
Analytical Morphology: Mathematical Morphology of Decision Tables
Upload
khangminh22Category
view
0download
0
GROWTH AND MORPHOLOGY OF RABBIT MARGINAL VESSEL
ENDOTHELIUM IN CELL CULTURE
P. M. DAVISON, K. BENSCH, and M. A. KARASEK
From the Departments of Dermatology and Pathology, Stanford University School ofMedicine,Stanford, California 94305
ABSTRACT
A procedure for the isolation and cultivation of endothelium from the marginalvessels of the rabbit ear is described . Endothelial cells, isolated by slow perfusionwith a trypsin solution, are cultured in minimal essential medium supplementedwith 10% fresh rabbit serum for up to 6 mo . In primary culture, marginal vesselendothelial cells grow in an expanding circular pattern with closely apposed cellmembranes . Weibel-Palade bodies, subcellular organelles unique to endothelialcells in situ, are present in both primary and in serially cultivated cells (12passages) . In intact skin, Weibel-Palade (W-P) bodies are observed in the peri-nuclear cytoplasm in close proximity to the cell membrane facing the vascularlumen. 8-16 tubules of 200 A diameter are present in each body. In primary andsubcultured cells, W-P bodies of identical size are seen in the vicinity of the Golgiapparatus and in close proximity to the outer cell membrane . At the optimumserum concentration (10%), a cell doubling time of 72-96 h is observed . Whengrowth in normal rabbit serum and in platelet-poor serum is compared, a slowergrowth rate is observed in the absence ofplatelets, suggesting that factors releasedby platelets affect endothelial cell proliferation . However, addition of crudeplatelet factor does not substitute for complete serum. Fibroblast growth factor isnot mitogenic for rabbit marginal vessel endothelium in vitro .
Valuable information on many aspects of endo-thelial cell function and biochemistry has beenobtained using the techniques developed in recentyears for the in vitro cultivation of endotheliumfrom large vessels such as the umbilical vein andaorta (8, 13, 31) . Attempts have been made toadapt these methods for microvascular endothe-lium (19, 34), but the long-term cultivation of suchcells has not been achieved. In the skin, the role ofthe dermal vasculature in the maintenance of ho-meostasis is becoming increasingly apparent . Forexample, sprouting of new vessels occurs duringrevascularization after skin injury (23), and exten-sive vascular networks evolve to support the thick-
J . CELT. BIOLOGY © The Rockefeller University Press - 0021-9525/80/05/0187/12 $1 .00
Volume 85
May 1980
187-198
ened epidermis seen in several skin disorders (22)and during tumor growth (6) . To date, studies onthe regulation ofthe dermal vasculature have beenlimited to in vivo models . Differences in the ultra-structure of endothelium from the various regionsof the vascular network, with respect to the num-ber of Weibel-Palade (W-P) bodies (30), type ofendothelial cell junctions (29), or the extent of thecytoplasmic microfilaments (15), have indicatedthat large vessel endothelium cultivated in vitromay not be a suitable model for study of micro-vascular function . Classically, the marginal vesselsofthe rabbit ear have been used as a model systemfor the study of microvessel proliferation and the
187

development of a vascular network (26, 32) . Tosimplify investigation of the role of endothelialcells in these processes, we have developed amethod to isolate and maintain in vitro endothelialcells from the marginal vessels . A mixed popula-tion of endothelial cells from the marginal arteryand vein together with microvessel endotheliumfrom arterioles, venules, and capillaries is obtainedafter perfusion of the vasculature with trypsinsolution . The identification of the released cells asendothelial in type by morphologic and ultrastruc-tural criteria, and their growth characteristics andrequirements in vitro are described . A preliminaryreport of this work has appeared (4) .
MATERIALS AND METHODSIsolation of Cells
Adult New Zealand rabbits (2-3.5 kg) were killed by barbi-turate overdose and the ears were removed . The pelage was
COLLECTINGPOINT
DROPS OF PERFUSATECONTAINING RELEASEDCELLS
188
ARTERY
FIGURE 1
Schematic representation of steps involvedin the perfusion and collection of endothelial cells fromthe marginal vessels ofthe rabbit . The vascular networkof the rabbit ear is perfused with a 1% trypsin solutionvia a polyethylene cannula inserted into the marginalvein . After incubation at 37°C a small volume oftrypsinsolution (1 .0 ml) is perfused through the vasculature.Drops of perfusate containing released endothelial cellsare collected (at a point cut at the base of the ear) in acentrifuge tube containing MEM and 10% calf serum .
THE JOURNAL OF CELL BIOLOGY " VOLUME 85, 1980
clipped closely with an electric clipper (Oster Corp ., Milwaukee,Wis.), and a collecting point was cut at the base of the ear asillustrated in Fig. 1 . The cut surface was immersed in sterileHanks' balanced salt solution containing penicillin (400 U/ml)and streptomycin (200 pg/ml) for 10 min . A small incision wasmade in the marginal vein, and a polyethylene cannula (poly-ethylene tubing Intramedic No . 7405, Clay-Adams, Div . Becton,Dickinson & Co ., Parsippany, N . J .; i.d . 0.015 inches, o .d. 0 .043inches) with a beveled edge was inserted and pushed -I cm intothe vein. Blood was flushed from the vascular network of the earwith -2 ml ofan isotonic salt solution (GKN) pH 7 .2 (per liter :I g glucose, 8 g NaCl, 0 .4 g KCI) . This was replaced with GKNcontaining 1% trypsin (ICN Pharmaceuticals, Inc., Life SciencesGroup, Cleveland, Ohio), and the cannulated ear was placed ina large petri dish and incubated at 37°C for 50 min . Afterremoval of the ear from the incubator, -1 ml (15-25 drops) oftrypsin solution was perfused gently through the ear vasculature .The perfusate was collected in a centrifuge tube containing 2 mlof minimal essential medium (MEM) (Grand Island BiologicalCo . [GIBCO] Grand Island, N . Y.) containing 10% calf serum(Irvine Scientific Co ., Santa Ana, Calif) and antibiotics as shownin Fig. 1, and the ear was returned to the incubator for anadditional 30-min incubation after which the collection of cellswas repeated. The cells released from several ears were pooledand collected by centrifugation, resuspended in MEM containing10% rabbit serum, penicillin (200 U/ml), and streptomycin (100fig/ml), and plated onto 35 x 10 mm plastic petri dishes (LuxScientific Corp ., Newbury Park, Calif.) at a cell density of - l to3 x 10° cells/dish .
SubcultivationPrimary cultures which achieved at least 50% confluence
within 2 wk were used for subculture . Cells were released fromthe petri dish surface by incubation with 0 .1% trypsin, 0.33%EDTA in phosphate-buffered saline (PBS), pH 7.3 (per liter : 8 gNaCl, 2 g KCI, 1 .15 g Na2 HPO,, 0 .2 g KH2POa) for 4 min at37°C, dispersed by gentle agitation with a Pasteur pipette, andcollected by centrifugation at 800 g for t min . The cells wereresuspended in MEM with 10% rabbit serum with antibiotics andplated at a cell density of 5-10 x 10" cells/35-mm petri dish. Assubcultured cells approached confluence, they were further seri-ally cultivated using the procedure described above.
Cell IdentificationOTHER CELL TYPES
For morphologic comparison, rabbit epidermal keratinocyteswere isolated and grown in vitro as described by Liu and Karasek(16) . Fibroblasts were isolated from cell outgrowths of rabbitskin organ cultures (3) .
PREPARATION OF TISSUE CELLS FORELECTRON MICROSCOPY
Cells were released from the culture surface by incubationwith 0.1% trypsin, 0 .33% EDTA in PBS as described above forsubcultivation, resuspended in MEM with 10% rabbit serum (2 .0
FIGURE 2
Attachment and growth of marginal vessel endothelium in primary culture . (A) 24 h afterplating; (B) day 3 in culture ; (C) day l 1 in culture . x 3,926 .


ml/dish), then fixed by the addition of 3.0 ml of 2.0% bufferedglutaraldehyde solution, pH 6.8 (9, 25) . After centrifugation (1min at 800 g), the cell pellet was covered with a layer of freshfixative . After rinsing in isotonic wash solution, the cell pelletwas postfixed in l% osmium tetroxide for l h, dehydrated, andembedded in Maraglas (Marblette Co., Div. of Allied ProductsCorp ., Long Island City, N. Y.) (5, IS). 1-Pm-thick sections ofthe plastic-embedded preparation were stained with metachro-matic dyes for examination by light microscopy (21) . Ultrathinsections stained with uranyl acetate and lead citrate were exam-ined in a Siemens Elmiskop 101 (33) .
Pieces of skin measuring 1-2 mm' were removed from therabbit ear, immediately fixed in 2% phosphate-buffered glutar-aldehyde solution (pH 6.8), and subsequently prepared for elec-tron microscope examination as described above for the isolatedcells.
Cell Proliferation Studies
GROWTH REQUIREMENTS
SERUM TYPE AND CONCENTRATION :
Freshly isolated orfirst-passage serially cultivated rabbit marginal vessel endothelialcells were suspended in MEM containing 10% heat-inactivatedrabbit serum prepared in the laboratory (see below) and platedonto 35-mm plastic petri dishes at a plating density of t to 5 x10' cells/dish . Cells were maintained in a humidified atmosphereof 94% air, 6% CO s at 37°C. The medium was removed 24 hlater and the cells were refed with either the same medium orMEM containing 10% calf or horse serum (Irvine Scientific Co .),pooled human serum (see below), or rabbit serum (GIBCO). Allsera were heat inactivated at 56'C for 30 min. For other experi-ments, cells were grown in MEMsupplemented with 0.5, 2.0, or10% rabbit serum . In all cases cells were fed twice weekly for 14d. Growth achieved during this period was determined by count-ing the cell number or by '"PO,' - incorporation as describedbelow.
PLATELET-POOR SERUM : Platelet-poor serum (PPS) wasprepared from fresh rabbit blood carefully collected via heartpuncture in a chilled plastic syringe . The blood was immediatelycooled to 4'C and spun in a refrigerated centrifuge at 1,000 g for10 min to sediment the blood elements. The plasma was removedand further centrifuged at 4°C at 10,000 g for 10 min. Theplasma was decanted and clotting was initiated by the introduc-tion ofa glass stirring rod. PPS was squeezed from the fibrin clotand filtered through a Millipore filter (Millipore Corp., Bedford,Mass.) before heat inactivation at 56°C . Fresh rabbit bloodallowed to clot before removal of the blood elements was used asthe control rabbit serum in these experiments . The absence ofplatelet factors in PPS was demonstrated by the failure of 3T3cells to proliferate in the presence of PPS alone.
PLATELET EXTRACT :
Aplatelet growth factor waspreparedfrom human platelets obtained from outdated platelet-richplasma no longer suitable for clinical use, by the method ofAntoniades and Scher (1). The protein content of this boiledplatelet extract was determined by the method of Lowry et al .(14) and its activity quantitated by use of a 3T3 cell assay (20) .Addition of the platelet extract (1 .5-3 tag protein) to cultures of
190
THE JOURNAL OF CELL BIOLOGY - VOLUME 85, 1980
3T3 cells maintained in PPS was found to support the growth of3T3 cells which failed to survive in PPS alone.
MEDIA :
The growth of rabbit marginal vessel endotheliumin MEMsupplemented with either serum or PPS was comparedto that achieved in Dulbecco's modified essential medium(DMEM) .FIBROBLASTGROWTH FACTOR :
Fibroblastgrowth factor(FGF) of bovine pituitary origin was kindly supplied by Dr .Dennis Gospodarowicz (Cancer Research Institute, University ofCalifornia, San Francisco). The lyophilized FGF was dissolvedin MEM and diluted such that final concentrations of between50 and 500 ng/ml culture medium were achieved .
DETERMINATION OF CELLPROLIFERATION
CELL NUMBER : Endothelial cells were released from theculture surface by brief trypsinization as described above forsubcultivation . The number of cells in an aliquot of the resultingcell suspension was counted with a haemocytometer and a Nikoninverted phase-contrast microscope .
INCORPORATION of '' .2 P0,' - : Primary and serially cul-tured endothelial cells were incubated in complete medium con-taining 21UCi/mf2pO;3- (carrier free, Schwartz/Mann Div., Bec-ton, Dickinson &Co., Orangeburg, N. Y.) .''LPO,'- incorporationinto TCA-precipitable material was determined by scintillationcounting as described previously (16).
RESULTS
Isolation of CellsAt trypsin concentrations below 1%, few endo-
thelial cells were released even after extended (2-h) incubation at 37°C . Trypsin infiltration of thesurrounding tissues was a common occurrence,resulting in the release of various other nonen-dothelial cell types . The maximum yield of endo-thelial cells essentially free of other cell types wasobtained after a 50-min incubation with higherconcentrations of trypsin solution (between 1 and2% in GKN) followed by a further 30-min incu-bation and a second endothelial cell harvest . Thepercentage of nonendothelial cell types increasedmarkedly if the incubation time was extended .
Identification of CellsPHASE MICROSCOPY
We could detect no morphological differencebetween the cells harvested from the first andsecond collections . In both cases, the majority ofthe cells that settle down and attach within 24 h
FIGURE 3
Acomparison of the typical in vitro morphology of confluent cultures of various cell typesfrom rabbit skin : (A) frbroblasts; (B) endothelial cells; (C epidermal keratinocytes. x 4,640.

191

192
THE JOURNAL OF CELL BIOLOGY " VOLUME óS, 1980

FIGURE 5
Higher magnification ofthe Golgi complex showing W-P bodies in various stages of assembly .The mature bodies tend to align near the cell membrane facing the vascular lumen (arrowheads) . L,vascular lumen. x 56,000.
was released as small cell sheets (Fig. 2A) whichgrew into circular colonies of closely apposed po-lygonal cells with well-defined nuclei and indis-tinct cell borders. This circular configurationwhich was maintained throughout primary culture(Fig. 2 B and C) differed markedly from the mor-phology of rabbit dermal fibroblasts or epidermal
keratinocytes, Fig. 3) . The long, spindle-shapedfibroblasts (Fig . 3A) grew in parallel arrays, con-trasting with the cobblestone appearance of mono-layer cultures of endothelial cells (Fig . 3 B) . Epi-dermal keratinocytes (Fig . 3 C) grew in multilay-ered sheets and were smaller and more granularin appearance than the larger endothelial cells .
FIGURE 4
Part of a venule lined by two endothelial cells . The numerous electron-opaque round andelliptical bodies in the cytoplasm (between the nucleus and vascular lumen in these cells) usually occupiedby the Golgi complex are W-P bodies . These bodies are present in much smaller concentrations in theattenuated parts of the endothelial cells (between the nuclei) . L, vascular lumen. x 12,000 . Inset : Highermagnification showing a single, tangentially sectioned W-P body located in the attenuated cytoplasm ofan endothelial cell . x 82,000 .
DAVISON ET AL.
Endothelial Cell Culture 193

IN TISSUE SECTION : The endothelial cellslining the blood vessels of the rabbit ear, particu-larly the venules, contained moderate numbers ofregular, rod-shaped structures, usually referred toas Weibel-Palade (W-P) bodies . These organelleswere present singly in the attenuated parts of thesecells (Fig . 4, inset) . However, most of these bodies,often in clusters, were encountered in the perinu-clear cytoplasm, usually in the vicinity of the Golgiapparatus, which in these cells was present in abulging mass of cytoplasm between the nucleusand the vessel lumen (Fig . 4) . Another notablefeature was the propensity of W-P bodies to belocated in close proximity to the cell membranefacing the vascular lumen, as demonstrated in Fig.5 . Although it was possible to determine the di-ameter of the W-P bodies (0.1-0 .2 pm), their meanaverage length could not be accurately measuredas very few of them were found lying in theirentirety in the plane of an ultrathin section . Esti-mates made from tangentially cut W-P bodies
194
FIGURE 6 (a) A primary culture (10 d in vitro) shows both W-P bodies in the Golgi zone and theiralignment near the cell surface . x 22,000 . Inset : A section cut perfectly perpendicular to the long axis ofa W-P body shows the body substructure which includes parallel tubules in the electron-opaque matrix ofthe core . x 190,000 . (b) W-P body (arrowhead) in a cell of a subculture after 10 passages (5 mo in vitro).x 68,000 .
ULTRASTRUCTURE
THE JOURNAL OF CELL BIOLOGY " VOLUME 85, 1980
suggested that these structures were in excess of 2pm in length .
IN CULTURED ENDOTHELIAL CELLS : Inprimary culture and after 12 serial passages, thecells retained the fine structural features exhibitedby endothelial cells in tissue sections (Fig . 6 a) .The mitochondria did not change in appearanceand number, but there was an increase in thenumber of cisternae, particularly of the roughendoplasmic reticulum, and in the number of freeribosomes. Microfilaments were conspicuous, par-ticularly when they formed a dense, often bundled,feltwork close to the part of the cell membrane incontact with the culture vessel . Pinocytotic vesiclesand the Golgi apparatus remained prominent inthe cultured cells . Present in the vicinity of thelatter, but also frequently close to the outer cellmembrane, were W-P bodies . The W-P bodies ofrabbit cells had an electron-opaque core which, oncross section, was seen to contain tubular substruc-tures running parallel to the long axis of the body(Fig . 6 a, inset) . Each of these tubules had anelectron-translucent center surrounded by a thin

wall which is only slightly more electron-opaquethan the core matrix of the W-P body . The diam-eter of these tubules, 8-16 of which are present ineach body, was -200 A . The content of the W-Pbodies was separated from the cytoplasm by a unitmembrane-type envelope with little or no electron-translucent space between it and the core . W-Pbodies were abundant in primary cell cultures, 5-10 W-P bodies per section being found in all cells .Their incidence decreased after repeated serialcultivation, but W-P bodies with features anddimensions as seen in endothelial cells in situ couldstill be identified in cells after 10 serial passages(Fig . 6) . There were only one to two W-P bodiesper section of one of ten cells in these cultures. Itshould be noted that one can obtain at least 100sections per cell, but that the same W-P bodieswould of course also appear in more than onesection .
Cell Proliferation Studies
REQUIREMENTS FOR GROWTH INVITRO
SERUMTYPE :
The growth ofprimary and firstpassage subcultured cells in 10% heat-inactivatedrabbit serum (prepared in the laboratory), calf andhorse serum (Irvine Scientific Co.), and humanserum (blood bank) is shown in Fig . 7 . All serasupported proliferation of rabbit marginal vesselendothelial cells, but growth in commerciallyavailable rabbit serum failed to consistently equalthat seen in fresh rabbit serum ; moreover, therewas considerable variability among batches . Whenfresh rabbit serum (10% in MEM) was used rou-tinely, the cells retained their characteristic endo-thelial morphology for extended periods in culture .With horse serum and with some batches of freshhuman serum, an increased number oflipoid gran-ules (as occur at higher rabbit serum concentra-tions) appeared .SERUM CONCENTRATION : The rate of
growth of marginal vessel endothelial cells wasdependent upon serum concentration (Fig . 8) . In0.5% serum, the cells failed to proliferate but re-mained in a quiescent state (at least for 14 d), asgrowth could be stimulated by the return to higherserum concentrations . The rate ofcell proliferationincreased with increasing serum concentration toa maximum in 10% serum, at which a populationdoubling time of between 72 and 96 h for first-passage cells was achieved . Higher serum concen-trations (20-30%) did not further stimulate prolif-
DAY AFTER PLATING
FIGURE 7 The proliferation rate of marginal vesselendothelial cells in various sera is compared . (O) 10%Rabbit serum, (O) 10% human serum, (A) 10% calfserum, (") 10% horse serum .
eration. The rate declined and dark lipoid granules(positive to oil red O stain) were formed and filledthe perinuclear region .MEDIA : There was no significant difference
between the proliferation rate of rabbit marginalvessel endothelium in MEM or DMEM. The sim-pler medium MEM was used routinely thereafter .PLATING DENSITY : Freshly isolated endo-
thelium suspended in MEMwith 10% rabbit serumwas plated into 16-mm wells at various cell den-sities from 0.5 to 15 times those routinely used .
DAVISON ET At. .
Endothelial Cell Culture
195

s
ái0
swm
z
wU
0
yóM0
a
zO_HamO
OUz
iMaáM
FIGURE 8
Growth of rabbit marginal vessel endothe-lium (first-passage subculture) at various rabbit serumconcentrations . 0.5% Serum (O-O, "-"), 2% serum(O---O, "---"), and 10% serum (O-0, "-") .
The plating efficiencies were above 80%, and thedegree of cell confluence at 24 h corresponded tothe plating density. The relative rate of growthwas maximal at the "normal" plating density,declining for both the two highest (15 and 7.5 xnormal) and the lowest (0 .5 x "normal") platingdensities. No other differences in the cell mor-phology or growth pattern of cells plated at thedifferent cell densities were noted.PROLIFERATION RATE IN WHOLE SERUM:The growth of primary and first-, second-,fourth-, and twelfth-passage endothelial cells in10% rabbit serum is compared in Fig. 9. Exposureof primary cells to 0.1% trypsin and 0.33% EDTAto produce the first-passage cells stimulated theproliferative rate so that the population doublingdecreased from -5 d (rate seen after an initial lagperiod) to -72 h. Thereafter the rate of prolifera-tion decreased with each successive subculture andincreased length of time in culture . Cells main-tained in culture for 6 mo (twelfth passage) still
196
THE JOURNAL OF CELL BIOLOGY " VOLUME 85, 1980
contained W-P bodies but the cells failed to pro-liferate .PP s:
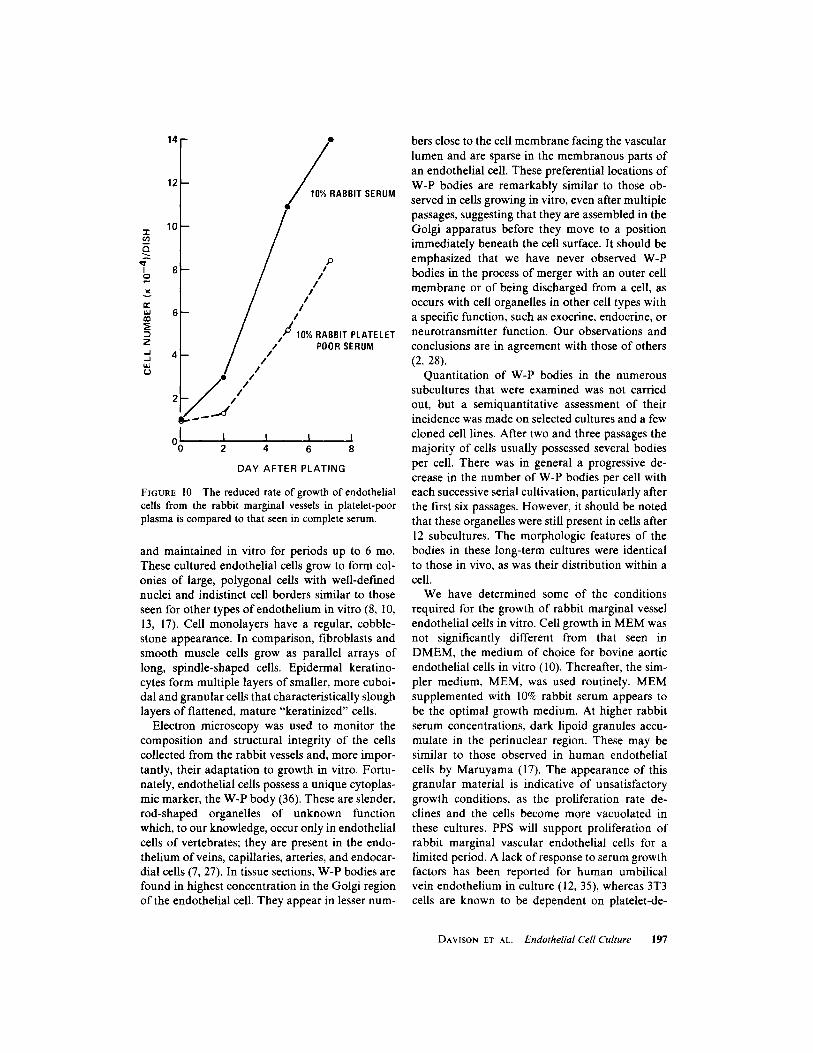
Rabbit marginal vessel endothelium wasfound to proliferate in MEM supplemented with10% PPS. The growth was not so rapid as in acomparable concentration of serum (Fig. 10). In-creasing the PPS concentration did not increasethe rate of growth. Cells maintained in PPS failedto reach confluence, and the cell number began todecline after 10 d in culture .PLATELET EXTRACT :
The addition of plate-let extract, 0.5 ml (15 .0 Pg protein/ml culturemedium), to cells maintained in PPS increased therate of growth above that in PPS alone (9 x 10'cells/dish on day 6 of culture, compared to 5 x10' cells/dish in PPS alone) . Early in the cultureperiod this growth rate approached that seen inwhole serum (10 x 10' cells/dish) on day 6. How-ever, the addition of platelet extract failed to ex-tend the proliferative period for cells maintainedin PPS, and by day 9 growth in PPS plus plateletfactor had fallen to 11 x 10' cells/dish comparedto 21 x 10' cells/dish in whole serum.FGF : FGF (50-500 ng/ml) failed to consist,
ently stimulate the growth of rabbit marginal ves-sel vascular endothelium maintained in whole se-rum or PPS. In contrast, FGF (10-50 ng/ml)stimulated the rate of growth of quiescent 3T3cells .
DISCUSSIONThe findings presented in this report show thatviable endothelial cells may be isolated from therabbit marginal vein, artery, and capillary network
s
ái0
mwm
zJLijU
DAY AFTER PLATING
FIGURE 9
Comparison of the proliferation rate of rab-bit marginal vessel endothelium in primary (ENP), first(ENS,), second (ENS2), fourth (ENS4), and twelfth(ENS,2) serial passage .
14
12
WU
4
2
DAY AFTER PLATING
FIGURE 10
The reduced rate of growth of endothelialcells from the rabbit marginal vessels in platelet-poorplasma is compared to that seen in complete serum .
and maintained in vitro for periods up to 6 mo .These cultured endothelial cells grow to form col-onies of large, polygonal cells with well-definednuclei and indistinct cell borders similar to thoseseen for other types of endothelium in vitro (8, 10,13, 17) . Cell monolayers have a regular, cobble-stone appearance . In comparison, fibroblasts andsmooth muscle cells grow as parallel arrays oflong, spindle-shaped cells . Epidermal keratino-cytes form multiple layers of smaller, more cuboi-dal and granular cells that characteristically sloughlayers of flattened, mature "keratinized" cells .
Electron microscopy was used to monitor thecomposition and structural integrity of the cellscollected from the rabbit vessels and, more impor-tantly, their adaptation to growth in vitro. Fortu-nately, endothelial cells possess a unique cytoplas-mic marker, the W-P body (36) . These are slender,rod-shaped organelles of unknown functionwhich, to our knowledge, occur only in endothelialcells of vertebrates; they are present in the endo-thelium ofveins, capillaries, arteries, and endocar-dial cells (7, 27). In tissue sections, W-P bodies arefound in highest concentration in the Golgi regionof the endothelial cell . They appear in lesser num-
hers close to the cell membrane facing the vascularlumen and are sparse in the membranous parts ofan endothelial cell . These preferential locations ofW-P bodies are remarkably similar to those ob-served in cells growing in vitro, even after multiplepassages, suggesting that they are assembled in theGolgi apparatus before they move to a positionimmediately beneath the cell surface . It should beemphasized that we have never observed W-Pbodies in the process of merger with an outer cellmembrane or of being discharged from a cell, asoccurs with cell organelles in other cell types witha specific function, such as exocrine, endocrine, orneurotransmitter function . Our observations andconclusions are in agreement with those of others(2, 28) .
Quantitation of W-P bodies in the numeroussubcultures that were examined was not carriedout, but a semiquantitative assessment of theirincidence was made on selected cultures and a fewcloned cell lines . After two and three passages themajority of cells usually possessed several bodiesper cell . There was in general a progressive de-crease in the number of W-P bodies per cell witheach successive serial cultivation, particularly afterthe first six passages . However, it should be notedthat these organelles were still present in cells after12 subcultures . The morphologic features of thebodies in these long-term cultures were identicalto those in vivo, as was their distribution within acell .We have determined some of the conditions
required for the growth of rabbit marginal vesselendothelial cells in vitro . Cell growth in MEM wasnot significantly different from that seen inDMEM, the medium of choice for bovine aorticendothelial cells in vitro (10) . Thereafter, the sim-pler medium, MEM, was used routinely . MEMsupplemented with 10% rabbit serum appears tobe the optimal growth medium . At higher rabbitserum concentrations, dark lipoid granules accu-mulate in the perinuclear region . These may besimilar to those observed in human endothelialcells by Maruyama (17) . The appearance of thisgranular material is indicative of unsatisfactorygrowth conditions, as the proliferation rate de-clines and the cells become more vacuolated inthese cultures . PPS will support proliferation ofrabbit marginal vascular endothelial cells for alimited period . A lack of response to serum growthfactors has been reported for human umbilicalvein endothelium in culture (12, 35), whereas 3T3cells are known to be dependent on platelet-de-
DAVISON ET AL .
Endothelial Cell Culture
197

rived growth factors (20) . Rabbit marginal vesselendothelium appears to require whole serum forextended in vitro cultivation . Although this issuggestive of a platelet factor requirement, addi-tion of one type of platelet preparation, a crudeboiled extract, does not provide a serum substitutefor these endothelial cells . A more purified plateletfactor preparation (24) may be more effective. Itis, however, likely that the growth requirements ofthe micro and small vessel endothelium may differfrom those reported for large vessel cells such ashuman umbilical vein or bovine aortic endothe-lium. The lack of response to FGF, a potentmitogen for bovine aortic endothelial cells in vitro(11), supports such a suggestion .
The authors wish to thank M . Ellen Charlton and IrmaDachne for their technical assistance.
This work was supported by grant 19595 of the U . S .Public Health Service .
Receivedfor publication 7 December 1979 .
REFERENCES
I . ANTONIADEs, H, N., and C. D. SCHER. 1977 . Radioimmunoassay of ahuman serum growth factor for Balb/c-3T3 cells: Derivation fromplatelets . Proc . Nail. Acad. Sci. U. S. A. 74:1973-1977 .
2. BURRI, P. H., and E. R. WEIBEL. 1968 . Beeinflussung eiher spezifichencytoplasmatischen Organelle von Endothelzellen durch Adrenalin . Z.Zellforch. Mikrosk. Anal . 88:426-440 .
3 . CRUICKSHANK, C. N. D. 1965 . In Cell and Tissues in Culture . E. N.Willner, editor . Academic Press, Inc., London. 549-568.
4. DAVISON, P., and M. A. KARASEx. 1978 . Factors affecting the growthand morphology of rabbit skin endothelial cells in vitro . Clin. Res . 26:208A .
5 . ERLANDSON, R. A. 1964 . Anew Maraglas, D. E. R. (R)732, embedmentfor electron microscopy . J. Cell Biol. 22 :704-709 .
6. FOLKMAN, J., and R. COTRAN . 1976 . Relation of vascular proliferationto tumor growth . Int. Rev. Exp. Palhol. 16 :207-248 .
7. FUCHS, A., and E. R. WEIBEL . 1966 . Morphometrische Untersuchungder Verteilung der Spezifischen Cytoplasmatischen Organelle in En-dothelzellen der Ratte. Z. Zellforsch. Mikrosk . Anal. 73:1-9 .
8 . GIMBRONE, M.A. . R. COTRAN, and J. FOLKMAN. 1974 . Human vascularendothelial cells in culture. J. Cell Biol. 60:673-684.
9. GORDON, G. B., L. R. MILLER, and K. B. BENSCH. 1963 . Fixation oftissue culture cells for ultrastructural cytochemistry . Exp. Cell Res. 31 :440443 .
10, GOSPODAROWIOz, D., J . MORAN, D. BRAUN, and C. R. BIRDWELL.1976 . Clonal growth of bovine vascular endothelial cells in tissueculture: Fibroblast growth factor as a survival agent. Proc. Nail. Acad.Sci. U. S. A. 73:4120-4124.
11 . GOSPODAROW ICz, D., G. GREENBURG, H. BIALE('KI, and B. R. ZETTER .1978 . Factors involved in the modulation of cell proliferation in vivoand in vitro . The role of Fiibroblast and epidermal growth factors in the
198
THE JOURNAL OF CELL BIOLOGY - VOLUME 85, 1980
proliferative response of mammalian cells . In Vitro (Rockville). 14:85-118 .
12 . HAUDENSCHILD. C. C., D. ZAHNISER, J. FOLKMAN, andM. KL .AGSBRUN .1976 . Human vascular endothelial cells in culture. Lack of response toserum growth factors . Exp. Cell. Res. 98:175-183 .
13 . JAFFE, E. A., R. L. N.ACHMAN, C. G. BF('KER, and R. MINICK . 1973 .Culture of human endothelial cells derived from umbilical veins. J.Clin . Invest. 52 :2745-2756 .
14 . LOWRY, O. H., N. J. ROSEBROUGH, A. L. FARR, and R. J . RANDALL.1951 . Protei n measurement with the Folin phenol reagent . J. Biol.Chem 193:265-275 .
15 . LUDATSCHER, R. H, 1978 . Ultrastructure of human dermal blood vesselswith special reference to endothelial filaments . Virehox-s Arch . B. Cell.Pathol. 27 :347-357 .
16 . LUI, S. C. . and M. A. KARASSK. 1978 . Isolatio n and serial cultivation ofrabbit skin epithelial cells . J. Invest. Dermatol. 70:288-293 .
17 . MARUYAMA, Y. 1963 . The human endothelial cell in tissue culture. Z.Zellforsch. Mikrosk . Anal. 60:69-79 .
18 . PALADE. G. E. 1952. A study of fixation for electron microscopy . J.Exp. Med. 95:285-298 .
19 . PANULA,P .,F.JOÓ,and L. RLCHARDT .1978.Evidence forthe presenceof viable endothelial cells in cultures derived from disassociated ratbrain . Experientia (Basel) . 34:95-96 .
20 . PLEDGER. W . J ., C. D. SCHER, P. MARTIN, H. N. ANFONIADEs, and C.D. STILES. 1977 . Growth requirements of normal and neoplastic cellcultures for platelet factor . J. Cell Biol. 75(2, Pt. 2): 401a.
21 . RICHARDSON, K. C., L. JARRETT, and E. H. FINKE. 1960. Embeddin g inepoxy resins for ultrathin sectioning in electron microscopy . StainTechnol. 35:313-323 .
22 . RYAN, T. J . 1975 . Pathophysiology of skin capillaries . Ins . J. Dermatol.14008-721 .
23 . RYAN, T. J . 1976 . Theblood vessels of the skin . J. Invest. Dermatol. 67 :1 to-118 .
24 . Ross, R., and A. VOGEL. 1978 . The platelet-derived growth factor . Cell .14:203-210 .
25 . SABATINI, D. D., K. BENSCH, and R. 1. B.ARRNETT. 1963 . Cytochemistryand electron microscopy . The preservation of cellular ultrastruetureand enzymatic activity by aldehyde fixation . J . Cell Biol. 17:19-58 .
26 . SANDISON, J. C. 1928 . The transparent chamber of the rabbit's eargiving a complete description of improved technique of constructionand introduction and general account of growth and behavior of bovinecells and tissues as seen with the microscope . Am . J. Anal. 41 :447-473 .
27 . SANTOLAN A, R. C., and F. BERTINI. 1970. Fine structure of endothelialcells of vertebrates. Distribution of dense granules. Z. Anat. Enrloick-lungsgesch. 131:148-155 .
28 . SENGEL_ A., and P. STOEBNER . 1970 . Golgi origin of tubular inclusionsin endothelial cells . J. Cell Biol. 44:223-225 .
29 . SIMIONESCU, M., N. SIMIONEscc . and G. E. PALADE . 1975 . Segmenta ldifferentiations of cell junctions in the vascular endothelium. J. Cell.Biol. 67 :863-885 .
30 . STEINSILPE. K. F. . and E. R. WEIBEL. 1970 . ElektronemikroskopischeUntersuchungen an Spezifischen Organellen von Endothaelzellen desFrosches (Rana temporaria) Z. Zellforsch . Mikrosk. Anal. 108:105-216 .
31 . THORGEIRSSON. G., and A. L. ROBERTSON . 1978 . The vascular endo-thelium pathobiologic significance . Am. J . Palhol. 93 :803-847 .
32. VAN DEN BRENK, H. A. S. 1959. Th e effect of ionizing radiation oncapillary sprouting and vascular remodelling in regenerative repairblastema observed in the rabbit ear chamber. Am. J Roentgenol.Radium Ther . 81 :859-884.
33 . VENABLE, 1. H. . and R. COGGERSH .ALL . 1965 . A simplified lead citrateslain for use in electron microscopy . J. Cell Biol. 25:407-408 .
34 . WAGNER, R. C., and M. A. MATTHEWS . 1975 . The isolation and cultureof capillary endothelium from epididymal fat . Microvasc . Res. 10:286-297.
35 . STRIKER, G. E. 1978 . Factors influencing endothelial cell proliferationin vitro. J. Cell Phvsiol . 96:203-214 .
36 . WEIBEL, E. R., and G. E. PALADE. 1964. Newcytoplasmic componentsin arterial endothelia . J. Cell Biol. 23:101-112,
Copyright © 2022 FDOKUMEN




















