
Parent-Offspring Conflict in Budgerigars
39
PARENT-OFFSPRING CONFLICT IN BUDGERIGARS by JUDY STAMPS'), ANNE CLARK, PAT ARROWOOD and BARBARA KUS (Department of Zoology, University of California, Davis, California 95616 U.S.A.) (With 7 Figures) (Acc. 23-VII-1984) Introduction Ten years ago, TRIVERS (1974) introduced the concept of genetic conflict between parents and offspring over the allocation of parental investment, and suggested that offspring might be selected to ask for (and receive) more investment than parents were selected to give. Subsequent theoretical studies have verified TRIVERS' assumption that such selfish offspring behaviour could evolve, even though the spread of selfish traits would tend to decrease the reproductive success of parents with selfish offspring (BLICK, 1977; PARKER & MACNAIR, 1978, 1979; STAMPS et al., 1978; MACNAIR & PARKER, 1979; METCALF et al., 1979; HARTUNG, 1980; ROBINSON, 1980; STAMPS, 1980; FELDMAN & ESHEL, 1982). However, this spate of theoretical studies has generated very few empirical studies on parent-offspring conflict, and most of these have focused on a single "prediction" of the theory [e. g. , that social hymenopterans will have a female-biased reproductive output (TRIVERS & HARE, 1976; NOONAN, 1981; PAGE & METCALF, 1982), or that conflict over food will increase near the end of the parental care period (e.g., DAVIES, 1976; BERGER, 1979; BURGER, 1981)]. Ten years after TRIVERS' paper, detailed studies of parent-offspring interactions from the viewpoint of parent-offspring conflict theory are still almost nonexistent (but see BENGTSSON& RYDEN, 1983; MocK, submitted, 2). The paucity of studies of parent-offspring conflict is not due to a lack of interest; TRIVERS (1974) is frequently cited and discussed. The problem may lie in the scarcity of testable hypotheses in the original TRIVERS paper. In order to prove the existence of genetic conflict, sensu TRIVERS, one would have to i) identify a species in which the offspring could affect 1) This research has been supported by grants from the Institute of Ecology and the Agricultural Experiment Station, University of California, Davis.
Transcript of Parent-Offspring Conflict in Budgerigars

PARENT-OFFSPRING CONFLICT IN BUDGERIGARS
by
JUDY STAMPS'), ANNE CLARK, PAT ARROWOOD and BARBARA KUS
(Department of Zoology, University of California, Davis, California 95616 U.S.A.)
(With 7 Figures) (Acc. 23-VII-1984)
Introduction
Ten years ago, TRIVERS (1974) introduced the concept of genetic conflict between parents and offspring over the allocation of parental investment, and suggested that offspring might be selected to ask for (and receive) more investment than parents were selected to give. Subsequent theoretical studies have verified TRIVERS' assumption that such selfish
offspring behaviour could evolve, even though the spread of selfish traits would tend to decrease the reproductive success of parents with selfish
offspring (BLICK, 1977; PARKER & MACNAIR, 1978, 1979; STAMPS et al., 1978; MACNAIR & PARKER, 1979; METCALF et al., 1979; HARTUNG, 1980; ROBINSON, 1980; STAMPS, 1980; FELDMAN & ESHEL, 1982). However, this
spate of theoretical studies has generated very few empirical studies on
parent-offspring conflict, and most of these have focused on a single "prediction" of the theory [e. g. , that social hymenopterans will have a female-biased reproductive output (TRIVERS & HARE, 1976; NOONAN, 1981; PAGE & METCALF, 1982), or that conflict over food will increase near the end of the parental care period (e.g., DAVIES, 1976; BERGER, 1979; BURGER, 1981)]. Ten years after TRIVERS' paper, detailed studies of parent-offspring interactions from the viewpoint of parent-offspring conflict theory are still almost nonexistent (but see BENGTSSON & RYDEN, 1983; MocK, submitted, 2).
The paucity of studies of parent-offspring conflict is not due to a lack of
interest; TRIVERS (1974) is frequently cited and discussed. The problem may lie in the scarcity of testable hypotheses in the original TRIVERS
paper. In order to prove the existence of genetic conflict, sensu TRIVERS, one would have to i) identify a species in which the offspring could affect
1) This research has been supported by grants from the Institute of Ecology and the Agricultural Experiment Station, University of California, Davis.

2
the allocation of parental care (e.g., by begging) ii) determine if offspring consistently varied in the extent of their demands iii) determine if the care
provided by parents as a result of offspring demands fit the definition of
parental investment [e. g. , decreased the future reproductive success of the
parent (TRIVERS, 1972)] and iv) see if parents with very demanding off-
spring had lower lifetime reproductive success than comparable parents with less demanding offspring. Such a study is currently impractical,
given the difficulty in measuring all the required variables. For example, with respect to (iii), investigators have only just determined that raising
any type of offspring affects future maternal reproductive success (mam- mals : ALTMAN et al., 1978; FUCHS, 1982; CLUTTON-BROCK, 1982; birds:
KLUYVER et al., 1977; SMITH & ROFF, 1980). It will be some time before
the effects of raising selfish and nonselfish offspring can be accurately determined and compared.
In the meanwhile, several of the theoretical models of parent-offspring conflict suggest indirect ways of investigating this phenomenon in natural
systems. A key element of these models, which was implied, but not
developed in TRIVERS' formulations, is that behavioural mechanisms
resulting from genetic conflict would often involve measurable costs to
the participants in the interactions. For example, if offspring were selfish
and parents evolved a behavioural counterstrategy to selfish behaviour, this counterstrategy might involve neural, energetic, temporal, or other
costs in comparison to a "laissez-faire" parental strategy. Alternately, the behaviours offspring used to acquire extra parental investment could involve substantial energetic or survival costs to those offspring (PARKER & MACNAIR, 1978, 1979; STAMPS et al., 1978; STAMPS, 1980). For exam-
ple, several workers have proposed that offspring begging behaviour may be both energetically costly and risky, if the begging attracted predators
(DAWKINS, 1976; ZAHAVI, 1977; PARKER & MACNAIR, 1979; STAMPS,
1980). If parents required expensive begging behaviours before giving extra parental investment, then the high costs of begging would reduce
the net energetic benefit to the young of obtaining those extra resources
(see STAMPS el al., 1978). The existence of both costly parental counterstrategies, and costly off-
spring demand behaviours is predicted by parent-offspring conflict
theory (reviews in PARKER & MACNAIR, 1979; and STAMPS, 1980). On the
other hand, if parents and offspring were in genetic "agreement" about the optimal amount of parental investment per offspring, then selection
should favor those parental care strategies and offspring demand behaviours which were most efficient for both participants. Hence, one

3
would have indirect evidence supporting the existence of genetic parent-
offspring conflict if a) one could demonstrate that an effective parental
counterstrategy existed, but that it was more costly to the parent than a
laissez-faire strategy and/or b) that offspring which demanded and
received extra resources grew and survived as well as offspring with lower
begging and resource accrual rates.
Budgerigars (Melopsittacus undulatus) afforded an excellent system for a
study of parent offspring conflict. The budgerigar's highly asynchronous
hatching pattern yielded pronounced differences in nestling sizes, and
hence competitive abilities, within broods. Sex differences in parental roles and feeding behaviour, as well as considerable inter-family varia-
tion in the paternal behaviour, resulted in two types of families: those in
which parents used an effective counterstrategy to counter offspring demands, and those in which offspring controlled food allocation via their
begging behaviour. Hence, it was possible to compare i) the efficiency of
a counterstrategic parental "style" with that of a laissez-faire "style" and ii) the growth, fledge ages and fledge weights of offspring under
parental control with those of offspring demanding and receiving extra
investment. A further advantage of this species is that it exhibits normal
reproductive patterns and breeding behaviours in captivity (see Discus-
sion). Hence it was possible to keep all individuals under comparable conditions, and to set resources such as food, water and nestboxes at
superabundant levels. In the field, parental "quality" and habitat
"quality" probably affect budgerigar breeding success; both factors
could be partially controlled in our laboratory study. For example, in the
field, distance from nest site to feeding site could affect food delivery rates; in captivity all pairs nested close to the food supply. Similarly,
parental differences in foraging efficiency and the ability to compete for
preferred nestsites were minimized by providing food and nestboxes ad
lib.
Methods
I. General methods.
Domestic budgerigars (Melopsittacus undulatus) were studied in large (2.5 x 3.2 x 6.3 m) outdoor flight cages at the University of California, Davis, California from 1978 to 1981. Each cage contained between 12 and 16 adults; during the breeding season of March to September densities temporarily increased up to 31 birds/cage, as young of the year fledged. These young were systematically removed to make room for fledglings of subse- quent broods. Adults and fledglings were marked with black Nyanzol-D dye marks on the head, neck, breast, wings, etc., and were banded for permanent identification. The birds were provided with a standard mix of canary and millet seeds ad lib., water with poultry

4
vitamins, as well as assorted vegetables (corn, chard, spinach etc.) and wild seed-bearing annuals.
Nestboxes with plexiglass backs and one-way viewers made with "Gila" brand car window film allowed observers to record behaviour within the nestboxes while remaining outside of the flight cages and invisible to the birds. Observations were recorded on tape and timed with an LED stopwatch. Observation periods usually lasted 0.5 hr (80% of 323 observations periods). Observations on a given nest were usually spaced at 2-3 day inter- vals, from the date the first offspring hatched to the date when the last offspring fledged; most new nestlings were observed within 24 hours of hatching. Nearly all (315) of the observations periods occurred during the morning activity period 0700-1100 hr. Inter- observer reliability scores could not be calculated because only one observer could watch any given box at a time, but comparisons of observations and recordings on the same boxes by different observers, and frequent discussions of methods and patterns revealed no major observer biases or differences. Stamps, Clark and Kus were responsible for 92 % of the observation periods; the remaining periods were covered by Arrowood and a highly trained graduate student.
A total of 20 nests were observed during the 1978, 1979 and 1981 breeding seasons for a total of 174 hr. Of these, 16 nests were observed intensively (X = 20 observation periods/ nest, X = 10.3 ± 3.4 hr per nest). Complete data from hatch to last fledge were available for 11 of these nests; in three other cases observations began mid-way into the nest cycle (first offspring 11 days old), and in two cases observations concentrated on the early por- tion of the nest cycle. The number of nestlings per nest varied from one to six in the observed nests. Data from small families (one and two nestlings) and very large families (six nestlings) were used in the analyses of parental feeding styles and nestling begging behaviours. However, tests of hypotheses relevant to parent-offspring conflict focused on eight nests with 3-5 offspring each (X = 11.0 ± 3.8 hr observation/nest). These nests are described in more detail in Results.
All nests in the flight cages were checked daily in the early morning. New eggs were date-marked and weighed to the nearest 0.1 g using Pesola scales, older eggs were checked, and any missing, broken or hatched eggs were noted. Hatchlings were weighed to the nearest 0.1 g, and marked on the head, neck, back or wings with gentian violet for identification during behavioural observations. Nestlings were reweighed and (if necessary) remarked each day, and were banded when they reached 10 g, after which they were weighed to the nearest 0.5 g.
Budgerigars in the flight cages rapidly habituated to the daily nest inspections; some females had to be removed manually from the boxes so that the contents could be weighed. Parents usually returned immediately to their boxes after the inspection. Even so, observers allowed at least 10 minutes to elapse between nest inspections and observa- tions of the nestboxes.
In addition to the twenty observed nests, egg and growth data were kept for a total of 94 other clutches in the same cages. These data were used to document the general reproduc- tive characteristics of budgerigars under our field cage conditions. Of these clutches, 19 were produced in 1978, 31 in 1979, 39 in 1980, and 5 in 1981.
II. Growth and fledging success.
Data on nestling growth and behaviour were compiled for four periods: Period 1: hatch date ( = day 1 ) to day 4, Period 2: day 5 to day 10, Period 3: day 11-22 and Period 4: day 23 to fledge date. For the purposes of this study, Periods 1 and 2 were usually combined (Period 1 & 2). Day 11 and day 22 were used to demarcate Periods 3 and 4 because preliminary analyses showed that most budgerigar nestlings achieved asymptotic weights at day 22, and day I1 corresponded to the age at which most nestlings achieved 50% of their asymptotic weight (CLARK, unpublished data).

5
Daily weights were used to compute growth rates for Period 1 & 2, Period 3 and Period 4. "Fledge weight" was the last recorded weight, taken within 24 hrs of fledging. Fledging was defined as a nestling leaving the nestbox; in the few cases when a nestling subsequent- ly returned to the nestbox, the first date it left was considered to be its fledge date.
Some analyses required relationships between weight variables (growth, fledge weights, weight at certain ages) and behavioural variables for Periods 1 & 2, 3 and 4. Such analyses required that our short observation periods accurately reflected the behaviour occurring over the nestling's entire growth period. In order to minimize possible errors due to inade- quate sampling, correlations between weight and behavioural data used only data from nestlings with at least four observation periods (2 hrs of observation) during the growth period. All nestlings exceeded this requirement for Period 1 & 2; 42/44 nestlings met or ex- ceeded the time requirement for Period 3, and 38/44 nestlings met or exceeded the time requirement for Period 4.
III. Behavioural observations.
This study focused on allo-feeding, begging and associated behaviours, although other ac- tivities and social interactions were also recorded. Allo-feeding in budgerigars is easily identified. As described by BROCKWAY (1964), budgerigar allo-feeding consists of i) head bobbing movements, followed by ii) beak grasping in which the bills are fitted together at longitudinally right angles, and iii) the regurgitation of food. A series of regurgitations received by a particular nestling was termed a feeding bout. Feeding bouts were con- sidered terminated when i) the parent stopped feeding any nestlings, ii) when the parent fed or attempted to feed a different nestling, iii) when the parent temporarily stopped feeding a particular nestling and engaged in self-preening, stretching or yawning, nest cleaning or nibbling other nestlings before returning to feed the original nestling, or iv) whenever 30 sec elapsed between sequential regurgitations to a particular nestling. Feeding bouts were in turn grouped into feeding episodes. A feeding episode began with the first feeding to any nestling, and was considered terminated when i) at least two min had elapsed between subsequent regurgitations to any nestlings or ii) the feeding parent left the nestbox between sequential regurgitations. If a feeding episode was still in pro- gress at the end of an observation period, we continued to record data until it was over, thus extending the duration of that observation period.
The time (to the nearest 5 sec) of all regurgitations from parents to young, from males to females, and from nestlings to other birds was measured. Preliminary analyses in- dicated that budgerigar parents always waited at least 3 sec between sequential regurgita- tions, and that most interfeed intervals were between 5 and 10 sec. Hence, there was ample time for the observers to see and record all of the regurgitations within each nestbox.
Begging in budgerigars consists of repeated high-pitched vocalizations, which in older nestlings are accompanied by various locomotor activities (see Results). Nestlings could beg before a feeding bout, or in the pauses between regurgitations after feeding had com- menced. Since we were primarily interested in the effect of begging on food allocation, we excluded cases in which nestlings vocalized during, but not prior to food delivery.
Because begging bouts varied in length, and because several nestlings often begged simultaneously, we concentrated on recording the number of begging bouts, rather than the total length of time each nestling spent in begging. A begging bout was defined as a series of begging calls given while facing a particular individual, and bouts were con- sidered to be terminated when i) the nestling stopped calling for at least one minute ii) the nestling redirected its calling to a different individual or iii) a nestling stopped calling when another bird interposed itself between the nestling and the potential feeder. Begging rates (bout per hr) were most accurate for families with only one nestling, and least ac- curate for large families when both parents were simultaneously feeding the nestlings. In such cases, we probably missed some bouts. In general, this type of bias would predict

6
results opposite to those statistically tested in the present study. For example, this bias would predict lower measured begging rates for families in which two parents simultaneously fed nestlings than in families in which one parent did all of the feeding. In fact, our results suggested, and statistics confirmed, that nestlings with two caretakers had higher begging rates than those with one feeding parent (see Results). Hence, if anything, methodological bias in begging rate would increase Type II statistical errors in the present study.
Results
I. Growth, feeding and begging behaviour in budgerigar families.
Hatching asynchrony, growth and fledging success.
Budgerigars in our flocks produced an average of 6.16 (S.D. ± 1.85) eggs
per clutch (N = 85 clutches) with an interval of 2.2 + 1.1 days between
successive eggs. Incubation began between the first and second egg, and
lasted 17.9+ 1.5 days (N = 176 hatched eggs). Additional asynchrony resulted from eggs which failed to hatch, and there was an interval of
2.6+ 1.5 days between the hatching of successive nestlings. For the 57
clutches which hatched at least one egg, the average brood size was
3.4+ 1.9 nestlings; 8.1 ± 3.2 days elapsed between the hatching of the
first and the last nestling. The size differences set up by hatching asyn-
chrony were maintained throughout the nestling period, and nestlings
fledged at 2.8 (± 2.5) day intervals (N = 49 clutches fledging young).
Hatching asynchrony led to appreciable differences in the sizes of
nestlings in the nest at any given time. For example, in a brood with 5
nestlings, the last nestling hatched at 1.1 g into a nest containing older
siblings ranging from 6.8 to 14 g (X = 10.4 g). In order to determine if the
youngest nestlings were disadvantaged by being in competition for food
with siblings so much larger than themselves, we compared the growth and fledging weights and ages of last hatched nestlings with i) those of
their oldest siblings and ii) the averages for all of their siblings. Although last hatched nestlings were underweight at day 11 relative to their older
siblings, they achieved comparable weights by day 23, and fledged at the
same ages and at significantly heavier weights than their siblings (Table
1). Hence there was no indication that last hatched young were disadvan-
taged by sharing a nest with much larger siblings. In most budgerigar families, there were negative relationships between
size at a given age and growth rate in the subsequent period. Twenty-
eight out of thirty-three families with three or more offspring had
negative Spearman rank correlations between size at day 11 and Period 3

7
growth, and only five had positive correlations (Binomial test, p < 0.001).
Similarly, twenty-nine families had negative correlations between size at
day 23 and Period 4 growth, while only two families had positive correla-
tions (Binomial test, p < 0.001). These negative correlations between size
and subsequent growth were the first indications that either parents or
offspring were influencing growth during the nestling period, such that
all of the siblings in a clutch eventually achieved comparable fledge
weights and ages.
TABLE 1. Weights and fledge ages of last hatched nestlings, in com-
parison to their older siblings (Thirty-three broods with 3 or more
offspring)
Nestling begging behaviour,.
Parents did not require very young hatchlings to beg before they would
feed them, and nestlings usually did not begin begging until the start of
Period 3 (X = 11.25 ± 4.4 days, N = 40 nestlings). In small nestlings, beg- ging consisted of vocalizations accompanied by waving of the head. Nest-
lings of approximately 6-8 days were able to stretch their necks up and
orient their heads in the parent's direction while wiggling their wings and
vocalizing. Later still, when nestlings became more ambulatory, they moved toward the parent, lifted their heads to reach the parent's beak,
attempted to grasp the parent's beak, and bobbed their heads up and
down while vocalizing (see BROCKWAY, 1964 for a description). Greater
size, agility and motor control allowed older nestlings to run towards the
parents, lunge for parent's beaks, and physically displace smaller off-
spring in proximity to parents (see below). Begging vocalizations seemed
to become louder as nestlings aged; while Period 1 & 2 nestlings could
barely be heard within 0.2 m of the nestboxes, begging Period 3 and 4
nestlings were audible from over 6 m away.
8
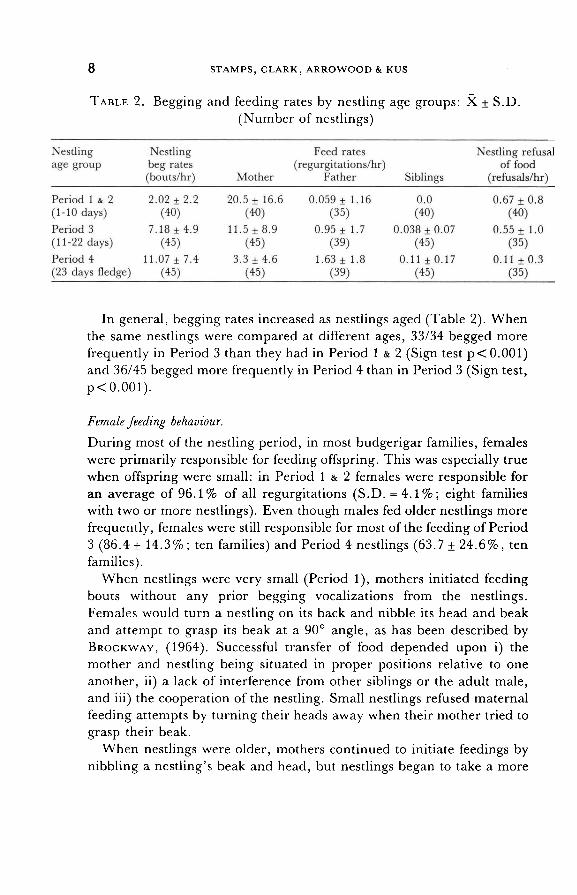
TABLE 2. Begging and feeding rates by nestling age groups: X ± S.D.
(Number of nestlings)
In general, begging rates increased as nestlings aged (Table 2). When
the same nestlings were compared at different ages, 33/34 begged more
frequently in Period 3 than they had in Period 1 & 2 (Sign test p < 0.001) and 36/45 begged more frequently in Period 4 than in Period 3 (Sign test,
p < 0.001).
Female feeding behaviour.
During most of the nestling period, in most budgerigar families, females
were primarily responsible for feeding offspring. This was especially true
when offspring were small: in Period 1 & 2 females were responsible for
an average of 96.1 % of all regurgitations (S.D. = 4.1 % ; eight families
with two or more nestlings). Even though males fed older nestlings more
frequently, females were still responsible for most of the feeding of Period
3 (86.4 + 14.3 % ; ten families) and Period 4 nestlings (63.7 ± 24.6 % , ten
families). When nestlings were very small (Period 1), mothers initiated feeding
bouts without any prior begging vocalizations from the nestlings. Females would turn a nestling on its back and nibble its head and beak
and attempt to grasp its beak at a 90° angle, as has been described by BROCKWAY, (1964). Successful transfer of food depended upon i) the
mother and nestling being situated in proper positions relative to one
another, ii) a lack of interference from other siblings or the adult male, and iii) the cooperation of the nestling. Small nestlings refused maternal
feeding attempts by turning their heads away when their mother tried to
grasp their beak.
When nestlings were older, mothers continued to initiate feedings by
nibbling a nestling's beak and head, but nestlings began to take a more

9
active role in the process. In response to the nibbling, nestlings would lift their heads to meet the parent, and sometimes begin to give begging vocalizations prior to beak grasping and food transfer. At this stage, nestlings refused feeding attempts by not lifting their heads in response to
nibbling, or by moving their heads away if the mother attempted to lift
them into feeding position. The importance of begging and nestling cooperation gradually in-
creased, so that by Period 3, most mothers required begging vocaliza-
tions before transferring food (see below). Females often continued to at-
tempt to initiate feedings by nibbling nestlings, and nestlings often con-
tinued to refuse feedings by actively evading maternal attempts to grasp their beaks. At this stage, nestlings also evaded maternal feeding at-
tempts by walking or running away from them.
By Period 4, maternal nibbling of beaks was rare, and offspring usual-
ly initiated feeding bouts by vigorous begging behaviours (see above). Two distinct types of behavioural conflict over food allocation between
mothers and nestling were observed in this study. First, there were the
cases in which mothers attempted to feed and offspring refused to accept the food. Second, there were cases in which offspring begged for food, and mothers refused to feed them. The general literature on avian
parent-offspring interactions suggested that the first type of conflict might be more common when nestlings were young, while the second type
might increase in importance when the nestlings were older (e.g. NORTON-GRIFFITHS, 1969; LBHRL, 1975; DAVIES, 1976; BENGTSSON &
RYDEN, 1981; HAFTORN, 1982). As expected, nestling refusals of food were most common when the
nestlings were young (Table 2). In Period 1 & 2 and Period 3 refusals oc-
curred at approximately the same rates (T = 0.55, 70 df, Table 2). However, Period 4 refusal rates were lower than in Period 1 & 2 (T = 3.5, 70 df, p < 0.001 ) or Period 3 (T = 2.47, 70 df, p < 0.01 ).
Conversely, mothers were less apt to refuse the demands of young
nestlings than those of older nestlings. Although mothers often fed Period
1 & 2 nestlings with no prior begging (see above), young nestlings that
begged more were fed more frequently than were quieter siblings of the
same age (nine out of nine families with three or more nestlings had
positive Spearman correlations between beg rates to the mother and feed
rates by the mother among Period 1 & 2 siblings: Binomial test,
p = 0.002). However, in Period 3 this responsiveness to offspring demands disappeared. In neither Period 3 nor Period 4 was there a
tendency within families for demanding offspring to be fed more often by

10
the mother than less demanding offspring at the same ages. Maternal
disregard of offspring begging was not complete, since most mothers re-
quired some begging from Period 3 offspring before they would feed
them. Nestlings 20-29 days old were compared during intervals when
they had been silent and when they had begged. Thirty-six out of 36
nestlings were fed more often by the mother after they had begged to her
then when they had been silent (p < 0.001, Sign test). In addition, by the
onset of Period 4 (day 23) no feedings were observed in which the off-
spring had not begged first. Since maternal feeding rates dramatically decreased as offspring aged while nestling beg rates increased over the
same period (Table 2), the result was a dramatic increase in the maternal
"refusal" rate as offspring aged. Given the spread of ages within budgerigar nests and the age-
dependence of both maternal responsiveness to begging and nestling beg-
ging rates, it should be no surprise that maternal allocation of food to in-
dividual nestlings on a daily basis was not correlated with the begging rates of those nestlings. Observation periods in which offspring begged and were fed were scored according to whether there was a positive or a
negative Spearman rank correlation between offspring begging rates and
maternal feeding rates. On 39 days there were positive relationships be-
tween begging and feeding, while on 40 days there were negative rela-
tionships. Three females had more positive than negative days, four
females had more negative than positive days, and three females had an
equal number of positive and negative days. Clearly, within an in-
dividual family on any given day, there was no tendency for nestlings
begging more often from females to be fed more often by them.
If female budgerigars did not feed according to offspring begging rate, what cues did they use? Available evidence indicates that females feed ac-
cording to some variable related to offspring age. Female budgerigars generally preferred to feed the youngest offspring
first. During the time when the entire brood was present in the nest (e.g., from last hatch to first fledge), we recorded the first offspring that a
female either beak-nibbled or fed for each feeding bout. Within eight out
of ten families, a maternal preference for younger chicks was reflected by positive Spearman correlations between hatch order and the number of
feeding bouts initiated by approaching that nestling first (Binomial test,
p = 0.055). Within three of these families, the Spearman rank correla- tions were significant at p < 0.01. Hence, most female budgerigars began
feeding bouts by approaching the youngest nestlings first, and only if
they refused food did the female go on to the older nestlings.

11
Feeding bout lengths and rates of feeding were also highly related to
offspring age. Both the number of regurgitations per feeding bout and the number of regurgitations per unit time declined as a function of offspring age, and both declined at a comparable rate for all offspring within the same broods. Feeding bout lengths averaged 9.26 regurgitations/bout for Period 1 & 2 nestlings (N = 40) nestlings; 3.01 regurgitations/bout for Period 3 nestlings (N = 35) and 2.18 regurgitations/per bout for Period 4
nestlings (N = 28). Twelve out of thirteen females had shorter average bout lengths for Period 3 than for younger nestlings (p = 0.002, Sign
test); and most females further reduced their average bout lengths for Period 4 as compared to Period 3 offspring (13/16 females, p = 0.011,
Sign test). Within a brood all offspring of a given age were treated the
same, and there was no relationship between birth order and either
feeding rates or bout lengths for any of the three periods. A steady decline in feeding rate with offspring age was also observed
(Table 2). Eleven out of 13 females fed Period 1 & 2 nestlings more fre-
quently than they fed those same nestlings in Period 3 (p = 0.011 Sign
test) and all of 16 females fed Period 4 nestlings less frequently than Period 3 nestlings (p < 0.001 Sign test). As in the case of bout length, there was no indication of any systematic change in feeding rates within families as a function of hatch order for any of the three periods.
These relationships between nestling age and maternal feeding behaviour suggested that some factor related to age might be used as a cue by female budgerigars. One possibility was offspring size. If size rather than age were important, then we would expect females to feed undersize offspring as if they were younger. Since feeding rates declined with offspring age, we looked at material feeding rates as a function of size when age was controlled. Size at day 11 was compared with maternal
feeding rates in the subsequent growth interval (Period 3); this age was chosen because both size at day 11 and maternal feeding rates in Period 3 were extremely variable both within and between families, and because later hatched nestlings were undersized at day 11 in comparison to their
siblings. A strong negative correlation was observed between offspring size at
day 11 and logio of the maternal regurgitation rate in Period 3 (r = -0.52, 36 df, p < 0.001, Fig. 1). Hence, females fed undersized 11 day olds almost 100 time more frequently than they fed the largest 11 day olds. An inverse relationship between size at 11 days and Period 3 maternal
feeding rate was observed within 7 out of 8 families (p = 0.04, Sign test), and within three families, the Spearman rank correlation coefficients be-

12
Fig. 1. Maternal feeding rate (regurgitations/hr) in Period 3 (age I 1-22 days) as a function of offspring weight (g) at 11 days.
tween size and feed rate were significant at p < 0.05. There was no in-
dication of a relationship between nestling begging rate to mothers and
size at day 11 (r = 0.04, 36 df; within families: 3 had positive nonsignifi- cant correlations between size at 11 days and beg rate, 5 had negative
nonsignificant correlations). Hence, undersized offspring were not being fed more because they were begging more often.
The preference for smallest offspring by female budgerigars was not
absolute. Several times offspring were lost from a brood, and in one case
we observed selective starvation of a youngest nestling. The pair in ques- tion hatched four young, and the female at first exhibited the normal
preference for, and high rates of feeding of, the fourth nestling. However, when the last nestling was four days old the female refused to feed it,
despite its vigorous begging, and instead began to preferentially feed the
next youngest (third) nestling. The fourth nestling continued to beg for
six more days, but was never observed to be fed again; it lost weight and

13
died on the twelfth day, while the female continued preferential feeding of the third nestling until it fledged.
Other observations suggest that a preference for smallest nestlings did
not require any prior experience with a brood. When one female was ex-
perimentally removed from a brood of four, her place was taken by a
"step mother". This second female had previously copulated with
several males, including the father of the brood, and had unsuccessfully
attempted to defend a box and incubate a brood. Although she had never
before been observed near the male's nestbox, within two days after the
first female's removal she moved into the box and began feeding his
nestlings. When presented with the half grown brood ranging from 13 to
19 days old, she immediately exhibited a preference for the younger
nestlings, and showed age dependent feeding rates and bout lengths com-
parable to those observed in females feeding their own broods.
As one might expect from the maternal disregard of begging and the
independence of bout lengths and birth order, females were not easily distracted if they were in the middle of a feeding bout with a particular
nestling. Interruptions of feeding bouts were often observed (231 obser-
vations, 12 families), and resulted from siblings (usually larger) interpos-
ing themselves between the mother and the nestling being fed. In most
such cases, females returned to the nestling previously being fed (148
cases) rather than shifting to the interrupting nestling (83 cases). Within
families, 9 out of 13 females were more apt to resume feeding the original
nestling than to feed the one which interrupted (p = 0.03, Sign test). This
maternal persistence was often time-consuming, as females separated from a nestling walked around the other siblings, ignoring their begging, returned to the previously fed nestling and resumed an interrupted bout.
Direct observations showed that female budgerigars faced with an ar-
ray of different-sized offspring simultaneously attended to both the size
and begging rate of each of their nestlings. We often saw a female initiate
a feeding bout by searching under a squirming pile of begging older nest-
lings until she found a nonbegging young nestling, nibble it to initiate a
feeding bout, and then transfer a long series of regurgitations to it while
fending off begging older siblings with her wings. Even when two or more
siblings were fed within seconds of each other, relative bout lengths and
feeding rates were as would be predicted from the nestling's ages. For ex-
ample, a female with a brood of five fed her begging 13 day old nestling
once, then evaded its begging efforts to reach and feed a nonbegging 7
day old 19 times. Hence, the complexities of female budgerigar feeding behaviour are evident even within the few minutes of a single feeding
episode.

14
Male feeding behaviour. r.
Male budgerigars usually allo-fed females, which then transferred food to
the nestlings, but some males also fed nestlings directly. When males fed
nestlings, they used the same behaviour patterns already described for
females. However, because males seldom fed very young nestlings (see
below), some behaviours, such as turning nestlings on their backs and
nibbling them prior to feeding were rarely observed in males.
In contrast to females, males tended to feed older offspring more often
than younger ones (Table 2). When average feed rates per period were
compared for individual males, there was no significant difference in
average feed rates for Period 1 & 2 and Period 3 offspring (X male feed
rate for Period 3 greater than for Period 1 & 2 offspring in 3 families, less
in 2 families, and in 7 families males didn't feed in either period). Most
males did feed Period 4 offspring at higher average rates than they fed
Period 3 offspring (11/12 males, p = 0.003 Sign test), and all males fed
Period 4 offspring more frequently, on average, than they had fed Period
1 & 2 offspring (N = 8 males, p = 0.004, Sign test). Males also differed from females in that their feeding bout lengths were
unrelated to offspring age. Male bout lengths were short for all offspring
(Regurgitations/bout: Period 1 and 2: 2.66 (N = 15 nestlings), Period 3:
1.63 (N = 19) Period 4: 1.86 (N = 23) and there were no significant dif-
ferences between average bout lengths for Periods 1 & 2, 3 or 4. In one
respect, however, males were similar to females. There was no tendency for male feeding rates or bout lengths to be related to offspring birth order
for any of the three offspring age groups. The average figures for male feeding tend to obscure the considerable
variance among males, some of which never fed their offspring directly, while others fed Period 4 nestlings up to 7.0/hr, and were responsible for
all of the regurgitations to some of their nestlings in Period 4. For conve-
nience in later tests, we focused on eight nests with comparable brood
sizes of 3-5 nestlings. In Period 4, four of these broods were rarely fed by the males (= female-fed families), while the other four were frequently fed by the males (= male-aided families, Fig. 2, Table 3). Additional data
on male feeding behaviour was available for eight other broods of dif-
ferent sizes (1-2 nestlings; 6 nestlings). Males that fed nestlings at a high rate in Period 4 also had fed their
nestlings in Period 3. Males' average feeding rates to Period 3 offspring were positively correlated with the average Period 4 feeding rates of those
same males (Spearman rank correlation coefficient = 0.91, n = 9,
p < 0.01 ). Period 4 and Period 1 & 2 average feeding rates were also
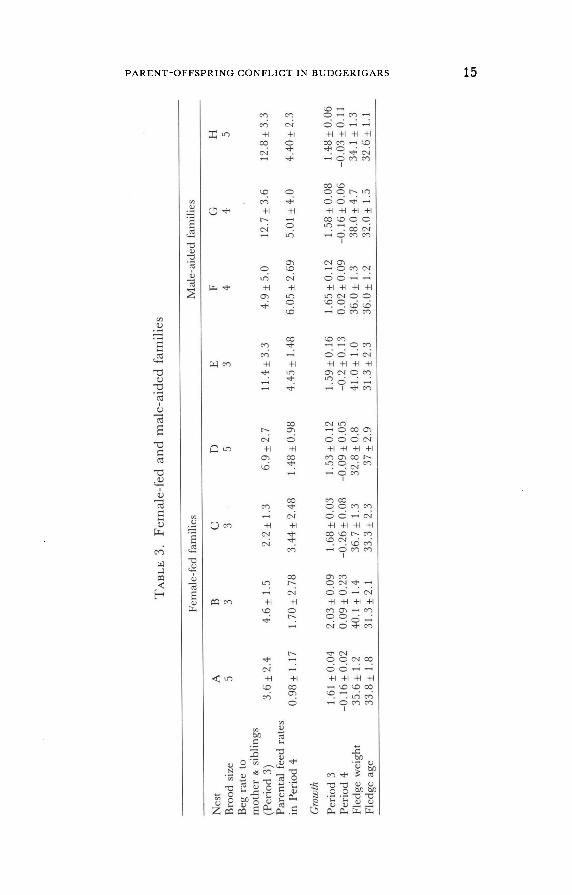
15

16
positively related (rs = 0.73, n=6, n.s.). Hence, the extent of male
feeding of Period 4 nestlings could be accurately predicted by observing their feeding behaviour earlier in the nest cycle (Periods 1, 2 and 3).
Another potentially useful predictor of male feeding behaviour was the
frequency with which males entered their nestboxes. For those nests with
two or more young, about half of male entries into their nestboxes resulted in the males' feeding their offspring (53 % of 188 entries resulted
in feeding of nestlings; X = 52 ± 20% for 9 males). The proportion of en-
tries resulting in feeding did not differ for male-aided and female-fed families (t test = 0.42, 7 df). However, the males of male-aided families
Fig. 2. Paternal feeding rates (regurgitations/hr) for Period 4 offspring in female-fed families (A-D) and male-aided families (E-H); X ± S. E. (number of nestlings).
entered the nestboxes while young were present more frequently than did
the males of female-fed families (male-aided families: 2.43 + 0.49
entries/hr; female-fed families: 0.47 ± 0.32 entries/hr, t = 6.64, 6 df,
p < 0.001 ). In fact, there was a strong positive correlation between male
entry rates and their average rate of feeding of Period 4 offspring
(r = 0.91, 7 df, p < 0.001 ). Hence, a male's inclination to feed offspring could be accurately assessed by observing the frequency with which he
entered his nestbox.
One possible reason for the considerable intermale variability in
feeding tendency was that all males responded equally to offspring beg-
ging, but that offspring in male-aided nests begged more often than those in female-fed nests. Because male interest in feeding nestlings was often evident quite early in the nest cycle, we noted the day that males were

17
first observed to feed nestlings, and then calculated the highest begging rates that we had observed on days prior to the male's first feeding. It is
likely that males heard these begging calls, because they frequently
perched on or near their nestboxes, and because begging offspring over
11 days old were audible to human observers at distances greater than the
maximum diagonal distances in the aviaries.
Data from the size-matched female-fed and male-assisted families
show that nonfeeding males (female-fed families) ignored offspring beg-
ging at extremely high rates, while feeding males (male-aided families)
jumped in the boxes to feed as soon as total begging rates emanating from
nests exceeded 4 begs/hr (female-fed Us male-aided families, p = 0.014,
Mann-Whitney U test, Fig. 3). Indeed, one male began to feed on the
same day as his nestlings were first heard to beg (family E, Fig. 3). Since
Fig. 3. Highest begging rates (no. bouts/hr) recorded on any day prior to the first day that a male entered the box to feed nestlings.
begging rates and intensities increased with nestling age in all families
(see section I), we also noted the age of the oldest nestling in each brood
on the first day a male was observed to feed. Again, feeding males started
to feed much earlier in the nest cycle than did males in female-fed
families (female-fed as male-aided families, p = .014, Mann-Whitney U
test). Hence, there was no indication that variance in nestling begging rates was responsible for the inter-male variance in feeding tendency.
Although offspring begging rate could not account for differences be-
tween males, it was an important determinate of intramale variation in
feeding behaviour. For those males which fed their young, nestling beg-

18
ging appeared to stimulate male entry into the nest, and, once in the
nest, males tended to feed the most vigorous beggers most frequently. In contrast to females, which spent long periods inside the nest
brooding, preening the nestlings, and cleaning the nestbox, males tended
to enter the nestboxes for brief periods, and most (56%, N = 260 entries) of male entrances resulted in the male's feeding nestlings and/or their
mate. Observations suggested that males tended to enter nestboxes when
they heard begging calls within; this was tested using the two nests with
the most data on male visits. For each nest we determined the number of
male visits that were preceded (< 0.5 min) by offspring begging vocaliza-
tions, and the number of visits when the offspring were silent. Expected
frequencies for each situation were computed by totaling all of the
minutes during which any nestlings begged, divided by the total minutes
each nest was observed. For example, nest H was observed a total of 992
minutes, during which one or more nestlings were begging for 130
minutes, or 13 % of the time. In this nest, however, nestling begging
preceeded male entry into the box on 19 occasions, while the male
entered the box with silent offspring only 6 times (p < 0. 00 1, X2 = 36.8). A
second feeder male (nest F) showed a comparable tendency to jump into
the nestbox when offspring were begging (p< 0.001, X2 = 15.3). For
neither of these two nests was there a significant tendency for females to
enter the nestbox following offspring begging. Once males had entered the nest boxes, they continued to respond to
nestling begging behaviour. In contrast to females, which fed or nibbled
smaller offspring first, males' first approaches were independent of off-
spring size. Instead, a male was most apt to feed first that offspring which
had reached the male and was begging from him. Further feedings in the
bout also tended to depend on offspring begging behaviour. As nestlings
managed to reach the male and beg from him, they were rewarded by a
feeding; because nestlings were constantly jockeying for position in front
of the male, bout lengths per nestling were short, and rarely did one
nestling get more than one or two regurgitations in sequence (see bout
lengths). The males' attention to begging is reflected by their distract-
ibility. When feeding one nestling "x" and interrupted by the begging of
a second nestling "y", six out of six males were more apt to feed y than to
return to x (p = 0.016, Binomial test). Recalling that females tended to
feed x in this situation, we compared the distractibilities of mated males
and females feeding the same nestlings: fathers were much more likely to
feed an interrupting nestling than were mothers (p < 0.001, Fisher com-
bined probabilities test).

19
If males attend to begging and not to offspring size, then on any given
day we would expect that those offspring begging more often would be
fed more often by their fathers. Since males fed infrequently, there were
only 22 observation periods in which more than one nestling begged and
were fed by the male. However, 15 of these observation periods had
positive correlations between offspring beg rate and male feed rate.
Moreover, six out of six males had more periods with positive relation-
ships between begging and feeding than periods with negative relation-
ships between these variables (p = 0.016, Binomial test). The data presented so far suggests that males simply distribute food in
response to beg rate. However, when presented with a choice between
begging nestlings and a begging mate, most males chose to feed the nestl-
ings. Out of 66 occasions when a female and one or more nestlings
simultaneously begged from a male, the male fed the nestling(s) in 46
cases. In these choice situations, eight out of nine males chose nestlings more often than they chose to feed their mates (p = 0.02, Binomial test). Since females begged as vigorously and as loudly as nestlings, it appears that males can discriminate between recipients while feeding, even
though they do not differentially feed the nestlings. As was noted in the previous section, the maternal feeding style in-
volved a number of time-consuming activities besides regurgitation. In
contrast, males appeared to feed the first available begging offspring. As
one would expect from the descriptive accounts, males were able to feed
nestlings at much higher rates within each feeding episode than were
females. On average, female budgerigars feeding alone delivered
0. 16 + 0.04 regurgitations/sec within feeding episodes (8 females, 308
bouts) while males feeding alone delivered food nearly twice as fast
(0. 28 + 0.15 regurgitations/sec, 5 males, 45 bouts). Males seemed more
likely than females to increase delivery rates as the nestlings grew older, but for neither sex was the trend significant. Five females had negative correlations between brood age (days since first hatch) and food delivery rates per episode, while three females had positive correlations between
these variables (average correlation for all eight females, r = 0.16 + 0.26,
average slope = 0.0017 ± 0.0033). Three out of five males had positive correlation between brood age and delivery rate (r = 0.30 + 0.41, average
slope = 0.011 ± 0.02). In nests where males and females feeding the same
offspring were compared, females fed more slowly in 5 out of 5 families
(p = 0.031, Sign test), and within three of these nests, the differences were
significant (Kolmogornov-Smirnov test, p<0.05, p<0.025 and
p < 0.005).

20
In the presence of a spouse simultaneously feeding the nestlings, the
feeding rates of males, but not females, decreased. Four out of five males'
rates decreased (X for all five males = 0.20 ± 0.06 regurgitations/sec), two
of them significantly (Kolmogornov-Smirnov test, p < 0.01, p < 0.025), while females' rates were comparable to the rates of females feeding alone
(X for all females = 0. 15 + 0.07 regurgitations/sec). The sex difference
underscores the temporal costs of discriminating between potential food
recipients: the presence of the male had little effect on females, which
already were using time consuming behaviours to selectively feed the
nestlings. However, the presence of the female forced males to
discriminate between spouse and offspring, and their feeding rates
declined when presented with even this fairly simple choice.
Nestling feeding behaaiour.
Budgerigar nestlings occasionally fed their siblings, using the same
behaviour patterns as adults. Feeding rates by siblings were low (Table 2); only 24 sib-sib regurgitations, involving 10 nestlings in four different
families, were observed in this study. All 24 observed sib-sib feedings followed either mutual or unidirec-
tional begging between the fed nestling and the feeding nestling. In-
terestingly, in 22/24 cases a nestling fed its younger sibling (p < 0.001,
Sign test), even though nestlings begged from both their older and
younger siblings. Since the number of available older and younger sib-
lings varies with birth order, we looked at the direction of begging for 15 5
middle nestlings (e.g., nestling # 2 in a brood of 3, nestlings # 2 and # 3
in broods of 4, etc. ). These nestlings had an equal number of older and
younger siblings, and directed 92 begs to younger siblings, and 125 begs to older siblings. Nine middle nestlings begged more often from older
siblings than younger ones; the reverse was true for the six others.
Hence, preferential begging was not entirely responsible for the
prevalence of food transfer from older to younger nestlings.
Parental feeding rates and nestling growth.
If parental feeding rates reflected the amount of food delivered to nest-
lings, then we would expect a positive relationship between parent
regurgitation rate in a given period and nestling growth rate in that same
period. In Period 1 & 2 there was no significant correlation between
parental regurgitation rate and nestling growth (r = -0.24, 32 df), but
significant positive correlations between these variables were obtained in

21
Periods 3 and 4. Period 3 nestlings exhibited a strongly positive, cur-
vilinear, saturating relationship between feeding rate and growth; the
best fit (highest r value) was obtained by
G = 1.3PO 1
where G = growth (g/d), and P = parental regurgitations per hour
(r = 0.50, 36 df, p < 0.01 ). This equation indicates that the effect of food
on growth was most dramatic (highest slope) for low feeding rates, and
that the slope gradually declined until above 20 feeds per hour there was
no perceptible effect of food on growth. In Period 4 there was also a
positive curvilinear relationship between parental regurgitation rates and
nestling growth; best fit was obtained by
G = 0
(r = 0.35, 36 df, p < 0.05). The lower r vaue for Period 4 reflects con-
siderable scatter in growth rates, especially at high feeding rates (see Fig.
7). Some of this variability depended on the sex of the feeding parent; for
furher analysis and discussion see Section II. These results suggest that
feeding rate can be used to estimate amount of food delivered to nestlings in Period 3 and 4.
II. Parent-offspring conflict in budgerigars.
Budgerigar parental care as parental investment.
Since parental investment by definition involves a cost in terms of subse-
quent parental reproductive success, we needed to test whether parental care in our captive situation fit this definition. Considering the
"optimal" resource conditions in the field cages (e.g., food ad lib., minimal travel distances from nests to food supply, water ad lib. lack of
competition for nestboxes, etc. ) it was possible that parent budgerigars
might be able to raise abnormally large numbers of nestlings with no
subsequent cost to future reproduction. Since in the field budgerigar nesting seasons are temporally restricted
(WYNDHAM, 1978), we reasoned that time intervals between subsequent clutches might be useful as an indication of the costs of rearing one brood
on the ability to rear a second brood through to fledging. Accordingly, we
looked for positive relationships between the number and biomass of
nestlings in broods raised by two parents and the time until the onset of
the laying of the first egg of the next clutch. Intervals between subsequent clutches were measured several ways: i) from the first egg of the first
clutch to the first egg of the next clutch (first egg), ii) from last egg of the

22
Fig. 4. Number of days between the first egg of a clutch and the first egg of the subsequent clutch, as a function of number of nestlings raised from the first clutch.
first clutch to the first egg of the next clutch (last egg), iii) from first fledge of the first clutch to the first egg of the next clutch (first fledge), iv) from
last fledge of the first clutch to first egg of the next clutch (last fledge). The first three of these variables were positively correlated to both the
number and biomass of nestlings in the first clutch, while "last fledge" was uncorrelated with either of these variables (Number of nestlings ver-
sus (a) first egg, r = 0.64, p< 0.001, N = 40 first clutches, Fig. 4, (b) last
egg, r = 0.53, p < 0.01, (c) first fledge, r = 0.68, p < 0.0005; Biomass of
nestlings versus (a) first egg, r = 0.57, p < 0.001, (b) last egg, r = 0.46,
p < 0.03; (c) first fledge, r = 0.74, p < 0.01 ). Interestingly, there were no
significant partial correlations between the number of eggs laid for the
first clutch and any time intervals to the next clutch when the linear ef-
fects of brood size (# nestlings) were removed. This implies that in our
captive flocks, egg production was much less costly to parents than was

23
raising young; under these conditions number of eggs laid does not fit the
definition of parental investment.
A few female budgerigars in our study managed to rear offspring without any male assistance. One such female produced no second clutch
after her first effort (interclutch interval > 1 year), while two other
females did eventually lay second clutches. In both cases, these females
took much longer to start on second clutches than did females with male
assistance which had raised the same number of offspring (Fig. 4). Hence, not only did parental care in budgerigars involve a cost to the
parents, but a male's allo-feeding of his mate and young shortened inter-
brood intervals by almost a month. It is supposed that if there were costs
to parents under captive conditions, greater costs would be experienced
by budgerigar parents in the wild.
The control of the allocation of food in budgerigar families.
Since the rearing of budgerigar broods seems to involve parental invest-
ment, and since feeding is the primary type of parental care provided by the parents, we next investigated the control of food allocation within
families. The previous section suggests that the control of food allocation
depends on the sex of the parent. Since females tend to ignore begging and allocate food by offspring size (or some variable related to size), in
female-offspring interactions the parent appears to control food alloca-
tion. In contrast, since males respond to offspring begging, in male-
offspring interactions the offspring may exercise some control over food
allocation. Given the variance in the extent of direct male provisioning of
young, one would expect that begging offspring would be more apt to be
rewarded during those periods when a large proportion of the feedings were delivered by the males.
For each nestling in Period 3 and 4, we calculated the proportion of all
observed regurgitations which were delivered by the male parent. Male
contributions were highest for Period 4 nestlings in male-aided families
(56.6 ± 27 % of regurgitations were delivered by males, N = 14 nestlings). The males of female-fed families provided a significantly lower propor- tion of the regurgitations to their Period 4 nestlings (26.9 + 41%, T = 2.33, 28 df, p < 0.025, one-tailed). Period 3 nestlings received a ma-
jority of their feedings from the female in both types of families, but
aiding males delivered a higher proportion of regurgitations than the
males of female-fed families (male-aided = 18.6 + 20.5 % , female-fed =
0.8 + 3.3 % , T = 3.42, 32 df, p < 0.005, one-tailed).

24
If male parents were more responsive to nestling begging, then the
strength of correlations between total beg rate and total feed rate should be related to the proportion of feedings contributed by the males. That is, total beg rate (= beg rate to both parents and to siblings) and total feed rate (= feed rate by parents and siblings) should be most strongly positively correlated for Period 4 offspring in male-aided families, and
progressively less-strongly related for Period 4, female-fed families, Period 3 male-aided families and Period 3 female-fed families.
As expected, the strongest positive correlation between total beg rate and total feed rate was obtained for Period 4 nestlings in male-aided families (r = 0.71, p < 0.01, 14 df). Correlations between these variables declined in the expected order (Period 4 female-fed nestlings: r = 0.15, n.s.; Period 3 male-aided nestlings: r=-0.04, n.s. ; Period 3 female-fed
nestlings: r=-0.26, n.s.). Hence offspring obtaining most of their
feedings from the male do receive more feedings if they beg more fre-
quently, while there is no benefit (and possibly is a cost) to extra begging for offspring primarily provisioned by the female.
Comparisons of female fed and male-aided families.
Since the allocation of parental care appeared to be primarily controlled
by the parents in female-fed families, and partially controlled by the off-
spring in male-fed families, a comparison of begging rates, feeding rates and fledging success in the two types of families allowed us to test several of the predictions of parent-offspring conflict theory. In particular we could determine if offspring control over parental investment lead to
potentially costly or inefficient escalated begging rates for the same
degree of physiological need, or if offspring demanded more food than
they required for maximal fledging success (see Introduction). As one might expect from the greater rewards of begging in male-aided
nests, offspring in those nests begged at higher rates in Period 3 and Period 4 than did offspring in female-fed nests (Fig. 5). A nested ANOVA analysis was used to control for differences among nests while
comparing male-aided with female-fed families. In Period 3, there were
significant differences in beg rate among nests (F6,24 = 3.24, p < 0.025), and between male-aided and female-fed families (F1 1,6 = 8.46, p < 0.05). In Period 4, there were no significant nest effects on begging rate, but male-aided nests again had higher begging rates than did female-fed families (F 1,6 = 13.3, p< 0.025). These elevated offspring begging rates did not appear to reflect greater offspring need in male-aided nests,

25
Fig. 5. Nestling beg rates (no. bouts/hr) during Period 3 and 4 in female-fed and male- aided families.
because there were no significant differences in Period 3 or 4 growth rates
in male-aided and female-fed nests (Table 3).
Offspring in male-aided nests not only begged more, but they were
more apt to beg at high rates to birds that would not reward their efforts.
For example, since neither mothers nor siblings were apt to respond to
begging by Period 3 offspring, most begging addressed to these birds was
wasted effort. Offspring in male-aided nests begged at higher rates from
their mothers and siblings in Period 3 than did the offspring in female-fed
nests (Table 3, nest effects = F6,2q = 3.8 p < 0.01; parental care effects:
F 1,6 = 7.2, p < 0. 05). Overall parental feeding rates were slightly, but not significantly,
higher for Period 3 offspring in male-aided nests (12.2 ± 9.6 feeds/hr) than in female-fed nests (9.1+6.6 feeds/hr), but there was a highly
significant difference in Period 4 feeding rates in the two types of families:
Fig. 6, nested ANOVA: nest effect = F1 q = 29.0, p<0.01). This in-
creased feeding was almost entirely due to extra male provisioning in
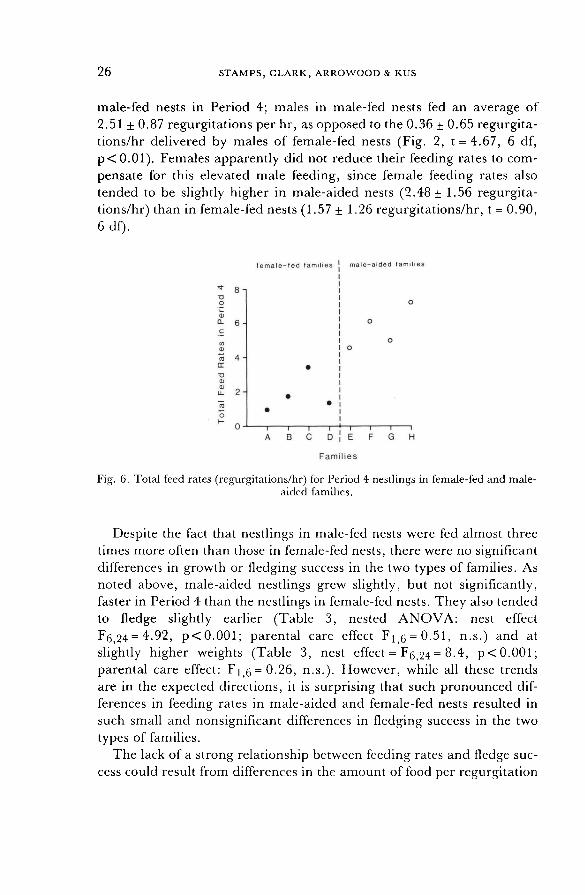
26
male-fed nests in Period 4; males in male-fed nests fed an average of
2.51 ± 0.87 regurgitations per hr, as opposed to the 0.36 ± 0.65 regurgita- tions/hr delivered by males of female-fed nests (Fig. 2, t = 4.67, 6 df,
p < 0.01 ). Females apparently did not reduce their feeding rates to com-
pensate for this elevated male feeding, since female feeding rates also
tended to be slightly higher in male-aided nests (2.48 + 1.56 regurgita-
tions/hr) than in female-fed nests (1.57 ± 1.26 regurgitations/hr, t = 0.90,
6 df).
Fig. 6. Total feed rates (regurgitations/hr) for Period 4 nestlings in female-fed and male- aided families.
Despite the fact that nestlings in male-fed nests were fed almost three
times more often than those in female-fed nests, there were no significant differences in growth or fledging success in the two types of families. As
noted above, male-aided nestlings grew slightly, but not significantly, faster in Period 4 than the nestlings in female-fed nests. They also tended
to fledge slightly earlier (Table 3, nested ANOVA: nest effect
F6,24=4.92, p<0.001; parental care effect F1,6=0.51, n.s.) and at
slightly higher weights (Table 3, nest effect = F6,24 = 8.4, p < 0.001;
parental care effect: F1,6=0.26, n.s.). However, while all these trends are in the expected directions, it is surprising that such pronounced dif-
ferences in feeding rates in male-aided and female-fed nests resulted in
such small and nonsignificant differences in fledging success in the two
types of families. The lack of a strong relationship between feeding rates and fledge suc-
cess could result from differences in the amount of food per regurgitation

27
delivered by parents in the two types of families. This possibility was
tested by comparing the regressions of growth versus feed rate for male-
aided and female-fed families. If the parents in male-aided families pro- vided less food per regurgitation than those in female-fed families, then
the intercept of feeding on growth would be lower in male-aided families.
Similarly, if male-aided families had a decelerating curvilinear relation-
ship between feeding rate and growth (e.g., less food/regurgitation at high
feeding rates), while female-fed families had a linear relationship, then
male-aided families should have an overall lower slope of growth on feed
rate than female-fed families.
In fact, there were no significant differences in either slope or intercept for regressions of Period 4 growth on total Period 4 feed rate for male-
aided and female-fed families (Fig. 7; male-aided families: growth
(g/d) = 0.022 (regurgitations/hr) -0.16, r = 0.31 ; female-fed families:
Fig. 7. Growth rate (g/day) in Period 4 as a function of type of parental care. Dark sym- bols : female-fed families, open symbols: male-aided families. Triangles: male-majority
fed (MMF) nestlings, Circles: female-majority fed (FMF) nestlings.

28
growth (g/d) = 0.034 (regurgitations/hr) -0.18, r = 0.30). Instead, Fig. 7
suggests that variance in growth rate as a function of feeding rate tends to
increase in both family types at feeding rates above 4/hr, and that more
male-aided than female-fed families end up in this zone of variability as a
result of their elevated feeding rates.
Another possible reason for the apparent lack of efficiency of male-fed
nests is that male parents might deliver smaller amounts of food per
regurgitation than do female parents. Since this trend could be obscured
by categorizing whole nests as male or female-fed, we determined for
each nestling whether it received the majority of its Period 4 feedings from the female (= female majority-fed, FMF) or from the male (= male-
majority-fed, MMF). While most FMF nestlings occurred in female-fed
families (and vice-versa for MMF nestlings) there were enough cross-
overs to make the analysis worthwhile (Fig. 7). A comparison of regressions between feeding rate and growth for
MMF and FMF nestlings suggested that males fed more per regurgita- tion than females, especially at higher feeding rates (Fig. 7, MMF:
growth = 0.054 regurgitations/hr -0.271; FMF: growth = 0.008 regurgi- tations/hr -0.124). There was no significant difference between the in-
tercepts for the two types of nestlings, but the slope for MMF nestlings was significantly higher than that for FMF nestlings (FI,28= 7.27, p < 0.025). In addition, while MMF nestlings had a strong significant
relationship between growth and feeding rate (r = 0. 71, p < 0.01 ), FMF
nestlings had a much more variable relationship between these variables
(r = 0.13, 14 df, Fig. 7). Hence, if anything, males fed more (not less) per
regurgitation than females, and the amount of food per regurgitation varied less among males than among females. In summary, there is no
indication that nestlings in male-aided families or male-fed nestlings received less food per regurgitation than nestlings in female-fed families
or female-fed nestlings. Hence, the different regurgitation rates in the
two types of families probably reflect true differences in the amount of
food delivered to the nestlings. The final question is whether the extra food delivered to the offspring
in male-aided nests constituted a cost in terms of future parental
reproductive success. As a first approximation, we used "first egg"
(defined above) to compare inter-clutch intervals for male-aided families
and female-fed families. While the differences were not significant, male-
aided families tended to start next clutches sooner than female-fed
families (male-aided = 58 + 2.6 days, female-fed 65 ± 7.1 days, t = 1.85, 6
df). Other results suggest that males eager to feed their nestlings also fed

29
their mates more often (CLARK & STAMPS, unpublished data), so these dif-
ferences may reflect variance in degree of male participation in allo-
feeding rather than the costs (or lack of them) of higher food delivery rates to the nestlings. In any case, there was no evidence under the condi-
tions of this study that the increased food delivery to male-aided Period 4
nestlings depressed future parental reproductive success in male-aided
families.
Discussion
The present study provides strong indirect evidence for the existence of
genetic parent-offspring conflict in budgerigars. First, offspring in this
species appear to be capable of "selfish" behaviour. When nestlings could influence parental feeding rates via their begging behaviour, then
they asked for and received nearly three times more regurgitations per unit time than offspring in comparable families in which parents deter-
mined food allocation. However, there was no significant difference in
growth rates, fledge ages or fledge sizes for these demanding offspring and their quieter counterparts in other nests. We assume that under
natural conditions, a threefold increase in the amount of food delivered
per nestling would involve temporal, energetic and/or survival costs for
the parents. As is also predicted by conflict theory, offspring escalate their begging
rates, for the same degree of need, when begging is positively reinforced
by parental feeding. Further analysis has shown that escalation of beg rates can occur within a day after males begin to feed (STAMPS & CLARK, in prep.), hence budgerigar nestlings are impressively sensitive to
changes in the potential rewards of solicitation. This behavioural profile of budgerigar nestlings fits the predictions of conflict theory, but is in-
congruent with the hypothesis that offspring honestly signal their
physiological needs via their begging rates.
On the other side of the equation, female budgerigars have evolved a
complex behavioural feeding strategy that effectively counters selfish off-
spring demands. Females allocate food primarily on the basis of offspring
age and size, and devalue beg rates according to offspring age and size.
This strategy effectively overrides any potential advantage to larger off-
spring in the extremely asynchronous budgerigar broods, and explains
why all nestlings within budgerigar broods have comparable growth rates
and fledging success. The sophistication of the female strategy, and its

30
proven efficacy in curtailing escalated beg rates and efficiently converting food into nestling biomass, argue that the female feeding mode has
evolved to counter selfish offspring behaviour. In addition, we have
shown that the female feeding strategy is temporally more costly than the
usual laissez-faire style of the male, since females can feed offspring at
only one half the rate, per episode, as can males. It is also clear that the
process of choosing among potential food recipients, rather than any in-
herent sexual difference in feeding style, is responsible for the slower
rates among females. When males had to choose between feeding females
and young, their feeding dropped to rates comparable to females'. A
100% increase in the time required to feed young could be extremely im-
portant in natural populations, if foraging were time-limited. The ex-
istence of a neurologically sophisticated and ecologically costly parental
counterstrategy is predicted by conflict theory, but not by the hypothesis that parents use the most efficient method to deliver food to young.
The theoretical models of parent-offspring conflict have outlined
several stable evolutionary outcomes of coevolution between selfish off-
spring and their parents (STAMPS et al., 1978; PARKER & MACNAIR, 1979). Male-aided budgerigar families seem to conform best to the "pro rata"
ESS discussed by PARKER & MACNAIR (1979), in which offspring ask for
and receive more than they require, but parents stop short of filling all
offspring demands. In this compromise scenario, neither participant "wins", since parents provide more than the optimal allocation, but off-
spring obtain less resources than they are requesting. In budgerigars, while older nestlings were able to obtain extra resources in male-aided
families, they almost never obtained sufficient resources to cut off their
begging behaviour. It is of course possible that Period 4 budgerigar nest-
lings would continue to beg regardless of the amount eaten, but this
seems unlikely, in view of the lack of begging and refusals of food in well-
fed younger nestlings, and the low beg rates of some singleton nestlings in
Period 4 (STAMPS & CLARK, in prep.). Hence male-fed budgerigar families generally seem to operate in a "compromise" zone of invest-
ment, in which nestlings receive more food than they require but less
than they request. Indirect evidence for a "pro rata" ESS in another avian species has
been provided by BENGTSSON & RYDEN (1983). Parent great tits (Parus
major) are sensitive to offspring begging, and food delivery rates can be
artificially increased by playback of taped nestling begging calls
(KHAYUTIN & DMITRIEVA, 1979; BENGTSSON & RYDEN, 1983) or by briefly starving a brood (BENGTSSON & RYDEN, 1983). However, when taped beg-

31
ging was used to increase parental feeding rates, and more food was
eaten by the nestlings, there was no significant increase in growth rate
within the brood (BENGTSSON & RYDEN, 1983). Again, nestlings seem
willing to eat more food than was required for efficient growth. On the
other hand, while parent tits did respond to begging, they rarely (if ever) fed offspring enough to stop their begging. In this case we know that
parents stop short of satiating the young because artificially provisioned broods ate enough to become silent (BENGTSSON & RYDEN, 1983). In-
terestingly, parental tits responded with alarm to artificially provisioned, silent broods, suggesting that this situation was a novel one for Parus
major parents! In addition to following a "pro rata" program, as described by conflict
theories, Great Tit parents use a proximal feeding strategy which could
help to limit the expression of selfish offspring behaviour. The theoretical
models assume that the extra resources obtained as a result of selfish
behaviour would all go to the selfish offspring (review in STAMPS, 1980). However, while parent tits did go on more foraging trips, and delivered
more food, as a result of hearing elevated begging rates, they did not
necessarily deliver more food to those offspring which begged most
vigorously. Instead, relatively quiet siblings obtained some of the extra
food, especially if they were larger than the vociferous nestlings (RYDEN &
BENGTSSON, 1980; BENGTSSON & RYDEN, 1981, 1983). If begging carried a
cost (e.g., in terms of energy), then any dilution of the benefits of extra
begging would tend to limit the evolution of escalated begging among the
offspring (see STAMPS et al., 1978). Puffins may be another species in which a pro rata solution to conflict
is tempered by a relatively high cost of begging. Puffin chicks have the
capacity to be selfish, and in captivity they will demand and eat twice as
much food as is required for maximal growth (HUDSON, 1979). Parents
are sensitive to cues emanating from their chicks, and will respond to
taped begging calls by increasing the rate of food provisioning, and to
substitutions of old chicks by younger ones by extending the duration of
care and fledging the changelings at normal ages and weights (HARRIS,
1983). However, puffin chicks are normally rather quiet at all ages,
perhaps because predatory gulls are attracted to the nest burrows and to
the unattended young by elevated begging calls (HARRIS, 1983). Female-fed budgerigar families seem to conform to a second
theoretical outcome for conflict, in which parents use some "honest cue"
for investment allocation (e.g., offspring size) and ignore offspring solicitation. This is a "parent wins" solution in that offspring do not ob-

32
tain extra parental investment, but parents could incur future reproduc- tive losses if the strategy involved substantial costs to the parents (STAMPS et al., 1978). In addition to the possible ecological cost of a doubling in food delivery time, the female budgerigar strategy might lead to uneven
growth rates, as females ignored larger young that were truly hungry in favor of smaller offspring that were nearly replete. The dangers of ignor- ing offspring signals, discussed by DAWKINS (1976) and PARKER & MACNAIR (1979) may only be affordable in a species like budgerigars, in which offspring have flexible developmental schedules and a long hatch-
fledge period. In budgerigars, nestlings can have retarded growth for four to five days, but catch up later and fledge at normal ages and weights
(CLARK, unpublished data). The current study illustrates the extreme difficulty of testing conflict by
measuring the costs and benefits of parental investment. The resolution of this study was fine enough to show a linear relationship between the number of young raised and the onset of the next clutch. Thus, raising budgerigar nestlings fits the definition of parental investment. However, the costs and benefits of raising selfish and nonselfish offspring could not be accurately determined. In fact, under the conditions of this study, the extra resources obtained by the "selfish" offspring in the male-aided families fulfilled neither of the criteria for parental investment: there was no indication that the extra food increased the fitness of the offspring that obtained it, nor was there any indication that it decreased the future
reproductive success of the parents providing it. Both negative results
may be related to the food ad lib conditions in captivity. It is likely that these results would have been different under the natural conditions of
long distances from nest to feeding areas, predation on adults and young, and temporally restricted breeding seasons (WYNDHAM, 1978, 1981, 1982; pers. comm.). However, it would also be difficult to measure
significant costs and benefits in a natural population of budgerigars, given the annual and intra-population variation in resource levels, mor-
tality, and reproduction (WYNDHAM, 1978, 1980a and b, 1981, 1982;
pers. comm.). Hence, the presence or absence of behavioural mechanisms predicted by conflict is currently a more useful approach than attempting to measure the reproductive payoffs predicted by con- flict.
Although the present study was conducted on domestic budgerigars in a captive situation, many of the results probably can be applied to the natural situation. For one thing, it is unlikely that the complex female
feeding strategy or the nestling's facilitative begging behaviour evolved

33
de novo in domestic birds. Female preference for smallest nestlings has been observed in other domestic and feral budgerigars (KURT BANZ, ANNE WENNER, pers. comm.), and field observations, while limited,
agree with the patterns observed in this study. For example, average clutch sizes, fledge ages, degree of hatching asynchrony, and hatch-order
independence of growth rate and fledging success comparable to those in our flight cages have been observed in natural populations (WYNDHAM, 1978; pers. comm.; WENNER, pers. comm.). In addition, variance in the
frequency of male entry into nestboxes, which in this study was highly correlated to male feeding of nestlings, has been seen in wild budgerigars (KURT BANZ, pers. comm.), and in other Australian psittacines [e. g. , the white-tailed black cockatoo, (SAUNDERS, 1982)]. In the field, as in our
study, female budgerigars incubate the young, and are allo-fed by their
mates, males are more apt to enter nestboxes at the end of the nest cycle, and neither parent is apt to feed young after they fledge (WYNDHAM, 1978, 1980b). The ad lib resource situation and high densities in the
flight cages also mirror conditions for some wild populations, in which
large flocks arrive in areas with temporarily abundant food supplies, and breed colonially with small inter-nest distances (WYNDHAM, 1978, 1980a,
1981). The size-dependent feeding strategy used by female budgerigars has
not been documented for any other avian species, but there are hints that it may exist elsewhere. Hatch asynchrony in most altricial birds is much less extreme than that of budgerigars (CLARK & WILSON, 1981), leading to smaller ranges of nestling size within broods. Size-dependent feeding would be much more difficult to observe in such species, and this prob- lem would be accentuated if the correlations between feeding rate and
offspring size were less extreme than the logio relationship found in
budgerigars. Hence, it is impressive that at least one of the handful of
passerine parent-offspring studies has found a strong inverse correlation between parental feeding rate and nestling weight [the fieldfare, Turdus
pilaris, (RYDEN & BENGTSSON, 1980)]. There are also hints that parrots other than budgerigars may exhibit
size-dependent feeding rates. Reproductive traits such as 2 day laying in- tervals and initiation of incubation with the first egg are conservative in
psittacines (SMITH, 1975) and often lead to pronounced hatch asynchrony (SMITH, 1972). However, weight and fledge data indicate that youngest nestlings are not disadvantaged relative to their older siblings (SMITH, 1975; CACCAMISE, 1976; SAUNDERS, 1982; WYNDHAM, unpublished data; STAMPS, pers. obs.), suggesting that one or both parents may be select-
ively feeding the much smaller, late-hatched nestlings.

34
Size-dependent feeding is only one of several proximal mechanisms
allowing parent birds to control food allocation, and prevent larger nest-
lings in asynchronous clutches from monopolizing all of the food
resources. In great tits (Parus major) and blackbirds (Turdus merula), RYDEN & BENGTSSON have found that food is normally evenly distributed
among the members of the brood, despite the fact that larger nestlings have priority access to the best feeding positions and are most apt to first
contact the parent's beak. In part, this unexpectedly egalitarian feeding is due to differential nestling behaviour; light nestlings beg more fre-
quently and vigorously than their siblings, and parental attention to this
begging compensates for their size disadvantage (RYDEN & BENGTSSON,
1980; BENGTSSON & RYDÉN, 1981; 1983). In addition, parents may refuse
to feed an individual nestling more than once in succession; this alterna-
tion ensures that all of the food will not go to the largest, most competitive
young [blackbirds, (RYDGN & BENGTSSON, 1980); field sparrows, (BEST,
1977)]. Another effective parental strategy is to use many alternate
landing sites and feeding positions around the nest cup. As R YDÉN &
BENGTSSON (1980) have shown for fieldfares, this technique prevents
larger nestlings from anticipating and monopolizing the best feeding sites, and tends to equalize food delivery rates among the nestlings. Of
course, this strategy is only available to open-nesters, since large nest-
lings can more easily monopolize the single entrance and access to food in hole or covered-nesting species [e.g., great tits: (BENGTSSON & RYDEN,
1983)]. This may help explain why the runts of hole-nesting passerines seem more apt to die than those of open-nesting species 1968); see also SLAGSVOLD, 1982 for an alternate explanation)].
The forgoing discussion suggests that many parent birds may set up
nestling size differences via changes in laying and incubation schedules, and then use feeding strategies that compensate for the competitive
disadvantages of the smaller, younger nestlings. These observations seem
inconsistent with the classical explanation for hatch asynchrony: i. e. , it
enables parents to starve the last hatched (smallest) young selectively if
sufficient food is unavailable to raise the entire brood (LACK, 1956;
RICKLEFS, 1965). There are at least two possible explanations for the ap-
parent discrepancy. First, asynchrony may not function in brood reduc-
tion, and second, complex parental behaviours may allow better control
of whether and when brood reduction occurs.
A number of alternate hypotheses for hatching asynchrony have been
discussed in the ecological literature (see reviews in CLARK & WiLSOrr,
1981; SLAGSVOLD, 1982; RICHTER, 1982). Data supporting many of these

35
hypotheses are indirect or inferential, but in some cases brood reduction
can be ruled out as an adaptive function of hatch asynchrony. For exam-
ple, in house martins, hatch asynchrony is primarily a result of food
shortages that delay egg laying, and late hatched nestlings often die early even when food is abundant during the nestling period (BRYANT, 1978a
and b). In fieldfares, experimental manipulation of brood asynchrony resulted in higher nesting survival and fledge success for synchronous than for asynchronous broods, largely because the last-hatched asyn- chronous young died early in the nestling period (SLAGSVOLD, 1982). Both synchronous and asynchronous broods were larger than the normal
brood sizes, so brood reduction (as a result of food shortage) would have
predicted higher fledging success in the asynchronous broods
(SLAGSVOLD, 1982). To the extent that hypotheses other than brood reduction account for hatching asynchrony, parents will be disadvan-
taged by losing late-hatched nestlings. Hence, we might expect to
sometimes see parental feeding strategies which counter the size advan-
tages of larger nestlings, and allow small nestlings to grow at normal, or
even at higher rates than their siblings (e.g. NEUB, 1979; budgerigars, see
above). Even if brood reduction were the primary function of hatching asyn-
chrony, parent-offspring conflict theory predicts that parents and off-
spring would not always agree on whether or when loss of the smallest
nestlings is appropriate (O'CONNOR, 1978). As was pointed out by early workers, the rule "feed the head which is lifted the highest" would cer-
tainly result in brood reduction, since large nestlings would have to be
satiated before their smaller siblings had a chance to eat (LOCKIE, 1955;
LACK, 1956; RICKLEFS, 1965; and LoHRr., 1968). However, if the older
siblings were selfish, sensu TRIVERS (1974), then they could take more
than their share before relinquishing access to food to their smaller sib-
lings. This would result in an ever-widening size discrepancy between the
nestlings, and early starvation of the younger nestlings under any but
superabundant food conditions. In contrast, the more sophisticated
parental feeding behaviours described above would allow avian parents
greater control over the proximal mechanisms of brood reduction; young
nestlings could be maintained in the brood despite competition from their
older siblings, while the size discrepancies maintained in the brood would
allow efficient starvation of the younger nestlings should the food situa-
tion deteriorate. In great tits, for example, food was evenly distributed
among the nestlings under normal circumstances, but after a short period of brood starvation, the largest nestlings obtained virtually all of the food

36
(RYDIN & BENGTSSON, 1980; BENGTSSON & RYDEN, 1983). In budgerigars, females usually feed all of their nestlings at the samp size-dependent rates, but this does not preclude a female's shifting her preference to the
penultimate nestling, and selectively starving her youngest nestling. Hence, complex feeding strategies favoring or maintaining small (young) nestlings are not only consistent with brood reduction, but may be re-
quired if parents are to retain control over the conditions for brood reduc- tion in asynchronous broods.
Summary
Despite widespread theoretical interest in genetic conflict between parents and offspring, there is little empirical evidence that it exists in nature. Theoretical models suggest two outcomes of conflict not predicted by alternate theories: (1) offspring that control the allocation of parental investment might show escalated demand behaviours (e.g., begging) and demand more resources than they could efficiently use and (2) parents might evolve behavioural counterstrategies which prevented offspring from obtaining extra resources, but which were more costly than a "laissez-faire" parental strategy allowing offspring control. These predictions were tested in budgerigars, (Melopsittacus undulatus) in large flight cages at Davis, California.
Budgerigar clutches hatch extremely asynchronously, yet all nestlings grew at similar rates and fledged at similar sizes and ages. This independence of hatch order and perfor- mance seemed due primarily to the mother budgerigar's allofeeding strategy: females allofed offspring mainly on the basis of size, and only secondarily attended to begging rate. Offspring of a given age and size were treated the same by their mothers regardless of hatch order, and offspring undersized for their age were fed as if they were younger. In contrast, male budgerigars attended to offspring begging rates. Males tended to initiate feeding bouts when offspring begged, and to allofeed vigorous beggers more often.
Variance in male allofeeding behaviour allowed comparisons of size-matched families in which females performed nearly all of the allofeeds to nestlings (= female-fed families) with families in which males and females both allofed nestlings (= male-aided families). The parent controlled the allocation of food in female-fed families, while offspring had greater control over food allocation in male-fed families. As was predicted by conflict theory, the female counterstrategy was effective but potentially costly: the food delivery rate of females was only half as fast as males'. Conversely, offspring control resulted in an escalation of beg rate for the same degree of need (as measured by size and growth), and male-aided offspring obtained nearly three times more regurgitations than female-fed nestlings, yet grew at comparable rates and fledged at comparable sizes and ages. Hence, offspring demanded and obtained more food than they could effectively use.
Other avian parents also seem to use effective but potentially costly counterstrategies, and other avian offspring may demand more food than they require. Even if initial hatch asynchronies functioned in brood reduction, the parental strategies described here would allow parents to retain control over the timing and conditions for offspring loss.
Literature cited
ALTMANN, J., ALTMANN, S. A. & HAUSFATER, G. (1978). Primate infant's effects on mother's future reproduction. - Science 201, p. 1028-1030.
BENGTSSON, H. & RYDÉN, O. (1981). Development of parent-young interaction in asynchronously hatched broods of altricial birds. - Z. Tierpsychol. 56, p. 255-272.

37
-- & -- (1983). Parental feeding rate in relation to begging behavior in asynchro- nously hatched broods of the great tit Parus major. - Behav. Ecol. Sociobiol. 12, p. 243-251.
BERGER, J. (1979). Weaning conflict in desert and mountain bighorn sheep (Ovis canaden- sis) : an ecological interpretation. - Z. Tierpsychol. 50, p. 188-200.
BEST, L. B. (1977). Field sparrow nestling biology. - Auk 94, p. 308-319. BLICK, J. (1977). Selection for traits which lower individual reproduction. - J. Theor.
Biol. 67, p. 597-601. BROCKWAY, B. F. (1964). Ethological studies of the budgerigar (Melopsittacus undulatus).
Reproductive behavior. - Behaviour 22, p. 294-324. BRYANT, D. M. (1978a). Establishment of weight hierarchies in broods of house martins,
(Pelichion urbica). - Ibis 120, p. 16-26. -- (1978b). Environmental influences on growth rate and survival of nestling house
martins. - Ibis 120, p. 271-283. BURGER, J. (1981). On becoming independent in herring gulls: parent-young conflict. -
Am. Nat. 117, p. 444-456. CACCAMISE, D. F. (1976). Growth rate in the monk parakeet. - Wilson Bull. 88,
p. 495-497. CLARK, A. B. & WILSON, D. S. (1981). Avian breeding adaptions: hatching asynchrony,
brood reduction, and nest failure. - Q. Rev. Biol. 56, p. 255-277. CLUTTON-BROCK, T. H. (1983). The costs of reproduction to red deer hinds. - J. Anim.
Ecol. 52, p. 367-384. DAVIES, N. B. (1976). Parental care and the transition to independent feeding in the
young spotted flycatcher (Muscicapa striata). - Behaviour 59, p. 280-293. -- (1978). Parental meanness and offspring independence: an experiment with hand-
reared great tits Parus major. - Ibis 120, p. 509-514. DAWKINS, R. (1976). The selfish gene. - Oxford University Press, Oxford. FELDMAN, M. W. & ESHEL, 1. (1982). On the theory of parent-offspring conflict: a two-
locus genetic model. - Am. Nat. 119, p. 285-292. FUCHS, S. (1982). Optimality of parental investment: the influence of nursing on repro-
ductive success of mother and female house mice. - Behav. Ecol. & Sociobiol. 10, p. 39-51.
HAFTORN, S. (1982). Parental care of nestlings by the goldcrest (Regulus regulus). - Ornis Fennica 59, p. 123-134.
HARRIS, M. P. (1983). Parent-young communication in the puffin Fratercula arctica. - Ibis 125, p. 109-114.
HARTUNG, J. (1980). Parent-offspring conflict - a retraction. - J. Theor. Biol. 87, p. 815-817.
HUDSON, P. J. (1979). The parent-chick feeding relationship of the puffin, Fratercula arctica. - J. Anim. Ecol. 48, p. 889-898.
KHAYUTIN, S. N. & DMITRIEVA, L. P. (1979). Sensory organization of nest behavior in Parus major (Aves). - Zool. Zh. 58, p. 1835-1847.
KLUYVER, H. N., BALEN, J. M. VAN & CAVÉ, A. J. (1977). The occurrence of time-saving mechanisms in the breeding of biology of the great tit Parus major. - In: Evolutionary ecology. (B. STONEHOUSE & C. PERRINS, eds). MacMillan, London.
LACK, D. (1956). Further notes on the breeding biology of the swift (Apus apus). - Ibis 98, p. 606-617.
LOCKIE, J. D. (1955). The breeding and feeding of jackdaws and rooks. - Ibis 97, p. 341-369.
LÖHRL, L. B. (1968). Das Nesthäkchen als biologisches Problem. - J. Ornithol. 109, p. 383-395.
MACNAIR, M. R. & PARKER, G. A. (1979). Models of parent-offspring conflict. III. Intrabrood conflict. - Anim. Behav. 27, p. 1202-1209.

38
METCALF, R. A., STAMPS, J. A. & KRISHNAN, V. V. (1979). Parent-offspring conflict which is not limited by degree of kinship. - J. Theor. Biol, 76, p. 99-107.
MOCK, D. (submitted1). Sibilicidal aggression and resource monopolization in birds. -- (submitted2). Sibilicidal brood reduction: the prey-size hypothesis. NEUB, M. (1979). Brut biologische Konsequenzen des asynchronen Schlupfens bei
Kohlmeise (Parus major) und Blaumeise (Parus caeruleus). - J. Ornithol. 120, p. 196-214.
NOONAN, K. M. (1981). Sex ratios of parental investment in colonies of the social wasp (Polistes fuscatus). - Science 199, p. 1354-1356.
NORTON-GRIFFITHS, M. (1969). The organization, control and development of parental feeding in the oystercatcher (Haemotopus ostiolegus). - Behaviour 34, p. 55-114.
O'CONNOR, R. J. (1978). Brood reduction in birds: selection for fratricide, infanticide and suicide? - Anim. Behav. 26, p. 79-96.
PAGE, R. E. & METCALF, R. A. (1982). Multiple matings, sperm utilization, and social evolution. - Am. Nat. 119, p. 263-281.
PARKER, G. A. (1979). Models of parent-offspring conflict. IV. Suppression: evolutionary retaliation by the parent. - Anim. Behav. 27, p. 1210-1235.
- - & MACNAIR, M. R. (1978). Models of parent-offspring conflict. I. Monogamy. - Anim. Behav. 26, p. 97-111.
RICHTER, W. (1982). Hatching asynchrony: the nest failure hypothesis and brood reduc- tion. - Am. Nat. 120, p. 828-832.
RICKLEFS, R. E. (1965). Brood reduction in the curve-billed thrasher. - Condor 67, p. 505-510.
ROBINSON, J. V. (1980). A necessary modification of Trivers' parent-offspring conflict model. - J. Theor. Biol. 83, p. 533-535.
RYDÉN, O. & BENGTSSON, H. (1980). Differential and locomotory behavior by early and late hatched nestlings affecting the distribution of food in asynchronously hatched broods of altrical birds. - Z. Tierpsychol. 53, p. 209-224.
SAUNDERS, D. A. (1982). The breeding behaviour and biology of the short-billed form of the white-tailed black cockatoo, Catyptorhynchus funereus. - Ibis 124, p. 422-456.
SLAGSVOLD, T. (1982). Clutch size, nest size, and hatching asynchrony in birds: exper- iments with the fieldfare (Turdus pilaris). - Ecology 63, p. 1389-1399.
SMITH, G. A. (1972). Nesting and nestling parrots. - Avicult. Mag. 78, p. 155-164. -- (1975). Systematics of parrots. - Ibis 117, p. 18-68. SMITH, J. N. M. & ROFF, D. A. (1980). Temporal spacing of broods, brood size and
parental care in song sparrows (Melospiza melodia). - Can. J. Zool. 58, p. 1007-1015. STAMPS, J. A. (1980). Parent-offspring conflict. - In: Sociobiology: Beyond nature/
nurture? (G. W. BARLOW & J. SILVERBERG, eds), p. 589-618. Colorado, Westview Press.
--, METCALF, R. A. & KRISHNAN, V. V. (1978). A genetic analysis of parent-offspring conflict. - Behav. Ecol. & Sociobiol. 3, p. 369-392.
TRIVERS, R. L. (1972). Parental investment and sexual selection. - In: Sexual selection and the descent of man. (B. CAMPBELL, ed.), p. 136-197. Aldine, Chicago.
-- (1974). Parent-offspring conflict. - Am. Zool. 14, p. 249-264. - - & HARE, M. (1976). Haplodiploidy and the evolution of the social insects. - Science
191, p. 249-263. WERSCHKUL, D. F. & JACKSON, J. A. (1979). Sibling competition and avian growth rates.
- Ibis 121, p. 97-102. WYNDHAM, E. (1978). Ecology of the budgerigar Melopsittacus undulatus (Shaw) (Psittaci-
formes : Platycercidae). - Ph.D. Thesis, Univ. of New England, Armingdale, Australia 2351.
-- (1980a). Environment and food of the budgerigar (Melopsittacus undulatus. - Aust. J. Ecol. 5, p. 47-61.

39
-- (1980b). Diurnal cycle, behaviour and social organization of the budgerigar Melopsit- tacus undulatus. - Emu 80, p. 25-33.
-- (1981). Breeding and mortality of budgerigars (Melopsittacus undulatus). - Emu 81, p. 240-243.
-- (1982). Movements and breeding seasons of the budgerigar. - Emu 82, p. 276-282. ZAHAVI, A. (1977). Reliability in communication systems and the evolution of altruism.
- In: Evolutionary ecology (B. STONEHOUSE & C. M. PERRINS, eds). University Park Press, Baltimore.
Résumé
En dépit de l'intérêt largement répandu pour la théorie sur l'antagonisme génétique entre parents et progéniture, il y a peu de preuves expérimentales de l'existence d'un tel anta- gonisme dans la nature. Les modèles théoriques proposent deux solutions à ce conflit, qui ne sont pas prédites par les autres théories sur les rapports entre parents et progéniture: (1) la progéniture contrôle la répartition de l'investissement parental, et peut adopter des comportements de plus en plus exigeants (par exemple, solliciter) et demander des res- sources qui dépassent ce qu'elle peut utiliser efficacement; (2) les parents peuvent élabo- rer des contre-stratégies comportementales, qui empêchent la progéniture d'obtenir des ressources supplémentaires, mais qui sont plus coûteuses qu'une stratégie parentale de "laissez-faire", en permettant à la progéniture le contrôle. Ces prédictions ont été testées sur des perruches (Melopsittacus undulatus) dans de grandes volières, à Davis (Californi).
Les couvées de perruches éclosent de manière tout à fait asynchrone; néanmoins tous les oisillons étudiés grandissent à la même vitesse et acquièrent leurs plumes au même âge et pour une même taille. Cette indépendance de l'ordre d'éclosion et de la croissance sem- ble due principalement à la façon dont les mères perruches donnent la becquée: les femel- les nourrissent leurs oisillons d'abord d'après leur taille, et seulement ensuite prêtent l'attention à l'intensité de leurs sollicitations. Les oisillons d'un âge et d'une taille donnés sont traités de la même façon par leur mère, sans considération de leur ordre d'éclosion, et les oisillons sous-développés pour leur âge sont nourris comme s'ils étaient plus jeunes. Au contraire, les perruches mâles prêtent attention à l'intensité de la sollicitation des oisil- lons. Les mâles ont tendance à commencer à donner la becquée quand les oisillons sollici- tent, et à nourrir plus souvent ceux qui sollicitent avec plus de vigueur.
Cette différence dans le comportement de nourrissage des mâles a permis de comparer des couvées de taille égale dans lesquelles la femelle se charge presque exclusivement de l'alimentation des oisillons et d'autres dans lesquelles mâle et femelle donnent tous deux la becquée. Les parents contrôlent la répartition de la nourriture dans les couvées nourries par la femelle seule, tandis que les oisillons ont un contrôle plus grand sur la distribution de la nourriture dans les familles où le mâle nourrit aussi. Comme is avait été prédit par la théorie de l'antagonisme, la contre-stratégie de la femelle est efficace, mais peut être coûteuse: la vitesse à laquelle les femelles donnent la nourriture n'est que la moitié de celle des mâles. Réciproquement, le contrôle par les oisillons aboutit à une escalade dans l'intensité des sollicitations, pour le même degré de besoin (mesuré d'après la taille et la croissance); et les oisillons soignés par le mâle reçoivent presque 3 fois plus de régurgita- tions que les oisillons nourris par la femelle seule; mais ils grandissent à une vitesse com- parable et acquièrent leurs plumes à une taille et un âge comparables. Donc, les oisillons demandent et obtiennent plus de nourriture qu'ils ne peuvent en utiliser efficacement.
Dans d'autres espèces d'Oiseaux, les parents semblent également utiliser des contre- stratégies efficaces mais potentiellement coûteuses, et les oisillons peuvent demander plus




















