodextrine as a placebo for 6 months. After intervention plasma samples were collected. Total plasma concentrations of lambda and kappa Ig-fLC were analyzed using ELISA.Results: Total plasma levels of lambda and kappa Ig-fLC were lower in infants fed the prebiotic mixture (n=34) compared to the placebo (n=40; p<0.001).Conclusions: These data demonstrate that the specific prebiotic mixture is capable of preventing the lambda and kappa Ig-fLC plasma levels to increase in infants at high risk for allergies. This may have contributed to the reduced incidence of the atopic dermatitis observed.

97
5
Chapter 5
Introduction
The incidence of allergies is increasing in the western world and treatment strategies are limited. Prebiotics as well as probiotics are intensively studied for a possible preventive role due to their capacities to positively influence the immune system [1]. Prebiotics are non-digestible carbohydrates that are fermented by Bifidobacteria and Lactobacilli in the gastrointestinal tract [2]. Immunofortis® is a specific prebiotic mixture, adapted from the breast milk oligosaccharide composition, consisting of short chain Galacto-oligosaccharides (scGOS) and long chain Fructo-oligosaccharides (lcFOS) in a 9:1 ratio. It was shown that oral supplementation of this scGOS/lcFOS prebiotic mixture reduces the cumulative incidence of atopic dermatitis (AD) in infants at high risk for allergies in comparison to the placebo bottle fed infants [3]. In the 6 months follow-up of this study it was shown that total plasma levels of IgE, IgG
1, IgG2 and IgG3, but not IgG4, of the infants who received the prebiotic mixture were reduced. Furthermore, cow’s milk protein (CMP) specific IgG1 levels were decreased, whereas no effect on specific IgE was observed at six months of age [4]. In a preclinical mouse model for orally induced sensitization for cow’s milk protein, specific IgE levels against casein were almost not detected. A role for immunoglobulin free light-chain (Ig-fLC) in the acute allergic skin response was observed (Schouten et al., submitted). Ig-fLC are produced by B cells, bind to mast cells and cause degranulation after second allergen encounter causing an immediate type of allergic response [5]. Ig-fLC concentrations were measured in the same infants evaluated by Moro et al. [3] in a double blind placebo controlled study where neonates at risk for allergies were fed a formula containing a scGOS/lcFOS mixture. This prebiotic mixture was found to reduce plasma levels of both lambda and kappa Ig-fLC, which may relate to the reduced incidence of atopic dermatitis found in this study population.
Methods
Subjects and samples
The study setup has been described by Moro et al. [3, 4]. In short, the study was a randomized double blind placebo controlled trial performed in infants at risk for atopy. Neonates with a parental history of allergic diseases in either mother or father were enrolled in the study and started formula feeding within the first two weeks of life. Two formulas were used, based on extensively hydrolyzed cow’s milk whey protein supplemented either with 0.8 g scGOS/lcFOS (in a 9:1 ratio) or 0.8 g maltodextrin per 100 mL. AD was scored using the SCORAD figure for infants under 2 years by two clinicians. Plasma samples of subpopulation representing

98
5
Non-digestible oligosaccharides reduce immunoglobulin free light-chain levels in infants at risk for allergies
the whole population were obtained after 6 months of intervention. The study protocol was approved by the Ethical Committee of the Macedonio Melloni Maternity Hospital in Italy.
Lambda and kappa immunoglobulin (Ig) free light-chain levels
For this study, total levels of Ig-fLC kappa and lambda were determined in plasma as described previously [6]. For each sample, four different dilutions were incubated, and at least three of them within the linear portion of the standard curves were used to calculate the concentration.
Statistics
Kappa Ig-fLC and lambda Ig-fLC date were analyzed using two-tailed Mann Whitney U test. Statistics were calculated using GraphPad Prism 4.03. Data are represented as medians.
Results
Ig-fLC concentrations were determined in plasma samples that were previously analyzed for
Figure 1. Oral supplementation with short chain Galacto-oligosaccharides (GOS)/ long chain Fructo-oligosaccharides (FOS) reduces the plasma concentrations of both (a) lambda and (b) kappa immunoglobulin free light-chains in infants at high risk for allergy at six months of age. Placebo n=40 and GOS/FOS n=34. *** P < 0.001.

99
5
Chapter 5
immunoglobulin content [4]. Oral supplementation of the prebiotic scGOS/lcFOS mixture significantly reduced plasma levels of both lambda (placebo 8.17 (6.0-11) µg/mL vs scGOS/lcFOS 5.62 (4.7-7.9) µg/mL, p<0.001) and kappa (placebo 7.58 (5.8-9.2) µg/mL vs scGOS/lcFOS 5.81 (4.0-6.7) µg/mL, p<0.001) Ig-fLC in infants at high risk for allergies (Figure 1). In this subgroup of the original study population, 19 out of 40 infants in the placebo group and 6 out of 34 infants in the GOS/FOS intervention group developed AD. The severity of the AD was mild (SCORAD 5.2-24.7).
Conclusions
Evidence for non-digestible carbohydrates as immunomodulatory components is increasing. However, the underlying mechanism of action is still under debate. This study shows that prebiotics affect the humoral immune response in infants at risk for developing allergic diseases. Ig-fLC concentrations were measured in plasma samples derived from a clinical study, described by Moro et al. [3], in which a scGOS/lcFOS mixture was found to reduce the cumulative incidence of AD. The GOS/FOS diet was found to reduce Ig-fLC in infants at risk for allergies.
The presented results are in accordance with the findings of Van Hoffen et al. [4]. In plasma samples of the same study they found GOS/FOS to reduce total levels of immunoglobulins (IgE, IgG
1) and CMP specific levels of IgG1. The prebiotics did not affect CMP specific IgE, however the plasma levels were very low in both groups.
Up to 40% of CMP allergic patients have almost no detectable CMP specific IgE levels [7]. Therefore another mechanism might be involved and Ig-fLC is a potential candidate. Furthermore, there is increasing evidence for the clinical relevance of Ig-fLC in allergic diseases. For example, it has been shown that in individuals with allergic and non-allergic asthma Ig-fLC concentrations are increased when compared to healthy controls [6], and in non-allergic rhinitis with eosinophilic syndrome patients Ig-fLC are enhanced in plasma as well as in nasal secretions (Powe et al., JACI accepted). In this study, GOS/FOS was found to reduce Ig-fLC, Moro et al. [3] have shown GOS/FOS to reduce significantly the cumulative incidence of AD. This indicates a possible role for Ig-fLC in the pathophysiology of AD. Indeed, Sediva et al. [8] showed that in severe AD patients (n=100; SCORAD 50-80) both lambda and, especially, kappa Ig-fLC were enhanced in serum when compared to healthy age matched controls (n=102).
These data demonstrate that dietary intervention with a specific prebiotic GOS/FOS mixture reduces both lambda and kappa plasma Ig-fLC concentrations in infants at risk for allergies.

100
5
Non-digestible oligosaccharides reduce immunoglobulin free light-chain levels in infants at risk for allergies
The reduced Ig-fLC concentrations may have contributed to the reduced incidence of AD in infants fed GOS/FOS.
References
1. Kukkonen, K., et al., Probiotics and prebiotic galacto-oligosaccharides in the prevention
of allergic diseases: A randomized, double-blind, placebo-controlled trial. J Allergy Clin
Immunol, 2007. 119(1): p. 192-8.
2. Hoppu, U., et al., Breast milk--immunomodulatory signals against allergic diseases. Allergy,
2001. 56 Suppl 67: p. 23-6.
3. Moro, G., et al., A mixture of prebiotic oligosaccharides reduces the incidence of atopic
dermatitis during the first six months of age. Arch Dis Child, 2006. 91(10): p. 814-9.
4. van Hoffen, E., et al., A specific mixture of short-chain galacto-oligosaccharides and long-
chain fructo-oligosaccharides induces a beneficial immunoglobulin profile in infants at high
risk for allergy. Allergy, 2009. 64(3): p. 484-7.
5. Redegeld, F.A., et al., Immunoglobulin-free light chains elicit immediate hypersensitivity-like
responses. Nat Med, 2002. 8(7): p. 694-701.
6. Kraneveld, A.D., et al., Elicitation of allergic asthma by immunoglobulin free light chains.
Proc Natl Acad Sci U S A, 2005. 102(5): p. 1578-83.
7. Shek, L.P., et al., Humoral and cellular responses to cow milk proteins in patients with milk-
induced IgE-mediated and non-IgE-mediated disorders. Allergy, 2005. 60(7): p. 912-9.
8. Sediva, A., J. Kayserova, and E. Vernerova, Increase of serum immunoglobulin free light
chains in children with atopic dermatitis. Allergy, 2008. 63: p. 157.

101
5
Chapter 5


6
Cow’s milk allergy symptoms are reduced in mice fed dietary synbiotics during oral sensitization
with whey
Bastiaan Schouten1
Betty C.A.M. van Esch1,2
Gerard A. Hofman1
Suzan A. C. M. van Doorn2
Jan Knol2
Alma J. Nauta2
Johan Garssen1,2
Linette E.M. Willemsen1
Léon M.J. Knippels2
1 Department of Pharmacology and Pathophysiology, Utrecht Institute for Pharmaceutical
Sciences, Faculty of Science, Utrecht University, Utrecht, The Netherlands2 Danone Research – Centre for Specialised Nutrition, Wageningen, The Netherlands
The Journal of Nutrition 2009;139(7):1398-1403

104
6
Cow’s milk allergy symptoms are reduced in mice fed dietary synbiotics during oral sensitization with whey
Abstract
Background: Cow’s milk allergy is the most common food allergy in children. So far, no effective treatment is available to prevent or cure food allergy. The purpose of this study was to compare effects of dietary supplementation with a prebiotic mixture (Immunofortis®), a probiotic strain (Bifidobacterium breve M-16V), or a synbiotic diet combining both on the outcome of the allergic response when provided during oral sensitization with whey in mice.Methods: Mice were fed diets containing 2% (wt:wt) Immunofortis® and/or the B. breve M-16V (n = 6/group). The acute allergic skin response was determined by measuring ear swelling. Antigen-induced anaphylaxis was scored. Furthermore, whey-specific serum immunoglobulins and mouse mast cell protease-1 (mMCP-1) were determined.Results: In mice fed the synbiotic mixture, the allergic skin response and the anaphylactic reaction were strongly reduced compared with whey-sensitized mice fed the control diet (p<0.01). Immunofortis® or B. breve M-16V alone were significantly less effective in reducing the allergic skin response than the synbiotic diet and did not reduce the anaphylactic reaction. The whey-specific IgE and IgG
1 responses were not affected; however, IgG2a was greater in all treated groups than in the control group (p<0.05). Serum mMCP-1 concentrations, reflecting mucosal mast cell degranulation, were lower in mice fed synbiotics compared with those fed the control diet (p<0.01).Conclusions: Dietary supplementation with Immunofortis®, B. breve M-16V, and particularly the synbiotic mixture, provided during sensitization, reduces the allergic effector response in a murine model of IgE-mediated hypersensitivity that mimics the human route of sensitization. This model shows the potential for dietary intervention with synbiotics in reducing the allergic response to food allergens.

105
6
Chapter 6
Introduction
The incidence of food allergy is rising in the western world [1]. Approximately 2–5% of children in the western world are affected with cow’s milk allergy (CMA). Many studies suggest that food allergy in early life is associated with the development of respiratory allergy and even asthma later in life [2-4]. So far, no effective preventive strategy or treatment protocol is available for CMA [5]. Avoidance of the affecting food in the diet is the single effective solution. Some studies suggest that breastfeeding protects the infant from developing allergic disease. Breast-feeding supports maturation of the immune system of the young infants and protects the child from infections [6, 7]. In human milk, there are many components with immunological properties, e.g. non-digestible oligosaccharides, secretory IgA (sIgA), mucins, cytokines, long-chain PUFA, and hormones. In particular, oligosaccharides may support immunocompetence, a prerequisite for adequate capacity to induce oral tolerance, either via direct effects or via stimulation of a beneficial intestinal microbiota [8-10].
Non-digestible prebiotic oligosaccharides are defined as “food ingredients that beneficially affect the host by selectively stimulating the growth and/or activity of 1 or a limited number of bacterial species already resident in the colon, and thus attempt to improve host health” [11]. Non-digestible carbohydrates, which mimic the oligosaccharides in human milk, are known to have immune regulatory properties [12]. In a mouse vaccination model, the vaccine-specific T helper cell (T
H1) response was improved when a prebiotic diet was given starting 1 wk prior to the first vaccination, whereas it failed when the diet was provided after the first vaccination [13]. This indicated that the prebiotics are especially effective during the priming phase of an immune response. Therefore, the prebiotic concept was suggested to be a relevant tool for the prevention of immune disorders such as allergies. In clinical preventive studies in children, early dietary intervention resulted in reduced incidence of allergic manifestations and infections [14], improved microbiota [15], increased fecal sIgA [16], and reduced incidence of atopic dermatitis (AD) [17]. Prebiotic oligosaccharides are known to stimulate the growth and/or activity of probiotic bacteria. Bifidobacteria together with Lactobacilli are well known for their health-promoting functions and selected species are used as probiotics [18-20]. Neonates receiving breastfeeding have higher counts in Bifidobacteria compared with babies fed standard infant formula. When added to standard milk formula, the prebiotic mixture Immunofortis® enhanced Bifidobacteria fecal counts in bottle-fed infants to levels similar to those found in breast-fed infants [21]. In a double-blind, placebo-controlled study, a mixture of probiotics containing Lactobacillus rhamnosus GG, Bifidobacterium breve, and Propionobacterium freudenreichii spp. Shermanii JS alleviated atopic eczema/dermatitis syndrome in IgE-sensitized infants [22]. In a recent review by Savilahti et al. [23], the positive effects of probiotics on allergy prevention and treatment

106
6
Cow’s milk allergy symptoms are reduced in mice fed dietary synbiotics during oral sensitization with whey
have been evaluated. They concluded the effects are highly variable and that optimal treatment remains unsettled. Because prebiotic oligosaccharides selectively stimulate the growth and/or activity of bacterial species such as B. breve, synbiotic concepts are currently introduced combining pro- and prebiotics enabling optimized function of the pro- and prebiotics. The potential of selective pre-, pro-, and synbiotics to prevent the development of allergic disease has been shown in some clinical studies [17, 20, 24-26]. However, the outcomes of these studies vary and questions remain concerning the mechanisms of action. In the present study, we used a mouse model of orally induced CMA to assess preventive effects of dietary intervention with Immunofortis®, a probiotic diet, or a synbiotic diet [27, 28].
Methods
Chemicals
Whey proteins were obtained from DMV International. Cholera toxin (CT) was purchased from Quadratech Diagnostics. PBS was from Cambrex Bio Science, and biotin-labeled rat anti-mouse IgE, IgG1, IgG2a, and IgA were from BD Biosciences. All other chemicals were obtained from Sigma-Aldrich-Chemie.
Diets
Semi purified cow milk protein-free, AIN-93G-based diets were composed and mixed with pro-, pre-, or synbiotics by Research Diet Services [29]. The probiotic strain, B. breve M-16V (freeze-dried on maltodextrin carrier and conserved at -20oC), was obtained from Morinaga (Morinaga Milk Industry) and 2% (wt:wt) 2 X 109 colony forming units/g was mixed through the diets and pressed into pellets. Viability of B. breve M-16V after diet manufacturing was confirmed by plating on MRS agar. The prebiotic diet contained 2% (wt:wt) Immunofortis®, a 9:1 (wt:wt) mixture of short-chain galacto-oligosaccharides (GOS; obtained by enzymatic elongation of lactose with galactose by β-galactosidase, with a degree of polymerization of 3–8) and long-chain fructo-oligosaccharides (FOS; derived from chicory inulin with a mean degree of polymerization of >23). Immunofortis® is a spray-dried powder of GOS (Vivinal GOS, FrieslandCampina Domo, Zwolle, The Netherlands) and long-chain FOS (Raftiline HP). Immunofortis® consists of ~50% GOS and FOS, 19% is maltodextrin (glucidex 2, Roquette), 16% lactose, 14% glucose, and 1% galactose (13). Immunofortis® was exchanged against 2% (wt:wt) of total carbohydrates present in the diet. In total, 4 diets were prepared: control,

107
6
Chapter 6
probiotic, prebiotic, and synbiotic (containing 2% Immunofortis® and B. breve M-16V). The diets were stored at -20oC prior to use.
Oral sensitization and challenge of mice
Three-week-old specific pathogen-free female C3H/HeOuJ mice were purchased from Charles River Laboratories. Animal care and use were performed in accordance with the guidelines of the Dutch Committee of Animal Experiments. Sham- and whey-sensitized mice were fed a control, B. breve M-16V, GOS/FOS, or synbiotic diet (n = 6/group). Two weeks before the first sensitization, the dietary intervention was started and continued during the whole study protocol. Mice were sensitized i.g. with 0.5 mL homogenized whey (40 g/L PBS) with CT (20 mg/L PBS) as an adjuvant using a blunt needle. Control mice received CT alone. Mice were boosted weekly for a period of 5 wk as described previously (27). One week after the last sensitization, feces were collected and mice were challenged with whey intradermally (i.d.) in the ear to determine the acute allergic skin response and anaphylaxis scores. After 24 h, mice were challenged i.g. with 1.0 mL homogenized whey (100 g/L PBS) and 30 min later, blood samples were collected in Minicollect Z Serum Sep vials (Greiner bio-one). Collected blood was centrifuged at 16,000 X g; 15 min and sera were stored at –70oC. The weight of the mice was monitored weekly.
Acute allergen-specific skin response
The acute allergen-specific skin response was measured after i.d. injection with 20 mL homogenized whey protein (0.5 g/L in PBS) in the ear pinnae. Ear thickness was measured in duplicate using a digital micrometer (Mitutoyo) at t = 0 and 1 h after challenge. Delta ear swelling (μm) was calculated by subtracting the specific whey-induced ear swelling in sensitized mice after 1 h from the basal (t = 0 h) ear thickness. Additionally, the mean Δ ear swelling from the group of sham-sensitized mice fed the corresponding diet was subtracted.
Systemic anaphylaxis
Anaphylaxis was scored after i.d. challenge in the ear. Symptoms of systemic anaphylaxis were scored (adapted from Li et al. [30]): 0 = no symptoms; 1 = pilar erecti, scratching

108
6
Cow’s milk allergy symptoms are reduced in mice fed dietary synbiotics during oral sensitization with whey
and rubbing around the nose and head; 2 = pilar erecti, reduced activity; 3 = activity after prodding and lowered body temperature; 4 = no activity after prodding, labored respiration, and lowered body temperature; and 5 = death.
Measurement of specific serum Ig and mouse mast cell protease-1
Concentrations of whey-specific IgE, IgG1 and IgG2a, were determined in serum using ELISA as described previously [27]; specific IgA was measured in feces. In short, microlon plates (Greiner) were coated with 100 μL whey (20 mg/L), washed, and blocked with 5% bovine serum albumin. Samples were applied in several dilutions; for determinations in feces, 1.0 g feces was dissolved in 10 mL PBS and centrifuged at 16,000 X g; 5 min. The supernatant was collected for IgA measurements. Samples were incubated for 2 h at room temperature. Plates were washed and incubated with biotin-labeled rat anti-mouse IgE, IgG
1, IgG2a, or IgA (1 mg/L), washed, and incubated with streptavidin-horseradish peroxidase, washed, and developed with o-phenylendiamine. The reaction was stopped with 4 mol/L H2SO4 and absorbance was measured at 490 nm on a Benchmark microplate reader (Bio-Rad). Results were expressed as arbitrary units with pooled sera from whey- and alum-intraperitoneally–immunized mice used as a positive reference serum to make the titration curve. Serum concentrations of mouse mast cell protease-1 (mMCP-1) were determined according to the manufacturer’s protocol using a commercially available ELISA kit (Moredun Scientific).
Preparation of fecal samples for PCR of B. breve M-16V
For PCR analysis, 0.1 g feces was placed in a Lysing Matrix E tube with 1.0 mL stool lysis buffer (Qiagen). These tubes were placed in a mini-BeadBeater (BioSpec Products) for 3 min, heated for 5 min at 95oC, and centrifuged at 14,000 X g; 5 min. Then 200 μL Lysozym (10 g/L MiliQ) was added to the supernatant, heated for 30 min at 37oC, and centrifuged at 4000 X g; 10 min. DNA was extracted from the collected supernatant using a NucliSENS easyMAG (BioMerieux). PCR was performed as described previously [21] using B. breve M-16V-specific primers. Briefly, primers and probes of the B. breve M-16V were used in a temperature profile consisting of 2 min at 50oC and 10 min at 95oC, followed by 45 cycles of 15 s at 95oC and 60oC for 1 min.

109
6
Chapter 6
Statistics
All data were analyzed using 1-way ANOVA and post hoc Bonferroni’s multiple comparison test except for the anaphylaxis data, these were analyzed using the Kruskal-Wallis test. Statistical analyses were conducted using GraphPad Prism software (version 4.03). P-values <0.05 were considered significant. Data are represented as means ± SEM.
Results
Acute allergic skin response
One hour after dermal challenge, the Δ ear swelling in the whey-sensitized mice was 117.6 ± 8.6 μm (remaining ear swelling upon subtraction of ear swelling in sham-sensitized mice fed control diet, see Methods section). The ear swelling of the whey-sensitized mice fed the pro- and prebiotic diet was lower than in those fed the control diet (p<0.05). In mice fed the synbiotic diet, this effect was even more pronounced (p<0.001; Figure 1). The ear swelling of mice fed the synbiotic diet was reduced (p<0.01) compared with mice fed the pro- and prebiotic diets. The diets themselves did not affect ear swelling in sham-sensitized control mice (data not shown).
Figure 1. Acute allergic skin response of whey-sensitized mice fed a control, B. breve, GOS/FOS, or synbiotic diet. The acute ear swelling response was measured 1 h after i.d. challenge with whey. Values are means ± SEM, n=6. Means without a common letter differ, p<0.05.

110
6
Cow’s milk allergy symptoms are reduced in mice fed dietary synbiotics during oral sensitization with whey
Figure 2. Anaphylactic symptom scores of sham-sensitized mice fed a control diet and whey-sensitized mice fed a control, B. breve, GOS/FOS, or synbiotic diet. Values are medians, n=6. Medians without a common letter differ, p<0.01.
Figure 3. Serum mMCP-1 concentrations of sham-sensitized mice fed a control diet and whey-sensitized mice fed a control, B. breve, GOS/FOS, or synbiotic diet. Values are means ± SEM, n=6. Means without a common letter differ, p<0.01.
Systemic anaphylaxis after i.d. challenge
The mice were i.d. challenged in the ear with whey 1 wk after the last sensitization. In the whey-sensitized group fed the control diet, the anaphylaxis score was enhanced compared with control sham-sensitized mice (p<0.01; Figure 2). In the synbiotic group, anaphylaxis scores were lower than whey-sensitized mice fed the control diet (p<0.01). Neither the pre- nor the probiotic diet alone affected shock scores.

111
6
Chapter 6
Decreased mMCP-1 levels in serum
To assess the effect of dietary intervention on mucosal mast cell degranulation, mMCP-1 concentrations were determined in the serum. Serum mMCP-1 was lower in mice fed the synbiotic diet than in those fed control diet (p<0.01; Figure 3) but was unaffected in mice fed the prebiotic or probiotic diet.
Whey-specific Ig levels in serum or feces
Thirty minutes after oral challenge, serum was obtained from all mice. Whey-specific serum IgE, IgG1, IgG2a, and fecal IgA were analyzed using ELISA. In whey-sensitized mice, specific IgE and IgG1 increased compared with sham-sensitized mice (p<0.01; Figure 4a, b). Overall, dietary intervention did not affect serum IgE and IgG1 levels. In contrast, the probiotic, prebiotic, and synbiotic diets enhanced whey-specific IgG2a levels in whey-sensitized mice (p<0.01) compared with sham-sensitized mice (Figure 4c). The IgG2a levels in whey-sensitized mice fed the control diet were not significantly enhanced compared with sham-sensitized mice. Specific IgA levels were increased in feces of whey-sensitized mice fed the control or B. breve M-16V diet (p<0.01; Figure 4d) and in mice fed the GOS/FOS or synbiotic diet (p<0.001) compared with sham-sensitized mice fed the control diet.
Figure 4. Whey-specific Ig of sham-sensitized mice fed a control diet and whey-sensitized mice fed a control, B. breve, GOS/FOS, or synbiotic diet. Whey-specific serum IgE a, serum IgG
1 b, serum IgG2a c, and fecal IgA D levels were measured. Values are means ± SEM, n=4–6. Means without a common letter differ, p<0.01.

112
6
Cow’s milk allergy symptoms are reduced in mice fed dietary synbiotics during oral sensitization with whey
PCR analysis
PCR analysis on the fecal samples was performed to verify whether the B. breve M-16V was present in the feces of the control or pro-, pre-, and synbiotic treated mice. Indeed, the presence of B. breve M16-V was confirmed only in the mice fed the pro- and synbiotic diets, whereas feces of control mice and mice fed Immunofortis®did not contain B. breve M-16V (data not shown).
Discussion
Both pro- and prebiotics modulate immune function and have been shown to reduce the risk of developing allergic disease in human studies [17, 31, 32]. Host factors, bacterial strain differences, dosage, and timing of supplementation may underlie the differences in effectiveness. There is general agreement that, although there are promising effects, more data are needed [23, 33]. In the present study, dietary intervention with synbiotics combining probiotic strain B. breve M-16V with prebiotic GOS/FOS mixture Immunofortis® was shown to protect against the development of allergic symptoms in mice orally sensitized with whey. Synbiotics effectively reduced the acute allergic skin response, the anaphylaxis scores, and mast cell-derived mMCP-1 concentrations. Pro- and prebiotics alone were less effective. The skin response is a robust parameter and was associated with enhanced serum IgE and mast cell-derived mMCP-1 in previous studies [27]. Mice fed pro- and prebiotic diets had a significantly reduced acute allergic skin response upon whey challenge in the ear compared with whey-sensitized mice fed the control diet. The synbiotic diet was even more effective and almost completely prevented occurrence of the acute skin response as well as anaphylactic shock reactions. The clinical effectiveness of the prebiotic mixture used in this study has previously been shown to reduce the incidence of AD in infants at risk. The newborn infants were randomly assigned to standard formula milk or formula milk containing Immunofortis® until 6 mo of age, reducing the incidence of AD by >50% [17]. Furthermore, infants at high risk for allergy supplemented with the prebiotic mixture had a slight reduction in total IgE and IgG
1–3 and specific IgG1 [34]. IgG4 tended to increase, which might be suggestive for tolerance induction/outgrowing food allergy. Also probiotics have been shown to reduce AD development in infants. When pregnant mothers with a family history of allergic disease and their babies were supplemented with Lactobacillus GG, the cumulative incidence of atopic eczema was still reduced after 7 y by 30% compared with placebo controls [35]. Although the incidence of eczema was reduced, IgE levels were not affected. In a similar study, Lactobacillus reuteri reduced the incidence of IgE-associated eczema and the occurrence of a positive skin prick test for almost 50%, whereas IgE was not affected [32]. As in the human studies, the allergic effector response in the current study

113
6
Chapter 6
using mice orally sensitized with whey was reduced upon dietary intervention with pro-, pre-, and synbiotics, whereas IgE levels remained enhanced. In young infants, development of AD can objectively be scored. Approximately one-third of these children are affected with food allergy. Nevertheless, the occurrence of eczema in AD patients and the acute allergic skin response as measured in mice form reliable measures for skin responses to allergens and, in this respect, this parameter in mice might reflect, at least in part, some aspects of AD. Indeed, a synbiotic diet has been shown to reduce AD-like skin lesions in mice [24].
Dietary intervention with pro-, pre-, and, in particular, synbiotics reduced the acute allergic skin response and the anaphylaxis scores very effectively, which could not be explained by a reduction in whey-specific T
H2 type IgE or IgG1 levels. However, increased levels of whey-specific IgG2a and enhanced fecal type IgA levels may reflect improved TH1 or regulatory T-cell responses in the mice fed a pro-, pre-, or synbiotic diet [16, 36, 37]. In a preclinical murine model, heat-treated L. rhamnosus GG reduced the development of AD probably through the induction of interleukin (IL)-10 [38]. These studies and our study are in accordance with at least 2 clinical studies in which probiotic treatment enhanced (fecal) IgA and IL-10 and alleviated allergic symptoms [23, 39]. Hence, dietary intervention with pre-, pro-, and synbiotics in mice orally sensitized with whey may have supported T
H1 responses and/or regulatory T-cell function.
Besides the acute allergic skin response, anaphylactic scores upon i.d. challenge were dramatically reduced in whey-sensitized mice fed the synbiotic diet. Although in humans the shock usually appears after an oral challenge, at least for food allergens, anaphylaxis in our mouse study occurred after i.d. challenge with whey proteins. Eigenmann et al. [40] studied allergic shock in a murine model of orally induced β-lactoglobulin sensitization. In this study, the anaphylactic shock reaction occurred after oral challenge. Oral inoculation with an avirulent Salmonella typhimurium strain prior to allergic sensitization partially prevented anaphylaxis. This is in accordance with the effects of the B. breve M-16V on anaphylaxis as observed in the current study. So far, to our knowledge, there is no literature available about the preventive effects of pre- and synbiotics on allergen induced anaphylaxis. The dietary effects on the acute allergic skin response and the anaphylactic shock coincided. Mast cell degranulation most probably underlies both symptom scores. Indeed, mMCP-1 serum concentrations reflecting mucosal mast cell degranulation were reduced, particularly in mice fed the synbiotic diet. Hence, the diets may have altered mast cell function. In a murine model of ovalbumin-induced allergy, oral administration of different bacterial strains prevented mast cell degranulation [41]. IgE receptor expression on mast cells can be diminished by CD4+CD25+ regulatory T-cells upon direct contact or via the secretion of IL-10 and transforming growth factor-β [42]. Because whey-specific fecal IgA was enhanced upon dietary intervention, this may be an indication of enhanced regulatory T-cell function. In a clinical study with healthy infants receiving a GOS/FOS diet, fecal sIgA increased and

114
6
Cow’s milk allergy symptoms are reduced in mice fed dietary synbiotics during oral sensitization with whey
it was suggested to have a regulatory function and a positive effect on mucosal immunity [16]. Alternatively, IgG-blocking antibodies may have inhibited IgE mediated anaphylaxis in mice, as was shown by a study from Strait et al. [43]. IgG2a that was enhanced upon dietary intervention may have a function as blocking antibody.
The current study shows very promising effects of the synbiotic concept in prevention of the acute allergic skin response, reducing the incidence and severity of anaphylaxis and reducing mMCP-1 a marker for mast cell degranulation. Little is known about the use of synbiotics to prevent or cure allergic disease. A synbiotic mixture of Lactobacillus casei and dextran reduced pollen allergy in humans [24]. Kukkonen et al. [25] showed that a synbiotic mixture containing L. rhamnosus GG and LC705, B. breve Bb99, and Propionibacterium freudenreichii ssp shermanii combined with GOS supplied to newborn infants was safe, reduced the cumulative incidence of IgE-associated (atopic) diseases, and decreased the occurrence of respiratory infections. In the present study, a preventive dietary intervention study was performed comparing the effects of pro-, pre-, and synbiotics in mice orally sensitized to whey. In particular, the synbiotic combination of Immunofortis® and the probiotic strain B. breve M-16V, and to a lesser extent the pro- and prebiotic diets, reduced the acute allergic skin response and anaphylactic scores, which are clinical relevant allergic parameters. Although the exact mechanism of action is not yet known, dietary intervention enhanced whey-specific T
H1 type serum IgG2a and specific IgA in feces while reducing mast cell degranulation. Synbiotics comprise a promising concept that may be more effective in reducing allergic symptoms than single preparations of pre- or probiotics.
References
1. Bjorksten, B., The epidemiology of food allergy. Curr Opin Allergy Clin Immunol, 2001. 1(3):
p. 225-7.
2. Ozol, D. and E. Mete, Asthma and food allergy. Curr Opin Pulm Med, 2008. 14(1): p. 9-12.
3. Skripak, J.M., et al., The natural history of IgE-mediated cow’s milk allergy. J Allergy Clin
Immunol, 2007. 120(5): p. 1172-7.
4. Sampson, H.A., Update on food allergy. J Allergy Clin Immunol, 2004. 113(5): p. 805-19; quiz
820.
5. Sicherer, S.H. and D.Y. Leung, Advances in allergic skin disease, anaphylaxis, and
hypersensitivity reactions to foods, drugs, and insects in 2007. J Allergy Clin Immunol, 2008.
121(6): p. 1351-8.
6. Vandenplas, Y., Oligosaccharides in infant formula. Br J Nutr, 2002. 87 Suppl 2: p. S293-6.
7. Eigenmann, P.A., Breast-feeding and atopic eczema dermatitis syndrome: protective or
harmful? Allergy, 2004. 59 Suppl 78: p. 42-4.

115
6
Chapter 6
8. Sudo, N., et al., The requirement of intestinal bacterial flora for the development of an IgE
production system fully susceptible to oral tolerance induction. J Immunol, 1997. 159(4): p.
1739-45.
9. Field, C.J., The immunological components of human milk and their effect on immune
development in infants. J Nutr, 2005. 135(1): p. 1-4.
10. Newburg, D.S., Oligosaccharides in human milk and bacterial colonization. J Pediatr
Gastroenterol Nutr, 2000. 30 Suppl 2: p. S8-17.
11. Gibson, G.R. and M.B. Roberfroid, Dietary modulation of the human colonic microbiota:
introducing the concept of prebiotics. J Nutr, 1995. 125(6): p. 1401-12.
12. Vos, A.P., et al., Immune-modulatory effects and potential working mechanisms of orally
applied nondigestible carbohydrates. Crit Rev Immunol, 2007. 27(2): p. 97-140.
13. Vos, A.P., et al., A specific prebiotic oligosaccharide mixture stimulates delayed-type
hypersensitivity in a murine influenza vaccination model. Int Immunopharmacol, 2006. 6(8):
p. 1277-86.
14. Arslanoglu, S., et al., Early dietary intervention with a mixture of prebiotic oligosaccharides
reduces the incidence of allergic manifestations and infections during the first two years of
life. J Nutr, 2008. 138(6): p. 1091-5.
15. Bakker-Zierikzee, A.M., et al., Effects of infant formula containing a mixture of galacto- and
fructo-oligosaccharides or viable Bifidobacterium animalis on the intestinal microflora
during the first 4 months of life. Br J Nutr, 2005. 94(5): p. 783-90.
16. Scholtens, P.A., et al., Fecal secretory immunoglobulin A is increased in healthy infants
who receive a formula with short-chain galacto-oligosaccharides and long-chain fructo-
oligosaccharides. J Nutr, 2008. 138(6): p. 1141-7.
17. Moro, G., et al., A mixture of prebiotic oligosaccharides reduces the incidence of atopic
dermatitis during the first six months of age. Arch Dis Child, 2006. 91(10): p. 814-9.
18. Boehm, G., et al., Supplementation of a bovine milk formula with an oligosaccharide mixture
increases counts of faecal bifidobacteria in preterm infants. Arch Dis Child Fetal Neonatal Ed,
2002. 86(3): p. F178-81.
19. Furrie, E., Probiotics and allergy. Proc Nutr Soc, 2005. 64(4): p. 465-9.
20. Kukkonen, K., et al., Probiotics and prebiotic galacto-oligosaccharides in the prevention
of allergic diseases: A randomized, double-blind, placebo-controlled trial. J Allergy Clin
Immunol, 2007. 119(1): p. 192-8.
21. Haarman, M. and J. Knol, Quantitative real-time PCR assays to identify and quantify fecal
Bifidobacterium species in infants receiving a prebiotic infant formula. Appl Environ
Microbiol, 2005. 71(5): p. 2318-24.
22. Viljanen, M., et al., Probiotics in the treatment of atopic eczema/dermatitis syndrome in
infants: a double-blind placebo-controlled trial. Allergy, 2005. 60(4): p. 494-500.
23. Savilahti, E., M. Kuitunen, and O. Vaarala, Pre and probiotics in the prevention and treatment
of food allergy. Curr Opin Allergy Clin Immunol, 2008. 8(3): p. 243-8.
24. Ogawa, T., et al., A new synbiotic, Lactobacillus casei subsp. casei together with dextran,

116
6
Cow’s milk allergy symptoms are reduced in mice fed dietary synbiotics during oral sensitization with whey
reduces murine and human allergic reaction. FEMS Immunol Med Microbiol, 2006. 46(3): p.
400-9.
25. Kukkonen, K., et al., Long-term safety and impact on infection rates of postnatal probiotic
and prebiotic (synbiotic) treatment: randomized, double-blind, placebo-controlled trial.
Pediatrics, 2008. 122(1): p. 8-12.
26. Lenoir-Wijnkoop, I., et al., Probiotic and prebiotic influence beyond the intestinal tract. Nutr
Rev, 2007. 65(11): p. 469-89.
27. Schouten, B., et al., Acute Allergic Skin Reactions and Intestinal Contractility Changes in Mice
Orally Sensitized against Casein or Whey. Int Arch Allergy Immunol, 2008. 147(2): p. 125-
134.
28. Boehm, G., et al., Prebiotic concept for infant nutrition. Acta Paediatr Suppl, 2003. 92(441):
p. 64-7.
29. Reeves, P.G., Components of the AIN-93 diets as improvements in the AIN-76A diet. J Nutr,
1997. 127(5 Suppl): p. 838S-841S.
30. Li, X.M., et al., A murine model of IgE-mediated cow’s milk hypersensitivity. J Allergy Clin
Immunol, 1999. 103(2 Pt 1): p. 206-14.
31. Kalliomaki, M., et al., Probiotics in primary prevention of atopic disease: a randomised
placebo-controlled trial. Lancet, 2001. 357(9262): p. 1076-9.
32. Abrahamsson, T.R., et al., Probiotics in prevention of IgE-associated eczema: a double-blind,
randomized, placebo-controlled trial. J Allergy Clin Immunol, 2007. 119(5): p. 1174-80.
33. Prescott, S.L. and B. Bjorksten, Probiotics for the prevention or treatment of allergic diseases.
J Allergy Clin Immunol, 2007. 120(2): p. 255-62.
34. Lam, H.Y., et al., Cow’s milk allergy in adults is rare but severe: both casein and whey proteins
are involved. Clin Exp Allergy, 2008. 38(6): p. 995-1002.
35. Kalliomaki, M., et al., Probiotics during the first 7 years of life: a cumulative risk reduction of
eczema in a randomized, placebo-controlled trial. J Allergy Clin Immunol, 2007. 119(4): p.
1019-21.
36. Adel-Patient, K., et al., Oral administration of recombinant Lactococcus lactis expressing
bovine beta-lactoglobulin partially prevents mice from sensitization. Clin Exp Allergy, 2005.
35(4): p. 539-46.
37. Katayama, S. and Y. Mine, Quillaja saponin can modulate ovalbumin-induced IgE allergic
responses through regulation of Th1/Th2 balance in a murine model. J Agric Food Chem,
2006. 54(9): p. 3271-6.
38. Sawada, J., et al., Ingestion of heat-treated Lactobacillus rhamnosus GG prevents
development of atopic dermatitis in NC/Nga mice. Clin Exp Allergy, 2007. 37(2): p. 296-303.
39. Marschan, E., et al., Probiotics in infancy induce protective immune profiles that are
characteristic for chronic low-grade inflammation. Clin Exp Allergy, 2008. 38(4): p. 611-8.
40. Eigenmann, P.A., K.E. Asigbetse, and C.P. Frossard, Avirulant Salmonella typhimurium strains
prevent food allergy in mice. Clin Exp Immunol, 2008. 151(3): p. 546-53.
41. Kim, H., et al., Oral probiotic bacterial administration suppressed allergic responses in an

117
6
Chapter 6
ovalbumin-induced allergy mouse model. FEMS Immunol Med Microbiol, 2005. 45(2): p.
259-67.
42. Kashyap, M., et al., Cutting edge: CD4 T cell-mast cell interactions alter IgE receptor
expression and signaling. J Immunol, 2008. 180(4): p. 2039-43.
43. Strait, R.T., S.C. Morris, and F.D. Finkelman, IgG-blocking antibodies inhibit IgE-mediated
anaphylaxis in vivo through both antigen interception and Fc gamma RIIb cross-linking. J Clin
Invest, 2006. 116(3): p. 833-41.


7
Oligosaccharide induced whey-specific CD25+ regulatory T-cells are crucial in the suppression of
cow’s milk allergy in mice
Bastiaan Schouten1
Betty C.A.M. van Esch1,2
Gerard A. Hofman1
Louis Boon3
Léon M.J. Knippels2
Linette E.M. Willemsen1 Johan Garssen1,2
1 Department of Pharmacology and Pathophysiology, Utrecht Institute for Pharmaceutical Sciences, Faculty of Science, Utrecht University, Utrecht, The Netherlands
2 Danone Research – Centre for Specialised Nutrition, Wageningen, The Netherlands3 Bioceros BV, Utrecht, The Netherlands
Submitted for publication

120
7
Oligosaccharide induced whey-specific CD25+ regulatory T-cells are crucial in the suppression of cow’s milk allergy in mice
Abstract
Background: Dietary intervention with a unique prebiotic non-digestible carbohydrate mixture has been shown to reduce the development of allergic disease in infants at risk. In this study the involvement of CD25+ regulatory T-cells in the carbohydrate induced effects was investigated in mice orally sensitized with whey using adoptive transfer experiments.Methods: Donor mice were sensitized with whey and fed a diet containing short-chain galacto-, long-chain fructo- and acidic-oligosaccharides or control diet starting two weeks before sensitization. The acute allergic skin reaction upon intra dermal whey challenge was determined and whey-specific immunoglobulins were measured. Splenocytes of the donor mice were transferred to naïve recipient mice after ex vivo depletion of CD25+ splenocytes.Results: The prebiotic diet clearly diminished the acute allergic skin reaction. Whey-sensitized recipient mice receiving splenocytes from whey-sensitized prebiotic fed donor mice displayed almost complete prevention of the acute allergic skin reaction compared to mice receiving cells from sham-sensitized prebiotic fed donor mice. Depletion of CD25+ cells inhibited these effects, although IgE sensitization was not prevented.Conclusion: This study points to a crucial role for whey-specific CD25+ regulatory T-cells in suppression of the allergic effector response induced by dietary intervention with prebiotics.

121
7
Chapter 7
Introduction
The gastro-intestinal tract is exposed to a wide variety of dietary proteins. In general, these proteins are tolerized by the gastro-intestinal immune system resulting in local and systemic non-responsiveness. This orally induced tolerance may be a result of anergy or active suppression, which is determined by both antigen dosage [1] and age [2]. In food allergic patients the oral tolerance to specific dietary proteins is broken or not established [3]. Currently, there is no specific therapy for the treatment of food allergy. The only option to avoid symptoms is to circumvent exposure to the responsible food. The most common food allergy in childhood is cow’s milk allergy (CMA) which affects about 2-3% of all infants [4]. It is caused by an allergic inflammatory response to milk proteins [5]. Cow’s milk proteins (3.5% w/v in milk) consist of casein and whey proteins. Whey proteins comprise β-lactoglobulin (the main allergen), α-lactalbumin, albumin, lactoferrin, immunoglobulins and proteosepeptone [6]. In Europe most of the infant formula feedings are whey based and are supplemented with prebiotics.
In food allergy the delegate balance between oral tolerance and hypersensitivity is regulated by cells of the immune system. Cells with regulatory properties include TGF-β-producing T
H3 cells, IL-10-producing Tr1 cells and NKT cells [7-9]. In addition, the TGF-β and IL-10 producing naturally occurring CD4+CD25+Foxp3+ regulatory T-cells (Treg) were described to be important for maintaining peripheral tolerance and for suppression of proliferation of CD4+ and CD8+ T-cells [10, 11]. Treg represent 5-10% of the CD4+ T lymphocyte population and express the transcription factor forkhead box p3 (Foxp3), which is confirmed as a key gene for generation and maintenance of Treg. Adaptive regulatory T-cells have similar surface markers, however their inhibitory capacities act via cell-cell contact instead of cytokines. Currently, the involvement of Treg in tolerance induction by dietary components is unknown. Human milk consists of numerous components, including but not limited to oligosaccharides, which may have many immunological properties and might facilitate tolerance [12, 13]. Prebiotic fibers manufactured to resemble some of these oligosaccharides are defined as “non-digestible food ingredients that beneficially affect the host by selectively stimulating the growth and/or activity of one or a limited number of bacterial species already resident in the colon, and thus attempt to improve host health” [14]. Short-chain galacto-oligosaccharides (scGOS) and long-chain fructo-oligosaccarides (lcFOS) are dominating the prebiotic literature. Beneficial immunological properties of these scGOS and lcFOS are confirmed by an improved vaccination response in a murine model [15] and lowered incidence of atopic dermatitis in children [16]. In addition, a similar prebiotic mixture reduced the acute allergic skin response in mice orally sensitized with whey [17]. Besides scGOS and lcFOS, the pectin derived acidic-oligosaccharides (pAOS) were identified as a potential prebiotic fiber [18].

122
7
Oligosaccharide induced whey-specific CD25+ regulatory T-cells are crucial in the suppression of cow’s milk allergy in mice
In the present study a mouse model for orally induced sensitization to whey proteins was used to study the immunological mechanisms that might underlie the induction of tolerance against whey using prebiotics [19]. The role of Treg in the preventive capacities of dietary intervention with a prebiotic mixture (2 w/w% (9:1:1) scGOS, lcFOS and pAOS) was assessed using adoptive transfer experiments. It was demonstrated that CD25+ regulatory T-cells play a pivotal role in the prebiotic dietary induced inhibition of cow’s milk allergy in mice.
Materials and methods
Chemicals
Whey proteins were obtained from DMV International (Veghel, The Netherlands). Cholera toxin was purchased from Quadratech Diagnostics (Epsom, UK). PBS from Cambrex Bio Science (Verviers, Belgium). Biotin-labeled rat anti-mouse IgE, IgG1 and IgG2a from BD Biosciences (Alphen aan den Rijn, The Netherlands). PC61 (anti-CD25), a monoclonal antibody from a rat PC61 hybridoma cell line, was a kindly gift of Bioceros BV (Utrecht, The Netherlands). All other chemicals were obtained from Sigma-Aldrich-Chemie (Zwijndrecht, The Netherlands).
Diets
Semi-purified cow’s milk protein free AIN-93G-based diets were composed and mixed with prebiotics by Research Diet Services (Wijk bij Duurstede, The Netherlands). The prebiotic diet contained 2 w/w% (9:1:1) short-chain GOS (scGOS; enzymatic elongation of lactose with galactose by β-galactosidase), long-chain FOS (lcFOS; from chicory inulin) and pAOS (produced from pectin). scGOS (Vivinal GOS, FrieslandCampina Domo, Zwolle, The Netherlands) contains oligosaccharides with a degree of polymerisation (dp) of 3–8. lcFOS (Raftiline HP, Orafti, Wijchen, The Netherlands) contains oligosaccharides with an average degree of polymerisation of >23. The scGOS/lcFOS spray-dried powder consists of approximately 50% scGOS and lcFOS, 19% maltodextrin (glucidex 2, Roquette, France), 16% lactose, 14% glucose and 1% galactose [15]. The pAOS powder (Südzucker AG, Mannheim, Germany) have a degree of polymerisation of 1-20, consisting of approximately 75% galacturonic acid oligomers, 10% monomers and 15% of moisture and ash [18]. All prebiotics were exchanged for the same amount of total carbohydrates, this resulted in a comparable overall carbohydrate composition in the diets.

123
7
Chapter 7
Oral sensitization and challenge of mice
Three-week-old specific pathogen free female C3H/HeOuJ mice were purchased from Charles River Laboratories (Maastricht, The Netherlands) and housed in the animal facility. Animal care and use were performed in accordance with the guidelines of the Dutch Committee of Animal Experiments. Two weeks before the first sensitization animals received the different diets; mice remained on these diets during the whole study protocol. Mice were sensitized intra gastric (i.g.) with 0.5 mL homogenized whey (40 mg/mL PBS) with cholera toxin (CT, 20 μg/mL PBS) as an adjuvant, using a blunt needle. Control mice received CT alone. Mice were boosted weekly for a period of 5 weeks as described previously [19]. One week after the last sensitization feces were collected and mice were challenged intra dermally (i.d) in the ear to determine the acute allergic skin response. After 24 hours blood samples were collected in Minicollect® Z Serum Sep vials (Greiner bio-one) and mice were sacrificed by cervical dislocation. Collected blood was centrifuged 15 min at 13500 rpm and sera were stored at –70oC.
Acute allergen specific skin reaction
The acute allergen specific skin response was measured after i.d. injection with 20 μL homogenized whey protein (0.5 mg/mL in PBS) in the ear pinnae. Ear thickness was measured in duplicate using a digital micrometer (Mitutoyo, Veenendaal, The Netherlands), at t=0 and 1 hour after challenge. The ear swelling (μm) was calculated by subtracting the basal (t=0 hour) from the specific whey induced ear swelling after 1 hour ear thickness and additionally (Δ ear swelling) subtracting the mean ear swelling from the group of sham-sensitized mice fed the corresponding diet.
Adoptive transfer experiments
To test whether splenocytes are able to transfer prebiotic induced tolerance adoptive transfer experiments were carried out. After the prebiotic supplemented (donor) mice were sacrificed, the spleens were obtained and pooled per group (n=12). Single cell suspensions of the four groups (sham, sham prebiotic, whey and whey prebiotic) were i.v. injected (100 μL, 1x107 cells/mL) in naïve mice, in total eight recipient groups of n=6/group were formed (Table 1 experiment A). The recipient mice were fed the control diet and sensitized (sham or whey) as described above. The acute allergic skin reaction was measured and whey-specific serum immunoglobulins were measured to determine allergic sensitization.

124
7
Oligosaccharide induced whey-specific CD25+ regulatory T-cells are crucial in the suppression of cow’s milk allergy in mice
In another set of experiments single spleen cell suspensions of whey-sensitized donor mice fed the control or prebiotic diet were pooled per group (n=16) and half of the cells were treated with 50 μg per spleen rat anti-CD25 (PC61). Two mL Biomag Goat anti-rat IgG (Bangs Laboratories Inc., Fishers, USA) was added to the treated cells and placed on a rotator for at 45 min, the bead-cell mixture was placed in a magnetic separator for 5 min and supernatant was transferred and again placed on the magnetic separator for 5 min. Depletion of CD25 positive cells was analyzed using flow cytometric analysis. The splenocytes of the four groups were i.v. injected (100 μL, 1x107 cells/mL) in recipient mice, in total eight groups were formed (n=6/group) (Table 1 experiment B). The recipient mice were sham- or whey-sensitized as described above. The acute allergic skin reaction was measured and whey-specific serum immunoglobulins were measured to determine allergic sensitization.
Flow cytometry
Single cell suspensions were prepared from mouse spleens. Cells were counted using a Coulter Z1 particle counter (Beckman Coulter) and diluted to a concentration of 2x106 cells/mL. For identification of CD4+CD25+Foxp3+ cells, anti-CD4-FITC (1:100) and anti-CD25-PE (1:100) were used (BD, Alphen aan den Rijn, The Netherlands). Foxp3 staining was performed following the manufacturer’s protocol (eBioscience, Breda, The Netherlands). Flow cytometry analysis was performed using FACSCalibur (BD, Alphen aan den Rijn, The Netherlands), CellQuest Pro Analysis software (BD, Alphen aan de Rijn, The Netherlands) and isotype controls were subtracted.
Measurement of specific serum immunoglobulins
Concentrations of whey-specific IgE, IgG1, IgG2a were determined in serum by means of ELISA as described before [19]. In short, plates were coated with 100 µL whey (20 mg/L. Plates were incubated with biotin-labeled rat anti-mouse IgE, IgG1 and IgG2a (1 mg/L), and incubated with streptavidin-horseradish peroxidase (HRP). Results were expressed as arbitrary units (AU), calculations were based on titration curves of pooled sera from whey and alum-i.p. immunized mice as a positive reference serum.
Statistics
All data were analyzed using the one way ANOVA and post hoc Bonferroni’s Comparison

125
7
Chapter 7
Experiment Sensitization of donors
Diet Treatment of splenocytes
Sensitization of recipients
Diet
A sham control - sham control
control - whey control
prebiotic - sham control
prebiotic - whey control
whey control - sham control
control - whey control
prebiotic - sham control
prebiotic - whey control
B whey control - sham control
control anti-CD25 sham control
control - whey control
control anti-CD25 whey control
prebiotic - sham control
prebiotic anti-CD25 sham control
prebiotic - whey control
prebiotic anti-CD25 whey control
Table 1. Overview of experimental set-up.
Test or Mann-Whitney U test. For the immunoglobulin levels an ANOVA was performed on log transformed data. Statistical analyses were conducted using GraphPad Prism software (version 4.03). P-values of * p<0.05, ** p<0.01 and *** p<0.001 were indicated when analyses reached statistically significance. Data are represented as means ± SEM.
Results
Acute allergic skin reaction of donor mice
To investigate the preventive effects of the prebiotic diet on sensitization, whey-sensitized mice and sham-sensitized mice were challenged i.d. in the ear pinnae with whey, one week after the last sensitization. The delta ear swelling in the whey-sensitized mice was 157 ± 11 μm, the prebiotic diet largely prevented the acute allergic skin reaction of the whey-sensitized mice (67 ± 5.8 μm, p<0.001, Figure 1). The diet did not affect ear swelling of sham-sensitized mice (data not shown).
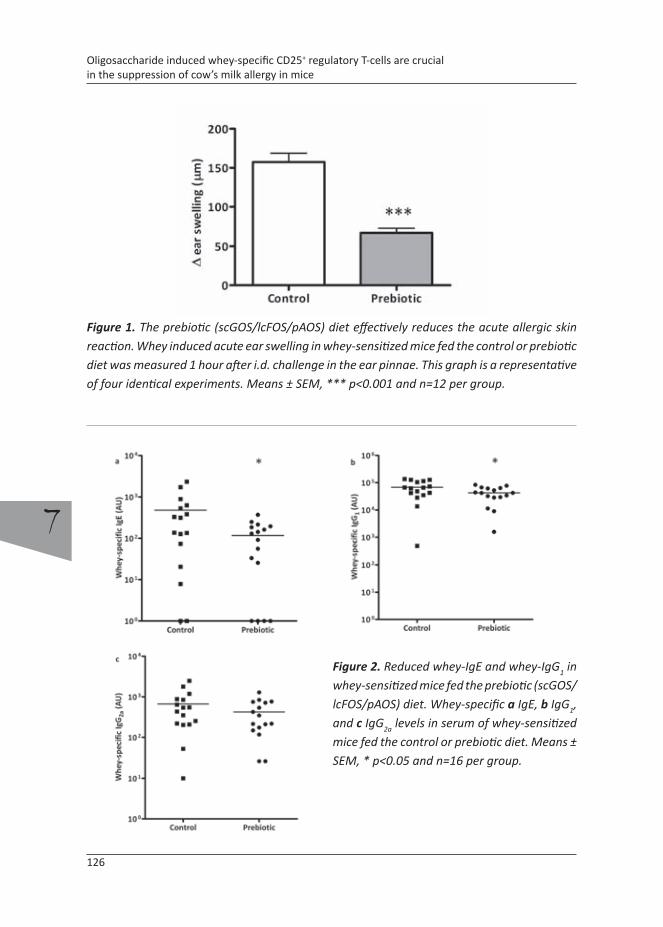
126
7
Oligosaccharide induced whey-specific CD25+ regulatory T-cells are crucial in the suppression of cow’s milk allergy in mice
Figure 1. The prebiotic (scGOS/lcFOS/pAOS) diet effectively reduces the acute allergic skin reaction. Whey induced acute ear swelling in whey-sensitized mice fed the control or prebiotic diet was measured 1 hour after i.d. challenge in the ear pinnae. This graph is a representative of four identical experiments. Means ± SEM, *** p<0.001 and n=12 per group.
Figure 2. Reduced whey-IgE and whey-IgG1 in whey-sensitized mice fed the prebiotic (scGOS/lcFOS/pAOS) diet. Whey-specific a IgE, b IgG1, and c IgG2a levels in serum of whey-sensitized mice fed the control or prebiotic diet. Means ± SEM, * p<0.05 and n=16 per group.

127
7
Chapter 7
Whey-specific immunoglobulin levels of donors
Whey-specific serum IgE, IgG1 and IgG2a levels were increased in whey-sensitized mice compared to sham-sensitized mice (data not shown). Whey-specific IgE was reduced in whey sensitized mice fed the prebiotic diet compared to mice fed the control diet (116 ± 27 vs 477 ± 167 AU, p<0.05, Figure 2). Also whey-specific IgG1 was reduced by the prebiotic diet (68120 ± 10650 vs 42560 ± 6045 AU, p<0.05). Dietary intervention did not affect whey-specific IgG2a levels in whey-sensitized mice fed control diet compared to prebiotic diet (665 ± 167 vs 425 ± 89 AU).
Acute allergic skin reactions in recipient mice adoptively transferred with splenocytes
Sham- as well as whey-sensitized recipient mice receiving splenocytes (i.v.) of whey-sensitized
Figure 3. Adoptive transfer of splenocytes of whey-sensitized mice fed the prebiotic (scGOS/lcFOS/pAOS) diet protects recipient mice from developing an acute allergic skin response. Recipient mice receiving splenocytes from whey-sensitized mice fed the control diet developed an acute skin response if sham- or whey-sensitized. Transfer of splenocytes of whey-sensitized mice fed prebiotic diet prevented these responses. However the acute skin response still developed in whey-sensitized recipients transferred with prebiotic fed donor mice that were sham-sensitized. The delta ear swelling was calculated by subtracting ear swelling of sham-sensitized recipient mice injected with cells of sham-sensitized donor mice fed corresponding control or prebiotic diet. Means ± SEM, * p<0.05, *** p<0.001 and n=6 per group.

128
7
Oligosaccharide induced whey-specific CD25+ regulatory T-cells are crucial in the suppression of cow’s milk allergy in mice
donor mice fed the control diet had severe allergic skin reactions in the ears 1 h after whey injection (Figure 3). In contrast, the transfer of donor splenocytes of whey-sensitized mice fed the prebiotic diet protected against development of an acute allergic skin reaction in sham-sensitized as well as whey-sensitized recipient mice (12 ± 5 and 22 ± 8 µm, p<0.001 compared to recipients transferred with control diet fed donor splenocytes). This protection was whey-specific since whey-sensitized recipient mice injected with splenocytes of sham-sensitized donors receiving the prebiotic diet still developed a severe ear swelling response (125 ± 12 µm).
Whey-specific immunoglobulin levels of recipients
Serum samples were collected of the recipient mice and analyzed for whey-specific immunoglobulins. Whey-specific IgE levels of recipient mice receiving splenocytes of sham-sensitized donor mice fed the control diet were enhanced upon whey-sensitization compared to sham-sensitization (352 ± 243 vs 6.0 ± 3.0 AU, p<0.01, Figure 4a). Similar results were obtained when recipients were transferred with splenocytes from sham-sensitized donor mice fed the prebiotic diet (256 ± 112 vs 5.2 ± 2.4 AU, p<0.01). Whey-specific IgE levels tended to increase in sham-sensitized recipient mice adoptively transferred with splenocytes of whey-sensitized mice. In contrast, transfer of splenocytes of whey-sensitized mice lowered the capacity to develop whey-specific IgE in recipients upon sensitization with whey. However, no difference in IgE levels were observed between recipients transferred with donor cells of mice fed the control or prebiotic diet.
Similar results were found for IgG1 and for IgG2a (Figure 4b and 4c). Whey-sensitized
recipient mice transferred with cells of sham-sensitized donor mice, regardless of the donor diet had enhanced IgG1 and IgG2a levels compared to sham-sensitized recipients transferred with splenocytes of sham-sensitized donors. Sham-sensitized recipients transferred with splenocytes of whey-sensitized mice fed either the control or prebiotic diet, had the lowest IgG1 and IgG2a levels. The IgG1 and IgG2a levels of recipient whey-sensitized mice transferred with splenocytes of whey-sensitized mice, regardless of the received diet, were of intermediate levels.
Overall, no differences in sensitization of the recipient mice transferred with cells of donor mice receiving control or prebiotic diet were observed regarding the antibody levels.

129
7
Chapter 7
Figure 4. Whey-specific immunoglobulin levels in recipient mice of which the donor mice were fed control or prebiotic (scGOS/lcFOS/pAOS) diet. No differences were observed between diet groups. a Whey-specific IgE b whey-specific IgG1 and c whey-specific IgG2a. Means ± SEM, * p<0.05, ** p<0.01, *** p<0.001 and n=5-6 per group.

130
7
Oligosaccharide induced whey-specific CD25+ regulatory T-cells are crucial in the suppression of cow’s milk allergy in mice
Figure 5. Ex vivo depletion of CD25+ Treg in spleen cell suspension of donor mice using PC61 antibody and magnetic beads. a Percentage of CD4+CD25+Foxp3+ Tregs. b Representative dot plots of CD25+Foxp3+ cells in the CD4+ population. These spleen cell fractions were used for adoptive transfer experiments. Means ± SEM, ** p<0.01, n=3-6.
CD25
Foxp3
rat IgG1 anti‐CD25
6.5% 3.6%
b
0.0%
isotype
7.9% 3.9%
preb
iotic
control

131
7
Chapter 7
Figure 6. Depletion of Treg in transferred spleen cell suspensions of whey-sensitized donors fed the prebiotic (scGOS/lcFOS/pAOS) diet abolished the protection against the development of an acute allergic skin reaction. Donor cells of whey-sensitized mice fed the control diet did not protect against the development of the acute allergic skin response in recipient mice and Treg depletion did not alter this response. Means ± SEM, ns = not significant, * p<0.05, ** p<0.01, *** p<0.001 and n=5-6 per group.
Acute allergic skin reaction of recipients transferred with Treg depleted donor cells
Splenocytes from whey-sensitized mice fed the control or prebiotic diet were transferred with or without prior ex vivo CD25+ cell depletion using the PC61 antibody. There was no difference in number of CD4+CD25+Foxp3+ Treg in the spleen of whey-sensitized mice fed the prebiotic diet (6.1 ± 0.3%, Figure 5) compared to the control diet (6.4 ± 0.3%). In both groups the ex vivo Treg depletion was similar, resulting in a 67% reduction in the prebiotic and 61% reduction in the control diet donor spleen suspension compared to the non-depleted controls (p<0.01).
Acute allergic skin reaction of recipients using Treg depleted donor cells
To investigate the role of Treg in the transfer of dietary prebiotics induced tolerance, splenocytes of whey-sensitized mice fed the control or prebiotic diet were ex vivo depleted for Treg cells and transferred. The acute allergic skin reaction in recipient mice was determined. Upon whey-sensitization the ear swelling response of recipient mice transferred with complete or Treg depleted spleen cell suspensions from whey-sensitized

132
7
Oligosaccharide induced whey-specific CD25+ regulatory T-cells are crucial in the suppression of cow’s milk allergy in mice
Figure 7. Whey-specific immunoglobulin levels in serum of recipient mice after adoptive transfer with sham or Treg depleted splenocytes. There are no differences between control and prebiotic (scGOS/lcFOS/pAOS) fed groups. a whey-specific IgE b whey-specific IgG1 and c whey-specific IgG2a. Means ± SEM, ns = not significant, * p<0.05, ** p<0.01 and n=6 per group.

133
7
Chapter 7
donor mice fed the control diet significantly enhanced compared to the sham-sensitized recipient mice (149 ± 8.1 or 179 ± 22 µm vs 78 ± 10 µm, p<0.01, Figure 6). Donor cells from whey-sensitized mice fed the prebiotic diet effectively prevented the occurrence of an acute skin response in recipients upon whey-sensitization compared to sham-sensitization and the ear swelling response was significantly lower compared to recipients transferred with cells of mice fed control diet (149 ± 8.1 vs 90 ± 15 µm; p<0.05). Treg depletion prior to transfer completely abolished the preventive effect of adoptively transferred donor cells of mice fed the prebiotic diet resulting in severe ear swelling in recipient mice upon whey-sensitization (224 ± 11 µm, p<0.001).
Whey-specific immunoglobulin levels of recipients of Treg depleted donor cells
Sham- or whey-sensitized recipient mice receiving spleen cell suspensions of control or prebiotic diet fed whey-sensitized donor mice showed similar whey-specific IgE levels. Recipient mice receiving Treg depleted spleen cell suspensions of whey-sensitized donor mice fed the control diet had higher whey-specific IgE levels in comparison with recipient mice receiving non-Treg depleted spleen cell suspensions (12.5 ± 3.6 vs 169 ± 71 AU, p<0.05, Figure 7a). A similar tendency was found in recipient mice receiving Treg depleted splenocytes of donor mice fed the prebiotic diet (32.5 ± 9.8 vs 170 ± 79 AU).
Also whey-specific IgG1 or IgG2a levels enhanced when recipient mice were transferred
with Treg depleted donor spleen cell suspensions, however no differences were observed between recipient mice transferred with donor cells of mice fed the control or prebiotic diet either in the presence or absence of Treg (Figure 7b and 7c).
Discussion
Genetic predisposition, environmental factors, timing, duration and extent of exposure are important determinants of allergic sensitization [20]. So far, no effective treatment for allergy is available, although murine and human studies have investigated the effectiveness of prebiotics to prevent the development of allergic disease. Dietary supplementation of oligosaccharides in babies and young infants has been found to reduce the incidence of atopic dermatitis [16] and infections in infants at risk [21]. Recently, we have shown a preventive effect of dietary prebiotics on the allergic effector response in an oral mouse model for whey allergy [22]. In the current study this model was used to investigate the

134
7
Oligosaccharide induced whey-specific CD25+ regulatory T-cells are crucial in the suppression of cow’s milk allergy in mice
contribution of CD25+ regulatory T-cells (Treg) in the development of tolerance to whey induced by the prebiotic diet using adoptive transfer experiments and ex vivo depletion of CD25+ Treg.
The prebiotic mixture of scGOS/lcFOS/pAOS used in this murine model for cow’s milk allergy ameliorated the severity of allergic symptoms, since the acute allergic skin response in mice fed the prebiotic diet during whey-sensitization was strongly reduced. A slight reduction in TH2 immunoglobulin levels (whey-specific IgE and IgG1) was observed, whereas TH1 type immunoglobulin IgG2a levels remained unaffected high. Previously, these prebiotics have been shown to improve the vaccination response in mice, which is mainly TH1 driven [18]. The results suggest an improved TH1/TH2 balance upon dietary intervention with prebiotics which could partly explain their effect, but not conclusively.
In mice orally sensitized with whey the acute allergic skin response determines the level of systemic response as a result of a break in oral tolerance [19]. This allergen-specific ear swelling response has similarities with the human skin prick test. Since the prebiotic diet was found to reduce the severity of the acute allergic skin response, adoptive transfer studies were performed to determine whether this tolerance could be transferred using splenocytes. Adoptive transfer of splenocytes from whey-sensitized donor mice fed the control diet transferred the allergy to recipient mice that were sham-sensitized, since they developed a whey-specific acute skin response. Guttormsen et al. [23] also found adoptively transferred whole spleen single cells to transfer immunologic memory for a vaccine. In contrast to the whey-sensitized recipient mice receiving splenocytes of whey-sensitized mice fed the control diet, the splenocytes of prebiotic fed whey-sensitized mice protected against the development of allergic symptoms. The whey-specific acute allergic skin reaction was prevented, hence tolerance for whey induced in the donor mice by the prebiotic diet could be transferred to the recipient mice.
We hypothesized the CD4+CD25+ Treg, which occurs naturally or can be induced, to be a potential cellular candidate mediating the prebiotic induced oral tolerance to whey. Two phases of oral tolerance can be distinguished, clonal deletion or active suppression of the immune response by Treg cells [24]. The prebiotic diet did not enhance the number of Treg cells in Peyer’s patches, mesenteric lymph nodes and spleen when mice were fed the prebiotic diet (data not shown). However, in vivo anti-CD25 depletion revealed that Treg cells play an important role in the oral sensitization with whey in mice (Van Esch et al., submitted). Furthermore Gri et al.[25] showed that Treg cells are able to inhibit mast cell degranulation and acute allergic responses which may relate to the reduced acute allergic skin reaction observed in the present study in mice fed the prebiotic diet.
The involvement of Treg in the induction of tolerance by prebiotics was studied using

135
7
Chapter 7
adoptive transfer of splenocytes of whey-sensitized mice fed the prebiotic diet that were ex vivo depleted for CD25+ Treg cells using an anti-CD25 antibody (PC61). Indeed, Treg depletion prior to transfer of these donor cells abrogated the transfer of tolerance which protected the recipient mice from developing a severe allergic skin reaction after whey-sensitization. It has been shown that the anti-CD25 mAb selectively depletes the CD25high cells which are CD4+ [26]. Indeed, in this study we confirmed the ex vivo depletion of CD4+CD25+Foxp3+ cells to be more than 60%. Also in other studies it was shown that complete depletion is not necessary to be effective and can be used to investigate Treg function [27-30]. Our data implicate the CD25+ Treg cells to be involved in the adoptive transfer of the tolerizing effect of dietary prebiotics. The transfer of tolerance observed in the recipient mice was whey-specific since transfer of cells of non-sensitized donor mice receiving the prebiotic diet did not protect recipient mice. This suggests involvement of an antigen-specific Treg cell population and there is growing evidence of a subset of Treg cells that have antigen-specific suppression characteristics [31, 32].
To determine allergic sensitization, usually plasma levels of allergen-specific IgE are measured. However, we and others found that this parameter is not conclusive for the generation of allergic symptoms [33]. In contrast to the acute allergic skin reaction in whey-sensitized recipient mice no differences in immunoglobulin levels were observed after adoptive transfer with cells of control or prebiotic fed mice. In a mouse ovalbumin model, transfer of Treg cells reduced the allergic response and reduced numbers of eosinophils in the lungs. Furthermore, airway remodeling was prevented. However, also in this study IgE levels remained unaffected high [34]. Given the extensive long half-life of the measured antibodies, the actual experimental period might be too short to identify potential difference on the long term in these levels caused by the intervention. However, the acute allergic skin response reflects clinical symptoms which occur upon degranulation of allergic effector cells. It can be concluded that the reduced skin response cannot be explained by differences in antibody titers between recipients transferred with cells of prebiotic or control diet fed mice.
In the present study it is shown that whey-sensitization is transferable using adoptive transfer of single spleen cell suspensions while also the protective effect of a prebiotic mixture on allergic symptoms could be transferred. The prebiotic diet provided only a minor reduction in whey-specific Igs and the reduced acute allergic skin response of the recipient mice transferred with donor cells of mice fed the prebiotic diet could not be explained by a reduction in immunoglobulin levels. Ex vivo Treg cell depletion abrogated the protective effects of the adoptive transfer with donor cells of whey-sensitized mice fed the prebiotic diet. The results suggest the involvement of whey-specific Treg cells in the transfer of tolerance.

136
7
Oligosaccharide induced whey-specific CD25+ regulatory T-cells are crucial in the suppression of cow’s milk allergy in mice
References
1. Friedman, A. and H.L. Weiner, Induction of anergy or active suppression following oral
tolerance is determined by antigen dosage. Proc Natl Acad Sci U S A, 1994. 91(14): p. 6688-
92.
2. Lundin, B.S., et al., Oral tolerization leads to active suppression and bystander tolerance in
adult rats while anergy dominates in young rats. Scand J Immunol, 1996. 43(1): p. 56-63.
3. Burks, A.W., S. Laubach, and S.M. Jones, Oral tolerance, food allergy, and immunotherapy:
implications for future treatment. J Allergy Clin Immunol, 2008. 121(6): p. 1344-50.
4. Hol, J., et al., The acquisition of tolerance toward cow’s milk through probiotic
supplementation: a randomized, controlled trial. J Allergy Clin Immunol, 2008. 121(6): p.
1448-54.
5. Crittenden, R.G. and L.E. Bennett, Cow’s milk allergy: a complex disorder. J Am Coll Nutr,
2005. 24(6 Suppl): p. 582S-91S.
6. Monaci, L., et al., Milk allergens, their characteristics and their detection in food: a review.
European Food Research and Technology, 2005(Volume 223, Number 2 / June, 2006): p.
149-179.
7. van Wijk, F., et al., CD4+CD25+ T cells regulate the intensity of hypersensitivity responses
to peanut, but are not decisive in the induction of oral sensitization. Clin Exp Allergy, 2007.
37(4): p. 572-81.
8. Akbari, O. and D.T. Umetsu, Role of regulatory dendritic cells in allergy and asthma. Curr
Opin Allergy Clin Immunol, 2004. 4(6): p. 533-8.
9. O’Garra, A. and P. Vieira, Regulatory T cells and mechanisms of immune system control. Nat
Med, 2004. 10(8): p. 801-5.
10. Kashyap, M., et al., Cutting edge: CD4 T cell-mast cell interactions alter IgE receptor
expression and signaling. J Immunol, 2008. 180(4): p. 2039-43.
11. Izcue, A. and F. Powrie, Special regulatory T-cell review: Regulatory T cells and the intestinal
tract--patrolling the frontier. Immunology, 2008. 123(1): p. 6-10.
12. Field, C.J., The immunological components of human milk and their effect on immune
development in infants. J Nutr, 2005. 135(1): p. 1-4.
13. Vos, A.P., et al., Immune-modulatory effects and potential working mechanisms of orally
applied nondigestible carbohydrates. Crit Rev Immunol, 2007. 27(2): p. 97-140.
14. Gibson, G.R. and M.B. Roberfroid, Dietary modulation of the human colonic microbiota:
introducing the concept of prebiotics. J Nutr, 1995. 125(6): p. 1401-12.
15. Vos, A.P., et al., A specific prebiotic oligosaccharide mixture stimulates delayed-type
hypersensitivity in a murine influenza vaccination model. Int Immunopharmacol, 2006. 6(8):
p. 1277-86.
16. Moro, G., et al., A mixture of prebiotic oligosaccharides reduces the incidence of atopic
dermatitis during the first six months of age. Arch Dis Child, 2006. 91(10): p. 814-9.

137
7
Chapter 7
17. Schouten, B., et al., Cow milk allergy symptoms are reduced in mice fed dietary synbiotics
during oral sensitization with whey. J Nutr, 2009. 139(7): p. 1398-403.
18. Vos, A.P., et al., Dietary supplementation of neutral and acidic oligosaccharides enhances
Th1-dependent vaccination responses in mice. Pediatr Allergy Immunol, 2007. 18(4): p. 304-
12.
19. Schouten, B., et al., Acute Allergic Skin Reactions and Intestinal Contractility Changes in Mice
Orally Sensitized against Casein or Whey. Int Arch Allergy Immunol, 2008. 147(2): p. 125-
134.
20. Kimber, I. and R.J. Dearman, Factors affecting the development of food allergy. Proc Nutr
Soc, 2002. 61(4): p. 435-9.
21. Arslanoglu, S., et al., Early dietary intervention with a mixture of prebiotic oligosaccharides
reduces the incidence of allergic manifestations and infections during the first two years of
life. J Nutr, 2008. 138(6): p. 1091-5.
22. Schouten, B., et al., Cow Milk Allergy Symptoms Are Reduced in Mice Fed Dietary Synbiotics
during Oral Sensitization with Whey. J Nutr, 2009.
23. Guttormsen, H.K., et al., Immunologic memory induced by a glycoconjugate vaccine in a
murine adoptive lymphocyte transfer model. Infect Immun, 1998. 66(5): p. 2026-32.
24. Tsuji, N.M. and A. Kosaka, Oral tolerance: intestinal homeostasis and antigen-specific
regulatory T cells. Trends Immunol, 2008. 29(11): p. 532-40.
25. Gri, G., et al., CD4+CD25+ regulatory T cells suppress mast cell degranulation and allergic
responses through OX40-OX40L interaction. Immunity, 2008. 29(5): p. 771-81.
26. Amu, S., et al., B-cell CD25 expression in murine primary and secondary lymphoid tissue.
Scand J Immunol, 2006. 64(5): p. 482-92.
27. Shreffler, W.G., et al., Association of allergen-specific regulatory T cells with the onset of
clinical tolerance to milk protein. J Allergy Clin Immunol, 2009. 123(1): p. 43-52 e7.
28. Kohm, A.P., et al., Cutting Edge: Anti-CD25 monoclonal antibody injection results in the
functional inactivation, not depletion, of CD4+CD25+ T regulatory cells. J Immunol, 2006.
176(6): p. 3301-5.
29. Lewkowich, I.P., et al., CD4+CD25+ T cells protect against experimentally induced asthma
and alter pulmonary dendritic cell phenotype and function. J Exp Med, 2005. 202(11): p.
1549-61.
30. Matsushita, N., et al., Comparative methodologies of regulatory T cell depletion in a murine
melanoma model. J Immunol Methods, 2008. 333(1-2): p. 167-79.
31. Zhang, Z.X., et al., Identification of a previously unknown antigen-specific regulatory T cell
and its mechanism of suppression. Nat Med, 2000. 6(7): p. 782-9.
32. Koenen, H.J. and I. Joosten, Antigen-specific regulatory T-cell subsets in transplantation
tolerance regulatory T-cell subset quality reduces the need for quantity. Hum Immunol,
2006. 67(9): p. 665-75.
33. van der Gugten, A.C., et al., Usefulness of specific IgE levels in predicting cow’s milk allergy.
J Allergy Clin Immunol, 2008. 121(2): p. 531-3.

138
7
Oligosaccharide induced whey-specific CD25+ regulatory T-cells are crucial in the suppression of cow’s milk allergy in mice
34. Kearley, J., D.S. Robinson, and C.M. Lloyd, CD4+CD25+ regulatory T cells reverse established
allergic airway inflammation and prevent airway remodeling. J Allergy Clin Immunol, 2008.
122(3): p. 617-24 e6.

139
7
Chapter 7


8
Dietary non-digestible carbohydrates inhibit the allergic effector response via induction of
casein-specific CD25+ regulatory T-cells in orally sensitized mice
Bastiaan Schouten1
Betty C.A.M. van Esch1,2
Gerard A. Hofman1
Sander de Kivit1
Louis Boon3
Léon M.J. Knippels2
Johan Garssen1,2
Linette E.M. Willemsen1
1 Department of Pharmacology and Pathophysiology, Utrecht Institute for Pharmaceutical
Sciences, Faculty of Science, Utrecht University, Utrecht, The Netherlands2 Danone Research – Centre for Specialised Nutrition, Wageningen, The Netherlands
3 Bioceros BV, Utrecht, The Netherlands
Submitted for publication

142
8
Dietary non-digestible carbohydrates inhibit the allergic effector response via induction of casein-specific CD25+ regulatory T-cells in orally sensitized mice
Abstract
Background: Dietary prebiotics have been shown to prevent the development of orally induced cow’s milk allergy in mice. In this study the contribution of CD25+ regulatory T-cells in casein allergy prevention by prebiotics was investigated using in vivo CD25+ depletion and adoptive transfer studies. Methods: Mice were sensitized with casein and fed a prebiotic diet containing 2% short-chain galacto-, long-chain fructo-, and acidic-oligosaccharides or a control diet. Anti-CD25 (PC61) or isotype control antibody was injected intraperitoneally two times during the dietary intervention. In adoptive transfer experiments, splenocytes of donor mice sensitized with casein and fed the prebiotic or control diet were transferred to naïve recipients. CD25+ cells were ex vivo depleted using PC61. Recipient mice were casein- or sham-sensitized and received the control diet. The acute allergic skin reaction upon intra dermal casein challenge was determined and casein-specific serum immunoglobulins were measured. Furthermore, T
H1 and TH2 cell numbers were analyzed in the mesenteric lymph nodes (MLN) using flow cytometry.Results: The prebiotic diet strongly reduced development of the casein-specific acute allergic skin reaction (p<0.001) and in vivo anti-CD25 treatment abrogated this. The diet was found to enhance the percentage of T
H1 and tended to reduce the percentage of TH2 cells in MLN of casein-sensitized mice. In casein-sensitized recipient mice, transferred with splenocytes from casein-sensitized prebiotic fed donor mice, development of the acute allergic skin reaction was prevented (p<0.001), while cells from sham-sensitized prebiotic fed donor mice were not able to down regulate the allergic response in recipients. This highlights the antigen specificity of the anti-allergic effects of the prebiotic diet. Ex vivo depletion of CD25+ regulatory T-cells prevented this transfer of tolerance without affecting casein specific immunoglobulin levels. Conclusion: CD25+ regulatory T-cells play a pivotal role in the anti-allergic effects induced by a unique mixture of non-digestible dietary carbohydrates.

143
8
Chapter 8
Introduction
Immune modulation by dietary intervention is a growing field in immunological sciences. In particular the effect of non-digestible carbohydrates is currently under investigation by many research groups. Non-digestible carbohydrates such as short-chain galacto oligosaccharides (scGOS) and long-chain fructo oligosaccharides (lcFOS) are added to clinical nutrition and infant formulas since they may have health and immune promoting properties [1, 2]. Acidic oligosaccharides derived from pectin (pAOS) are recently developed non-digestible carbohydrates that mimic properties of human acidic oligosaccharides such as present in breast milk [3]. Recent research is focusing on the influence of prebiotic carbohydrates on the development of allergic disease, dietary intervention in most studies started immediately after birth. In a mouse model for orally induced cow’s milk allergy (CMA), a specific mixture of scGOS and lcFOS (Immunofortis®) has been shown to reduce the allergic effector response [4]. Furthermore, in a clinical study with infants at high risk for allergy the same carbohydrate mixture reduced the incidence of atopic dermatitis significantly [5]. Oligosaccharides may exert their immunomodulatory effects via direct interaction with the mucosal immune system and/or via adaptation of the intestinal microbiota, since oligosaccharides such as scGOS and lcFOS are selectively fermented by lactic acid producing Bifidobacteria and Lactobacilli in the gut [6-8].
One of the first manifestations of atopic disease is the development of CMA in young infants. There are two major allergenic fractions in cow’s milk: casein and whey. Casein, and in particular αS1-casein, is the main allergenic protein [9, 10]. Normally the immune system is generating local and systemic tolerance to harmless cow’s milk proteins, however in some infants who develop cow’s milk allergy this oral tolerance is not induced, resulting in an allergic response to these proteins. Oral tolerance induction is in majority generated within the gut associated lymphoid tissue in particular the Peyer’s patches (PP) and the mesenteric lymph nodes (MLN). From the MLN tolerance can be transferred to the systemic immune system. Especially regulatory T-cells play a crucial role in this process [11, 12]. Cells with regulatory properties include TGF-β-producing T
H3 cells, IL-10-producing Tr1 cells and iNKT cells [13-16]. However, CD4+CD25+Foxp3+ naturally occurring de novo generated regulatory T-cells (Treg) were described to be important for maintenance of peripheral tolerance or acquired tolerance for e.g food antigens, respectively [17-19]. Treg cells represent 5-10% of the CD4+ T lymphocyte population and express the transcription factor forkhead box p3 (Foxp3), which is confirmed as a key gene for generation and maintenance of Treg cell functionality. The contribution of CD25+ Treg cells in the induction of oral tolerance has been shown in children who have outgrown cow’s milk allergy, however also other mechanisms may be involved as well [20-22].

144
8
Dietary non-digestible carbohydrates inhibit the allergic effector response via induction of casein-specific CD25+ regulatory T-cells in orally sensitized mice
The underlying mechanism of action of allergy prevention by non-digestible carbohydrates is largely unknown. In the present study, mice were fed a 2% scGOS/lcFOS/pAOS prebiotic mixture during oral sensitization with casein which was found to reduce the allergic effector response. In this model the contribution of CD25+ Treg cells to this preventive effect of the diet was studied using in vivo anti-CD25 depletion and adoptive transfer of splenocytes ex vivo depleted for CD25+ Treg cells.
Materials and Methods
Chemicals
Casein proteins were obtained from DMV International (Veghel, The Netherlands). Cholera toxin was purchased from Quadratech Diagnostics (Epsom, UK). PBS from Cambrex Bio Science (Verviers, Belgium). Biotin-labeled rat anti-mouse IgE, IgG1 and IgG2a from BD Biosciences (Alphen aan den Rijn, The Netherlands). PC61 (anti-CD25), a rat monoclonal antibody PC61 hybridoma cell line, was a kindly gift of Bioceros BV (Utrecht, The Netherlands). All other chemicals were obtained from Sigma-Aldrich-Chemie (Zwijndrecht, The Netherlands).
Diets
Semi-purified cow’s milk protein free AIN-93G-based diets were composed and mixed with the oligosaccharides by Research Diet Services (Wijk bij Duurstede, The Netherlands). The scGOS/lcFOS/pAOS diet contained 2 w/w% (9:1:1) short-chain GOS (enzymatic elongation of lactose with galactose by β-galactosidase), long-chain FOS (from chicory inulin) and pAOS (produced from pectin). GOS (Vivinal GOS, FrieslandCampina Domo, Zwolle, The Netherlands) contains oligosaccharides with a degree of polymerisation (dp) of 3–8. FOS (Raftiline HP, Orafti, Wijchen, The Netherlands) contains oligosaccharides with an average dp of >23. The GOS/FOS spray-dried powder consists of approximately 50% GOS and FOS, 19% maltodextrin (glucidex 2, Roquette, France), 16% lactose, 14% glucose and 1% galactose [23]. The pAOS powder (Südzucker AG, Mannheim, Germany) have a dp of 1-20, consisting of approximately 75% galacturonic acid oligomers, 10% monomers and 15% of moisture and ash [24]. All oligosaccharides were exchanged for the same amount of total carbohydrates, which resulted in a comparable overall carbohydrate composition in the diets.

145
8
Chapter 8
Oral sensitization and challenge of mice
Three-week-old specific pathogen free female C3H/HeOuJ mice were purchased from Charles River Laboratories (Maastricht, The Netherlands) and housed in the animal facility. Animal care and use were performed in accordance with the guidelines of the Dutch Committee of Animal Experiments. Mice were fed the control or scGOS/lcFOS/pAOS diet during the whole study period, the diet was started two weeks before the first sensitization. Mice were sensitized intra gastric (i.g.) with 0.5 mL homogenized casein (40 mg/mL PBS) with cholera toxin (CT, 20 μg/mL PBS) as an adjuvant, using a blunt needle. Control mice received CT alone. Mice were boosted weekly for a period of 5 weeks as described previously [25]. One week after the last sensitization feces were collected and mice were challenged intra dermally (i.d) in the ear pinnae to determine the acute allergic skin response. After 24 hours blood samples were collected in Minicollect® Z Serum Sep vials (Greiner bio-one) and mice were sacrificed by cervical dislocation. Collected blood was centrifuged 15 min at 13,500 rpm and sera were stored at –70oC.
Acute allergen specific skin reaction
The acute allergen specific skin response was measured after i.d. injection with 20 μL homogenized casein protein (0.5 mg/mL in PBS) in the ear pinnae. Ear thickness was measured in duplicate using a digital micrometer (Mitutoyo, Veenendaal, The Netherlands), at t=0 and 1 hour after challenge. The ear swelling (μm) was calculated by subtracting the basal (t=0 hour) from the specific casein induced ear swelling after 1 hour ear thickness and additionally (Δ ear swelling) subtracting the mean ear swelling from the group of sham-sensitized mice fed the corresponding diet.
Anti-CD25 depletion studies
In the first set of experiments in vivo depletion of CD25+ regulatory T-cells was performed (Table 1). One day before the first sensitization, sham- and casein-sensitized mice fed the control or scGOS/lcFOS/pAOS diet were i.p. injected with 100 µL (2 mg/mL) rat anti-mouse CD25 mAb or rat IgG1 isotype control antibody as a control. A second CD25+ depletion was carried out one day before the third (day 14) sensitization.

146
8
Dietary non-digestible carbohydrates inhibit the allergic effector response via induction of casein-specific CD25+ regulatory T-cells in orally sensitized mice
Table 1. Overview of experimental set-up of adoptive transfer experiments.
Sensitization of donors
Diet Treatment of splenocytes
Sensitization of recipients
Diet
sham control - sham control
control - casein control
control anti-CD25 sham control
control anti-CD25 casein control
scGOS/lcFOS/pAOS - sham control
scGOS/lcFOS/pAOS - casein control
scGOS/lcFOS/pAOS anti-CD25 sham control
scGOS/lcFOS/pAOS anti-CD25 casein control
casein control - sham control
control - casein control
control anti-CD25 sham control
control anti-CD25 casein control
scGOS/lcFOS/pAOS - sham control
scGOS/lcFOS/pAOS - casein control
scGOS/lcFOS/pAOS anti-CD25 sham control
scGOS/lcFOS/pAOS anti-CD25 casein control
Adoptive transfer experiments
To test whether splenocytes are able to transfer scGOS/lcFOS/pAOS induced prevention of the allergic effector response, the contribution of CD25+ regulatory T-cells was studied in adoptive transfer experiments. After the scGOS/lcFOS/pAOS supplemented (donor) mice were sacrificed, the spleens were obtained and pooled per group (n=18). Single cell suspensions of the four groups (sham, sham scGOS/lcFOS/pAOS, casein and casein scGOS/lcFOS/pAOS) were pooled. Per group 2/3 part of the spleen cell suspension was treated with 600 μg anti-CD25 (PC61) mAb in 60 mL RPMI with 5% FCS for half an hour. After washing with RPMI with 5% FCS two mL Biomag Goat anti-rat IgG magnetic beads (Bangs Laboratories Inc., Fishers, USA) were added to the treated cells and placed on a rotator for at 45 min. The bead-cell mixture was placed in a magnetic separator (Bangs Laboratories Inc., Fishers, USA) for 5 min and supernatant was transferred and again placed on the magnetic separator for 5 min. Depletion of CD25 positive cells was analyzed using flow cytometric analysis. Cells were counted using the Coulter Z1 particle counter (Beckman Coulter) and the splenocytes of the donor mice ex vivo depleted for CD25+ cells or complete 1/3 part non-depleted suspensions were i.v. injected (100 μL, 1x107 cells/mL) in sixteen different groups of recipient mice, n=6/group (Table 1). The recipient mice were sham- or casein-sensitized as described above. The

147
8
Chapter 8
acute allergic skin reaction was measured and casein-specific serum immunoglobulins were measured to determine allergic sensitization.
Flow cytometry
Single cell suspensions were prepared from mouse spleens and blocked for 30 min in PBS containing 1% BSA and 5% FCS. In addition, for the isolation of MLN lymphocytes, MLN were treated with RPMI containing collagenase D and DNAse (Roche Diagnostics, Almere, The Netherlands) and incubated for 30 min at 37oC twice. Collagenase D and DNAse were inactivated using RPMI supplemented with 5% FCS. Cells were counted using a Coulter Z1 particle counter (Beckman Coulter) and diluted to a concentration of 2x106 cells/mL. For identification of different subsets of cells, cells were incubated for 30 min at 4oC with different Abs (all from BD, Alphen aan den Rijn, The Netherlands, unless otherwise stated). Anti-CD4-FITC (1:100), anti-CD4-PerCP-CY5.5 (1:50), anti-CD25-PE (1:100), anti-CD69-FITC (1:50), anti-CD69-PE (1:50), anti-CXCR3-PE (1:50) and anti-T1/ST2-FITC (1:50, MD Biosciences, St. Paul, Minnesota, USA) and isotype controls. Foxp3 staining was performed following the manufacturer’s protocol (eBioscience, Breda, The Netherlands). Flow cytometry analysis was performed using FACSCalibur (BD, Alphen aan den Rijn, The Netherlands) and CellQuest Pro Analysis software (BD, Alphen aan de Rijn, The Netherlands) and isotype controls were subtracted.
Measurement of specific serum immunoglobulins
Concentrations of casein-specific IgE, IgG1 and IgG2a were determined in serum by means of ELISA as described before [25]. In short, plates were coated with 100 µL casein (20 mg/L), washed and several serum dilutions were applied for 1 h. Plates were washed and incubated with biotin-labeled rat anti-mouse IgE, IgG1 and IgG2a (1 mg/L), washed and incubated with streptavidin-horseradish peroxidase (HRP). For detection A490 nm absorption was determined using spectrophotometer (Bio-Rad). Results were expressed as arbitrary units (AU), calculations were based on titration curves of pooled sera from casein and alum-i.p. immunized mice as a positive reference serum.

148
8
Dietary non-digestible carbohydrates inhibit the allergic effector response via induction of casein-specific CD25+ regulatory T-cells in orally sensitized mice
Statistics
All data were analyzed using the one way ANOVA and post hoc Bonferroni’s Comparison Test or Mann-Whitney U test. For the immunoglobulin levels the ANOVA was performed on log transformed data. Statistical analyses were conducted using GraphPad Prism software (version 4.03). P-values of *<0.05, ** p<0.01 and *** p<0.001 were indicated when analyses reached statistically significance. Data are represented as means ± SEM.
Results
Adoptive transfer experiments; donor mice
Two weeks prior to the first sensitization mice were fed control or scGOS/lcFOS/pAOS diet. One day before the first sensitization mice were i.v. injected with anti-CD25 or isotype control. To study CD25+ Treg cells involvement in the preventive effects of the scGOS/lcFOS/pAOS diet adoptive transfer experiments were performed. Initially, four groups of donor mice were casein- or sham-sensitized and fed the control diet or scGOS/lcFOS/pAOS (Table I) and the acute allergic skin reaction was determined (Figure 1). The scGOS/lcFOS/pAOS diet effectively reduced the acute allergic skin reaction of casein-sensitized donor mice when compared to the control diet (109 ± 4.3 vs 61 ± 5.0 µm, p<0.001). To explore underlying mechanistic pathways of scGOS/lcFOS/pAOS the T
H1 and TH2 cells in the MLN were analyzed using flow cytometry (Figure 2). The percentage of activated TH1 cells (CD4+CD69+CXCR3+) was enhanced in the scGOS/lcFOS/pAOS fed casein-sensitized mice when compared to control fed casein-sensitized mice (p<0.05, Figure 2a). The percentage of activated TH2 cells tended to be reduced (Figure 2b).
Acute allergic skin reaction
To study the contribution of CD25+ regulatory T-cells to the preventive effects of the scGOS/lcFOS/pAOS diet on the acute allergic skin reaction, sham- and casein-sensitized mice were injected (i.p.) with anti-CD25 antibody or isotype control. The delta ear swelling in the isotype control casein-sensitized mice was 71 ± 5.5 μm and the scGOS/lcFOS/pAOS diet largely prevented this (28 ± 6.0 μm, p<0.001, Figure 3). The diet did not affect ear swelling of sham-sensitized mice. The protective effect of the scGOS/lcFOS/pAOS diet was completely abrogated by anti-CD25 mAb treatment (76 ± 5.4 vs 78 ± 4.9 μm).
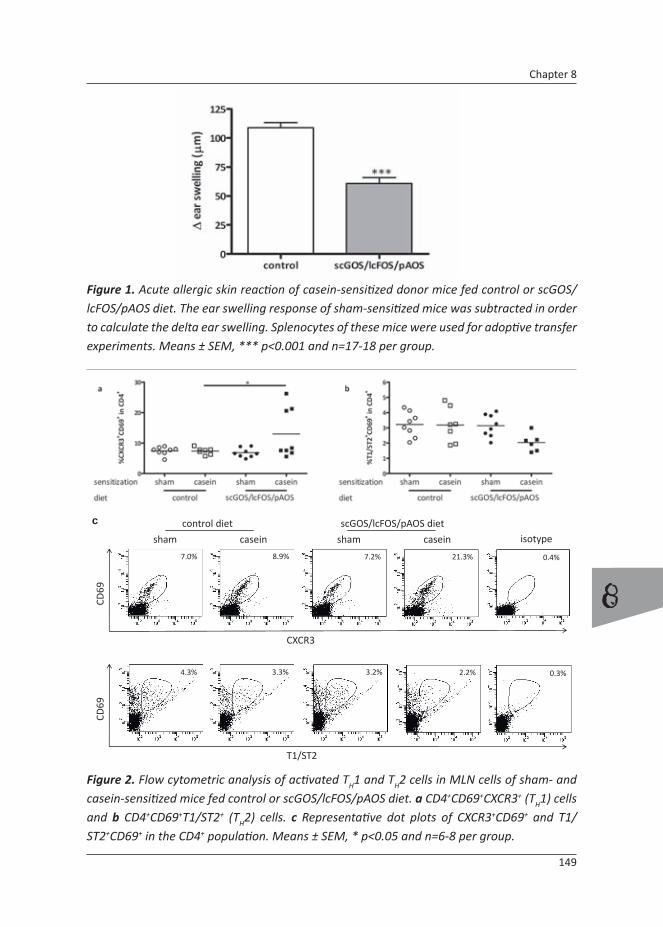
149
8
Chapter 8
Figure 1. Acute allergic skin reaction of casein-sensitized donor mice fed control or scGOS/lcFOS/pAOS diet. The ear swelling response of sham-sensitized mice was subtracted in order to calculate the delta ear swelling. Splenocytes of these mice were used for adoptive transfer experiments. Means ± SEM, *** p<0.001 and n=17-18 per group.
7.0%
CXCR3
CD69
4.3%
T1/ST2
CD69
control diet scGOS/lcFOS/pAOS diet
sham shamcasein casein
8.9% 7.2% 21.3%
2.2%3.3% 3.2%
c
0.4%
isotype
0.3%
Figure 2. Flow cytometric analysis of activated TH1 and TH2 cells in MLN cells of sham- and casein-sensitized mice fed control or scGOS/lcFOS/pAOS diet. a CD4+CD69+CXCR3+ (TH1) cells and b CD4+CD69+T1/ST2+ (TH2) cells. c Representative dot plots of CXCR3+CD69+ and T1/ST2+CD69+ in the CD4+ population. Means ± SEM, * p<0.05 and n=6-8 per group.

150
8
Dietary non-digestible carbohydrates inhibit the allergic effector response via induction of casein-specific CD25+ regulatory T-cells in orally sensitized mice
Figure 3. The scGOS/lcFOS/pAOS diet effectively reduced the acute allergic skin reaction which was abrogated by anti-CD25 mAb treatment. Casein induced acute ear swelling in casein-sensitized mice fed the control of scGOS/lcFOS/pAOS diet was measured 1 hour after i.d. challenge. Anti-CD25 or isotype control Abs were i.p. injected. This graph is a representative of multiple identical experiments. Means ± SEM, *** p<0.001 and n=6 per group.
CD25
Foxp3
rat IgG1 anti‐CD25
5.4% 2.9%
b
0.0%
isotype
Figure 4. a Confirmation of in vivo depletion of CD4+CD25+Foxp3+ regulatory T-cells in the spleen after the second anti-CD25 treatment. b Representative dot plots of CD25+Foxp3+ cells in the CD4+ population following isotype or anti-CD25 treatment. The percentage of CD4+CD25+Foxp3- cells was unaffected. Means ± SEM, ** p<0.01 and n=5-6 per group.

151
8
Chapter 8
Confirmation depletion of CD25+ cells using flow cytometry
Two groups of casein-sensitized mice were sacrificed at day 15, two days after the second anti-CD25 or isotype control treatment and one day after the third sensitization. The splenocytes were analyzed to confirm CD25+ Treg cell depletion. CD4+CD25+Foxp3+ regulatory T-cell percentages were declined by more than 65% (Figure 4, p<0.01). A difference was observed in the percentage of the CD4+ cells between isotype rat IgG1 and anti-CD25 treated groups (12.9 ± 1.0 vs 10.1 ± 0.3% of total lymphocytes, p<0.05, data not shown), anti-CD25 treatment did not reduce CD4+CD25+Foxp3- cells which includes the effector T-cell population (5.1 ± 0.4 vs 5.5 ± 0.5% of CD4+ cells; data not shown).
Casein-specific immunoglobulin levels of in vivo Treg cells depleted mice
Serum samples were collected and analyzed for casein-specific immunoglobulin levels. As described before [25] casein-specific IgE levels were not enhanced in comparison with sham-sensitized mice. However, after Treg cell depletion casein-specific IgE levels were enhanced (p<0.05, Figure 5a) in comparison with sham-sensitized mice. No differences were observed between the control diet and scGOS/lcFOS/pAOS diet with respect to casein-specific IgE levels in isotype or anti-CD25 treated mice.
Casein-specific IgG1 levels of mice fed the control diet were not enhanced due to two non-
responders. However, after Treg cell depletion casein-specific IgG1 levels were enhanced compared to sham-sensitized mice (p<0.01). Casein-sensitized mice fed scGOS/lcFOS/pAOS diet had low casein-specific IgG1 levels. However, this was not significant lowered as compared to mice fed the control diet. After Treg cell depletion casein-specific IgG1 levels were significantly enhanced as compared to both sham- as casein-sensitized mice fed the scGOS/lcFOS/pAOS diet (p<0.05, Figure 5b).
No differences in casein-specific IgG2a levels of mice fed the control diet were found. Casein-specific IgG2a levels of casein-sensitized mice fed the scGOS/lcFOS/pAOS diet were enhanced as compared to casein-sensitized control fed mice upon anti-CD25 treatment (p<0.05, Figure 5c). However, also sham-sensitized mice fed the scGOS/lcFOS/pAOS tended to be enhanced (p<0.05, Figure 5c). After Treg cell depletion, in both diet groups, casein-specific IgG2a levels were enhanced, although only the scGOS/lcFOS/pAOS group reached significance compared to sham sensitized controls (p<0.01).

152
8
Dietary non-digestible carbohydrates inhibit the allergic effector response via induction of casein-specific CD25+ regulatory T-cells in orally sensitized mice
Figure 5. Casein-specific immunoglobulin levels in sham- or casein-sensitized mice. Mice were i.p. injected with anti-CD25 or isotype control antibodies and fed control or scGOS/lcFOS/pAOS diet. a Casein-specific IgE, a casein-specific IgG1 and c casein-specific IgG2a. Means ± SEM, * p<0.05, ** p<0.01 and n=6 per group.

153
8
Chapter 8
Figure 6. Adoptive transfer of splenocytes in absence or presence of ex vivo depleted CD25+ cells. Casein-sensitized recipient mice adoptively transferred with splenocytes from casein-sensitized scGOS/lcFOS/pAOS fed donor mice were protected against the development of an acute allergic skin reaction whereas sham-sensitized donor cells did not transfer this tolerance. Ex vivo depletion of CD25+ Treg cells from splenocytes of scGOS/lcFOS/pAOS fed donor mice abolished this casein-specific protection. Donor cells of casein-sensitized mice fed the control diet did not protect against the development of the acute allergic skin response in recipient mice and Treg cell depletion did not alter this response. Delta ear swelling was calculated, subtracting the mean ear swelling of corresponding sham-sensitized recipient mice. Means ± SEM, ** p<0.01, *** p<0.001 and n=4-6 per group.
Acute allergic skin response of recipient mice in adoptive transfer experiments
Splenocytes of sham- or casein-sensitized mice fed control or scGOS/lcFOS/pAOS diet were adoptively transferred to recipient mice (Table I). Recipient mice, fed the control diet, were sensitized and the acute allergic skin response was measured after i.d. challenge with casein. All recipient mice developed an acute allergic skin reaction, which was not prevented using splenocytes of casein-sensitized mice fed the control diet (Figure 6). In contrast, adoptive transfer of splenocytes of casein-sensitized mice fed the scGOS/lcFOS/pAOS diet almost completely prevented development of the acute allergic skin reaction. This effect was casein specific since splenocytes of sham-sensitized mice fed the control diet did not transfer this tolerance. Ex vivo CD25+ Treg cell depletion completely abrogated the protective effect (p<0.001).

154
8
Dietary non-digestible carbohydrates inhibit the allergic effector response via induction of casein-specific CD25+ regulatory T-cells in orally sensitized mice
Casein-specific immunoglobulins of recipient mice in adoptive transfer experiments
Casein-specific IgE levels of sham-sensitized recipient mice receiving splenocytes from sham-sensitized donor mice was low (control diet 4.0 ± 0.22, scGOS/lcFOS/pAOS 4.8 ± 0.71 AU; data not shown) and did not enhance upon sensitization (Figure 7a). However, when the casein-sensitized recipient mice were transferred with splenocytes from casein-sensitized donors (control as well as scGOS/lcFOS/pAOS fed) the casein-specific IgE levels tended to enhance. Ex vivo CD25+ Treg cell depletion did not affect this. A similar tendency was found for casein-specific IgG1 and IgG2a levels (Figure 7b and 7c). Casein-sensitization of recipient mice transferred with splenocytes from sham-sensitized donor mice resulted in enhanced levels of casein-specific IgG1 when compared to sham-sensitization (control fed 105 ± 4.4 and scGOS/lcFOS/pAOS fed 103 ± 20 AU; data not shown; Figure 7b). Transfer with splenocytes of casein-sensitized donors tended to enhance this response. Ex vivo CD25+ Treg cell depletion did not affect this and no differences were found between recipients transferred with splenocytes of mice fed the control or scGOS/lcFOS/pAOS diet. Casein-specific IgG2a levels were not enhanced upon sensitization (data not shown). However, casein-specific IgG2a levels of casein-sensitized recipient mice receiving splenocytes from casein-sensitized control diet fed donor mice were enhanced as compared to recipients receiving splenocytes of sham-sensitized control diet fed donor mice (p<0.001; Figure 7c). The scGOS/lcFOS/pAOS diet showed the same tendency and ex vivo CD25+ Treg cell depletion did not alter this response.
In general, no differences in casein-specific immunoglobulin levels were found between recipient mice receiving splenocytes of control or scGOS/lcFOS/pAOS fed mice and ex vivo CD25+ Treg cell depletion did not affect immunoglobulin levels.
Discussion
In this study, pivotal involvement of CD25+ regulatory T-cells (Treg) is demonstrated in the protective effects of a specific oligosaccharide mixture on the induction of cow’s milk allergy in mice orally sensitized with casein. Dietary intervention with scGOS/lcFOS/pAOS was started two weeks prior to the first sensitization and continued during the sensitization phase and was found to reduce the acute allergic skin response. These findings are in accordance with a study in infants at high risk for allergies; when these children were fed hypo-allergic milk formulas supplemented with scGOS and lcFOS during the first two years of life, the incidence of atopic dermatitis decreased and reduced allergic manifestations and infections were observed compared to control formula fed infants [5, 26]. We hypothesized that the

155
8
Chapter 8
Figure 7. Casein-specific immunoglobulin levels in serum of recipient mice after adoptive transfer with whole spleen suspension or ex vivo CD25+ Treg cells depleted splenocytes. No differences in immunoglobulin levels were observed in mice transferred with splenocytes of control and scGOS/lcFOS/pAOS fed mice and ex vivo Treg cell depletion did not affect immunoglobulin levels. a Casein-specific IgE b casein-specific IgG1 c casein-specific IgG2a. Means ± SEM, * p<0.05, ** p<0.01 and n=6 per group.

156
8
Dietary non-digestible carbohydrates inhibit the allergic effector response via induction of casein-specific CD25+ regulatory T-cells in orally sensitized mice
diet affects the mucosal immune system and actively interferes in oral tolerance induction. Allergies develop as a consequence of a disparity in the TH1/TH2 balance towards TH2 [27]. Different Treg cell subsets actively regulate this balance locally in the gastro-intestinal tract. Antigens, or dendritic cells loaded with antigen, travel from the Peyer’s patches to the mesenteric lymph nodes (MLN) and are presented to T-cells. Antigen-responsive T-cells (CD4+) travel subsequently through the bloodstream into the mucosa or to the peripheral immune system (e.g. spleen) [11, 12]. Antigen specific T-cells from the MLN in this respect can relocate systemic tolerance. In the present study the scGOS/lcFOS/pAOS mixture was found to affect the TH1/TH2 balance in the MLN of casein-sensitized mice. The percentage of activated TH1 cells was enhanced 24 h after oral challenge with casein while the number of activated TH2 cells tended to reduce. This effect was only observed in the scGOS/lcFOS/pAOS fed casein-sensitized mice and associated with a reduced systemic acute allergic skin response. The percentage of TH1 in casein-sensitized mice fed control diet was unaltered compared to sham-sensitized mice. This may indicate active regulation of the mucosal immune response as consequence of dietary intervention with scGOS/lcFOS/pAOS occurring in the presence of casein. These effects in the MLN may relate to the peripheral reduction of acute allergic effects. Local induced tolerance can be transferred to the periphery resulting in peripheral tolerance, in particular Treg cells have been indicated to contribute to this phenomenon [12]. Sun et al. [28] showed that the oral administration of Ag induced oral tolerance both locally (mucosal lymphoid organs) and systemically (spleen) was induced by CD4+CD25+Foxp3+ cells.
To study the contribution of CD25+ Treg cells in the prevention of the acute allergic skin reaction by the diet in vivo depletion studies were performed. Monoclonal anti-CD25 antibody (mAb) treatment resulted in a decrease of regulatory CD4+CD25+Foxp3+ cells in the spleen, whereas CD4+CD25+ effector cells were unaffected. It has been shown that the PC61 anti-CD25 mAb selectively depletes the CD25high cells which are CD4+ [29]. Indeed, in the current study the in vivo depletion of CD4+CD25+Foxp3+ cells was confirmed to be more than 65%. Also in other studies it was shown that complete in vivo depletion is not necessary in order to investigate Treg cell function [30-33]. In the present study in vivo depletion of CD25+ Treg cells was found to abrogate the protective effects of the scGOS/lcFOS/pAOS diet, when treated with anti-CD25 scGOS/lcFOS/pAOS fed mice did develop a severe acute allergic skin response. Indicating a role for CD25+ Treg cells in the preventive effects caused by the diet.
However, the depletion of CD25+ Treg cells also resulted in enhanced casein-specific immunoglobulin levels. This was in particular found in mice fed the scGOS/lcFOS/pAOS diet, since the diet tended to suppress casein specific TH2 type IgE and IgG1 levels upon sensitization. In vivo Treg cell depletion enhanced casein specific IgG1 and tended to enhance IgE in these mice compared to treatment with isotype control antibodies, indicating an exacerbated TH2 response upon in vivo anti-CD25 treatment. The imbalance in favor of TH2

157
8
Chapter 8
in allergy is regulated by Treg cells resulting in a suppression of the allergic TH2 response, depletion of Treg cells can therefore result in exacerbation of allergic sensitization. Also Sumi et al. [34] showed that in a murine model for allergic conjunctivitis treatment with anti-CD25 mAb resulted in increased levels of specific IgE as well as IgG1. Therefore, in vivo anti-CD25 mAb treatment may have enhanced the capacity of B-cells to produce antibodies since regulation by Treg cells was disrupted. The increased antibody levels upon in vivo anti-CD25 mAb treatment could have underlain abrogation of the protective effect of the scGOS/lcFOS/pAOS diet.
To discriminate between these interfering effects adoptive transfer experiments with splenocytes were conducted to confirm the contribution of Treg cells in the protective effects of the diet. Splenocytes, untreated or ex vivo depleted for CD25+ cells, from sham- and casein-sensitized donor mice fed control and scGOS/lcFOS/pAOS diet were transferred to naïve recipient mice that were sham- or casein-sensitized and fed control diet. Casein-sensitized recipients receiving splenocytes from scGOS/lcFOS/pAOS fed casein-sensitized donor mice were protected from the development of an acute allergic casein-specific skin response, which was abrogated after ex vivo anti-CD25 mAb treatment of the splenocytes. These findings were in line with our previous finding using in vivo CD25+ Treg cell depletion. However ex vivo depletion did not alter the levels of casein-specific immunoglobulins when compared to recipient mice transferred with whole spleen cell suspensions of casein-sensitized donor mice. Hence, the adoptive transfer studies confirmed Treg cells to be involved in the protective effects provided by the scGOS/lcFOS/pAOS diet.
In casein-sensitized recipients transferred with splenocytes of casein-sensitized donor mice, IgG
2a levels were increased and IgE and IgG1 showed similar tendency, when compared to recipients transferred with sham-sensitized donor splenocytes. Guttormsen et al. [35] already found that whole spleen single cell suspensions were able to transfer immunologic memory for a vaccine in adoptive transfer experiments. This is comparable to the observations in the casein allergic mice.
The donor diets did not differentially alter the immunoglobulin levels of the recipient mice and Treg cell depletion also was found to leave antibody levels unaffected high. Typically, the acute allergic skin response was completely prevented in recipient mice transferred with cells of casein-sensitized donor mice fed the scGOS/lcFOS/pAOS diet. This suggests a Treg cell mediated suppression of the effector immune response independent of antibody production. Gri et al.[36] showed that Treg cells are able to inhibit acute allergic responses which may relate to the reduced allergic skin reaction observed in the present study in mice fed the scGOS/lcFOS/pAOS diet. Mast cells are most probably involved in this acute reaction and Treg cells are believed to alter mast cell degranulation by IL-10 production or cell-cell contact [36, 37]. In addition, Treg cells have been shown to suppress Fc-receptor expression

158
8
Dietary non-digestible carbohydrates inhibit the allergic effector response via induction of casein-specific CD25+ regulatory T-cells in orally sensitized mice
on mast cells which would reduce the capacity of these cells to degranulate and induce an acute skin response [38].
Both the in vivo and ex vivo CD25+ Treg cell depletion studies suggest a central participation of CD25+ Treg cells in the preventive effects of scGOS/lcFOS/pAOS diet on casein induced acute allergic responses. Additionally, these data suggest the existence of antigen-specific CD25+ Treg cells, since only splenocytes from casein-sensitized and not splenocytes from sham-sensitized donor mice fed the scGOS/lcFOS/pAOS diet could prevent the acute allergic response in recipient mice. In a review by Koenen and Joosten [39], protocols to identify and proliferate antigen-specific Treg cells are provided and the use of these cells in preventing solid organ transplant rejection is hypothesized. Multiple studies have speculated on an antigen-specific Treg cell population and their application in different diseases [39-41].
In the present study it is shown that dietary intervention with scGOS/lcFOS/pAOS reduces the development of an acute allergic response upon antigen challenge, although specific immunoglobulins levels remain high. Both in vivo as well as ex vivo depletion of CD25+ Treg cell abrogated the diminished acute allergic response and imply crucial involvement of antigen specific CD25+ Treg cells in the suppression of the allergic effector response.
References
1. Fanaro, S., et al., Galacto-oligosaccharides and long-chain fructo-oligosaccharides as
prebiotics in infant formulas: a review. Acta Paediatr Suppl, 2005. 94(449): p. 22-6.
2. Boehm, G., et al., Prebiotic carbohydrates in human milk and formulas. Acta Paediatr Suppl,
2005. 94(449): p. 18-21.
3. Fanaro, S., et al., Acidic oligosaccharides from pectin hydrolysate as new component for infant
formulae: effect on intestinal flora, stool characteristics, and pH. J Pediatr Gastroenterol
Nutr, 2005. 41(2): p. 186-90.
4. Schouten, B., et al., Cow milk allergy symptoms are reduced in mice fed dietary synbiotics
during oral sensitization with whey. J Nutr, 2009. 139(7): p. 1398-403.
5. Moro, G., et al., A mixture of prebiotic oligosaccharides reduces the incidence of atopic
dermatitis during the first six months of age. Arch Dis Child, 2006. 91(10): p. 814-9.
6. Vandenplas, Y., Oligosaccharides in infant formula. Br J Nutr, 2002. 87 Suppl 2: p. S293-6.
7. Haarman, M. and J. Knol, Quantitative real-time PCR assays to identify and quantify fecal
Bifidobacterium species in infants receiving a prebiotic infant formula. Appl Environ
Microbiol, 2005. 71(5): p. 2318-24.
8. Boehm, G., et al., Supplementation of a bovine milk formula with an oligosaccharide mixture
increases counts of faecal bifidobacteria in preterm infants. Arch Dis Child Fetal Neonatal Ed,

159
8
Chapter 8
2002. 86(3): p. F178-81.
9. Crittenden, R.G. and L.E. Bennett, Cow’s milk allergy: a complex disorder. J Am Coll Nutr,
2005. 24(6 Suppl): p. 582S-91S.
10. Monaci, L., et al., Milk allergens, their characteristics and their detection in food: a review.
European Food Research and Technology, 2005(Volume 223, Number 2 / June, 2006): p.
149-179.
11. Mowat, A.M., Anatomical basis of tolerance and immunity to intestinal antigens. Nat Rev
Immunol, 2003. 3(4): p. 331-41.
12. Tsuji, N.M. and A. Kosaka, Oral tolerance: intestinal homeostasis and antigen-specific
regulatory T cells. Trends Immunol, 2008. 29(11): p. 532-40.
13. van Wijk, F., et al., CD4+CD25+ T cells regulate the intensity of hypersensitivity responses
to peanut, but are not decisive in the induction of oral sensitization. Clin Exp Allergy, 2007.
37(4): p. 572-81.
14. Akbari, O. and D.T. Umetsu, Role of regulatory dendritic cells in allergy and asthma. Curr
Opin Allergy Clin Immunol, 2004. 4(6): p. 533-8.
15. O’Garra, A. and P. Vieira, Regulatory T cells and mechanisms of immune system control. Nat
Med, 2004. 10(8): p. 801-5.
16. Jukes, J.P., K.J. Wood, and N.D. Jones, Natural killer T cells: a bridge to tolerance or a pathway
to rejection? Transplantation, 2007. 84(6): p. 679-81.
17. Izcue, A. and F. Powrie, Special regulatory T-cell review: Regulatory T cells and the intestinal
tract--patrolling the frontier. Immunology, 2008. 123(1): p. 6-10.
18. Akdis, C.A. and M. Akdis, Mechanisms and treatment of allergic disease in the big picture of
regulatory T cells. J Allergy Clin Immunol, 2009. 123(4): p. 735-46; quiz 747-8.
19. Huehn, J., J.K. Polansky, and A. Hamann, Epigenetic control of FOXP3 expression: the key to
a stable regulatory T-cell lineage? Nat Rev Immunol, 2009. 9(2): p. 83-9.
20. Sletten, G.B., et al., Memory T Cell Proliferation in Cow’s Milk Allergy after CD25+ Regulatory
T Cell Removal Suggests a Role for Casein-Specific Cellular Immunity in IgE-Mediated but
Not in Non-IgE-Mediated Cow’s Milk Allergy. Int Arch Allergy Immunol, 2006. 142(3): p. 190-
198.
21. Karlsson, M.R., J. Rugtveit, and P. Brandtzaeg, Allergen-responsive CD4+CD25+ regulatory T
cells in children who have outgrown cow’s milk allergy. J Exp Med, 2004. 199(12): p. 1679-
88.
22. Tiemessen, M.M., et al., Cow’s milk-specific T-cell reactivity of children with and without
persistent cow’s milk allergy: key role for IL-10. J Allergy Clin Immunol, 2004. 113(5): p. 932-
9.
23. Vos, A.P., et al., A specific prebiotic oligosaccharide mixture stimulates delayed-type
hypersensitivity in a murine influenza vaccination model. Int Immunopharmacol, 2006. 6(8):
p. 1277-86.
24. Vos, A.P., et al., Dietary supplementation of neutral and acidic oligosaccharides enhances
Th1-dependent vaccination responses in mice. Pediatr Allergy Immunol, 2007. 18(4): p. 304-

160
8
Dietary non-digestible carbohydrates inhibit the allergic effector response via induction of casein-specific CD25+ regulatory T-cells in orally sensitized mice
12.
25. Schouten, B., et al., Acute Allergic Skin Reactions and Intestinal Contractility Changes in Mice
Orally Sensitized against Casein or Whey. Int Arch Allergy Immunol, 2008. 147(2): p. 125-
134.
26. Arslanoglu, S., et al., Early dietary intervention with a mixture of prebiotic oligosaccharides
reduces the incidence of allergic manifestations and infections during the first two years of
life. J Nutr, 2008. 138(6): p. 1091-5.
27. Romagnani, S., Immunologic influences on allergy and the TH1/TH2 balance. J Allergy Clin
Immunol, 2004. 113(3): p. 395-400.
28. Sun, J.B., et al., Oral tolerance induction with antigen conjugated to cholera toxin B subunit
generates both Foxp3+CD25+ and Foxp3-CD25- CD4+ regulatory T cells. J Immunol, 2006.
177(11): p. 7634-44.
29. Amu, S., et al., B-cell CD25 expression in murine primary and secondary lymphoid tissue.
Scand J Immunol, 2006. 64(5): p. 482-92.
30. Shreffler, W.G., et al., Association of allergen-specific regulatory T cells with the onset of
clinical tolerance to milk protein. J Allergy Clin Immunol, 2009. 123(1): p. 43-52 e7.
31. Kohm, A.P., et al., Cutting Edge: Anti-CD25 monoclonal antibody injection results in the
functional inactivation, not depletion, of CD4+CD25+ T regulatory cells. J Immunol, 2006.
176(6): p. 3301-5.
32. Lewkowich, I.P., et al., CD4+CD25+ T cells protect against experimentally induced asthma
and alter pulmonary dendritic cell phenotype and function. J Exp Med, 2005. 202(11): p.
1549-61.
33. Matsushita, N., et al., Comparative methodologies of regulatory T cell depletion in a murine
melanoma model. J Immunol Methods, 2008. 333(1-2): p. 167-79.
34. Sumi, T., et al., Thymus-derived CD4+ CD25+ T cells suppress the development of murine
allergic conjunctivitis. Int Arch Allergy Immunol, 2007. 143(4): p. 276-81.
35. Guttormsen, H.K., et al., Immunologic memory induced by a glycoconjugate vaccine in a
murine adoptive lymphocyte transfer model. Infect Immun, 1998. 66(5): p. 2026-32.
36. Gri, G., et al., CD4+CD25+ regulatory T cells suppress mast cell degranulation and allergic
responses through OX40-OX40L interaction. Immunity, 2008. 29(5): p. 771-81.
37. Bundoc, V.G. and A. Keane-Myers, IL-10 confers protection from mast cell degranulation in a
mouse model of allergic conjunctivitis. Exp Eye Res, 2007. 85(4): p. 575-9.
38. Kashyap, M., et al., Cutting edge: CD4 T cell-mast cell interactions alter IgE receptor
expression and signaling. J Immunol, 2008. 180(4): p. 2039-43.
39. Koenen, H.J. and I. Joosten, Antigen-specific regulatory T-cell subsets in transplantation
tolerance regulatory T-cell subset quality reduces the need for quantity. Hum Immunol,
2006. 67(9): p. 665-75.
40. Zhang, Z.X., et al., Identification of a previously unknown antigen-specific regulatory T cell
and its mechanism of suppression. Nat Med, 2000. 6(7): p. 782-9.

161
8
Chapter 8
41. Ghoreishi, M., et al., Expansion of antigen-specific regulatory T cells with the topical vitamin
d analog calcipotriol. J Immunol, 2009. 182(10): p. 6071-8.


9
iNKT cells contribute to the allergic response in cow’s milk protein sensitized mice
Bastiaan Schouten1
Betty C.A.M. van Esch1,2
Stephanie M. Lim1
Gerard A. Hofman1
Léon M.J. Knippels2
Linette E.M. Willemsen1
Johan Garssen1,2
Arjan P. Vos2
1 Department of Pharmacology and Pathophysiology, Utrecht Institute for Pharmaceutical
Sciences, Faculty of Science, Utrecht University, Utrecht, The Netherlands2 Danone Research – Centre for Specialised Nutrition, Wageningen, The Netherlands
To be submitted for publication

164
9
iNKT cells contribute to the allergic response in cow’s milk protein sensitized mice
Abstract
Background: Little is known about the contribution of the invariant natural killer T (iNKT)-cells in the onset of food allergy. Using a mouse model for cow’s milk allergy the function of iNKT cells was investigated. α-Galactosylceramide (αGalCer), a specific iNKT cell agonist, was used to stimulate iNKT cells.Methods: Female C3H/HeOuJ mice were sensitized five times orally with casein or whey proteins. One hour before the sensitizations mice were treated intraperitoneally with αGalCer (2 µg/mice) or control. One week after the last sensitization mice were challenged intradermally in the ear and acute allergic skin reactions were measured. Sera were analyzed for specific immunoglobulins. Furthermore, in liver, spleen and mesenteric lymph nodes (MLN) percentages of iNKT cells were analyzed and liver lymphocyte restimulation assays were performed.Results: Mice treated with αGalCer showed enhanced acute allergic skin reactions in comparison with sham treated casein- or whey-sensitized mice. This finding was associated with increased levels of whey-specific immunoglobulins, while casein-specific immunoglobulins were not affected. The percentage of iNKT cells in the liver of casein- and whey-sensitized mice was reduced when compared to sham-sensitized mice. However, in whey-sensitized mice IL-4, IFN-γ and IL-10 secretion upon ex vivo αGalCer restimulation was unaffected. The iNKT cell frequency was enhanced in the spleen and MLN of whey- and not in casein-sensitized mice. Strikingly, αGalCer treatment was found to deplete iNKT cells in the liver of sensitized as well as sham-sensitized mice, and the hepatocytes did not respond to ex vivo restimulation with αGalCer.Conclusion: iNKT cell frequencies were reduced in the liver of casein- and whey-sensitized mice and enhanced in the spleen and MLN of whey allergic mice. Due to repeated application of αGalCer, hepatocytes were functionally depleted for iNKT cells. This resulted in an increased allergic effector response which was most pronounced in whey-sensitized mice and associated with enhanced whey-specific immunoglobulin levels. In conclusion, iNKT cells may suppress cow’s milk allergic symptoms in mice and may differentially regulate oral sensitization for casein and whey.

165
9
Chapter 9
Introduction
Cow’s milk allergy (CMA) affects 3-5% of the infants and is outgrown in most of the cases within 3-5 years [1, 2]. IgE-mediated CMA predisposes infants to other food allergies and even asthma later in life [3, 4]. In most individuals the condition comprises skin, respiratory tract and gastrointestinal tract symptoms and in some cases, although rare, even systemic anaphylaxis [5, 6]. Oral tolerance is believed to be induced in the mesenteric lymph nodes (MLN) and Peyer’s patches (PP) by regulatory T-(Treg)cells [7, 8]. However, invariant Natural Killer T- (iNKT) cells play a pivotal role as well in oral tolerance by inducing Treg cells and in addition, by clonally deleting antigen-specific CD4+ T-cells [9]. Natural Killer T- (NKT) cells are a subset of cells that express a T cell receptor (TCR), which makes them T-lymphocytes. There are several subsets of NKT cells, however the most studied NKT cell is the invariant NKT (iNKT), or type I NKT cell, which express an invariant Vα14Jα18 TCR chain in combination with a Vβ2, Vβ7 and Vβ8 in mice [10]. iNKT-cells, T-cells and NK-cells share cell surface markers like CD161 and NKR-P1 [11]. Whereas T-cells interact with peptides presented on MHC-class I or II molecules by antigen presenting cells, iNKT cells do not interact with MHC-class I or II molecules. However, iNKT cells recognize glycolipids presented by CD1d, a non-classical antigen presenting molecule [11, 12]. Therefore, iNKT cells can generate an adaptive immune response towards a class of antigens which is not covered by conventional T-cells. These antigens include mycobacterial cell wall antigens, gangliosides, tumor-derived phospholipids and nonlipidic molecules [13, 14]. The ability of iNKT cells to show flexibility in their cytokine response have led to the proposition that iNKT cells have regulatory properties and might be involved in homeostasis of the mucosal immune system [12, 15, 16]. In mice, these cells are present at low frequencies in the thymus, bone marrow, spleen, MLN and PP (<2.0%) [17] (Schouten et al., unpublished data). In contrast, hepatic lymphocytes consist of approximately 20% iNKT cells. The best characterized and potent specific iNKT cell agonist is a glycolipid isolated from the marine sponge Agelas mauritianus, α-galactosylceramide (αGalCer) [18-20]. The complex of CD1d plus αGalCer binds to the iNKT cell TCR and activates the cell. After iNKT stimulation interleukin (IL)-4 and interferon (IFN)-γ are rapidly produced and secreted. More recently, it has been shown that iNKT cells produce also T
H17 cytokines, both IL-17 and IL-22 [21, 22].
The function of iNKT cells has been studied in several diseases including diabetes [23], multiple sclerosis [23], colitis [24] and cancer [25]. However, the contribution of iNKT cells, using αGalCer, in the onset of allergy is, to our knowledge, only studied in allergic asthma models [26-28]. With regard to food allergy and in particular cow’s milk allergy so far, no data are available on the role of iNKT cells in induction or maintenance of allergy to food ingredients and/or tolerance. Therapeutic approaches include the elimination of cow’s milk proteins from the diet and several forms of immunotherapy [29-31]. To assess the

166
9
iNKT cells contribute to the allergic response in cow’s milk protein sensitized mice
mechanisms underlying CMA, the present study focused on the contribution of iNKT cells in casein and whey protein induced hypersensitivity. The effects of αGalCer application, as an iNKT-specific stimulus, were studied during oral sensitization in a murine CMA model.
Methods
Chemicals
Casein and whey proteins were obtained from DMV International (Veghel, The Netherlands). Cholera toxin was purchased from Quadratech Diagnostics (Epsom, UK). PBS from Cambrex Bio Science (Verviers, Belgium). Biotin-labeled rat anti-mouse IgE, IgG1, IgG2a and IgA from BD Biosciences (Alphen aan den Rijn, The Netherlands). αGalCer was a kindly gift from Kyowa Hakko Kirin Co., Ltd (Japan). All other chemicals were obtained from Sigma-Aldrich-Chemie (Zwijndrecht, The Netherlands).
Oral sensitization, treatment and challenge of mice
Three-week-old specific pathogen free female C3H/HeOuJ mice were purchased from Charles River Laboratories (Maastricht, The Netherlands), bred and raised on a cow’s milk protein free diet and housed in the animal facility. Animal care and use were performed in accordance with the guidelines of the Dutch Committee of Animal Experiments. Mice were sensitized intragastrically (i.g.) with 0.5 mL homogenized casein or whey (40 mg/mL PBS) with cholera toxin (CT, 20 μg/mL PBS) as an adjuvant, using a blunt needle. Control mice received CT alone. Mice were boosted weekly for a period of 5 weeks as described previously [32]. One hour before every sensitization mice were intraperitoneally (i.p.) treated with 100 μL αGalCer (20 µg/mL PBST) or control (PBS and 0.5% Tween). One day after the last sensitization feces were collected and one week after the last sensitization mice were challenged intradermally (i.d) in the ear pinnae to determine the acute allergic skin reaction and challenged i.g. with 0.5 mL homogenized casein or whey (100 mg/mL PBS). After 16 hours blood samples were collected in Minicollect® Z Serum Sep vials (Greiner bio-one) and mice were sacrificed by cervical dislocation. Collected blood was centrifuged 15 min at 16000 x g and sera were stored at –70oC.

167
9
Chapter 9
Acute allergen specific skin reaction
The acute allergen specific skin reaction was measured after i.d. injection with 20 μL homogenized casein or whey protein (0.5 mg/mL in PBS) in the ear pinnae. Ear thickness was measured in duplicate using a digital micrometer (Mitutoyo, Veenendaal, The Netherlands), at t=0 and 1 hour after challenge. The ear swelling (μm) was calculated by subtracting the basal (t=0 hour) from the specific casein or whey induced ear swelling after 1 hour.
Measurement of specific serum immunoglobulins
Concentrations of casein- and whey-specific IgE, IgG1 and IgG2a were determined in serum whereas specific IgA was measured in feces by means of ELISA as described before [32]. In short, plates were coated with 100 µL casein or whey (20 mg/L) and after washing several serum or fecal water dilutions were added. Bound whey or casein specific antibodies were detected using biotin-labeled rat anti-mouse IgE, IgG
1, IgG2a or IgA (1 mg/L) and streptavidin-horseradish peroxidase (HRP). Results were expressed as arbitrary units (AU), calculations were based on titration curves with pooled reference sera from casein or whey alum-i.p. immunized mice.
Flow cytometry
Single cell suspensions were prepared from mouse livers, liver lymphocytes were isolated using 40% Percoll gradient, spleens and MLN and blocked for 30 min in PBS containing 1% BSA and 5% FCS. Cells were counted using a Coulter Z1 particle counter (Beckman Coulter) and diluted to a concentration of 2x106 cells/mL. For identification of iNKT cells, cells were incubated for 30 min at 4oC with anti-CD3-PerCP-CY5.5 (1:100), anti-CD4-FITC (1:100), anti-CD19-APC and CD1d-tetramer loaded with αGalCer (1:100). Antibodies were purchased from BD (Alphen aan den Rijn, The Netherlands), except for CD1d-tetramer preloaded with αGalCer, which were purchased from ProImmune Ltd. (Oxford, UK). iNKT cells were gated using CD3+CD19-CD4+CD1d/αGalCer tetramer+. Flow cytometric analysis was performed using FACSCalibur (BD, Alphen aan den Rijn, The Netherlands) and CellQuest Pro Analysis software (BD, Alphen aan de Rijn, The Netherlands).

168
9
iNKT cells contribute to the allergic response in cow’s milk protein sensitized mice
Restimulation assay
In hepatocyte culture supernatants of single cell suspensions (1x106/mL) restimulated using αGalCer (100ng/ml) or control were harvested at 48 h and concentrations of granulocyte macrophage colony-stimulating factor (GM-CSF), IFN-γ, tumor growth factor (TNF)-α, IL-2, IL-4, IL-5, IL-10 and IL-12p70 were determined by multiplex cytokine bead assay (Bio-Rad, Veenendaal, The Netherlands) using a Luminex 200 Analyzer (Nuclilab, Ede, The Netherlands). Data were analyzed using xPonent software 3.0.380.0 (Luminex Corporation, Austin, USA).
Statistics
All data were analyzed using the one way ANOVA and post hoc Bonferroni’s Comparison Test or Mann-Whitney U test. Analyses of the immunoglobulin levels and cytokine concentrations were performed on log transformed data. Statistical analyses were conducted using GraphPad Prism software (version 4.03). P-values of * p<0.05, ** p<0.01 and *** p<0.001 were indicated when analyses reached statistically significance. Data are represented as means ± SEM.
Figure 1. The acute allergic skin reaction of sham- casein- or whey-challenged mice, control or αGalCer treated. Mice were sensitized i.g. with cholera toxin (CT) and whey or casein for 5 times during weekly intervals, CT alone was used for sham-sensitization. Before each sensitization mice were i.p injected using αGalCer or control. Means ± SEM, * p<0.05 and *** p<0.001 and n=5-6 per group.

169
9
Chapter 9
Results
Acute allergic skin reaction
One week after the last sensitization mice were challenged i.d. in the ear with casein or whey. After one hour the acute allergic skin reaction was determined. αGalCer treatment did not affect the ear swelling reaction of sham-sensitized mice (60.7 ± 3.3 vs 76.0 ± 10.1 µm for casein challenge and 53.7 ± 4.6 vs 54.5 ± 8.0 µm for whey challenge). However, both casein- and whey-sensitized mice developed an increased acute allergic skin reaction after αGalCer treatment (137.2 ± 4.5 vs 162.5 ± 7.5 µm for casein-sensitized mice, p<0.05 and 136.4 ± 12.7 vs 209.9 ± 10.2 µm for whey-sensitized mice; p<0.001; Figure 1).
Allergen-specific immunoglobulin levels
Sixteen hours after oral challenge, serum was obtained from all mice. Casein-specific IgE, IgG1 and IgA levels were enhanced in casein- and whey-sensitized mice when compared to sham-sensitization (Figure 2a, b, d, e, f and h). Specific IgG2a was only enhanced in whey-sensitized mice and tended to enhance in casein-sensitized mice (Figure 2c, g). αGalCer treatment did not affect immunoglobulin levels in casein-sensitized mice but further enhanced whey-specific IgE, IgG1, IgG2a, but not IgA in whey-sensitized mice (Figure 2e, f, g and h).
Flow cytometry
The effect of sensitization and αGalCer treatment on iNKT cell frequency was determined using flow cytometry. Casein- (14.6 ± 2.5%) and whey-sensitization (14.0 ± 2.8%) was found to reduce the percentage of hepatic iNKT cells in comparison with sham-sensitized mice (22.9 ± 0.6%; p<0.05; Figure 3a). In addition, αGalCer treatment was found to deplete hepatic iNKT cells in both sham- and casein- or whey-sensitized mice in comparison with control treated mice (p<0.001; Figure 3a). Low percentages of iNKT cells were found in the spleen and the MLN possibly as a consequence of migration. Whey-sensitization was found to enhance iNKT cell percentages in the spleen of whey-sensitized mice (2.4 ± 0.2%) as compared to sham- (1.4 ± 0.2) and casein-sensitized (1.2 ± 0.1%) mice and αGalCer treatment was found to abrogate this (1.5 ± 0.1%; p<0.001; Figure 3b). In addition, whey-sensitization was found to enhance iNKT cell percentages in the MLN of whey-sensitized mice (0.92 ± 0.04%) as
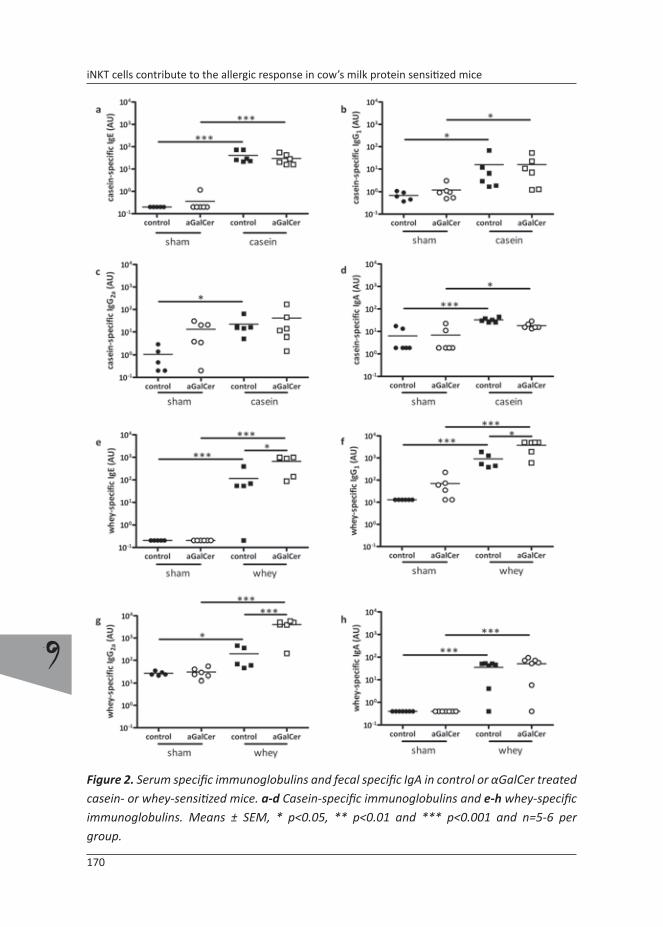
170
9
iNKT cells contribute to the allergic response in cow’s milk protein sensitized mice
Figure 2. Serum specific immunoglobulins and fecal specific IgA in control or αGalCer treated casein- or whey-sensitized mice. a-d Casein-specific immunoglobulins and e-h whey-specific immunoglobulins. Means ± SEM, * p<0.05, ** p<0.01 and *** p<0.001 and n=5-6 per group.

171
9
Chapter 9
CD4
CD1d loaded with αGalCer
αGalCer1.2%
d
spleen
sham21.7%
liver
11.7%
sham13.2%
sham αGalCer0.9%
casein‐sensitized whey‐sensitizedsham‐sensitized
1.9% 0.9% 1.2% 1.4% 2.4% 1.4%
MLN
0.4% 0.3% 0.45% 0.6% 0.85% 0.75%
αGalCer0.8%
Figure 3. Flow cytometric analysis of iNKT cells in control or αGalCer treated sham- casein- or whey-sensitized mice. Cells were isolated and stained with CD3, CD4 and αGalCer-loaded CD1d tetramers. Percentages are calculated within the CD3+ population and representative dot plots are shown. a iNKT in the liver b in the spleen and c in the mesenteric lymph nodes. d Representative dot plots of the CD4+CD1d tetramers loaded with αGalCer+ in the CD3+ population. Means ± SEM, * p<0.05 and *** p<0.001 and n=5-6 per group.
compared to sham- (0.4 ± 0.0%) and casein-sensitized mice (0.47 ± 0.02% (p<0.05, p<0.01; Figure 3c). The frequency of MLN iNKT in whey-sensitized mice was higher than casein-sensitized mice (p<0.05). αGalCer treatment was not found to affect the percentage of NKT cells in whey-sensitized mice but did increase MLN iNKT cells of casein-sensitized mice vs control treated casein-sensitized mice (0.7 ± 0.09 vs 0.5 ± 0.02%; p<0.05; Figure 3c).
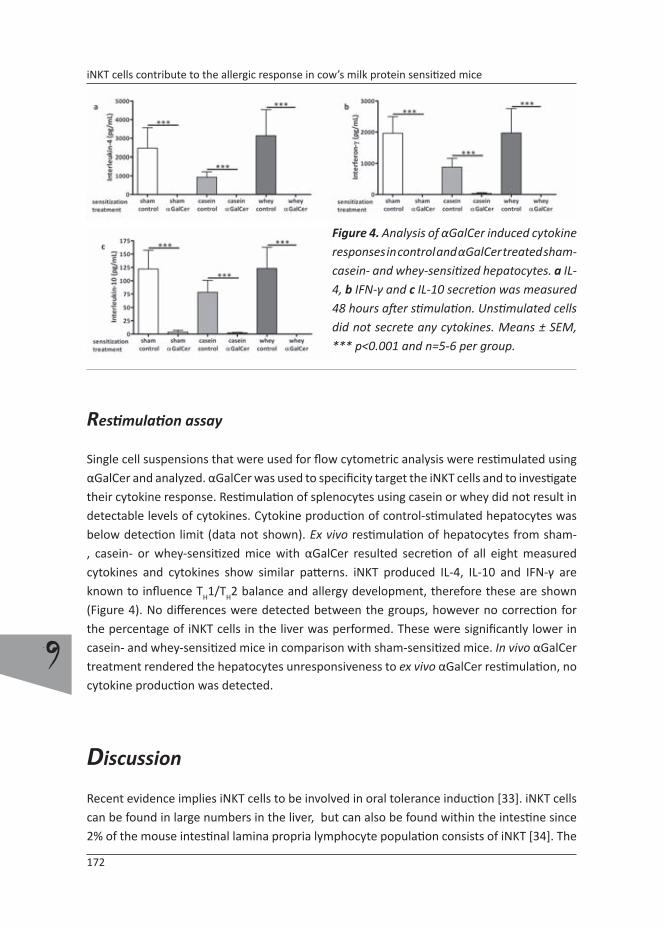
172
9
iNKT cells contribute to the allergic response in cow’s milk protein sensitized mice
Restimulation assay
Single cell suspensions that were used for flow cytometric analysis were restimulated using αGalCer and analyzed. αGalCer was used to specificity target the iNKT cells and to investigate their cytokine response. Restimulation of splenocytes using casein or whey did not result in detectable levels of cytokines. Cytokine production of control-stimulated hepatocytes was below detection limit (data not shown). Ex vivo restimulation of hepatocytes from sham-, casein- or whey-sensitized mice with αGalCer resulted secretion of all eight measured cytokines and cytokines show similar patterns. iNKT produced IL-4, IL-10 and IFN-γ are known to influence T
H1/TH2 balance and allergy development, therefore these are shown (Figure 4). No differences were detected between the groups, however no correction for the percentage of iNKT cells in the liver was performed. These were significantly lower in casein- and whey-sensitized mice in comparison with sham-sensitized mice. In vivo αGalCer treatment rendered the hepatocytes unresponsiveness to ex vivo αGalCer restimulation, no cytokine production was detected.
Discussion
Recent evidence implies iNKT cells to be involved in oral tolerance induction [33]. iNKT cells can be found in large numbers in the liver, but can also be found within the intestine since 2% of the mouse intestinal lamina propria lymphocyte population consists of iNKT [34]. The
Figure 4. Analysis of αGalCer induced cytokine responses in control and αGalCer treated sham- casein- and whey-sensitized hepatocytes. a IL-4, b IFN-γ and c IL-10 secretion was measured 48 hours after stimulation. Unstimulated cells did not secrete any cytokines. Means ± SEM, *** p<0.001 and n=5-6 per group.

173
9
Chapter 9
contribution of iNKT cells in the pathophysiology of cow’s milk allergy (CMA) is unknown, therefore the effect of oral sensitization on iNKT cell populations in the liver, spleen and MLN was investigated. Furthermore, αGalCer, a specific iNKT agonist, was used to intervene with the development of allergy during oral sensitization with casein or whey in mice.
Both casein- and whey-sensitization resulted in decreased iNKT cell percentages in the liver of cow’s milk allergic mice. Typically, in whey-sensitized mice iNKT cells percentages were increased in the spleen and MLN, this was not observed in casein-sensitized mice. These data show that the iNKT cell population is affected during sensitization and may imply that these cells contribute to the development of allergic disease or are involved in the suppression of (food) allergic disorders. iNKT cells have been shown to induce or break oral tolerance [27, 33] and may suppress or enhance allergic disease in mice [18, 35]. These dual mechanisms may partly be explained by the existence of different subsets of iNKT. In humans it was shown that part of the population of αGalCer reactive iNKT cells produced high amounts of IL-4, IL-10 and IFN-γ and induced IL-12 producing DC in vitro, resulting in T
H1 skewing of the effector response while another subset produced relatively low amounts of IL-4 and IL-10 and induced DC cytolysis and a TH2 polarized immune response [36]. Typically, in particular iNKT derived IL-4 and IL-10 were found to contribute to oral tolerance induction [33].
To further explore the involvement of iNKT in allergic sensitization for cow’s milk proteins, agonist studies using αGalCer were performed. αGalCer is the best characterized and potent specific iNKT cell agonist [18-20]. iNKT treatment was intended to restore the frequency and/or activity of hepatic iNKT cells in casein- or whey-sensitized mice in order to inhibit the acute allergic skin reaction. However, multiple i.p. injections of αGalCer resulted in an almost complete depletion of iNKT cells in the liver, both in sensitized as well as in control mice, and exacerbation of the allergic response in sensitized mice. A strong decrease in hepatic iNKT cells could be explained by migration of these cells towards other lymphoid organs or the periphery. However, FACS analysis did not show enhanced iNKT numbers in spleen or MLN, suggesting that the iNKT cells were depleted instead of stimulated. Parekh et al. [37, 38] demonstrated that a single high dose administration of αGalCer (5 µg/mouse) causes anergy of iNKT cells for at least one month. Matsuda et al. [20] as well as Hachem et al. [18] showed that a single lower dose of αGalCer (2 µg/mouse) resulted in an inhibition of allergic airway inflammation in an ovalbumin model. More in line with our results is the study by Meyer et al. [39] who showed an increased airway hyper reactivity after two doses of αGalCer (2 times 2 µg/mouse), although the number of iNKT cells were not analyzed in this study and the study by Kim et al. [28] who showed that intranasally application of αGalCer (3 times 1 µg/mouse) worsened allergic asthma in mice.
Although oral sensitization resulted in reduced numbers of iNKT cells in the liver, these cells functionally reacted upon restimulation with αGalCer, since several cytokines were

174
9
iNKT cells contribute to the allergic response in cow’s milk protein sensitized mice
produced, such as IL-4, IFN-γ and IL-10 which are known to contribute to either TH1 skewing or tolerance induction [33, 36]. This increase in cytokine production upon ex vivo αGalCer restimulation is in agreement with multiple other studies [26, 35, 40]. The produced cytokine concentrations in restimulated hepatocyte iNKT cells can be corrected for the percentage of iNKT cells in the liver. Lower frequencies of hepatic iNKT cells in control treated whey-sensitized mice produced the same amounts of cytokines as sham-sensitized mice, whereas equal numbers would have produced higher levels of cytokines. Therefore, whey-sensitized mice produced relatively more IL-4, IL-10 and IFN-γ, whereas the cytokine production per iNKT cell was probably not altered in casein-sensitized mice. The observation that the iNKT cells were depleted by the in vivo αGalCer treatment was in correspondence with the cytokine production of hepatocytes restimulated with αGalCer. An almost complete reduction of all cytokines was found in comparison with control treated mice, indicating iNKT unresponsiveness or absence in association with an increased allergic reaction. αGalCer treatment in particular was found to enhance the acute allergic skin response in whey-sensitized mice. In these mice reduced iNKT numbers in the liver were compensated by enhanced cytokine production per cell. Furthermore, in the spleen and MLN of whey-sensitized mice the iNKT cell numbers were enhanced which was abrogated in the spleen by in vivo αGalCer treatment. This may imply a contribution of iNKT in the suppression of CMA in particular in whey-sensitized mice.
Differences between casein- and whey-sensitized mice in cytokine production by liver iNKT cells and spleen and MLN iNKT frequencies indicate a different role for iNKT in casein and whey allergy in mice. The semi-invariant T-cell receptor of NKT cells in particular recognizes CD1d presented glycolipid antigens presented by antigen presenting cells or intestinal epithelial cells [34]. Casein and whey proteins differ with regard to physical, physiological and dietary properties [41-44]. The three dimensional shape of the proteins differ. Individual caseins (11.6 - 25.2 kDa) are crosslinked and form aggregates, these nanoclusters form large structures called micelles, whereas whey (14.2 - 76.2 kDa) forms linear structures of mostly monomers and dimers. Furthermore, the glycolipid composition of whey and casein may differ. Antigen presentation therefore might be different for either casein or whey affecting iNKT-cell properties.
The dramatic augmentation of the acute allergic skin response in αGalCer treated whey-sensitized mice was found to be associated with significantly enhanced whey-specific IgE and IgG
1 when compared to control treated whey-sensitized mice. In casein-sensitized mice the enhancement of the acute skin response was much less severe and casein-specific immunoglobulins were not further enhanced upon αGalCer treatment. αGalCer treatment also significantly increased TH1 or regulatory T-cell-related immunoglobulin IgG2a levels in whey-sensitized mice and not in casein-sensitized mice. Specific IgA levels remained increased in both casein-sensitized mice as well as whey-sensitized mice in comparison to

175
9
Chapter 9
their sham-sensitized control mice, with no further enhancement after αGalCer treatment. These immunoglobulin data suggest an immunoglobulin dependent mechanism for whey-sensitized mice, whereas for casein-sensitized mice another mechanism may underlie the increase in acute allergic skin response due to in vivo αGalCer treatment. Other studies showed that the acute allergic skin response in casein-sensitized mice shows immunoglobulin free light chain dependency (Schouten et al., submitted). Immunoglobulin free light chains are believed to form an alternative pathway to mast cell degranulation [45-48].
It can be concluded that iNKT cells might play a regulatory role in the onset of CMA in mice. Multiple i.p. injections of αGalCer with weekly intervals depleted hepatic iNKT cells and to a lesser extent spleen and MLN iNKT cells. Depletion of iNKT cells exacerbated the allergic response in mice orally sensitized with cow’s milk proteins, this was most pronounced for whey.
References
1. Schafer, T., et al., Epidemiology of food allergy/food intolerance in adults: associations with
other manifestations of atopy. Allergy, 2001. 56(12): p. 1172-9.
2. Host, A., Frequency of cow’s milk allergy in childhood. Ann Allergy Asthma Immunol, 2002.
89(6 Suppl 1): p. 33-7.
3. Rhodes, H.L., et al., Early life risk factors for adult asthma: a birth cohort study of subjects at
risk. J Allergy Clin Immunol, 2001. 108(5): p. 720-5.
4. Schroeder, A., et al., Food allergy is associated with an increased risk of asthma. Clin Exp
Allergy, 2009. 39(2): p. 261-70.
5. Bahna, S.L., Cow’s milk allergy versus cow milk intolerance. Ann Allergy Asthma Immunol,
2002. 89(6 Suppl 1): p. 56-60.
6. Kalach, N., et al., Intestinal permeability in children: variation with age and reliability in the
diagnosis of cow’s milk allergy. Acta Paediatr, 2001. 90(5): p. 499-504.
7. Tsuji, N.M. and A. Kosaka, Oral tolerance: intestinal homeostasis and antigen-specific
regulatory T cells. Trends Immunol, 2008. 29(11): p. 532-40.
8. Mowat, A.M., Anatomical basis of tolerance and immunity to intestinal antigens. Nat Rev
Immunol, 2003. 3(4): p. 331-41.
9. Kim, H.J., et al., NKT cells play critical roles in the induction of oral tolerance by inducing
regulatory T cells producing IL-10 and transforming growth factor beta, and by clonally
deleting antigen-specific T cells. Immunology, 2006. 118(1): p. 101-11.
10. Delovitch, T.L., The “End of the beginning” of the immunobiology of iNKT cells in health and
disease. Int Rev Immunol, 2007. 26(1-2): p. 1-3.
11. Godfrey, D.I., et al., NKT cells: what’s in a name? Nat Rev Immunol, 2004. 4(3): p. 231-7.

176
9
iNKT cells contribute to the allergic response in cow’s milk protein sensitized mice
12. Kronenberg, M. and L. Gapin, The unconventional lifestyle of NKT cells. Nat Rev Immunol,
2002. 2(8): p. 557-68.
13. Park, J.E., et al., Fine specificity of natural killer T cells against GD3 ganglioside and
identification of GM3 as an inhibitory natural killer T-cell ligand. Immunology, 2008. 123(1):
p. 145-55.
14. Brutkiewicz, R.R., CD1d ligands: the good, the bad, and the ugly. J Immunol, 2006. 177(2): p.
769-75.
15. Gumperz, J.E., et al., Functionally distinct subsets of CD1d-restricted natural killer T cells
revealed by CD1d tetramer staining. J Exp Med, 2002. 195(5): p. 625-36.
16. Oling, V., et al., Circulating CD4+CD25 high regulatory T cells and natural killer T cells in
children with newly diagnosed type 1 diabetes or with diabetes-associated autoantibodies.
Ann N Y Acad Sci, 2007. 1107: p. 363-72.
17. Ikarashi, Y., et al., Phenotypical and functional alterations during the expansion phase
of invariant Valpha14 natural killer T (Valpha14i NKT) cells in mice primed with alpha-
galactosylceramide. Immunology, 2005. 116(1): p. 30-7.
18. Hachem, P., et al., Alpha-galactosylceramide-induced iNKT cells suppress experimental
allergic asthma in sensitized mice: role of IFN-gamma. Eur J Immunol, 2005. 35(10): p. 2793-
802.
19. Burdin, N., et al., Selective ability of mouse CD1 to present glycolipids: alpha-
galactosylceramide specifically stimulates V alpha 14+ NK T lymphocytes. J Immunol, 1998.
161(7): p. 3271-81.
20. Matsuda, H., et al., alpha-Galactosylceramide, a ligand of natural killer T cells, inhibits allergic
airway inflammation. Am J Respir Cell Mol Biol, 2005. 33(1): p. 22-31.
21. Goto, M., et al., Murine NKT cells produce Th17 cytokine interleukin-22. Cell Immunol, 2009.
254(2): p. 81-4.
22. Stronge, V.S., et al., A closer look at CD1d molecules: new horizons in studying NKT cells.
Trends Immunol, 2007. 28(10): p. 455-62.
23. Lehuen, A., et al., Overexpression of natural killer T cells protects Valpha14- Jalpha281
transgenic nonobese diabetic mice against diabetes. J Exp Med, 1998. 188(10): p. 1831-9.
24. Saubermann, L.J., et al., Activation of natural killer T cells by alpha-galactosylceramide in
the presence of CD1d provides protection against colitis in mice. Gastroenterology, 2000.
119(1): p. 119-28.
25. Schmieg, J., et al., Superior protection against malaria and melanoma metastases by a
C-glycoside analogue of the natural killer T cell ligand alpha-Galactosylceramide. J Exp Med,
2003. 198(11): p. 1631-41.
26. Kim, H.Y., et al., The development of airway hyperreactivity in T-bet-deficient mice requires
CD1d-restricted NKT cells. J Immunol, 2009. 182(5): p. 3252-61.
27. Chang, J.H., et al., Functional maturation of lamina propria dendritic cells by activation of
NKT cells mediates the abrogation of oral tolerance. Eur J Immunol, 2008. 38(10): p. 2727-
39.

177
9
Chapter 9
28. Kim, J.O., et al., Asthma is induced by intranasal coadministration of allergen and natural
killer T-cell ligand in a mouse model. J Allergy Clin Immunol, 2004. 114(6): p. 1332-8.
29. Staden, U., et al., Specific oral tolerance induction in food allergy in children: efficacy and
clinical patterns of reaction. Allergy, 2007. 62(11): p. 1261-9.
30. Niederberger, V., Allergen-specific immunotherapy. Immunol Lett, 2009. 122(2): p. 131-3.
31. Skripak, J.M. and H.A. Sampson, Towards a cure for food allergy. Curr Opin Immunol, 2008.
20(6): p. 690-6.
32. Schouten, B., et al., Acute Allergic Skin Reactions and Intestinal Contractility Changes in Mice
Orally Sensitized against Casein or Whey. Int Arch Allergy Immunol, 2008. 147(2): p. 125-
134.
33. Nowak, M. and J. Stein-Streilein, Invariant NKT cells and tolerance. Int Rev Immunol, 2007.
26(1-2): p. 95-119.
34. Zeissig, S., et al., Role of NKT cells in the digestive system. III. Role of NKT cells in intestinal
immunity. Am J Physiol Gastrointest Liver Physiol, 2007. 293(6): p. G1101-5.
35. Akbari, O., et al., Essential role of NKT cells producing IL-4 and IL-13 in the development of
allergen-induced airway hyperreactivity. Nat Med, 2003. 9(5): p. 582-8.
36. Liu, T.Y., et al., Distinct subsets of human invariant NKT cells differentially regulate T helper
responses via dendritic cells. Eur J Immunol, 2008. 38(4): p. 1012-23.
37. Parekh, V.V., et al., Glycolipid antigen induces long-term natural killer T cell anergy in mice. J
Clin Invest, 2005. 115(9): p. 2572-83.
38. Sullivan, B.A. and M. Kronenberg, Activation or anergy: NKT cells are stunned by alpha-
galactosylceramide. J Clin Invest, 2005. 115(9): p. 2328-9.
39. Meyer, E.H., et al., Glycolipid activation of invariant T cell receptor+ NK T cells is sufficient to
induce airway hyperreactivity independent of conventional CD4+ T cells. Proc Natl Acad Sci
U S A, 2006. 103(8): p. 2782-7.
40. Askenase, P.W., et al., TLR-dependent IL-4 production by invariant Valpha14+Jalpha18+ NKT
cells to initiate contact sensitivity in vivo. J Immunol, 2005. 175(10): p. 6390-401.
41. Calbet, J.A. and J.J. Holst, Gastric emptying, gastric secretion and enterogastrone response
after administration of milk proteins or their peptide hydrolysates in humans. Eur J Nutr,
2004. 43(3): p. 127-39.
42. Dangin, M., et al., Influence of the protein digestion rate on protein turnover in young and
elderly subjects. J Nutr, 2002. 132(10): p. 3228S-33S.
43. Hall, W.L., et al., Casein and whey exert different effects on plasma amino acid profiles,
gastrointestinal hormone secretion and appetite. Br J Nutr, 2003. 89(2): p. 239-48.
44. Toden, S., et al., Differential effects of dietary whey, casein and soya on colonic DNA damage
and large bowel SCFA in rats fed diets low and high in resistant starch. Br J Nutr, 2007. 97(3):
p. 535-43.
45. Redegeld, F.A., et al., Immunoglobulin-free light chains elicit immediate hypersensitivity-like
responses. Nat Med, 2002. 8(7): p. 694-701.
46. Redegeld, F.A. and F.P. Nijkamp, Immunoglobulin free light chains and mast cells: pivotal role

178
9
iNKT cells contribute to the allergic response in cow’s milk protein sensitized mice
in T-cell-mediated immune reactions? Trends Immunol, 2003. 24(4): p. 181-5.
47. Groot Kormelink, T., et al., Atopic and non-atopic allergic disorders: current insights into the
possible involvement of free immunoglobulin light chains. Clin Exp Allergy, 2009. 39(1): p.
33-42.
48. Thio, M., et al., Free immunoglobulin light chains: a novel target in the therapy of
inflammatory diseases. Trends Pharmacol Sci, 2008. 29(4): p. 170-4.

179
9
Chapter 9


10
General discussion

182
10
General discussion
Status quo
The incidence of allergic diseases is growing and therefore the need for accurate diagnostics as well as therapeutics increases. Food allergies are diagnosed by e.g. titers of specific IgE using the radioallergosorbent test and/or a positive skin prick test. In addition, the golden standard, a double-blind placebo-controlled oral food challenge can be performed [1-3]. Until now there is no satisfying treatment available and the treatment so far comprises strict allergen avoidance, several forms of immunotherapy and treatment of symptoms by pharmaceuticals.
Allergen avoidance, for cow’s milk allergic patients, can be realized using partial or extensive hydrolysates, or in the case of persistent and severe clinical reactions, an amino acid based formula. Time consuming immunotherapies involve subcutaneous injections of allergen extracts, incrementally increased swallowed allergen (oral immunotherapy) or incrementally increased allergen doses held under the tongue and then swallowed (sublingual immunotherapy) [4-6]. For non-invasive preventive purposes several immune modulatory dietary components are added to infant milk formulas which have been shown to have high potential. One clinical study has shown to reduce the incidence of atopic diseases by using non-digestible oligosaccharides [7]. Clear underlying immunological mechanisms are still unknown [8].
In healthy individuals, harmless food proteins induce oral tolerance. In contrast, in allergic patients a serious effector response is being generated resulting in allergic inflammation upon allergen challenge. The current insights in the mechanisms underlying allergic diseases point to an imbalance of T
H1 and TH2 effector responses, polarizing towards TH2 and too little suppressive capacities of regulatory T-cells (Figure 1). However, it is largely unknown why this imbalance exists and what is needed for the induction of oral tolerance. It has been hypothesized that a lower exposure to microbes might cause hypersensitivity of the immune system to normally harmless agents such as food ingredients. Another risk factor for developing an allergic disease is the genetic predisposition [9]. A child with either a mother or father (or both) with a history of allergic rhinitis, asthma or atopic eczema has a higher risk of developing an allergy than a child without any parental history of allergy.
This thesis provides preclinical data concerning the effectiveness of several dietary immune modulatory components (e.g. prebiotics, probiotics and synbiotics) and, in part, describes their underlying mechanisms of action.

183
10
Chapter 10
Figure 1. Schematic representation of the organization of a humoral immune response by cells of the murine adaptive immune system resulting in mast cell sensitization and degranulation responsible for the allergic symptoms. TH: T helper cell; iTreg: inducible regulatory T-cell; nTreg natural regulatory T-cell; NKT; natural killer T-cell; Tr1: T regulatory type 1 cell.

184
10
General discussion
A mouse model for cow’s milk allergy
To provide in the requirements for preclinical efficacy testing of components that may prevent or treat cow’s milk allergy and to study underlying mechanisms, a reliable model is essential. Therefore we adapted an existing mouse model for orally induced cow’s milk allergy (CMA) and extended the available parameters to measure the allergic outcome [10-12]. In Europe, infant formulas are whey protein based, whereas in the USA these formulas are based on caseins. Oral sensitization of mice using whey or casein in the absence of cholera toxin results in oral tolerance for the given proteins and therefore a mucosal adjuvant is needed to break oral tolerance. Cholera toxin is an enterotoxin produced by Vibrio cholera with strong adjuvant function at the mucosal surface [13]. The T
H2 polarizing effect of cholera toxin is diverse. The most important are the enhanced antigen presentation by various antigen presenting cells and increased permeability of the mucosal epithelium resulting in the production of TH2 related immunoglobulins (IgE and IgG1) and TH2 produced cytokines, i.e. IL-4, IL-5, IL-6 and IL-10 [14].
The whey and casein allergic models were optimized and validated and the acute allergic skin response was used as a sensitive and stable major parameter throughout all the experiments. The mice were sensitized (leading to casein- and whey-specific IgE and IgG1 serum levels) and developed a systemic allergic reaction. To determine the challenge reaction, the acute allergic skin response (ear swelling upon i.d. allergen challenge) was measured as an equivalent of the skin prick test in humans. The acute allergic skin response was significantly enhanced in both casein- as well as whey-sensitized mice if challenged with the specific allergen. However, an important difference between the casein and the whey model was that only in whey-sensitized mice increased levels of specific IgE were demonstrated, whereas both models exhibit increased levels of specific IgG
1. Upon allergen challenge, serum concentrations of mouse mast cell protease-1 were enhanced. In addition, the numbers of mast cells were reduced in the colon of both casein- as well as whey-sensitized mice as demonstrated by histological procedures. These data indicate that the mast cell is a key player in this acute food allergy model. However also other cell types (especially basophils, although also monocytes, eosinophils, Langerhans cells and dendritic cells [15]) display FcεRI and may have a contribution to in the acute allergic effector response as well. However, the precise role of these cells in this model for cow’s milk allergy remains to be analyzed in more detail.
Gastro-intestinal discomfort is one of the most prominent symptoms of CMA, which can lead to abdominal pain, diarrhea or sometimes constipation [16, 17]. In our models we used the labor intensive isometric organ bath technique to investigate contraction of colon segments. Casein-sensitized mice had significantly reduced motility when compared to sham- or whey-

185
10
Chapter 10
sensitized mice. Both casein- as well as whey-sensitized mice had slightly increased water levels in their feces, reflecting diarrhea. Differences between casein- and whey-induced allergy in mice was not only observed in the humoral immune response also functional intestinal differences were found. Casein and whey proteins differ with regard to physical, physiological and dietary properties [18-21]. The three dimensional shape of the proteins differ significantly. Individual caseins (11.6 - 25.2 kDa) are cross-linked and form aggregates. These nanoclusters form large structures called micelles. In contrast, whey (14.2 - 76.2 kDa) form linear structures of mostly monomers and dimers. Antigen presentation by dendritic cells and B-cells might be different for either protein, as whey proteins are smaller when compared to casein micelles affecting the establishment of the adaptive immune response. As a consequence, this may have led to different local intestinal allergic responsiveness resulting in altered contractility in casein-sensitized mice in combination with the specific physicochemical characteristics of the casein or whey proteins.
Focus on discrepancy
The surprising difference in antibody response between the whey and casein model led us to investigate this phenomenon in more detail. Coincidently, part of our group was interested in an alternative pathway of mast cell degranulation via immunoglobulin free light chains (Ig-fLC) [22, 23]. Therefore Ig-fLC levels were determined and blocking experiments were performed using F991, a specific 9 mer peptide Ig-fLC antagonist. In casein-sensitized mice total levels of Ig-fLC were enhanced. Whereas only in the casein-sensitized mice the acute allergic skin response was blocked using F991 (i.v.). Furthermore, to provide evidence that this mechanism was humoral and not cellular, concentrated spleen supernatants were transferred i.v. into naïve recipient mice. Ear swelling, as well as blocking of the ear swelling by F991, was confirmed. The relevance for Ig-fLC in diseases is under investigation. For example, in patients suffering from multiple sclerosis free kappa light chains correlate with disability prognosis [24] and in rheumatoid arthritis, there is a significant correlation between kappa and lambda fLC and the disease activity score [25]. Furthermore, in non-IgE mediated rhinitis patients both lambda and kappa Ig-fLC levels are increased in nasal secretions (Powe et al., JACI in press). Notably, children with severe AD have higher levels of lambda and kappa light chain in serum [26]. Plasma samples from infants at high risk for allergy who participated in a prebiotic intervention study were measured for both kappa and lambda Ig-fLC levels. The infants supplemented with the specific short-chain galacto-oligosaccharide (scGOS) and long-chain fructo-oligosaccharide (lcFOS) prebiotic mixture (Immunofortis®) had a reduced incidence of atopic dermatitis and infections when compared to the control group [7, 27, 28]. In line with our preclinical findings both kappa and lambda concentrations were diminished in infants with AD in comparison with infants without AD (Schouten et

186
10
General discussion
al., submitted). The groups were heterogeneous, as the inclusion criteria were a parental history of atopic eczema, allergic rhinitis or asthma. Therefore more clinical study data with cow’s milk allergic infants or other forms of allergies are necessary to further confirm these findings and are currently under analysis.
Biotics
After establishing the murine model of CMA, dietary intervention studies were conducted to determine the preventive effects of pre-, pro-, and synbiotics on the allergic outcome. Two weeks prior the first sensitization mice were fed the specific prebiotic diet (Immunofortis®), the probiotic diet (Bifidobacterium breve M-16V) or the synbiotic diet (the combination of both) (Figure 2). A preventive instead of a treatment set up was chosen to obtain maximum efficacy. As hypothesized, all three diets were capable, in some extent, to prevent the allergic outcome. However, the synbiotic diet showed the best results. The acute allergic skin response and anaphylaxis were almost completely prevented and mouse mast cell protease levels were lower as compared to control mice. Nevertheless, sensitization could not be prevented as both specific IgE and specific IgG
1 serum levels remained elevated, although both specific-fecal IgA and specific-serum IgG2a levels, indicative of a regulatory and/or TH1 type immune responses, were enhanced, which might indicate an improved TH1/TH2 balance and/or Treg function. In addition, it was shown that supplementation of UV-killed bacteria of the same strain had similar effects (Schouten et al., unpublished data). These findings are in accordance with the outcome of clinical studies using pre- and/or probiotics for the prevention of atopic disease [29-32]. When pregnant mothers, with a family history of allergic disease, and their babies were supplemented with Lactobacillus GG the cumulative incidence of atopic eczema was still reduced after 7 years by 30% when compared to placebo controls [29]. Although the incidence of eczema was reduced, no effects were found on IgE levels. In a similar study Lactobacillus reuteri reduced the incidence of IgE-associated eczema and the occurrence of a positive skin prick test for almost 50%, whereas no effect on IgE was observed [30]. On the other hand, several studies show little effect of dietary intervention with pre-, pro- or synbiotics [32, 33]. Generally, various factors influence the outcome of these studies, especially dietary composition, duration of treatment, differences in probiotic (e.g. single strain or mixtures) that have been used and doses play an important role. This is one of the first studies that show additional effects of the combination of prebiotics (scGOS/lcFOS) and probiotics (B. breve M-16V). Ogawa et al. [34] showed additional effects of Lactobacillus casei together with dextran in the clinical skin severity scores in a mouse model. In contrast, Passeron et al. [35] did not find an additional effect of synbiotics over prebiotics in children with AD. Taken together, the pre-, pro- and in particular the synbiotic diets effectively reduced the allergic outcome of mice sensitized

187
10
Chapter 10
with whey proteins. However, underlying mechanisms (e.g. production of specific IgA and IgG2a) are still largely unknown and the remaining studies in this thesis focused on the role of the CD25+ regulatory T (Treg)-cell.
Three of a kind
Besides scGOS and lcFOS another potent prebiotic, a pectin-derived acidic oligosaccharide (pAOS), was manufactured in order to represent the acidic oligosaccharide fraction which is present in human breast milk. Conjugation of acidic oligosaccharides with bovine β-lactoglobulin showed less allergenicity in both in vitro experiments and after i.p. immunization in mice [36, 37]. Furthermore, in a vaccination model and in a model for allergic asthma the combination of scGOS, lcFOS and pAOS suppressed allergy in mice [38, 39]. In addition, the combination of the three non-digestible carbohydrates were well tolerated and did not result in differences of growth, crying, vomiting and regurgitation pattern when added to milk formula as compared to control formula nourished infants [40, 41].
In both casein- and whey-sensitized mice the scGOS/lcFOS/pAOS mixture effectively prevented the acute allergic skin response. However, sensitization was not prevented and both specific-IgE and IgG
1 levels remained enhanced, although whey-specific immunoglobulins were slightly lower when compared to control fed mice. The discrepancy between the observed clinical symptoms (acute allergic skin response) and antigen-specific serum levels is difficult to explain. In chapter 3, a correlation between IgE and/or IgG1 and the acute allergic skin response was shown in whey- sensitized mice fed control diet. However, after dietary intervention this correlation was lost and the symptoms were reduced whereas the specific
Figure 2. A schematic overview of the dietary intervention, sensitization and challenge protocol and the main parameters that are analyzed.
0 7 33282114 34‐14 ‐7
Control or intervention diet
Sensitization by gavage with whey + CT or CT alone (control)
serum
ear swellinganaphylaxisFACS
days

188
10
General discussion
immunoglobulin levels remain elevated. In human studies, without any dietary intervention, the correlation between high specific IgE levels and clinical reactivity is complex as well and depends e.g. on the age of the patient [42-44]. This makes reliable diagnosis very difficult. Furthermore, desensitization is associated with elevated levels of specific IgG4. It is known that inhibition of mast cell activation occurs after coengagement of IgG and IgE receptors and therefore, at least, partly prevents mast cell degranulation [45, 46].
Regulatory affairs
The preventive capacities of scGOS/lcFOS/pAOS on the clinical outcome of disease was demonstrated at least in several mouse models for allergy. However, the mechanism of action was still unidentified. In 2007 Van Wijk et al. [47] showed that CD4+CD25+ Treg cells regulate the intensity of a hypersensitivity response in a murine model of peanut allergy. To investigate this Van Wijk et al. used a rat anti-CD25 monoclonal antibody (mAb; PC61) for in vivo depletion of the Treg, which resulted in an exacerbation of the allergic response to peanut. Although, several other studies showed that the use of PC61 is effective enough and can be used to investigate Treg cell function [48-51]. Recent studies showed that the combination of both PC61 and another anti-CD25 mAb 7D4 was more effective in depletion or inactivation of Treg cells [52]. Furthermore, debate is ongoing whether using either of the mAbs depletes or inactivates only the Treg cells.
Also in CMA clinically reactive patients, functional CD4+CD25+ Treg cells were found to regulate the allergen specific response since in vitro Treg depletion was shown to increase allergen specific T-cell proliferation [53]. In addition, children who outgrew their allergy had higher frequencies of Treg cells in circulation, whereas (again) depletion of Treg cells resulted in a fivefold induced proliferation response ex vivo [54]. Therefore we hypothesized that the preventive effect of the scGOS/lcFOS/pAOS diet on the acute allergic skin response to cow’s milk proteins could be due to improved Treg cell function (Figure 1).
To test this hypothesis, mice were injected i.p. with αCD25 mAb 1 day before the first oral sham- or casein-sensitization (which was 13 days after starting the scGOS/lcFOS/pAOS diet) and the Treg cell depletion was repeated 1 day before the 3th oral sensitization. One week after the 5th and last oral sensitization the acute allergic skin response was measured. In mice fed the scGOS/lcFOS/pAOS diet the acute allergic skin response was reduced, whereas in vivo Treg depletion abolished this protective effect, although immunoglobulin levels were increased as well in the Treg depleted mice. In vivo anti-CD25 mAb treatment may have enhanced the capacity of B-cells to produce antibodies since regulation by Treg cells was disrupted. The increased antibody levels upon in vivo anti-CD25 mAb treatment could

189
10
Chapter 10
have resulted in an enhanced acute skin response or the depletion of Treg as such indeed effectively abrogated the protective effect of the diet. To pin down the Treg cells as crucial cell type by which the scGOS/lcFOS/pAOS diet mechanistically acts, adoptive transfer studies were performed. After transfer of splenocytes of sensitized mice fed the scGOS/lcFOS/pAOS diet into naïve recipient mice fed the control diet and thereafter sensitized to casein or whey, the acute allergic skin response was still effectively suppressed upon casein or whey challenge. In addition, the allergic memory of sensitized mice fed the control diet was transferred as well. These experiments showed that a cellular mechanism was involved and that using splenocytes for adoptive transfer is adequate. Splenocytes were already shown to be effective in transferring immunological memory for a vaccine [55]. Furthermore, our experiments revealed that this prevention was antigen specific, since splenocytes of sham-sensitized mice fed the scGOS/lcFOS/pAOS diet did not prevent the acute allergic skin response in the recipient mice. There is growing evidence, in particular in transplantation biology, that although few in numbers, antigen-specific Treg cells can suppress both T
H1 and TH2 [56, 57].
The following step was to repeat the experiments; although before the transfer, splenocytes were depleted for Treg cells, using PC61. Recipient mice receiving splenocytes from scGOS/lcFOS/pAOS fed donor mice depleted for Treg cells were no longer able to protect against the induction of the acute allergic skin response after intra dermal allergen challenge. Both casein- and whey-sensitized mice showed comparable results with regard to adoptive transfer experiments and involvement of Treg cells, although some slight differences were observed. In whey-sensitized recipients, transferred with ex vivo Treg depleted splenocytes of whey-sensitized mice, an increased T
H1 and TH2 immunoglobulins were observed, whereas casein-sensitized mice did not show this increase. This effect is indifferent of the diet the donors received, once more suggesting differences in the adaptive immune response developed as a consequence of sensitization with either casein or whey. Another mechanism of, probably, B-cell activation or switch takes place, explaining the differences observed in sensitization, although the net overall effect, the acute allergic skin response, was similar. In this set of experiments transfer of the peripheral immune cells (e.g. splenocytes) showed that the effect of the scGOS/lcFOS/pAOS diet acts systemically. However, currently it is unknown whether local immune cells (e.g. MLN or Peyer’s patches) have the same capabilities. In these organs oral tolerance is induced and can be transferred to the systemic compartment via the MLN involving Treg cells [58]. In a murine ovalbumin model MLN cells were able to transfer allergy to naïve mice and diarrhea developed upon oral ovalbumin challenge [59]. In addition, transfer of MLN or Peyer’s patches cell suspensions of mice orally sensitized to β-lactoglobulin into naïve mice resulted in an increased production of specific IgG
1, indicative for transfer of sensitization [60]. Furthermore, MLN cells of ovalbumin tolerized mice could transfer oral tolerance in naïve mice [61]. These experiments suggest a crucial contribution for local lymphoid organs in the onset and regulation of allergy and tolerance.

190
10
General discussion
Additional experiments are needed to pinpoint the specific T cell subset in the regulation of oral tolerance in cow’s milk allergy.
Lethal regulation
Invariant Natural Killer T (iNKT) cells form another subset of cells with regulatory properties and might be involved in homeostasis of the mucosal immune system. In mice these cells are present at low frequencies in the spleen, MLN and Peyer’s patches (<1.0%) [62] (Schouten et al., submitted). However, hepatic lymphocytes consist of approximately 20% iNKT cells and therefore the liver was chosen to study iNKT cells. The best characterized and potent specific iNKT cell agonist is a glycolipid isolated from the marine sponge Agelas mauritianus and is called α-galactosylceramide (αGalCer) [63-65]. We found that after oral sensitization with either whey or casein the percentage of hepatic iNKT cells, using flow cytometry, was decreased. Therefore it was hypothesized that after stimulation of these cells the severity of the acute allergic skin reaction might be reduced. However, stimulation of the iNKT cells with αGalCer (2 µg/mouse) 1 h before oral sensitization resulted, unexpectedly, in an increased acute allergic skin response and a much further decrease in hepatic iNKT counts, in control mice as well. We speculated on a migration of these cells towards the other lymphoid organs, although this was not confirmed by FACS analyses, determining iNKT cell numbers in Peyer’s patches and MLN, and gave rise to in the interpretation that the iNKT cells were depleted instead of stimulated. This interpretation was supported after analyzing cytokine production of hepatocytes restimulated with αGalCer. Hepatocytes from mice treated with αGalCer showed almost a complete reduction of cytokine profiles in comparison with control treated mice. Parekh et al. [66] demonstrated that a single high dose administration of αGalCer (5 µg/mouse) causes anergy of iNKT cells for at least one month. In contrast, Matsuda et al. [65] as well as Hachem et al. [63] showed that a single lower dose of αGalCer (2 µg/mouse) resulted in an inhibition of allergic airway inflammation in an ovalbumin model. More in line with our results is the study by Meyer et al. who showed that the airway hyper reactivity increased after two doses of αGalCer (2 times 2 µg/mouse), although the number of iNKT cells were not analyzed in this study.
iNKT cells might play a role in the onset of CMA in mice, although further studies are needed to confirm these findings. A single low dose before the first sensitization might stimulate the iNKT cells effectively and might partially prevent the development of an acute allergic skin response. In addition, the contribution of iNKT cells in the immune modulatory effect of dietary components like scGOS/lcFOS/pAOS requires further investigation.
It is difficult to combine our research on Treg cells and iNKT cells. However, Kim et al. [67]

191
10
Chapter 10
showed that iNKT cells play a pivotal role in oral tolerance by inducing Treg cells and in addition by clonally deleting antigen-specific CD4+ T cells. Therefore it can be argued that, in line with our results, both Treg cells and iNKT cells are involved in the regulation of CMA in mice. However, studies which investigate both the role of Treg cells as well as iNKT cells in CMA are needed to further assess this regulatory mechanism.
To conclude the discussion
The studies performed in this thesis have resulted in a better understanding of the immunological mechanisms underlying the pathophysiology in mice orally sensitized with caseins or whey proteins. It was found that sensitization with cow’s milk proteins can lead to both IgE- and Ig-fLC-dependent allergic hypersensitivity responses. Immune modulation by dietary intervention with probiotics, synbiotics, and in particular the prebiotic scGOS/lcFOS/pAOS diet was investigated and small parts of their working mechanisms are unraveled. Treg cells have been shown to contribute to the immune modulatory effects of prebiotics. These results should be further confirmed in clinical studies which are currently being performed.
References
1. Monaci, L., et al., Milk allergens, their characteristics and their detection in food: a review.
European Food Research and Technology, 2005(Volume 223, Number 2 / June, 2006): p.
149-179.
2. Pourpak, Z., et al., The role of cow milk allergy in increasing the severity of atopic dermatitis.
Immunol Invest, 2004. 33(1): p. 69-79.
3. Sicherer, S.H. and H.A. Sampson, Cow’s milk protein-specific IgE concentrations in two age
groups of milk-allergic children and in children achieving clinical tolerance. Clin Exp Allergy,
1999. 29(4): p. 507-12.
4. Staden, U., et al., Specific oral tolerance induction in food allergy in children: efficacy and
clinical patterns of reaction. Allergy, 2007. 62(11): p. 1261-9.
5. Niederberger, V., Allergen-specific immunotherapy. Immunol Lett, 2008.
6. Skripak, J.M. and H.A. Sampson, Towards a cure for food allergy. Curr Opin Immunol, 2008.
20(6): p. 690-6.
7. Moro, G., et al., A mixture of prebiotic oligosaccharides reduces the incidence of atopic
dermatitis during the first six months of age. Arch Dis Child, 2006. 91(10): p. 814-9.
8. Boehm, G. and G. Moro, Structural and functional aspects of prebiotics used in infant
nutrition. J Nutr, 2008. 138(9): p. 1818S-1828S.

192
10
General discussion
9. Bjorksten, B., Genetic and environmental risk factors for the development of food allergy.
Curr Opin Allergy Clin Immunol, 2005. 5(3): p. 249-53.
10. Schouten, B., et al., Acute Allergic Skin Reactions and Intestinal Contractility Changes in Mice
Orally Sensitized against Casein or Whey. Int Arch Allergy Immunol, 2008. 147(2): p. 125-
134.
11. Frossard, C.P., C. Hauser, and P.A. Eigenmann, Antigen-specific secretory IgA antibodies in
the gut are decreased in a mouse model of food allergy. J Allergy Clin Immunol, 2004. 114(2):
p. 377-82.
12. Li, X.M., et al., A murine model of IgE-mediated cow’s milk hypersensitivity. J Allergy Clin
Immunol, 1999. 103(2 Pt 1): p. 206-14.
13. Lycke, N., The mechanism of cholera toxin adjuvanticity. Res Immunol, 1997. 148(8-9): p.
504-20.
14. Holmgren, J., et al., Mucosal adjuvants and anti-infection and anti-immunopathology
vaccines based on cholera toxin, cholera toxin B subunit and CpG DNA. Immunol Lett, 2005.
97(2): p. 181-8.
15. Kinet, J.P., The high-affinity IgE receptor (Fc epsilon RI): from physiology to pathology. Annu
Rev Immunol, 1999. 17: p. 931-72.
16. Furrie, E., Probiotics and allergy. Proc Nutr Soc, 2005. 64(4): p. 465-9.
17. Kalach, N., et al., Intestinal permeability in children: variation with age and reliability in the
diagnosis of cow’s milk allergy. Acta Paediatr, 2001. 90(5): p. 499-504.
18. Calbet, J.A. and J.J. Holst, Gastric emptying, gastric secretion and enterogastrone response
after administration of milk proteins or their peptide hydrolysates in humans. Eur J Nutr,
2004. 43(3): p. 127-39.
19. Dangin, M., et al., Influence of the protein digestion rate on protein turnover in young and
elderly subjects. J Nutr, 2002. 132(10): p. 3228S-33S.
20. Hall, W.L., et al., Casein and whey exert different effects on plasma amino acid profiles,
gastrointestinal hormone secretion and appetite. Br J Nutr, 2003. 89(2): p. 239-48.
21. Toden, S., et al., Differential effects of dietary whey, casein and soya on colonic DNA damage
and large bowel SCFA in rats fed diets low and high in resistant starch. Br J Nutr, 2007. 97(3):
p. 535-43.
22. Redegeld, F.A., et al., Immunoglobulin-free light chains elicit immediate hypersensitivity-like
responses. Nat Med, 2002. 8(7): p. 694-701.
23. Kraneveld, A.D., et al., Elicitation of allergic asthma by immunoglobulin free light chains.
Proc Natl Acad Sci U S A, 2005. 102(5): p. 1578-83.
24. Rinker, J.R., 2nd, K. Trinkaus, and A.H. Cross, Elevated CSF free kappa light chains correlate
with disability prognosis in multiple sclerosis. Neurology, 2006. 67(7): p. 1288-90.
25. Gottenberg, J.E., et al., Serum immunoglobulin free light chain assessment in rheumatoid
arthritis and primary Sjogren’s syndrome. Ann Rheum Dis, 2007. 66(1): p. 23-7.
26. Sediva, A., J. Kayserova, and E. Vernerova, Increase of serum immunoglobulin free light
chains in children with atopic dermatitis. Allergy, 2008. 63: p. 157.

193
10
Chapter 10
27. Arslanoglu, S., G.E. Moro, and G. Boehm, Early supplementation of prebiotic oligosaccharides
protects formula-fed infants against infections during the first 6 months of life. J Nutr, 2007.
137(11): p. 2420-4.
28. Arslanoglu, S., et al., Early dietary intervention with a mixture of prebiotic oligosaccharides
reduces the incidence of allergic manifestations and infections during the first two years of
life. J Nutr, 2008. 138(6): p. 1091-5.
29. Kalliomaki, M., et al., Probiotics during the first 7 years of life: a cumulative risk reduction of
eczema in a randomized, placebo-controlled trial. J Allergy Clin Immunol, 2007. 119(4): p.
1019-21.
30. Abrahamsson, T.R., et al., Probiotics in prevention of IgE-associated eczema: a double-blind,
randomized, placebo-controlled trial. J Allergy Clin Immunol, 2007. 119(5): p. 1174-80.
31. Rosenfeldt, V., et al., Effect of probiotic Lactobacillus strains in children with atopic dermatitis.
J Allergy Clin Immunol, 2003. 111(2): p. 389-95.
32. Kukkonen, K., et al., Probiotics and prebiotic galacto-oligosaccharides in the prevention
of allergic diseases: A randomized, double-blind, placebo-controlled trial. J Allergy Clin
Immunol, 2007. 119(1): p. 192-8.
33. Sistek, D., et al., Is the effect of probiotics on atopic dermatitis confined to food sensitized
children? Clin Exp Allergy, 2006. 36(5): p. 629-33.
34. Ogawa, T., et al., A new synbiotic, Lactobacillus casei subsp. casei together with dextran,
reduces murine and human allergic reaction. FEMS Immunol Med Microbiol, 2006. 46(3): p.
400-9.
35. Passeron, T., et al., Prebiotics and synbiotics: two promising approaches for the treatment of
atopic dermatitis in children above 2 years. Allergy, 2006. 61(4): p. 431-7.
36. Yoshida, T., et al., Reduced T cell response to beta-lactoglobulin by conjugation with acidic
oligosaccharides. J Agric Food Chem, 2005. 53(17): p. 6851-7.
37. Hattori, M., et al., Reduced immunogenicity of beta-lactoglobulin by conjugation with acidic
oligosaccharides. J Agric Food Chem, 2004. 52(14): p. 4546-53.
38. Vos, A.P., et al., Dietary supplementation of neutral and acidic oligosaccharides enhances
Th1-dependent vaccination responses in mice. Pediatr Allergy Immunol, 2007. 18(4): p. 304-
12.
39. Vos, A.P., et al., Dietary supplementation with specific oligosaccharide mixtures decreases
parameters of allergic asthma in mice. Int Immunopharmacol, 2007. 7(12): p. 1582-7.
40. Fanaro, S., et al., Acidic oligosaccharides from pectin hydrolysate as new component for infant
formulae: effect on intestinal flora, stool characteristics, and pH. J Pediatr Gastroenterol
Nutr, 2005. 41(2): p. 186-90.
41. Fanaro, S., et al., Galacto-oligosaccharides are bifidogenic and safe at weaning: a double-
blind randomized multicenter study. J Pediatr Gastroenterol Nutr, 2009. 48(1): p. 82-8.
42. Celik-Bilgili, S., et al., The predictive value of specific immunoglobulin E levels in serum for
the outcome of oral food challenges. Clin Exp Allergy, 2005. 35(3): p. 268-73.
43. van der Gugten, A.C., et al., Usefulness of specific IgE levels in predicting cow’s milk allergy.

194
10
General discussion
J Allergy Clin Immunol, 2008. 121(2): p. 531-3.
44. Garcia-Ara, C., et al., Specific IgE levels in the diagnosis of immediate hypersensitivity to
cows’ milk protein in the infant. J Allergy Clin Immunol, 2001. 107(1): p. 185-90.
45. Ruiter, B., et al., Maintenance of tolerance to cow’s milk in atopic individuals is characterized
by high levels of specific immunoglobulin G4. Clin Exp Allergy, 2007. 37(7): p. 1103-10.
46. Malbec, O. and M. Daeron, The mast cell IgG receptors and their roles in tissue inflammation.
Immunol Rev, 2007. 217: p. 206-21.
47. van Wijk, F., et al., CD4+CD25+ T cells regulate the intensity of hypersensitivity responses
to peanut, but are not decisive in the induction of oral sensitization. Clin Exp Allergy, 2007.
37(4): p. 572-81.
48. Shreffler, W.G., et al., Association of allergen-specific regulatory T cells with the onset of
clinical tolerance to milk protein. J Allergy Clin Immunol, 2009. 123(1): p. 43-52 e7.
49. Kohm, A.P., et al., Cutting Edge: Anti-CD25 monoclonal antibody injection results in the
functional inactivation, not depletion, of CD4+CD25+ T regulatory cells. J Immunol, 2006.
176(6): p. 3301-5.
50. Lewkowich, I.P., et al., CD4+CD25+ T cells protect against experimentally induced asthma
and alter pulmonary dendritic cell phenotype and function. J Exp Med, 2005. 202(11): p.
1549-61.
51. Matsushita, N., et al., Comparative methodologies of regulatory T cell depletion in a murine
melanoma model. J Immunol Methods, 2008. 333(1-2): p. 167-79.
52. Couper, K.N., et al., Incomplete depletion and rapid regeneration of Foxp3+ regulatory T cells
following anti-CD25 treatment in malaria-infected mice. J Immunol, 2007. 178(7): p. 4136-
46.
53. Sletten, G.B., et al., Memory T Cell Proliferation in Cow’s Milk Allergy after CD25+ Regulatory
T Cell Removal Suggests a Role for Casein-Specific Cellular Immunity in IgE-Mediated but
Not in Non-IgE-Mediated Cow’s Milk Allergy. Int Arch Allergy Immunol, 2006. 142(3): p. 190-
198.
54. Karlsson, M.R., J. Rugtveit, and P. Brandtzaeg, Allergen-responsive CD4+CD25+ regulatory T
cells in children who have outgrown cow’s milk allergy. J Exp Med, 2004. 199(12): p. 1679-
88.
55. Guttormsen, H.K., et al., Immunologic memory induced by a glycoconjugate vaccine in a
murine adoptive lymphocyte transfer model. Infect Immun, 1998. 66(5): p. 2026-32.
56. Koenen, H.J. and I. Joosten, Antigen-specific regulatory T-cell subsets in transplantation
tolerance regulatory T-cell subset quality reduces the need for quantity. Hum Immunol,
2006. 67(9): p. 665-75.
57. Zhang, Z.X., et al., Identification of a previously unknown antigen-specific regulatory T cell
and its mechanism of suppression. Nat Med, 2000. 6(7): p. 782-9.
58. Tsuji, N.M. and A. Kosaka, Oral tolerance: intestinal homeostasis and antigen-specific
regulatory T cells. Trends Immunol, 2008. 29(11): p. 532-40.
59. Knight, A.K., et al., CD4 T cells activated in the mesenteric lymph node mediate gastrointestinal

195
10
Chapter 10
food allergy in mice. Am J Physiol Gastrointest Liver Physiol, 2007. 293(6): p. G1234-43.
60. Frossard, C.P., et al., Lymphocytes in Peyer patches regulate clinical tolerance in a murine
model of food allergy. J Allergy Clin Immunol, 2004. 113(5): p. 958-64.
61. Hauet-Broere, F., et al., Functional CD25- and CD25+ mucosal regulatory T cells are induced
in gut-draining lymphoid tissue within 48 h after oral antigen application. Eur J Immunol,
2003. 33(10): p. 2801-10.
62. Ikarashi, Y., et al., Phenotypical and functional alterations during the expansion phase
of invariant Valpha14 natural killer T (Valpha14i NKT) cells in mice primed with alpha-
galactosylceramide. Immunology, 2005. 116(1): p. 30-7.
63. Hachem, P., et al., Alpha-galactosylceramide-induced iNKT cells suppress experimental
allergic asthma in sensitized mice: role of IFN-gamma. Eur J Immunol, 2005. 35(10): p. 2793-
802.
64. Burdin, N., et al., Selective ability of mouse CD1 to present glycolipids: alpha-
galactosylceramide specifically stimulates V alpha 14+ NK T lymphocytes. J Immunol, 1998.
161(7): p. 3271-81.
65. Matsuda, H., et al., alpha-Galactosylceramide, a ligand of natural killer T cells, inhibits allergic
airway inflammation. Am J Respir Cell Mol Biol, 2005. 33(1): p. 22-31.
66. Parekh, V.V., et al., Glycolipid antigen induces long-term natural killer T cell anergy in mice. J
Clin Invest, 2005. 115(9): p. 2572-83.
67. Kim, H.J., et al., NKT cells play critical roles in the induction of oral tolerance by inducing
regulatory T cells producing IL-10 and transforming growth factor beta, and by clonally
deleting antigen-specific T cells. Immunology, 2006. 118(1): p. 101-11.


Miscellaneous
SummaryNederlandse samenvatting
DankwoordCurriculum Vitae
List of publicationsAbbreviations

198
M
Summary
Summary
The prevalence of allergic diseases is rising to almost 20%. This includes one percent of the population affected with an adverse immune reaction to food components. In developed countries 2-4% and in some countries even higher percentages of infants exhibit cow’s milk allergy (CMA). Although most infants outgrow CMA before their fifth year, immunoglobulin (Ig)E-mediated CMA predisposes the development of other (food) allergies and even asthma later in life. Furthermore CMA is one of the leading causes of food allergy in adults. Approximately 0.1-0.5% have clinical symptoms after ingestion of milk containing products. Clinical symptoms may involve the skin, respiratory tract and gastrointestinal tract, and can even lead to a systemic anaphylactic reaction. Cow’s milk contains two main protein classes, which are the caseins (30 g/L) and whey proteins (5 g/L). The caseins consist mainly of αS1-, αS2-, κ- and β-casein, whereas whey proteins comprise of β-lactoglobulin, α-lactalbumin, bovine serum albumin, serum immunoglobulins and lactoferrin. Large population studies with cow’s milk allergic infants have shown that the major allergens are β-lactoglobulin, αS1-casein and α-lactalbumin. Animal models of CMA provide a tool to reveal mechanisms involved in the pathophysiology of CMA and to explore new preventive and/or therapeutic approaches. This thesis focuses on the use of dietary intervention with pre-, pro- or synbiotics to prevent the development of cow’s milk allergy in mice. In most of the animal models currently available to study food allergy, sensitization is not performed via the oral route but via intra peritoneal injection of the allergen. In reality humans are mostly sensitized upon ingestion of the inflicting food. In the current thesis a mouse model for orally induced CMA was introduced using whey or casein proteins. Furthermore, new tools were developed to address pathological changes in the casein and whey allergic mice. Measurement of the acute allergic skin response, as an equivalent of the human skin prick test, was introduced to reflect systemic allergy as one of the consequences of oral sensitization (chapter 3).
Striking differences were observed comparing the sensitizing capacities of whey and casein (chapter 3). These proteins have different characteristics e.g. their three-dimensional structures, whey proteins are linear and casein proteins are globular. Both whey and casein allergic mice induced systemic acute allergic symptoms as reflected by an increase in ear swelling upon intradermal challenge with the specific antigen. However in casein allergic mice specific IgE titers were not detectable. In addition, only in casein sensitized mice intestinal contractility changes occurred, reflecting local pathophysiological functional changes. Since casein sensitized mice appeared to have allergic symptoms in the absence of casein specific IgE it was speculated that another mechanisms might be involved in the onset of the acute allergic response, e.g. immunoglobulin free light chain (Ig-fLC). Ig-fLC is produced by B cells and it has been shown to induce an acute allergic response. Indeed, using a specific Ig-fLC antagonist (F991) a complete reduction of the allergic skin response

199
M
Miscellaneous
was established in the casein sensitized mice only. This was confirmed in a passive transfer study in which spleen supernatant was injected into naïve recipient mice. F991 protected these mice from the development of an acute allergic skin response, when challenged in the ear with casein (chapter 4).
To confirm the relevance for Ig-fLC in our preclinical model, sera from a cohort of term born infants at risk for allergy were analyzed for Ig-fLC. A difference in Ig-fLC concentrations was found between infants that developed atopic dermatitis (AD) after six months when compared to infants that remained unaffected thus with no detectable AD (chapter 4). The infants of this cohort received a control milk formula or Immunofortis® (for details see next paragraph) supplemented milk formula. The addition of the Immunofortis® mixture was found to lower the incidence of AD by 50%. In the sera of these Immunofortis® treated infants Ig-fLC concentrations were found to be significantly reduced which may relate to the lower incidence of AD (chapter 5).
The clinical study, as described partially in chapter 4 and 5, showed a clear preventive effect of these specific prebiotics. In other human as well as animal studies pre-, pro- and synbiotics have been shown to be effective in the reduction of allergic disorders as well. In order to investigate the underlying mechanisms of action preventive intervention studies were carried out using the established mouse model for CMA. In order to investigate the preventive effects mice were fed a control or short-chain galacto-oligosaccharides (scGOS) and long-chain fructo-oligosaccharides (lcFOS) containing diet during oral whey-sensitization. This scGOS/lcFOS (ratio 9:1) mixture (Immunofortis®) contains non-digestible carbohydrates that selectively stimulate the growth of bacteria that support the health of the host. This prebiotic mixture appeared to inhibit the allergic symptoms highly significantly. If probiotic bacteria were added allergic symptoms were downregulated as well. The best preventive concept, however, was a mixture of the prebiotic oligosaccharides and a probiotic Bifidobacterium breve M-16V strain. This latter mixture is called a synbiotic. In sum, dietary intervention with the pre-, pro- and in particular the synbiotics was found to diminish the acute allergic skin response and anaphylactic scores upon allergen challenge. However, no reduction in specific IgE and IgG
1 levels was observed and the number of regulatory T-cells (CD4+CD25+FoxP3+) in the spleen, mesenteric lymph nodes and Peyer’s patches was unaltered. IgG2a, indicative for a regulatory T-cell or TH1 response, was affected positively by the different diets (chapter 6).
In order to examine the role of regulatory T-cells in the prebiotic effects the functional contribution of regulatory T-cells using in vivo regulatory T-cell depletion and adoptive transfer experiments were performed. For this purpose pectin-derived acidic-oligosaccharides (pAOS), a third group of non-digestible carbohydrates simulating the acidic oligosaccharide structures present in breast milk, were added to the scGOS/lcFOS mixture.

200
M
Summary
Both whey- and casein-sensitization was performed using dietary intervention with a scGOS/lcFOS/pAOS (ratio 9:1:1) mixture. The outcome was comparable to the effect of scGOS/lcFOS. Dietary intervention was found to reduce the acute allergic skin response in whey- as well as casein-sensitized mice, however there was still no significant effect on the serological parameters and the number of regulatory T-cells. Nevertheless it was hypothesized that the regulatory T-cells played an important functional role in the allergic outcome. Therefore a different set of experiments was initiated using a monoclonal antibody to deplete the regulatory T-cells in vivo (PC61, an anti-CD25 monoclonal antibody). From these studies it was concluded that regulatory T-cells played a crucial role in the preventive effects of scGOS/lcFOS/pAOS (chapter 7 and 8).
To add additional evidence regarding the involvement of the regulatory T-cells a set of adoptive transfer experiments was initiated. Single cell suspensions of spleens from allergic (either whey or casein) or non-allergic mice fed a control or prebiotic diet were transferred to naïve recipient mice. In addition, splenocytes were transferred which were ex vivo depleted for CD25+ regulatory T-cells using magnetic beads. Using these experiments it was shown that the dietary induced inhibition of allergy could be transferred, while selective depletion of CD4+CD25+Foxp3+ regulatory T-cells abrogated this protective effect. Furthermore these results showed that the induction of tolerance by the prebiotic diet was antigen specific, since single cell suspensions of control sensitized mice fed the prebiotic diet did not protect recipient mice from developing allergic symptoms (chapter 7 and 8).
Another potentially important regulatory cell type in allergy is the invariant Natural Killer T (iNKT)-cell. Initial experiments showed that the number of iNKT-cells in the liver of casein- as well as whey-sensitized mice was diminished. Therefore we hypothesized that after stimulation with α-galactosylceramide (αGalCer), a specific iNKT-cell agonist, the number of iNKT-cells would be restored or at least their activity would increase, protecting the mice from developing CMA. However, the acute allergic skin response was worsened instead of improved, which was most pronounced in whey sensitized mice. In addition, whey-specific IgE, IgG
1 and IgG2a were enhanced as well, while in casein-sensitized mice αGalCer treatment did not alter casein-specific immunoglobulin levels. Flow cytometric analysis revealed an almost complete decrease in number of iNKT-cells. In addition, hepatocyte suspensions of αGalCer treated mice failed to produce cytokines upon ex vivo restimulation. Therefore we hypothesized that over-stimulation of the iNKT-cell population resulted in a depletion instead of activation of these cells. Hence data presented in this chapter suggest iNKT-cells to have immunomodulatory capacities, in particular relevant in mice orally sensitized with whey (chapter 9).
In this thesis a new murine model for whey as well as casein has been described and differences in underlying mechanism of action between the two allergenic proteins were

201
M
Miscellaneous
revealed. Different dietary interventions showed preventive effects in the onset of CMA. The synbiotic diet was most effective. The induction of protein specific regulatory T-cells was revealed to be part of the immunological mechanism involved in both whey- and casein-sensitized mice. Furthermore, the contribution of iNKT cells to the pathophysiology was investigated and might be a new potential target for dietary intervention in particular in case of whey-sensitization. Future studies using the newly developed murine models for whey and casein allergy will focus on further unraveling the underlying mechanisms of CMA and to study new dietary interventions aimed at the prevention and/or treatment of CMA.

202
M
Nederlandse samenvatting
Nederlandse samenvatting
De prevalentie van allergische aandoeningen is 20%, hiervan is ongeveer 1% gericht tegen eiwitten uit voeding. Koemelkallergie (KMA) vormt een serieus probleem in de westerse wereld. Twee tot drie procent van de baby’s krijgt last van KMA in de eerste levensjaren. Gelukkig groeit 70 tot 80% van deze kinderen over de allergie heen voordat ze vijf jaar oud zijn. Een bijkomend probleem is echter dat deze kinderen een verhoogd risico lopen op het ontwikkelen van andere allergieën en astma in hun latere leven. De belangrijkste symptomen van KMA zijn huidreacties (eczeem), luchtwegklachten, buikpijn en in sommige gevallen zelfs een anafylactische shock. Allergie is een ziekte van het afweersysteem die zich kenmerkt door de aanwezigheid van ongewenste specifieke immunoglobuline E (IgE) antilichamen in het bloed. Kinderen, maar ook volwassenen, met koemelkallergie hebben veelal IgE dat zich richt tegen koemelkallergenen. Om KMA te diagnosticeren wordt dit in het serum gemeten. De twee belangrijke allergene eiwitgroepen in koemelk zijn caseïne- (30 g/L) en wei-eiwitten (5 g/L). De caseïne-eiwitten worden onderverdeeld in αS1-, αS2-, κ- en β-caseïne, terwijl de wei-eiwitten onderverdeeld worden in β-lactoglobuline, α-lactalbumine, serum albumine en lactoferrine. Grote klinische studies hebben uitgewezen dat de belangrijkste allergenen waarop de mens reageert β-lactoglobuline, αS1-caseïne en α-lactalbumine zijn.
Diermodellen voor KMA kunnen helpen bij het onderzoek naar de betrokken mechanismen die leiden tot de ontwikkeling van KMA en bij het toetsen van nieuwe preventieve en therapeutische behandelmogelijkheden. Een nadeel van de meeste bestaande diermodellen is dat de dieren niet gesensibiliseerd (allergisch gemaakt) worden op de manier waarop mensen allergisch raken. Vaak worden proefdieren namelijk op een kunstmatige wijze gesensibiliseerd (bijvoorbeeld via een injectie in de buikholte) en niet via orale toediening van het allergene eiwit. Om onderliggende mechanismen en nieuwe nutritionele concepten voor preventie en/of behandeling van KMA te onderzoeken hebben we een diermodel ontwikkeld waarin muizen via orale toediening van de koemelkeiwitten caseïne- of wei-allergisch worden gemaakt. In deze muizen zijn de pathologische veranderingen bestudeerd en middels dieetinterventies is onderzocht of de ontwikkeling van allergische symptomen kon worden voorkomen. De allergische oortest (vergelijkbaar met de huidpriktest bij mensen) is geïntroduceerd om sensibilisatie aan te tonen. Hierbij wordt caseïne of wei in de huid van het oor gespoten. Als het dier allergisch is voor het eiwit zwelt het oor op en deze verdikking van het oor wordt gemeten (hoofdstuk 3).
De muismodellen voor wei- en voor caseïne-geïnduceerde koemelkallergie ontwikkelden een allergeen specifieke oorzwellingsrespons. Dit suggereert eenzelfde werkingsmechanisme in beide modellen. Er werden echter opmerkelijke verschillen gevonden. Patiёnten met koemelkallergie kunnen last hebben van diarree maar ook van constipatie. Als maat voor

203
M
Miscellaneous
darmmotiliteit werd in orgaanbadstudies de contractiliteit van de darmspieren gemeten. Met name in de dikke darm van muizen die allergisch waren gemaakt voor caseїne bleek de mate van carbachol (een stof die het samentrekken van spieren opwekt) geïnduceerde spiercontractie verhoogd ten opzichte van wei-allergische en controle dieren. Er is meer onderzoek nodig om vast te kunnen stellen wat de reden van deze verschillen is.
Zoals eerder beschreven is allergie een ziekte van het afweersysteem die zich kenmerkt door allergeen specifiek IgE in het bloed. In het wei-model bleek dit precies zo te zijn, maar muizen die met caseïne gesensibiliseerd werden hadden een allergische respons (oorzwelling) in afwezigheid van specifiek IgE. Onze hypothese was dat een ander mechanisme eenzelfde reactie veroorzaakt. In een IgE gemedieerde allergische respons bindt een eiwit, bijvoorbeeld wei, aan IgE, wat gebonden zit aan receptoren op mestcellen. Door receptor crosslinking ontvangt deze allergische effectorcel een signaal waarna hij zijn inhoud (o.a. histamine) uitscheidt in de omgeving. Hierdoor vindt er oedeemvorming en huidirritatie plaats en worden andere cellen aangetrokken; de allergische reactie. Echter, bij caseïne kan dit proces waarschijnlijk niet via IgE antilichamen lopen, omdat deze niet of in erg lage hoeveelheden aanwezig zijn in deze muizen. Op onze afdeling wordt onderzoek gedaan naar een alternatieve route voor het uitlokken van een allergische respons en het betrokken molecuul wordt immunoglobuline vrije lichte keten genoemd (light chain). IgE bestaat uit twee zware (grote) ketens en twee lichte (kleine) ketens die aan elkaar vastzitten. De alternatieve route van allergeenbinding en mestcelldegranulatie zou mogelijk via deze lichte ketens plaats kunnen vinden zonder dat ze aan de zware ketens vastzitten. In het caseïne model is onderzocht of deze immunoglobuline vrije lichte ketens verhoogd aanwezig waren in het bloed, dit bleek inderdaad het geval. Vervolgens injecteerden we F991 in de muizen voordat de oorzwellingstest werd uitgevoerd. F991 is een zogenaamde antagonist en vangt vrije lichte ketens weg. Deze zijn daardoor niet meer actief in de muizen. In F991 behandelde caseïne-allergische muizen vond hierdoor bijna geen oorzwelling meer plaats, terwijl gebruik van F991 geen gevolg had voor de oorzwelling van wei-allergische muizen. Om dit te bevestigen zijn miltcellen van een aantal caseïne-allergische muizen in kweek gebracht. Deze cellen produceren antilichamen, waaronder vrije lichte ketens. Het supernatant van deze miltcellen werd in een niet-allergische (naïeve) muis gespoten en daarna hebben we bestudeerd of er een oorzwelling optrad na injectie van caseïne in het oor. De antilichamen geproduceerd door miltcellen van caseine-allergische muizen bleken inderdaad naïeve muizen allergisch te maken. De oorzwelingsrespons bleek immunoglobuline vrije lichte keten afhankelijk, want F991 kon het optreden van de oorzwelling voorkomen (hoofdstuk 4).
Om te bestuderen of immunoglobuline vrije lichte ketens ook een rol kunnen spelen bij allergische mensen zijn in een groep van zuigelingen die risico lopen om een allergie te ontwikkelen, immunoglobuline vrije lichte ketens gemeten in het serum. In de groep

204
M
Nederlandse samenvatting
van zuigelingen die atopisch eczeem ontwikkelden (vaak ontstaan door een allergische reactie) lagen de waarden van de immunoglobuline vrije lichte ketens hoger dan in de groep van zuigelingen die geen atopisch eczeem hadden. Verder bleken de concentraties van immunoglobuline vrije lichte ketens lager in de groep zuigelingen die Immunofortis® (betekenis wordt in detail in onderstaande alinea uitgelegd) in hun flesvoeding kregen. Juist deze groep ontwikkelde minder vaak atopisch eczeem, in vergelijking met zuigelingen die normale flesvoeding zonder toevoeging van Immunofortis® ontvingen (hoofdstuk 5).
Om de sensibilisatie te verminderen zijn muizenproeven uitgevoerd met verschillende dieetinterventies. Als eerste zijn in het muis-allergiemodel voor wei-eiwitten de dieren gevoerd met verschillende diëten. Een van de diëten bevatte niet-verteerbare koolhydraten (een soort suiker), zogenaamde oligosacchariden. Deze oligosacchariden komen van nature ook voor in moedermelk en in de beschreven studies is een mix gebruikt die erg lijkt op bepaalde oligosacchariden in moedermelk. De gebruikte mix bevatte in totaal 2% korte keten galacto-oligosacchariden (scGOS) en lange keten fructo-oligosacchariden (lcFOS; verhouding 9:1, bekend als Immunofortis®). Het is bekend dat deze mix bepaalde goede bacteriën in de darm stimuleert en op deze manier de gezondheid van het individu ondersteunt. Dit wordt een prebiotisch effect genoemd. De niet-verteerbare koolhydraten worden prebiotica genoemd, de bacteriën zelf probiotica en de combinatie synbiotica. Als potentieel effectieve probiotica is een bacteriestam geselecteerd, de Bifidobacterium breve M-16V. In onze preventieve studie gebruikten we drie diëten: het prebiotische dieet, een probiotisch dieet en de combinatie, een synbiotisch dieet. Alle drie de diëten bleken in staat om de allergische reactie te onderdrukken, maar het synbiotische dieet deed dit het beste. De oorzwelling was sterk verminderd en de anafylactische shock was terug op controleniveau. We vonden geen verschillen in IgE (of IgG
1) antilichaam-niveaus en het aantal regulatoire T-cellen. Deze regulatoire T-cellen zijn te herkennen aan bepaalde receptoren op hun oppervlak (celmembraan), in dit geval CD4, CD25 en FoxP3, en bewaken de immunologische balans zodat deze niet doorslaat in de richting van allergieën of auto-immuunziekten. De niveaus van IgG
2a antilichamen waren verhoogd door de diëten. Dit duidt op een verbeterde balans van de immuunrespons, mogelijk veroorzaakt door de activiteit van regulatoire T-cellen (hoofdstuk 6).
Een derde groep van niet-verteerbare koolhydraten die erg lijkt op die uit moedermelk werd toegevoegd aan de oligosaccharide mix; deze zure oligosaccharide is afgeleid van pectine (pAOS). In een andere experimentenreeks hebben we deze in combinatie met de vorige prebiotische mix getest (2% scGOS/lcFOS/pAOS in een verhouding van 9:1:1). Het preventieve effect op de allergie van de muizen was wederom positief. De oorzwelling werd bijna volledig voorkomen, maar er was nog steeds nauwelijks tot geen effect te zien op de antilichaamniveaus en het aantal regulatoire T-cellen. Toch was onze hypothese dat het dieet een effect had op de regulatoire T-cellen die de ontwikkeling van KMA mogelijk

205
M
Miscellaneous
kunnen afremmen. Om dit te bevestigen zijn deze regulatoire T-cellen voor en tijdens de sensibilisatie verwijderd uit de muizen met een monoklonaal antilichaam gericht tegen de CD25-receptor die aanwezig is op deze cellen. Voor zowel wei- als caseïne-allergische muizen deden we dezelfde ontdekking: als de muizen waarvan de regulatoire T-cellen verwijderd waren gevoerd werden met het prebiotische dieet, kregen ze wel weer een sterke oorzwelling. Hierdoor is de conclusie gerechtvaardigd dat regulatoire T-cellen betrokken zijn bij de gunstige effecten van het prebiotische dieet (hoofdstuk 7 en 8).
Om meer zekerheid te verkrijgen omtrent deze bevinding, hebben we zogenaamde adoptieve transfer-experimenten uitgevoerd. Cellen uit de milt van zowel wei- als caseïne-allergische muizen die een sterk verminderde allergische reactie lieten zien doordat ze het prebiotische dieet hadden ontvangen, zijn geïnjecteerd in naïeve muizen die nog niet allergisch waren gemaakt. We ontdekten dat deze andere muizen niet allergisch gemaakt konden worden en dat de cellen die verantwoordelijk zijn voor deze tolerantie, in de milt aanwezig zijn. Vervolgens voerden we de experimenten een tweede keer uit, maar vóórdat we de miltcellen injecteerden verwijderden we de regulatoire T-cellen uit de celsuspensie. Nu bleek dat de ontvangende muizen niet meer beschermd waren tegen de sensibilisatie met wei of caseïne, ze werden dus allergisch. Verder zijn er aanwijzingen dat bij deze doorbroken bescherming wei- of caseïne-specifieke regulatoire T-cellen betrokken waren. Dit volgt uit het experiment waarin we muizen wel voerden met het prebiotische dieet, deze muizen werden niet gesensibiliseerd met caseine of wei. De miltcellen van deze niet-allergisch gemaakte muizen werden geïnjecteerd in naïeve muizen. Deze muizen waren echter niet beschermd tegen het ontwikkelen van allergie. Het oligosaccharide dieet ondersteunt dus de ontwikkeling van wei- of caseïne-specifieke regulatoire T-cellen en juist deze antigeen-specifieke cellen kunnen bescherming bieden tegen het ontwikkelen van allergie (hoofdstuk 7 en 8).
Naast de regulatoire T-cel waren we ook geïnteresseerd in de bijdrage van een ander type regulatoire cel, namelijk de ‘Natural Killer T’ (NKT)-cel in de bescherming tegen koemelkallergie. De eerste experimenten toonden aan dat zowel wei- als caseïne-allergische muizen een verlaagd aantal van deze NKT cellen in de lever hadden. Uit dit gegeven ontstond de hypothese dat wanneer we deze cellen zouden activeren of laten vermenigvuldigen, de allergie zou verminderen. De activatie vond plaats door middel van de stof α-galactosylceramide (kortweg αGalCer). αGalCer kan selectief de NKT-cel stimuleren. We vonden echter geen verlaagde allergische respons, zoals verwacht, maar juist een verhoging van de acute huidrespons (voornamelijk in de wei-allergische dieren). We onderzochten de NKT-cellen in de lever en deze bleken zeer sterk in aantal verminderd door de toediening van αGalCer. Ook migratie van deze cellen konden we vrijwel uitsluiten omdat we ook in andere organen deze cellen hebben geteld (milt en mesenterische lymfeknopen). Uit deze bevinding concludeerden we dat we, door de dieren een aantal maal met αGalCer te stimuleren, de NKT-cellen niet geactiveerd, maar juist verwijderd hadden. De consequentie

206
M
Nederlandse samenvatting
was dat de allergie toenam in plaats van te verminderen. De data die zijn beschreven in dit hoofdstuk suggereren dat NKT-cellen immunomodulerende capaciteiten bevatten, met name in muizen die wei-allergisch zijn (hoofdstuk 9).
In dit proefschrift zijn twee muismodellen voor koemelkallergie ontwikkeld. De muizen zijn oraal gesensibiliseerd tegen wei- of caseïne-eiwitten en de verschillen in onderliggende mechanismen tussen deze twee modellen zijn bestudeerd. Verschillende dieetinterventies lieten een preventief beschermend effect zien op het ontstaan van KMA, vooral het synbiotische dieet leek hierbij veelbelovend. De betrokkenheid van eiwit-specifieke regulatoire T-cellen in het beschermende effect door de diëten is aangetoond in de wei- en caseïne-allergische muizen. Daarnaast is de bijdrage van NKT-cellen aan de allergische respons tegen wei en caseïne onderzocht. NKT-cellen lijken een mogelijk target te zijn voor dieetinterventie. De ontwikkelde muismodellen bieden de mogelijkheid om meer mechanistisch onderzoek te verrichten naar het ontstaan van KMA. De hieruit opgedane kennis kan in de toekomst mogelijk gebruikt worden om nieuwe therapeutische of preventieve (voedings)interventies te ontwikkelen met als doel de klachten van KMA te voorkomen of mogelijk zelfs te verhelpen.

207
M
Dankwoord
Dankwoord
Zo, het zit erop! Na vier jaar van denken, uitvoeren, analyseren en schrijven, heel veel schrijven, is het af. Wat kun je veel doen in vier jaar en toch vliegt de tijd voorbij. Nu alleen het meest gelezen stuk van het proefschrift nog: het dankwoord. Het tot stand komen van dit ‘boekje’ is zeker geen soloactie en daarom neem ik de gelegenheid om iedereen te bedanken voor zijn of haar bijdrage! Natuurlijk ga ik iemand vergeten, maar jij ook bedankt!
Zoals elke promotie begint, zo begon het ook bij mij: met een promotor. Johan, ik blijf me verbazen over de hoeveelheid dingen die jij voor elkaar krijgt. En dan heb je zelfs nog tijd om op het lab te helpen. Jij kunt als geen ander mensen motiveren voor onderzoek. Ik ben je dankbaar dat je mij inwijdde in de wereld van de immunologie. Wat zal ik onze gesprekken missen tijdens de ITH’s (meer in detail zal ik niet treden).
Linette, als co-promotor is het je taak om de dagelijkse begeleiding van je AIO’s te verzorgen. Jij hebt dit heel letterlijk genomen en waar ik juist de grote lijnen zie en wel denk te weten hoe het zit, ga jij een stap verder en wil je echt elk detail weten. Jij hebt het perfectionisme tot een kunst verheven! Wat hebben wij een hoge mate van complementariteit. Zonder jouw inzet en motivatie was de helft nog niet op papier gekomen.
Léon, als tweede co-promotor kwam je iets later bij de club, maar je was absoluut een aanwinst voor ons. Je inzet en je kennis van allergiemodellen, samen met je relativeringsvermogen, hebben zeer zeker bijgedragen aan dit proefschrift. Nu zijn we eindelijk formeel collega’s. Bedankt voor je wijze input!
I own many thanks to the reading committee who had to judge about the thoroughness of this manuscript. I was optimistic about your decision, though you were really enthusiastic! Professor Joost Ruitenberg, Professor Georg Kraal, Professor Henk van Loveren, Professor Günther Boehm en Dr. Maarten Hoekstra: thank you all for reading this manuscript!
Maandelijks hadden we overleg met het Utrecht Center for Food Allergy. Deze mensen kwam ik elke keer ook weer tegen als ik ergens op congres was… Edward, Els, Carla, Maarten, André, Suzanne, Henk, Raymond, Marissa, Marianne, Joost, Janine, Geert, Jolanda en alle anderen, bedankt voor jullie waardevolle discussies!
Louis, jou wil ik ook bedanken voor je input, maar vooral voor je antilichamen. Zonder de PC61 had ik de mooiste hoofdstukken niet gehad!
En dan natuurlijk het lab @ UIPS, daar is het echte werk uitgevoerd en daar heb ik het meest

208
M
Dankwoord
gezeten. Allereerst Betty, wat had ik zonder jou gemoeten? Je hebt ervoor gezorgd dat we een vliegende start konden maken en de data bleven maar binnenstromen. We hadden zelfs zoveel dat er al snel besloten werd dat je voor jezelf zou gaan beginnen. Ik zie uit naar jouw boekje! Het meest zal ik onze gezamenlijke congresbezoeken missen. Altijd gezellig, altijd even de stad in om iets van de sfeer op te snuiven. Ik vind het super dat je mijn paranimf wilt zijn!
Een kunstenaar in wat je doet ben jij, Gerard. Nooit te beroerd om iets uit te voeren en bedankjes wilde je niet (‘doe maar in de emmer’). Hierbij dan toch: bedankt!
Dank ben ik ook verschuldigd aan mijn mede-AIO’s: Sven, Esmaeil, Anneke, Yousef en Hadi, jullie gingen mij voor op het nobele pad der promovendi. Gelukkig gingen er niet alleen mensen weg. Marco kwam de gelederen versterken (ook op foto-gebied!) en vervolgens kwamen de Sassen (Saskia O. en Saskia B.) erbij (wanneer gaan we eigenlijk weer verder met Janeway?). Bedankt voor de toffe tijd, ik mis jullie nu al.
Mijn UIPS-kamergenoten wil ik ook hartelijk danken! Anneke, je was mijn vraagbaak tijdens de eerste twee jaar van mijn AIO-tijd en met jouw vertrek raakte ik mijn voorbeeld kwijt. Gelukkig bleef ik niet alleen achter in mijn sobere multimediale verblijf. Daphne en Tom (weer een AIO!), jullie kwamen mij vergezellen. Bedankt voor de goede gesprekken! Tom, samen de wereld van Light chain verkennen was super! Veel dank aan the Light Chain team onder aanvoering van Frank, dankzij jullie heb ik twee extra hoofdstukken in mijn boekje! En Bart, het wordt toch lastig om mijn dankwoord langer te laten worden dan de rest, maar ik doe mijn best! Het blotten voor mij is voorbij, denk ik…
En dan de rest van de FFF. Jullie hebben mij vier jaar lang als Fijne Farmacologie Familie een echt tweede thuis gegeven! Frans, Gert, Aletta, Paul, Andries, Ferdi, Marjolein, Judith, Maaike, Hans, Els, Dinij, Thea, Ingrid, Astrid, Naomi, Kim, Marije, Liz, Pim, Bin, Sel Gillina, Abdi, Nahid, Simone, Tamara, Karin: bedankt!
Verder wil ik graag mijn studenten bedanken. Ook jullie hebben bijgedragen aan de kennis die we nu hebben over koemelkallergie. Ik heb veel van jullie geleerd en ik hoop jullie ook van mij. Ben, Richard, Darya, Karin, Louise, Simone, Nicole, Feike, Michael en Stephanie: super bedankt. Mijn dank geldt ook de studenten van anderen die mij zijdelings hielpen: Sandra, Diana, Tea, Roderick en anderen. Nu denken Lieke, Sander en Laura dat ik hen vergeten ben. Nou, dat lukt me niet en ik zou het ook niet willen! Lieke; eerst onze bachelorstudent, toen onze masterstudent en nu collega-AIO! Ik was blij dat je mijn in vitro taken kwam overnemen, want ik voelde me steeds schizofrener worden. Hetzelfde geldt voor Sander, ik was blij dat je onze koemelkgroep kwam versterken, ook eerst als student en nu als mede-AIO. En Laura, na je stage bleef je maar komen en toen mocht je dan soort van blijven! Nog

209
M
Miscellaneous
een UIPS-AIO die werkt aan het interessante project voedselallergie.
Naast de FFF was er nog een afdeling waar ik gedurende het vorderen van mijn promotie steeds meer aanwezig was: Danone. Mijn flex-plek werd langzaamaan omgezet in een vaste plek, bij Joyce en Anita op de kamer. Ik wil jullie bedanken voor het kritisch meedenken, zowel inhoudelijk als over de vorm. Gelukkig hebben jullie voorkomen dat er allemaal pyjamaplaatjes in mijn boekje staan! En jij Joyce, super bedankt voor het proof readen van mijn proefschrift, je zult je niet verveeld hebben tijdens de treinreis van 2x5 uur naar Duitsland.Veel dank ook aan de andere Danoontjes: Paul (straks nog een persoonlijk woord voor jou), Sander, Niki, Prescilla, Nicole, Nienke, Jaqueline, Miranda, Jeroen, Bea, Alma, Karen, Hilde, Suzan, en naast immunologen nog: Jan, Kaouther, Monique, Thijmen, Martijn, Eline V. Veel PR voor mijn proefschrift is verzorgd door Mirjam; bedankt voor alle interviews. Misschien gaan mensen mij op straat nog eens herkennen.En dan de laatste groep van Danone, mijn nieuwe collega’s. Mieke en Patricia, jullie maakten al eerder de switch van immunology naar klinische studies. Ik wil jullie bedanken voor de emotionele hulp in de eindstrijd en het inwijden in de wonderlijk gestructureerde wereld van de klinische studies. Verder de rest van het baby team: Jürgen, Paul, Tarik, Jenny, Dineke, Martine, Xuchen, Stefanie, Sarah, Claudia, Carolien, Flemming, Liandré, Gerda. En natuurlijk ook alle anderen van medical, ik voel mij al helemaal thuis!
Er is nog een groep mensen die mij al jaren door dik en dun steunt en mijn gedachten af en toe wist los te rukken van mijn promotie: mijn vrienden. Wat moest ik zonder jullie, bedankt voor de vriendschap! Leendert, Nelleke, Jesse en Chaja, bedankt voor het gebruik van jullie tuin! We hopen er binnenkort zelf een te hebben, maar we zullen blijven komen!Gerrit en Dinella, de langste avond van het jaar houden jullie ons gezelschap, maar zelfs dan komen we nauwelijks toe aan een potje Kolonisten. Nu jullie een eigen huisje hebben gaan we zeker vaker bij jullie langskomen! Christianne, jij bent een beetje de zus die ik nooit gehad heb en veel mensen zien ons ook zo. Timon en Jolien, bedankt voor de gesprekken, ook die over slechte docenten, ik hoop dat ik een betere was voor mijn studenten…Af en toe is het prettig om met een AIO uit een ander vakgebied te praten over hoe het project loopt. Jan Harm en Myra, bedankt hiervoor en jij JH, succes met de laatste loodjes!Maarten en Suze, het was erg leuk en handig om een adresje in Utrecht te hebben voor als ik moest ‘overblijven’. Dat is nu voorbij, maar we houden contact. Jaap, Janneke, Lydia, Jort en Thirza; Paul Peter, Jolanda en Nathan: ik hoop dat onze kids nog veel speelplezier samen zullen hebben.Dan heb ik ook nog mijn volleybal vrienden: Christian, Tessa, Dirk, Annemien, Paul (alweer), Bart en Erik, bij jullie kon ik wekelijks mijn frustratie eruit slaan.

210
M
Dankwoord
Het hele volleybalteam bestaat uit mensen uit de NGK en een aantal vervult een dubbelrol; de kring gaf op de woensdagavonden een welkome ontspanning en in de muziek vond ik ook afleiding om zaken te kunnen relativeren. Jan, Rense, Annemien, Christian, Christine, Annelie, (kleine) Rense, Sandor, Lisette, Jaap, Petra, Jordy, Kimberly, Paul, Geja, Karlan, Mincke, Johan, Ineke, Evert en Gerlies: bedankt voor de goede momenten, het is duidelijk waar het in het leven om gaat.Collega, volley-teamgenoot, kringleden, vrienden… jullie passen overal bij: Paul en Geja en natuurlijk jullie kids, Noah en Florian. Jullie weten wat er voor nodig is om te promoveren en Paul is in meerdere opzichten een voorganger en voorbeeld voor mij geweest. Tof dat we zoveel met elkaar delen, laten we dat blijven doen! En Paul, bedankt dat je mij blijft steunen tot en met de promotie zelf, super dat jij ook mijn paranimf wilt zijn.
Pa en Ma, bedankt voor de mogelijkheden die jullie mij gegeven hebben. Jullie stonden en staan altijd voor ons klaar met tijd en ook met ruimte in meerdere opzichten!Maarten en Susanne, broer en (schoon)zus, ik heb er echt een zus bij gekregen! Bedankt voor jullie vriendschap! En dan nu de belangrijkste mensen in mijn leven. Adriëlle, levensgezellin. Wat hebben wij al veel beleefd. Telkens weer denken we dat de drukte wel voorbij is en dan verzinnen we weer iets. Ik ben dankbaar voor je liefde en je hulp tijdens deze fase, zonder jou had ik het niet gered! Met name voor het corrigeren van de Nederlandse teksten ben ik je veel dank verschuldigd. Ik hou van jou!!!Natuurlijk noem ik hier ook Joël en Micha en ook Kimberly. Jullie zijn samen het beste (en vermoeiendste) dat mij is overkomen. Joël, je zei wel eens: ‘Papa, je oeft vandaa niete werke…’. Dan was het niet makkelijk om de deur uit te stappen! Bedankt voor jullie liefde!
Als laatste geef ik alle eer aan mijn Hemelse Vader. Hij schiep alles zo fantastisch, dat we
Ben ik in het schrijven soms te kort en bondig, dit dankwoord is allerminst kort te noemen. Toch sluit ik pas af nadat ik nog een aantal voor mij zeer belangrijke mensen bedankt heb.Allereerst mijn schoonfamilie: Pa, Ma, Wiggert, Beate, Make en Opa Velema. Ik heb het getroffen met jullie!Opa en Oma Van Ham, al de keren dat ik vroeger bij jullie speelde hebben jullie mij laten zien hoe je op een degelijke manier dingen maakt. Hiermee hebben jullie de basis gelegd voor het doen van goed onderzoek, bedankt!Oma Schouten, wat zou Opa dit graag meegemaakt hebben! Hij wist als geen ander te analyseren en door te dringen tot de kern van de zaak. Precies een jaar geleden is hij overleden. We denken nog vaak aan hem, en natuurlijk ook aan u!
zelfs na jaren van intensieve studie nog nauwelijks weten hoe het immuunsysteem in elkaar zit.

211
M
Miscellaneous
Zie ik de hemel, het werk van Uw vingers, de maan en de sterren door U daar bevestigd, wat is de sterveling dat U aan hem denkt, het mensenkind dat U naar hem omziet?
Psalm 8:4 en 5

212
M
Curriculum Vitae
Curriculum Vitae
Bastiaan Schouten was born in Utrecht on 27 February 1981 and graduated from the Menso Alting College in Hoogeveen in 2000 and decided to study Biology at the Wageningen University and Research Centre. During his first student internship he studied the estrogenic potency of different food-packaging-associated plasticizers and antioxidants under supervision of Dr. M.G.R. ter Veld, Prof. Dr. A.J. Murk and Prof. Dr. Ir. I.M.C.M. Rietjens at the sub-department of Toxicology of the Wageningen University and Research Centre. During his traineeship at Numico Research B.V. in Wageningen, he studied central fatigue during chemotherapy in rats, under supervision of Dr. E.M. van der Beek and Dr. L.M. Broersen. His second student internship concerned the biological, ethical, philosophical and theological aspects of biometric identification under supervision of Prof. Dr. Ir. E. Schuurman of Reformational Philosophy at the Applied Philosophy group of the Wageningen University and Research Centre. During his study the author was chairman of the student corps Ichthus Wageningen in 2004-2005.
After graduating, the author started as a PhD student at the division of Pharmacology and Pathophysiology of the Utrecht Institute of Pharmaceutical Sciences (UIPS) of the Utrecht University in September 2005. The project was entitled: “Role for pre- and probiotics in immune-modulation, especially during allergic reactions to food components” under supervision of Prof. Dr. J. Garssen, Dr. L.E.M. Willemsen (UIPS) and Dr. L.M.J. Knippels (Danone Research) in collaboration with Danone Research – Centre for Specialised Nutrition in Wageningen. During his PhD, the author was member of the PhD-council of the Utrecht Institute of Pharmaceutical Sciences graduate school, member of the PhD-council of the Eijkman graduate school, member of the Graduate School of Life Sciences and co-founder and member of the PhD-council of the faculty of Science.
Currently, the author is employed as a Clinical Study Manager in the Clinical Studies Platform of Danone Research – Centre for Specialised Nutrition, headed by Dr. Ir. J. van der Mooren.
He is married to Adriëlle Schouten-Loonstra and together they have two sons, Joël (2006) and Micha (2008).

213
M
List of publications
List of publications
Full papers
Cow’s milk allergy symptoms are reduced in mice fed dietary synbiotics during oral sensitization with wheySchouten B., Van Esch B.C.A.M., Hofman G.A., Van Doorn S.A.C.M., Nauta A.J., Garssen J., Willemsen L.E.M., Knippels L.M.J.2009 The Journal of Nutrition139(7):1398-1403
Acute allergic skin response as a new tool to evaluate the allergenicity of whey hydrolisates in a mouse model of orally induced cow’s milk allergyVan Esch B.C.A.M., Schouten B., Hofman G.A., Van Baalen T., Nijkamp F.P., Knippels L.M.J., Willemsen L.E.M., Garssen J. 2009 Pediatric Allergy and Immunology, in press
Exposure of intestinal epithelial cells to UV killed Lactobacillus GG but not Bifidobacteria breve enhances the effector immune response in vitroVan Hoffen E., Korthagen N.M., De Kivit S., Schouten B., Bardoel B., Duivelshof A., Knol J., Garssen J., Willemsen L.E.M.2009 International Archives of Allergy and Immunology, in press
Acute allergic skin reactions and intestinal contractility changes in mice orally sensitized against casein or wheySchouten B., Van Esch E.C.A.M., Hofman G.A., Van den Elsen L.W.J., Willemsen L.E.M., Garssen J.2008 International Archives of Allergy and Immunology 147(2): p. 125-134
Non-digestible oligosaccharides reduce immunoglobulin free light-chain levelsSchouten B., Van Esch B.C.A.M., Groot Kormelink T., Moro G.E., Boehm G., Arslanoglu A., Willemsen L.E.M., Knippels L.M.J., Garssen J. Submitted
Contribution of IgE and Immunoglobulin free light chain in the allergic reaction to cow’s milk proteinsSchouten B., Van Esch B.C.A.M., Blokhuis B., Groot Kormelink T., Hofman G.A., Moro G.E., Boehm G., Arslanoglu A., Willemsen L.E.M., Knippels L.M.J., Garssen J. Submitted

214
M
List of publications
Dietary non-digestible carbohydrates inhibit the allergic effector response via induction of casein-specific CD25+ regulatory T-cells in orally sensitized miceSchouten B., Van Esch B.C.A.M., Hofman G.A., De Kivit S., Boon L., Knippels L.M.J., Garssen J., Willemsen L.E.M. Submitted
Oligosaccharide induced whey-specific CD25+ regulatory T-cells are crucial in the suppression of cow’s milk allergy in miceSchouten B., Van Esch B.C.A.M., Hofman G.A., Boon L., Knippels L.M.J., Willemsen L.E.M., Garssen J.Submitted
Nutrition and food allergySchouten B., Van Esch B.C.A.M., Knippels L.M.J., Willemsen L.E.M., Garssen J. Review in preparation
iNKT cells contribute to the allergic response in cow’s milk protein sensitized miceSchouten B., Van Esch B.C.A.M., Lim S., Hofman G.A., Knippels L.M.J., Willemsen L.E.M., Garssen J., Vos A.P. In preparation
CD25+ regulatory T-cells play a prominent role in immune modulation by specific oligosaccharidesVan ‘t Land B., Schijf M., Van Esch B.C.A.M., Van Bergenhenegouwen J., Schouten B., Bastiaans J., Boon L., Garssen J. In preparation
Estrogenic potency of food-packaging-associated plasticizers and antioxidants as detected in ERalpha and ERbeta reporter gene cell linesTer Veld M.G.R., Schouten B., Louisse J., Van Es D.S., Van der Saag P.T., Rietjens I.M.C.M., Murk A.J.2006. J Agric Food Chem 54(12): p. 4407-16
Abstract publications
Immunoglobulin free light chains play a pivotal role in casein allergy in mice and might be a biomarker for atopic dermatitis in infantsSchouten B., Van Esch B., Blokhuis B., Groot Kormelink T., Hofman G., Moro G., Boehm G., Arslanoglu S., Knippels L., Willemsen L., Redegeld F., Garssen J.

215
M
Miscellaneous
2009 Allergy, suppl 90, vol 64: p54
A probiotic mixture reduces the acute allergic skin response via regulatory T cells in mice orally sensitised to wheySchouten B., Van Esch B., Hofman G., Boon L., Knippels L., Willemsen L., Garssen J.2009 Allergy, suppl 90, vol 64: p63
Induction of acute systemic anaphylaxis in a murine model of cow's milk allergyNauta A.J., Schouten B., Van Doorn S.A.C.M., Van Esch B.C.A.M., Willemsen L.E.M., Garssen J., Knippels L.M.J. 2008 Allergy, suppl 88, vol 63: p9
Prebiotic oligosaccharides reduce casein sensitization in mice: a possible role for regulatory T cellsSchouten B., Van Esch B.C.A.M., Hofman G.A., Boon L., Knippels L.M.J., Willemsen L.E.M., Garssen J. 2008 Allergy, suppl 88, vol 63: p62
Anti-CD25 treatment in a mouse model for cow's milk allergy modifies the acute skin response from IgE- into Ig free light chain-dependentVan Esch B.C.A.M., Schouten B., Blokhuis B.R.J., Hofman G.A., Boon L., Knippels L.M.J., Willemsen L.E.M., Garssen J., Redegeld F.A. 2008 Allergy, suppl 88, vol 63: p. 181
A specific mixture of short chain galacto-oligosaccharides and long chain fructo-oligosaccharides and a probiotic strain suppresses the allergic reaction in cow's milk allergic miceSchouten B., Van Esch B.C.A.M., Nauta A.J., Van Doorn S.A.C.M., Hofman G.A., Knol J., Garssen J., Willemsen L.E.M., Knippels L.M.J. 2008 Allergy, suppl 88, vol 63: p406
T-helper 1 skewing of the effector immune response through intestinal epithelial cells exposed to Toll like receptor agonistsDe Kivit S., Korthagen N., Schouten B., Van Hoffen E., Garssen J., Willemsen L.E.M. 2008 Allergy, suppl 88, vol 63: p409
Acute skin response in mice as a measure for cow's milk hydrolysate induced oral tolerance to whey proteinsVan Esch B.C.A.M., Schouten B., Hofman G.A., Van Baalen T., Nauta A.J., Knippels L.M.J., Willemsen L.E.M., Garssen J.

216
M
List of publications
2008 Allergy, suppl 88, vol 63: p461
A prebiotic oligosaccharide mixture inhibits pathophysiological changes in mice orally sensitized against casein or wheySchouten B., Van Esch B.C.A.M., Hofman G.A., Willemsen L.E.M., Garssen J. 2008 Nederlands Tijdschrift voor Allergie, vol 8(1)
Intestinal epithelial cells exposed to probiotics or TLR/NOD2 agonists enhance Th1 type cytokine secretionWillemsen L.E.M., Korthagen N., De Kivit S., Schouten B., Duivelshof A., Knol J., Garssen J. 2008 Nederlands Tijdschrift voor Allergie, vol 8(1)
A new animal model for cow’s milk allergy: useful tool for epitope testingVan Esch B.C.A.M., Schouten B., Hofman G.A., Willemsen L.E.M., Knol E., Van Hoffen E., Van Baalen T., Knippels L.M.J., Garssen J. 2007 Allergy, suppl 83, vol 32, p57, 139
Differential pathophysiological changes in mice orally sensitized to casein or whey proteinsSchouten B., Van Esch B.C.A.M., Hofman G.A., Van den Elsen L.W.J., Willemsen L.E.M., Garssen J. 2007 Gastroenterology, suppl 1, vol 132, nr 4, p460, M2156
Dietary intervention reduces inflammation and depression relevant to Alzheimer’s DiseaseDe Wilde M.C., Schouten B., Balvers M., Kamphuis P.J., Van der Beek E.M., Broersen L.M. 2006 Alzheimer’s & Dementia vol 2(3): S237 p2-016

217
M
Abbreviations
Abbreviations
AD: atopic dermatitis CM: cow’s milk CMA: cow’s milk allergy CT: cholera toxin DBPCFC: double-blind placebo controlled food challenge dp: degree of polymerization FA: food allergy fLC: free light chains GM-CSF: granulocyte macrophage colony-stimulating factor I.d.: intra dermal IFN-γ: interferon-γ Ig: immunoglobulin I.g.: intra gastric IL: interleukin iNKT: invariant natural killer T (cells) lcFOS: long-chain fructo oligosaccharides MLN: mesenteric lymph node mMCP-1: mouse mast cell protease-1 pAOS: acidic oligosaccharides derived from pectin PP: Peyer’s patch PUFA: polyunsaturated fatty acids scGOS: short-chain galacto oligosaccharides sIgA: secretory immunoglobulin A SPT : skin prick test TCR: T cell receptor T
H: T helper cell TNF-α: Tumor necrosis factor-α Treg: regulatory T (cells)




















