
offspring of rabbit mothers with chronic secondary hypertension
38
1 Gender differences in post-natal growth and renal development in offspring of rabbit mothers with chronic secondary hypertension. D Maduwegedera 1 , MM Kett 1 , RL Flower 1 , GW Lambert 2 , JF Bertram 3 , EM Wintour 1 , KM Denton 1 . 1 Department of Physiology Monash University, 2 Baker Heart Institute and 3 Department of Anatomy and Cell Biology Monash University, Melbourne, Australia. Running Title: Impact of maternal hypertension on offspring Correspondence: Dr Kate Denton Department of Physiology Monash University Victoria Australia. Ph: 613 99059553 Fax: 613 99055666 Email: [email protected] Page 1 of 38 Copyright © 2006 by the American Physiological Society.
Transcript of offspring of rabbit mothers with chronic secondary hypertension

1
Gender differences in post-natal growth and renal development in
offspring of rabbit mothers with chronic secondary hypertension.
D Maduwegedera1, MM Kett1, RL Flower1, GW Lambert2, JF Bertram3, EM Wintour1, KM
Denton1.
1Department of Physiology Monash University, 2 Baker Heart Institute and 3Department of
Anatomy and Cell Biology Monash University, Melbourne, Australia.
Running Title: Impact of maternal hypertension on offspring
Correspondence:
Dr Kate Denton
Department of Physiology
Monash University
Victoria
Australia.
Ph: 613 99059553
Fax: 613 99055666
Email: [email protected]
Page 1 of 38
Copyright © 2006 by the American Physiological Society.

2
Abstract
Previously we demonstrated that adult blood pressure was increased in offspring of rabbit
mothers with chronic secondary renal hypertension. Our study identified sex-specific
differences in the programming of hypertension, with female not male offspring having
increased blood pressure at 30 weeks of age. The aim of this study was to characterise
the maternal hypertension during pregnancy to determine potential programming stimuli.
Further, we examined the impact of chronic maternal hypertension on offspring birth
weight, nephron number and renal noradrenaline content (as an index of renal innervation
density). Three groups of mothers and their offspring were studied; two-kidney, one-wrap
(2K-1W, n = 9 mothers) hypertensive, two-kidney, two-wrap (2K-2W, n = 8) hypertensive,
and a sham–operated group (n = 9). Mean arterial blood pressure was increased by ~20
mmHg throughout pregnancy in both hypertensive groups as compared to sham mothers
(PG < 0.001). Plasma renin activity (PRA; PG < 0.05) and aldosterone (PG < 0.05) levels
were increased during gestation in the 2K-1W, but not the 2K-2W mothers. Birth weight
was increased by ~20% in offspring of both groups of hypertensive mothers (PT < 0.001),
though this was associated with a reduction in litter size. Renal noradrenaline content was
increased (~40%, P < 0.05) at 5 weeks of age in female 2K-1W offspring as compared to
sham offspring. Glomerular number was not reduced in offspring of either group of
hypertensive mothers, however glomerular tuft volume was reduced in female 2K-2W
offspring (P < 0.05), indicative of a reduction in glomerular filtration surface area. In
conclusion, the two models of renal hypertension produced differential effects on the
offspring. The impact of a stimulated maternal renin-angiotensin system in the 2K-1W
model of hypertension may influence development of the renal sympathetic nerves and
contribute to programming of adult hypertension.
Page 2 of 38

3
Introduction
Chronic hypertension during pregnancy is an increasing problem as women are tending to
have children later in life when the incidence of obesity, diabetes and renal disease
associated hypertension is increased. The babies of women with chronic hypertension not
only have acute problems perinatally, but more disturbingly, may have an increased long-
term burden in the form of future cardiovascular risk (22, 53). There is now compelling
evidence, both epidemiological and experimental, to support the hypothesis that events
occurring in fetal life can have life-long consequences for the health of the adult (3, 24).
The importance of this issue has been recently highlighted in a NHLBI working group
report recommending increased focus on the study of hypertensive disorders in pregnancy
and in particular the possibility of future cardiovascular risk to offspring (51).
Previously, we demonstrated that adult blood pressure was increased in offspring of rabbit
mothers with pre-existing mild secondary hypertension (2K-1W) (13). Our study identified
sex-specific differences in the programming of hypertension, with female not male
offspring having increased blood pressure at 30 weeks of age. However, in this study
neither maternal blood pressure during pregnancy or birth weight was determined.
Therefore in the current study we documented mean arterial pressure (MAP) and plasma
renin activity (PRA) in a 2K-1W model of hypertension throughout pregnancy.
Furthermore, we incorporated a second model of maternal hypertension in our studies, the
2-kidney, 2-wrap model (2K-2W), with the aim of producing a more severe degree of
hypertension in the mothers, as previously reported in males (10, 11). We hypothesised
that the increase in MAP in 2K-2W mothers would be greater than the 2K-1W model and
that the degree of elevation in blood pressure would be maintained in both hypertensive
models during pregnancy.
Page 3 of 38

4
Low birth weight has been associated with adult hypertension in several models of
adverse intrauterine environments, though accumulating evidence suggests that altered
fetal development can also occur independently of low birth weight (45). It is well
recognised that hypertension in pregnancy is associated with low birth weight in humans
(22, 53). Therefore, we also addressed the hypothesis that birth weight would be reduced
in the offspring of these hypertensive pregnancies.
Two potential mechanisms that might contribute to the programming of hypertension in
adulthood were also examined. A reduced nephron endowment / filtration surface area
has been linked to adult hypertension (32). Thus nephron number was determined,
stereologically, in kidneys from 5 week old offspring, an age when nephrogenesis is
complete (42). We hypothesised that glomerular number would be decreased by maternal
hypertension. Strong evidence implicates overactivity of the renal sympathetic nerves in
the pathogenesis of hypertension (20, 28). Studies in the spontaneously hypertensive rat
(SHR) have shown altered renal nerve development (25). Total noradrenaline levels were
increased in SHR when compared to controls, consistent with increased sympathetic
innervation density (25). We therefore also aimed to identify whether the renal
sympathetic innervation was altered in the offspring of hypertensive mothers; a possible
mechanism by which offspring are rendered vulnerable to developing increased blood
pressure in later life. It was hypothesised that renal noradrenaline content would be
increased (indicative of an increased sympathetic innervation) in the offspring of
hypertensive mothers.
Page 4 of 38

5
Methods
Animals
English crossbred rabbits were used. Experiments were performed in accordance with the
Australian Code of Practice for the Care and Use of Animals for Scientific Purposes and
were approved in advance by the Monash University Standing Committee on Ethics in
Animal Experimentation.
Mothers
Nulliparous female rabbits (n = 26, 16 ± 1 weeks of age, 2.7 ± 0.1 kg) were housed
individually in a room maintained at a temperature of 23-25 °C with a 12h light/dark cycle.
Rabbits had free access to water and were fed high fibre, low starch rabbit pellets ad
libitum (Glen Forrest Stockfeeders, WA, Australia).
Arterial blood pressure and body weight were measured in rabbits on a control day.
Catheters were placed in the central ear arteries under local anesthetic (Xylocaine; Astra
Pharmaceuticals, NSW, Australia) and conscious arterial pressure was measured for 30
minutes (13). An arterial blood sample (total volume ~5 ml) was then collected for
hematocrit, plasma creatinine, plasma renin activity (PRA) and aldosterone measurement.
Surgery was then performed on three groups of rabbits; 2 kidney, 1 wrapped (2K-1W, n =
9) and 2 kidney, 2 wrapped (2K-2W, n = 8) hypertensive and sham-operated (n = 9)
normotensive rabbits. In brief, anesthesia was induced with propofol (10 mg/kg IV;
Diprivan, Zeneca) and maintained with isoflurane (Forthane, Abbot). In all animals
incisions were made on both flanks to expose left and right kidneys. In the sham-operated
group the kidneys were untouched. In the 2K-1W model the left kidney was wrapped in
cellophane (15 x 12 cm) and in the 2K-2W group both kidneys were wrapped in cellophane
Page 5 of 38

6
(10, 11, 13). Four weeks following surgery at gestational age 0 (GA0) blood pressure and
blood samples were repeated after which the rabbits were mated with a normotensive
male. Blood pressure and blood sampling measurements were then repeated at GA18
and GA28 and 5 weeks post-partum.
Offspring
Kittens were spontaneously delivered between 31-32 days gestation. Total number of
offspring (both live and stillborn) and birth weight was recorded. On the day of birth
kidneys were collected from 2 kittens from each litter, which were killed by anesthetic
overdose (Pentobarbitone 25 mg/kg i.p.). Kidneys were collected, weighed, snap frozen in
liquid nitrogen and stored at –70 oC for later noradrenaline content assay.
The remaining offspring were weighed weekly and at 5 weeks of age arterial blood
pressure was measured in the conscious animals via an intra-arterial catheter inserted into
the central ear artery. Arterial blood pressure, hematocrit, plasma creatinine and PRA
measurements were made as described above in the mothers. The rabbit offspring were
than anesthetised with pentobarbitone (60 mg/kg, i.p.) and the kidneys exposed via a
midline incision. Ligatures were placed around the left renal vessels, the kidney excised,
weighed and snap frozen for later noradrenaline content measurement. A catheter was
placed in the lower abdominal aorta below the remaining right kidney in preparation for
fixation. The kidney, was cleared by rinsing with phosphate buffer and then perfusion fixed
at arterial pressure with 4% paraformaldehyde in 0.1M phosphate buffer for ten minutes as
previously described (12).
Glomerular Stereology
Right kidneys were cut in half and then into quarters, each quarter was then sliced into 1.5
Page 6 of 38

7
mm slices. Due to the existing variation in size of slices, they were arranged from smallest
to largest and every tenth slice taken for further sampling, with the first slice chosen at
random. Tissue blocks were processed and embedded in glycolmethacrylate (Technovit
7100, Haraeus Kulzer Gmbh, Germany). Blocks were exhaustively sectioned at 20 µm
with every 10th and 11th section sampled, the first of which was chosen at random from
the first 10 sections. Sections were stained with periodic acid-Schiff reagent (PAS).
Total kidney volume was estimated by the Cavalieri principle. In brief, every 10th section
was placed under a microfiche with a superimposed grid (3 x 3 mm) and points falling on
kidney tissue counted. The following formula was used to calculate kidney volume (Vkid):
Vkid = F1 × F2 × t × a(p) × Ps
Where F1 is the inverse of the first sampling fraction (F1 = 10; i.e. every 10th slice was
sampled) and F2 is the inverse of the second sampling fraction (F2 = 10, i.e. every 10th pair
of sections was analysed), t is the average section thickness, a(p) is the area associated
with each grid point, and Ps is the total number of points hitting kidney tissue.
Total glomerular number (Nglom, kid) was estimated using physical dissectors as described
in detail previously (19). In brief, total glomerular number was estimated using:
Nglom, kid = 10 x 10 x PS/2PF x Q-
where 10 was the inverse of the first sampling fraction (1/10 of the slices) and 10 was the
inverse of the second section sampling fraction (1/10 of the sections), PS/2PF was the
fraction of the section area used for counting glomeruli, and Q- was the actual number of
glomeruli counted. Approximately, 100-150 glomeruli were counted for each kidney.
Mean glomerular tuft (Vglom) and renal corpuscle (Vcorp) volumes were estimated as:
Page 7 of 38

8
Vglom = VV(glom,kid)/NV(glom,kid),
and
Vcorp = VV(corp,kid) /NV(glom,kid),
Where VV(glom,kid) and NV(glom,kid) are volume density and numerical density, respectively, of
glomeruli in kidney and VV(corp,kid) is the volume density of renal corpuscles in kidney (31).
Total glomerular tuft (Vglom(tot)) and renal corpuscle (Vcorp(tot)) volumes were also estimated
as:
Vglom(tot) = Vglom x Nglom,kid
and
Vcorp(tot) = Vcorp x Nglom,kid
Noradrenaline Assay
In offspring at birth and 5 weeks of age noradrenaline content was determined in the left
kidney by HPLC, as previously described (43).
Statistical Analysis
One-way ANOVA with tukey post-hoc comparisons were performed to test for differences
between the groups on the control day in the mothers (see Table 1) and to compare
differences in glomerular number and noradrenaline content between the groups of
offspring. One-way repeated ANOVA was used to test for statistical differences between
the maternal groups during gestation, with factors time (PT; GA0, GA18, GA28 and 5
weeks post-partum) and group (PG; sham vs. 2K-1W; sham vs. 2K-2W; or 2K-1W vs. 2K-
2W) and their interaction (PGT). P values were conservatively adjusted using the
Page 8 of 38

9
Greenhouse-Geisser correction (36). P < 0.05 was considered to be statistically
significant.
Results
Mothers
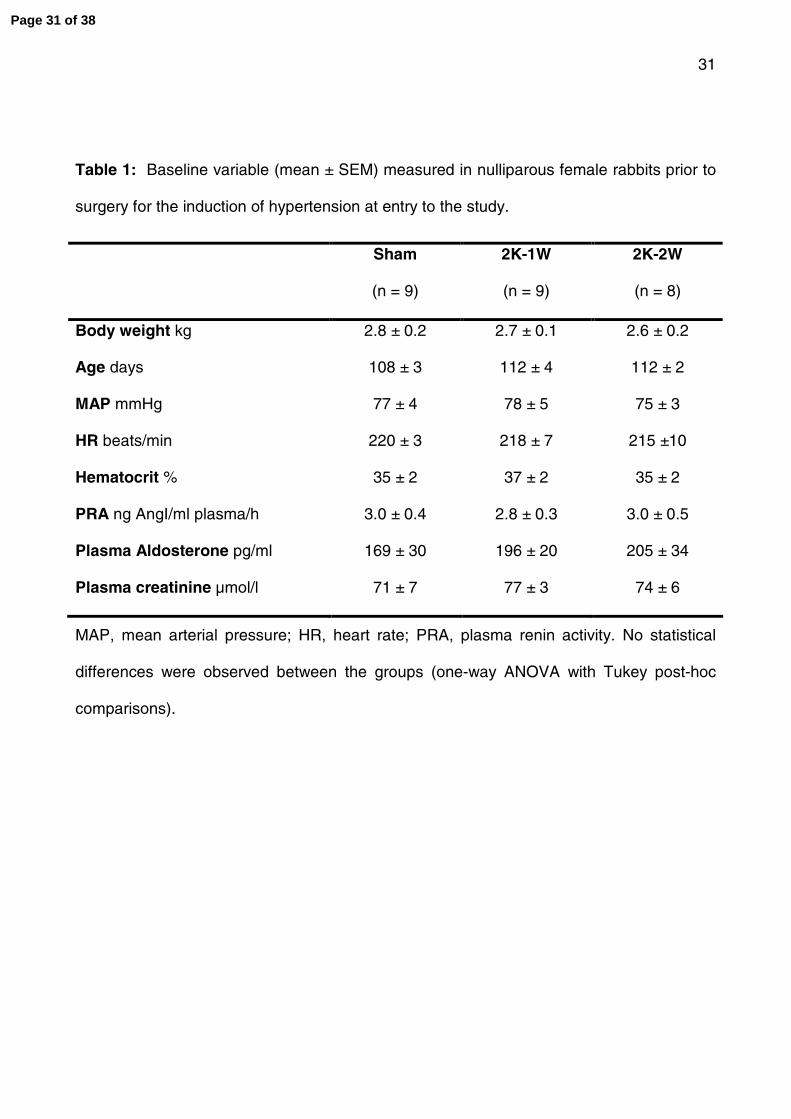
There were no significant differences in any variable measured between the groups of
female rabbits (prospective mothers) at entry to the study (Table 1).
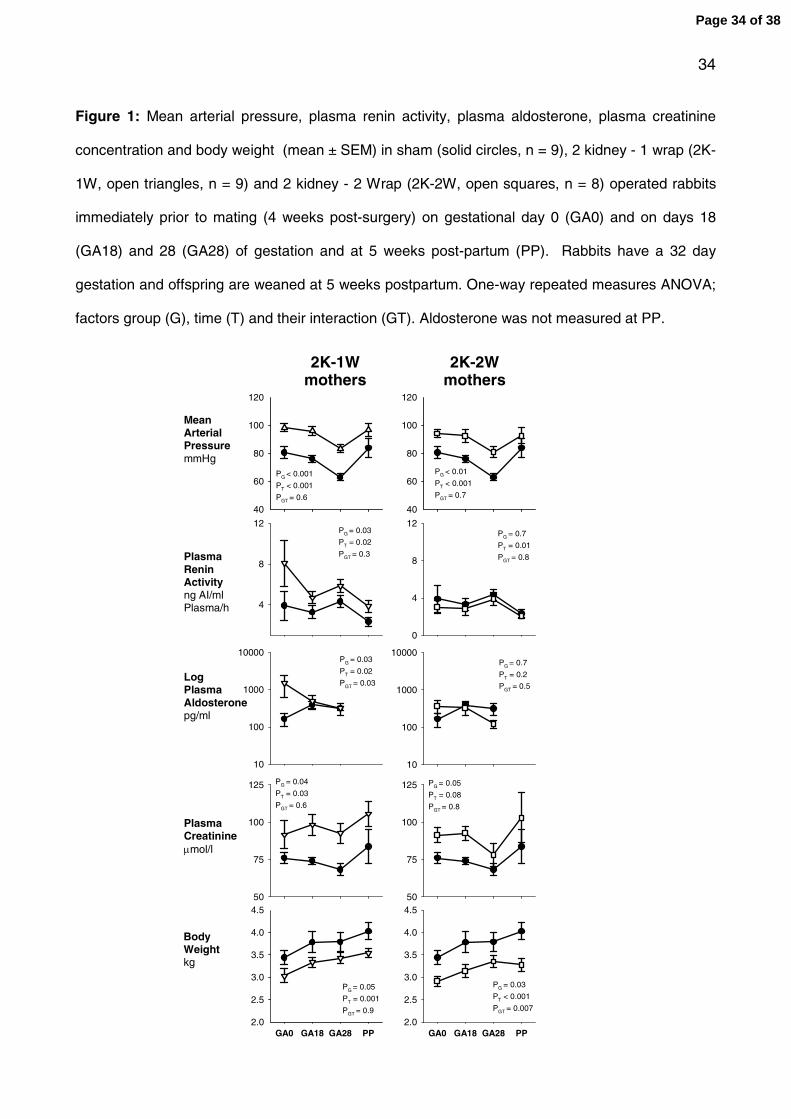
Mean arterial pressure (MAP) was significantly greater in both the 2K-1W (PG < 0.001) and
2K-2W (PG < 0.01) groups as compared to the sham-operated group throughout gestation
and the post-partum period (fig 1). Surprisingly, in the two models of renal hypertension
the increase in blood pressure was not different. MAP rose by 21 ± 5 mmHg (P < 0.001)
and 24 ± 3 mmHg (P < 0.001) in both the 2K-1W and 2K-2W groups 4 weeks post-
surgery, respectively. Comparison between the 2K-1W and 2K-2W groups revealed no
statistical difference in MAP between the 2 models of hypertension at any time. There was
a significant dip in MAP throughout gestation in all groups reaching a nadir at GA28 (fig 1).
The decrease in MAP from GA0 to GA28 was of a similar degree being 18 ± 6 mmHg, 15
± 5 mmHg and 15 ± 2 mmHg in sham, 2K-1W and 2K-2W groups, respectively. MAP
rebounded to pre-pregnancy levels at 5 weeks post-partum in all groups following birth (fig
1). Heart rate was not significantly different between the groups throughout the study.
Hematocrit was not significantly different between the groups though a significant fall in
hematocrit (~4%) was observed during gestation in all groups.
Plasma renin activity was significantly greater in the 2K-1W hypertensive mothers
throughout gestation and the post-partum period as compared to the sham-operated group
(fig 1, PG = 0.03). There was no difference in the PRA levels between the 2K-2W and
Page 9 of 38

10
sham groups (fig 1). PRA was also significantly greater in the 2K-1W group as compared
to the 2K-2W group throughout gestation and the post-partum period (PG = 0.03). Plasma
aldosterone rose significantly during gestation in the sham-operated group increasing by
250 ± 110 % (P < 0.05) and 110 ± 87 % (P < 0.05) at GA18 and GA28 as compared to
GA0, respectively. The plasma aldosterone level was significantly increased at GA0 in the
2K-1W group (PG = 0.03) as compared to the sham group during pregnancy, but not in the
2K-2W mothers (fig 1)
Plasma creatinine levels were significantly elevated by ~15% in both models of
hypertension following surgery and throughout gestation, though the degree of increase
was not different between the 2 models as compared to the sham group (fig 1). The
degree of increase in plasma creatinine in the 2K-1W and 2K-2W groups was not further
altered during pregnancy (fig 1).
Body weights were not different between the three groups at the commencement of the
study (Table 1). All three groups of rabbits gained weight following surgery. Normotensive
sham rabbits gained significantly more weight (+610 ± 128 g) than the 2K-1W (+278 ± 41
g, P < 0.05) and 2K-2W (+320 ± 64 g, P < 0.05) hypertensive rabbits by GA0 following
surgery. Body weight remained significantly lower in the 2K-1W hypertensive mothers
than the sham-operated mothers throughout pregnancy, though the rate of increase was
not different; 0.52 ± 0.05 kg vs. 0.59 ± 0.04 kg, respectively from GA0 to PP (fig 1; PGT =
0.9). In the 2K-2W group, body weight was also significantly less (PG = 0.03), than the
sham-operated group throughout pregnancy. However, the rate of increase during
pregnancy was also significantly less in the 2K-2W (0.34 ± 0.05 kg) as compared to the
sham group, from GA0 to PP (fig 1; PGT = 0.007).
Page 10 of 38

11
Offspring
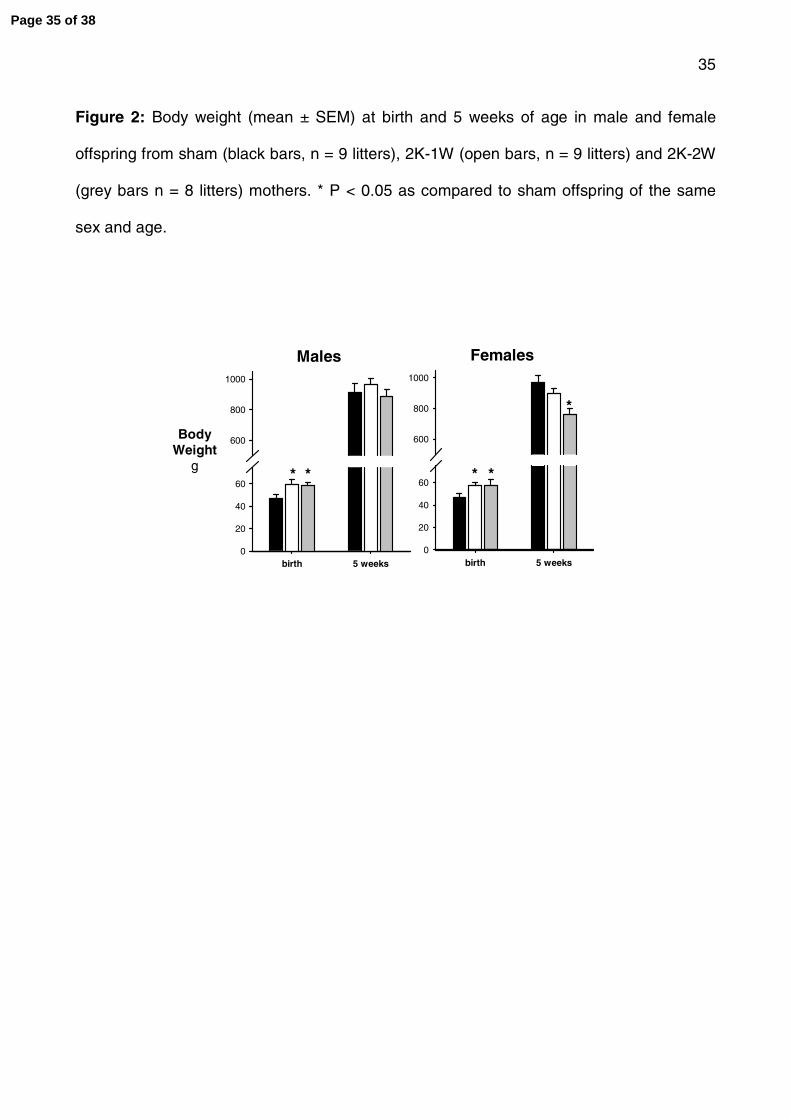
At birth and 5 weeks of age
A total of 149 kittens were born to 26 dams (Table 2). Birth weight was significantly greater
(~25%) in male and female offspring in both models of hypertensive mothers as compared
to the offspring of normotensive sham-operated mothers (fig 2). By 5 weeks of age
differences in body weight were no longer apparent in all offspring except the female
offspring of 2K-2W mothers, which were significantly smaller than the age matched
offspring of sham mothers (fig 2). Litter size and sex ratio for each group are given in
Table 2. Litter size was significantly reduced in the 2K-1W (P < 0.05), as compared to the
sham group offspring. Litter size was also smaller in the 2K-2W group offspring but did not
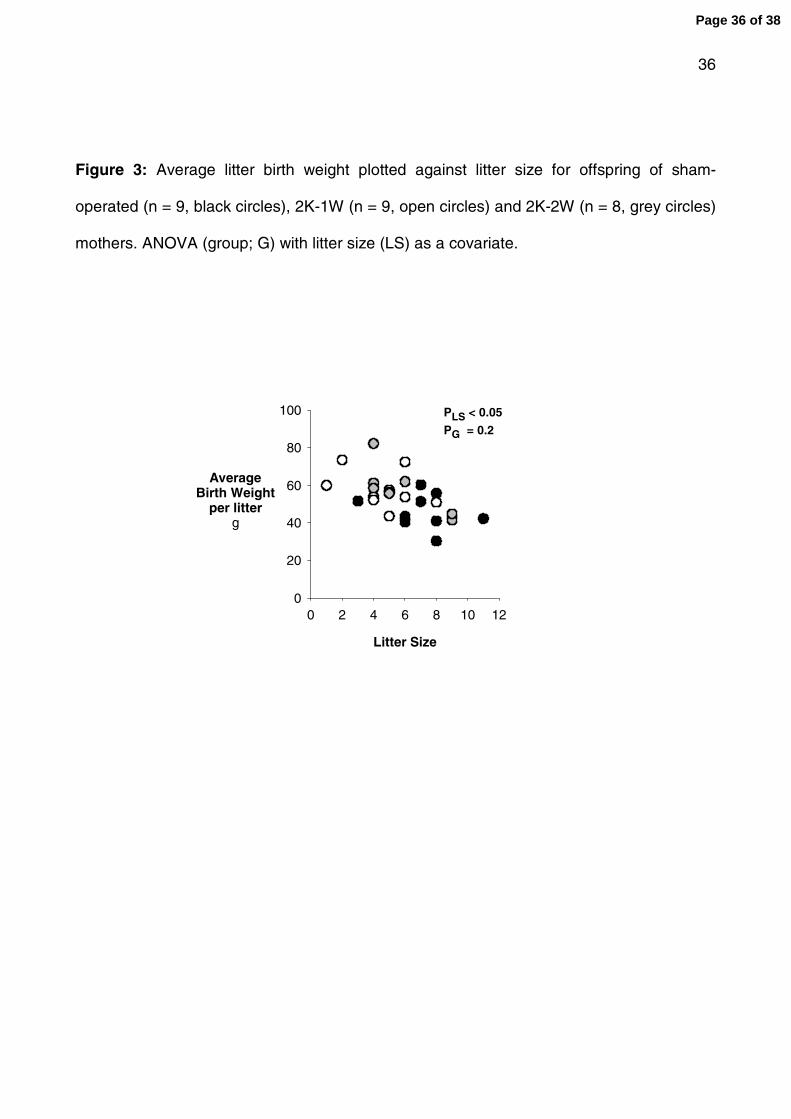
reach statistical significance (Table 2). When litter size was included as a covariate in the
analysis of birth weight, a significant effect of litter size was observed but birth weight was
no longer different between the groups (fig 3). Organ to body weight ratios were not
significantly different at birth in offspring of both models of hypertension as compared to
the offspring of sham-operated mothers (data not shown). There was no difference in the
ratio of male to female offspring between the groups (Table 2).
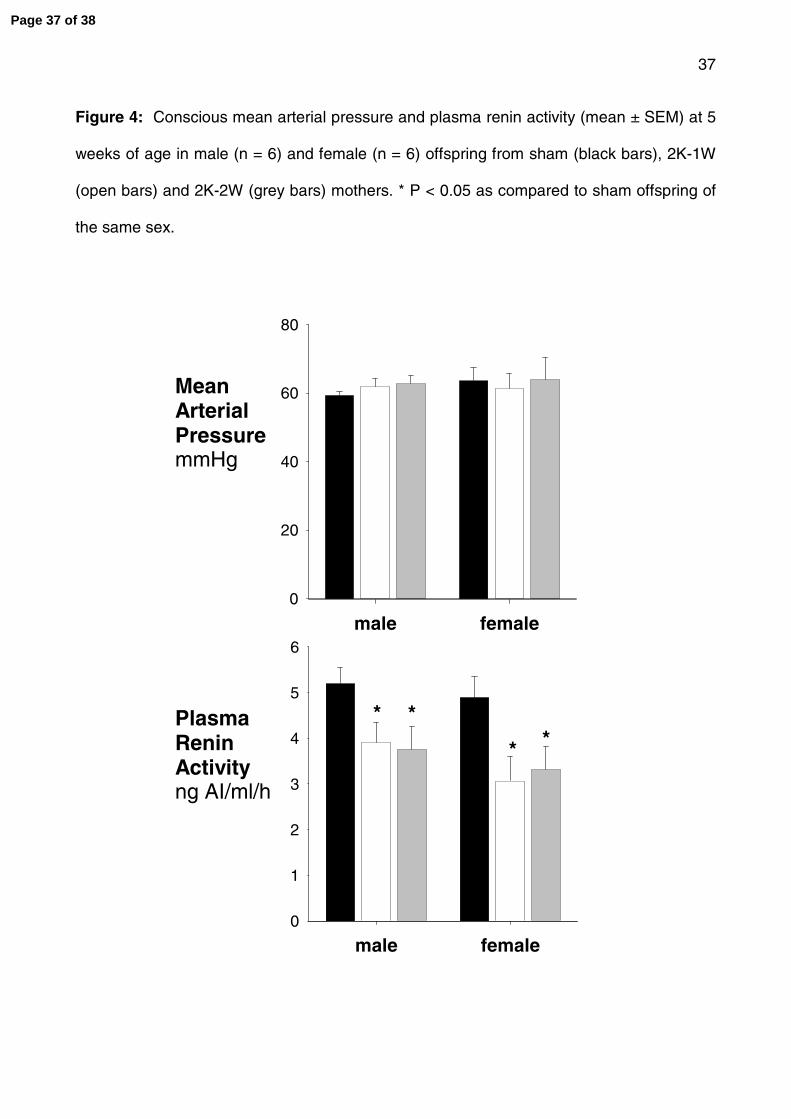
Conscious intra-arterial blood pressure was not significantly different between the sham,
2K-1W and 2K-2W offspring at 5 weeks of age (fig 4). PRA was significantly reduced in
both the 2K-1W (~28%; P < 0.05) and 2K-2W (~27%; P < 0.05) offspring at 5 weeks of age
as compared to the offspring of sham-operated mothers (fig 4).
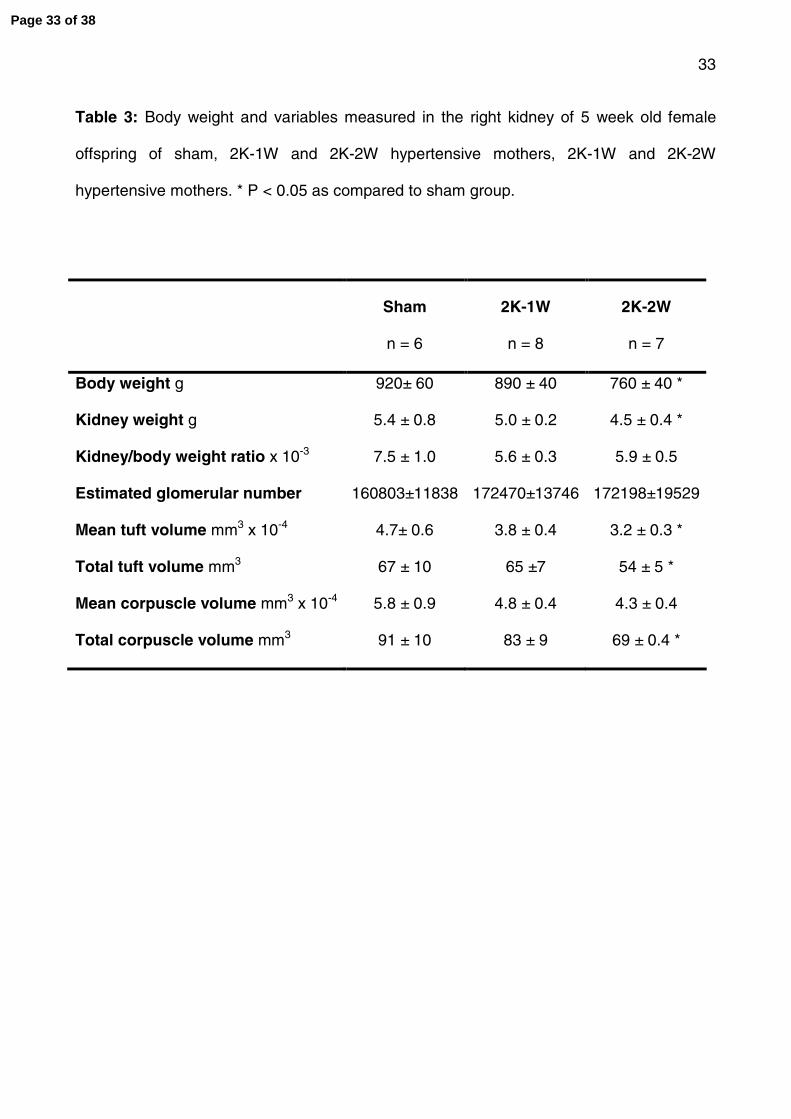
Glomerular number
Glomerular number was determined in right kidneys from female offspring at 5 weeks of
age; an age when nephrogensis is complete in the rabbit. Estimated nephron number
Page 11 of 38

12
averaged 160,803 ± 11,838, 172,470 ± 13,746 and 172,198 ±19,529 in the sham, 2K-1W
and 2K-2W female offspring, not significantly different. Glomerular tuft and corpuscle
volumes were also not significantly different between the 2K-1W and sham groups (Table
3). However, mean glomerular tuft volume (P= 0.04), total glomerular tuft volume (P =
0.04) and total corpuscle volume (P = 0.04) were significantly smaller in the 2K-2W as
compared to the sham offspring (Table 3). Mean corpuscle volume was smaller but this
did not reach statistical significance (P = 0.1).
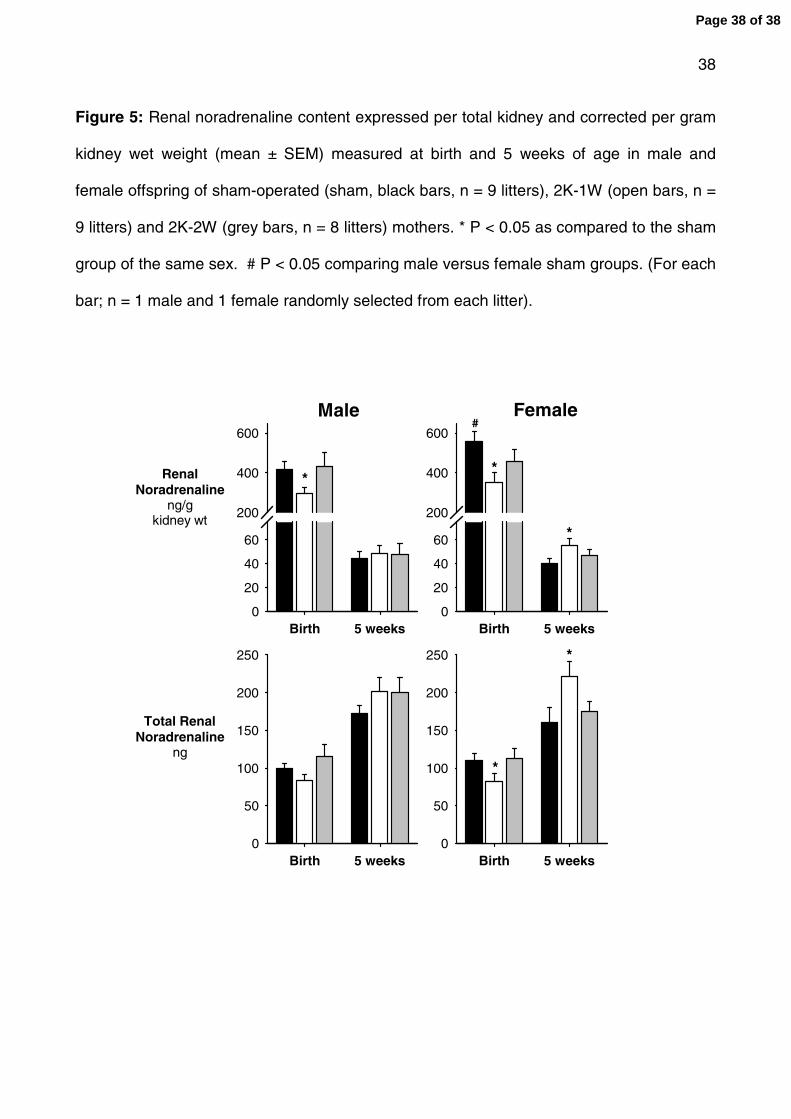
Renal noradrenaline content
Renal noradrenaline content was determined as an index of renal sympathetic innervation
density. At birth total noradrenaline content was ~20% lower in female (P = 0.02) and
~15% lower in male (not significant; P = 0.2) 2K-1W offspring as compared to sham
offspring. When corrected for kidney weight the decrease in noradrenaline was ~30% (per
g KW) in the female (P = 0.01) and male (P = 0.04) 2K-1W offspring compared to sham
offspring (fig 5). At 5 weeks of age both total noradrenaline and noradrenaline per gram
kidney weight was ~40% higher (P = 0.05) in female but not male 2K-1W offspring
compared to sham offspring. (fig 5). No significant differences in noradrenaline content
were seen between the 2K-2W and sham offspring (fig 5). At birth renal noradrenaline
content was greater in female than male offspring of sham-operated mothers (fig 5). By 5
weeks of age noradrenaline content was on longer significantly different between the
sexes in the sham offspring (fig 5).
Discussion
MAP was elevated throughout pregnancy in both 2K-1W and 2K-2W models of
hypertension. Activation of the renin-angiotensin-system (RAS) was also evident, with
Page 12 of 38

13
PRA and plasma aldosterone significantly increased in the 2K-1W, but not the 2K-2W
mothers. These two models of chronic maternal hypertension differentially affected the
offspring. Birth weight was not reduced in offspring from either model of maternal
hypertension, but renal noradrenaline content (an index of renal innervation) was
increased in the 5 week old female 2K-1W offspring, while a reduction in glomerular
volume (indicative of filtration surface area) was indicated in the 5 week old female 2K-2W
offspring. Potentially, both these alterations in offspring development may increase the risk
of hypertension in later life.
Mothers
Several maternal factors may have influenced fetal and post-natal development, some of
which were differentially effected in the models of renal hypertension utilized in the current
study. The similar degree of hypertension (~20 mmHg) in the two models was unexpected,
as was the indication that renal function was reduced to a similar extent (~20% increase in
plasma creatinine levels) since we have previously reported in males an increased severity
of hypertension and reduction in renal function in the 2K-2W model as compared to the
2K-1W model (10, 11, 15). The reasons for the similar increase in blood pressure in these
models in the females are unknown, though it is well recognised that hypertension is
attenuated in females as compared to males in many experimental models of hypertension
(49). This increase in blood pressure may influence fetal development and programme
adult blood pressure. However, against this argument a previous study in which blood
pressure was increased by central infusion of aldosterone during pregnancy, did not
increase blood pressure in adult offspring (27). Compromised renal function, with altered
extracelluar fluid homeostasis in maternal plasma may in turn influence fetal development
(21). In many cases, human hypertension is accompanied by renal insufficiency, which
Page 13 of 38

14
may contribute to disturbances in extracellular fluid homeostasis and an altered hormonal
milieu, as observed in our model.
Differential changes in the renin-angiotensin-aldosterone system were observed in the 2
models of hypertension. The 2K-1W mothers had elevated PRA and plasma aldosterone
levels during gestation while the 2K-2W mothers did not. Neither renin or angiotensin II
cross the placenta (5). However, the uteroplacental circulation has a local RAS that plays
important roles in placental angiogenesis and in modulating placental production of
cytokines, growth factors and vasoactive substances, which also influence fetal
development (47). It is our premise that due to the elevated maternal RAS activity, the
placental RAS may be down regulated, thereby altering production of placental factors
important for both placental and fetal development. It is also possible that elevated
Angiotensin II (Ang II) may result in reduced uterine blood flow. Previous studies where
chronic Ang II was infused into pregnant rabbits (4) and ewes (41) have confirmed this.
Evidence also suggests that uteroplacental perfusion is reduced in humans (53) and
animal models (30, 39) with chronic hypertension. Normally, the uterine artery is
particularly insensitive to Ang II due to the predominance of angiotensin type 2 receptors
(AT2R) during pregnancy (52); however following chronic Ang II infusion uterine artery
AT2R density decreases which may reduce uterine blood flow (6). Thus placental nutrient
transfer may be affected due to reduced uterine blood flow via this mechanism during
pregnancy when maternal Ang II levels are increased.
Ang II has also been shown to decrease 11beta-hydroxysteroid dehydrogenase (11-
βHSD2) in cultured human placental cells acting via AT2 receptors (34). If 11-βHSD2 is
decreased in our model this would potentially increase fetal exposure to maternal
glucocorticoids, which have been shown to influence the growth and development of the
Page 14 of 38

15
fetus (18). Aldosterone can cross the placenta and may therefore influence fetal
development directly. Minuth and colleagues (44) have investigated differentiation of the
beta-type intercalated cells in the collecting ducts of newborn rabbit kidney by labeling the
cultured epithelia with peanut agglutinin (PNA). They demonstrated that incubation of new
born kidney collecting duct with aldosterone increased the number of PNA labelled cells to
72% as compared to 8% in the control. Although the concentration of aldosterone used in
the culture was much higher than normal physiological levels, one can speculate that long
term exposure to elevated aldosterone (as seen in our 2K-1W model) may result in altered
renal development.
Another potential mechanism by which fetal development may have been altered is
through malnutrition. The hypertensive mothers were relatively undernourished compared
to the normotensive sham mothers as less weight was gained following surgery to induce
hypertension. This was aggravated in the 2K-2W mothers, and the rate of increase in
body weight during pregnancy was less in the 2K-2W mothers and this was associated
with lower post-natal growth rates in the offspring of these pregnancies. Nutrient restriction
is the most commonly used model of an adverse intrauterine environment and has been
shown to programme adult hypertension in many studies (33) and may contribute to
programming of offspring in our studies, particularly in the 2K-2W model.
Thus chronic renal hypertension is a complex model in which many adverse stimuli may
potentially have an adverse impact on fetal development with long-term consequences for
the adult. The rabbit as a model in these studies has many advantages. We have shown
that the hemodynamic changes that occur in a rabbit during pregnancy very closely
resemble those that occur in human pregnancy. The rabbit is also comparable to the
human in that its placentation is similar. The rabbit, like humans has a discoid
Page 15 of 38

16
hemochorial placenta, the primary difference between the two being that the exchange
surface is of a villous nature in the human placenta, but is labyrinthine in the rabbit
placenta (9, 23). Each fetus has its own individual placenta with a single trophoblast layer
separating maternal and fetal blood in the placental labyrinth (9). The visceral yolk sac (a
pre-placental organ) and the extraembryonic membranes more closely resemble those in
the human than do those of rats and mice (23). Also, the sequence of organ development
(Carnegie stages) is similar in the rabbit and human (23). PRA in the sham operated
normotensive rabbits increased towards late gestation (P < 0.08). Again this is similar to
the situation during normal pregnancy in humans, sheep and rats, when PRA rises
progressively, reaching a peak at birth and decreasing thereafter (54). Aldosterone levels,
in line with the elevated PRA also increased towards late gestation in the normotensive
rabbits, in agreement with studies in humans (55).
Offspring
Hypertensive pregnancies in humans, even mild hypertension, are generally associated
with reduced birth weight (22, 53), though there are reports of increased birth weight (59-
62). It is arguable in this study as to whether the offspring from both models of maternal
hypertension had an increased birth weight or the hypertensive mothers had smaller sized
litters and therefore relatively larger offspring. Taking litter size into account in the
statistical analysis as a covariate removed the difference in body weight between the
groups. This supports the conclusion that smaller litter size was responsible for the relative
increase in birth weight.
In our original study we demonstrated that at 10 weeks of age there was no difference in
MAP (in both sexes) and suppression of PRA in offspring of 2K-1W mothers (13). We now
report a similar situation at 5 weeks of age, with no difference in blood pressure and
Page 16 of 38

17
reduced PRA in the offspring of 2K-1W mothers. In addition we demonstrated similar
findings in the offspring of 2K-2W mothers. Whether the reduced PRA reflects altered
development and/or function of the renin-angiotensin system or a compensatory response
counteracting a rise in blood pressure in the offspring is unknown.
In the female offspring of 2K-1W mothers, renal noradrenaline content was reduced by
~20% at birth but was increased by ~40% at 5 weeks of age, providing evidence of altered
renal sympathetic nerve growth and development. These alterations in noradrenaline
content occurred predominantly in female offspring, which corresponds with our earlier
demonstration that adult blood pressure was increased in female not male 2K-1W
offspring at 30 weeks of age (13). No differences in renal noradrenaline content were
observed in the offspring of the 2K-2W mothers at birth or 5 weeks of age. This suggests
that activation of the renin-angiotensin-aldosterone system in the 2K-1W model of
hypertension, not the increase in maternal arterial pressure, may have impinged upon the
development of the renal sympathetic nerves. In the male offspring similar trends were
seen in noradrenaline content at birth, but were no longer apparent by 5 weeks of age. A
possible explanation for this sexually-dimorphic effect may be that the ontogeny of renal
sympathetic innervation is different in males and females as suggested by the higher renal
noradrenaline content at birth in females as compared to males from normotensive
mothers.
Given the compelling evidence implicating the renal sympathetic nerves in the
development of essential hypertension (20, 28, 46, 50), altered growth of the renal nerves
may underlie the programming of high blood pressure in adult offspring in the 2K-1W
model of maternal hypertension. In further support of this hypothesis, altered renal nerve
development; with increased noradrenaline content being observed from as early as 1
Page 17 of 38

18
week of age, has been demonstrated in spontaneously hypertensive rats (25). Also, renal
denervation abolished hypertension in low-birth weight offspring from pregnant rats with
reduced uterine perfusion demonstrating a causative role for the renal nerves in the
development of programmed hypertension (1). However, another study in offspring from
pregnancies with reduced uterine perfusion demonstrated no effect on adult blood
pressure or activity of the sympathetic nervous system (29).
If the kidney’s of female offspring of 2K-1W mothers are hyper-innervated one might
expect that renin release be enhanced, however this was not the case PRA was reduced
in these offspring. We offer two possible explanations for this finding. Firstly, neural
stimulation of PRA may be increased but other mechanisms may be counter-regulating
this effect (ie tubulo-glomerular feedback) in the pre-hypertensive phase in this model.
Secondly, evidence suggests that at least 2 populations of functionally distinct nerves
innervate the kidney (Type I and II) (14, 16, 17, 37, 38). We speculate that if it is Type I
nerves that are increased in the female offspring (predominantly innervating the smooth
muscle of the afferent arteriole) this might also explain why PRA is not increased.
However, there is some evidence to suggest that the renin-angiotensin system is playing a
role in the development of the hypertension at least in the maintenance phase since at 30
weeks of age PRA is normal when you might expect PRA to be suppressed due to the
increase in arterial pressure (13).
How the renin-dependent 2K-1W hypertensive maternal environment might influence renal
nerve growth is not known as few studies have examined the development of the renal
sympathetic nerves in disease models or for that matter under normal conditions (2, 48).
However, much work has focused on the growth and survival of postganglionic
sympathetic neurons in the broader setting of the periphery (7, 26, 35). Briefly, neuronal
Page 18 of 38

19
network development is guided by a finely orchestrated sequence of attractive and
repulsive molecular cues that determine axon extension, neuron survival or removal and
generation of dendrites, branches (arborisation) or synapses. These molecular cues, (e.g.
vascular endothelial growth factor, nerve growth factor, bone morphogenic proteins, Wnts,
Ephrins and Slits to name a few (7, 26, 35) maybe up or down regulated or miss-timed
causing an alteration in final development of the sympathetic nervous system.
The rabbit, like the rat and mouse is altricial (development continues during the postnatal
period), thus completion of nephrogenesis occurs around 2-3 weeks after birth (42). A
reduction in nephron number has consistently been associated with the programming of
adult hypertension in other models (8, 33, 56-58). Glomerular number was estimated,
using unbiased stereology, only in female rabbits in this study, as we previously found that
only female rabbits had a significant increase in arterial blood pressure by 30 weeks of age
(13). We demonstrated that total glomerular number was not reduced in female offspring
of either model of chronic hypertension indicating that a reduced nephron endowment is
not involved in the programming of hypertension in offspring of hypertensive mothers.
However, Bowman’s capsule and glomerular capillary tuft volumes were decreased in
female offspring of 2K-2W mothers. Therefore female offspring of 2K-2W mothers had the
same number of glomeruli but these glomeruli were on average smaller in size, indicative
of a reduction in filtration surface area. A reduction in filtration surface area at birth has
been postulated to predispose an individual to hypertension in later life (32, 40). Thus
there is evidence of altered renal development in the female offspring of 2K-2W mothers,
however considering these animals also demonstrated a reduced bodyweight at 5 weeks
of age, the significance of the reduced filtration surface area for adult cardiovascular
disease remains to be seen.
Page 19 of 38

20
In conclusion, birth weights of offspring from two models of hypertension were not
reduced. The fact that maternal blood pressure was increased by a similar level in these
models but that PRA was only elevated in the 2K-1W mothers may contribute to the
differential effects on offspring development observed. We can only speculate whether it
was the hypertension, activation of the maternal renin-angiotensin system or their
combination that drives programming of high blood pressure in the offspring of 2K-1W
mothers. We suggest that permanent structural and/or functional changes in the renal
sympathetic control of kidney function in offspring of 2K-1W hypertensive mothers may
contribute to the programming of hypertension in adulthood. The reduction in postnatal
growth and glomerular volume (indicative of a reduced filtration surface area) in the 2K-2W
model of chronic maternal hypertension, warrants future investigation, as it has not been
determined if hypertension is programmed in these offspring as adults. Hypertension
during pregnancy affects 1 in 10 women with the cause in the majority of cases unknown
(51). Our studies show that individuals may be differentially affected during fetal
development depending on the aetiology of the maternal hypertension.
Page 20 of 38

21
Acknowledgements:
This study was supported by the National Health and Medical Research Council of
Australia Grant 236821 and a National Foundation of Australia grant-in-aid G03M1143.
Page 21 of 38

22
References
1. Alexander BT, Hendon AE, Ferril G, and Dwyer TM. Renal denervation abolishes
hypertension in low-birth weight offspring from pregnant rats with reduced uterine
perfusion. Hypertension 45: 754-758, 2005.
2. Barajas L and Liu L. The renal nerves in the newborn rat. Pediatr Nephrol 7: 657-
666, 1993.
3. Barker DJP. Mothers, babies and health in later life. Edinburgh: Churchill
Livingstone, 1998.
4. Binder ND, Laird MR, and Faber JJ. Interrelationships between the renin
angiotensin system and uteroplacental blood flow--a recent perspective. Reprod Fertil Dev
7: 1437-1442., 1995.
5. Broughton Pipkin F, Benjamin N, and Macallan C. Placental transfer of large
angiotensin fragment in the guinea pig. Am J Obstet Gynecol 128: 904-906, 1977.
6. Burrell JH, Hegarty BD, McMullen JR, and Lumbers ER. Effects of gestation on
ovine fetal and maternal angiotensin receptor subtypes in the heart and major blood
vessels. Exp Physiol 86: 71-82, 2001.
7. Carmeliet P and Tessier-Lavigne M. Common mechanisms of nerve and blood
vessel wiring. Nature 436: 193-200, 2005.
8. Celsi G, Kistner A, Aizman R, Eklof AC, Ceccatelli S, de Santiago A, and
Jacobson SH. Prenatal dexamethasone causes oligonephronia, sodium retention, and
higher blood pressure in the offspring. Pediatr Res 44: 317-322., 1998.
9. Del Mar Yllera M, Alexandre-Pires GM, and Cifuentes JM. Placenta:
Page 22 of 38

23
regularization of neovascularization. Microvascularization pattern of the rabbit term
placenta. Microsc Res Tech 60: 38-45, 2003.
10. Denton KM and Anderson WP. Role of angiotensin II in renal wrap hypertension.
Hypertension 7: 893-898., 1985.
11. Denton KM, Anderson WP, and Korner PI. Renal blood flow and glomerular
filtration rate in renal wrap hypertension in rabbits. J Hypertens 1: 351-355., 1983.
12. Denton KM, Anderson WP, and Sinniah R. Effects of angiotensin II on regional
afferent and efferent arteriole dimensions and the glomerular pole. Am J Physiol Regul
Integr Comp Physiol 279: R629-638., 2000.
13. Denton KM, Flower RL, Stevenson KM, and Anderson WP. Adult rabbit offspring
of mothers with secondary hypertension have increased blood pressure. Hypertension 41:
634-639., 2003.
14. Denton KM, Luff SE, Shweta A, and Anderson WP. Differential neural control of
glomerular ultrafiltration. Clin Exp Pharmacol Physiol 31: 380-386, 2004.
15. Denton KM, Maduwegedera DWK, Flower RL, and Anderson WP. Effects of
angiotensin converting enzyme inhibtion in two-kidney, one-wrap hypertensive rabbits.
FASEB Journal March: 1164-APS, 5330, 2003.
16. Denton KM, Shweta A, and Anderson WP. Preglomerular and postglomerular
resistance responses to different levels of sympathetic activation by hypoxia. J Am Soc
Nephrol 13: 27-34., 2002.
17. Denton KM, Shweta A, Flower RL, and Anderson WP. Predominant
postglomerular vascular resistance response to reflex renal sympathetic nerve activation
during ANG II clamp in rabbits. Am J Physiol Regul Integr Comp Physiol 287: R780-786,
Page 23 of 38

24
2004.
18. Dodic M, Moritz K, and Wintour EM. Prenatal exposure to glucocorticoids and
adult disease. Arch Physiol Biochem 111: 61-69, 2003.
19. Douglas-Denton R, Moritz KM, Bertram JF, and Wintour EM. Compensatory
renal growth after unilateral nephrectomy in the ovine fetus. J Am Soc Nephrol 13: 406-
410, 2002.
20. Esler M. Sympathetic nervous system: contribution to human hypertension and
related cardiovascular diseases. J Cardiovasc Pharmacol 26: S24-28., 1995.
21. Feig DI, Nakagawa T, Karumanchi SA, Oliver WJ, Kang D-H, Finch J, and
Johnson RJ. Hypothesis: Uric acid, nephron number, and the pathogenesis of essential
hypertension. Kidney Int 66: 281-287, 2004.
22. Ferrer RL, Sibai BM, Mulrow CD, Chiquette E, Stevens KR, and Cornell J.
Management of mild chronic hypertension during pregnancy: a review. Obstet Gynecol 96:
849-860., 2000.
23. Foote RH and Carney EW. The rabbit as a model for reproductive and
developmental toxicity studies. Reprod Toxicol 14: 477-493, 2000.
24. Fowden AL, Giussani DA, and Forhead AJ. Intrauterine programming of
physiological systems: causes and consequences. Physiology (Bethesda) 21: 29-37,
2006.
25. Gattone VHn, Evan AP, Overhage JM, and Severs WB. Developing renal
innervation in the spontaneously hypertensive rat: evidence for a role of teh sympathetic
nervous system in renal damage. J Hypertens 8: 423-428, 1990.
26. Glebova NO and Ginty DD. Growth and survival signals controlling sympathetic
Page 24 of 38

25
nervous system development. Annu Rev Neurosci 28: 191-222, 2005.
27. Gomez-Sanchez EP and Gomez-Sanchez CE. Maternal hypertension and
progeny blood pressure: role of aldosterone and 11beta-HSD. Hypertension 33: 1369-
1373., 1999.
28. Grisk O and Rettig R. Interactions between the sympathetic nervous system and
the kidneys in arterial hypertension. Cardiovasc Res 61: 238-246, 2004.
29. Jansson T and Lambert GW. Effect of intrauterine growth restriction on blood
pressure, glucose tolerance and sympathetic nervous system activity in the rat at 3-4
months of age. J Hypertens 17: 1239-1248., 1999.
30. Karlsson K, Ljungblad U, and Lundgren Y. Blood flow of the reproductive system
in renal hypertensive rats during pregnancy. Am J Obstet Gynecol 142: 1039-1044., 1982.
31. Kett MM, Alcorn D, Bertram JF, and Anderson WP. Glomerular dimensions in
spontaneously hypertensive rats: Effects of AT1 antagonism. J Hypertens 14: 107-113,
1996.
32. Kett MM and Bertram JF. Nephron endowment and blood pressure: what do we
really know? Curr Hypertens Rep 6: 133-139, 2004.
33. Langley-Evans SC. Fetal origins of adult disease. Br J Nutr 81: 5-6., 1999.
34. Lanz B, Kadereit B, Ernst S, Shojaata K, Causevic M, Frey B, Frey FJ, and
Mohaupt MG. Angiotensin II regulates 11B-hydroxysteroid dehydrogenase type 2 via AT2
receptors. Kidney International 64: 970-977, 2003.
35. Low LK and Cheng HJ. A little nip and tuck: axon refinement during development
and axonal injury. Curr Opin Neurobiol 15: 549-556, 2005.
36. Ludbrook J. Repeated measurements and multiple comparisons in cardiovascular
Page 25 of 38

26
research. Cardiovasc Res 28: 303-311., 1994.
37. Luff SE, Hengstberger SG, McLachlan EM, and Anderson WP. Distribution of
sympathetic neuroeffector junctions in the juxtaglomerular region of the rabbit kidney.
Journal of the autonomic nervous system 40: 239 -254, 1992.
38. Luff SE, Young SB, and Anderson WP. Two structurally distinct types of
sympathetic axons innervating the rabbit renal cortex differ in their peptide content. Proc
Aust Neurosci Soc 11: 17, 2001.
39. Lumbers ER, Burrell JH, Stevens AD, and Weir BA. Effects of one-clip, one-
kidney hypertension in chronically catheterized pregnant ewes. Clin Exp Pharmacol
Physiol 24: 336-343., 1997.
40. McKenzie H, Lawler EV, and Brenner B. Congenital oligonephropathy; the flaw in
essential hypertension? Kidney Int 100: S30-s34, 1996.
41. McMullen JR, Gibson KJ, Lumbers ER, and Burrell JH. Selective down-
regulation of AT2 receptors in uterine arteries from pregnant ewes given 24-h intravenous
infusions of angiotensin II. Regul Pept 99: 119-129, 2001.
42. McVary KT and Maizels M. Urinary obstruction reduces glomerulogenesis in the
developing kidney: a model in the rabbit. J Urol 142: 646-651; discussion 667-648, 1989.
43. Medvedev OS, Esler MD, Angus JA, Cox HS, and Eisenhofer G. Simultaneous
determination of plasma noradrenaline and adrenaline kinetics. Responses to
nitroprusside-induced hypotension and 2-deoxyglucose-induced glucopenia in the rabbit.
Naunyn-Schmiedeberg's Arch Pharmacol 341: 192-199, 1990.
44. Minuth WW, Fietzek W, Kloth S, Aigner J, Herter P, Rockl W, and Kubitza M.
Aldosterone modulates PNA binding cell isoforms within renal collecting duct epithelium.
Page 26 of 38

27
Kidney Int 44: 537-544, 1993.
45. Moritz KM, Dodic M, and Wintour EM. Kidney development and the fetal
programming of adult disease. Bioessays 25: 212-220., 2003.
46. Orth SR, Amann K, Strojek K, and Ritz E. Sympathetic overactivity and arterial
hypertension in renal failure. Nephrol Dial Transplant 16: 67-69., 2001.
47. Poisner AM. The human placental renin-angiotensin system. Front
Neuroendocrinol 19: 232-252, 1998.
48. Pupilli C, Gomez RA, Tuttle JB, Peach MJ, and Carey RM. Spatial association of
renin-containing cells and nerve fibers in developing rat kidney. Pediatr Nephrol 5: 690-
695, 1991.
49. Reckelhoff JF. Gender differences in the regulation of blood pressure.
Hypertension 37: 1199-1208, 2001.
50. Ritz E, Adamczak M, and Zeier M. Kidney and hypertension-causes. Update 2003.
Herz 28: 663-667, 2003.
51. Roberts JM, Pearson G, Cutler J, and Lindheimer M. Summary of the NHLBI
Working Group on Research on Hypertension During Pregnancy. Hypertension 41: 437-
445., 2003.
52. Rosenfeld CR. Mechanisms regulating angiotensin II responsiveness by the
uteroplacental circulation. Am J Physiol Regul Integr Comp Physiol 281: R1025-1040,
2001.
53. Shah DM. Perinatal implications of maternal hypertension. Semin Pediatr Neurol 8:
108-119., 2001.
54. Skinner SL. The renin system in fertility and normal human pregnancy. In: The
Page 27 of 38

28
renin-angiotensin system (1st ed.), edited by Robertson JI and Nicholls MG. London:
Gower Medical Publishing, 1993, p. 50.51-50.16.
55. Wintour EM, Coghlan JP, Oddie CJ, Scoggins BA, and Walters WA. A
sequential study of adrenocorticosteroid level in human pregnancy. Clin Exp Pharmacol
Physiol 5: 399-403, 1978.
56. Wintour EM, Moritz KM, Johnson K, Ricardo S, Samuel CS, and Dodic M.
Reduced nephron number in adult sheep, hypertensive as a result of prenatal
glucocorticoid treatment. J Physiol 549: 929-935, 2003.
57. Woods LL, Ingelfinger JR, Nyengaard JR, and Rasch R. Maternal protein
restriction suppresses the newborn renin-angiotensin system and programs adult
hypertension in rats. Pediatr Res 49: 460-467., 2001.
58. Woods LL, Weeks DA, and Rasch R. Programming of adult blood pressure by
maternal protein restriction: role of nephrogenesis. Kidney Int 65: 1339-1348, 2004.
59. Xiong X, Demianczuk NN, Buekens P, and Saunders LD. Association of
preeclampsia with high birth weight for age. Am J Obstet Gynecol 183: 148-155, 2000.
60. Xiong X, Demianczuk NN, Saunders LD, Wang FL, and Fraser WD. Impact of
preeclampsia and gestational hypertension on birth weight by gestational age. Am J
Epidemiol 155: 203-209, 2002.
61. Xiong X and Fraser WD. Impact of pregnancy-induced hypertension on birthweight
by gestational age. Paediatr Perinat Epidemiol 18: 186-191, 2004.
62. Xiong X, Mayes D, Demianczuk N, Olson DM, Davidge ST, Newburn-Cook C,
and Saunders LD. Impact of pregnancy-induced hypertension on fetal growth. Am J
Obstet Gynecol 180: 207-213, 1999.
Page 28 of 38

29
Table and Figure Legends
Table 1: Baseline variables (mean ± SEM) measured in nulliparous female rabbits prior to
surgery for the induction of hypertension at entry to the study. MAP, mean arterial
pressure; HR, heart rate; PRA, plasma renin activity. No statistical differences were
observed between the groups (one-way ANOVA with Tukey post-hoc comparisons).
Table 2: Total number of births and stillbirths, average litter size, range of litter size and
sex ratio of litters from sham-operated normotensive, 2K-1W and 2K-2W hypertensive
mothers. Values are mean ± SEM. * P < 0.05 as compared to sham operated
normotensive mothers.
Table 3: Body weight and variables measured in the right kidney of 5 week old female
offspring of sham, 2K-1W and 2K-2W hypertensive mothers, 2K-1W and 2K-2W
hypertensive mothers. * P < 0.05 as compared to sham group.
Figure 1: Mean arterial pressure, plasma renin activity, plasma aldosterone, plasma
creatinine concentration and body weight (mean ± SEM) in sham (solid circles, n = 9), 2
kidney - 1 wrap (2K-1W, open triangles, n = 9) and 2 kidney - 2 Wrap (2K-2W, open
squares, n = 8) operated rabbits immediately prior to mating (4 weeks post-surgery) on
gestational day 0 (GA0) and on days 18 (GA18) and 28 (GA28) of gestation and at 5
weeks post-partum (PP). Rabbits have a 32 day gestation and offspring are weaned at 5
weeks postpartum. One-way repeated measures ANOVA; factors group (G), time (T) and
their interaction (GT). Aldosterone was not measured at PP.
Figure 2: Body weight (mean ± SEM) at birth and 5 weeks of age in male and female
offspring from sham (black bars, n = 9 litters), 2K-1W (open bars, n = 9 litters) and 2K-2W
(grey bars, n = 8 litters) mothers. * P < 0.05 as compared to sham offspring of the same
Page 29 of 38

30
sex and age.
Figure 3: Average litter birth weight plotted against litter size for offspring of sham-
operated (n = 9, black circles), 2K-1W (n = 9, open circles) and 2K-2W (n = 8, grey circles)
mothers. ANOVA (group; G) with litter size (LS) as a covariate.
Figure 4: Conscious mean arterial pressure and plasma renin activity (mean ± SEM) at 5
weeks of age in male (n = 6) and female (n = 6) offspring from sham (black bars), 2K-1W
(open bars) and 2K-2W (grey bars) mothers. * P < 0.05 as compared to sham offspring of
the same sex.
Figure 5: Renal noradrenaline content expressed per total kidney and corrected per gram
kidney wet weight (mean ± SEM) measured at birth and 5 weeks of age in male and
female offspring of sham-operated (sham, black bars, n = 9 litters), 2K-1W (open bars, n =
9 litters) and 2K-2W (grey bars, n = 8 litters) mothers. * P < 0.05 as compared to the sham
group of the same sex. # P < 0.05 comparing male versus female sham groups. (For each
bar; n = 1 male and 1 female randomly selected from each litter).
Page 30 of 38
31
Table 1: Baseline variable (mean ± SEM) measured in nulliparous female rabbits prior to
surgery for the induction of hypertension at entry to the study.
Sham
(n = 9)
2K-1W
(n = 9)
2K-2W
(n = 8)
Body weight kg 2.8 ± 0.2 2.7 ± 0.1 2.6 ± 0.2
Age days 108 ± 3 112 ± 4 112 ± 2
MAP mmHg 77 ± 4 78 ± 5 75 ± 3
HR beats/min 220 ± 3 218 ± 7 215 ±10
Hematocrit % 35 ± 2 37 ± 2 35 ± 2
PRA ng AngI/ml plasma/h 3.0 ± 0.4 2.8 ± 0.3 3.0 ± 0.5
Plasma Aldosterone pg/ml 169 ± 30 196 ± 20 205 ± 34
Plasma creatinine µmol/l 71 ± 7 77 ± 3 74 ± 6
MAP, mean arterial pressure; HR, heart rate; PRA, plasma renin activity. No statistical
differences were observed between the groups (one-way ANOVA with Tukey post-hoc
comparisons).
Page 31 of 38

32
Table 2: Total number of births and stillbirths, average litter size, range of litter size and
sex ratio of litters from sham-operated normotensive, 2K-1W and 2K-2W hypertensive
mothers. Values are mean ± SEM. * P < 0.05 as compared to sham operated
normotensive mothers.
Sham 2K-1W 2K-2W
Mothers 9 9 8
Total offspring 61 44 44
Total still births 2 3 2
Average litter size 6.8 ± 0.7 4.8 ± 0.6 * 5.5 ± 1.9
Litter size range 3 - 11 1- 8 4 - 9
Sex ratio (male : female) 4 ± 1 : 3 ± 1 3 ± 1 : 3 ± 1 3 ± 1 : 3 ± 1
Page 32 of 38
33
Table 3: Body weight and variables measured in the right kidney of 5 week old female
offspring of sham, 2K-1W and 2K-2W hypertensive mothers, 2K-1W and 2K-2W
hypertensive mothers. * P < 0.05 as compared to sham group.
Sham
n = 6
2K-1W
n = 8
2K-2W
n = 7
Body weight g 920± 60 890 ± 40 760 ± 40 *
Kidney weight g 5.4 ± 0.8 5.0 ± 0.2 4.5 ± 0.4 *
Kidney/body weight ratio x 10-3 7.5 ± 1.0 5.6 ± 0.3 5.9 ± 0.5
Estimated glomerular number 160803±11838 172470±13746 172198±19529
Mean tuft volume mm3 x 10-4 4.7± 0.6 3.8 ± 0.4 3.2 ± 0.3 *
Total tuft volume mm3 67 ± 10 65 ±7 54 ± 5 *
Mean corpuscle volume mm3 x 10-4 5.8 ± 0.9 4.8 ± 0.4 4.3 ± 0.4
Total corpuscle volume mm3 91 ± 10 83 ± 9 69 ± 0.4 *
Page 33 of 38
34
Figure 1: Mean arterial pressure, plasma renin activity, plasma aldosterone, plasma creatinine
concentration and body weight (mean ± SEM) in sham (solid circles, n = 9), 2 kidney - 1 wrap (2K-
1W, open triangles, n = 9) and 2 kidney - 2 Wrap (2K-2W, open squares, n = 8) operated rabbits
immediately prior to mating (4 weeks post-surgery) on gestational day 0 (GA0) and on days 18
(GA18) and 28 (GA28) of gestation and at 5 weeks post-partum (PP). Rabbits have a 32 day
gestation and offspring are weaned at 5 weeks postpartum. One-way repeated measures ANOVA;
factors group (G), time (T) and their interaction (GT). Aldosterone was not measured at PP.
0
4
8
12
40
60
80
100
120
PlasmaReninActivityng AI/mlPlasma/h 4
8
12
2K-1Wmothers
2K-2Wmothers
MeanArterialPressuremmHg
40
60
80
100
120
PlasmaCreatinineµmol/l
50
75
100
125
50
75
100
125
GA0 GA18 GA28 PP2.0
2.5
3.0
3.5
4.0
4.5
GA0 GA18 GA28 PP
BodyWeightkg
2.0
2.5
3.0
3.5
4.0
4.5
PG = 0.03
PT = 0.02
PGT = 0.3
PG = 0.05
PT = 0.001
PGT = 0.9
PG = 0.05
PT = 0.08
PGT = 0.8
PG = 0.04
PT = 0.03
PGT = 0.6
PG = 0.7
PT = 0.01
PGT = 0.8
PG < 0.01
PT < 0.001
PGT = 0.7
PG < 0.001
PT < 0.001
PGT = 0.6
PG = 0.03
PT < 0.001
PGT = 0.007
LogPlasmaAldosteronepg/ml
10
100
1000
10000PG = 0.03
PT = 0.02
PGT = 0.03
PG = 0.7
PT = 0.2
PGT = 0.5
10
100
1000
10000
Page 34 of 38
35
Figure 2: Body weight (mean ± SEM) at birth and 5 weeks of age in male and female
offspring from sham (black bars, n = 9 litters), 2K-1W (open bars, n = 9 litters) and 2K-2W
(grey bars n = 8 litters) mothers. * P < 0.05 as compared to sham offspring of the same
sex and age.
Males
birth 5 weeks
BodyWeight
g
0
20
40
60
600
800
1000
Females
birth 5 weeks0
20
40
60
600
800
1000
* * * *
*
Page 35 of 38
36
Figure 3: Average litter birth weight plotted against litter size for offspring of sham-
operated (n = 9, black circles), 2K-1W (n = 9, open circles) and 2K-2W (n = 8, grey circles)
mothers. ANOVA (group; G) with litter size (LS) as a covariate.
Litter Size
0 2 4 6 8 10 12
AverageBirth Weight
per litterg
0
20
40
60
80
100 PLS < 0.05
PG = 0.2
Page 36 of 38
37
Figure 4: Conscious mean arterial pressure and plasma renin activity (mean ± SEM) at 5
weeks of age in male (n = 6) and female (n = 6) offspring from sham (black bars), 2K-1W
(open bars) and 2K-2W (grey bars) mothers. * P < 0.05 as compared to sham offspring of
the same sex.
male female
MeanArterialPressuremmHg
0
20
40
60
80
male female
PlasmaReninActivityng AI/ml/h
0
1
2
3
4
5
6
* *
* *
Page 37 of 38
38
Figure 5: Renal noradrenaline content expressed per total kidney and corrected per gram
kidney wet weight (mean ± SEM) measured at birth and 5 weeks of age in male and
female offspring of sham-operated (sham, black bars, n = 9 litters), 2K-1W (open bars, n =
9 litters) and 2K-2W (grey bars, n = 8 litters) mothers. * P < 0.05 as compared to the sham
group of the same sex. # P < 0.05 comparing male versus female sham groups. (For each
bar; n = 1 male and 1 female randomly selected from each litter).
Birth 5 weeks
Total RenalNoradrenaline
ng
0
50
100
150
200
250
Birth 5 weeks0
50
100
150
200
250
*
*
Male
Birth 5 weeks
RenalNoradrenaline
ng/gkidney wt
0
20
40
60
200
400
600
Female
Birth 5 weeks0
20
40
60
200
400
600
*
#
*
*
Page 38 of 38




















